Retrospective Selection in Visual and
Tactile Working Memory Is Mediated
by Shared Control Mechanisms
Tobias Katus1,2 and Martin Eimer1
Abstracto
■ Selective attention regulates the activation of working mem-
ory (W.M.) representaciones. Retro-cues, presented after memory
sample stimuli have been stored, modulate these activation
states by triggering shifts of attention to task-relevant samples.
Aquí, we investigated whether the control of such attention
shifts is modality-specific or shared across sensory modalities.
Participants memorized bilateral tactile and visual sample stimuli
before an auditory retro-cue indicated which visual and tactile
stimuli had to be retained. críticamente, these cued samples were
located on the same side or opposite sides, thus requiring
spatially congruent or incongruent attention shifts in tactile and
visual WM. To track the attentional selection of retro-cued sam-
ples, tactile and visual contralateral delay activities (tCDA and
CDA components) were measured. Clear evidence for spatial
synergy effects from attention shifts in visual WM on concurrent
shifts in tactile WM were observed: Tactile WM performance was
impaired, and tCDA components triggered by retro-cues were
strongly attenuated on opposite-sides relative to same-side trials.
These spatial congruency effects were eliminated when cued
attention shifts in tactile WM occurred in the absence of simulta-
neous shifts within visual WM. Results show that, en contraste con
other modality-specific aspects of WM control, concurrent atten-
tional selection processes within tactile and visual WM are medi-
ated by shared supramodal control processes. ■
INTRODUCCIÓN
Working memory ( W.M.) is responsible for the mainte-
nance of perceptual information that is no longer physi-
cally present and for making this information accessible
to other cognitive and response-related processes (p.ej.,
Baddeley, 2012). It is generally believed that spatial atten-
tion plays a central role during the encoding and storage
of sensory signals in WM (see Awh, Vogel, & Oh, 2006, para
discussion). WM maintenance relies on the same fronto-
parietal networks that are also involved in the control of
spatial attention (Awh & En Jonia, 2001). Decreasing the dis-
tance between memorized stimuli reduces WM precision
and increases binding errors in WM (Ahmad et al., 2017;
Emrich & Ferber, 2012), reflecting a competition for spatial
attention during encoding and WM maintenance of visual
objects. This suggests that spatial attention is responsible
for the binding of stimulus features and the formation of
object-based visual representations not only in perception
(p.ej., Robertson, 2003) but also in WM.
If spatial attention is critical for the encoding of sen-
sory stimuli into WM, these encoding processes should
be affected by the spatial location of these stimuli. Este
factor should be particularly important for the concurrent
encoding of to-be-memorized stimuli from different
1Birkbeck, University of London, 2University of Aberdeen
© 2019 Instituto de Tecnología de Massachusetts
sensory modalities in multisensory WM tasks. It has been
suggested that WM for stimuli from different sensory
modalities relies on shared top–down attentional con-
trol mechanisms (p.ej., Cowan, 2011). If this were the
caso, spatial synergies should be observed when to-
be-memorized sample stimuli from different modalities
are presented simultaneously. Por ejemplo, when vi-
sual and tactile sample stimuli appear on opposite
sides, concurrent attention shifts in opposite directions
are required during the encoding of these stimuli. Si
the underlying attentional control processes are linked
across vision and touch, this should result in costs rela-
tive to situations where these stimuli appear on the same
lado. Such spatial synergies have been demonstrated in
previous behavioral and electrophysiological exper-
iments on crossmodal links in spatial attention (p.ej.,
Eimer, van Velzen, & Conductor, 2002; Spence & Conductor,
1996) for the perceptual processing of multisensory
estímulos. Allocating attention to concurrent visual, audi-
conservador, or tactile stimuli was more efficient when these
stimuli appeared at the same location than when they
were presented on opposite sides, suggesting that these
attention shifts are controlled by shared central mecha-
nisms (see Eimer & Conductor, 2001, for further discussion).
Sin embargo, two recent bimodal WM studies from our lab
(Katus & Eimer, 2016, 2019b) have failed to find any
evidence for such spatial congruency effects for the
Revista de neurociencia cognitiva 32:3, páginas. 546–557
https://doi.org/10.1162/jocn_a_01492
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
.
F
t
/
oh
norte
0
5
METRO
a
y
2
0
2
1
encoding and maintenance of visual and tactile sample
estímulos. The goal of the present experiments was to inves-
tigate whether such crossmodal spatial synergies might
emerge in tasks where attention has to be allocated ret-
rospectively to visual and tactile items that are already
stored in WM.
In our first previous study (Katus & Eimer, 2016), vi-
sual and tactile samples were presented bilaterally, pero
only stimuli on one of these sides had to be memorized
in each modality. In different blocks, task-relevant visual
and tactile samples were either located on the same side
or on opposite sides. To assess whether the spatial con-
gruency of these samples affected visual and tactile WM
encoding and maintenance processes, EEG was recorded
during task performance to measure the visual contra-
lateral delay activity (CDA) and its tactile equivalent, el
tCDA component. The CDA and tCDA are elicited contra-
lateral to the side of task-relevant visual sample stimuli
(p.ej., Vogel & Machizawa, 2004) or tactile sample stimuli
(p.ej., Katus & Eimer, 2015). Both components are sensi-
tive to WM load and individual differences in WM capacity
(Katus, Grubert, & Eimer, 2015; Vogel & Machizawa,
2004) and have modality-specific topographies over vi-
sual and somatosensory cortex, respectivamente. Esto indica
that they reflect the activation of WM representations in
modality-specific perceptual cortical areas, as proposed
by the sensory recruitment account of WM (Postle, 2006;
En Jonia, Lacey, & Nee, 2005). In our study (Katus & Eimer,
2016), visual CDA and tCDA components were elicited
over the same hemisphere in same-side blocks and over
different hemispheres in opposite-sides blocks, contra-
lateral to the respective task-relevant visual and tactile
sample items. críticamente, no crossmodal spatial congruency
effects were found; visual CDA and tCDA were elicited at
the same time and were identical in amplitude in same-
side and opposite-sides blocks, and there were also no
costs for WM performance in opposite-sides blocks.
Similar results were observed in our second bimodal
tactile/visual WM study (Katus & Eimer, 2019b). Aquí,
participants memorized task-relevant visual and tactile
stimuli in a first memory sample set (S1) before encoding
relevant samples in a second sample set (S2). críticamente,
relevant visual or tactile S2 samples appeared unpredict-
ably on the same side or on the side opposite to the task-
relevant S1 samples. De este modo, the foci of tactile and visual
attention could either be maintained on the same side
or had to be redirected to opposite sides in the period
following the S2 samples. As expected, CDA and tCDA
components reversed polarity on trials where visual or
tactile S1 and S2 samples appeared on opposite sides, re-
flecting shifts of spatial attention on these trials. Sin embargo
and importantly, there were no crossmodal interactions in
spatial selection: The visual CDA was entirely unaffected
by concurrent attention shifts in tactile WM, y el
tCDA was insensitive to shifts of attention in visual WM.
Visual and tactile WM performance was also not modu-
lated by attention shifts in the other modality. En general,
the results of these two studies suggest that the control
processes that allocate spatial attention during the en-
coding and subsequent maintenance of visual and tactile
sample stimuli are not linked but operate in an entirely
independent modality-specific fashion (see also Katus &
Eimer, 2018, 2019a, for analogous evidence for indepen-
dent capacities of spatial WM in touch and vision).
In these previous experiments, attention shifts could
already take place during the encoding of the sample dis-
plays into WM, as the identity of the task-relevant target
samples was known in advance (es decir., before the samples
were presented). Sin embargo, selective spatial attention
does not only mediate encoding but can also selectively
modulate the activation states of representations that are
already stored in WM. This has been demonstrated in ex-
periments where retro-cues presented during the delay
period specified a subset of task-relevant visual sample
estímulos (p.ej., Lepsien & Nobre, 2006; Griffin & Nobre,
2003). These retro-cues produced clear benefits for WM
actuación, demonstrating that attention modulates
WM representations in line with task goals. ERP studies
have further shown that retro-cues trigger visual CDA
(kuo, alimenta, & Nobre, 2012; Eimer & Kiss, 2010) y
tCDA (Katus, Müller, & Eimer, 2015) componentes, reflejar-
ing changes in the attentional activation states of visual
and tactile WM representations in bimodal memory tasks
(Katus, Grubert, & Eimer, 2017).
Previous retro-cue studies demonstrate that spatial at-
tention can be selectively allocated to representations
that are already stored in WM. This raises the question
whether the retrospective selection of information in
WM is mediated by processes that operate in a strictly
modality-specific fashion or whether these processes
are linked across different modalities, resulting in cross-
modal interactions during spatial selection. The goal of
the current study was to investigate this question. Nosotros
employed bimodal WM tasks where visual/tactile sample
stimuli were followed by retro-cues that marked a subset
of the visual and tactile samples as relevant for a com-
parison with a subsequent test stimulus (unpredictably
vision or touch). In Experiment 1, participants had to
initially encode four visual samples presented in both vi-
sual fields and two tactile samples presented to both
manos. These samples were either filled or unfilled (ver
Methods section for details), and an auditory retro-cue
presentado 700 msec after the sample stimuli indicated
whether filled or unfilled visual and tactile samples had to
be retained (ver figura 1). Once the retro-cue appeared,
attention could be allocated to the relevant visual and tactile
sample items, and irrelevant samples could be dropped
from WM. The critical manipulation concerned the spa-
tial congruency of the visual and tactile samples that had
to be maintained. On same-side trials, these samples
had been presented on the same side (both left or both
bien). On opposite-sides trials, their spatial relationship
was incongruent (visual samples on the left and tactile
samples on the right, or vice versa).
Katus and Eimer
547
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
t
F
.
/
oh
norte
0
5
METRO
a
y
2
0
2
1
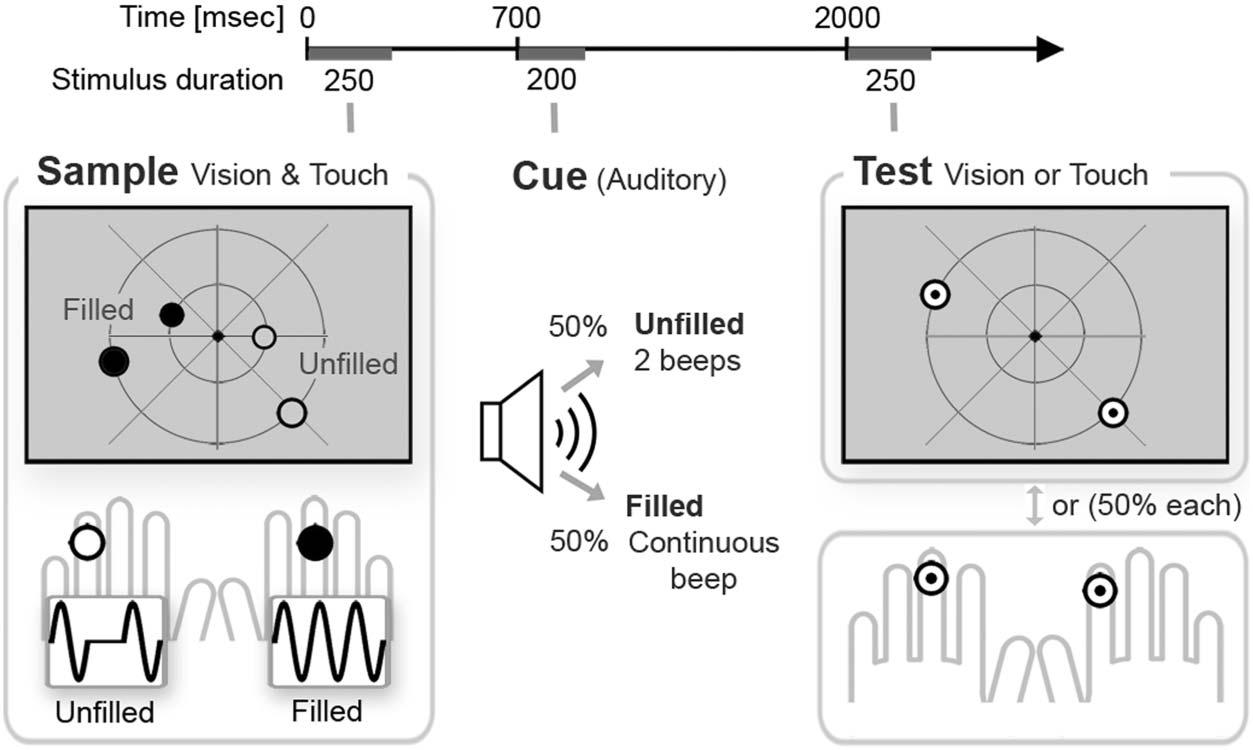
Cifra 1. Bimodal retro-cue
working memory task in
Experimento 1. A bimodal
visual/tactile sample set was
followed by an auditory
retro-cue and a unimodal tactile
or visual (50% cada) prueba
stimulus set. The tactile samples
included one filled and one
unfilled stimulus, presented to
opposite hands. lo visual
samples included pairs of filled
and unfilled circles, presentado
in opposite visual hemifields.
The auditory retro-cue signaled
whether filled or unfilled
sample stimulus locations
(50% cada) had to be retained
for comparison with the test
stimulus. The side where filled
and unfilled sample stimuli
were presented was determined independently for vision and touch, Resultando en 50% same-side trials (filled/unfilled tactile and visual samples on the
same side) y 50% opposite-sides trials (filled tactile sample on the left and filled visual samples on the right, or vice versa). Memory was
unpredictably tested for either vision or touch, with test stimulus sets that always contained one stimulus on the left side and one on the right
lado. Experimento 2 used analogous procedures, except that the task-relevant side for visual samples was specified in advance and remained
constant for six blocks.
CDA and tCDA components were measured during the
intervals between samples and retro-cues and between
retro-cues and test displays, separately for same-side
and opposite-sides trials, and WM performance was also
compared between these two types of trials. El critico
question was whether spatial congruency would affect
the selective activation of retrospectively cued visual
and tactile WM representations and subsequent WM per-
rendimiento. If retrospective attention shifts within visual
and tactile WM are mediated by shared top–down control
procesos, spatial synergies should emerge. This should
be reflected by attenuated visual CDA and tCDA com-
ponents in response to retro-cues on opposite-sides trials
relative to same-side trials and by impaired visual and tactile
WM performance on opposite-sides trials. Alternativamente,
the absence of such spatial congruency effects would
suggest that the allocation of attention to visual and tac-
tile representations that are already stored in WM is not
controlled by a central supramodal system but by inde-
pendent modality-specific processes.
EXPERIMENT 1
Métodos
Participantes
Sixteen participants (edad media = 29 años; nine women,
one left-handed) took part in Experiment 1. Three addi-
tional participants were excluded from statistical analysis
because of excessive alpha activity and EEG artifacts that
led to the exclusion of more than 30% of trials. All par-
ticipants were neurologically unimpaired and gave in-
formed written consent before testing. The experiment
was conducted in accordance with the Declaration of
Helsinki and was approved by the Psychology Ethics
Committee of Birkbeck, University of London.
Stimulus Material and Setup
Participants were seated in a dimly lit recording chamber
with their hands covered from sight. Headphones played
continuous pink noise during EEG recordings to mask any
sounds produced by tactile stimulation. Tactile stimuli
(100 Hz sinusoids, intensidad 0.37 norte, duración 250 mseg)
were delivered by eight mechanical stimulators that were
attached to the left and right hands’ distal phalanges of the
índice, middle, ring, and little fingers. The stimulators were
driven by custom-built amplifiers, controlled by MATLAB
routines (The MathWorks) via an eight-channel sound
card (M-Audio, Delta 1010LT). There were two types of
tactile sample stimuli (filled and unfilled). For filled stim-
uli, a continuous vibration was presented for 250 mseg.
Unfilled stimuli consisted of two 20-msec pulses, sepa-
rated by a 210-msec delay. Tactile memory test stimuli
consisted of two 60-msec pulses separated by a 130-msec
delay.
Visual stimuli were shown at a viewing distance of 100 cm
against a dark gray background on a 22-in. monitor
(Samsung SyncMaster 2233; 100-Hz refresh rate, 16-msec re-
sponse time). All visual stimuli were presented for 200 mseg.
Analogous to the tactile stimulus material, three types of
visual stimuli were used (filled samples, unfilled samples,
test stimuli). These visual stimuli were presented against a
background of black crosshairs (four lines at 0°, 45°, 90°,
and 135° of polar angle; horizontal/vertical eccentricity:
3.49° of visual angle) and two concentric black rings around
the fixation dot. This background remained constantly on-
screen during task performance. The eccentricity of the
548
Revista de neurociencia cognitiva
Volumen 32, Número 3
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
t
F
.
/
oh
norte
0
5
METRO
a
y
2
0
2
1
two rings was 2.52° and 5.04°. Visual sample and test stimuli
were presented on top of these rings (ver figura 1). Su
size was scaled for eccentricity (0.34° vs. 0.46° for stimuli on
the inner vs. outer ring, respectivamente). Visual sample stimuli
were filled white circles or unfilled white outline rings (line
width: 0.12 y 0.16 cm, for inner and outer stimuli). Visual
test stimuli were white outline rings with a small white dot in
the ring’s center (diameter: 0.12 y 0.16 cm for inner and
outer stimuli).
Two different auditory retro-cues (600 Hz, 200-mseg
duration with 5-msec ramps, presented via headphones)
signaled the stimulus type in the bimodal sample set
(filled or unfilled) that had to be retained. Task-relevant
filled tactile/visual stimuli were indicated by a continuous
tono. Task-relevant unfilled stimuli were signaled by a
retro-cue consisting of two 50-msec tones separated by
a 100-msec interval. Responses were given vocally during
the 1800-msec period after the presentation of the test
stimulus set (“a” for match and “e” for mismatch, ver
abajo). They were recorded by a headset microphone,
were analyzed online with custom-written voice-key rou-
tines, and were manually rechecked for correctness after
the experiment.
Task Design and Stimulation Procedure
Simultaneously presented tactile and visual sample stim-
uli (250 msec duration) were followed after 700 msec by
an auditory retro-cue (200 msec duration) and after
2000 msec by a unimodal tactile (50%) or visual (50%)
test stimulus set (250 msec duration). Tactile WM load
was one item per side, and visual WM load was two
items per side. Tactile sample sets included one filled
and one unfilled stimulus, presented to randomly deter-
mined fingers of the left and right hand. A pair of filled
visual samples was presented in one hemifield, y un
pair of unfiled samples was presented in the other hemi-
campo. The precise locations of these visual sample stimuli
were sampled from 102 angular positions (in polar co-
ordinates, left side: 130°–230°, right side: 310°–50°), con
the constraint that the sampled positions were at least
25° apart. For each side, the two selected positions were
randomly assigned to the inner and outer rings on the
monitor, with one sample being shown on top of each
ring (ver figura 1). Fundamentalmente, tactile and visual sample
stimuli of the same type (filled or unfilled) were either
located on the same side or on opposite sides. Same-side
and opposite-sides trials were equally likely and varied
unpredictably within each block.
Participants first had to encode all visual and tactile
sample stimuli on both sides. Following the retro-cue
(a continuous tone or two tones with a gap, presentado
with equal probability and in random order in each
block), they had to selectively maintain the locations of
the cued task-relevant tactile and visual samples (filled
or unfilled) solo. Memory was unpredictably tested for
touch or vision (50% cada), with test stimulus sets
containing one stimulus on the left side and one on
the right side. Participants had to decide whether the
location of a cued visual or tactile sample matched the
location of a (visual or tactile) test stimulus. This was
the case on match trials (50%). On mismatch trials
(50%), the test stimulus location differed from the mem-
orized sample location. For tactile mismatch trials, el
stimulus was randomly delivered to one of the three
fingers where no sample had been presented. For visual
mismatch trials, the position of one of the two relevant
samples was randomly shifted either upward or down-
ward by 30° on its ring in the test display. The task-
irrelevant uncued visual or tactile samples could also
spatially match or mismatch with the stimuli on the irrel-
evant (uncued) side of the test stimulus. Spatial matches
or mismatches between sample and tests were indepen-
dently randomized for the cued (relevant) and uncued
(irrelevante) test stimulus locations.
The experiments comprised 528 trials each that were
run in 12 bloques. There were four experimental condi-
ciones (spatial congruency: relevant visual and tactile
samples on the same vs. opposite sides; tested modality:
touch vs. visión) con 132 trials each, which unpre-
dictably alternated within each block. Training was run
before each experiment (depending on individual perfor-
mance between ∼20 and 44 ensayos). Feedback about the
proportion of correct responses was given after each
block.
Acquisition and Preprocessing of EEG Data
EEG data, sampled at 500 Hz using a BrainVision ampli-
fier, were DC recorded from 64 Ag/AgCl active electrodes
at standard locations of the extended 10–20 system. Two
electrodes at the eyes’ outer canthi monitored horizontal
eye movements (horizontal EOG, HEOG). Continuous
EEG was acquired with left mastoid reference and re-
referenced to the arithmetic mean of both mastoids for
data preprocessing. Data were offline submitted to a
20-Hz low-pass filter (Blackman window, filter order
1000). Epochs were extracted for the 2-sec period follow-
ing the sample set and were corrected for a 200-msec
prestimulus baseline that was taken before the onset of
the tactile/visual sample stimuli.
Artifact Rejection and Correction
Trials with saccades were rejected using a differential
step function that ran on the bipolarized HEOG (step
width 200 mseg, límite 30 μV). Independent compo-
nent analysis (Delorme, Sejnowski, & Makeig, 2007) era
subsequently used to correct for frontal artifacts such as
eye blinks and residual traces of horizontal eye move-
ments that had not been detected by the step function.
We rejected trials in which difference values for corre-
sponding left- minus right-hemispheric electrodes ex-
ceeded a fixed threshold of ±50 μV for at least two
Katus and Eimer
549
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
.
F
/
t
oh
norte
0
5
METRO
a
y
2
0
2
1
electrode pairs. We also excluded trials where amplitudes
at any electrode exceeded a fixed 150-μV threshold. El
remaining epochs entered Fully Automated Statistical
Thresholding for EEG Artifact Rejection (Nolan, Whelan,
& Reilly, 2010) for the interpolation of noisy electrodes
and were subsequently converted to current source densi-
corbatas (CSDs: iterations = 50, m = 4, lambda = 10−5; Tenke
& Kayser, 2012); 98.4% of epochs remained for statistical
analysis on average. Statistical tests were based on correct
and incorrect trials, as the exclusion of incorrect trials did
not change the pattern of results but would have de-
creased the signal-to-noise ratio of EEG data.
Selection of electrodes and time windows; topographical
maps. We separately averaged CSDs across three adja-
cent electrodes contralateral and ipsilateral to the task-
relevant side. As in prior studies (Katus & Eimer, 2016,
2018, 2019a), the tCDA and visual CDA components were
measured at lateral central (tCDA: C3/4, FC3/4, CP3/4)
and occipital scalp regions (CDA: PO7/8, PO3/4, O1/2).
Statistical tests were conducted on difference values of
contra- minus ipsilateral CSDs averaged between 1050
y 2000 msec after the sample set, corresponding to a
time window of 350–1300 msec following the retro-cue
(cf. McCants, Katus, & Eimer, 2020; Katus, Müller, et al.,
2015).
Spline-interpolated voltage maps illustrate the topo-
graphical distribution of lateralized activity in the period
after the cue. These maps were obtained by subtracting
ipsilateral CSDs from contralateral CSDs, with contra-/
ipsilateral referring to the side where the task-relevant vi-
sual sample stimuli had been presented. To collapse data
across blocks where the visual samples on the left versus
right side were task-relevant, electrode coordinates were
flipped over the midline for visual left-side memory trials.
Por lo tanto, in the topographical maps, a negative poten-
tial over the left hemisphere indicates the presence of
delay activity contralateral to the visual targets.
Statistical analyses. Data were analyzed with paired
t tests and repeated-measures ANOVAs. Error bars in
graphs indicate confidence intervals for the true popula-
tion mean. De este modo, error bars that do not overlap with the
zero axis ( y ≠ 0) inform about statistically significant
tCDA/CDA components.
Bayesian t tests (Rojo, Speckman, Sol, Morey, &
Iverson, 2009) and the software JASP ( JASP Team,
2016) were used to calculate Bayes factors for each main
effect/interaction in our statistical designs. The Bayes
factor denotes the relative evidence for the alternative hy-
pothesis as compared with the null hypothesis and thus
allows for statistical inferences regarding the presence or
absence of a modulation. The Bayes factor for the null hy-
pothesis (BF01) corresponds to the inverse of the Bayes
factor for the alternative hypothesis (BF10) and indexes
the relative evidence in the data that an effect is absent
rather than present. We report the numerically larger BF
and categorize the evidence as substantial (for BFs > 3
y < 10), strong (BFs > 10 y < 30), very strong
(BFs > 30 y < 100), or decisive (BFs > 100) according
to the convention suggested by Jeffreys (1961).
Resultados
Behavioral Performance
Accuracy data were analyzed using an ANOVA with the
factors Tested Modality (touch or vision) and Spatial Congru-
ency (same-side trials, opposite-sides trials). Actuación
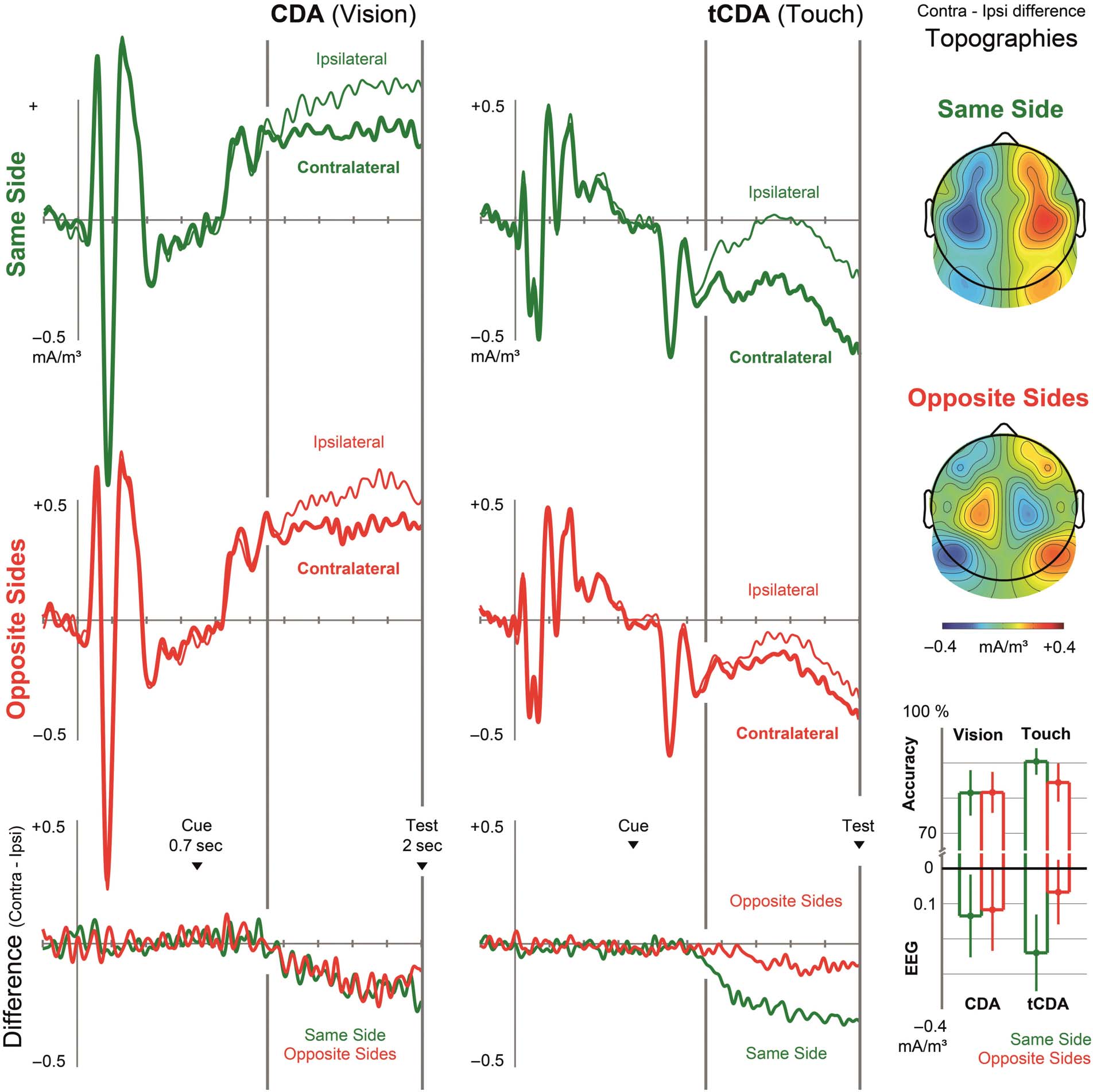
(significar 84.5% correcto) was lower in the visual as compared
with tactile task (81.6% vs. 87.4%, Tested Modality: F(1,
15) = 9.074, pag = .009, BF10 = 6.107), and was generally
reduced on opposite-sides relative to same-side trials
(83.0% vs. 86.0%, Spatial Congruency: F(1, 15) = 9.261,
pag = .008, BF10 = 6.434). The interaction between both fac-
tors was significant (F(1, 15) = 7.842, pag = .014, BF10 =
4.288). Follow-up analyses conducted separately for trials
where tactile or visual WM was tested showed that tactile
WM accuracy was impaired on opposite-sides relative to
same-side trials (84.4% vs. 90.4%, t(15) = 4.357, pag <
.001, BF10 = 62.856). In contrast, no such spatial congru-
ency effect was found for visual WM performance (81.6%
vs. 81.5% for opposite-side vs. same-side trials, t(15) =
0.084, p = .934, BF10 = 3.903).
EEG Data
Amplitudes of the tCDA and CDA components were mea-
sured between 350 and 1300 msec after the onset of the
retro-cue (see Methods section for electrode locations
used to compute the tCDA and CDA). We examined
whether the amplitudes of these components were sensi-
tive to the spatial congruency of cued task-relevant visual
and tactile sample stimuli in WM (same-side vs. opposite-
sides trials). We obtained statistically significant visual CDA
components in both types of trials (same-side: t(15) =
2.447, p = .027, BF10 = 2.425; opposite-sides: t(15) =
2.144, p = .049, BF10 = 1.530). The CDA did not differ in
size between same-side and opposite-sides trials (t(15) =
0.254, p = .803, BF01 = 3.805), as CDA components of
equal size were obtained on both types of trials (see bar
graphs in Figure 2). In contrast, the tCDA was smaller on
opposite-sides relative to same-side trials (t(15) = 4.394,
p < .001, BF10 = 67.012). In fact, tCDA amplitudes were
not significantly different from zero on opposite-sides trials
(t(15) = 1.562, p = .139), although the evidence for the null
hypothesis was not conclusive in this case (BF01 = 1.428).
In contrast, the tCDA in same-side trials was highly reli-
able (t(15) = 4.691, p < .001, BF10 = 112.039).
Discussion
We used retro-cue procedures to investigate whether
concurrent attention shifts to visual and tactile WM
550
Journal of Cognitive Neuroscience
Volume 32, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
5
-
4
p
6
d
f
2
0
/
1
3
3
2
1
/
3
3
6
/
5
o
4
c
6
n
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
2
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
0
5
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
5
-
4
p
6
d
f
2
0
/
1
3
3
2
1
/
3
3
6
/
5
o
4
c
6
n
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
2
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
f
.
o
n
0
5
M
a
y
2
0
2
1
Figure 2. Experiment 1. CSDs were measured over visual and somatosensory scalp regions contralateral and ipsilateral (thick vs. thin line) to
the cued tactile/visual target samples. The top and middle rows show CSDs recorded in same-side and opposite-sides trials, respectively. The bottom
row shows contra- minus ipsilateral difference waves. Topographies display the scalp distribution of lateralized effects elicited between 350 and
1300 msec after the onset of the retro-cue (plotted relative to the task-relevant side of the visual samples, note the polarity reversal of the tCDA
on opposite-sides trials). Bar graphs show average accuracy in the visual and tactile tasks and mean amplitudes of lateralized components over
visual and somatosensory cortical regions in the period after the retro-cue in same-side trials (green) and opposite-sides trials (red). Error bars
indicate 95% confidence intervals.
representations are controlled independently for these
two modalities or whether there are spatial synergies be-
tween these attentional shifts, indicative of crossmodal
links. The results of Experiment 1 were clear but some-
what unexpected. The allocation of attention to retro-
cued visual WM representations was entirely unaffected
by the spatial congruency of simultaneous attention shifts
toward task-relevant tactile WM representations. Visual
CDA components emerged in the interval after a retro-
cue had been presented, confirming that attention was
directed toward visual WM representations that were sig-
naled as task-relevant by these cues. CDA amplitudes did
not differ between same-side and opposite-sides trials,
and visual WM performance was also virtually identical
on these two types of trials. These results provide strong
evidence that the control of attention shifts within visual
WM operates independently of any concurrent attention
shifts in tactile WM. In marked contrast, these visual
Katus and Eimer
551
attention shifts produced clear spatial synergy effects for
the concurrent activation of tactile WM representations
during the period after the retro-cue. The amplitude of
tCDA components measured during this period was
strongly attenuated on opposite-sides relative to same-
side trials, and the tCDA was not significantly different
from zero on opposite-sides trials. Tactile WM perfor-
mance was also impaired on these trials.
If the control of attention shifts within WM was shared
between vision and touch, one would have predicted
symmetrical spatial synergy effects for both modalities.
In fact, these effects were only found for the allocation
of attention to tactile but not visual WM representations
in Experiment 1, indicating that attention shifts toward
cued samples in visual WM affected concurrent shifts in
tactile WM more strongly than vice versa. This asymmetry
between sensory modalities could reflect a generic bias in
the attentional control of WM toward vision (cf. Katus
et al., 2017) but could also be a result of the specifics
of the bimodal WM task used in this experiment. As
WM load was higher in the visual task than in the tactile
task (two items vs. one item), WM maintenance demands
were higher for vision than for touch, which is also
suggested by the observation that WM performance
was lower in the visual task. Determining the identity
of retrospectively cued targets, in contrast, was relatively
straightforward in vision, where selection was based on a
simple feature (filled versus unfilled dots). The tactile tar-
gets and distracters were harder to distinguish because
the tactile filled and unfilled stimuli were initially identi-
cal, with the gap that defined unfilled samples appearing
only after 20 msec. Because WM load was higher in vision
and the attribute relevant for retrospective selection was
more salient, participants may have prioritized atten-
tional shifts in visual WM, resulting in spatial synergy ef-
fects from vision to touch but not vice versa.
Regardless of the exact factors responsible for this
asymmetry, the fact that there were strong behavioral
and electrophysiological crossmodal spatial congruency
effects for tactile WM provides clear evidence that the
attentional control processes activated in response to
the retro-cues were linked across sensory modalities.
These results differ markedly from our previous visual/
tactile WM experiments that did not involve retro-cues,
where no such links between the attentional control of
visual and tactile WM were found (Katus & Eimer, 2016,
2019b). This striking dissociation suggests that the en-
coding of sensory stimuli into WM and their subsequent
maintenance on the one hand and the selective allocation
of attention to WM representations that are already stored
in WM on the other are controlled by qualitatively distinct
mechanisms (see also Katus, Andersen, & Müller, 2012).
The former operate in a modality-specific fashion, whereas
the latter are shared between sensory modalities. If this
was the case, evidence for crossmodal links between visual
and tactile WM should only be found in tasks where atten-
tion shifts to retro-cued visual and tactile WM represen-
tations are activated at the same time in both modalities.
When such shifts within WM are only required in touch,
they should not be affected by the spatial congruency of
simultaneously maintained visual WM representations.
Experiment 2 was conducted to test this prediction.
EXPERIMENT 2
To determine whether spatial congruency effects on at-
tention shifts to retro-cued tactile WM representations
are eliminated in the absence of concurrent attention
shifts within visual WM, we slightly modified the pro-
cedures used in Experiment 1. Stimulus presentation
procedures and the timing of events remained the same,
but the retro-cues were now only relevant for touch. In
Experiment 2, the location of task-relevant visual sample
stimuli (left or right visual hemifield) was specified be-
fore the first experimental block and remained constant
for six successive blocks before it was reversed. As the
location of task-relevant visual sample stimuli was known
in advance, participants could immediately encode these
stimuli into WM before the arrival of the retro-cue. For
this reason, visual CDA components were expected to
emerge already during the interval between the sample
display and the retro-cue, reflecting the attentional acti-
vation of visual WM representations during this period.
However, the retro-cue still indicated whether tactile sam-
ple stimuli on the left or right side had to be retained.
Thus, attention shifts to task-relevant tactile WM represen-
tations could only be initiated after the retro-cue was pre-
sented, as in Experiment 1. Therefore, tCDA components
should again only be present during the interval between
the retro-cue and the test display.
The critical manipulation in Experiment 2 again con-
cerned the spatial relationship between task-relevant visual
and tactile sample stimuli. On same-side trials, retro-cued
tactile samples were located on the side that was consis-
tently task-relevant for the visual WM task. On opposite-
sides trials, these tactile samples were located on the
other side. Thus, the attention shifts within tactile WM
prompted by the retro-cues were spatially congruent or
incongruent with the representations in visual WM that
had already been activated before the retro-cues had
been presented. The central question was whether the
spatial congruency effects found for tactile WM in Ex-
periment 1 would also be found in Experiment 2. If these
effects reflect crossmodal links that are specific to the
control of retrospective selection processes in tactile
and visual WM, they should now be absent, as no shifts
of attention within visual WM were required in response
to the retro-cues. Alternatively, the allocation of attention
to retro-cued tactile WM representations may be more
generally affected by the ongoing maintenance of visual
WM representations on the same versus opposite side. In
this case, Experiment 2 should find similar crossmodal
spatial congruency effects for tactile WM as were ob-
served in Experiment 1.
552
Journal of Cognitive Neuroscience
Volume 32, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
5
-
4
p
6
d
f
2
0
/
1
3
3
2
1
/
3
3
6
/
5
o
4
c
6
n
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
2
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
t
/
o
n
0
5
M
a
y
2
0
2
1
Methods
Participants
Sixteen neurologically unimpaired participants (mean
age = 32 years; nine women, all right-handed) took part
in Experiment 2. One additional participant was ex-
cluded because of excessive EEG artifacts. Informed
consent and ethics approval procedures were the same
as in Experiment 1.
Stimulus Material, Setup, Task Design, and
Stimulation Procedure
These were all identical to Experiment 1, with one impor-
tant exception: In Experiment 2, the retro-cue was only
relevant for the tactile modality. The side of the task-
relevant visual samples (left or right hemifield) was
now indicated via verbal instruction and on the computer
screen before the first experimental block and remained
constant until the seventh block. The visual sample stim-
uli on the task-relevant side were equiprobably filled (in
which case they were accompanied by unfilled distracters
on the opposite side), or they were unfilled (and pre-
sented along with filled distracters on the opposite side).
Whether the left- or right-side visual samples were rele-
vant in the first or second experimental half was randomly
determined for each participant. Participants therefore had
to encode visual sample locations only on the relevant side
and tactile sample locations on both sides before the retro-
cue indicated which tactile sample (filled or unfilled) had to
be retained in WM. Note that this blocking of relevant
visual sample locations did not affect the spatial congru-
ency manipulation (same-side vs. opposite-sides trials),
which still varied unpredictably on a trial-by-trial basis,
depending on which tactile sample was indicated as rel-
evant by the retro-cue.
Acquisition, Preprocessing of EEG Data, Artifact
Rejection, and Correction
These were all identical to Experiment 1. After artifact re-
jection, 98.7% of epochs remained for statistical analysis
on average.
Results
Behavioral Performance
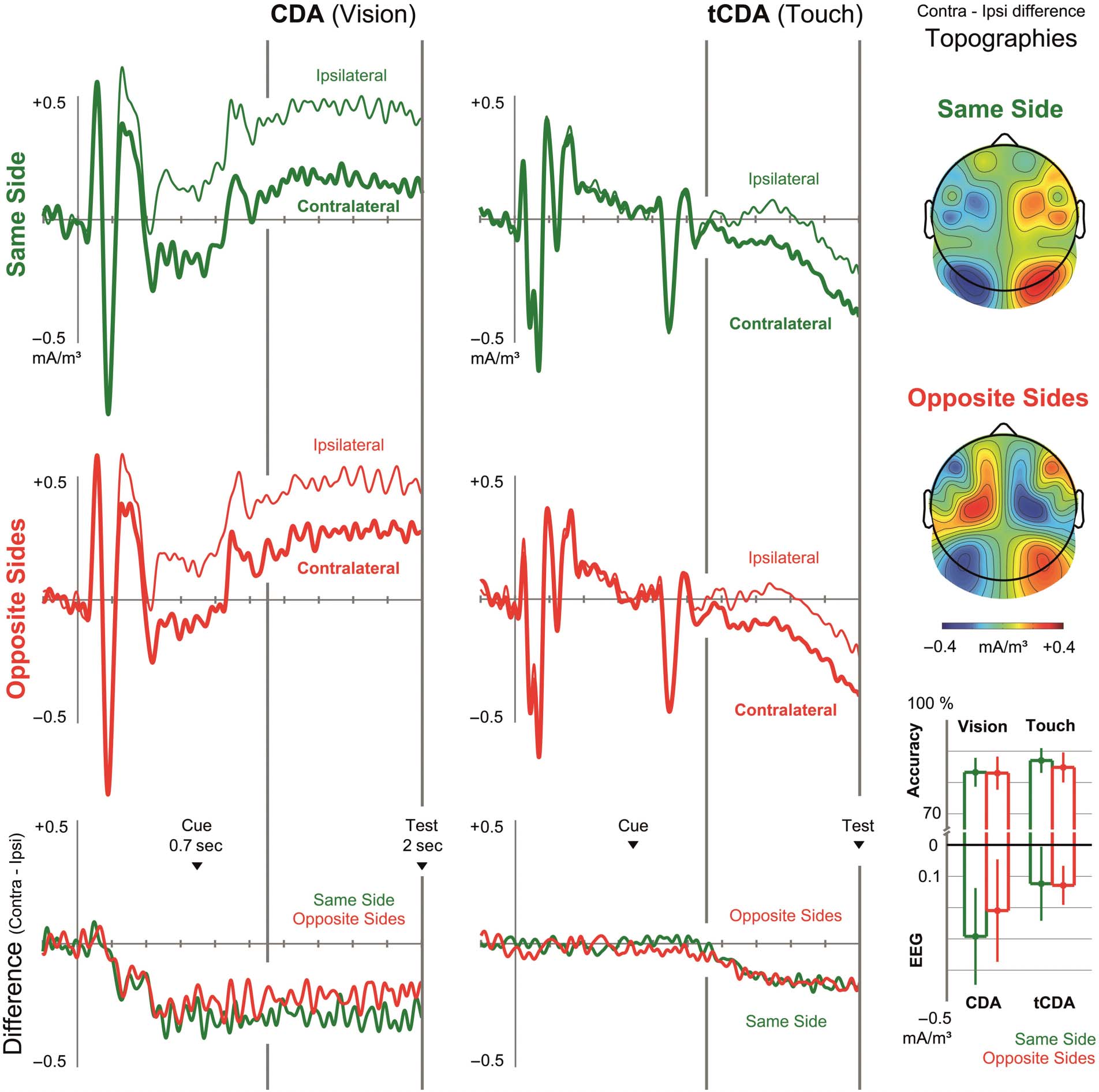
Participants correctly responded in 84.5% of all trials. An
ANOVA did not yield any significant main effects or inter-
action. Performance did not significantly differ between
trials where touch versus vision was tested (85.9% vs.
83.1% correct; Tested Modality: F(1, 15) = 1.020, p =
.329, BF01 = 2.521), there was no significant difference in
performance between opposite-sides versus same-side trials
(83.9% vs. 85.1%; Spatial Congruency: F(1, 15) = 2.090, p =
.169, BF01 = 1.635). In contrast to Experiment 1, the inter-
action between these two factors was not significant (F(1,
15) = 1.942, p = .184, BF01 = 1.733). Although tactile
WM performance tended to be lower on opposite-sides rel-
ative to same-side trials (84.8% vs. 87.0%), this difference
was not significant (t(15) = 1.711, p = .108, BF01 = 1.187).
EEG Data
Because the location of the task-relevant visual (but not
tactile) samples was already known before the onset of
the retro-cues in Experiment 2, the visual CDA compo-
nent was expected to emerge shortly after the presenta-
tion of the sample set, whereas the tCDA should only
appear after the retro-cue, as in Experiment 1. These pre-
dictions were confirmed. In the period before the retro-
cue (i.e., 300–700 msec after presentation of the sample
set), a reliable CDA was elicited (t(15) = 4.144, p = .001,
BF10 = 43.307), whereas there was no evidence for the pres-
ence of a contralateral negativity over somatosensory cortex
(t(15) = 0.115, p = .910, BF01 = 3.892; see Figure 3). The
absence of any lateralized effect at central electrodes (i.e., no
tCDA) during the period where a strong visual CDA was
already present over posterior electrodes demonstrates
that our CSD analysis method was successful in prevent-
ing volume conduction of electrical activity over visual
areas to more anterior sites.
During the period after the retro-cue, both the tCDA and
CDA components were present in all experimental condi-
tions (tCDA same-side: t(15) = 2.223, p = .042, BF10 =
1.720; tCDA opposite-sides: t(15) = 4.408, p = .001,
BF10 = 68.684; CDA same-side: t(15) = 4.025, p =
.001, BF10 = 35.187; CDA opposite-sides: t(15) = 2.730,
p = .016, BF10 = 3.8165; see also Figure 3, where statis-
tically significant lateralized effects are marked by error
bars that do not overlap with the zero axis). As in Ex-
periment 1, the CDA amplitudes did not differ between
same-side and opposite-sides trials (t(15) = 0.972, p =
.346, BF01 = 2.601). But in marked contrast to Exper-
iment 1, there was now also no longer a significant
difference between tCDA amplitudes on opposite- and
same-side trials (t(15) = 0.092, p = .928, BF01 = 3.900).
Comparison between Experiments
To formally assess whether the congruency effects (for
same- vs. opposite-sides trials) in tCDA amplitudes and
tactile WM accuracy differed between the two experi-
ments, we analyzed the behavioral and ERP results ob-
tained in Experiments 1 and 2. We used mixed ANOVAs
with the between-subject factor Experiment and the
within-subject factor Spatial Congruency. For tCDA ampli-
tudes, we found no main effect of Experiment (F(1, 30) =
0.316, p = .578, BF01 = 2.874) and only a marginal main
effect of Spatial Congruency (F(1, 30) = 3.800, p = .061,
BF10 = 1.376). However, the interaction was statistically
significant (Experiment × Spatial Congruency: F(1, 30) =
4.295, p = .047, BF10 = 2.242), indicating that spatial con-
gruency effects on tCDA amplitudes were indeed more
Katus and Eimer
553
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
5
-
4
p
6
d
f
2
0
/
1
3
3
2
1
/
3
3
6
/
5
o
4
c
6
n
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
2
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
5
-
4
p
6
d
f
2
0
/
1
3
3
2
1
/
3
3
6
/
5
o
4
c
6
n
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
2
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
Figure 3. Experiment 2. CSDs over visual and somatosensory scalp regions were measured contralateral and ipsilateral (thick vs. thin line) to the
task-relevant tactile/visual target samples. In Experiment 2, the task-relevant side was blocked for vision, so that retro-cues were only relevant for the
tactile WM task. The top and middle rows show CSDs recorded contralateral and ipsilateral to task-relevant visual and tactile sample stimuli in
same-side and opposite-sides trials, respectively. The bottom row shows contra-/ipsilateral difference waves. Topographies display the scalp
distribution of lateralized effects during the time period after the retro-cue (plotted relative to the task-relevant side of visual samples, hence
the polarity reversal of the tCDA for opposite-sides trials). Bar graphs show average accuracy in the visual and tactile tasks and mean amplitudes of
the CDA and tCDA in the period after the retro-cue in same side trials (green) and opposite sides (red) trials, with error bars indicating 95%
confidence intervals.
pronounced in Experiment 1 relative to Experiment 2.
For tactile WM accuracy, an analogous mixed ANOVA re-
vealed a reliable main effect of Spatial Congruency (F(1,
30) = 18.169, p < .001, BF10 = 67.174) and a significant
interaction between Experiment × Spatial Congruency
(F(1, 30) = 4.976, p = .033, BF10 = 2.019), again reflecting
a larger congruency effect in Experiment 1. The main
effect of Experiment was not significant for tactile WM
performance (F(1, 30) = 0.267, p = .609, BF01 = 1.764).
Analogous analyses were also conducted for CDA ampli-
tudes and visual WM performance. As expected, there were
no significant effects for CDA amplitudes (all ps > .1) o
visual WM accuracy (all ps > .6), confirming that no spatial
congruency effects were present for these measures in
either experiment. Although CDA amplitudes were numer-
ically larger in Experiment 2, this difference between exper-
iments was not reliable (F(1, 30) = 2.772, pag = .106, BF01 =
2.710).1
554
Revista de neurociencia cognitiva
Volumen 32, Número 3
Discusión
The results of Experiment 2 were clear-cut. In marked
contrast to Experiment 1, there were now no longer
any spatial congruency effects for tactile WM. The ampli-
tudes of tCDA components elicited in the period after
the retro-cue was presented were essentially identical
on same-side and opposite-sides trials. There was also
no significant impairment of tactile WM performance
on opposite-sides trials. The presence of spatial con-
gruency effects on tactile WM in Experiment 1 y el
absence of these effects in Experiment 2 were substanti-
ated by significant interactions between experiment and
spatial congruency in the between-experiment analyses
reported above. The critical factor that changed between
these two experiments was the point in time when atten-
tion could be allocated to task-relevant visual WM repre-
sentaciones. In Experiment 2, the to-be-attended side for
vision was blocked, so that task-relevant visual samples
could be attended and encoded into WM immediately
after the sample display was presented. The presence
of reliable CDA components in the period between the
samples and the retro-cues confirms that this was indeed
the case. Following the retro-cues, attention shifts to-
ward task-relevant sample stimuli were required within
tactile WM only, whereas the previously established focus
of attention in visual WM could be maintained. De este modo, el
absence of spatial congruency effects for tactile WM in
Experimento 2 and the presence of such effects in Exper-
mento 1 were associated with the absence versus presence
of concurrent attention shifts toward retro-cued WM repre-
sentations in both modalities. These findings support
the hypothesis that crossmodal links in multimodal WM
tasks specifically affect the mechanisms that control the
selective attentional activation of representations that
are already stored in WM.
GENERAL DISCUSSION
The question whether the processes involved in the atten-
tional control of WM are modality specific or shared across
sensory modalities remains the subject of considerable
debate (p.ej., Fougnie, Zughni, Godwin, & Marois, 2015;
Cowan, Saults, & Blume, 2014; Cowan, 2011; Fougnie
& Marois, 2011; Cocchini, Logie, Della Sala, MacPherson,
& Baddeley, 2002). In our own previous work (Katus &
Eimer, 2016, 2018, 2019b), we used ERP markers of concur-
rent visual and tactile WM maintenance processes and
found no evidence for crossmodal links. Visual CDA and
tCDA components did not differ between trials where to-
be-encoded visual and tactile sample stimuli were pre-
sented on the same versus opposite sides, and there were
also no differences in visual and tactile WM performance.
The absence of any crossmodal spatial congruency effects
suggests that the WM encoding and subsequent main-
tenance processes in vision and touch are mediated by
entirely modality-specific attentional control processes.
en este estudio, we investigated a different aspect of the
attentional control of WM. Using retro-cue procedures,
we tested whether there are crossmodal links between
processes that control the retrospective selection of vi-
sual and tactile WM representations. In Experiment 1,
we found clear evidence for crossmodal spatial congru-
ency effects. Tactile WM performance was impaired,
and tCDA components were strongly attenuated on trials
where retro-cues prompted attention shifts into opposite
directions within visual and tactile WM. Esto indica
that the control of such retrospective selection processes
is not fully modality specific but is based on mechanisms
that are at least partially shared across sensory modalities.
The results of Experiment 2 confirmed that these shared
mechanisms are specifically involved in the control of
concurrent attentional allocation processes in visual and
tactile WM. The behavioral and electrophysiological
spatial congruency effects observed for tactile WM in
Experimento 1 were eliminated when shifts of attention
within tactile WM triggered by retro-cues took place in
the absence of simultaneous shifts in visual WM, y esto
was confirmed by additional between-experiment analy-
ses.2 Although visual WM representations that had been
encoded previously were still selectively maintained at
the point in time when attention was shifted toward
retro-cued tactile WM representations, the spatial congru-
ency between these representations did not have any
systematic effect on tactile WM. This suggests that the
crossmodal spatial synergy effects found in Experiment 1
for tactile WM were not the generic result of a spatial
mismatch between simultaneously active visual and
tactile WM representations but were more specifically
associated with the control of retrospective selection
processes within WM. The absence of such effects in
Experimento 2 also underlines that the initial encoding
and storage of sensory stimuli and the subsequent selec-
tive activation of a subset of these stimuli are not just
temporally, but also functionally, dissociable (see also
miers, alimenta, & Nobre, 2017, for discussion of an anal-
ogous distinction between maintenance and retrospec-
tive selective activation processes in WM).
The current findings and the results of our previous
studies on the control of multimodal WM (Katus &
Eimer, 2016, 2018, 2019a, 2019b) indicate that encoding
and storage on the one hand and the attentional selec-
tion of existing WM representations on the other are
regulated by distinct attentional control mechanisms.
The former type of control appears to operate in a strictly
modality-specific fashion, whereas the latter is shared
across sensory modalities. If this was the case, the ob-
vious question is why such a difference in the modality
specificity of these two attentional control mechanisms
for WM should exist. Actualmente, we can only offer specula-
tions with regard to this question, rather than definitive
answers. One possibility is that this apparent dissociation
reflects a qualitative difference in the prospective versus
retrospective attentional control of WM. In situations
Katus and Eimer
555
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
.
/
F
t
oh
norte
0
5
METRO
a
y
2
0
2
1
where the spatial or nonspatial properties of to-be-
memorized events are known in advance, representaciones
of these properties (attentional templates; Duncan &
Humphreys, 1989) can be activated in a prospective fash-
ion before the onset of memory sample sets. These tem-
plates represent specific sensory attributes of task-relevant
sample stimuli and may therefore guide the selection and
encoding of these stimuli in a sensory-specific fashion. En
contrast, because no such preparatory attentional tem-
plates are involved when relevant representations in WM
are selected retrospectively, these selection processes may
be mediated primarily by modality-unspecific control pro-
cesses. Another possibility is that the difference between
the control of WM encoding/storage and of retrospective
selection mechanisms within WM is related to the dis-
tinction between external versus internal attention (Chun,
Golomb, & Turk-Browne, 2011). External attention controls
the selective processing of sensory stimuli in the outside
world, and internal attention regulates the selective activa-
tion of internal representations held in different types of
memory stores. The selective processing of sensory stimuli
in sample displays and the encoding of these stimuli into
WM primarily involves external attention, whereas the selec-
tive activation of stored WM representations signaled by
retro-cues requires internal attention. Because the demands
and constraints on external and internal attention are
considerably different (see Myers et al., 2017, for further
discussion), this could be reflected in differences in the
underlying control processes. Por ejemplo, the main func-
tion of external attention is to rapidly detect and select task-
relevant sensory objects and to exclude irrelevant objects
from further processing. This is believed to be achieved
by “winner-takes-all” competitive interactions between sen-
sory objects that are modulated by top–down task goals
(p.ej., Desimone & Duncan, 1995). Because such biased
competition processes in external attention are imple-
mented in modality-specific sensory-perceptual areas, es
plausible to assume that they are controlled by modality-
specific mechanisms. A diferencia de, internal attention operates
within a much more limited number of currently stored
mental representations from different sensory modalities,
and the selective prioritization of some of these repre-
sentations does not necessarily involve the loss of others
(p.ej., Lewis-Peacock, Drysdale, Oberauer, & Postle, 2012).
The allocation of attention to such internal WM repre-
sentations might therefore be mediated by higher-level
control processes in pFC that are shared and coordinated
across sensory modalities.
Although these suggestions are in line with the
results of the current visual/tactile WM experiments,
they cannot easily be reconciled with previous evidence
for crossmodal links in perceptual attention. In these
estudios, external attention was allocated to sensory stim-
uli in different modalities, and target-defining attributes
were known in advance (p.ej., Eimer & Conductor, 2001;
Spence & Conductor, 1996). Further research is clearly needed
to further dissociate the aspects of WM that are con-
trolled by modality-specific versus modality-nonspecific
mechanism.
Conclusión
The current study was the first to use retro-cue proce-
dures in a bimodal WM task to investigate the concurrent
allocation of spatial attention to representations stored in
visual and tactile WM. We found behavioral and electro-
physiological crossmodal spatial synergy effects for such
attention shifts within WM, indicating that the underlying
control mechanisms are shared across sensory modali-
corbatas. These supramodal mechanisms appear to be specific
to the control of attentional selection processes within
W.M., whereas other WM functions are regulated by
modality-specific processes.
Expresiones de gratitud
This work was funded by the Leverhulme Trust (grant RPG-2015-
370). We thank Laura Katus for proofreading the manuscript.
Reprint requests should be sent to Tobias Katus, School of
Psicología, University of Aberdeen, AB24 3FX Aberdeen,
Reino Unido, o por correo electrónico: t.katus@abdn.ac.uk.
Notas
1. As expected on the basis of previous observations (van Ede,
Chekroud, & Nobre, 2019), the retro-cues employed in the pres-
ent experiments triggered small but systematic deviations of eye
gaze toward the side where cued task-relevant visual sample
stimuli had been presented. Sin embargo, an analysis of mean
HEOG amplitudes between 350 y 1300 msec after the retro-
cue showed that this effect only reached statistical significance
in same-side trials of Experiment 1 (t(15) = 2.824, pag = .013,
BF10 = 4.455), but not for opposite-sides trials, and neither for
same-side nor opposite-sides trials in Experiment 2 (all ps >
.05). En tono rimbombante, a mixed ANOVA across both experiments
found no reliable differences in the size of these residual eye
movements between experiments (F(1, 30) = 0.037, pag = .850,
BF01 = 3.145) and no reliable interaction between Experi-
ment × Spatial Congruency (F(1, 30) = 0.738, pag = .397, BF01 =
2.217); the main effect of Spatial Congruency did also not
reach the significance threshold (F(1, 30) = 3.900, pag = .058,
BF10 = 1.672). Analogous results were obtained when this
analysis was conducted for HEOG data that had not been
corrected using independent component analysis. De este modo, cualquier
tendency to move the eyes toward cued visual sample loca-
tions is unlikely to account for the presence of Spatial Con-
gruency effects on tCDA amplitudes in Experiment 1 pero no
in Experiment 2.
2.
It should be noted that tactile WM accuracy and tCDA am-
plitudes were numerically enhanced on same-side trials and
reduced on opposite-sides trials in Experiment 1 as compared
to Experiment 2, suggesting that shifts of attention in visual WM
triggered by retro-cues in Experiment 1 produced both costs
and benefits for concurrent attention shifts in tactile WM on
opposite-sides and same-side trials, respectivamente. Sin embargo, di-
rect comparisons of tactile WM performance and tCDA ampli-
tudes between Experiments 1 y 2, conducted separately for
same-side and opposite-sides trials, yielded no reliable differ-
ences (all t(30) < 1.55, all p > .13).
556
Revista de neurociencia cognitiva
Volumen 32, Número 3
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
.
/
F
t
oh
norte
0
5
METRO
a
y
2
0
2
1
REFERENCIAS
Ahmad, J., Swan, GRAMO., Bowman, h., Wyble, B., Nobre, A. C.,
Shapiro, k. l., et al. (2017). Competitive interactions affect
working memory performance for both simultaneous and
sequential stimulus presentation. Informes Científicos, 7, 4785.
Awh, MI., & En Jonia, j. (2001). Overlapping mechanisms of
attention and spatial working memory. Tendencias en Cognitivo
Ciencias, 5, 119–126.
Awh, MI., Vogel, mi. K., & Oh, S.-H. (2006). Interactions between
attention and working memory. Neurociencia, 139, 201–208.
Baddeley, A. (2012). Working memory: Theories, modelos, y
controversies. Annual Review of Psychology, 63, 1–29.
Chun, METRO. METRO., Golomb, j. D., & Turk-Browne, norte. B. (2011). A
taxonomy of external and internal attention. Annual Review
of Psychology, 62, 73–101.
Cocchini, GRAMO., Logie, R. h., Della Sala, S., MacPherson, S. MI., &
Baddeley, A. D. (2002). Concurrent performance of two
tareas de memoria: Evidence for domain-specific working
memory systems. Memoria & Cognición, 30, 1086–1095.
Cowan, norte. (2011). The focus of attention as observed in visual
working memory tasks: Making sense of competing claims.
Neuropsicología, 49, 1401–1406.
Cowan, NORTE., Saults, j. S., & Blume, C. l. (2014). Central and
peripheral components of working memory storage. Diario
of Experimental Psychology: General, 143, 1806–1836.
Delorme, A., Sejnowski, T., & Makeig, S. (2007). Enhanced detection
of artifacts in EEG data using higher-order statistics and
independent component analysis. Neuroimagen, 34, 1443–1449.
Desimone, r., & Duncan, j. (1995). Neural mechanisms of selective
visual attention. Revisión anual de neurociencia, 18, 193–222.
Duncan, J., & Humphreys, GRAMO. W.. (1989). Visual search and
stimulus similarity. Revisión psicológica, 96, 433–458.
Eimer, METRO., & Conductor, j. (2001). Crossmodal links in endogenous
and exogenous spatial attention: Evidence from event-related
brain potential studies. Neurociencia & Biobehavioral
Reseñas, 25, 497–511.
Eimer, METRO., & Kiss, METRO. (2010). An electrophysiological measure
of access to representations in visual working memory.
Psychophysiology, 47, 197–200.
Eimer, METRO., van Velzen, J., & Conductor, j. (2002). Cross-modal
interactions between audition, tocar, and vision in
endogenous spatial attention: ERP evidence on preparatory
states and sensory modulations. Revista de Cognitivo
Neurociencia, 14, 254–271.
Emrich, S. METRO., & Ferber, S. (2012). Competition increases binding
errors in visual working memory. Revista de visión, 12, 12.
Fougnie, D., & Marois, R. (2011). What limits working memory
capacity? Evidence for modality-specific sources to the
simultaneous storage of visual and auditory arrays. Diario de
Psicología experimental: Aprendiendo, Memoria, y
Cognición, 37, 1329–1341.
Fougnie, D., Zughni, S., Godwin, D., & Marois, R. (2015).
Working memory storage is intrinsically domain specific.
Revista de Psicología Experimental: General, 144, 30–47.
Griffin, I. C., & Nobre, A. C. (2003). Orienting attention to
locations in internal representations. Revista de Cognitivo
Neurociencia, 15, 1176–1194.
JASP Team. (2016). JASP (Versión 0.7.5.5) [Computer software].
Retrieved from https://jasp-stats.org/.
Jeffreys, h. (1961). Theory of probability (3tercera ed.). Oxford
Prensa universitaria.
En Jonia, J., Lacey, S. C., & Nee, D. mi. (2005). Processes of
working memory in mind and brain. Direcciones actuales en
ciencia psicológica, 14, 2–5.
Katus, T., andersen, S. K., & Müller, METRO. METRO. (2012). Maintenance
of tactile short-term memory for locations is mediated by
spatial attention. Biological Psychology, 89, 39–46.
Katus, T., & Eimer, METRO. (2015). Lateralized delay period activity
marks the focus of spatial attention in working memory:
Evidence from somatosensory event-related brain potentials.
Revista de neurociencia, 35, 6689–6695.
Katus, T., & Eimer, METRO. (2016). Multiple foci of spatial attention
in multimodal working memory. Neuroimagen, 142, 583–589.
Katus, T., & Eimer, METRO. (2018). Independent attention
mechanisms control the activation of tactile and visual
working memory representations. Revista de Cognitivo
Neurociencia, 30, 644–655.
Katus, T., & Eimer, METRO. (2019a). The sources of dual-task costs in
multisensory working memory tasks. Revista de Cognitivo
Neurociencia, 31, 175–185.
Katus, T., & Eimer, METRO. (2019b). Shifts of spatial attention in
visual and tactile working memory are controlled by
independent modality-specific mechanisms. Cerebral
Corteza. https://doi.org/10.1093/cercor/bhz088.
Katus, T., Grubert, A., & Eimer, METRO. (2015). Electrophysiological
evidence for a sensory recruitment model of somatosensory
working memory. Corteza cerebral, 25, 4697–4703.
Katus, T., Grubert, A., & Eimer, METRO. (2017). Intermodal
attention shifts in multimodal working memory. Diario de
Neurociencia Cognitiva, 29, 628–636.
Katus, T., Müller, METRO. METRO., & Eimer, METRO. (2015). Sostenido
maintenance of somatotopic information in brain regions
recruited by tactile working memory. Diario de
Neurociencia, 35, 1390–1395.
kuo, B.-C., alimenta, METRO. GRAMO., & Nobre, A. C. (2012). Atención
modulates maintenance of representations in visual short-
term memory. Revista de neurociencia cognitiva, 24, 51–60.
Lepsien, J., & Nobre, A. C. (2006). Cognitive control of attention
en el cerebro humano: Insights from orienting attention to
mental representations. Brain Research, 1105, 20–31.
Lewis-Peacock, j. A., Drysdale, A. T., Oberauer, K., & Postle,
B. R. (2012). Neural evidence for a distinction between short-
term memory and the focus of attention. Diario de
Neurociencia Cognitiva, 24, 61–79.
McCants, C. w., Katus, T., & Eimer, METRO. (2020). Task goals
modulate the activation of part-based versus object-based
representations in visual working memory. Cognitivo
Neurociencia, 11, 92–100.
miers, norte. MI., alimenta, METRO. GRAMO., & Nobre, A. C. (2017). Prioritizing
information during working memory: Beyond sustained
internal attention. Tendencias en Ciencias Cognitivas, 21, 449–461.
Nolan, h., Whelan, r., & Reilly, R. B. (2010). FASTER: Fully
automated statistical thresholding for EEG artifact rejection.
Journal of Neuroscience Methods, 192, 152–162.
Postle, B. R. (2006). Working memory as an emergent property
of the mind and brain. Neurociencia, 139, 23–38.
Robertson, l. C. (2003). Binding, spatial attention and perceptual
awareness. Naturaleza Reseñas Neurociencia, 4, 93–102.
Rojo, j. NORTE., Speckman, PAG. l., Sol, D., Morey, R. D., &
Iverson, GRAMO. (2009). Bayesian t tests for accepting and
rejecting the null hypothesis. Boletín psiconómico &
Revisar, 16, 225–237.
Spence, C., & Conductor, j. (1996). Audiovisual links in endogenous
covert spatial attention. Revista de Psicología Experimental:
Percepción y desempeño humanos, 22, 1005–1030.
Tenke, C. MI., & Kayser, j. (2012). Generator localization by
current source density (CSD): Implications of volume
conduction and field closure at intracranial and scalp
resolutions. Clinical Neurophysiology, 123, 2328–2345.
van Ede, F., Chekroud, S. r., & Nobre, A. C. (2019). Humano
gaze tracks attentional focusing in memorized visual space.
Nature Human Behavior, 3, 462–470.
Vogel, mi. K., & Machizawa, METRO. GRAMO. (2004). Neural activity predicts
individual differences in visual working memory capacity.
Naturaleza, 428, 748–751.
Katus and Eimer
557
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
norte
t
pag
oh
:
a
/
d
/
mi
d
metro
i
F
r
t
oh
pag
metro
r
C
h
.
pag
s
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
2
C
yo
3
mi
5
–
4
pag
6
d
F
2
0
/
1
3
3
2
1
/
3
3
6
/
5
oh
4
C
6
norte
/
_
a
1
_
8
0
6
1
1
4
0
9
7
2
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
2
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
t
F
/
.
oh
norte
0
5
METRO
a
y
2
0
2
1