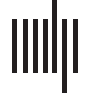ARTÍCULO DE REVISIÓN
The Role of Sensory Feedback in Developmental
Stuttering: A Review
un acceso abierto
diario
Abigail R. Bradshaw1
, Daniel R.. Lametti2
, and Carolyn McGettigan1
1Department of Speech, Hearing & Phonetic Sciences, University College London, Reino Unido
2Department of Psychology, Acadia University, Canada
Palabras clave: developmental stuttering, speech motor control, altered feedback paradigm, sensorimotor
integración, sensory feedback
ABSTRACTO
Developmental stuttering is a neurodevelopmental disorder that severely affects speech fluency.
Multiple lines of evidence point to a role of sensory feedback in the disorder; this has led to a
number of theories proposing different disruptions to the use of sensory feedback during speech
motor control in people who stutter. The purpose of this review was to bring together evidence
from studies using altered auditory feedback paradigms with people who stutter, in order to
evaluate the predictions of these different theories. This review highlights converging evidence for
particular patterns of differences in the responses of people who stutter to feedback perturbations.
The implications for hypotheses on the nature of the disruption to sensorimotor control of speech
in the disorder are discussed, with reference to neurocomputational models of speech control
(predominantly, the DIVA model; Guenther et al., 2006; Tourville et al., 2008). While some
consistent patterns are emerging from this evidence, it is clear that more work in this area is needed
with developmental samples in particular, in order to tease apart differences related to symptom
onset from those related to compensatory strategies that develop with experience of stuttering.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
Sensory feedback processing is known to play a crucial role in speech production, and forms a
central part of many models of speech motor control (Guenther et al., 2006; Hickok et al., 2011;
Parrell, Ramanarayanan, et al., 2019). Auditory feedback is thought to play a particularly impor-
tant role in learning to produce speech sounds early in development, as well as continuing to
guide and maintain accurate speech production throughout the lifetime (Guenther, 2016). Para
ejemplo, not only do congenitally deaf individuals struggle to develop typical speech production
(Herrero, 1975; Svirsky et al., 2004), but those who lose their hearing as adults can also show
gradual changes in articulation over time (Cowie & Douglas-Cowie, 1992). Disruptions to
somatosensory feedback have been shown to have even more profound effects on speech pro-
duction in the short term, with temporary oral anaesthesia resulting in highly inaccurate articu-
lación (Ringel & Steer, 1963; Scott & Ringel, 1971). Such evidence has led researchers to propose
that the internal targets that guide speech motor control are sensory in nature, and to stress the
importance of feedback loops that compare the intended/predicted and actual sensory conse-
quences of a speech production (Guenther et al., 2006; Parrell & Houde, 2019).
Citación: Bradshaw, A. r., Lametti, D. r.,
& McGettigan, C. (2021). El rol de
sensory feedback in developmental
stuttering: A review. Neurobiology of
Idioma, 2(2), 308–334. https://doi.org
/10.1162/nol_a_00036
DOI:
https://doi.org/10.1162/nol_a_00036
Supporting Information:
https://doi.org/10.1162/nol_a_00036
Recibió: 2 Octubre 2020
Aceptado: 16 Marzo 2021
Conflicto de intereses:
The authors have declared that no
competing interests exist.
Autor correspondiente:
Abigail R. Bradshaw
a.bradshaw@ucl.ac.uk
Editor de manejo:
Kate Watkins
Derechos de autor: © 2021
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
Stuttering and sensory feedback
Feedback perturbations:
Refer to experimentally induced
changes in the real-time sensory
feedback a speaker receives during
speech production.
Sensory Feedback and Developmental Stuttering
Developmental stuttering is a disorder that involves impairment in speech fluency. The speech of
people who stutter (PWS) is characterised by frequent repetitions and prolongations of syllables,
as well as tense pauses in which a speech sequence fails to be initiated (known as blocks). El
onset of the disorder typically occurs in early childhood, with a prevalence of up to 8% in pre-
school children (Yairi & Ambrose, 2013); sin embargo, the majority (alrededor 80%) spontaneously
recover during childhood, resulting in an incidence of chronic lifetime stuttering of around 1%
in the general population (Craig et al., 2002). Stuttering has been studied at multiple levels, de
genes to behaviour, yet the causes of stuttering remain unknown. One prominent account of the
speech motor control aspects of the disorder proposes that stuttering involves disruption to sen-
sorimotor integration. The most overt evidence for this comes from observations that dramatic
increases in the fluency of PWS can be achieved temporarily by altering auditory feedback during
speech (para una revisión, see Lincoln et al., 2006). In these studies, PWS speak into a microphone and
their speech is played back to them in real-time over headphones. Alterations of the feedback—
típicamente, delays, frequency shifts, or masking—can reduce stuttering frequency by up to 90%
(Bloodstein & Ratner, 2008; Foundas et al., 2013; Kalinowski et al., 1993). Sin embargo, estos
fluency-enhancing effects are temporary; effects can start to wear off as the speaker “adapts” to
the particular feedback alteration, and they do not persist once feedback is returned to normal.
Such observations have led to great interest in the role of sensory feedback in stuttering. A num-
ber of authors have proposed that disruption in the use of auditory feedback during speech motor
control may contribute to speech dysfluencies (p.ej., Max et al., 2004). There has also been some
interest in the idea that somatosensory feedback processing may be disrupted in PWS (Archibald
& De Nil, 1999; Loucks & De Nil, 2006). Neural evidence further points to a possible disruption
in sensorimotor integration, with reports of altered structure and function in multiple relevant
brain areas in PWS, such as the left inferior frontal gyrus (IFG), primary motor cortex (PMC),
and posterior superior temporal gyrus STG; (Brown y cols., 2005; Watkins et al., 2008; para una revisión,
see Chang et al., 2019).
Sin embargo, the precise nature of the disruption to sensory feedback processing in PWS remains
unknown. Sensory feedback has multiple roles to play in speech production, from calibration of
stored sensory targets to the guidance of online adjustments to an unfolding motor programme
(Parrell & Houde, 2019). Respectivamente, multiple theoretical perspectives have been suggested
that propose different sites of disruption to auditory feedback processing in stuttering (Chang
& Guenther, 2020; máx. & Daliri, 2019; Max et al., 2004). These make contrasting predictions
as to the behaviour of PWS in situations in which sensorimotor integration is required.
The Altered Feedback Paradigm
One major paradigm for investigating auditory-motor integration during speech is the altered
feedback paradigm (Houde & Jordán, 1998). This involves perturbation of the auditory speech
feedback a speaker hears in real time, typically in the form of a shift in fundamental frequency
(F0) or formant frequencies. These feedback perturbations can be either unexpected, with upward
and downward shifts applied randomly to utterances (Burnett et al., 1998); or sustained, semejante
that the same type and level of perturbation is consistently applied across multiple utterances
(Houde & Jordán, 1998). In both unexpected and sustained cases, participants are found to
demonstrate compensatory adjustments to their speech productions so as to oppose the effects
of the perturbation (ver figura 1 for more details). This has the effect of moving the auditory
feedback from utterances closer to baseline (pre-perturbation) niveles. Fundamentalmente, the responses
induced by unexpected and sustained perturbations of auditory feedback are proposed
Neurobiology of Language
309
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Cifra 1. Unexpected and sustained perturbation paradigms. (A) Unexpected perturbation para-
digms typically require participants to sustain phonation of vowel sounds for a duration of several
artículos de segunda clase. On a proportion of trials, feedback perturbations are unexpectedly applied with random
onset during vocalisation, typically with both upward and downward shift trials (upwards F0 pertur-
bation pictured). Speakers typically display reactive changes to the current vocalisation so as to
oppose this perturbation during the trial, termed the compensation response. (B) Sustained perturba-
tion paradigms typically require participants to repeat words (with normal vocalisation timing) across
many trials. After a baseline period with normal feedback, a feedback perturbation is then applied in
a consistent fashion across trials (upwards F1 perturbation pictured). Across many trials, the speaker
typically starts to gradually change their productions so as to oppose the perturbation, termed the
adaptation response. Feedback is typically then returned to normal for a final block of trials, to look
at after-effects of adaptation. Adapted from Burnett et al. (1998) and Lametti et al. (2018).
to reflect the operation of different underlying subsystems within speech motor control
(mira la sección, Feedforward versus Feedback control). These paradigms therefore offer much poten-
tial for teasing apart the nature of the possible deficits in auditory-motor integration in PWS.
AIMS OF THE CURRENT REVIEW
Over the last decade, there has been a growing number of studies using such altered feedback
paradigms with samples of PWS. Despite the prominence of the idea of disrupted sensorimotor
integration in stuttering, there has been no previous effort to bring together this evidence in order
to explicitly test and evaluate the predictions made by different theories. The current paper
therefore aims to provide the first review of studies using altered feedback paradigms with PWS,
with specific reference to predictions made by different theories of speech motor disruption in
stuttering. By reviewing this evidence, we aimed to identify if PWS show consistent differences
in their responses to altered auditory feedback that may reveal how sensory feedback processing
is disrupted in the disorder.
This review will begin by summarising the key theories that propose a disruption to the use of
auditory feedback in speech motor control in PWS. This will highlight the similarities and differ-
ences between theories, and their predictions for responses of PWS in altered feedback exper-
elementos. We will then review evidence from studies using the altered feedback paradigm with
PWS with reference to these predictions, in order to evaluate which theory is best supported
by the evidence. In so doing, we will identify key phenomena that must be explained by such
teorías, as well as areas that require further theory development.
Neurobiology of Language
310
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Feedback control:
A mode of motor control in which
sensory feedback is used to control a
movement online.
Feedforward control:
A mode of motor control involving
pre-planned motor commands that
are not corrected by sensory feedback
en línea.
Introduction to Speech Motor Control: The DIVA Model
Before proceeding to summarise key theories of feedback processing disruption in stuttering,
it is useful to provide a brief introduction to core concepts and terminology used within the
field of speech (and indeed non-speech) motor control.
Multiple formal models of speech motor control have been presented which, although draw-
ing on many of the same concepts, have distinct functional architectures. These include the
Directions into the Velocities of Articulators (DIVA) modelo (Guenther et al., 2006; Tourville
et al., 2008); the Task Dynamics framework (Saltzman & Kelso, 1987; Saltzman & Munhall,
1989); State Feedback Control (SFC) (Hickok et al., 2011; Houde & Nagarajan, 2011); y
the Feedback Aware Control of Tasks in Speech (FACTS) modelo (Parrell, Ramanarayanan,
et al., 2019). A recent comparative review of these different speech motor control model archi-
tectures was led by Parrell, Lammert, et al. (2019). The current review, sin embargo, will focus on the
DIVA model, because this is the only framework that has been applied to stuttering; en efecto, el
majority of the theories and studies reviewed in the current paper draw directly on the DIVA
modelo. To aid understanding throughout discussion of such research in the following sections,
we will therefore begin with a brief outline of some core concepts within computational models
of speech motor control, and how these are implemented in the DIVA model.
Feedforward versus Feedback control
At the centre of computational approaches to motor control is a primary distinction between two
types of control systems: feedback control and feedforward control. These two systems use sen-
sory feedback in different ways. The feedback control system uses sensory feedback for online
control of movements. During speech production, the sensory consequences of an articulatory
gesture are compared to the predicted/desired sensory outcome; any discrepancy between these
generates an error signal that is used to correct the ongoing movement trajectory online. en contra-
contraste, a purely feedforward control system issues pre-planned motor commands that are not cor-
rected online; sensory feedback is instead used to train and update these feedforward commands
offline, so that future movements are performed accurately from the outset. The DIVA model
combines these two control systems, with the feedback controller being further subdivided into
two modality specific controllers, one for somatosensory feedback and one for auditory feed-
atrás. There are thus three key components of speech motor control in DIVA: the feedforward
controller, the auditory feedback controller, and the somatosensory feedback controller. Discurso
motor commands are generated by summing the outputs of these three controllers. This control
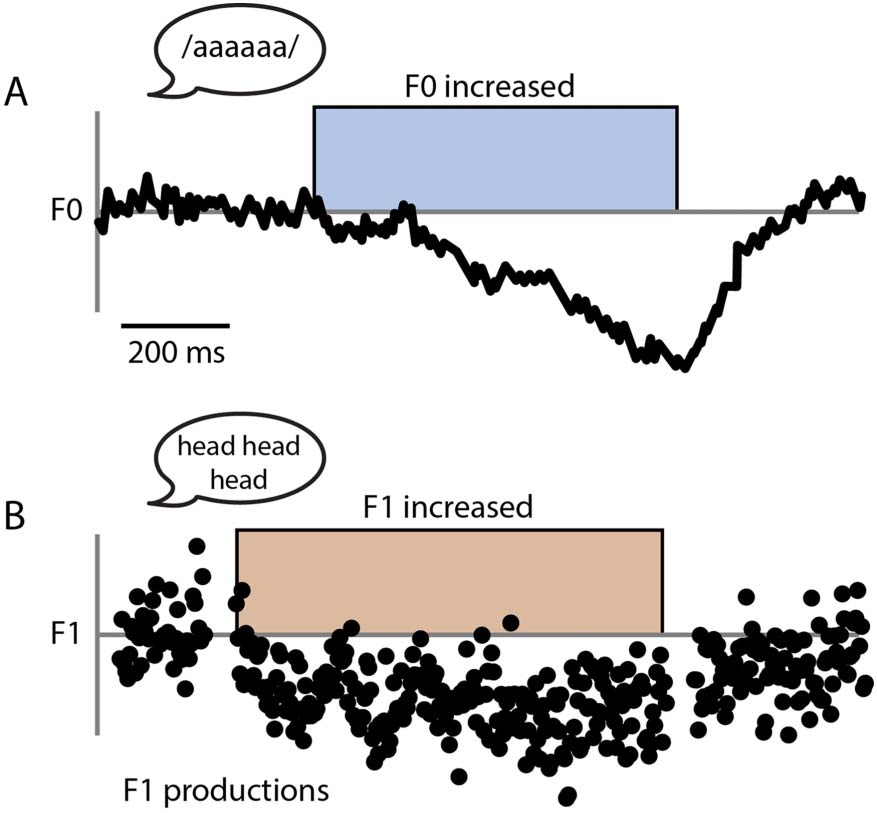
system is illustrated in Figure 2.
According to DIVA, speech motor control early in speech development relies exclusively on
feedback control, with motor commands being generated online using sensory (predominantly
auditory) comentario. Over time, the output from the feedback controller is used to train the feed-
forward controller, so that accurate feedforward commands can be learnt. This corresponds to
the formation of speech sound map nodes, which encode the motor, auditory, and somatosen-
sory target trajectories associated with each speech sound. Once these become sufficiently re-
liable, there is a shift in balance between the systems, such that the feedforward system takes
over as the dominant form of speech motor control. Sin embargo, the presence of the feedback con-
troller means that speech motor control remains sensitive to errors in sensory feedback. El
DIVA model can thus explain the ability of speakers to compensate for artificially induced per-
turbations of speech feedback during altered feedback experiments.
Feedback and feedforward modes of motor control have different associated benefits and
limitations. A feedback control system can allow for sensitivity to changes in auditory feedback,
Neurobiology of Language
311
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Cifra 2.
Simplified schematic of the DIVA model control system. Motor commands sent to the
vocal tract are the result of the summed output of the three control subsystems: the Feedforward
controller, the Auditory Feedback controller, and the Somatosensory Feedback controller. The feed-
back controllers compare sensory feedback to their corresponding sensory targets for a speech pro-
ducción, and if needed issue online corrective motor commands based on detected errors. Dashed
arrows indicate the use of these corrective commands to update the feedforward commands offline.
Adapted from Guenther (2016).
and as such is critical for supporting the initial learning of speech sounds as outlined above. A
speech motor control system that continued to rely purely on feedback control, sin embargo, would
be severely limited in the range of movement speeds it could handle; processing of sensory feed-
back involves delays of up to 150 EM, prohibiting rapid speech movements. These delays can
also result in greater instability in motor control, since feedback-based corrections to ongoing
movements are likely to be triggered too late in the speech sequence, leading to overshoots and
potentially oscillatory behaviour. A diferencia de, the feedforward system can generate motor com-
mands for an utterance before sensory feedback from that production is available, enabling the
production of fast and precise articulatory movements that characterise much of our connected
speech. The feedforward system, sin embargo, relies on the feedback system to detect any sensory
errors in produced speech, in order for stored feedforward commands to be updated so as to
reduce errors in future utterances. In this way, a control scheme that combines these two con-
trollers can balance their relative advantages and disadvantages.
En tono rimbombante, the two major types of altered feedback paradigm (unexpected versus sustained)
are proposed to differentially engage the two control systems. Unexpected perturbations are
thought to engage the feedback control system, which supports the online correction of sensory
errors during production of the perturbed utterance (hereafter termed compensation responses).
En cambio, sustained perturbations are proposed to induce sensorimotor learning via the feed-
forward system, in which there is a gradual updating of stored motor plans based on the consis-
tent errors in sensory feedback (hereafter termed adaptation responses). Evidence that such
changes in stored feedforward commands have occurred is found in the existence of aftereffects
of adaptation, in which changes to speech persist for a time after the perturbation has been re-
emocionado, with a gradual “wash-out” and return to baseline levels over time (Purcell & Munhall,
2006). As highlighted above, sin embargo, this updating of feedforward commands relies on the
detection of consistent errors in speech feedback by the feedback control system; de este modo, adapta-
tion responses in sustained perturbation paradigms reflect the joint operation of the feedback
and feedforward control systems. Some experiments have attempted to isolate the contribution
of the feedforward system during sustained perturbations, such as including noise-masked trials
in which feedback control is not possible (Houde & Jordán, 1998), or restricting analysis to an
Compensation responses:
The changes a speaker makes to an
ongoing vocalisation to counteract an
unexpected perturbation of sensory
comentario.
Adaptation responses:
Correspond to the gradual changes
made across repeated speech
productions to counteract a sustained
perturbation of sensory feedback.
Neurobiology of Language
312
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Forward models:
Used in motor control to translate
motor commands into their predicted
or desired sensory consequences.
Inverse models:
Used in motor control to translate
sensory outcomes into the motor
commands needed to achieve them.
early time-window within each utterance before feedback-based corrections can be initiated
(Parrell et al., 2017). Keeping this point in mind, it can still be seen that contrasting these two
perturbation paradigms can allow for comparisons of the operation of the feedback versus
feedforward control systems.
Internal models in speech motor control
Both the feedback and feedforward control systems make use of what are termed internal
modelos. These act to translate between motor commands and their associated sensory out-
comes. Two types of internal models are described in the speech motor control literature:
forward models, which translate motor commands into predicted sensory consequences; y
inverse models, which translate sensory outcomes into motor commands. Inverse models are
used by the feedforward system to enable transformation of a desired sensory goal into the motor
commands needed to achieve that goal. The feedback system uses forward models to allow a
comparison between incoming sensory feedback from a speech articulatory gesture and some
kind of prediction of that feedback. The source of that prediction, sin embargo. varies between dif-
ferent theories. One long-standing idea is that the prediction is obtained from an “efference
copy” of the motor commands (Wolpert et al., 1995); específicamente, when motor commands
are sent to the articulators, a prediction of their sensory consequences is simultaneously sent
to sensory cortex via reafference, in order to allow a “subtraction” of incoming sensory input
from this sensory prediction. This process of prediction generation through efference copy is
used in the SFC and FACTS model frameworks, in order to help provide an internal estimate
of the current state of the articulators (see Parrell & Houde, 2019, para una revisión). Comparison
of this prediction with incoming sensory feedback can then be used to update this estimate.
Although early versions of the DIVA model also used efference copy as a source of sensory pre-
dictions in forward models (Guenther, 1995; Guenther et al., 1998), the most recent versions
propose that these predictions come from the sensory target/goal for the intended utterance.
Eso es, the forward model allows a comparison between the intended and actual incoming sen-
sory feedback. In DIVA, inverse models in the feedback system are then used to translate any
detected sensory error into corrective motor commands. De nuevo, sin embargo, it should be noted that
this proposal is unique to DIVA; the motor control models from which these terms originated do
not consider inverse models to be part of the feedback system or to receive any information on
sensory errors (Wolpert et al., 1998). En general, sin embargo, it can be seen that sensory feedback has
a key role to play in establishing the sensory-motor mappings that underpin these internal
modelos.
Neural basis of the DIVA model
A key feature of the DIVA model is that it is not only mathematically defined at the computational
nivel, but also neurally specified. De este modo, in the model, different components of the feedback
and feedforward control systems have been attributed to different neural substrates. These are
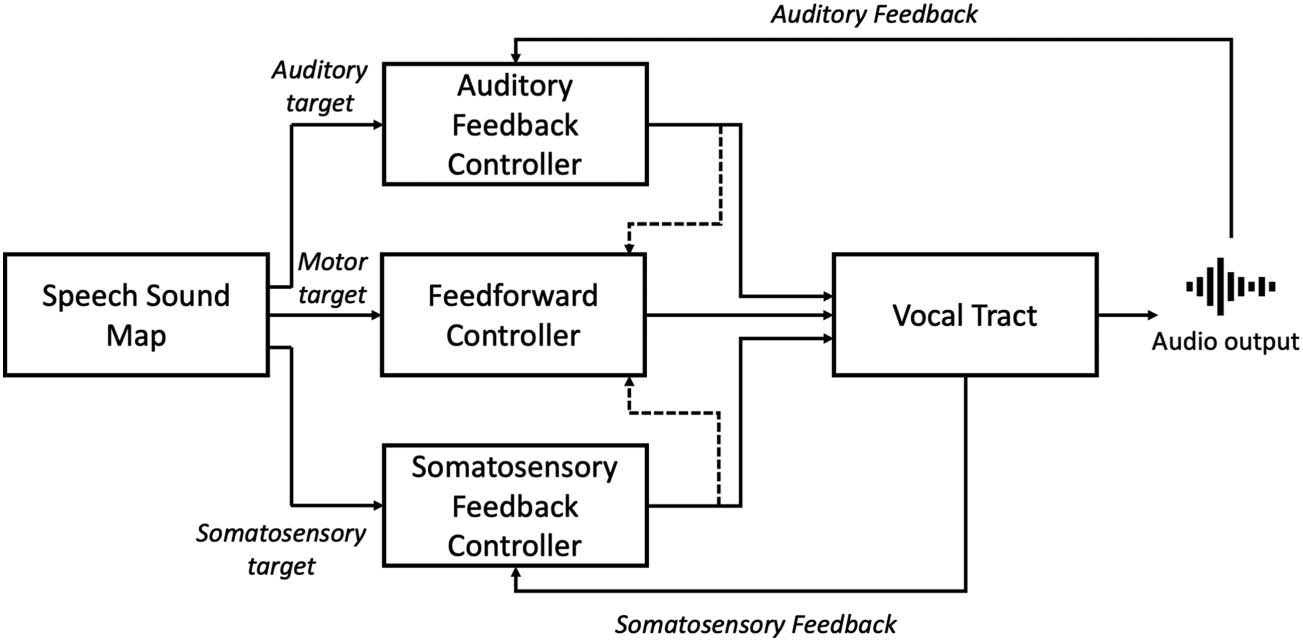
illustrated in Figure 3. Production of a speech sound begins with activation of a speech sound
map node in the left ventral PMC; these are activated by initiation nodes in the supplementary
motor area (SMA), in turn activated by either the pre-SMA or basal ganglia. Projections from the
speech sound map to the articulator map in bilateral primary motor cortex then constitute the
feedforward motor commands for a speech sound. In the feedback control system, projections
from the speech sound map to posterior auditory cortex (pSTG) act as a forward model that
predicts the intended sensory consequences of the target speech sound. Posterior auditory
cortex then facilitates comparison of this sensory target with actual sensory feedback, cual
may result in activation of an auditory error map. Projections from posterior auditory cortex to
Neurobiology of Language
313
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Cifra 3. The DIVA model of speech motor control. Schematic shows different components involved in speech motor control and their
hypothesised neural bases according to the DIVA model. Note that loops through subcortical structures are not shown on this diagram.
Adapted from Guenther (2016).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
a feedback control map in right vPMC translate this auditory error into corrective motor com-
mands (via inverse models), which are sent on to the articulator map in motor cortex.
Across these various points of processing, the DIVA model proposes the use of both cortico-
cortical and cortico-subcortico-cortical connections, via structures such as the basal ganglia and
cerebellum. The DIVA model has also been extended to account for the planning, timing, y
coordination of multisyllabic speech sequences; this is the Gradient Order DIVA (GODIVA)
modelo (Bohland et al., 2010). This incorporates processing within a range of brain regions in-
volved in working memory and motor sequencing, such as the posterior inferior frontal sulcus,
vPMC, SMA, pre-SMA, and basal ganglia. Curiosamente, brain imaging evidence from samples of
PWS has reported abnormalities in many of the regions implicated in the DIVA and GODIVA
modelos, including the left inferior frontal gyrus (IFG), the pSTG, and the basal ganglia (for a
revisar, see Chang et al., 2019).
Theories of Disruption to Feedback Processing in Stuttering
Multiple authors have suggested disruptions to the normal use of auditory feedback in speech
motor control in stuttering, often based in disturbances of the relative balance of dominance
between the feedforward and feedback control systems. These theories will be reviewed in
the following sections, ending with a summary of their key predictions for responses to alter-
ations of auditory feedback during speech production in PWS.
The Speech Motor Skills account
The Speech Motor Skills (SMS ) view of stuttering proposes that PWS are at the lower end of a
motor skill continuum (van Lieshout et al., 1996a, 1996b). In typical individuals, execution of
movements gradually shifts from explicit to implicit control with practice, resulting in the
movement becoming automated. Other hallmarks of skilled movement control include being
Neurobiology of Language
314
Stuttering and sensory feedback
energy efficient, having the ability to flexibly adapt to changing task requirements, and being
able to optimise movements achieving specific goals. A diferencia de, limited motor skill would
be reflected in error prone movements that are more variable, more slowly executed, menos
flexible, and that show limited improvement with practice. According to the SMS account of
stuttering, limited speech motor skill in PWS thus results in speech movements that are less auto-
matised and efficient (es decir., less implicit). This view is based upon a body of evidence reporting less
proficient motor performance in PWS, across both speech and nonspeech tasks. En particular, PWS
appear less able to benefit from practice during motor skill learning, as measured during finger
tapping (Smits-Bandstra & De Nil, 2007), nonsense word learning (Namasivayam & camioneta
Lieshout, 2008), and syllable sequencing tasks (Smits-Bandstra et al., 2006).
In order to compensate for this reduction in motor skill, PWS are proposed to increase depen-
dence on sensory feedback during speech motor control. Por ejemplo, this can be achieved by
slowing the rate of speech, something that has been reported in the fluent speech of PWS
(Zimmermann, 1980) and is frequently employed in therapeutic interventions (Bloodstein &
Ratner, 2008; Onslow & Ingham, 1987). De este modo, according to this view, differences in the use
of sensory feedback for speech motor control in stuttering are part of a compensatory strategy;
eso es, feedback control itself is not disrupted and can be used to help prevent stuttering.
Sin embargo, there are costs involved with this increased reliance on feedback control:
Movements are more time-consuming, they place greater demands on attentional resources,
and the range of movement speeds that can be dealt with effectively by the system is restricted
(es decir., slower movements are favoured). Relating this to the DIVA model framework, this account
can be conceptualised as a reduced reliance on feedforward planning and an increased reliance
on feedback control.
Unstable or insufficiently activated internal models
In a seminal paper, Max et al. (2004) presented two hypotheses regarding potential causes of
speech dysfluency in stuttering, based within the DIVA model framework. The first of these hy-
potheses proposed that stuttering involves disruption to the internal models that facilitate the
transformation between motor commands and sensory consequences. In this account, stuttering
is proposed to involve an impairment in the ability to use auditory feedback early in develop-
ment to establish such bidirectional mappings between motor commands and sensory conse-
quences. The authors suggest that this could be underpinned by a disruption to cerebellar or
basal ganglia based learning.
Disruption to the learning, retention, and updating of both types of internal models will have
multiple implications for speech motor control. En primer lugar, disrupted inverse models will result in
inaccurate feedforward motor commands, increasing the need for feedback-based correction
of errors. On top of this, disruption to forward models will result in inaccurate prediction of
the expected sensory consequences of those commands within the feedback control system.
In this way, there is both an increase in production errors, and simultaneously an impairment
in the ability of the feedback system to anticipate and correct for such errors. En efecto, such faulty
forward model predictions could result in error signals being erroneously generated, triggering a
correction of otherwise correctly executed movements.
Por último, the system will be forced to rely more on a purely afferent feedback control strat-
uno (es decir., reliant on actual sensory feedback without any forward modelling or prediction of that
comentario). Such over-reliance on feedback control is associated with increased instability, pendiente
to delays inherent in feedback processing (see previous section, Feedforward versus Feedback
control). Like the speech motor skills account, this theory thus proposes that slowing of speech
Neurobiology of Language
315
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
can act as a compensatory strategy for PWS. Longer movement durations would allow the sys-
tem to make better use of afferent feedback processing, in the face of faulty modelling of feed-
atrás. Por lo tanto, slowed speech is again seen as a compensatory mechanism, rather than as a
symptom of an underlying limitation.
A slight variation on these ideas was proposed by Hickok et al. (2011), in relation to their
hierarchical state feedback control model of speech production. They similarly suggested that
stuttering involves inaccurate forward predictions of the sensory consequences of a speech ges-
tura. Sin embargo, this was attributed to noisiness in the mapping between an internal estimate of
the state of the vocal tract and the sensory system. This will result in inaccurate generation of
error signals, as proposed above. They suggested that noisiness in this mapping was caused by
disruption to area Spt (a region located at the parietal-temporal boundary in the left posterior
Sylvian fissure).
It is worth highlighting here that while the DIVA model assumes the use of inverse models in
the feedback system to translate detected sensory errors into corrective motor commands, este
view is not shared by the Unstable Internal Models hypothesis; por lo tanto, discussion of faulty
inverse models in the feedback system is not included in this theory.
Overreliance on feedback control
The second hypothesis put forward by Max et al. (2004) proposed that stuttering involves weak-
ened feedforward control that leads to an overreliance on feedback control. Eso es, early in
childhood, stuttering individuals fail to show the normal transition from greater reliance on feed-
back control to increased reliance on the feedforward system for mature speech motor control.
Unlike in the SMS account of stuttering, sin embargo, this increased reliance on sensory feedback is
not considered to help compensate for stuttering, but instead is a cause of speech dysfluency.
Específicamente, the delays inherent in afferent feedback cause speech motor control to become un-
stable. Although similar to the first hypothesis presented by these authors (reviewed above), crit-
ically this hypothesis does not assume any disruption to internal models or to the feedback
system itself. En cambio, the core site of disruption is within feedforward control, forcing the system
into an overreliance on feedback control of speech.
This theory has been further developed in subsequent papers by Guenther and colleagues.
Civier et al. (2010) proposed that an overreliance on feedback leads to an accumulation of large
sensorimotor errors, due to the time-delays associated with detection and correction of errors in
the feedback system. These errors are considered to be present in the fluent speech of PWS;
sin embargo, when they become sufficiently large, a hypothesised “monitoring subsystem” detects
the excessive error. This triggers a motor reset in which the system attempts to repair the error by
restarting the syllable, resulting in a repetition. According to this model, each repetition consti-
tutes an unsuccessful production (es decir., a production with high error); corrective commands sent
to the articulators should mean that each successive repetition involves progressively less audi-
tory error. The intended utterance can then proceed (es decir., fluent speech continues) cuando el
error has been reduced to a sufficiently low level.
Simulations within the DIVA and GODIVA models have been used to provide support for this
cuenta (Civier et al., 2010; Civier et al., 2013). Using the DIVA model, Civier et al. (2010) eran
able to replicate multiple features of speech in PWS by simulating reduced gain of feedforward
and increased gain of feedback control. Específicamente, this “stuttering DIVA” simulation produced
repetitions of syllables, as well as auditory errors in fluent speech in the form of delayed formant
transitions. This was especially seen on tokens that involved bilabial consonants followed by
high-F2 vowels (p.ej., /bid/). This latter finding replicates findings of slowed and delayed formant
Neurobiology of Language
316
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
transitions in PWS, again typically on transitions involving high-F2 vowels (Chang et al., 2002;
Robb & Blomgren, 1997). According to Civier et al. (2010), overreliance on the feedback system
would be expected to have a particularly detrimental effect on production of such tokens, as they
require the production of rapid transition rates that cannot be supported by the sluggish feedback
sistema. Curiosamente, simulations of slowed speech and masking of auditory feedback were
found to reduce dysfluencies in the model. Further simulations by Civier et al. (2013) usando
the GODIVA model suggested that the nature of the disruption to the feedforward system may
involve either a disconnection of cortico-striatal pathways or a dysregulation of the dopamine
sistema. Both these simulated disruptions were found to result in stuttering behaviour due to
delayed readout of the motor programme for the next syllable in a speech sequence.
De nuevo, it should be stressed that in this account both the feedback control system and the
sensitivity of PWS to sensorimotor errors is considered to be normal; the only site of impairment
is in the read-out of feedforward commands, which causes a bias towards feedback-based con-
controlar. Sin embargo, even though the feedback system is functioning normally, it is itself the cause of
speech errors due to its associated limitations, Por ejemplo, in the range of movement speeds that
it can handle. This is proposed to underlie the slowing of speech typically seen in PWS; eso es,
rather than being seen as an intentional compensatory strategy to aid fluency, slowness of
speech is construed as an inevitable result of a dysfluency-inducing weakness (eso es, overreli-
ance on feedback control). As previously described, this limited speed of speech can itself then
lead to increased sensorimotor errors, as some formant transitions do not occur at their target
velocidad. The efficacy of masking auditory feedback in improving fluency is attributed to a reduced
ability to detect small errors in sensory feedback, and thus a reduced likelihood of a repetition
being triggered. In this way, any conditions that lead to a reduction in the detection of speech
errors by the feedback control system would be predicted by this account to improve speech
fluidez.
Impaired left-hemisphere basal ganglia motor loop
Recientemente, Guenther and colleagues have proposed a variation on these ideas concerning the role
of auditory feedback processing in stuttering (Chang & Guenther, 2020; Guenther, 2016). En esto
cuenta, stuttering is rooted in disruption of the basal ganglia motor loop, thus building on pre-
vious models implicating this group of structures in the pathology underlying stuttering (Alm,
2004; Giraud et al., 2008). After considering a number of possible disturbances to basal ganglia
functioning, Guenther and colleagues focus on an account that considers interactions between
auditory feedback and the basal ganglia “initiation circuit.” This account draws on the GODIVA
model’s proposed role of the basal ganglia in generating initiation and termination signals for
moving between motor programmes representing different phonemes within a speech se-
quence. Fundamentalmente, this process is guided by monitoring of the current cognitive, motor, and sen-
sory context; when the basal ganglia detect a match between the current context and that
required by an upcoming speech gesture, they signal the SMA to initiate the next phoneme in
the sequence.
According to this account, the core deficit in stuttering is in left hemisphere cortical areas
involved in feedforward control, which results in small articulatory errors in speech. These errors
affect the fluid initiation and termination of speech motor commands by the basal ganglia.
Específicamente, detection of these errors in auditory feedback means that the current sensory con-
text does not match that required for initiating the next motor programme. The basal ganglia are
consequently unable to move forward in the speech sequence, resulting in dysfluency.
Por lo tanto, although the core disruption in this account is in the feedforward system, it is process-
ing within the feedback system that ultimately leads to breakdowns in speech fluency.
Neurobiology of Language
317
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
In this way, involvement of auditory feedback control mechanisms is considered by this ac-
count to be maladaptive and a major contributing factor to stuttering, not a compensatory strat-
uno. En cambio, it is proposed that PWS may in fact try to suppress processing of auditory feedback
in order to prevent the detection of feedback errors that contribute to speech dysfluencies. Esto es
based on findings of reduced activity in auditory cortical regions in PWS, identified by meta-
analyses as one of the most common findings across imaging studies in stuttering (Marrón
et al., 2005; Budde et al., 2014). Por lo tanto, in contrast to previous proposals related to over-
reliance on feedback, this account predicts a gradual reduction in reliance on feedback with
experience of stuttering, as a learnt compensatory strategy. Fluency enhancing conditions that
involve alterations of speech feedback (p.ej., delaying, masking) are also proposed to reduce or
prevent detection of articulatory errors, allowing speech sequences to unfold unimpeded.
Disrupted auditory prediction and feedback monitoring
A distinct view of the role of auditory feedback in stuttering was presented in a recent paper by
Max and Daliri (2019). In contrast to previously reviewed accounts that tend to assume no dis-
ruption to the processing of feedback itself, this account argues for impaired auditory feedback
processing in PWS. Específicamente, they proposed that stuttering involves a disruption in the use of
auditory prediction to prime the auditory system prior to onset of a speech movement. Semejante
priming may optimise the tuning characteristics of auditory cortex, in order to prepare for pro-
cessing of upcoming auditory feedback. A failure to do so may lead to aberrant feedback-driven
corrective commands, triggering unnecessary repairs of speech movements to result in a break-
down of fluency during speech production. In this account, por lo tanto, it is not the process of
feedback monitoring that is affected per se, but processes involved in priming the auditory
cortices prior to movement onset that have a knock-on effect for their upcoming role in sensory
error detection.
This hypothesis was based on a series of studies reporting a lack of pre-speech auditory mod-
ulación (PSAM) in people who stutter (Daliri & máx., 2015a, 2015b, 2018). Específicamente, in peo-
ple who do not stutter (PWNS), the auditory evoked potential (AEP) measured in response to an
auditory probe was significantly smaller when presented during a period of speech motor plan-
ning in a delayed-response speech task, than when presented at the same point in time in a silent
reading condition. En cambio, PWS were found to lack this PSAM effect. This normal reduction
of the AEP in PSAM was not interpreted as a general suppression of the auditory system, pero
rather an enhancement of its sensitivity to auditory feedback, and specifically, to errors in that
comentario. Curiosamente, the same pattern of results was found when investigating auditory mod-
ulation during a listening condition, in which a written word stimulus was followed by a record-
ing of the participant’s voice speaking the word (Daliri & máx., 2015a). De nuevo, PWNS showed
significant modulation of the AEP in this listening condition relative to silent reading, mientras
this modulation effect was absent in a group of PWS. They thus argued that reduced PSAM in
PWS reflects a general disruption to prediction of upcoming auditory inputs, whether self- o
externally-generated, rather than a disruption to motor command planning.
According to these authors, production of simple monosyllabic words in isolation is assumed
to be reliant on feedforward control mechanisms; production of longer more complex multisyl-
labic utterances is by contrast considered to place greater demands on feedback monitoring and
error correction, thus increasing the likelihood of a breakdown in fluency. This is in contrast to
previously reviewed theories, which consider the feedback system to be restricted in its ability to
handle faster movement speeds as in fluid connected speech utterances, thus benefiting from a
slowing of speech utterances. The authors also propose that the mechanisms involved in PSAM
Neurobiology of Language
318
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
may contribute more to online feedback-driven corrections, being less involved in longer-term
speech adaptation and updating of stored internal models.
En general, the functional relevance of PSAM is as yet unproven. Promising evidence for its con-
tribution to feedback control, sin embargo, is suggested by the finding that PSAM is reduced in
PWNS when speaking with predictable delays in auditory feedback (Daliri & máx., 2018); este
is consistent with the idea that PSAM-related mechanisms for enhancing feedback monitoring
may disengage when auditory feedback is rendered non-informative.
Comparison of theories and predictions
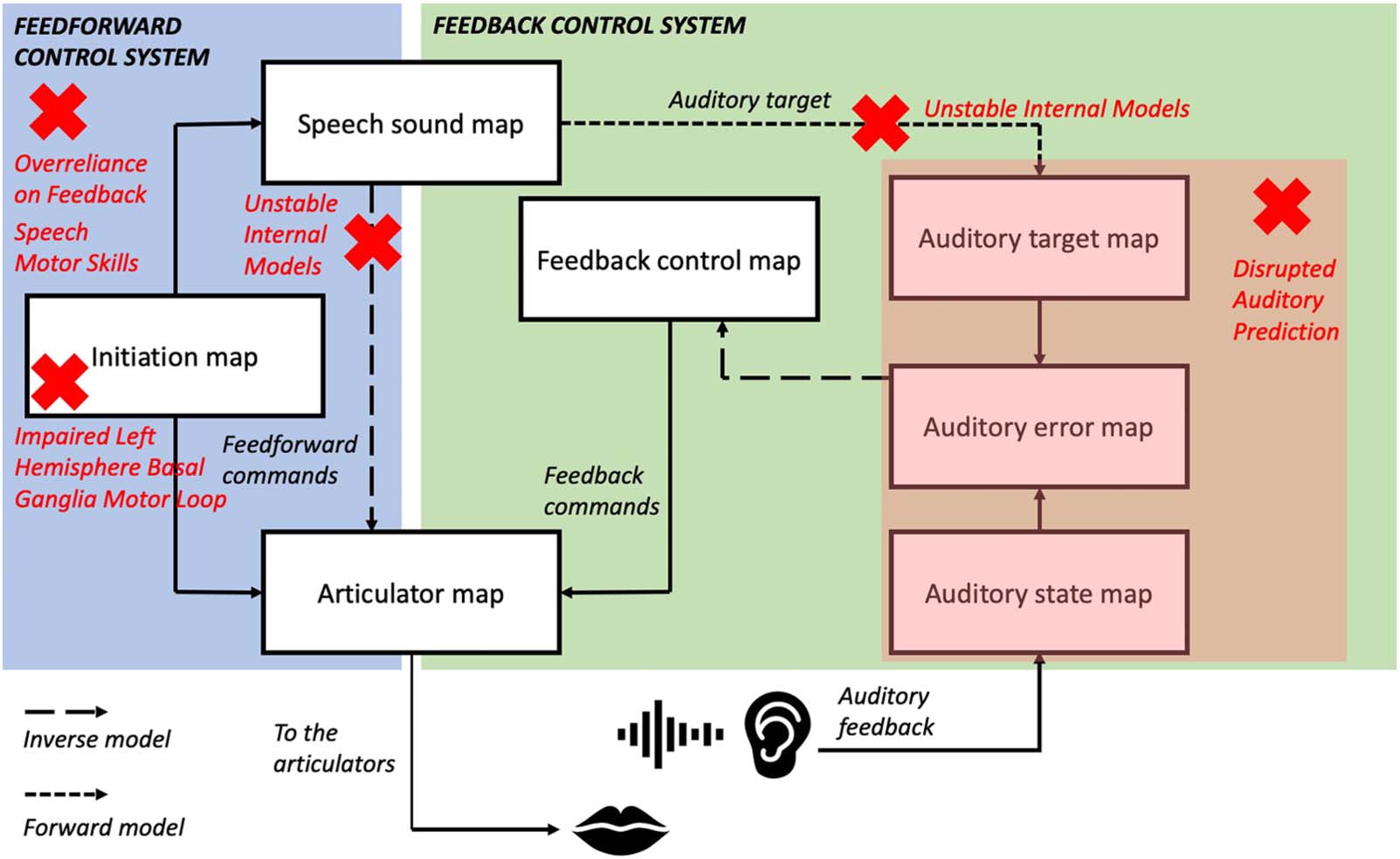
To aid comparison of these different sensorimotor accounts of stuttering, Cifra 4 illustrates the
major sites of disruption proposed by each account projected onto the DIVA model framework.
It can be seen that the different theories propose disruptions to a wide variety of areas within
sensorimotor control of speech. It should be kept in mind when viewing this figure that not all
of these theories use the DIVA model as their framework for speech motor control, and so map-
ping their sites of disruption onto this model is not always straightforward. En particular, el
Unstable Internal Models hypothesis does not assume the presence of inverse models in the
feedback control system, and so this connection in the DIVA model is not marked with a red
cross for this theory.
A major question that differentiates these theoretical perspectives concerns whether a change
in auditory feedback processing reflects a primary site of disruption in stuttering, or a secondary
downstream consequence of disruption to a different neural system. The SMS account and the
Disrupted Auditory Prediction account both consider dysfluencies to be the result of the im-
paired functioning of one of the two control subsystems in isolation; eso es, the faulty operation
of the feedforward or feedback control system directly causes stuttered speech. A diferencia de, otro
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4. Comparison of sensorimotor accounts of stuttering within the DIVA model framework. Red crosses denote main areas of disruption
proposed by different theories, within a simplified DIVA model framework. Note that the somatosensory feedback controller has been removed
due to the focus on auditory feedback in these theories. Adapted from Guenther (2016).
Neurobiology of Language
319
Stuttering and sensory feedback
accounts propose that it is the interactions between the feedback and feedforward control sys-
tems that result in disruptions to fluent speech. Por ejemplo, in the Overreliance on Feedback
hypothesis, even though impairments in feedforward control lead to increased articulatory er-
rors, these by themselves are not a major impediment to fluency if speech is controlled solely by
the feedforward control system; crucialmente, it is the operation of the feedback controller on these
speech errors that results in dysfluencies. Similarmente, in the Impaired Left Hemisphere Basal
Ganglia Motor Loop account, disruption to the readout of feedforward motor commands leads
to the detection of sensory errors in feedback that prevent the proper generation of initiation and
termination signals by the basal ganglia. In this way, while auditory feedback processing plays a
role in all of the reviewed theories of stuttering, the theories differ in whether feedback process-
ing is specifically impaired (es decir., the primary source of the problem), or simply a secondary con-
sequence of some other disruption.
There are several other divisions that can be made between these theories that flow from this
central distinction. The first concerns whether recruitment of feedback control in speech is seen
as compensatory or maladaptive. In the SMS and Unstable Internal Models accounts, slowing of
speech in order to enhance processing of sensory feedback is seen as a strategy employed to help
speech motor control. En cambio, although the Overreliance on Feedback hypothesis also pre-
dicts increased use of sensory feedback in speech, this is itself a cause of dysfluencies, due to the
accumulation of sensorimotor error. Feedback control of speech is similarly seen as maladaptive
in the accounts proposed by Guenther (2016) and Max and Daliri (2019).
A further related distinction that can be made between theories concerns whether changes in
feedback control are seen prior to stuttering onset, or whether these develop with experience of
stuttering. The Unstable Internal Models hypothesis, Overreliance on Feedback hypothesis, y
the Disrupted Auditory Prediction account all propose changes to auditory feedback control
processes or their relative weighting in speech motor control as part of the original pathology
in stuttering; these theories thus predict that differences in feedback processing should be seen
from the onset of the disorder. En cambio, the SMS and Impaired Left Hemisphere Basal Ganglia
Motor Loop accounts both propose that changes to feedback processing develop with experi-
ence of stuttering, suggesting a compensatory increase or decrease in reliance on sensory feed-
back over time respectively. Por lo tanto, these accounts would predict that changes in sensitivity
to auditory feedback should only be seen in individuals with significant experience of stuttering,
and not from stuttering onset.
Predictions for performance on altered feedback paradigms
The different assumptions made by these theories have implications for their predictions regard-
ing the compensation and adaptation responses of PWS during perturbations of auditory feed-
atrás. A summary of these predictions along with other key features of these accounts are
presented in Table 1. It can be seen that altered feedback paradigms thus have the potential
for testing the predictions of these theories, in order to start evaluating which provide the best
fit to observed behaviour in stuttering.
Impaired compensation for unexpected perturbations of auditory feedback would be predicted
by theories that assume disruption within the feedback control system. The Unstable Internal
Models account would predict that disruption to forward models should result in an impaired
ability to detect and correct for sensory errors online, resulting in weaker or more variable com-
pensation responses. The Disrupted Auditory Prediction account would similarly predict that a
failure to properly prime the auditory system for processing of upcoming speech feedback would
result in impaired detection of auditory errors, and thus weaker compensation responses. El
Neurobiology of Language
320
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
norte
mi
tu
r
oh
b
oh
oh
gramo
y
i
yo
avance
control
disrupted
Sí
Sí
Sí
Sí
Mesa 1.
Summary of theories and their predictions
Feedback
control
disrupted
No
Sí
No
Feedback control
compensatory or
maladaptive
Changes in
feedback control
developmental or
acquired after onset
Predicts impairment
in compensation for
unexpected feedback
perturbations
Predicts impairment
in adaptation to
sustained feedback
perturbations
Compensatory
Acquired
No (may be enhanced
Sí?
in adults)
Compensatory
Developmental
Sí
Maladaptive
Developmental
No (may be
enhanced)
Sí
Sí
No
Maladaptive
Acquired
Sí (weakened
in adults only)
Sí (possibly more
affected in adults)
No
Sí
Maladaptive
Developmental
Sí
Sí?
Teoría
Speech Motor Skills
cuenta (van Lieshout
et al., 1996a, 1996b)
Unstable Internal
Models hypothesis
(Max et al., 2004)
Overreliance on
Feedback hypothesis
(Civier et al., 2010;
Max et al., 2004)
Impaired Left
Hemisphere Basal
Ganglia Motor Loop
cuenta (Chang &
Guenther, 2020;
Guenther, 2016)
Disrupted Auditory
Prediction account
(máx. & Daliri, 2019)
oh
F
l
a
norte
gramo
tu
a
gramo
mi
3
2
1
S
t
tu
t
t
mi
r
i
norte
gramo
a
norte
d
s
mi
norte
s
oh
r
y
F
mi
mi
d
b
a
C
k
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Impaired Left Hemisphere Basal Ganglia account would also predict weaker compensation re-
sponses to unexpected feedback perturbations, but only in adults or individuals with significant
experience of stuttering who have learned to reduce reliance on sensory feedback as a compen-
satory strategy. En cambio, children who stutter should not show differences in compensation
responses relative to children who do not stutter. In contrast to these theories, the Overreliance
on Feedback hypothesis does not assume any disruption within the feedback system itself, y
so would predict intact or possibly even enhanced compensation responses in both children
and adults who stutter.
En cambio, impaired adaptation to sustained perturbations would be predicted by theories
that propose malfunctioning within the feedforward control system for speech. The Unstable
Internal Models account would predict disruption to the adaptation response, due to problems
in the updating of stored inverse models within the feedforward system. The Overreliance on
Feedback hypothesis and Impaired Left Hemisphere Basal Ganglia Motor Loop account both
propose disruption within the feedforward system, and so would predict abnormalities of the
adaptation response. In both theories, sin embargo, it is the readout of feedforward commands that
is affected, and not the updating of stored inverse models on which sensorimotor learning relies.
It is therefore unclear exactly how the adaptation response would be affected, and whether there
would be no learning at all. Sin embargo, Chang and Guenther’s (2020) suggestion of a decreased
reliance on sensory feedback over time would presumably also affect the adaptation response;
eso es, sensory errors would not be incorporated into stored inverse models, leaving feedforward
commands unchanged to result in no learning. This account would thus be compatible with a
difference in the adaptation response between children and adults who stutter.
The Disrupted Auditory Prediction account does not assume impairments in the feedforward
sistema, and the authors suggest that the PSAM effect may have greater functional relevance for
online corrections of speech gestures via the feedback control system (máx. & Daliri, 2019).
Sin embargo, it is difficult to see how offline updating of internal models within the feedforward
system could remain unaffected by disruption to the auditory system’s ability to detect sensory
errores. The DIVA model explicitly assumes that such updating of feedforward models underlying
the adaptation response relies on detection of errors by the feedback system (see dashed arrows
En figura 2). While there is evidence that the adaptation response can be selectively impaired
without disruption to compensation responses (p.ej., Parrell et al., 2017), to our knowledge there
is currently no evidence that a successful adaptation response can occur in the absence of an
intact compensation response. Disrupted auditory prediction would therefore likely result in an
impairment in the adaptation response.
The predictions made by the SMS hypothesis regarding differences in compensation and ad-
aptation responses in PWS are less clear, since this theory is not rooted in a sensorimotor control
framework as explicitly as the other theories. Sin embargo, on the premise that this view is consistent
with an impairment in automated feedforward speech motor control that triggers increased re-
liance on sensory feedback, we can assume this theory would predict weakened or more vari-
able adaptation responses but enhanced compensation responses in PWS.
It can therefore be seen that these theories make different predictions regarding patterns of
behaviour of PWS in feedback perturbation studies. In the last decade, several studies have be-
gun investigating compensation and adaptation responses in PWS. Reviewing this body of ev-
idence can thus start to provide insight into which of the theoretical accounts best explains the
pattern of behaviour of PWS in such experiments. The following section will therefore present
the evidence to date using altered feedback paradigms with PWS, highlighting where findings
support particular theories over others.
Neurobiology of Language
322
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Review of Evidence from Altered Feedback Experiments
Pitch shift studies
Early studies of compensation responses to pitch perturbations in PWS reported small effects,
often in underpowered samples. Bauer et al. (2007) reported a trend for PWS to show slower
compensation responses to unexpected pitch shifts than PWNS, although in a small sample of
just four PWS. Específicamente, when averaging the response across trials within-subjects, PWS
showed longer mean latencies and delayed response peak times. Loucks et al. (2012) reported
similar effects in a larger sample (norte = 14), with PWS showing significantly delayed average onset
latencies in their compensation responses. This study also reported weaker average compensa-
tion responses in PWS, although this was based on qualitative description of trends, and not
statistically significant differences. A more recent study by Sares et al. (2018) was able to dem-
onstrate a statistically significant reduction in the average response (area under the curve) a
pitch shifts in a sample of 19 PWS; sin embargo, further analyses were presented to challenge a
straightforward interpretation of this effect. They argued that the weaker averaged compensa-
tion response in PWS was attributable to (1) the decreased number of opposing trials (versus
following trials or trials with no compensation) shown in PWS, y (2) the greater variability in
the timing of their opposing responses. Fundamentalmente, when looking at only those trials on which an
opposing response was made, no significant group difference in the magnitude of compensa-
tion was found between PWS and PWNS.
These results suggest that the feedback control system of PWS is able to detect and appropri-
ately compensate for auditory errors online, but the timing of the implementation of such cor-
rective commands involves increased variability. This could mean that on some trials, hay
not sufficient time for the delayed response to reach its peak magnitude, resulting in an overall
reduced compensation magnitude when averaged across trials. In a recent fMRI study, Sares
et al. (2020) were not able to replicate the group difference in the average compensation re-
sponse when the paradigm was conducted in the scanner, despite using a subset of the same
participants from their previous study. They attributed this to aspects of the in-scanner environ-
ment such as the external noise; sin embargo, it could also simply highlight the unreliability of this
efecto.
Despite this lack of behavioural differences, Sares et al. (2020) did nevertheless find differ-
ences in underlying neural activation between PWS and control participants who did not stutter.
PWS were found to show deactivation of the middle temporal gyrus (MTG) and superior frontal
gyrus (SFG) in the right hemisphere for shifted versus unshifted trials; conversely, PWNS showed
increased activity in these areas for shifted versus unshifted trials. The authors proposed that
these two areas may form a processing circuit in which the MTG informs the SFG about auditory
comentario, which in turn engages a compensatory response. Además, independent compo-
nents analysis identified differences in the functional networks engaged in the two groups; mientras
controls demonstrated engagement of a network that integrated both auditory and motor com-
ponents, auditory and motor networks were dissociated in PWS during vocalisation. Esta sugerencia-
gests weaker communication or coordination between auditory and motor regions, which could
underlie the increased variability in timing of the compensation response.
Teóricamente, this increased variability in timing of the compensatory response perhaps best
fits the Unstable Internal Models hypothesis. Instability in forward and inverse models within the
feedback control system could mean that while in some cases appropriate corrective commands
can be generated, instability of modelling at other points results in a failure to initiate the com-
pensation response sufficiently quickly. Such instability in the formation of internal models
could be underwritten by the weakened communication between auditory and motor regions
Neurobiology of Language
323
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
reported by Sares et al. (2020). Similarmente, this would fit with the account proposed by Hickok
et al. (2011), which suggests that noisy/variable forward predictions result from disruption to
area Spt which coordinates the mapping between motor and sensory areas.
Formant perturbation studies
Unexpected perturbations. Studies of compensation responses to unexpected formant perturba-
tions have also reported some contradictory findings. A study by Cai et al. (2012) found reduced
compensation responses to unexpected perturbations of F1 in 21 PWS. In further analyses, fue
demonstrated that these reduced responses were not attributable to (1) increased variability in
responses across trials, (2) delayed response latencies, o (3) impaired auditory acuity for per-
ceiving differences in the F1 of vowels. Además, the proportion of compensating trials did
not differ between groups. This pattern of results thus differs from that reported by Sares et al.
(2018) with unexpected pitch shifts, in that weaker responses could not be attributed to differ-
ences in timing of the responses.
En cambio, a further study from this same group failed to find evidence of weaker compen-
sation to unexpected formant perturbations. Cai et al. (2014) investigated compensation to
unexpected F2 perturbations during production of the multisyllabic phrase “I owe you a
yo-yo.” In this task, PWS were found to demonstrate normal compensatory responses that
looked similar to those of controls. Although the authors described a trend for these responses
to have a slower onset in PWS, this was not manifest in any significant group differences. Este
discrepancy in findings between Cai et al. (2012) and Cai et al. (2014) could be explained by
multiple methodological differences between the studies, such as the nature of the perturbation
(a sudden step-like increase in F1 versus a gradual ramp up in F2) and the target utterance (single
words versus a multisyllabic utterance).
In a second experiment reported by Cai et al. (2014), sin embargo, a significant group difference
was found in response to a temporal perturbation. This involved random acceleration or decel-
eration of auditory feedback (específicamente, advancing or delaying the timing of the perceived F2
transition). Curiosamente, controls failed to demonstrate compensation for either type of pertur-
bation; instead they showed following responses on deceleration trials only, in which they
further lengthened syllable productions. PWS, por otro lado, lacked any significant timing
adjustments in response to the temporal perturbations. The authors interpreted this as evidence
that stuttering involves slowness in auditory-motor integration for temporal control. Sin embargo,
interpretation of this behaviour is made difficult given that the response of controls was not to
compensate for the temporary perturbation. En cambio, their so-called “following responses”
suggest that timing of speech motor control was disrupted by the deceleration of feedback.
En cambio, the speech of PWS appeared to be resistant to the disruptive effects of this auditory
feedback perturbation. This pattern echoes the effects of delayed auditory feedback on speech
motor control, resulting in dysfluent and disrupted speech in controls but improved fluency in
PWS.
The lack of compensation for these temporal perturbations in controls makes it difficult to
relate this behaviour to an error-based correction mechanism within the feedback control sys-
tema. En cambio, this perturbation is more likely to disrupt the proper generation of timing cues by
the basal ganglia as outlined in the GODIVA account (Bohland et al., 2010); eso es, since audi-
tory feedback is effectively delayed, the generation of a termination and initiation cue for the
next phoneme will be delayed also, resulting in a slowing of speech. The behaviour of PWS
in this study thus fits with Guenther’s (2016) proposal that a deficit in the use of auditory feed-
back to guide generation of internal timing cues leads to a reduced reliance on auditory
Neurobiology of Language
324
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
comentario. De este modo, the timing of speech motor control in PWS is less affected by temporal pertur-
bations of auditory feedback. In this way, this difference in response of PWS to this type of
auditory feedback perturbation may more likely reflect compensatory changes in sensitivity to
auditory feedback, rather than a central point of disruption.
It is interesting to note that when citing these two studies by Cai and colleagues, authors often
appear to present their findings as converging evidence for weaker compensation responses in
PWS. As is apparent from the current review, sin embargo, the findings from these two studies in
fact demonstrate quite different forms of disruption to sensorimotor responses to altered
feedback in PWS and, in the case of compensation to unexpected perturbations of F2, informe
conflicting results. It will therefore be important for future literature to ensure the findings from
these studies are accurately represented when making claims about compensation responses
in PWS.
Sustained perturbations. Preliminary suggestions that adaptation responses may be abnormal in
PWS were found in a serendipitous finding reported by Cai et al. (2012). Although their paradigm
involved unexpected perturbations, a cross-trial adaptation effect was found in control partici-
pants, in which an early “following” response was seen in perturbed trials that were immediately
preceded by a perturbed trial of the opposite direction (p.ej., a downward shift trial preceded by
an upward shift trial). This was interpreted as evidence that some offline updating of internal
models had occurred in response to that previous perturbation. Curiosamente, such cross-trial
adaptation was entirely absent in PWS, suggesting problems in the updating of stored sensori-
motor models.
Respectivamente, subsequent studies reported weaker adaptation responses in PWS in sustained
perturbation paradigms. Sengupta et al. (2016) reported reduced adaptation in response to a
sustained perturbation of F1 and F2 in a sample of eight PWS participants. Similar results were
found by Daliri et al. (2018), who reported significantly reduced adaptation in a group of 14
adults who stutter compared to controls. Curiosamente, this was not found for a group of children
who stutter, who showed significant adaptation to the same extent as control children. The au-
thors therefore argued that abnormalities in the speech adaptation response in adult PWS were
likely the result of a compensatory strategy learned over many years of stuttering, rather than the
result of a primary deficit that contributes to stuttering onset. Eso es, experience of stuttering
throughout childhood and into adulthood may lead to a reduced reliance on auditory feedback
for speech motor control.
Sin embargo, recent work by Kim et al. (2020) provides contradictory evidence to this previous
estudiar. Fundamentalmente, significantly weaker adaptation to a formant perturbation was seen in both
adults and children who stutter compared to non-stuttering aged matched controls. En efecto, en
este estudio, the difference in extent of adaptation between stuttering and non-stuttering partici-
pants was even greater in the child sample compared to the adult sample. This discrepancy in
findings from Daliri et al. (2018) may reflect the type of formant perturbation employed; mientras
Daliri et al. perturbed F1 and F2 in opposite directions so as to induce a shift in the phonemic
category of the vowel sound, Kim et al. employed a global formant perturbation in which all
formants were perturbed. This results in a change in the perception of the size of the vocal tract
that produced the vowel. Kim et al. highlighted how sensitivity to this type of perturbation of
auditory feedback would be highly relevant for adapting to the rapid maturational changes in
the biomechanics of the vocal apparatus that occur during childhood; a failure to update one’s
internal models to reflect these developmental changes in vocal anatomy would be problematic
for accurate sensorimotor control during speech. It should be highlighted also that while Daliri
et al.’s study included children aged 6 a 11 años, Kim et al. included children as young as
Neurobiology of Language
325
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
three years (ranging up to 9 años). En efecto, in this study the most severe impairments in adap-
tation were seen in the youngest children who stutter, with adaptation improving with age in this
grupo. They therefore concluded that impaired speech adaptation is not something that
develops with experience of stuttering, but is in fact present very early in development, close
to the onset of the disorder. This is consistent with the view that this disruption to the adaptation
response may be causally relevant for the disorder.
As well as comparing adults and children who stutter, this study by Kim et al. (2020) también
compared adaptation responses within-subjects in response to formant perturbations that were
either introduced gradually in a ramp-like fashion, or suddenly (es decir., the maximum perturbation
is employed in a single-step). In the group of adults who stutter, a greater reduction in adaptation
compared to controls was found in the sudden perturbation condition compared to the gradual
perturbation condition. En efecto, significant adaptation at the group level was seen in the adults
who stutter in response to a gradual upward (but not downward) perturbation (although this was
still significantly reduced in magnitude compared to PWNS). De este modo, a gradual ramping up of the
formant perturbation appeared to aide adaptation in adults who stutter. This was, sin embargo, no
the case in children who stutter, who failed to show significant adaptation for either the gradual
or sudden perturbation condition. The authors speculated that this difference in adults may
reflect differential impairment of cortical, basal ganglian, and cerebellar circuits, with some
evidence that these structures may be differentially recruited for these different types of per-
turbation (Criscimagna-Hemminger et al., 2010; Robertson & Miall, 1999; Venkatakrishnan
et al., 2011).
A study by Kim and Max (2020) aimed to further investigate the reasons for reduced adapta-
tion in PWS. En primer lugar, they replicated this finding in an F1 perturbation paradigm with a greater
number of perturbation trials (120 ensayos) than had been used in previous studies; adaptation in
PWS thus does not seem to benefit from extended exposure to altered feedback. En segundo lugar, ellos
investigated the potential contribution of an explicit component to speech adaptation in PWS
and PWNS. In visuomotor adaptation for reaching movements, participants are known to em-
ploy explicit aiming strategies in order to combat large sensory errors caused by sudden pertur-
bations, in addition to implicit updating of internal models (Vínculo & taylor, 2015). In their study,
Kim and Max (2020) found that neither group reported explicit awareness of intentions to change
their speech when repeatedly asked after each trial (with the exception of one PWNS partici-
pant). This accords with other evidence suggesting that speech adaptation to formant perturba-
tions indeed only involves an implicit component (Lametti et al., 2020; Munhall et al., 2009).
Finalmente, this study also looked at whether participants’ perceptual targets for the spoken words
were changed during the task, given evidence that speech motor learning can induce changes in
the perception of speech sounds (Lametti et al., 2014; Shiller et al., 2009). It is possible that re-
duced adaptation in PWS could be due to greater shifts in their perceptual targets in the direction
of the perturbation, reducing perceived sensory error and thus adaptation. Los participantes fueron
asked to select an acoustic stimulus that best represented each of the test words at repeated
time-points throughout the task; sin embargo, neither group demonstrated any change in the F1
of their selected targets after adaptation. En general, por lo tanto, these authors concluded that re-
duced adaptation in PWS reflects disruption to implicit sensorimotor learning that relies on
the updating of internal models.
In order to further understand the nature of the disruption to adaptation responses in PWS,
several studies have employed EEG measures in concert with speech motor learning para-
digms. Previous work with PWNS has reported changes in phase coherence in the theta-band
during speech adaptation, which were attributed to the formation of a new feedforward map
with learning (Sengupta & Nasir, 2015). Building on these findings, and alongside their
Neurobiology of Language
326
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
reporting of behavioural group differences, Sengupta et al. (2016) compared phase coherence
changes during speech adaptation between PWS and PWNS, and found widespread group
differences across various frequency bands. Específicamente, PWS showed abnormalities in alpha
coherence early in the perturbation phase (higher alpha-beta and alpha-gamma coherence
than controls), whereas abnormalities in theta coherence were seen late in the perturbation
phase (theta-beta coherence increased over adaptation in PWNS but not in PWS). Based on
the timing of these differences, alpha coherence abnormalities early in adaptation were sug-
gested to reflect abnormal processing of feedback error, while theta coherence abnormalities
late in adaptation were suggested to reflect abnormal learning of a new feedforward map.
They therefore concluded that their behavioural findings of weakened adaptation responses
in PWS (as discussed previously) may reflect aberrant communication within sensorimotor
networks that affects both the detection of sensory errors and the updating of internal feedfor-
ward models.
EEG measures were also related to adaptation responses in PWS by Daliri and Max (2018),
who investigated the potential relationship with PSAM (mira la sección, Disrupted auditory predic-
tion and feedback monitoring). As previously reviewed, the PSAM effect has been reported to be
absent in PWS (Daliri & máx., 2015a, 2015b); this raises the possibility that reduced adaptation
responses in PWS could be caused by a failure to appropriately prime the auditory system to
process sensory errors in auditory feedback (reflected in reduced PSAM). Although they were
able to replicate both reduced speech adaptation and reduced PSAM in a sample of 13 PWS,
surprisingly a negative correlation between the magnitude of these effects was found in the PWS
grupo, with no significant relationship at all in the control group. Eso es, PWS who showed
greater PSAM showed reduced speech adaptation. This is not consistent with a view in which
PSAM enhances sensitivity of the auditory system to sensory errors, and is difficult to interpret
given that both PSAM and adaptation were weaker in PWS. Por lo tanto, although these altered
responses at a group level in PWS both support the notion of disrupted sensorimotor integration,
how they may relate to one another remains unclear.
En general, findings of weaker adaptation responses in PWS are consistent with a number of
theories that propose disruption to feedforward control in stuttering (Chang & Guenther,
2020; Civier et al., 2010; Max et al., 2004), or to auditory prediction that would affect
updating of internal models in the feedforward system (máx. & Daliri, 2019). Evidencia de
Kim et al. (2020) demonstrates that this disruption can be seen early in childhood, near to the
onset of the disorder. They related this evidence to Max et al.’s (2004) Unstable Internal
Models account, arguing that a failure to learn accurate or stable internal models would
seriously hamper the speech sensorimotor system’s ability to flexibly adjust to maturational
changes in the vocal apparatus that occur throughout childhood. Their global formant pertur-
bation was specifically selected to mirror this type of maturational change by inducing a
change in perception of the size of the vocal tract (of course over a much shorter timescale
than in development itself ). Sin embargo, Daliri et al.’s (2018) finding of intact adaptation in
niños (but not adults) who stutter in response to a formant perturbation that affected the
identity of a vowel sound suggests that some aspects of auditory-motor learning for speech
may be relatively unaffected in childhood, instead becoming progressively worse with expe-
rience of stuttering into adulthood. This is perhaps more consistent with the Impaired Left
Hemisphere Basal Ganglia Motor Loop account (Chang & Guenther, 2020; Guenther,
2016). Although this proposes a core deficit in the left hemisphere feedforward system, también
acknowledges that the use of auditory feedback in guiding speech motor control may change
over time with experience of stuttering, as PWS learn to suppress auditory feedback as a com-
pensatory strategy. Why responding to these two types of formant perturbations and their
Neurobiology of Language
327
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
corresponding auditory feedback errors should be differentially affected in children who
stutter, sin embargo, cannot be readily explained by any current theories.
DISCUSIÓN
This review of evidence from altered feedback studies has found that there are differences in the
way in which PWS use auditory feedback to guide speech production. Compensation responses
to unexpected perturbations appear to be disrupted; sin embargo, this evidence is not always
consistent and may reflect issues with the timing of the response rather than a straightforward
weakening. This is not always adequately conveyed when these articles are cited in the literature;
future studies citing this work should ensure that it is interpreted correctly. Adaptation responses
to sustained perturbations appear to be consistently weaker across studies in adults who stutter,
as well as in children who stutter when certain formant perturbations are used. En general, allá-
delantero, PWS appear to be less effective in the use of feedback to guide their production of speech
sounds, across both unexpected and sustained perturbation paradigms.
Evaluating Theories of the Use of Feedback in Stuttering
The overall pattern of evidence speaks directly against the Overreliance on Feedback hypoth-
esis, despite this being a highly influential theory in the field. En cambio, this theory appears to fit
much better with the pattern of behaviour shown by Parkinson’s patients in altered feedback
experimentos. Específicamente, compensation for unexpected perturbations is enhanced (Chen
et al., 2013; Huang et al., 2016; Liu et al., 2012), whereas adaptation to sustained perturbations
is reduced (Abur et al., 2018; Mollaei et al., 2013). This clear dissociation between responses in
the two paradigms fits with the idea of a core disruption to feedforward control causing an over-
reliance on feedback control. Parkinson’s disease is a limb motor control disorder caused by
degeneration of dopaminergic neurons in the substantia nigra (pars compacta) of the basal gan-
glia. Curiosamente, a number of speech disturbances including stuttering-like dysfluencies are
often reported in patients (Juste et al., 2018). It is possible therefore that an overreliance on feed-
back control may contribute to stuttering-like behaviour in some cases.
Contrasting this evidence from Parkinson’s patients with the currently reviewed findings from
PWS highlights how the pattern is much more complex in developmental stuttering. Esta sugerencia-
gests that a simple account of impairment in one control system and overreliance on the other is
not sufficient in this case. This likely reflects the fact that stuttering is a developmental and not an
acquired disorder. Eso es, it is important to recognise that the impairments to speech motor con-
trol seen in developmental stuttering are not the result of damage to the mature system.
Asombrosamente, this point is apparently often overlooked. Por ejemplo, the DIVA model simulations
presented by Civier et al. (2010) involved inducing a shift in balance between feedback and
feedforward control after learning in the model was complete. Tal como, this does not provide
a viable model of a developmental disorder. It is vital that any theory of sensory feedback dis-
ruption in stuttering considers the importance of dynamic interactions between different parts of
the speech motor control system as it develops.
Evidence from studies of sustained feedback perturbations demonstrate that the adaptation
response is disrupted (weaker) in adults who stutter. Además, this disruption can be seen
early in childhood and thus potentially contributes to the cause of the disorder (Kim y cols.,
2020). Sin embargo, there is also evidence that the severity of this impairment may change across
desarrollo. Kim et al. (2020) reported an improvement in the adaptation response to a global
formant perturbation with increasing age in children who stutter, with the worst impairment seen
in an early age group. En cambio, the evidence from Daliri et al. (2018) suggests that adaptation
Neurobiology of Language
328
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
to a formant perturbation that affects the identity of a vowel gets worse over time in a person who
stutters, remaining intact in childhood but becoming impaired by adulthood. It is difficult to
know how to reconcile these two findings. Kim et al. (2020, pag. 11) argued that the Unstable
Internal Models hypothesis does not assume that children who stutter would not be able to
“correct their productions when auditory feedback from previous trials indicates that they
produced the words with a completely wrong sound.” However, it is hard to imagine how
children who stutter would be able to show any kind of speech adaptation if their core deficit
was in the feedforward system and internal models.
En general, the evidence reviewed here reveals abnormalities of both compensation and
adaptation responses in PWS. This suggests that the disruption to sensorimotor learning
processes in stuttering has repercussions for the operation of both the feedback and the feed-
forward control systems. This is consistent with the Unstable Internal Models hypothesis (máx.
et al., 2004), in which there is a failure to appropriately update both inverse models in the feed-
forward system and forward models in the feedback system. This latter disruption to forward
models that facilitate prediction of the sensory consequences of a speech motor gesture would
also be consistent with the Disrupted Auditory Prediction account (máx. & Daliri, 2019). A failure
to accurately predict the sensory consequences of speech movements would result in erroneous
sensory error signals, affecting both online compensation responses to unexpected feedback
perturbations, and consequently the appropriate updating of inverse models in the feedforward
system when perturbations are sustained. More work is needed, sin embargo, to better understand
how these disruptions may change over time, and how this may manifest differently in response
to different types of errors in auditory feedback. En particular, these accounts will need to rec-
oncile the finding of intact adaptation in children who stutter in response to perturbations that
affect perceived vowel identity (Daliri et al., 2018). This finding itself would benefit from repli-
cation in a sample including younger-aged children, in order to determine whether such an ad-
aptation response remains intact at an age closer to typical onset of stuttering.
Role of Development and Potential Heterogeneity
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
This discussion highlights the difficulty of interpreting disrupted responses to altered feedback
measured in the mature system. These responses reflect the end point of an abnormal devel-
opmental trajectory, and as such represent the combined result of both primary disruptions
and their secondary consequences, including compensatory strategies. In this way, it can be
difficult to tease apart these different aspects when relying on evidence from a single point in
tiempo. Longitudinal studies of responses to altered feedback in children who stutter, incluido
comparisons between those who go on to recover and those whose stuttering persists, will
likely provide better insight into how dynamic interactions between different subparts of the
system may be critical for development of fluent speech.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
It is also important to consider that the causes of speech motor control problems in develop-
mental stuttering may be heterogeneous across the population. Eso es, stuttering may emerge
when there is a disruption to the balance across subsystems, but the exact pattern of this disrup-
tion may vary across individuals. This could explain some of the inconsistencies in the reviewed
evidencia, such as the failure of all studies to find overall weakened responses to unexpected
perturbations of auditory feedback in adult PWS.
Comparing Auditory and Somatosensory Feedback Control
It will be important for future theorising on the role of sensory feedback in stuttering to integrate
evidence across different modalities. The current review focused on theories and evidence
Neurobiology of Language
329
Stuttering and sensory feedback
considering the role of auditory feedback in stuttered speech, because this has been most exten-
sively studied in altered feedback and sensorimotor learning paradigms. Sin embargo, there is a sep-
arate literature considering the role played by somatosensory feedback in stuttering (Archibald &
De Nil, 1999; Loucks & De Nil, 2006). PWS have been found to show reduced accuracy and in-
creased variability in jaw movements, exacerbated by an absence of visual feedback and increased
time-pressure (Loucks & De Nil, 2006). There is also some evidence that their motor control system
is less able to take jaw size into account during movement planning (Daliri et al., 2013).
Sin embargo, to our knowledge there are currently no studies of sensorimotor learning with
somatosensory feedback perturbations in PWS. Where manipulations of somatosensory feed-
back have been used with PWS, these have typically involved alterations such as tendon vibra-
ción, equivalent to masking of auditory feedback. In order to consider disruptions to the use of
auditory and somatosensory feedback within a common framework, it would be of interest to
investigate the ability of PWS to compensate for jaw perturbations in paradigms comparable to
those reviewed here in the auditory domain (p.ej., Tremblay et al., 2003). Eso es, are the com-
pensation and adaptation responses shown by PWS during somatosensory perturbations similar
to those seen in the auditory domain?
This would be of interest to questions concerning the development of stuttering, given that
somatosensory feedback is thought to play a less important role in early typical speech devel-
opment (Tourville & Guenther, 2011; Trudeau-Fisette et al., 2019). En efecto, it has been shown
that individuals can vary in their relative weighting of these two sources of feedback during
speech motor control (Lametti et al., 2012). It would be of interest to consider whether the same
is true in PWS, or whether as a group they are more likely to use one particular form of feedback
control over the other. In the DIVA model, distinct auditory and somatosensory feedback con-
trollers are proposed, which would allow for their selective disruption; alternatively, it is possible
that a more general disruption (p.ej., to internal modelling) may affect the operation of both feed-
back controllers in a similar way. More work is therefore needed to compare auditory-based and
somatosensory-based feedback control in PWS.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
Summary and Conclusions
This review has sought to provide insights into the role of sensory feedback in developmental
stuttering, by bringing together evidence on responses of PWS in altered auditory feedback par-
adigms. This evidence mostly favours a theoretical perspective that proposes a disruption to the
updating and use of internal models in speech motor control in stuttering, affecting both feed-
back and feedforward control of speech. En general, this field of research would benefit from an
improved focus on the developmental aspects of the disorder, and consideration of interactions
between feedback processing across auditory and somatosensory modalities.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INFORMACIÓN DE FINANCIACIÓN
Carolyn McGettigan, Leverhulme Trust (https://dx.doi.org/10.13039/501100000275), Award
ID: RL-2016-013.
CONTRIBUCIONES DE AUTOR
Abigail R. Bradshaw: Conceptualización: Lead; Investigación: Lead; Escritura – borrador original: Lead;
Escritura – revisión & edición: Lead. Daniel Lametti: Conceptualización: Equal; Supervisión: Equal;
Escritura – revisión & edición: Equal. Carolyn McGettigan: Conceptualización: Equal; Fondos
adquisición: Lead; Supervisión: Equal; Escritura – revisión & edición: Equal.
Neurobiology of Language
330
Stuttering and sensory feedback
REFERENCIAS
Abur, D., Lester-Smith, R. A., Daliri, A., Lupiani, A. A., Guenther, F. h.,
& Stepp, C. mi. (2018). Sensorimotor adaptation of voice fundamental
frequency in Parkinson’s disease. MÁS UNO, 13(1). DOI: https://doi
.org/10.1371/journal.pone.0191839, PMID: 29373589, PMCID:
PMC5786318
Alm, PAG. A. (2004). Stuttering and the basal ganglia circuits: A critical
review of possible relations. Journal of Communication Disorders,
37(4), 325–369. DOI: https://doi.org/10.1016/j.jcomdis.2004.03
.001, PMID: 15159193
Archibald, l., & De Nil, l. F. (1999). The relationship between stut-
tering severity and kinesthetic acuity for jaw movements in adults
who stutter. Journal of Fluency Disorders, 24(1), 25–42. DOI:
https://doi.org/10.1016/S0094-730X(98)00023-0
Bauer, j. J., Hubbard Seery, C., LaBonte, r., & Ruhnke, l. (2007).
Voice F0 responses elicited by perturbations in pitch of auditory
feedback in individuals that stutter and controls. The Journal of
the Acoustical Society of America, 121(5), 3201. DOI: https://doi
.org/10.1121/1.4782465
Bloodstein, o., & Ratner, norte. (2008). A handbook on stuttering
(6th ed.). Clifton Park.
Bohland, j. w., Bullock, D., & Guenther, F. h. (2010). Neural rep-
resentations and mechanisms for the performance of simple
speech sequences. Revista de neurociencia cognitiva, 22(7),
1504–1529. DOI: https://doi.org/10.1162/jocn.2009.21306,
PMID: 19583476, PMCID: PMC2937837
Vínculo, k. METRO., & taylor, j. A. (2015). Flexible explicit but rigid
Diario
learning in a visuomotor adaptation task.
implicit
of Neurophysiology, 113(10), 3836–3849. DOI: https://doi.org
/1 0.1 15 2/jn.0 00 09 .2 01 5, PM I D: 2 58 55 69 0, PM CI D:
PMC4473515
Marrón, S., Ingham, R. J., Ingham, j. C., Laird, A. r., & Fox, PAG. t.
(2005). Stuttered and fluent speech production: An ALE meta-
analysis of functional neuroimaging studies. Human Brain
Cartografía, 25(1). DOI: https://doi.org/10.1002/ hbm.20140,
PMID: 15846815, PMCID: PMC6871755
Budde, k. S., barrón, D. S., & Fox, PAG. t. (2014). Stuttering, induced
fluidez, and natural fluency: A hierarchical series of activation
likelihood estimation meta-analyses. Brain and Language, 139,
99–107. DOI: https://doi.org/10.1016/j.bandl.2014.10.002,
PMID: 25463820, PMCID: PMC4405378
Burnett, t. A., Freedland, METRO. B., larson, C. r., & Hain, t. C. (1998).
Voice F0 responses to manipulations in pitch feedback. Diario
of the Acoustical Society of America, 103(6), 3153–3161. DOI:
https://doi.org/10.1121/1.423073, PMID: 9637026
Cai, S., Beal, D. S., Ghosh, S. S., Guenther, F. h., & Perkell, j. S.
(2014). Impaired timing adjustments in response to time-varying
auditory perturbation during connected speech production in
persons who stutter. Brain and Language, 129(1), 24–29. DOI:
https://doi.org/10.1016/j.bandl.2014.01.002, PMID: 24486601,
PMCID: PMC3947674
Cai, S., Beal, D. S., Ghosh, S. S., Tiede, METRO. K., Guenther, F. h., &
Perkell, j. S. (2012). Weak responses to auditory feedback per-
turbation during articulation in persons who stutter: Evidencia
for abnormal auditory-motor transformation. MÁS UNO, 7(7).
DOI: https://doi.org/10.1371/journal.pone.0041830, PMID:
22911857, PMCID: PMC3402433
Chang, S.-E., Garnett, mi. o., Etchell, A., & Perro chino, h. METRO. (2019).
Functional and neuroanatomical bases of developmental stuttering:
Current insights. neurocientífico, 25(6), 566–582. DOI: https://doi
.org/10.1177/1073858418803594, PMID: 30264661, PMCID:
PMC6486457
Chang, S.-E., & Guenther, F. h. (2020). Involvement of the cortico-
basal ganglia-thalamocortical loop in developmental stuttering.
Fronteras en psicología, 10, 3088. DOI: https://doi.org/10.3389
/fpsyg.2019.03088, PMID: 32047456, PMCID: PMC6997432
Chang, S.-E., Ohde, R. NORTE., & Conture, mi. GRAMO. (2002). Coarticulation
and formant transition rate in young children who stutter. Diario
of Speech, Idioma, and Hearing Research, 45(4), 676–688.
DOI: https://doi.org/10.1044/1092-4388(2002/054)
Chen, X., Zhu, X., Wang, mi. P., Chen, l., li, w., Chen, Z., & Liu, h.
(2013). Sensorimotor control of vocal pitch production in Parkinson’s
enfermedad. Brain Research, 1527, 99–107. DOI: https://doi.org/10
.1016/j.brainres.2013.06.030, PMID: 23820424
Civier, o., Bullock, D., máx., l., & Guenther, F. h. (2013).
Computational modeling of stuttering caused by impairments
in a basal ganglia thalamo-cortical circuit involved in syllable se-
lection and initiation. Brain and Language, 126(3), 263–278.
DOI: https://doi.org/10.1016/j.bandl.2013.05.016, PMID:
23872286, PMCID: PMC3775364
Civier, o., Tasko, S. METRO., & Guenther, F. h. (2010). Overreliance on
auditory feedback may lead to sound/syllable repetitions:
Simulations of stuttering and fluency-inducing conditions with
a neural model of speech production. Journal of Fluency
Disorders, 35(3), 246–279. DOI: https://doi.org/10.1016/j.jfludis
.2010.05.002, PMID: 20831971, PMCID: PMC2939043
Cowie, r., & Douglas-Cowie, mi. (1992). Postlingually acquired deaf-
ness: Speech deterioration and the wider consequences. Moutón
de Gruyter. DOI: https://doi.org/10.1515/9783110869125
Craig, A., Hancock, K., Tran, y., Craig, METRO., & Peters, k. (2002).
Epidemiology of stuttering in the community across the entire life
span. Journal of Speech, Idioma, and Hearing Research, 45(6),
1097–1105. DOI: https://doi.org/10.1044/1092-4388(2002/088)
Criscimagna-Hemminger, S. MI., Bastian, A. J., & Shadmehr, R.
(2010). Size of error affects cerebellar contributions to motor
aprendiendo. Revista de neurofisiología, 103(4), 2275–2284. DOI:
https://doi.org/10.1152/jn.00822.2009, PMID: 20164398,
PMCID: PMC2853280
Daliri, A., & máx., l. (2015a). Electrophysiological evidence for a
general auditory prediction deficit in adults who stutter. Cerebro
and Language, 150, 37–44. DOI: https://doi.org/10.1016/j
.bandl.2015.08.008, PMID: 26335995, PMCID: PMC4663101
Daliri, A., & máx., l. (2015b). Modulation of auditory processing
during speech movement planning is limited in adults who stutter.
Brain and Language, 143, 59–68. DOI: https://doi.org/10.1016/j
.bandl.2015.03.002, PMID: 25796060, PMCID: PMC4380808
Daliri, A., & máx., l. (2018). Stuttering adults’ lack of pre-speech
auditory modulation normalizes when speaking with delayed au-
ditory feedback. Corteza, 99, 55–68. DOI: https://doi.org/10.1016
/j.cortex.2017.10.019, PMID: 29169049, PMCID: PMC5801108
Daliri, A., Prokopenko, R. A., & máx., l. (2013). Afferent and effer-
ent aspects of mandibular sensorimotor control in adults who
stutter. Journal of Speech, Idioma, and Hearing Research, 56(6),
1774–1788. DOI: https://doi.org/10.1044/1092-4388(2013/12
-0134), PMID: 23816664, PMCID: PMC3795963
Daliri, A., Wieland, mi. A., Cai, S., Guenther, F. h., & Chang, S.-E.
(2018). Auditory-motor adaptation is reduced in adults who
stutter but not in children who stutter. Developmental Science,
21(2), e12521. DOI: https://doi.org/10.1111/desc.12521, PMID:
28256029, PMCID: PMC5581739
Foundas, A. l., Mock, j. r., Corey, D. METRO., Golob, mi. J., & Conture, mi. GRAMO.
(2013). The SpeechEasy device in stuttering and nonstuttering
adultos: Fluency effects while speaking and reading. Brain and
Neurobiology of Language
331
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Idioma, 126(2), 141–150. DOI: https://doi.org/10.1016/ J
.BANDL.2013.04.004, PMID: 23712191
Giraud, A.-L., Neumann, K., Bachoud-Levi, A.-C., von Gudenberg,
A. w., Euler, h. A., Lanfermann, h., & Preibisch, C. (2008).
Severity of dysfluency correlates with basal ganglia activity in
persistent developmental stuttering. Brain and Language, 104(2),
190–199. DOI: https://doi.org/10.1016/j.bandl.2007.04.005,
PMID: 17531310
Guenther, F. h. (1995). Speech sound acquisition, coarticulation,
and rate effects in a neural-network model of speech production.
Revisión psicológica, 102(3), 594–621. DOI: https://doi.org/10
.1037/0033-295X.102.3.594, PMID: 7624456
Guenther, F. h. (2016). Neural control of speech. La prensa del MIT.
DOI: https://doi.org/10.7551/mitpress/10471.001.0001
Guenther, F. h., Ghosh, S. S., & Tourville, j. A. (2006). Neural
modeling and imaging of the cortical interactions underlying syl-
lable production. Brain and Language, 96(3), 280–301. DOI:
https://doi.org/10.1016/j.bandl.2005.06.001, PMID: 16040108,
PMCID: PMC1473986
Guenther, F. h., Hampson, METRO., & Johnson, D. (1998). A theoretical
investigation of reference frames for the planning of speech
movimientos. Revisión psicológica, 105(4), 611–633. DOI:
https://doi.org/10.1037/0033-295X.105.4.611-633, PMID:
9830375
Hickok, GRAMO., Houde, J., & Rong, F. (2011). Sensorimotor integration
in speech processing: Computational basis and neural organiza-
ción. Neurona, 69(3), 407–422. DOI: https://doi.org/10.1016/j
.neuron.2011.01.019, PMID: 21315253, PMCID: PMC3057382
Houde, j. F., & Jordán, METRO. I. (1998). Sensorimotor adaptation in
speech production. Ciencia, 279(5354), 1213–1216. DOI:
https://doi.org/10.1126/science.279.5354.1213, PMID: 9469813
Houde, j. F., & Nagarajan, S. S. (2011). Speech production as state
feedback control. Frontiers in Human Neuroscience, 5(Octubre).
DOI: https://doi.org/10.3389/fnhum.2011.00082, PMID:
22046152, PMCID: PMC3200525
Huang, X., Chen, X., yan, NORTE., jones, j. A., Wang, mi. P., Chen, l.,
guo, Z., li, w., Liu, PAG., & Liu, h. (2016). The impact of
Parkinson’s disease on the cortical mechanisms that support
auditory-motor integration for voice control. Human Brain
Cartografía, 37(12), 4248–4261. DOI: https://doi.org/10.1002
/hbm.23306, PMID: 27400999, PMCID: PMC6867337
Juste, F. S., Sassi, F. C., Costa, j. B., & de Andrade, C. R. (2018).
Frequency of speech disruptions in Parkinson’s disease and de-
velopmental stuttering: A comparison among speech tasks. PLOS
ONE, 13(6). DOI: https://doi.org/10.1371/journal.pone.0199054,
PMID: 29912919, PMCID: PMC6005540
Kalinowski, J., Armson, J., Stuart, A., & Gracco, V. l. (1993). Effects
of alterations in auditory feedback and speech rate on stuttering
frequency. Language and Speech, 36(1), 1-dieciséis. DOI: https://doi
.org/10.1177/002383099303600101, PMID: 8345771
kim, k. S., Daliri, A., Flanagan, j. r., & máx., l. (2020). Dissociated
development of speech and limb sensorimotor learning in stutter-
En g: Speech auditory-motor learning is impaired in both children
and adults who stutter. Neurociencia, 451, 1–21. DOI: https://
d o i . o r g / 1 0 . 1 0 1 6 / j . n e u r o s c i e n c e . 2 0 2 0 . 1 0 . 0 1 4 , P M I D :
33091464, PMCID: PMC7704609
kim, k. S., & máx., l. (2020). Speech auditory-motor adaptation
lacks an explicit component: Reduced adaptation in adults
who stutter reflects limitations in implicit sensorimotor learning.
bioRxiv. DOI: https://doi.org/10.1101/2020.09.06.284638
Lametti, D. r., Nasir, S. METRO., & Ostry, D. j. (2012). Sensory preference in
speech production revealed by simultaneous alteration of auditory
and somatosensory feedback. Revista de neurociencia, 32(27),
9351–9358. DOI: https://doi.org/10.1523/JNEUROSCI.0404-12
.2012, PMID: 22764242, PMCID: PMC3404292
Lametti, D. r., Quek, METRO. Y. METRO., Prescott, C. B., Bretaña, J.-S., &
watkins, k. mi. (2020). The perils of learning to move while
speaking: One-sided interference between speech and visuo-
motor adaptation. Boletín psiconómico & Revisar, 27(3),
544–552. DOI: https://doi.org/10.3758/s13423-020-01725-8,
PMID: 32212105
Lametti, D. r., Rochet-Capellan, A., Neufeld, MI., Shiller, D. METRO., &
Ostry, D. j. (2014). Plasticity in the human speech motor system
drives changes in speech perception. Revista de neurociencia,
34(31), 10339–10346. DOI: https://doi.org/10.1523/JNEUROSCI
.0108-14.2014, PMID: 25080594, PMCID: PMC4115140
Lametti, D. r., Herrero, h. J., Freidin, PAG. F., & watkins, k. mi. (2018).
Cortico-cerebellar networks drive sensorimotor learning in speech.
Revista de neurociencia cognitiva, 30(4), 540–551. DOI: https://doi
.org/10.1162/jocn_a_01216, PMID: 29211651
Lincoln, METRO., Packman, A., & Onslow, METRO. (2006). Altered auditory
feedback and the treatment of stuttering: A review. Diario de
Fluency Disorders, 31(2), 71–89. DOI: https://doi.org/10.1016/j
.jfludis.2006.04.001, PMID: 16750562
Liu, h., Wang, mi. P., Metman, l. v., & larson, C. R. (2012). Vocal
responses to perturbations in voice auditory feedback in individ-
uals with Parkinson’s disease. MÁS UNO, 7(3). DOI: https://doi
.org/10.1371/journal.pone.0033629, PMID: 22448258, PMCID:
PMC3308986
Loucks, t. METRO. J., Chon, h., & Han, W.. (2012). Audiovocal integra-
tion in adults who stutter. International Journal of Language &
Communication Disorders, 47(4), 451–456. DOI: https://doi.org
/10.1111/j.1460-6984.2011.00111.x, PMID: 22788230
Loucks, t. METRO. J., & De Nil, l. F. (2006). Oral kinesthetic deficit in
adults who stutter: A target-accuracy study. Journal of Motor
Comportamiento, 38(3), 238–246. DOI: https://doi.org/10.3200/JMBR
.38.3.238-247, PMID: 16709563
máx., l., & Daliri, A. (2019). Limited pre-speech auditory modula-
tion in individuals who stutter: Data and hypotheses. Diario de
Speech Language and Hearing Research, 62(8S), 3071–3084.
DOI: https://doi.org/10.1044/2019_JSLHR-S-CSMC7-18-0358,
PMID: 31465711, PMCID: PMC6813031
máx., l., Guenther, F. h., Gracco, V. l., Ghosh, S. S., & Wallace,
METRO. mi. (2004). Unstable or insufficiently activated internal models
and feedback-biased motor control as sources of dysfluency: A
theoretical model of stuttering. Contemporary Issues in
Communication Science and Disorders, 31(Primavera), 105–122.
DOI: https://doi.org/10.1044/cicsd_31_S_105
Mollaei, F., Shiller, D. METRO., & Gracco, V. l. (2013). Sensorimotor
adaptation of speech in Parkinson’s disease. Movement
Disorders, 28(12), 1668–1674. DOI: https://doi.org/10.1002
/mds.25588, PMID: 23861349, PMCID: PMC3812368
Munhall, k. GRAMO., macdonald, mi. NORTE., Byrne, S. K., & Johnsrude, I.
(2009). Talkers alter vowel production in response to real-time
formant perturbation even when instructed not to compensate.
The Journal of the Acoustical Society of America, 125(1), 384–390.
DOI: https://doi.org/10.1121/1.3035829, PMID: 19173425, PMCID:
PMC2658635
Namasivayam, A. K., & van Lieshout, PAG. (2008). Investigating
speech motor practice and learning in people who stutter.
Journal of Fluency Disorders, 33(1), 32–51. DOI: https://doi.org
/10.1016/j.jfludis.2007.11.005, PMID: 18280868
Onslow, METRO., & Ingham, R. j. (1987). Speech quality measurement
and the management of stuttering. Journal of Speech and Hearing
Disorders, 52(1), 2–17. DOI: https://doi.org/10.1044/jshd.5201
.02, PMID: 3543496
Neurobiology of Language
332
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Parrell, B., Agnew, Z., Nagarajan, S., Houde, J., & Ivry, R. B. (2017).
Impaired feedforward control and enhanced feedback control of
speech in patients with cerebellar degeneration. Diario de
Neurociencia, 37(38), 9249–9258. DOI: https://doi.org/10.1523
/ JNEUROSCI.3363-16.2017, PMID: 28842410, PMCID:
PMC5607467
Parrell, B., & Houde, j. (2019). Modeling the role of sensory feed-
back in speech motor control and learning. Journal of Speech
Language and Hearing Research, 62(8S), 2963–2985. DOI:
https://doi.org/10.1044/2019_JSLHR-S-CSMC7-18-0127, PMID:
31465712, PMCID: PMC6813034
Parrell, B., Lammert, A. C., Ciccarelli, GRAMO., & Quatieri, t. F. (2019).
Current models of speech motor control: A control-theoretic
overview of architectures and properties. Journal of the
Acoustical Society of America, 145(3), 1456–1481. DOI:
https://doi.org/10.1121/1.5092807, PMID: 31067944
Parrell, B., Ramanarayanan, v., Nagarajan, S., & Houde, j. (2019).
The FACTS model of speech motor control: Fusing state estima-
tion and task-based control. PLOS Computational Biology, 15(9).
DOI: https://doi.org/10.1371/journal.pcbi.1007321, PMID:
31479444, PMCID: PMC6743785
Purcell, D. w., & Munhall, k. GRAMO. (2006). Adaptive control of vowel
formant frequency: Evidence from real-time formant manipula-
ción. The Journal of the Acoustical Society of America. DOI:
https://doi.org/10.1121/1.2217714, PMID: 16938984
Ringel, R. l., & Steer, METRO. D. (1963). Some effects of tactile and au-
ditory alterations on speech output. Journal of Speech and
Hearing Research, 6(4), 369–378. DOI: https://doi.org/10.1044
/jshr.0604.369, PMID: 14071874
Robb, METRO., & Blomgren, METRO. (1997). Analysis of F2 transitions in the
speech of stutterers and nonstutterers. Journal of Fluency Disorders,
22(1), 1-dieciséis. DOI: https://doi.org/10.1016/S0094-730X(96)00016-2
Robertson, mi. METRO., & Miall, R. C. (1999). Visuomotor adaptation during
inactivation of the dentate nucleus. NeuroReport, 10(5), 1029–1034.
DOI: https://doi.org/10.1097/00001756-199904060-00025, PMID:
10321480
Saltzman, mi. l., & Kelso, j. A. S. (1987). Skilled actions: A task-
dynamic approach. Revisión psicológica, 94(1), 84–106. DOI:
https://doi.org/10.1037/0033-295X.94.1.84, PMID: 3823306
Saltzman, mi. l., & Munhall, k. GRAMO. (1989). A dynamical approach to
gestural patterning in speech production. Ecological Psychology,
1(4), 333–382. DOI: https://doi.org/10.1207/s15326969eco0104_2
Sares, A. GRAMO., Deroche, METRO. l. D., Ohashi, h., Shiller, D. METRO., &
Gracco, V. l. (2020). Neural correlates of vocal pitch compensa-
tion in individuals who stutter. Frontiers in Human Neuroscience,
14, 18. DOI: https://doi.org/10.3389/fnhum.2020.00018, PMID:
32161525, PMCID: PMC7053555
Sares, A. GRAMO., Deroche, METRO. l. D., Shiller, D. METRO., & Gracco, V. l.
(2018). Timing variability of sensorimotor integration during
vocalization in individuals who stutter. Informes Científicos, 8(1),
16340. DOI: https://doi.org/10.1038/s41598-018-34517-1,
PMID: 30397215, PMCID: PMC6218511
Scott, C. METRO., & Ringel, R. l. (1971). Articulation without oral sen-
sory control. Journal of Speech and Hearing Research, 14(4),
804–818. DOI: https://doi.org/10.1044/jshr.1404.804, PMID:
5163911
Sengupta, r., & Nasir, S. METRO. (2015). Redistribution of neural phase
coherence reflects establishment of feedforward map in speech mo-
tor adaptation. Revista de neurofisiología, 113(7), 2471–2479.
DOI: https://doi.org/10.1152/jn.00731.2014, PMID: 25632078,
PMCID: PMC4416577
Sengupta, r., Shah, S., Sangre, K., Loucks, T., & Nasir, S. METRO. (2016).
Anomaly in neural phase coherence accompanies reduced
sensorimotor integration in adults who stutter. Neuropsicología,
93(A), 242–250. DOI: https://doi.org/10.1016/j.neuropsychologia
.2016.11.004, PMID: 27833009
Shiller, D. METRO., Sato, METRO., Gracco, V. l., & Baum, S. R. (2009).
Perceptual recalibration of speech sounds following speech
motor learning. The Journal of the Acoustical Society of
America, 125(2), 1103–1113. DOI: https://doi.org/10.1121/1
.3058638, PMID: 19206885
Herrero, C. R. (1975). Residual hearing and speech production in
deaf children. Journal of Speech and Hearing Research, 18(4),
DOI: https://doi.org/10.1044/jshr.1804.795, PMID: 1207108
Smits-Bandstra, S., & De Nil, l. F. (2007). Sequence skill learning in
persons who stutter: Implications for cortico-striato-thalamo-cortical
dysfunction. Journal of Fluency Disorders, 32(4), 251–278. DOI:
https://doi.org/10.1016/j.jfludis.2007.06.001, PMID: 17963936
Smits-Bandstra, S., De Nil, l. F., & Saint-Cyr, j. A. (2006). Discurso
and nonspeech sequence skill learning in adults who stutter.
Journal of Fluency Disorders, 31(2), 116–136. DOI: https://doi
.org/10.1016/j.jfludis.2006.04.003, PMID: 16735062
Svirsky, METRO. A., Teoh, S. w., & Neuburger, h. (2004). Desarrollo
of language and speech perception in congenitally, profoundly
deaf children as a function of age at cochlear implantation.
Audiology and Neuro-Otology, 9(4), 224–233. DOI: https://doi
.org/10.1159/000078392, PMID: 15205550
Tourville, j. A., & Guenther, F. h. (2011). The DIVA model: A neural
theory of speech acquisition and production. Language and
Cognitive Processes, 26(7), 952–981. DOI: https://doi.org/10.1080
/01690960903498424, PMID: 23667281, PMCID: PMC3650855
Tourville, j. A., Reilly, k. J., & Guenther, F. h. (2008). Neural mecha-
nisms underlying auditory feedback control of speech. NeuroImagen,
39(3), 1429–1443. DOI: https://doi.org/10.1016/J.NEUROIMAGE
.2007.09.054, PMID: 18035557, PMCID: PMC3658624
Tremblay, S., Shiller, D. METRO., & Ostry, D. j. (2003). Somatosensory
basis of speech production. Naturaleza, 423(6942), 866–869. DOI:
https://doi.org/10.1038/nature01710, PMID: 12815431
Trudeau-Fisette, PAG., Ito, T., & Ménard, l. (2019). Auditory and somato-
sensory interaction in speech perception in children and adults.
Frontiers in Human Neuroscience, 13, 344. DOI: https://doi.org
/10.3389/fnhum.2019.00344, PMID: 31636554, PMCID:
PMC6788346
van Lieshout, PAG., Hulstijn, w., & Peters, h. F. METRO. (1996a). De
planning to articulation in speech production: What differenti-
ates a person who stutters from a person who does not stutter?
Journal of Speech, Idioma, and Hearing Research, 39(3),
546–564. DOI: https://doi.org/10.1044/jshr.3903.546, PMID:
8783133
van Lieshout, PAG., Hulstijn, w., & Peters, h. F. METRO. (1996b). Discurso
production in people who stutter: Testing the motor plan assem-
bly hypothesis. Journal of Speech and Hearing Research, 39(1),
76–92. DOI: https://doi.org/10.1044/jshr.3901.76, PMID:
8820700
Venkatakrishnan, A., Banquet, j. PAG., Burnod, y., & Contreras-Vidal, j. l.
(2011). Parkinson’s disease differentially affects adaptation to gradual
as compared to sudden visuomotor distortions. Human Movement
Ciencia, 30(4), 760–769. DOI: https://doi.org/10.1016/j.humov
.2010.08.020, PMID: 21414678, PMCID: PMC3203839
watkins, k. MI., Herrero, S. METRO., davis, S., & Howell, PAG. (2008). Structural
and functional abnormalities of the motor system in developmental
stuttering. Cerebro, 131(1), 50–59. DOI: https://doi.org/10.1093/brain
/awm241, PMID: 17928317, PMCID: PMC2492392
Wolpert, D. METRO., Ghahramani, Z., & Jordán, METRO. I. (1995). An internal
model for sensorimotor integration. Ciencia, 269(5232), 1880–1882.
DOI: https://doi.org/10.1126/science.7569931, PMID: 7569931
Neurobiology of Language
333
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Stuttering and sensory feedback
Wolpert, D. METRO., Miall, R. C., & Loco, METRO. (1998). Internal models
in the cerebellum. Tendencias en Ciencias Cognitivas, 2(9), 338–347.
DOI: https://doi.org/10.1016/S1364-6613(98)01221-2, PMID:
21227230
Yairi, MI., & Ambrose, norte. (2013). Epidemiology of stuttering: 21st
century advances. Journal of Fluency Disorders, 38(2), 66–87.
DOI: https://doi.org/10.1016/j.jfludis.2012.11.002, PMID:
23773662, PMCID: PMC3687212
Zimmermann, GRAMO. (1980). Articulatory behaviours associated with
stuttering: A cinefluorographic analysis. Journal of Speech and
Hearing Research, 23(1), 108–121. DOI: https://doi.org/10
.1044/jshr.2301.108, PMID: 7442171
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
2
3
0
8
1
9
2
5
1
8
0
norte
oh
_
a
_
0
0
0
3
6
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurobiology of Language
334