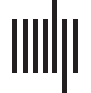ARTÍCULO DE INVESTIGACIÓN
Grammatical Parallelism in Aphasia:
A Lesion-Symptom Mapping Study
William Matchin1
, Dirk-Bart den Ouden1
, Alexandra Basilakos1
,
Brielle Caserta Stark2
, Julius Fridriksson1
, and Gregory Hickok3
1Department of Communication Sciences and Disorders, University of South Carolina, Columbia, CAROLINA DEL SUR, EE.UU
2Department of Speech, Language and Hearing Sciences, Program for Neuroscience,
Indiana University Bloomington, Bloomington, EN, EE.UU
3Department of Cognitive Sciences, Department of Language Science, Universidad de California, Irvine, Irvine, California, EE.UU
Palabras clave: syntax, aphasia, agrammatism, paragrammatism, lesion-symptom mapping
ABSTRACTO
Sentence structure, or syntax, is potentially a uniquely creative aspect of the human mind.
Neuropsychological experiments in the 1970s suggested parallel syntactic production and
comprehension deficits in agrammatic Broca’s aphasia, thought to result from damage to
syntactic mechanisms in Broca’s area in the left frontal lobe. This hypothesis was
sometimes termed overarching agrammatism, converging with developments in linguistic
theory concerning central syntactic mechanisms supporting language production and
comprensión. Sin embargo, the evidence supporting an association between receptive
syntactic deficits, expressive agrammatism, and damage to frontal cortex is equivocal. En
addition, the relationship between a distinct grammatical production deficit in aphasia,
paragrammatism, and receptive syntax has not been assessed. We used lesion-symptom
mapping in three partially overlapping groups of left-hemisphere stroke patients to
investigate these issues: grammatical production deficits in a primary group of 53 subjects
and syntactic comprehension in larger sample sizes (norte = 130, 218) that overlapped with
the primary group. Paragrammatic production deficits were significantly associated with
multiple analyses of syntactic comprehension, particularly when incorporating lesion
volume as a covariate, but agrammatic production deficits were not. The lesion correlates
of impaired performance of syntactic comprehension were significantly associated with
damage to temporal lobe regions, which were also implicated in paragrammatism, pero no
with the inferior and middle frontal regions implicated in expressive agrammatism. Nuestro
results provide strong evidence against the overarching agrammatism hypothesis. Por
contrast, our results suggest the possibility of an alternative grammatical parallelism
hypothesis rooted in paragrammatism and a central syntactic system in the posterior
temporal lobe.
INTRODUCCIÓN
Agrammatism and Syntactic Parallelism in Aphasia
Syntax, or the ability to combine words into hierarchical structures, enables a core component of
human linguistic creativity: the ability to make novel sentences of unbounded size and number
(Chomsky, 1965; von Humboldt, 1836). The nature of syntactic deficits in aphasia and the role
of Broca’s area in such deficits has a long and complicated history. From the beginning of the
un acceso abierto
diario
Citación: Matchin, w., den Ouden, D.-B.,
Basilakos, A., Rígido, B. C., Fridriksson,
J., & Hickok, GRAMO. (2023). Grammatical
parallelism in aphasia: A lesion-
symptom mapping study. Neurobiología
of Language. Publicación anticipada.
https://doi.org/10.1162/nol_a_00117
DOI:
https://doi.org/10.1162/nol_a_00117
Supporting Information:
https://doi.org/10.1162/nol_a_00117
Recibió: 3 Noviembre 2022
Aceptado: 19 Julio 2023
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
William Matchin
matchin@mailbox.sc.edu
Editor de manejo:
Jenny Crinion
Derechos de autor: © 2023
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
.
/
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
Syntax:
The structure of sentences that
supports combinatorial and creative
use of language.
Agrammatism:
A language production disorder
involving the omission of functional
elementos, the overall simplification of
sentence structure, and telegraphic
speech.
20th century until the 1970s, agrammatism in people with nonfluent Broca’s aphasia was
defined as the systematic reduction of syntactic complexity and omission of functional elements
(such as auxiliary verbs and articles) in speech production; syntactic comprehension was
assumed to be intact (Forster, 1919; Isserlin, 1922). This was consistent with the classical model
of language in the brain espoused by Wernicke, Lichtheim
, and later Geschwind, positing that
frontal damage, which was linked to Broca’s aphasia, produced expressive language deficits in
the absence of notable receptive deficits (Geschwind, 1970, 1972, 1979; Wernicke, 1874).
The received view of impaired production but preserved comprehension in agrammatism
was questioned in the 1970s, when some studies (typically with small numbers of participants
and an absence of detailed lesion analysis) revealed apparent syntactic comprehension deficits
in people with agrammatic Broca’s aphasia (Caramazza & Zurif, 1976; Zurif et al., 1972;
patients with conduction aphasia, without agrammatic production deficits, showed the same
comprehension pattern, a fact rarely discussed in current literature). A highly influential study
by Caramazza and Zurif (1976) assessed the syntactic comprehension abilities of English-
speaking people with agrammatic Broca’s aphasia using semantically reversible sentences
with noncanonical word order. In both (1) y (2), abajo, the sentences have a noncanonical
word order, in which the initial noun phrase and participant of the main clause, the man in (1)
and the apple in (2), are the object of the verbs in the embedded clauses. Reversible sentences
consisted of those in which the doer and receiver of the action could switch positions while
the sentence still made sense (1), as opposed to semantically constrained sentences which
would be thematically impossible if the doer and receiver were switched (2).
1.
2.
The man that the woman is hugging is happy (reversible, noncanonical word order,
poor performance)
The apple that the boy is eating is red (semantically constrained, noncanonical word
orden, good performance)
This study showed that comprehension of semantically constrained sentences with
noncanonical word order (2) was good, but that comprehension of reversible sentences with
noncanonical word order (1) was poor. Caramazza and Zurif (1976) argued that people
with agrammatic Broca’s aphasia lacked normal syntactic ability not only for production but
also for comprehension, explaining the selective pattern of comprehension deficits. This pattern
could be explained via the preservation of word-level understanding and interpretive heuristics
based on semantic plausibility and assuming the first noun to be the agent of the action.
Such results shifted the standard assumption in aphasiology toward syntactic parallelism:
the idea that syntactic comprehension deficits accompanied syntactic production deficits in
agrammatic Broca’s aphasia (Berndt & Caramazza, 1980; Kean, 1977; Saffran et al., 1980;
Schwartz et al., 1980; Zurif, 1980). The syntactic parallelism hypothesis was sometimes
termed overarching agrammatism (Grodzinsky, 2000a; Swinney & Zurif, 1995), emphasizing
that the comprehension and production deficits in agrammatism resulted from disruption to a
common underlying central syntactic mechanism supporting both the production and compre-
hension of language (Chomsky, 1965, 1980). This represented a major shift in thinking regard-
ing aphasiology and the neurobiology of language. Instead of talking about language deficits
and neurological models in terms of production or comprehension functions, the vocabulary
of linguistic theory regarding central functions of syntax (assumed to be localized to Broca’s
área, or the left posterior inferior frontal gyrus) and semantics (assumed to be localized to
left posterior temporal cortex) was used to define the language–brain relationship (see also
Jakobson, 1956, and Jakobson & Halle, 1956, for early views on the application of linguistics
Neurobiology of Language
2
Q1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
.
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
to aphasiology). This was in part based on the assumed relationship between Broca’s aphasia
and Broca’s area, although we note that Broca’s aphasia may be caused by lesions not involv-
ing Broca’s area (Fridriksson et al., 2007), and damage to Broca’s area alone is insufficient to
cause Broca’s aphasia (Fridriksson et al., 2015; Gajardo-Vidal et al., 2021; Mohr et al., 1978).
The seemingly tight convergence between linguistic theory and aphasiology that this work
pointed toward began to unravel with demonstrations of intact receptive syntactic abilities in
people with agrammatic Broca’s aphasia in the 1980s. Específicamente, syntactic acceptability
judgments, subtle and direct tests of syntactic ability, were shown to be mostly intact in people
with Broca’s aphasia and expressive agrammatism (Linebarger et al., 1983; wilson & Saygın,
2004; Wulfeck & Bates, 1991). These authors proposed instead that the relationship between
the syntactic mechanisms identified in linguistic theory and language deficits in aphasia was
much more indirect, with agrammatic comprehension deficits in aphasia reflecting the impaired
ability to make use of intact syntactic representations for semantic interpretation. Some authors
sought to preserve aspects of the overarching agrammatism hypothesis by positing a more
restricted syntactic deficit, one tied to mechanisms devoted to the movement of sentence
elementos (Chomsky, 1981) that explained difficulties with noncanonical sentences (Grodzinsky,
1986; Hickok et al., 1993). Sin embargo, by the 1990s, many researchers in aphasiology had
abandoned the syntactic parallelism hypothesis (Goodglass et al., 1993; Hickok & Avrutin,
1995; Kean, 1995; Swinney & Zurif, 1995; para una revisión, see Matchin & Rogalsky, 2023).
Despite this, the hypothesis of overarching agrammatism never fully disappeared, and many
researchers have continued to focus on nonfluent aphasia patients, primarily those with Broca’s
aphasia and/or expressive agrammatism, and have continued to use noncanonical sentence
structures as a diagnostic tool for receptive syntax (Cho-Reyes & Thompson, 2012; Grodzinsky,
2000b; Grodzinsky & Finkel, 1998; Mesulam et al., 2015; Santi & Grodzinsky, 2007;
Thompson et al., 1997, 2013; Thompson & Shapiro, 2005; Tyler et al., 2011; wilson, Dronkers,
et al., 2010). Además, the arrival of functional neuroimaging in the 1990s breathed new life
into the overarching agrammatism hypothesis, with several studies of syntactic comprehension
showing effects in Broca’s area (Dapretto & Bookheimer, 1999; Embick et al., 2000; Friederici
et al., 2000; Moro et al., 2001; Stromswold et al., 1996). Finalmente, research on the neural basis of
word-level processes (lexical access) demonstrated associations within the left posterior tem-
poral lobe (Dronkers et al., 2004; Hickok & Poeppel, 2000; Lau et al., 2008; Levelt, 2001;
Levelt et al., 1999; Pinker & Ullman, 2002). Juntos, these findings reinforced a framework
for language in the brain that posited a syntactic computation function in Broca’s area and a
lexical storage function in left posterior temporal cortex (Friederici, 2002; Hagoort, 2005),
aligning two core aspects of language with the two classical language regions. De este modo, a pesar de
striking contradictory evidence from aphasiology, the overarching agrammatism hypothesis
and the concept of a central syntactic hub in Broca’s area continues to have substantial influ-
ence in psychology, linguistics, and neuroscience (Arbib, 2016; Bozic et al., 2015; D’Ausilio
et al., 2012; Friederici, 2017; Hagoort, 2013, 2016; Hagoort & Indefrey, 2014; Kuperberg,
2007; Menenti et al., 2011; Momma & Phillips, 2018; Ocampo & Kritikos, 2011; Pulvermüller
& Fadiga, 2010; Rilling, 2014; Tyler et al., 2010; wilson, Dronkers, et al., 2010).
Paragrammatism
Paragrammatism:
A language production disorder
involving the misuse of functional
elements and structures, without an
overall tendency toward omission or
reducción.
Agrammatism is not the only grammatical production deficit in aphasia. In the early 1900s,
clinicians described an expressive disorder termed paragrammatism, which is associated with
fluent aphasia and characterized by grammatical distortion but without the overall
reduction/simplification that is characteristic of agrammatism (Bonhoeffer, 1902; Butterworth
& Howard, 1987; Goodglass et al., 1993; Heeschen, 1985; Kleist, 1914). This disorder has
Neurobiology of Language
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
/
.
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
Lesion-symptom mapping:
A statistical technique that allows
researchers to associate patterns of
brain damage with behavioral
actuación.
received vanishingly little attention in the last decades. In recent work, Matchin et al. (2020)
found a double dissociation in the lesion distributions associated with agrammatism versus
paragrammatism: Agrammatism was associated with inferior and middle frontal damage
including Broca’s area, but not left temporal cortex, consistent with several previous studies
(den Ouden et al., 2019; Sapolsky et al., 2010; Wilson et al., 2011; wilson, Henry, et al.,
2010). Por el contrario, paragrammatism was associated with damage to the posterior temporal
corteza, but not frontal cortex, consistent with case studies (Yagata et al., 2017). Reciente
lesion-symptom mapping (LSM) and connectome-based LSM studies have identified a robust
association between syntactic comprehension deficits and damage to the middle posterior
temporal lobe, with minimal or no implication of damage to or disconnection of Broca’s area,
assessed via noncanonical sentence comprehension (Billot et al., 2022; den Ouden et al.,
2019; Dronkers et al., 2004; Kristinsson et al., 2020; Lukic et al., 2021; Lwi et al., 2021;
Matchin, Basilakos, et al., 2022; Matchin, den Ouden, et al., 2022; Rogalsky et al., 2018),
syntactic acceptability judgments (wilson & Saygın, 2004), and sentence comprehension with
word production ability as a covariate (Pillay et al., 2017; cf. Adezati et al., 2022, which found
that sentence comprehension deficits were associated with damage to both regions). Esta sugerencia-
gests that parallel grammatical deficits in aphasia (rooted in paragrammatism, en vez de
agrammatism) may result from common injury to the posterior temporal lobe. En efecto, alguno
studies assessing syntactic production deficits measured using the Northwestern Assessment of
Verbs and Sentences (NAVS) sentence priming production task have found that superior tem-
poral lobe damage, and not frontal damage, is associated with production deficits (den Ouden
et al., 2019; Lukic et al., 2021), although we note that this test critically involves correct com-
prehension to perform the task, conflating these results with comprehension ability.
Hypotheses and Predictions
Recent functional neuroimaging studies often find syntax-related activations in the inferior
frontal (typically along with the posterior temporal) lobe (Blank et al., 2016; Fedorenko
et al., 2016; Matchin, Liao, et al., 2019; Nelson et al., 2017; Zaccarella, Meyer, et al.,
2017; cf. several recent studies that highlight temporal and not frontal lobe, Brennan et al.,
2016; Flick & Pylkkänen, 2020; Matar et al., 2021; Matchin, Brodbeck, et al., 2019; Matchin,
İlkbaşaran, et al., 2022; Murphy et al., 2022; Stanojević et al., 2023). While these studies have
long suggested to researchers that frontal lobe structures support syntactic processing, the cor-
relational nature of functional imaging precludes an assessment of the extent to which these
structures are causally implicated. Lesion studies, por el contrario, can provide critical evidence
regarding the causal role of these regions (Rorden & Karnath, 2004). Respectivamente, many of
the aphasiological studies
reviewed above have concluded that frontal structures are causally
implicated in receptive syntax, in accordance with the “overarching agrammatism” hypothe-
hermana. Sin embargo, this question of syntactic parallelism, rooted in agrammatism, has not been revis-
ited and comprehensively addressed using modern LSM methods (cf. den Ouden et al., 2019;
Lukic et al., 2021, which assessed both production and comprehension and only found tem-
poral lobe damage associated with objective quantitative assessments. Den Ouden et al. found
that frontal damage was associated with expressive agrammatism as identified with perceptual
análisis, but did analyze the relationship of these categorizations with syntactic comprehen-
sion variables). Nor has a syntactic parallelism hypothesis rooted in paragrammatism been
investigated.
Here we systematically (re)assess these issues in three partially overlapping groups of peo-
ple with post-stroke aphasia. In order for syntactic parallelism to hold, the association between
syntactic comprehension and production deficits should be robust to various measures and
Neurobiology of Language
4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
.
/
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Q2
Grammatical parallelism in aphasia
analiza. De este modo, we assessed syntactic comprehension ability in a variety of ways. In the liter-
ature, different approaches have been used, with some authors using sentence comprehension
measures that implicate syntactic abilities that minimize lexical demands (Dronkers et al.,
2004; Gorno-Tempini et al., 2004; Matchin, den Ouden et al., 2022; Mesulam et al., 2015;
Thompson et al., 2013; wilson, Dronkers, et al., 2010), and others combining such measures
with covariates in order to more purely isolate syntactic abilities (Kristinsson et al., 2020;
Magnusdottir et al., 2013; Matchin, Basilakos, et al., 2022; Pillay et al., 2017; Rogalsky
et al., 2018). We adopted both approaches here, using two different sentence comprehension
measures to assess receptive syntax: the Sequential Commands subtest of the Western Aphasia
Battery—Revised ( WAB-R; Kertesz, 2006) and the comprehension of noncanonical sentence
estructuras (Cho-Reyes & Thompson, 2012; Magnusdottir, 2005), each with or without a covar-
iate to attempt to control for lexical-semantic processing, for a total of four analyses of syntac-
tic comprehension. The overarching agrammatism hypothesis predicts that both expressive
agrammatism and syntactic comprehension deficits follow from lesions to the same parts of
the frontal lobe. The syntactic parallelism hypothesis rooted in paragrammatism predicts that
both expressive paragrammatism and syntactic comprehension deficits will follow from
lesions to the same parts of the posterior temporal lobe. We expected that our results would
speak against the overarching agrammatism view and instead support the overarching para-
grammatism view, given previous suggestive neuropsychological data of intact syntactic com-
prehension in agrammatism and previous associations of syntactic comprehension deficits
with posterior temporal lobe lesions.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
Sequential Commands:
A subtest of the WAB-R that involves
asking the participant to perform
increasingly complex actions that
require comprehending sentence
estructura.
Western Aphasia Battery—Revised
( WAB-R):
A standard battery of language
comprensión, producción, y
repetition abilities; supports aphasia
classification and severity
evaluación.
yo
/
/
.
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
MATERIALES Y MÉTODOS
Participantes
Participants were drawn from a database of individuals with chronic, post-stroke aphasia who
have completed testing for various studies conducted at the University of South Carolina and
the Medical University of South Carolina over the last 15 años. All were native speakers of
Inglés, had suffered an ischemic stroke to the left hemisphere at least six months prior to the
estudiar, and presented with language difficulties (most participants were classified as aphasic
according to the WAB-R [Kertesz, 2006]; sin embargo, some scored outside of the aphasic range
by the time of examination. All participants had presented with aphasia in the acute phase
following their stroke, which formed the basis of their enrollment in the study). We performed
retrospective analyses in three overlapping groups of stroke survivors. Group 1 consistió en 53
Participantes (all of whom were included in Group 3 y 42 of whom were included in Group
2) who were perceptually assessed for agrammatism and paragrammatism. Group 2 consisted
of a subset of 130 participants who were assessed on one of two similar tests of sentence com-
prehension involving canonical and noncanonical sentence structures. Group 3 consistió en
218 participants who were assessed on the WAB-R. Mesa 1 provides demographic, lesion vol-
ume, and aphasia severity for each of the three groups (demographic information broken down
by perceptual ratings of AGRAMMATISM and PARAGRAMMATISM can be found in Table 1 of Matchin
et al., 2020). All procedures were approved by the internal review boards at each institution
and informed consent was obtained.
Measures and Procedure
Todo 218 participants were evaluated using the WAB-R (Kertesz, 2006) to determine the pres-
ence and severity of aphasia. The test was administered and scored by certified speech-
language pathologists with extensive experience evaluating individuals with aphasia. El
Neurobiology of Language
5
Grammatical parallelism in aphasia
Mesa 1.
Participant information for the three partially overlapping groups of participants.
Tasks
Group 1
Perceptual ratings of AGRAMMATISM
and PARAGRAMMATISM
Group 2
Noncanonical sentence
comprensión
Group 3
Western Aphasia Battery—
Revised ( WAB-R)
Number of participants
53
in group
Sex
130 (42 participants from
218 (all participants from
Group 1 incluido)
Groups 1 y 2 incluido)
35 masculino, 18 femenino
83 masculino, 47 femenino
133 masculino, 85 femenino
Pre-stroke handedness
45 right-handed, 2 left-handed,
6 unknown
108 right-handed, 8 izquierda-
handed, 14 unknown
178 right-handed, 9 izquierda-
handed, 31 unknown
Mean age at testing (años)
58.9 (DE = 12.2)
60.0 (DE = 10.7)
60.0 (DE = 11.4)
Mean months post-stroke
48.6 (DE = 53.5)
45.3 (DE = 50.4)
43.0 (DE = 48.4)
at initial testing
Mean education (años)
15.8 (DE = 2.3)
15.4 (DE = 2.4), *norte = 128
15.0 (DE = 2.3), *norte = 210
Mean lesion volume (mm3)
125,102 (DE = 85,490)
111,267 (DE = 92,645)
120,855 (DE = 97,488)
Mean WAB-R AQ
68.2 (DE = 16.7)
65.3 (DE = 26.9)
61.4 (DE = 28.1)
Nota. SD = standard deviation. AQ = aphasia quotient of the WAB-R, a summary measure of overall language ability, con 0 being the most severe score. El
WAB-R denotes that an AQ of <93.8 denotes presence of clinical aphasia.
* Education information was not available for all participants, the number for which education information indicated here.
Auditory Word Recognition:
A subtest of the WAB-R that involves
asking the participant to identify
common everyday objects after
hearing their labels.
WAB-R contains multiple subtests to evaluate production and comprehension ability. Here we
focus on the Sequential Commands and the Auditory Word Recognition subtests. The Sequen-
tial Commands subtest consists of 11 sentential instructions for actions to perform. It requires
the participant to process basic syntactic relations indicated with prepositional phrase modi-
fiers and connectives, while minimizing lexical demands (using repeated high frequency
nouns referring to common objects located in the testing area). A previous study also used
the Sequential Commands subtest of the WAB-R to assess syntactic comprehension (Gorno-
Tempini et al., 2004), but did so in conjunction with additional measures including complex
sentences with noncanonical word order. While some of the commands can be performed
correctly without syntactic analysis, relying on lexical comprehension alone (e.g., raise your
hand ), the bulk of the total score requires analyzing both the lexical items and their syntactic
arrangement to perform correctly (e.g., point to the comb with the pen). Indeed, Schwartz et al.
(1980) report that people with agrammatic Broca’s aphasia have difficulty comprehending sen-
tences of this type (recall that having Broca’s aphasia does not necessarily imply that damage
to Broca’s area is the cause of the deficits, Fridriksson et al., 2015). Participants can
receive
partial credit for correctly performing a subset of the actions indicated in a command; full
credit required performing all of the indicated actions in the correct order. We used raw scores
on this subtest for our sequential commands measure.
To control for lexical-semantic processing, in additional analyses we incorporated the Audi-
tory Word Recognition subtest as a covariate with the sequential commands measure to pro-
duce the SEQUENTIAL COMMANDSAUDWORDS analysis. This subtest involves asking the participant to
point to real-world objects or printed images as requested. Some of these objects are contained
within the Sequential Commands subtest. Participants are prompted with a sentence, for exam-
ple, “point to the __” or “show me the __”. The test involves multiple types of tested words,
Neurobiology of Language
6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Q3
Grammatical parallelism in aphasia
Noncanonical sentence
comprehension:
A standard metric of syntactic
comprehension abilities, involving
sentences with different word order
than is typically expected within the
language.
including real household objects (e.g., cup, comb), pictured objects (the same as real objects),
pictured shapes (e.g., square, circle), pictured letters (e.g., J, F), pictured numbers (e.g., 5, 61),
pictured colors (e.g., blue, red), real world furniture (e.g., window, chair), real world body parts
(e.g., ear, nose), real world fingers (e.g., thumb, index finger), and real world body parts on the
correct side (e.g., right ear, left knee). For each item the participant receives 1 point, for a total
of 60 points. Thus, for our SEQUENTIAL COMMANDSAUDWORDS analysis we assessed SEQUENTIAL COM-
MANDS incorporating the Auditory Word Recognition subtest as a covariate.
We included two additional analyses of syntactic comprehension, NONCANONICAL and NON-
CANONICALACTIVE, based on the standard approach to syntactic comprehension in aphasia cen-
tered around noncanonical sentence structures introduced by Caramazza and Zurif (1976)
and developed since then by many authors (Caplan et al., 1996; Cho-Reyes & Thompson,
2012; Dronkers et al., 2004; Magnusdottir, 2005; Magnusdottir et al., 2013). We used data
that has been previously reported in LSM analyses of these tasks in 130 people with chronic
stoke-based aphasia, one study reporting the lesion correlates of noncanonical sentence com-
prehension combined with active sentence comprehension as a covariate (Matchin, Basilakos,
et al., 2022), and one study reporting the lesion correlates of noncanonical sentence compre-
hension by itself (Matchin, den Ouden, et al., 2022). Some of these same participants have
also been reported in another study (den Ouden et al., 2019).
The NONCANONICAL measure was derived from the performance on noncanonical structures
from the NAVS (82 participants; Cho-Reyes & Thompson, 2012) or the Test of Syntax (ToS; 48
participants; Magnusdottir, 2005; Magnusdottir et al., 2013). The NAVS involves testing the
comprehension of a variety of canonical and noncanonical sentence types, each with five total
trials, assessed via pointing to the correct picture. The performance on the three noncanonical
sentence types of the NAVS combined has been used as an index of syntactic comprehension:
passives (with a by-phrase) e.g., the dog is chased by the cat, object-extracted WH-questions,
e.g., who is the cat chasing?, and object-relatives, e.g., Pete saw the boy who the girl is pulling,
for a maximum score of 15 (den Ouden et al., 2019; Mesulam et al., 2015; Thompson et al.,
2013), a procedure similar to that used with other tests. The ToS (Magnusdottir, 2005;
Magnusdottir et al., 2013) includes a similar set of canonical and noncanonical sentence
types, each with five total trials: passives (with a by-phrase), for example, “The boy is painted
by the girl”; object-extracted WH- questions, for example, “Which boy is the girl painting?”;
and object clefts, for example, “It is the girl that the boy paints.” The sentence types across the
two tasks are not strictly identical, but involve essentially the same structures with the same
degree of complexity, including the key factor of noncanonical object-first word order. There-
fore, for participants who were not assessed with the NAVS, we calculated the equivalent
scores on the ToS (correct noncanonical trials, out of 15 points). This score, correct noncanon-
ical trials out of 15, comprised the NONCANONICAL measure. The NONCANONICALACTIVE analysis was
derived by performing a linear regression on the noncanonical score, including performance
on simple active sentences as a covariate (ACTIVE SENTENCE COMPREHENSION, out of 5 points), which
was included in both the NAVS and ToS, to control for lexical-semantic processing.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Agrammatism/Paragrammatism Ratings
For our measures of syntactic production deficits, AGRAMMATISM and PARAGRAMMATISM, we used
the data from 53 participants reported in Matchin et al. (2020): categorical perceptual ratings,
formed as a consensus of four expert raters based on the patients’ unconstrained retelling of
the Cinderella Story in their own words, following the AphasiaBank protocol (MacWhinney
et al., 2011
). Full details of participant selection, testing, and evaluation are provided in
Q4
Neurobiology of Language
7
Grammatical parallelism in aphasia
Matchin et al. (2020). Briefly, 100 people with chronic stroke-based aphasia, who partook in
a broader study of aphasia recovery, were recorded as they performed the Cinderella Story
retelling task from the AphasiaBank protocol (MacWhinney et al., 2011). Recordings ranged
from a few seconds to several minutes. Four expert raters, blind to any information about
each participant other than their recording, watched the recordings as many times as needed
and rated each participant as AGRAMMATIC (11 participants), PARAGRAMMATIC (21 participants), BOTH
AGRAMMATIC and PARAGRAMMATIC (4 participants), or no grammatical deficit (17 participants).
Following this, a discussion was held among all four raters to develop a single consensus rating
for each participant. 47 participants were excluded due to severely limited speech output or
unintelligibility (45 participants) and poor audio recording quality (2 participants). The four
patients who were identified as exhibiting some features of both classifications were included
in analyses of both of these measures; thus, we had a total of 15 AGRAMMATIC participants and
25 PARAGRAMMATIC participants. Only 42 of these original 53 participants also performed the
canonical/noncanonical sentence comprehension tasks, thus the numbers for each subgroup
are as follows: AGRAMMATIC (7 participants), PARAGRAMMATIC (16 participants), no grammatical
deficit (15 participants), or both AGRAMMATIC and PARAGRAMMATIC (4 participants). Throughout
the paper, we express the general concepts discussed in the literature of agrammatism (and
agrammatic speech) and paragrammatism (and paragrammatic speech) using regular type-
face. We express the corresponding perceptual classification of these concepts as applied
to our participant groups using small capital typeface, that is, AGRAMMATISM/AGRAMMATIC and
PARAGRAMMATISM/PARAGRAMMATIC, and correspondingly, NOT-AGRAMMATIC and NOT-PARAGRAMMATIC
for patients that were classified as not having these grammatical production deficits.
Neuroimaging and Lesion Mapping
High-resolution magnetic resonance imaging (MRI) data (T1- and T2-weighted images) were
collected at University of South Carolina and the Medical University of South Carolina on
a 3T Siemens Trio scanner with a 12-element head coil. T1-weighted MRI images were
collected using an MP-RAGE sequence, voxel dimensions 1 mm3, 256 × 256 matrix, 9°
flip angle, TR 2,250 ms, either 160 slices with inversion time of 900 ms and echo time
of 4.52 ms, or 192 slices with inversion time of 925 ms and TE of 4.15 ms with parallel
imaging (GRAPPA = 2, 80 reference lines). T2-weighted MRI images were collected using a
sampling perfection with application optimized contrasts with a different flip angle evolu-
tion sequence (3D-SPACE). This 3D turbo spin echo scan has 192 slices 1 mm thick, TR of
2,800 ms, TE of 402 ms, variable flip angle, 256 × 256 matrix, with parallel imaging
(GRAPPA = 2, 120 reference lines).
Lesions were demarcated onto each participant’s T2 image by an expert neurologist
(Dr. Leonardo Bonilha) or an expert cognitive neuroscientist (Dr. Roger Newman-Norlund)
extensively trained by Dr. Bonilha (with consultation as needed with an expert on lesion
mapping, Dr. Chris Rorden),
both blind to the behavioral data. Lesion maps were then
aligned to the high resolution T1 image. Lesions were replaced with the corresponding
brain structure from the intact hemisphere, and this image as well as the lesion map in
participant space were subsequently warped to Montreal Neurological Institute (MNI)
space (Nachev et al., 2008) using SPM12 (Ashburner & Friston, 2005). The warped lesion
map was then binarized with a 50% probability threshold, which was used to perform
voxel-wise and region of interest (ROI) analyses. Figure 1 shows lesion overlap maps
for each of the three groups indicated in Table 1, indicating the distribution of lesions
and lesion coverage.
Magnetic resonance imaging (MRI):
A neuroimaging technique that
allows for precision mapping of both
brain structure and brain activity.
Neurobiology of Language
8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Q5
Grammatical parallelism in aphasia
Figure 1. Lesion overlap maps for the three groups of participants detailed in Table 1. The lower bound indicates the lower bound of the
lesion load threshold, i.e. the minimum number of participants with damage to a voxel required for statistical analysis; the upper bound indi-
cates maximum overlap. (Left) The primary group (Group 1) of 53 participants who were assessed for AGRAMMATISM and PARAGRAMMATISM. (Middle)
The group of 130 participants who were assessed for Noncanonical sentence comprehension (Group 2), including 42 of the participants from
Group 1. (Right) The broader set of 218 participants who were enrolled in all measures (Group 3).
Behavioral Analyses
In our first pass of behavioral analyses, we compared AGRAMMATIC and PARAGRAMMATIC participants
to their NOT-AGRAMMATIC and NOT-PARAGRAMMATIC counterparts on each analysis of syntactic com-
prehension using linear regression in JASP Team (2023). We performed
these analyses without
lesion volume as a covariate in order to replicate previous studies that have found associations
between agrammatic Broca’s aphasia and syntactic comprehension deficits that did not address
lesion volume as a confounding variable. We corrected for multiple comparisons using a
Bonferroni correction with an adjusted alpha threshold of p < 0.025 for the four comparisons
within each family of tests (treating AGRAMMATISM and PARAGRAMMATISM as separate families), using
one-sided tests (negative associations only), controlling the total family-wise error at p < 0.05.
In a second set of behavioral analyses, we incorporated lesion volume as a covariate in
linear regression. AGRAMMATIC participants in our sample have nearly twice the lesion volume
of the PARAGRAMMATIC participants (Matchin et al., 2020), thus presenting potential confounds.
The lesion volume differences are likely due to vasculature differences in these lesion distri-
butions (DeMarco & Turkeltaub, 2018). In particular, we suspected that some reports of syn-
tactic comprehension deficits in agrammatic patients might be due to their (comparatively
larger) frontal-based lesions encroaching into the temporal lobe, which is consistent with
the fact that chronic Broca’s aphasia reliably implicates posterior temporal as well as frontal
damage (Fridriksson et al., 2015). By incorporating lesion volume as a covariate into our anal-
yses, we controlled for this potentially confounding factor. We corrected for multiple compar-
isons using a Bonferroni correction with an adjusted alpha threshold of p < 0.025 for the four
comparisons within each family of tests (treating agrammatism and paragrammatism as sepa-
rate families), (negative associations only), controlling the total family-wise error at p < 0.05.
We also report supplementary analyses in parallel with those described above, incorporat-
ing age at testing, years of education, and WAB-R AQ as covariates (see Supporting
Information, available at https://doi.org/10.1162/nol_a_00117, for details), given that these
variables likely relate to better outcomes (Lwi et al., 2021).
Lesion Analyses
We performed two types of lesion-behavior analyses. First, we ran an exploratory whole-brain
analysis at the voxel level, reporting unthresholded results to provide an overall picture of the
Neurobiology of Language
9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
Q6
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Grammatical parallelism in aphasia
lesion distributions associated with all behavioral measures (AGRAMMATISM, PARAGRAMMATISM,
AUDITORY WORD RECOGNITION, SEQUENTIAL COMMANDS, SEQUENTIAL COMMANDSAUDWORDS, ACTIVE SENTENCE
COMPREHENSION, NONCANONICAL, NONCANONICALACTIVE). Then, we performed an atlas-based
analysis using the parcellation developed by Faria et al. (2012), which contains both gray
and white matter regions, in NITRC (2020) to identify
regions significantly associated with
agrammatism and paragrammatism, correcting for multiple comparisons using permutation
testing (10,000 permutations). For all of these analyses, to ensure reliable localization we
used a lesion load threshold of 10% of sample (22 participants for SEQUENTIAL COMMANDS and
SEQUENTIALCOMMANDSAUDWORDS, 13 participants for NONCANONICAL and NONCANONICALACTIVE, and 5
participants for AGRAMMATISM and PARAGRAMMATISM), and lesion volume was included as a covar-
iate in all analyses (DeMarco & Turkeltaub, 2018; Ivanova et al., 2021).
Second, we assessed the extent to which damage to the regions implicated in AGRAMMATISM
and PARAGRAMMATISM were associated with each of the four syntactic comprehension analyses.
We first created ROIs based on the significant results from the atlas-based analyses described
above, by combining the significant regions together. Thus, for AGRAMMATISM, the ROI consisted
of the combination of the inferior frontal gyrus, pars opercularis, the inferior frontal gyrus, pars
triangularis, and the posterior middle frontal gyrus, whereas for PARAGRAMMATISM, the ROI con-
sisted of the middle superior temporal gyrus, posterior superior temporal gyrus, and posterior
middle temporal gyrus. We then used these lesion distributions as ROIs for further analysis
of the four syntactic comprehension analyses. We first calculated proportion damage to each
ROI for each participant. We then adjusted the data using a rationalized arcsine transform in
order to deal with nonnormality and unequal variances associated with proportional data
(Studebaker, 1985). We then performed one-sided linear regression analyses relating the
damage values for each ROI and each behavioral analysis, separately, incorporating lesion
volume as a covariate. For the analyses of SEQUENTIALCOMMANDSAUDWORDS and NONCANONICALACTIVE,
we analyzed SEQUENTIALCOMMANDS and NONCANONICAL, incorporating the additional covariates of
Auditory Word Comprehension and Active Sentence Comprehension, respectively. We
corrected for multiple comparisons reflecting both the one-sided tests (negative associations
only) and a Bonferroni correction with an adjusted alpha threshold of p < 0.025 for each
family of tests (treating AGRAMMATISM and PARAGRAMMATISM as separate families), controlling the
total error at p < 0.05.
Finally, to illustrate concretely the fact that subjects with expressive agrammatism also tend
to have large lesions encroaching on the temporal lobe, we also computed a lesion overlap
map of subjects with expressive AGRAMMATISM who also showed the classic agrammatic com-
prehension profile, numerically worse performance on NONCANONICAL relative to canonical
structures (seven subjects). That is, we identified all subjects who performed worse by at least
one trial on the three noncanonical conditions relative to the three canonical conditions, and
then summed the lesion maps for all seven of these subjects, revealing the extent of lesion
overlap in each region within this group. We supplemented this with additional LSM analyses
of expressive agrammatism, with and without a lesion volume covariate (replicating figures
presented in Matchin et al., 2020).
RESULTS
Behavioral Data (No Lesion Volume Covariate)
We compared each group of grammatically impaired participants (AGRAMMATIC, PARAGRAMMATIC)
to their not grammatically impaired counterparts (NOT-AGRAMMATIC, NOT-PARAGRAMMATIC) with
t o o u r f o ur s y n t a c t i c c om p r e h e nsi o n an a l y s e s : S E Q U E N T I A L C O M M A N D S,
r e s p e c t
Neurobiology of Language
10
Q7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Grammatical parallelism in aphasia
SEQUENTIALCOMMANDSAUDWORDS (incorporating the AUDITORY WORD RECOGNITION covariate), NONCANON-
ICAL, NONCANONICALACTIVE (incorporating the ACTIVE SENTENCE COMPREHENSION covariate).
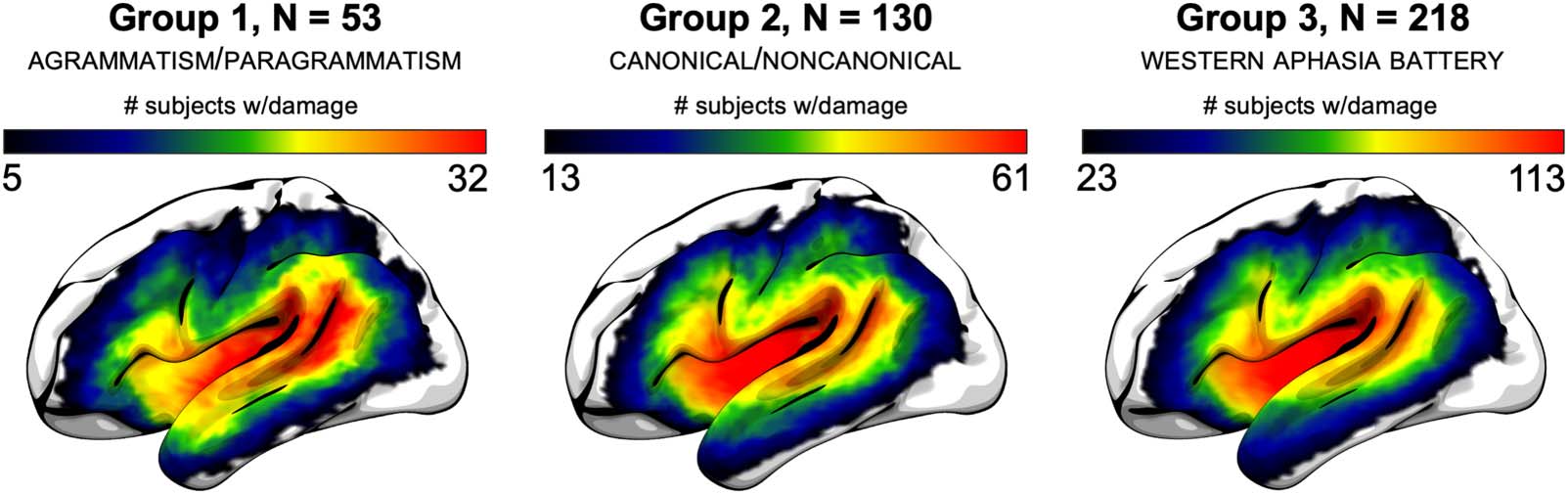
Figure 2 shows average performance in each group on the two primary syntactic compre-
hension measures (SEQUENTIAL COMMANDS and NONCANONICAL), Figure 3 shows average perfor-
mance in each group on the two behavioral covariates (AUDITORY WORD RECOGNITION and ACTIVE
SENTENCE COMPREHENSION), and Figure 4 shows average performance on all canonical and non-
canonical sentence structures from the NAVS and ToS. Table 2 shows statistical results for all of
the behavioral analyses involving these measures, not incorporating lesion volume as a covar-
iate. There were no significant effects of AGRAMMATISM on performance for any of the syntactic
comprehension measures. By contrast, there were significant effects of PARAGRAMMATISM on per-
formance for the analyses of SEQUENTIALCOMMANDSAUDWORDS and NONCANONICAL. Subtracting the sig-
nificant effect sizes (partial η2) for these two analyses for agrammatism from paragrammatism
revealed that the effect sizes were, following the guidelines of Cohen (1988), medium in
strength greater for paragrammatism than agrammatism (0.082 and 0.133, respectively for
the two behavioral measures). Supplementary analyses reported in Table S1 incorporating
age, education, and WAB-R AQ as covariates showed similar results, albeit statistically weak-
ened such that no analyses reached significance after correcting for multiple comparisons.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Behavioral performance on the two raw measures of syntactic comprehension (SEQUEN-
TIAL COMMANDS and NONCANONICAL), sorted by the categorical perceptual ratings of AGRAMMATISM and
PARAGRAMMATISM of Group 1 (total N = 53): AGRAMMATIC (N = 15) / NOT-AGRAMMATIC (N = 38), and PARA-
GRAMMATIC (25) and NOT-PARAGRAMMATIC (28). Dots indicate each individual data point within the rel-
evant group, boxes and hash marks indicate a box-and-whisker plot with median and upper and
lower quartiles, and curves represent estimated continuous distributions. (Top) SEQUENTIAL COMMANDS.
(Bottom) NONCANONICAL. The four participants classified as both AGRAMMATIC and PARAGRAMMATIC were
included in both groups for the analyses here.
Neurobiology of Language
11
Grammatical parallelism in aphasia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
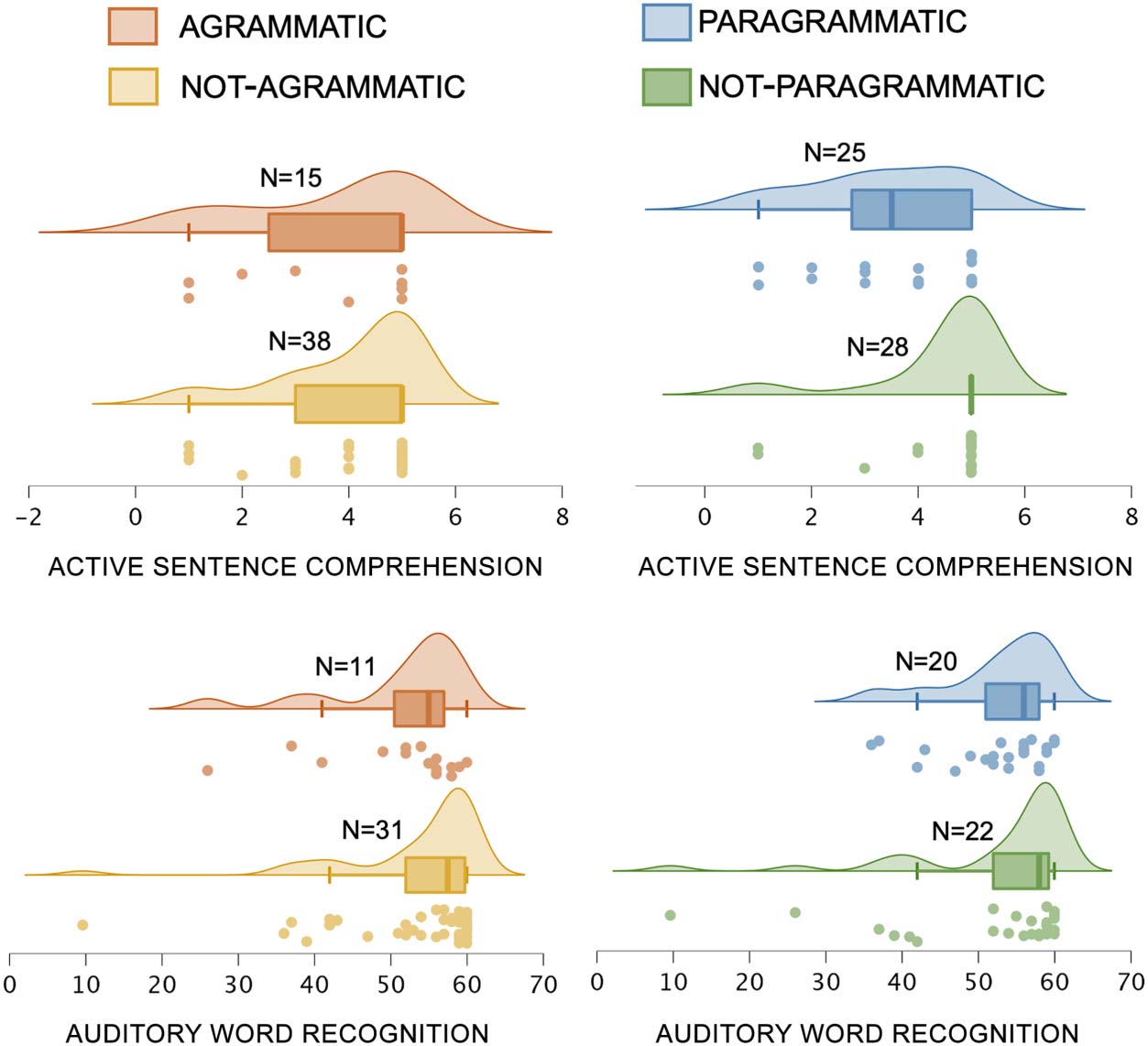
Figure 3. Behavioral performance on the two covariates (ACTIVE SENTENCE COMPREHENSION and AUDI-
TORY WORD RECOGNITION), sorted by the categorical perceptual ratings of AGRAMMATISM and PARAGRAMMA-
TISM of Group 1 (total N = 53): AGRAMMATIC (N = 15) / NOT-AGRAMMATIC (N = 38), and PARAGRAMMATIC (25)
and NOT-PARAGRAMMATIC (28). Dots indicate each individual data point within the relevant group,
boxes and hash marks indicate a box-and-whisker plot with median and upper and lower quartiles,
and curves represent estimated continuous distributions. (Top) ACTIVE SENTENCE COMPREHENSION. (Bot-
tom) AUDITORY WORD RECOGNITION. The four participants classified as both AGRAMMATIC and PARAGRAM-
MATIC were included in both groups for the analyses here.
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
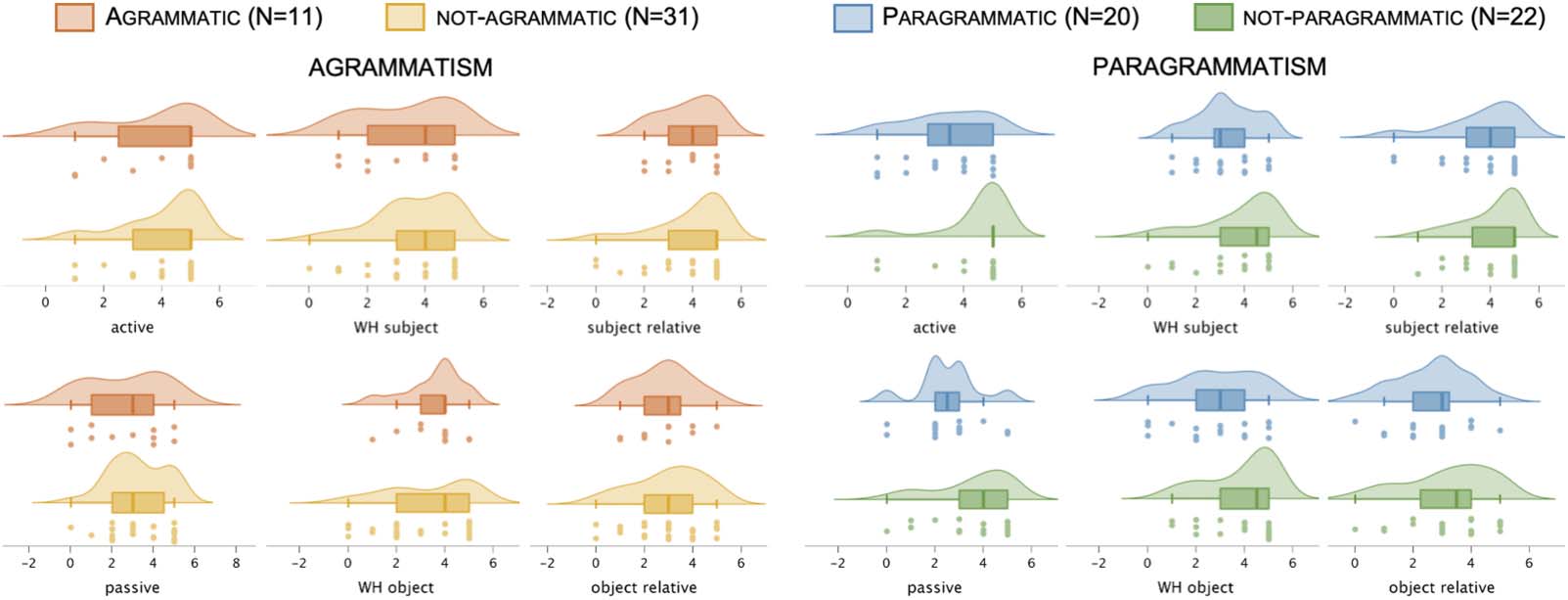
Figure 4. Behavioral performance on each individual canonical and noncanonical sentence type (N = 42), sorted by categorical perceptual
ratings of AGRAMMATISM and PARAGRAMMATISM. Dots indicate each individual data point within the relevant group, boxes and hash marks indicate a
box-and-whisker plot with median and upper and lower quartiles, and curves represent estimated continuous distributions.
Neurobiology of Language
12
Grammatical parallelism in aphasia
Table 2.
Statistical results for behavioral analyses (multivariate linear regression) without incorporating lesion volume as a covariate.
SEQUENTIAL COMMANDS
AGRAMMATISM
t = −1.307, p = 0.197, partial η2 = 0.032
PARAGRAMMATISM
t = −1.459, p = 0.151, partial η2 = 0.040
SEQUENTIAL COMMANDSAUDWORDS
t = −1.217, p = 0.229, partial η2 = 0.029
*t = −2.501, p = 0.016, partial η2 = 0.111
NONCANONICAL
NONCANONICALACTIVE
t = −0.405, p = 0.688, partial η2 = 0.004
*t = −2.524, p = 0.016, partial η2 = 0.137
t = −0.070, p = 0.945, partial η2 = 0.000
t = −1.305, p = 0.199, partial η2 = 0.042
Note. * = statistically significant result, using an adjusted alpha of p < 0.025, reflecting both the one-sided tests (negative associations only) and a Bonferroni
correction for four multiple comparisons within each family of tests (AGRAMMATISM and PARAGRAMMATISM considered as separate families).
Behavioral Data, Incorporating Lesion Volume as a Covariate
We next performed four linear regression analyses examining the relationship between the
scores for AGRAMMATISM and PARAGRAMMATISM and syntactic comprehension, incorporating lesion
volume as a covariate. Table 3 shows statistical results for each of these analyses. There were
no significant effects of AGRAMMATISM on performance for any of the syntactic comprehension
analyses. By contrast, there were significant effects of PARAGRAMMATISM on performance for the
analyses of SEQUENTIALCOMMANDSAUDWORDS and NONCANONICAL, and a trend toward significance for
the analysis of SEQUENTIAL COMMANDS. Subtracting the significant effect sizes (partial η2) for these
two analyses for agrammatism from paragrammatism revealed that the effect sizes were,
following the guidelines of Cohen (1988), large in strength greater for paragrammatism relative
to agrammatism (0.168 and 0.161, respectively for the two behavioral measures). Supplemen-
tary analyses reported in Table S2 incorporating age, education, and WAB-R AQ as covariates
showed similar results, albeit statistically weakened such that no analyses reached significance
after correcting for multiple comparisons.
Brain Lesion Data
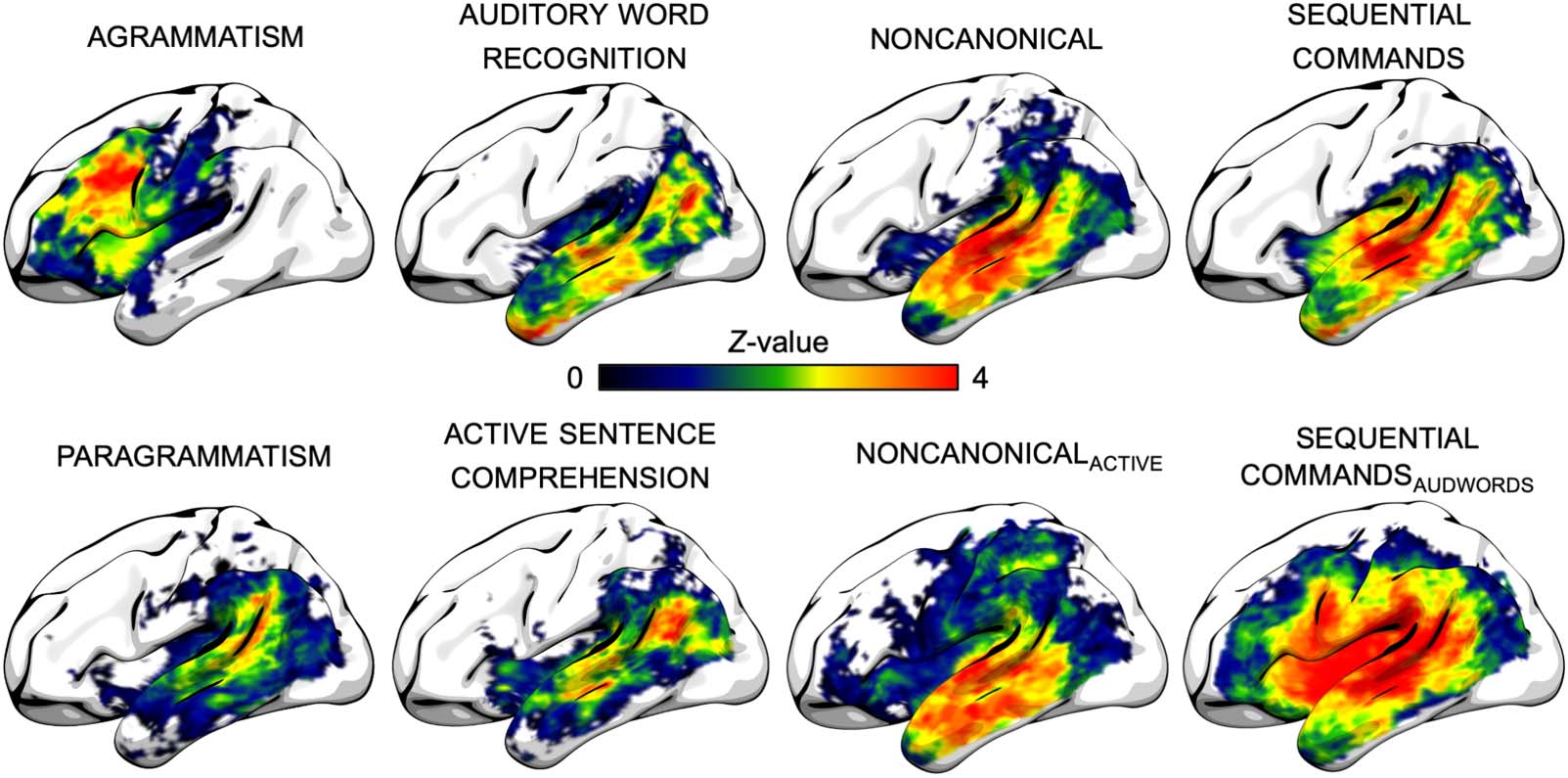
Figure 5 shows unthresholded voxel-wise whole-brain lesion maps associated with each
behavioral measure of interest. As reported in analyses of the same data set in Matchin
et al. (2020), AGRAMMATISM (Group 1) was associated with damage primarily to inferior and mid-
dle frontal areas and anterior insula, and PARAGRAMMATISM (Group 1) was associated with dam-
age primarily to posterior superior temporal and inferior parietal lobe. As reported in analyses
of the same data set in Matchin, Basilakos, et al. (2022), deficits in the lexical-semantic control
measure AUDITORY WORD RECOGNITION (Group 3) were associated with damage throughout the
temporal and parietal lobes, but particularly the inferior portion of the angular gyrus, middle
temporal lobe, and temporal pole. Deficits in the lexical-semantic control measure ACTIVE
SENTENCE COMPREHENSION (Group 2) were associated with damage to the inferior portion of the
Table 3.
Statistical results for behavioral analyses incorporating lesion volume as a covariate (multivariate linear regression).
SEQUENTIAL COMMANDS
AGRAMMATISM
t = 0.632, p = 0.530, partial η2 = 0.008
PARAGRAMMATISM
t = −2.278, p = 0.027, partial η2 = 0.094
SEQUENTIAL COMMANDSAUDWORDS
t = −0.002, p = 0.999, partial η2 = 0
*t = −3.150, p = 0.003, partial η2 = 0.168
NONCANONICAL
NONCANONICALACTIVE
t = 0.205, p = 0.839, partial η2 = 0.001
*t = −2.721, p = 0.010, partial η2 = 0.160
t = 0.700, p = 0.488, partial η2 = 0.013
t = −1.5, p = 0.142, partial η2 = 0.056
Note. * = statistically significant result, using an adjusted alpha of p < 0.025, reflecting both the one-sided tests (negative associations only) and a Bonferroni
correction for four multiple comparisons within each family of tests (AGRAMMATISM and PARAGRAMMATISM considered as separate families).
Neurobiology of Language
13
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Grammatical parallelism in aphasia
Figure 5. Whole-brain voxel-based analyses, unthresholded. All effects include lesion volume as a covariate.
angular gyrus and superior middle temporal lobe. As reported in analyses of a broader data set
(including the proper subset of data reported here) in Matchin, den Ouden, et al. (2022), def-
icits in NONCANONICAL (Group 2) were associated with damage throughout the temporal lobe,
extending into the border of the inferior parietal lobe. As reported on analyses of the same data
set in Matchin, Basilakos, et al. (2022), deficits in NONCANONICAL after incorporating ACTIVE
SENTENCE COMPREHENSION scores as a covariate, NONCANONICALACTIVE, (Group 2) were associated
with damage throughout the temporal lobe. Deficits in SEQUENTIAL COMMANDS (Group 3, data
not reported previously) were associated with somewhat similar patterns of damage to NONCA-
NONICAL, but extending into the temporal pole and the posterior insula. Deficits in SEQUENTIAL
COMMANDS after incorporating AUDITORY WORD RECOGNITION scores as a covariate, SEQUENTIAL
COMMANDSAUDWORDS, (Group 3, data not reported previously) were associated with damage to
the middle and posterior superior temporal lobe, posterior insula, and inferior precentral gyrus,
extending into the inferior frontal gyrus.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
/
.
l
We then determined the regions which were significantly associated with AGRAMMATISM and
PARAGRAMMATISM when correcting for multiple comparisons with permutation thresholding
(10,000 permutations). AGRAMMATISM (with lesion volume as a covariate) was associated with
damage to the posterior middle frontal gyrus (Z = 4.47), inferior frontal gyrus, pars opercularis
(Z = 3.82), and inferior frontal gyrus, pars triangularis (Z = 3.47). PARAGRAMMATISM (with
lesion volume as a covariate) was associated with damage to posterior superior temporal gyrus
(Z = 3.26), middle superior temporal gyrus (Z = 3.03), and posterior middle temporal gyrus
(Z = 2.87).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
We next examined the relationship between our measures of syntactic comprehension and
percent damage to ROIs defined by the combined regions significantly associated with AGRAM-
MATISM (posterior middle frontal gyrus, inferior frontal gyrus, pars opercularis, and inferior fron-
tal gyrus, pars triangularis combined) and PARAGRAMMATISM (posterior superior temporal gyrus,
middle temporal gyrus, and posterior middle temporal gyrus combined) using multivariate lin-
ear regression, incorporating lesion volume as a covariate in all analyses (Table 4). There were
no significant relationships between residual damage to the AGRAMMATISM ROI on performance
for any of the syntactic comprehension measures. By contrast, there were significant relation-
ships between residual damage to the PARAGRAMMATISM ROI on performance for all four syntactic
comprehension analyses. Subtracting the significant effect sizes (partial η2) for these analyses
Neurobiology of Language
14
Grammatical parallelism in aphasia
Table 4.
and each measure of syntactic comprehension.
Statistical results for lesion-deficit linear regression analyses between the brain regions implicated in grammatical production deficits
SEQUENTIAL COMMANDS
AGRAMMATISM ROI
t = 2.129, p = 0.034, partial η2 = 0.021
PARAGRAMMATISM ROI
*t = −5.896, p < 0.001, partial η2 = 0.139
SEQUENTIAL COMMANDSAUDWORDS
t = 0.054, p = 0.957, partial η2 = 0
*t = −5.081, p < 0.001, partial η2 = 0.108
NONCANONICAL
NONCANONICALACTIVE
t = 2.745, p = 0.007, partial η2 = 0.056
*t = −5.043, p < 0.001, partial η2 = 0.167
t = 2.360, p = 0.020, partial η2 = 0.042
*t = −3.787, p < 0.001, partial η2 = 0.102
Note. * = statistically significant result, using an adjusted alpha of p < 0.025, reflecting both the one-sided tests (negative associations only) and a Bonferroni
correction for four multiple comparisons within each family of tests (AGRAMMATISM and PARAGRAMMATISM considered as separate families).
for the agrammatism ROI from the paragrammatism ROI revealed that the effect sizes were,
following the guidelines of Cohen (1988), medium to large in strength greater for the para-
grammatism ROI relative to the agrammatism ROI (0.16, 0.108, 0.223, and 0.144, respectively
for the four behavioral measures).
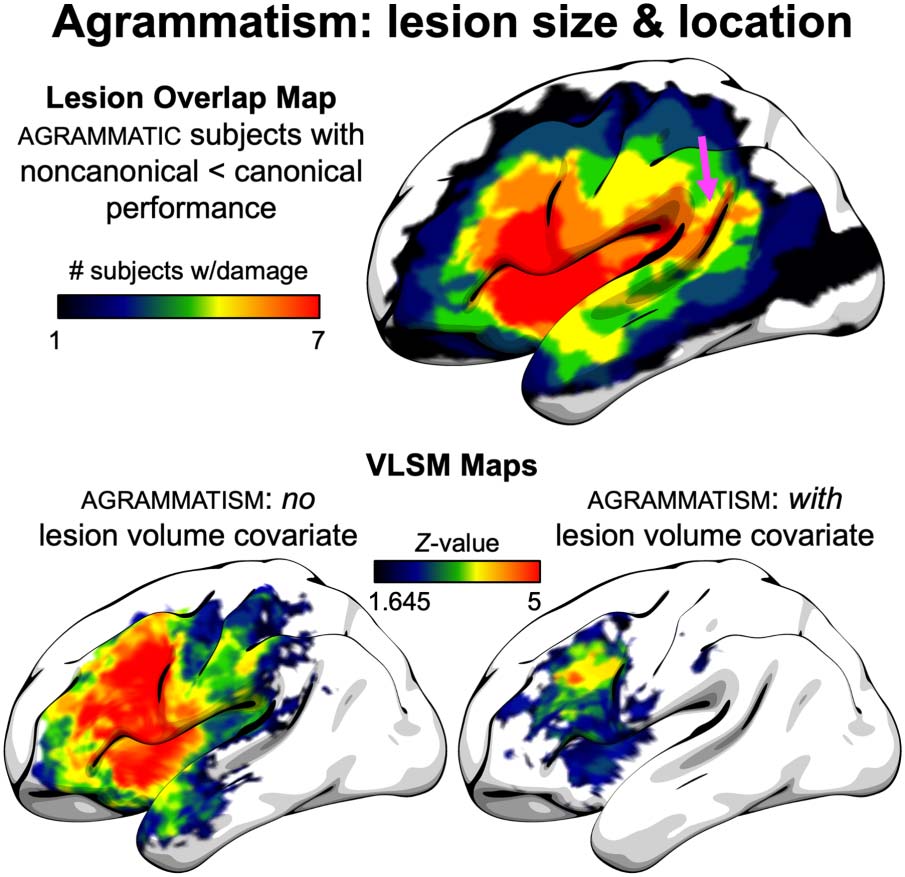
Analyses regarding lesion volume and expressive agrammatism are shown in Figure 6. The
lesion overlap map of agrammatic patients who also showed the classical pattern of agram-
matic comprehension, worse performance on noncanonical relative to canonical structures,
showed that all seven subjects with this profile had lesions in frontal and insular cortex, but
there was also a region of maximum overlap in the posterior superior temporal lobe, including
the posterior superior temporal sulcus. With respect to LSM analyses of expressive agramma-
tism, without including the lesion volume covariate, there were extensive effects throughout
the frontal and parietal lobes, with significant anterior temporal damage and damage extend-
ing into posterior superior temporal gyrus and sulcus. When including the lesion volume
covariate, the significant effects were almost entirely restricted to frontal lobe, with no tempo-
ral lobe involvement.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
n
o
_
a
_
0
0
1
1
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Agrammatism: lesion size and location. (Top) Lesion overlap map of subjects with
expressive agrammatism and agrammatic comprehension, defined as worse performance on non-
canonical relative to canonical structures. Arrow indicates area of maximum overlap in the poste-
rior superior temporal sulcus. (Bottom) LSM analysis maps, thresholded at voxel-wise p < 0.05
(Z-value > 1.645), both without (izquierda) y con (bien) a lesion volume covariate.
Neurobiology of Language
15
Grammatical parallelism in aphasia
DISCUSIÓN
Reassessing Overarching Agrammatism
We found little evidence to support the classical overarching agrammatism hypothesis, el
idea that parallel syntactic comprehension deficits coincide with expressive agrammatism
(Caramazza & Zurif, 1976; Grodzinsky, 2000a; Schwartz et al., 1980; Thompson et al.,
2013; Zurif, 1980). Given that patients with AGRAMMATISM and frontal damage had lesions much
larger than those with paragrammatism or no grammatical deficit (Matchin et al., 2020), nosotros
suspected that previous studies finding associations between agrammatic Broca’s aphasia
(with assumed damage to Broca’s area) and syntactic comprehension deficits, might be due
in part to large lesions in these patients, possibly encroaching on the temporal lobe (see also
Fridriksson et al., 2015). Our results support this account: With or without a lesion volume
covariate, there were no significant associations between expressive agrammatism and syntac-
tic comprehension deficits, and the two potentially suggestive trends present in the analyses
without the lesion volume covariate completely disappeared when the lesion volume covar-
iate was included. Además, including a lesion volume covariate, damage to regions asso-
ciated with AGRAMMATISM (Broca’s area and posterior middle frontal gyrus) was not associated
with any of the syntactic comprehension measures. En general, the data cast strong doubt on the
overarching agrammatism hypothesis.
The lack of any association in our data between expressive agrammatism and deficits in
noncanonical sentence comprehension seemingly contradicts previous studies (Caramazza
& Zurif, 1976; Cho-Reyes & Thompson, 2012; Schwartz et al., 1980; Thompson et al.,
2013), but it is important to note that this does not mean that our agrammatic patients had
no deficits in comprehending these sentence structures. Bastante, it illustrates that agrammatism
sí mismo, out of a group of people with aphasia, is not particularly strongly associated with such
deficits; eso es, people with aphasia but not agrammatism were not significantly different in
their ability to comprehend noncanonical sentence structures. This underscores the impor-
tance of an appropriate comparison group; some previous studies only examined agrammatic
patients (Caplan & Futter, 1986; Grodzinsky, 2000b; Schwartz et al., 1980) or compared
agrammatic patients to those with mild anomic aphasia (Cho-Reyes & Thompson, 2012),
which conflates expressive agrammatism with overall aphasia severity and/or lesion volume.
Note that our results do not imply that inferior frontal cortex regions never play any role in
sentence comprehension. En particular, frontal-motor systems are widely thought to play a key
role in working memory (Baddeley, 2003; Baddeley et al., 1981; Pettigrew & Hillis, 2014;
Rogalsky et al., 2008). Working memory is important for sentence comprehension, particularly
for difficult constructions and perhaps other demanding aspects of everyday natural commu-
nication (being presented with false starts and stops, garden-paths/misparsing, preparing to
respond to the interlocuter). En particular, the linear morpho-syntactic system supported by
the pars triangularis of Broca’s area, as posited by Matchin and Hickok (2020), is ideal for
assisting comprehension by reiterating the heard sequence of morphemes for reanalysis by
hierarchical syntactic mechanisms in the posterior temporal lobe. This is congruent with the
hypothesis of a syntactic working memory system (Fiebach et al., 2005; Matchin, 2018;
Rogalsky et al., 2015) consisting of looping interactions between inferior frontal cortex (typi-
cally used for linear sequencing during production) and posterior temporal cortex. Tal como, a
working memory deficit may help explain some of the reported associations between agram-
matic Broca’s aphasia and deficits in the comprehension of semantically reversible, complex,
noncanonical sentences (although we note that a recent LSM study assessed this possibility
and did not find any support for it (Rogalsky et al., 2018). Además, working memory
Neurobiology of Language
16
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
.
/
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
deficits (along with deficits in visual-motor processing) might account for the fact that agram-
matism was very weakly (and nonsignificantly) associated with deficits in SEQUENTIAL COMMANDS
and SEQUENTIALCOMMANDSAUDWORDS when lesion volume was not taken into account, given that the
Sequential Commands subtest of the WAB-R requires participants to keep a series of actions in
memory to correctly perform more complex items.
A role for frontal-motor systems in syntactic working memory may also explain some of the
syntax-related activations that are observed in inferior frontal cortex in neuroimaging studies
(Rogalsky & Hickok, 2011). While early studies found evidence for syntactic comprehension
effects primarily in Broca’s area, recent neuroimaging studies have found that posterior
temporal cortex shows equally reliable syntactic comprehension effects (Blank et al., 2016;
Diachek et al., 2020; Fedorenko et al., 2012, 2016; Goucha & Friederici, 2015; Matchin
et al., 2017; Meyer & Friederici, 2016; Nelson et al., 2017; Pallier et al., 2011; Shain et al.,
2020; Zaccarella, Meyer, et al., 2017; Zaccarella, Schell, et al., 2017); En realidad, some studies
have shown that posterior temporal lobe shows an activation profile more consistent with
fuller aspects of abstract hierarchical structure building than frontal cortex (Brennan, 2016;
Fedorenko et al., 2020). The fact that both of these regions reliably exhibit syntactic effects is
well-explained by attributing a syntactic function to both of these regions (Matchin & Hickok,
2020), with the frontal contribution reflecting production-related morpho-syntactic resources
that assist comprehension in demanding contexts, but that are not necessary for combining
words into structured phrases and sentences, a role reserved for the posterior temporal lobe.
Some LSM studies have found an association between damage and/or degeneration of infe-
rior frontal cortex and deficits in comprehension of complex, noncanonical sentence structure,
typically in addition to posterior temporal damage (Adezati et al., 2022; Amici et al., 2007;
Fridriksson et al., 2018; Kristinsson et al., 2020; Magnusdottir et al., 2013; Mesulam et al.,
2015; Tyler et al., 2011; Wilson et al., 2011). Sin embargo, there are several reasons to question
whether these results reflect necessary resources for normal syntactic comprehension.
Primero, most of these studies did not incorporate lesion volume as a covariate, which is an
important variable to ensure accurate localization in LSM (DeMarco & Turkeltaub, 2018;
Ivanova et al., 2021). This is a particularly acute issue given that agrammatic production is
associated with large lesions (Matchin et al., 2020) that encroach on the temporal lobe
(Fridriksson et al., 2015). Our lesion overlap analysis showed that 100% of agrammatic sub-
jects who also showed the traditional agrammatic comprehension profile (worse performance
on noncanonical relative to canonical sentences) had lesions extending into the posterior
temporal lobe. It is possible that syntactic comprehension deficits in agrammatic patients in
older studies may have been due to temporal lobe damage rather than the frontal lobe lesions
that were often assumed.
Segundo, many LSM studies have reported no association between syntactic comprehension
and frontal lobe structures even for complex structures (den Ouden et al., 2019; Dronkers
et al., 2004; Matchin, Basilakos, et al., 2022; Matchin, den Ouden, et al., 2022; Rogalsky
et al., 2018; Thothathiri et al., 2012). Tercero, complex noncanonical structures critically involve
substantial working memory resources (Rey & Just, 1991; Pettigrew & Hillis, 2014; Rogalsky
et al., 2008), which could be the source of the associations with frontal networks as noted.
Cuatro, LSM of sentence comprehension in general does not highlight Broca’s area, but rather
posterior temporal and inferior parietal cortex, similar to the areas we identified here (Baldo &
Dronkers, 2007; Dronkers et al., 2004; Fridriksson et al., 2018; Pillay et al., 2017; Rogalsky
et al., 2018; Thothathiri et al., 2012), and most sentences, regardless of their complexity and
canonicity, likely draw on at least some syntactic resources in order to establish the basic
Neurobiology of Language
17
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
/
.
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
thematic relations of sentences. Finalmente, LSM studies have shown that damage to posterior tem-
poral lobe, but not Broca’s area, is associated with syntactic acceptability judgment deficits
(Fahey et al., 2021; wilson & Saygın, 2004). Sin embargo, these studies have limited numbers
of participants. Large-scale LSM studies using a variety of alternative measures of syntactic
comprehension ability, including acceptability judgments, are needed to more firmly assess
the extent to which the posterior temporal lobe is engaged in receptive syntactic processing
that cannot be accounted for by individual word-level conceptual-semantic and/or phonolo-
gical deficits.
Curiosamente, a recent cortical stimulation study in six participants did find that stimulation
of Broca’s area (primarily pars opercularis) resulted in deficits in comprehension of passive
(but not active) oraciones (Riva et al., 2022), which aligns with a case study of acute stroke
resulting in relatively focal hypoperfusion of Broca’s area with impaired syntactic comprehen-
sión (Davis et al., 2008). This is consistent with the possibility that Broca’s area plays a sup-
porting role that, when acutely disrupted, results in dysfunction that resolves over time,
whereas posterior temporal lobe plays a fundamental syntactic structure building function.
Paragrammatism, Syntactic Comprehension Deficits and the Posterior Temporal Lobe
By contrast to our investigation of the relationship between expressive agrammatism and syn-
tactic comprehension deficits, both our behavioral measure of PARAGRAMMATISM, and damage to
the PARAGRAMMATISM ROI (middle and posterior superior temporal gyrus and posterior middle
temporal gyrus) were significantly associated with some syntactic comprehension deficits,
whether lesion volume was included as a covariate or not. These results suggest that while
both posterior temporal and frontal cortex play important roles in syntax, they do so asymmet-
rically: Posterior temporal cortex is critically involved in both comprehension and production,
whereas frontal cortex is only critically involved in production. A functional-anatomical asym-
metry is supported by recent fMRI studies which have found that these regions diverge accord-
ingly with respect to syntactic measures in production and comprehension (Giglio et al., 2022;
Matchin & Wood, 2020). This is consistent with previous functional-anatomical asymmetries
identified in the phonological domain (Hickok & Poeppel, 2004).
These results suggest the possibility of a syntactic parallelism hypothesis rooted in para-
grammatism rather than agrammatism. Roots of this idea extend back to Wernicke (1874),
who argued that the temporal lobe subserved both receptive and expressive function at the
speech-sound level. These ideas were adapted to syntax in the framework advanced by
Matchin and Hickok (2020): The posterior temporal lobe (crucially including ventral superior
temporal sulcus) underlies a hierarchical lexical syntactic function for interfacing with brain
systems involved in processing meaning, whereas the pars triangularis of Broca’s area and per-
haps a more dorsal region in the middle frontal gyrus underlies a morpho-syntactic sequencing
function that interfaces with the motor system. De este modo, it is possible that the more abstract idea
underlying overarching agrammatism is viable—that a central syntactic system can be local-
ized to particular part of the brain and supports both production and comprehension, pero en
the posterior temporal lobe and not the frontal lobe.
Limitaciones
We focused on the question of whether or not an overarching agrammatism hypothesis is
apoyado, in accordance with traditional claims in the literature, and whether or not an
alternative overarching paragrammatism hypothesis is supported. We are not claiming that
the significant effects we identified in paragrammatism are stronger than those in
Neurobiology of Language
18
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
.
/
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
agrammatism, only that we can identify an association between grammatical production and
comprehension deficits only for paragrammatism, and not for agrammatism. En tono rimbombante, mientras
we did find evidence of a relationship between PARAGRAMMATISM and deficits in some syntactic
comprehension measures, this was not overwhelmingly robust, and some of the measures did
not reach significance. We did not have handedness or education information for all subjects,
which are important variables that are known to relate to aphasia outcome (Lwi et al., 2021).
En tono rimbombante, while we did find evidence of a relationship between PARAGRAMMATISM and deficits in
some syntactic comprehension measures, this was not overwhelmingly robust, and some of the
measures did not reach significance (and the statistical results were weaker when incorporating
edad, education, and WAB-R AQ as covariates; see Supplementary Data).
Además, while there was a statistically strong relationship between damage to the
regions implicated in PARAGRAMMATISM and syntactic comprehension deficits, the whole-brain
lesion maps for these measures showed some important differences in the lesion distributions
for these measures. The SEQUENTIAL COMMANDS and SEQUENTIALCOMMANDSAUDWORDS analyses, mientras
designed to avert some of the limitations of working memory demands required of complex
estructuras, likely still involved some degree of phonological working memory resources which
shifted the lesion distribution superiorly away from the middle temporal gyrus. Asimismo, el
analyses of NONCANONICAL and NONCANICALACTIVE likely taxed working memory resources and/or
conceptual-semantic processing in addition to syntax. Además, the speech of patients with
fluent aphasia, who are overrepresented in the PARAGRAMMATIC group analyzed here relative to
the AGRAMMATIC group, frequently contains phonological and semantic errors, some of which
may contribute to the overall impression of paragrammatic speech output used for categori-
zation in this study. New reliable and valid measures of agrammatic and paragrammatic
speech production and syntactic comprehension abilities are needed to more clearly investi-
gate these issues.
Finalmente, although LSM provides important insights into the organization of the brain, com-
plementary to functional neuroimaging in healthy subjects, it also has complicating factors.
Por ejemplo, patterns of functional disruption due to stroke may be complex and difficult to
assess purely from the standpoint of lesions visually identifiable on MRI scans. Además,
particularly in chronic stroke there may be functional reorganization, which makes interpre-
tation of lesion-deficit correlation results difficult to interpret. De este modo, we continue to advocate
for the combination of methods, including both LSM and functional neuroimaging (and other
methods, such as brain stimulation), to provide the fullest insights into the organization of syn-
tax in the brain.
Conclusions
En este trabajo, we would like to reinforce two main points. Primero, behaviorally, paragrammatism,
and not expressive agrammatism, is associated with behavioral deficits in syntactic compre-
hension, when assessed from a variety of perspectives and covariates. Segundo, neurologically,
there is convergence in the brain lesions associated with syntactic comprehension and para-
grammatism but not agrammatism. Fundamentalmente, our results may conflict with previous studies
because we included a full range of patient types, rather than solely comparing agrammatic
patients to people without aphasia or only mild anomic aphasia (cf. Thompson et al., 2013,
which examined both stroke-based aphasia and primary progressive aphasia
, comparing
agrammatic patients to those with significantly less severe aphasia), and investigated the role
This provides strong evidence against the concept of “overarching agramma-
of lesion size.
tism” rooted in the frontal lobe, and provides some converging evidence for a role for posterior
Neurobiology of Language
19
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
/
.
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
Q8
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Q9
Q10
Grammatical parallelism in aphasia
temporal lobe in a central syntactic processing mechanism that supports both comprehension
and production.
EXPRESIONES DE GRATITUD
We wish to thank Leonardo Bonilha, Roger Newman-Norlund, and Chris Rorden for their
asistencia.
INFORMACIÓN DE FINANCIACIÓN
Alexandra Basilakos, National Institute on Deafness and Other Communication Disorders
(https://dx.doi.org/10.13039/100000055), Award ID: DC014435. Julius Fridriksson, National
Institute on Deafness and Other Communication Disorders (https://dx.doi.org/10.13039
/100000055), Award ID: DC014664. Julius Fridriksson, National Institute on Deafness and
Other Communication Disorders (https://dx.doi.org/10.13039/100000055), Award ID:
DC011739.
CONTRIBUCIONES DE AUTOR
William Matchin: Conceptualización; Análisis formal; Investigación; Administración de proyecto;
Visualización; Escritura – borrador original; Writing – review and editing. Dirk-Bart den Ouden:
Investigación; Administración de proyecto; Writing – review and editing. Alexandra Basilakos: Datos
curation; Investigación; Writing – review and editing. Brielle Caserta Stark: Investigación;
Writing – review and editing. Julius Fridriksson: Adquisición de financiación; Recursos. Gregory
Hickok: Conceptualización; Writing – review and editing.
DATA AVAILABILITY STATEMENT
All data are publicly available using the following link (use the grammaticalParallelism_
data_8.3.2022.xlsx spreadsheet): https://www.dropbox.com/home/Matchin_publiclyShared
_C-STAR_lesionData.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
.
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
REFERENCIAS
Adezati, MI., Thye, METRO., Edmondson-Stait, A. J., Szaflarski, j. PAG., &
Mirman, D. (2022). Lesion correlates of auditory sentence
comprehension deficits in post-stroke aphasia. NeuroImagen:
Informes, 2(1), Article 100076. https://doi.org/10.1016/j.ynirp
.2021.100076, PubMed: 35243477
Amici, S., Brambati, S. METRO., Wilkins, D. PAG., Ogar, J., Dronkers, norte. l.,
Molinero, B. l., & Gorno-Tempini, METRO. l. (2007). Anatomical corre-
lates of sentence comprehension and verbal working memory in
neurodegenerative disease. Revista de neurociencia, 27(23),
6282–6290. https://doi.org/10.1523/JNEUROSCI.1331-07.2007,
PubMed: 17554002
Arbib, METRO. A. (2016). Towards a computational comparative neuro-
primatology: Framing the language-ready brain. Physics of Life
Reseñas, 16, 1–54. https://doi.org/10.1016/j.plrev.2015.09.003,
PubMed: 26482863
Ashburner, J., & Friston, k. j. (2005). Unified segmentation. Neuro-
Image, 26(3), 839–851. https://doi.org/10.1016/j.neuroimage
.2005.02.018, PubMed: 15955494
Baddeley, A. (2003). Working memory: Looking back and looking
forward. Naturaleza Reseñas Neurociencia, 4(10), 829–839. https://
doi.org/10.1038/nrn1201, PubMed: 14523382
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Baddeley, A., Eldridge, METRO., & Luis, V. (1981). The role of subvo-
calisation in reading. The Quarterly Journal of Experimental Psy-
chology Section A, 33(4), 439–454. https://doi.org/10.1080
/14640748108400802
Baldo, j. v., & Dronkers, norte. F. (2007). Neural correlates of arith-
metic and language comprehension: A common substrate? nuevo-
ropsychologia, 45(2), 229–235. https://doi.org/10.1016/j
.neuropsychologia.2006.07.014, PubMed: 16997333
Berndt, R. S., & Caramazza, A. (1980). A redefinition of the syn-
drome of Broca’s aphasia: Implications for a neuropsychological
model of language. Applied Psycholinguistics, 1(3), 225–278.
https://doi.org/10.1017/S0142716400000552
Billot, A., Thiebaut de Schotten, METRO., Parrish, t. B., Thompson, C. K.,
Rapp, B., Caplan, D., & Kiran, S. (2022). Structural disconnections
associated with language impairments in chronic post-stroke apha-
sia using disconnectome maps. Corteza, 155, 90–106. https://doi
.org/10.1016/j.cortex.2022.06.016, PubMed: 35985126
Blank, I., Balewski, Z., Mahowald, K., & Fedorenko, mi. (2016). Syn-
tactic processing is distributed across the language system. nuevo-
roImage, 127, 307–323. https://doi.org/10.1016/j.neuroimage
.2015.11.069, PubMed: 26666896
Neurobiology of Language
20
Grammatical parallelism in aphasia
Bonhoeffer, C. (1902). Zur Kenntniss der Rückbildung motorischer
Aphasien. Mitteilung aus den Grenzgebieten der Medizin und
Chirurgie, 10, 203–224.
Bozic, METRO., Fonteneau, MI., Su, l., & Marslen-Wilson, W.. D. (2015).
Grammatical analysis as a distributed neurobiological function:
Bihemispheric grammatical analysis. Mapeo del cerebro humano,
36(3), 1190–1201. https://doi.org/10.1002/ hbm.22696,
PubMed: 25421880
Brennan, j. (2016). Naturalistic sentence comprehension in the
cerebro. Language and Linguistics Compass, 10(7), 299–313.
https://doi.org/10.1111/lnc3.12198
Brennan, j. r., Stabler, mi. PAG., Van Wagenen, S. MI., Luh, W.-M., &
Hale, j. t. (2016). Abstract linguistic structure correlates with
temporal activity during naturalistic comprehension. Brain and
Idioma, 157–158, 81–94. https://doi.org/10.1016/j.bandl
.2016.04.008, PubMed: 27208858
Butterworth, B., & Howard, D. (1987). Paragrammatisms. Cogni-
ción, 26(1), 1–37. https://doi.org/10.1016/0010-0277(87)90012
-6, PubMed: 3608394
Caplan, D., & Futter, C. (1986). Assignment of thematic roles to
nouns in sentence comprehension by an agrammatic patient.
Brain and Language, 27(1), 117–134. https://doi.org/10.1016
/0093-934X(86)90008-8, PubMed: 3947937
Caplan, D., Hildebrandt, NORTE., & Makris, norte. (1996). Location of
lesions in stroke patients with deficits in syntactic processing in
sentence comprehension. Cerebro, 119(3), 933–949. https://doi.org
/10.1093/brain/119.3.933, PubMed: 8673503
Caramazza, A., & Zurif, mi. B. (1976). Dissociation of algorithmic
and heuristic processes in language comprehension: Evidencia
from aphasia. Brain and Language, 3(4), 572–582. https://doi
.org/10.1016/0093-934X(76)90048-1, PubMed: 974731
Chomsky, norte. (1965). Aspects of the theory of syntax. CON prensa.
https://doi.org/10.21236/AD0616323
Chomsky, norte. (1980). Rules and representations. Columbia Univer-
sity Press.
Chomsky, norte. (1981). Lectures on government and binding. Foris.
Cho-Reyes, S., & Thompson, C. k. (2012). Verb and sentence pro-
duction and comprehension in aphasia: Northwestern Assess-
ment of Verbs and Sentences (NAVS). Aphasiology, 26(10),
1250–1277. https://doi.org/10.1080/02687038.2012.693584,
PubMed: 26379358
cohen, j. (1988). Statistical power analysis for the behavioral sci-
ences. Routledge.
Dapretto, METRO., & Bookheimer, S. Y. (1999). Form and content: Dis-
sociating syntax and semantics in sentence comprehension.
Neurona, 24(2), 427–432. https://doi.org/10.1016/S0896
-6273(00)80855-7, PubMed: 10571235
D’Ausilio, A., Craighero, l., & Fadiga, l. (2012). The contribution
of the frontal lobe to the perception of speech. Journal of Neuro-
linguistics, 25(5), 328–335. https://doi.org/10.1016/j.jneuroling
.2010.02.003
davis, C., Kleinman, j. T., Newhart, METRO., Gingis, l., Pawlak, METRO., &
Hillis, A. mi. (2008). Speech and language functions that require a
functioning Broca’s area. Brain and Language, 105(1), 50–58.
https://doi.org/10.1016/j.bandl.2008.01.012, PubMed:
18325581
DeMarco, A. T., & Turkeltaub, PAG. mi. (2018). A multivariate lesion
symptom mapping toolbox and examination of lesion-volume
biases and correction methods in lesion-symptom mapping.
Mapeo del cerebro humano, 39(11), 4169–4182. https://doi.org/10
.1002/hbm.24289, PubMed: 29972618
den Ouden, D.-B., Malyutina, S., Basilakos, A., Bonilha, l.,
Gleichgerrcht, MI., Yourganov, GRAMO., Hillis, A. MI., Hickok, GRAMO.,
Rorden, C., & Fridriksson, j. (2019). Cortical and structural-
connectivity damage correlated with impaired syntactic pro-
cessing in aphasia. Mapeo del cerebro humano, 40(7), 2153–2173.
https://doi.org/10.1002/hbm.24514, PubMed: 30666767
Diachek, MI., Blank, I., Siegelman, METRO., Affourtit, J., & Fedorenko, mi.
(2020). The domain-general multiple demand (Maryland) network
does not support core aspects of language comprehension: A
large-scale fMRI investigation. Revista de neurociencia, 40(23),
4536–4550. https://doi.org/10.1523/JNEUROSCI.2036-19.2020,
PubMed: 32317387
Dronkers, norte. F., Wilkins, D. PAG., Van Valin, R. D., Jr., Redfern, B. B., &
Jaeger, j. j. (2004). Lesion analysis of the brain areas involved in
language comprehension. Cognición, 92(1–2), 145–177. https://
doi.org/10.1016/j.cognition.2003.11.002, PubMed: 15037129
Embick, D., Marantz, A., Miyashita, y., O’Neil, w., & Sakai, k. l.
(2000). A syntactic specialization for Broca’s area. Actas
of the National Academy of Sciences, 97(11), 6150–6154.
https://doi.org/10.1073/pnas.100098897, PubMed: 10811887
Fahey, D., Igoe, J., Fridriksson, J., Hickok, GRAMO., & Matchin, W..
(2021). A lesion-symptom mapping study of syntactic accept-
ability judgments in chronic post-stroke aphasia [Poster]. Acad-
emy of Aphasia, Virtual. https://easychair.org/publications
/preprint_open/BQLM
Faria, A. v., joel, S. MI., zhang, y., Oishi, K., van Zjil, PAG. C. METRO., Molinero,
METRO. I., Pekar, j. J., & Mori, S. (2012). Atlas-based analysis of
resting-state functional connectivity: Evaluation for reproduc-
ibility and multi-modal anatomy–function correlation studies.
NeuroImagen, 61(3), 613–621. https://doi.org/10.1016/j
.neuroimage.2012.03.078, PubMed: 22498656
Fedorenko, MI., Blank, I. A., Siegelman, METRO., & Mineroff, z. (2020).
Lack of selectivity for syntax relative to word meanings through-
out the language network. Cognición, 203, Article 104348.
https://doi.org/10.1016/j.cognition.2020.104348, PubMed:
32569894
Fedorenko, MI., Nieto-Castañon, A., & Kanwisher, norte. (2012). Lexical
and syntactic representations in the brain: An fMRI investigation
with multi-voxel pattern analyses. Neuropsicología, 50(4),
499–513. https://doi.org/10.1016/j.neuropsychologia.2011.09
.014, PubMed: 21945850
Fedorenko, MI., Scott, t. l., brunner, PAG., Coon, W.. GRAMO., Pritchett, B.,
Schalk, GRAMO., & Kanwisher, norte. (2016). Neural correlate of the
construction of sentence meaning. Actas del Nacional
Academia de Ciencias, 113(41), E6256–E6262. https://doi.org/10
.1073/pnas.1612132113, PubMed: 27671642
Fiebach, C. J., Schlesewsky, METRO., Lohmann, GRAMO., por cramon, D. y.,
& Friederici, A. D. (2005). Revisiting the role of Broca’s area in
sentence processing: Syntactic integration versus syntactic
working memory. Mapeo del cerebro humano, 24(2), 79–91. https://
doi.org/10.1002/hbm.20070, PubMed: 15455462
Flick, GRAMO., & Pylkkänen, l. (2020). Isolating syntax in natural lan-
guage: MEG evidence for an early contribution of left posterior
temporal cortex. Corteza, 127, 42–57. https://doi.org/10.1016/j
.cortex.2020.01.025, PubMed: 32160572
Forster, mi. (1919). Agrammatismus (erschwerte Satzfindung) und
Mangel an Antrieb nach Stirnhirnverletzung. Monatsschrift für
Psychiatrie and Neurologie, 46(1), 22–43. https://doi.org/10
.1159/000190703
Fridriksson, J., Bonilha, l., & Rorden, C. (2007). Severe Broca’s
aphasia without Broca’s area damage. Behavioural Neurology,
18(4), 237–238. https://doi.org/10.1155/2007/785280, PubMed:
18430982
Fridriksson, J., den Ouden, D.-B., Hillis, A. MI., Hickok, GRAMO., Rorden,
C., Basilakos, A., Yourganov, GRAMO., & Bonilha, l. (2018). Anatomy
Neurobiology of Language
21
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
.
/
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
of aphasia revisited. Cerebro, 141(3), 848–862. https://doi.org/10
.1093/brain/awx363, PubMed: 29360947
Fridriksson, J., Fillmore, PAG., guo, D., & Rorden, C. (2015). Chronic
Broca’s aphasia is caused by damage to Broca’s and Wernicke’s
areas. Corteza cerebral, 25(12), 4689–4696. https://doi.org/10
.1093/cercor/bhu152, PubMed: 25016386
Friederici, A. D. (2002). Towards a neural basis of auditory sen-
tence processing. Tendencias en Ciencias Cognitivas, 6(2), 78–84.
https://doi.org/10.1016/S1364-6613(00)01839-8, PubMed:
15866191
Friederici, A. D. (2017). Language in our brain: The origins of a
uniquely human capacity. CON prensa. https://doi.org/10.7551
/mitpress/9780262036924.001.0001
Friederici, A. D., Meyer, METRO., & por cramon, D. Y. (2000). Auditory
language comprehension: An event-related fMRI study on the
processing of syntactic and lexical information. Brain and Lan-
guage, 74(2), 289–300. https://doi.org/10.1006/brln.2000.2313,
PubMed: 10950920
Gajardo-Vidal, A., Lorca-Puls, D. l., PLORAS Team, Warner, h.,
Pshdary, B., Crinion, j. T., Leff, A. PAG., Hope, t. METRO. h., Geva, S.,
Seghier, METRO. l., Verde, D. w., Bowman, h., & Precio, C. j.
(2021). Damage to Broca’s area does not contribute to
long-term speech production outcome after stroke. Cerebro,
144(3), 817–832. https://doi.org/10.1093/ brain/awaa460,
PubMed: 33517378
Geschwind, norte. (1970). The organization of language and the brain:
Language disorders after brain damage help in elucidating the
neural basis of verbal behavior. Ciencia, 170(3961), 940–944.
https://doi.org/10.1126/science.170.3961.940, PubMed:
5475022
Geschwind, norte. (1972). Language and the brain. Scientific American,
226(4), 76–83. https://doi.org/10.1038/scientificamerican0472-76,
PubMed: 5014017
Geschwind, norte. (1979). Specializations of the human brain. ciencia-
tific American, 241(3), 180–199. https://doi.org/10.1038
/scientificamerican0979-180, PubMed: 493918
Giglio, l., Ostarek, METRO., Weber, K., & Hagoort, PAG. (2022). Common-
alities and asymmetries in the neurobiological infrastructure for
language production and comprehension. Corteza cerebral,
32(7), 1405–1418. https://doi.org/10.1093/cercor/ bhab287,
PubMed: 34491301
Goodglass, h., Christensen, j. A., & Gallagher, R. (1993). Compar-
ison of morphology and syntax in free narrative and structured
pruebas: Fluent vs. nonfluent aphasics. Corteza, 29(3), 377–408.
https://doi.org/10.1016/S0010-9452(13)80250-X, PubMed:
8258282
Gorno-Tempini, METRO. l., Dronkers, norte. F., Rankin, k. PAG., Ogar, j. METRO.,
Phengrasamy, l., rosa, h. J., Johnson, j. K., Weiner, METRO. w., &
Molinero, B. l. (2004). Cognition and anatomy in three variants
of primary progressive aphasia. Annals of Neurology, 55(3),
335–346. https://doi.org/10.1002/ana.10825, PubMed:
14991811
Goucha, T., & Friederici, A. D. (2015). The language skeleton after
dissecting meaning: A functional segregation within Broca’s area.
NeuroImagen, 114, 294–302. https://doi.org/10.1016/j
.neuroimage.2015.04.011, PubMed: 25871627
Grodzinsky, Y. (1986). Language deficits and the theory of syntax.
Brain and Language, 27(1), 135–159. https://doi.org/10.1016
/0093-934X(86)90009-X, PubMed: 3947938
Grodzinsky, Y. (2000a). Overarching agrammatism. In Y.
Grodzinsky, l. Shapiro, & D. Swinney (Editores.), Language and the
cerebro: Representation and processing (páginas. 73–86). Académico
Prensa. https://doi.org/10.1016/B978-012304260-6/50005-0
Grodzinsky, Y. (2000b). The neurology of syntax: Language use
without Broca’s area. Ciencias del comportamiento y del cerebro, 23(1),
1–21. https://doi.org/10.1017/S0140525X00002399, PubMed:
11303337
Grodzinsky, y., & Finkel, l. (1998). The neurology of empty cate-
gories: Aphasics’ failure to detect ungrammaticality. Diario de
Neurociencia Cognitiva, 10(2), 281–292. https://doi.org/10
.1162/089892998562708, PubMed: 9555112
Hagoort, PAG. (2005). On Broca, cerebro, and binding: A new frame-
trabajar. Tendencias en Ciencias Cognitivas, 9(9), 416–423. https://doi
.org/10.1016/j.tics.2005.07.004, PubMed: 16054419
Hagoort, PAG. (2013). MUC (memory, unification, control) y
más allá de. Fronteras en psicología, 4, Article 416. https://doi.org
/10.3389/fpsyg.2013.00416, PubMed: 23874313
Hagoort, PAG. (2016). MUC (memory, unification, control): A model
on the neurobiology of language beyond single word processing.
In Neurobiology of language (páginas. 339–347). Elsevier. https://doi
.org/10.1016/B978-0-12-407794-2.00028-6
Hagoort, PAG., & Indefrey, PAG. (2014). The neurobiology of language
beyond single words. Revisión anual de neurociencia, 37(1),
347–362. https://doi.org/10.1146/annurev-neuro-071013
-013847, PubMed: 24905595
Heeschen, C. (1985). Agrammatism versus paragrammatism: A ficti-
tious opposition. In M.-L. Kean (Ed.), Agrammatism (páginas. 207–248).
Prensa académica. https://doi.org/10.1016/ B978-0-12-402830-2
.50012-2
Hickok, GRAMO., & Avrutin, S. (1995). Representation, referentiality, y
processing in agrammatic comprehension: Two case studies.
Brain and Language, 50(1), 10–26. https://doi.org/10.1006/brln
.1995.1038, PubMed: 7552227
Hickok, GRAMO., & Poeppel, D. (2000). Towards a functional neuroanat-
omy of speech perception. Tendencias en Ciencias Cognitivas, 4(4),
131–138. https://doi.org/10.1016/S1364-6613(00)01463-7,
PubMed: 10740277
Hickok, GRAMO., & Poeppel, D. (2004). Dorsal and ventral streams: A
framework for understanding aspects of the functional anatomy
of language. Cognición, 92(1–2), 67–99. https://doi.org/10.1016/j
.cognition.2003.10.011, PubMed: 15037127
Hickok, GRAMO., Zurif, MI., & Canseco-Gonzalez, mi. (1993). Structural
description of agrammatic comprehension. Brain and Language,
45(3), 371–395. https://doi.org/10.1006/ brln.1993.1051,
PubMed: 8269331
Isserlin, METRO. (1922). Über Agrammatisms. Zeitschrift für die gesamte
Neurologie and Psychiatrie, 75, 332–410. https://doi.org/10.1007
/BF02901581
Ivanova, METRO. v., Herron, t. J., Dronkers, norte. F., & Baldo, j. V. (2021).
An empirical comparison of univariate versus multivariate
methods for the analysis of brain–behavior mapping. Humano
Mapeo cerebral, 42(4), 1070–1101. https://doi.org/10.1002/hbm
.25278, PubMed: 33216425
Jakobson, R. (1956). Two aspects of language and two types of
aphasic disturbances. En R. Jakobson & METRO. Halle (Editores.), Funda-
mentals of language (páginas. 115–133). Moutón.
Jakobson, r., & Halle, METRO. (Editores.). (1956). Fundamentals of language.
Moutón.
JASP Team. (2023). JASP ( Versión 0.17.3) [Computer software].
Kean, M.-L. (1977). The linguistic interpretation of aphasic syn-
dromes: Agrammatism in Broca’s aphasia, an example. Cogni-
ción, 5(1), 9–46. https://doi.org/10.1016/0010-0277(77)90015-4
Kean, M.-L. (1995). The elusive character of agrammatism. Cerebro
and Language, 50(3), 369–384. https://doi.org/10.1006/ brln
.1995.1053, PubMed: 7583195
Neurobiology of Language
22
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
.
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Q11
Grammatical parallelism in aphasia
Kertesz, A. (2006). Western aphasia battery—Revised. Grune and
Stratton. https://doi.org/10.1037/t15168-000
Rey, J., & Just, METRO. A. (1991). Individual differences in syntactic pro-
cesando: The role of working memory. Journal of Memory and
Idioma, 30(5), 580–602. https://doi.org/10.1016/0749
-596X(91)90027-h
Kleist, k. (1914). Aphasie und geisteskrankheit. Münchener Medizi-
nische Wochenschrift, 61, 8–12.
Kristinsson, S., Thors, h., Yourganov, GRAMO., Magnusdottir, S., Hjaltason,
h., Rígido, B. C., Basilakos, A., den Ouden, D.-B., Bonilha, l.,
Rorden, C., Hickok, GRAMO., Hillis, A., & Fridriksson, j. (2020). Cerebro
damage associated with impaired sentence processing in acute
aphasia. Revista de neurociencia cognitiva, 32(2), 256–271.
https://doi.org/10.1162/jocn_a_01478, PubMed: 31596169
Kuperberg, GRAMO. R. (2007). Neural mechanisms of language compre-
hension: Challenges to syntax. Brain Research, 1146, 23–49.
https://doi.org/10.1016/j.brainres.2006.12.063, PubMed:
17400197
Lau, mi. F., Phillips, C., & Poeppel, D. (2008). A cortical network for
semantics: (De)constructing the N400. Nature Reviews Neurosci-
ence, 9(12), 920–933. https://doi.org/10.1038/nrn2532, PubMed:
19020511
Levelt, W.. j. METRO. (2001). Spoken word production: A theory of
lexical access. procedimientos de la Academia Nacional de Ciencias,
98(23), 13464–13471. https://doi.org/10.1073/pnas.231459498,
PubMed: 11698690
Levelt, W.. j. METRO., Roelofs, A., & Meyer, A. S. (1999). A theory of lex-
ical access in speech production. Ciencias del comportamiento y del cerebro,
22(1), 1–38. https://doi.org/10.1017/S0140525X99001776,
PubMed: 11301520
Linebarger, METRO. C., Schwartz, METRO. F., & Saffran, mi. METRO. (1983). Sensi-
tivity to grammatical structure in so-called agrammatic aphasics.
Cognición, 13(3), 361–392. https://doi.org/10.1016/0010
-0277(83)90015-X, PubMed: 6683142
Lukic, S., Thompson, C. K., Barbieri, MI., Chiappetta, B., Bonakdarpour,
B., Kiran, S., Rapp, B., Parrish, t. B., & Caplan, D. (2021).
Common and distinct neural substrates of sentence production
and comprehension. NeuroImagen, 224, Article 117374. https://
doi.org/10.1016/j.neuroimage.2020.117374, PubMed:
32949711
Lwi, S. J., Herron, t. J., Curran, B. C., Ivanova, METRO. v., Schendel, K.,
Dronkers, norte. F., & Baldo, j. V. (2021). Auditory comprehension
deficits in post-stroke aphasia: Neurologic and demographic
correlates of outcome and recovery. Frontiers in Neurology, 12,
Article 680248. https://doi.org/10.3389/fneur.2021.680248,
PubMed: 34456845
Magnusdottir, S. (2005). Setningafræðipróf (Test of syntax). Landspí-
tali University Hospital.
Magnusdottir, S., Fillmore, PAG., den Ouden, D.-B., Hjaltason, h.,
Rorden, C., Kjartansson, o., Bonilha, l., & Fridriksson, j.
(2013). Damage to left anterior temporal cortex predicts impair-
ment of complex syntactic processing: A lesion-symptom map-
ping study. Mapeo del cerebro humano, 34(10), 2715–2723. https://
doi.org/10.1002/hbm.22096, PubMed: 22522937
matar, S., Dirani, J., Marantz, A., & Pylkkänen, l. (2021). Left pos-
terior temporal cortex is sensitive to syntax within conceptually
matched Arabic expressions. Informes Científicos, 11(1), Article
7181. https://doi.org/10.1038/s41598-021-86474-x, PubMed:
33785801
Matchin, W..
(2018). A neuronal retuning hypothesis of
sentence-specificity in Broca’s area. Boletín psiconómico &
Revisar, 25(5), 1682–1694. https://doi.org/10.3758/s13423-017
-1377-6, PubMed: 28940045
Matchin, w., Basilakos, A., den Ouden, D.-B., Rígido, B. C., Hickok,
GRAMO., & Fridriksson, j. (2022). Functional differentiation in the lan-
guage network revealed by lesion-symptom mapping. Neuro-
Image, 247, Article 118778. https://doi.org/10.1016/j
.neuroimage.2021.118778, PubMed: 34896587
Matchin, w., Basilakos, A., Rígido, B. C., den Ouden, D.-B., Fridriksson,
J., & Hickok, GRAMO. (2020). Agrammatism and paragrammatism: A
cortical double dissociation revealed by lesion-symptom map-
ping. Neurobiology of Language, 1(2), 208–225. https://doi.org
/10.1162/nol_a_00010, PubMed: 34296193
Matchin, w., Brodbeck, C., Hammerly, C., & Lau, mi. (2019). El
temporal dynamics of structure and content in sentence compre-
hension: Evidence from fMRI-constrained MEG. Human Brain
Cartografía, 40(2), 663–678. https://doi.org/10.1002/hbm.24403,
PubMed: 30259599
Matchin, w., den Ouden, D.-B., Hickok, GRAMO., Hillis, A. MI., Bonilha,
l., & Fridriksson, j. (2022). The Wernicke conundrum revisited:
Evidence from connectome-based lesion-symptom mapping.
Cerebro, 145(11), 3916–3930. https://doi.org/10.1093/ brain
/awac219, PubMed: 35727949
Matchin, w., Hammerly, C., & Lau, mi. (2017). The role of the IFG and
pSTS in syntactic prediction: Evidence from a parametric study of
hierarchical structure in fMRI. Corteza, 88, 106–123. https://doi.org
/10.1016/j.cortex.2016.12.010, PubMed: 28088041
Matchin, w., & Hickok, GRAMO. (2020). The cortical organization of syn-
tax. Corteza cerebral, 30(3), 1481–1498. https://doi.org/10.1093
/cercor/bhz180, PubMed: 31670779
Matchin, w., İlkbaşaran, D., Hatrak, METRO., Roth, A., Villwock, A.,
Halgren, MI., & Mayberry, R. I. (2022). The cortical organization of
syntactic processing is supramodal: Evidence from American Sign
Idioma. Revista de neurociencia cognitiva, 34(2), 224–235.
https://doi.org/10.1162/jocn_a_01790, PubMed: 34964898
Matchin, w., Liao, C.-H., Gaston, PAG., & Lau, mi. (2019). Same words,
different structures: An fMRI investigation of argument relations
and the angular gyrus. Neuropsicología, 125, 116–128.
https://doi.org/10.1016/j.neuropsychologia.2019.01.019,
PubMed: 30735675
Matchin, w., & Rogalsky, C. (2023). Aphasia and syntax. In J.
Sprouse (Ed.), Handbook of experimental syntax. Oxford Univer-
sity Press. https://psyarxiv.com/m8pnd/
Matchin, w., & Wood, mi. (2020). Syntax-sensitive regions of the
posterior inferior frontal gyrus and the posterior temporal lobe
are differentially recruited by production and perception. Cere-
bral Cortex Communications, 1(1), Article tgaa029. https://doi
.org/10.1093/texcom/tgaa029, PubMed: 34296103
Menenti, l., Gierhan, S. METRO. MI., Segaert, K., & Hagoort, PAG. (2011).
Shared language: Overlap and segregation of the neuronal infra-
structure for speaking and listening revealed by functional MRI.
ciencia psicológica, 22(9), 1173–1182. https://doi.org/10
.1177/0956797611418347, PubMed: 21841148
Mesulam, M.-M., Thompson, C. K., Weintraub, S., & Rogalski, mi. j.
(2015). The Wernicke conundrum and the anatomy of language
comprehension in primary progressive aphasia. Cerebro, 138(8),
2423–2437. https://doi.org/10.1093/ brain/awv154, PubMed:
26112340
Meyer, l., & Friederici, A. D. (2016). Neural systems underlying the
processing of complex sentences. In G. Hickok & S. l. Pequeño
(Editores.), Neurobiology of language (páginas. 597–606). Elsevier.
https://doi.org/10.1016/B978-0-12-407794-2.00048-1
Mohr, j. PAG., Pessin, METRO. S., Finkelstein, S., Funkenstein, h. h.,
Duncan, GRAMO. w., & davis, k. R. (1978). Broca aphasia: Pathologic
and clinical. Neurología, 28(4), 311–324. https://doi.org/10.1212
/ WNL.28.4.311, PubMed: 565019
Neurobiology of Language
23
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
/
.
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
Momma, S., & Phillips, C. (2018). The relationship between parsing
y generación. Annual Review of Linguistics, 4(1), 233–254.
https://doi.org/10.1146/annurev-linguistics-011817-045719
Moro, A., Tettamanti, METRO., Perani, D., Donati, C., Cappa, S. F., &
Fazio, F. (2001). Syntax and the brain: Disentangling grammar
by selective anomalies. NeuroImagen, 13(1), 110–118. https://
doi.org/10.1006/nimg.2000.0668, PubMed: 11133314
Murphy, MI., Woolnough, o., Rollo, PAG. S., Roccaforte, z. J., Segaert,
K., Hagoort, PAG., & Tandon, norte. (2022). Minimal phrase composi-
tion revealed by intracranial recordings. Revista de neurociencia,
42(15), 3216–3227. https://doi.org/10.1523/ JNEUROSCI.1575
-21.2022, PubMed: 35232761
Nachev, PAG., Coulthard, MI., Jäger, h. r., Kennard, C., & Husain, METRO.
(2008). Enantiomorphic normalization of focally lesioned brains.
NeuroImagen, 39(3), 1215–1226. https://doi.org/10.1016/j
.neuroimage.2007.10.002, PubMed: 18023365
nelson, METRO. J., El Karoui, I., Giber, K., Cual, X., cohen, l.,
Koopman, h., Cash, S. S., Naccache, l., Hale, j. T., Pallier, C.,
& Dehaene, S. (2017). Neurophysiological dynamics of
phrase-structure building during sentence processing. Pro-
cesiones de
the National Academy of Sciences, 114(18),
E3669–E3678. https://doi.org/10.1073/pnas.1701590114,
PubMed: 28416691
NITRC. (2020). NiiStat [Software]. https://www.nitrc.org/projects
/niistat
Ocampo, B., & Kritikos, A. (2011). Interpreting actions: The goal
behind mirror neuron function. Brain Research Reviews, 67(1–2),
260–267. https://doi.org/10.1016/j.brainresrev.2011.03.001,
PubMed: 21396402
Pallier, C., Devauchelle, A.-D., & Dehaene, S. (2011). Cortical rep-
resentation of the constituent structure of sentences. Actas
of the National Academy of Sciences, 108(6), 2522–2527. https://
doi.org/10.1073/pnas.1018711108, PubMed: 21224415
Pettigrew, C., & Hillis, A. mi. (2014). Role for memory capacity in
sentence comprehension: Evidence from acute stroke. Aphasiol-
ogia, 28(10), 1258–1280. https://doi.org/10.1080/02687038.2014
.919436, PubMed: 25221377
Pillay, S. B., Binder, j. r., Humphries, C., Bruto, W.. l., & Book,
D. S. (2017). Lesion localization of speech comprehension defi-
cits in chronic aphasia. Neurología, 88(10), 970–975. https://doi
.org/10.1212/ WNL.0000000000003683, PubMed: 28179469
Pinker, S., & Ullman, METRO. t. (2002). The past and future of the past
tense. Tendencias en Ciencias Cognitivas, 6(11), 456–463. https://doi
.org/10.1016/S1364-6613(02)01990-3, PubMed: 12457895
Pulvermüller, F., & Fadiga, l. (2010). Active perception: Sensorimo-
tor circuits as a cortical basis for language. Nature Reviews Neu-
roscience, 11(5), 351–360. https://doi.org/10.1038/nrn2811,
PubMed: 20383203
Rilling, j. k. (2014). Comparative primate neurobiology and the
evolution of brain language systems. Current Opinion in Neuro-
biology, 28, 10–14. https://doi.org/10.1016/j.conb.2014.04.002,
PubMed: 24835547
Riva, METRO., wilson, S. METRO., Cai, r., Castellano, A., Jordán, k. METRO.,
Henry, R. GRAMO., Gorno Tempini, METRO. l., Berger, METRO. S., & Chang,
mi. F. (2022). Evaluating syntactic comprehension during awake
intraoperative cortical stimulation mapping. Journal of Neurosur-
gery, 138(5), 1403–1410. https://doi.org/10.3171/2022.8
.JNS221335, PubMed: 36208435
Rogalsky, C., Almeida, D., Sprouse, J., & Hickok, GRAMO. (2015). Oración
processing selectivity in Broca’s area: Evident for structure but
not syntactic movement. Idioma, Cognition and Neuroscience,
30(10), 1326–1338. https://doi.org/10.1080/23273798.2015
.1066831, PubMed: 27135039
Rogalsky, C., & Hickok, GRAMO. (2011). The role of Brocaʼs area in sen-
tence comprehension. Revista de neurociencia cognitiva, 23(7),
1664–1680. https://doi.org/10.1162/jocn.2010.21530, PubMed:
20617890
Rogalsky, C., LaCroix, A. NORTE., Chen, K.-H., anderson, S. w.,
Damasio, h., Amar, T., & Hickok, GRAMO. (2018). The neurobiology
of agrammatic sentence comprehension: A lesion study. Diario
of Cognitive Neuroscience, 30(2), 234–255. https://doi.org/10
.1162/jocn_a_01200, PubMed: 29064339
Rogalsky, C., Matchin, w., & Hickok, GRAMO. (2008). Broca’s area, sen-
tence comprehension, and working memory: Un estudio de resonancia magnética funcional.
Frontiers in Human Neuroscience, 2, Article 14. https://doi.org
/10.3389/neuro.09.014.2008, PubMed: 18958214
Rorden, C., & Karnath, H.-O. (2004). Using human brain lesions to
infer function: A relic from a past era in the fMRI age? Naturaleza
Reseñas Neurociencia, 5(10), 812–819. https://doi.org/10.1038
/nrn1521, PubMed: 15378041
Saffran, mi. METRO., Schwartz, METRO. F., & Marin, oh. S. METRO. (1980). The word
order problem in agrammatism: II. Production. Brain and Lan-
gu age, 10(2), 263 –280. https://doi.org/10 .10 16/0093
-934X(80)90056-5, PubMed: 7407547
Santi, A., & Grodzinsky, Y. (2007). Taxing working memory with
syntax: Bihemispheric modulations. Mapeo del cerebro humano,
28(11), 1089–1097. https://doi.org/10.1002/ hbm.20329,
PubMed: 17133392
Sapolski, D., Bakkour, A., Negreira, A., Nalipinski, PAG., Weintraub,
S., Mesulam, M.-M., Caplan, D., & Dickerson, B. C. (2010). Cor-
tical neuroanatomic correlates of symptom severity in primary
progressive aphasia. Neurología, 75(4), 358–366. https://doi.org
/10.1212/ WNL.0b013e3181ea15e8, PubMed: 20660866
Schwartz, METRO. F., Saffran, mi. METRO., & Marin, oh. S. METRO. (1980). The word
order problem in agrammatism. I. Comprehension. Brain and
Idioma, 10(2), 249–262. https://doi.org/10.1016/0093
-934X(80)90055-3, PubMed: 7407546
Shain, C., Blank, I. A., van Schijndel, METRO., Schuler, w., & Fedorenko,
mi. (2020). fMRI reveals language-specific predictive coding dur-
ing naturalistic sentence comprehension. Neuropsicología,
138, Article 107307. https://doi.org/10.1016/j.neuropsychologia
.2019.107307, PubMed: 31874149
Stanojević, METRO., Brennan, j. r., Dunagan, D., Steedman, METRO., & Hale,
j. t. (2023). Modeling structure-building in the brain with CCG
parsing and large language models. Ciencia cognitiva, 47(7),
Article e13312. https://doi.org/10.1111/cogs.13312, PubMed:
37417470
Stromswold, K., Caplan, D., alperto, NORTE., & Rauch, S. (1996). Local-
ization of syntactic comprehension by positron emission tomog-
raphy. Brain and Language, 52(3), 452–473. https://doi.org/10
.1006/brln.1996.0024, PubMed: 8653390
Studebaker, GRAMO. A., 1985. A “rationalized” arcsine transform. Diario
of Speech, Idioma, and Hearing Research, 28(3), 455–462.
https://doi.org/10.1044/jshr.2803.455, PubMed: 4046587
Swinney, D., & Zurif, mi. (1995). Syntactic processing in aphasia.
Brain and Language, 50(2), 225–239. https://doi.org/10.1006
/brln.1995.1046, PubMed: 7583188
Thompson, C. K., Lange, k. l., Schneider, S. l., & Shapiro, l. PAG.
(1997). Agrammatic and non-brain-damaged subjects’ verb
and verb argument structure production. Aphasiology, 11(4–5),
473–490. https://doi.org/10.1080/02687039708248485
Thompson, C. K., Meltzer-Asscher, A., Dar, S., Sotavento, J., Wieneke, C.,
Weintraub, S., & Mesulam, M.-M. (2013). Syntactic and morpho-
syntactic processing in stroke-induced and primary progressive
aphasia. Behavioural Neurology, 26(1–2), 35–54. https://doi.org
/10.3233/BEN-2012-110220, PubMed: 22713394
Neurobiology of Language
24
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
.
/
/
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grammatical parallelism in aphasia
Thompson, C. K., & Shapiro, l. (2005). Treating agrammatic apha-
sia within a linguistic framework: Treatment of underlying forms.
Aphasiology, 19(10–11), 1021–1036. https://doi.org/10.1080
/02687030544000227, PubMed: 17410280
Thothathiri, METRO., Kimberg, D. y., & Schwartz, METRO. F. (2012). The neu-
ral basis of reversible sentence comprehension: Evidencia de
voxel-based lesion symptom mapping in aphasia. Journal of Cog-
nitive Neuroscience, 24(1), 212–222. https://doi.org/10.1162
/jocn_a_00118, PubMed: 21861679
tyler, l. K., Marslen-Wilson, W.. D., Randall, B., Wright, PAG., Devereux,
B. J., Zhuang, J., Papoutsi, METRO., & Stamatakis, mi. A. (2011). Left infe-
rior frontal cortex and syntax: Function, structure and behaviour in
patients with left hemisphere damage. Cerebro, 134(2), 415–431.
https://doi.org/10.1093/brain/awq369, PubMed: 21278407
tyler, l. K., Shafto, METRO. A., Randall, B., Wright, PAG., Marslen-Wilson,
W.. D., & Stamatakis, mi. A. (2010). Preserving syntactic processing
across the adult life span: The modulation of the frontotemporal
language system in the context of age-related atrophy. Cerebral
Corteza, 20(2), 352–364. https://doi.org/10.1093/cercor/bhp105,
PubMed: 19505991
von Humboldt, W.. (1836). On language. Cambridge University
Prensa.
Wernicke, C. (1874). The symptom complex of aphasia: A psycho-
logical study on an anatomical basis. En R. S. cohen & METRO. W..
Wartofsky (Editores.), Boston studies in the philosophy of science
(páginas. 34–97). D. Reidel Publishing Company. https://doi.org/10
.1007/978-94-010-3378-7_2
wilson, S. METRO., Dronkers, norte. F., Ogar, j. METRO., Jang, J., Growdon, METRO. MI.,
Agosta, F., Henry, METRO. l., Molinero, B. l., & Gorno-Tempini, METRO. l.
(2010). Neural correlates of syntactic processing in the nonfluent
variant of primary progressive aphasia. Revista de neurociencia,
30(50), 16845–16854. https://doi.org/10.1523/JNEUROSCI.2547
-10.2010, PubMed: 21159955
wilson, S. METRO., Galantucci, S., Tartaglia, METRO. C., Rising, K., Patterson,
D. K., Henry, METRO. l., Ogar, j. METRO., DeLeon, J., Molinero, B. l., &
Gorno-Tempini, METRO. l. (2011). Syntactic processing depends on
dorsal language tracts. Neurona, 72(2), 397–403. https://doi.org
/10.1016/j.neuron.2011.09.014, PubMed: 22017996
wilson, S. METRO., Henry, METRO. l., Besbris, METRO., Ogar, j. METRO., Dronkers,
norte. F., Jarrold, w., Molinero, B. l., & Gorno-Tempini, METRO. l. (2010).
Connected speech production in three variants of primary pro-
gressive aphasia. Cerebro, 133(7), 2069–2088. https://doi.org/10
.1093/brain/awq129, PubMed: 20542982
wilson, S. METRO., & Saygın, A. PAG. (2004). Grammaticality judgment in
aphasia: Deficits are not specific to syntactic structures, aphasic
syndromes, or lesion sites. Revista de neurociencia cognitiva,
16(2), 238–252. https://doi.org/10.1162/089892904322984535,
PubMed: 15068594
Wulfeck, B., & Bates, mi. (1991). Differential sensitivity to errors of
agreement and word order in Broca’s aphasia. Journal of Cogni-
tive Neuroscience, 3(3), 258–272. https://doi.org/10.1162/jocn
.1991.3.3.258, PubMed: 23964841
Yagata, S. A., Yen, METRO., McCarron, A., Bautista, A., Lamair-Orosco,
GRAMO., & wilson, S. METRO. (2017). Rapid recovery from aphasia after
infarction of Wernicke’s area. Aphasiology, 31(8), 951–980.
https://doi.org/10.1080/02687038.2016.1225276, PubMed:
29051682
Zaccarella, MI., Meyer, l., Makuuchi, METRO., & Friederici, A. D. (2017).
Building by syntax: The neural basis of minimal linguistic struc-
turas. Corteza cerebral, 27(1), 411–421. https://doi.org/10.1093
/cercor/bhv234, PubMed: 26464476
Zaccarella, MI., Schell, METRO., & Friederici, A. D. (2017). Reviewing the
functional basis of the syntactic Merge mechanism for language:
A co o r d i n a t e – b a s e d a c t i v a t i o n l i k e l i h o o d e s t i m a t i o n
meta-analysis. Neurociencia & Revisiones de biocomportamiento, 80,
646–656. https://doi.org/10.1016/j.neubiorev.2017.06.011,
PubMed: 28743620
Zurif, mi. B. (1980). Language mechanisms: A neuropsychological
perspectiva: The effects of focal brain damage on the processing
of syntactic elements may provide an important clue to the man-
ner in which language is organized in the brain. American Scien-
tist, 68(3), 305–311. PubMed: 7406311
Zurif, mi. B., Caramazza, A., & Myerson, R. (1972). Grammatical
judgments of agrammatic aphasics. Neuropsicología, 10(4),
405–417. https://doi.org/10.1016/0028-3932(72)90003-6,
PubMed: 4657523
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
yo
/
/
.
1
0
1
1
6
2
norte
oh
_
a
_
0
0
1
1
7
2
1
5
5
7
0
0
norte
oh
_
a
_
0
0
1
1
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurobiology of Language
25