ARTÍCULO DE INVESTIGACIÓN
Predictive Coding and Internal Error Correction in
Speech Production
un acceso abierto
diario
Alex Teghipco1
, Kayoko Okada2, Emma Murphy2, and Gregory Hickok1
1Department of Cognitive Sciences, Universidad de California, Irvine, California, EE.UU
2Department of Psychology, Loyola Marymount University, Los Angeles, California, EE.UU
Palabras clave: speech production, resonancia magnética funcional, internal error correction, predictive coding, internal models,
imagined speech, overt speech, nonwords, taboo words, tongue twisters
ABSTRACTO
Speech production involves the careful orchestration of sophisticated systems, yet overt
speech errors rarely occur under naturalistic conditions. The present functional magnetic
resonance imaging study sought neural evidence for internal error detection and correction by
leveraging a tongue twister paradigm that induces the potential for speech errors while
excluding any overt errors from analysis. Previous work using the same paradigm in the
context of silently articulated and imagined speech production tasks has demonstrated forward
predictive signals in auditory cortex during speech and presented suggestive evidence of
internal error correction in left posterior middle temporal gyrus (pMTG) on the basis that this
area tended toward showing a stronger response when potential speech errors are biased
toward nonwords compared to words (Okada et al., 2018). The present study built on this prior
work by attempting to replicate the forward prediction and lexicality effects in nearly twice as
many participants but introduced novel stimuli designed to further tax internal error correction
and detection mechanisms by biasing speech errors toward taboo words. The forward
prediction effect was replicated. While no evidence was found for a significant difference in
brain response as a function of lexical status of the potential speech error, biasing potential
errors toward taboo words elicited significantly greater response in left pMTG than biasing
errors toward (neutral) palabras. Other brain areas showed preferential response for taboo words
as well but responded below baseline and were less likely to reflect language processing as
indicated by a decoding analysis, implicating left pMTG in internal error correction.
INTRODUCCIÓN
Speaking is a deceptively complex task involving several computational stages: selecting
words from a mental dictionary that can contain tens of thousands of entries, correctly acces-
sing and coding the sequence of sounds that could have many possible permutations, y
executing the motor commands to reproduce those sounds with coordinated movements of
several independent articulators within the vocal tract (Levelt, 1989). Given the system’s com-
plejidad, the ample opportunities for error, and the fact that speech is articulated at a rate of
approximately five syllables per second (Jacewicz et al., 2010), it is remarkable that the vast
majority of words are accurately produced (estimates put the number at approximately 99.9%;
ver, p.ej., Garnham et al., 1982; Levelt, 1992). How is this achieved? One possibility is that the
mechanism is so exquisitely tuned that it simply makes very few coding errors. Another pos-
sibility is that coding errors occur more frequently, but are unconsciously detected and
Citación: Teghipco, A., Okada, K.,
Murphy, MI., & Hickok, GRAMO. (2023).
Predictive coding and internal error
correction in speech production.
Neurobiology of Language, 4(1),
81–119. https://doi.org/10.1162
/nol_a_00088
DOI:
https://doi.org/10.1162/nol_a_00088
Recibió: 16 Marzo 2022
Aceptado: 2 Noviembre 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Alex Teghipco
ateghipc@uci.edu
Editor de manejo:
Kate Watkins
Derechos de autor: © 2023
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
Forward predictive coding:
The idea that motor plans lead to
predictions of sensory consequences,
facilitating error detection and
correction in motor control.
Internal (speech) error correction:
The correction of a speech error that
is committed prior to speech output.
Functional magnetic resonance
imaging (resonancia magnética funcional):
Neuroimaging method used to detect
changes in blood flow and oxygen in
the brain.
Lexicality effect:
Stronger neural response when
recitation is biased to increase the
potential for errors to be nonwords
than words.
Posterior middle temporal gyrus
(pMTG):
Brain region crucial for lexical
Procesando.
corrected internally, prior to ever being spoken (Hickok, 2012; Levelt, 1983; Nozari et al.,
2011). The latter possibility is consistent with the idea from the motor control literature that
the brain simulates the position and trajectory of the motor effector it is controlling, a so-called
forward internal model, as a mechanism for improving the speed and accuracy of movements
via predictive coding (Loco, 1999; Shadmehr & Krakauer, 2008; Wolpert et al., 1995). Enterrar-
nal predictive coding may provide a mechanism to detect and correct speech errors prior to
producing them (Hickok, 2012).
Although talkers can detect and correct overtly produced speech errors, evidence for inter-
nal error correction in speech is limited to inferences based on the timing of error corrections.
Por ejemplo, overtly realized error corrections, such as “v-horizontal,” have been argued to
occur too quickly to be accomplished using overt feedback alone, which in turn suggests the
existence of at least an internal mechanism to detect, if not correct errors (Nooteboom, 2005;
Nozari et al., 2011). Further evidence for internal error detection comes from electrophysio-
logical measures, which have identified signals that predict speech errors prior to their vocal-
ización (Möller et al., 2007). Strong direct evidence for internal error correction is sparse,
sin embargo.
In one fMRI study, Okada et al. (2018) reported suggestive evidence for the existence of an
internal error correction mechanism. In their experiment, the authors presented participants
with tongue twister sequences that were designed to bias speech errors toward either words
(REEF LEECH ➔ LEAF REACH) or nonwords ( WREATH LEAGUE ➔ LEATH REEG), a paradigm
previously investigated by Oppenheim and Dell (2008). Behavioral research on slips of the
tongue has shown that nonword errors are more rare than real word errors, the “lexical bias”
efecto (Baars et al., 1975; Dell, 1986; Levelt et al., 1999; Nooteboom, 2005). Several previous
behavioral studies have shown that the lexical bias effect holds even when subjects do not
phonate their speech and self-report their errors (Corley et al., 2011; Oppenheim & Dell,
2008, 2010). Although not the main focus of their study, a lexicality effect was reported by
Okada et al. (2018) in the left posterior middle temporal gyrus (pMTG), a region implicated
in lexical-level processes. Eso es, Okada and colleagues found greater activation in MTG
when participants recited tongue twisters that were biased to produce nonword errors com-
pared to word-errors, even on trials in which participants responded accurately. This is par-
ticularly interesting because no speech errors were committed on these trials—the activation
differences observed in MTG reflect the potential for a word versus nonword error. El
authors suggested that speech errors resulting in nonwords are more readily detectable and
therefore would be more likely to be internally corrected prior to speaking. The fact that they
found a clear effect of lexical status of the error bias even under conditions of accurate per-
formance demonstrates that the system detected the distinction internally, and this could be
the case only if in fact an internal error was committed and then corrected prior to accurate
producción. Although this is a very interesting finding, the pMTG activation in their study did not
reach statistical significance corrected for multiple comparisons, and as the authors note, este
work requires replication. Runnqvist et al. (2020) recently reported on a study that used a very
similar design to Okada et al. (2018), and while they reported evidence of internal error cor-
rection in the cerebellum, they did not detect an effect in the cerebral cortex.
The present research builds on this prior work by attempting a direct replication of Okada
et al. (2018) along with an extension to potentially render the paradigm more sensitive to
detecting evidence for an internal error correction mechanism. The experiment we introduce
uses the same tongue twister stimulus list and the same tasks (silent articulation and imaging)
as previous fMRI work, but additionally includes a set of stimuli designed to increase error
salience and therefore error detection and correction, thereby increasing our chances of
Neurobiology of Language
82
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
observing evidence for such a process. Previous work has shown that tongue twisters with the
potential to induce taboo word slips (p.ej., FULL BUD BUCK FUSS) elicit significantly fewer
slips than neutral tongue twisters, suggesting a higher rate of internal error detection and cor-
rection (Motley et al., 1982). We hypothesized that tongue twisters biased toward taboo word
errors would elicit greater activation in pMTG lexical networks compared to non-taboo word
errores. The basis for this is that detecting and correcting word-level speech errors presumably
drives activity to areas critical for lexical processing, which implicates pMTG (p.ej., Indefrey,
2011; Lau & Namyst, 2019), and that prior work has found suggestive evidence for internal
error correction in pMTG using tongue twisters (Okada et al., 2018; a possible mechanism of
this effect is explored in the Discussion section).
In addition to mapping an error correction mechanism in the brain, we aim to replicate
Okada et al.’s (2018) finding that motor-driven forward predictive signals are present in audi-
tory cortex in a speech production task. In their study, Okada et al. (2018) tuvo participantes
silently recite a sequence of tongue twisters in an fMRI experiment. Two speech production
conditions were included, one in which speech was articulated without phonating (silent artic-
ulación) and one in which speech production was imagined without articulation (imagined).
Both conditions were matched for acoustic input (es decir., no speech input). Previous behavioral
research has shown that these two tasks engage different levels of linguistic/motor planning.
Imagined speech engages lexical-level processes but not lower-level phonological processes
whereas silently articulating speech engages both levels of processing. Por lo tanto, engaging
motor-phonological processes should generate a forward prediction of the acoustic conse-
quences of the executed (silent) speech, whereas engaging lexical-level processes should
no. As expected, a contrast of silently articulated speech compared to imagined speech
revealed activity in left inferior frontal gyrus (IFG) and premotor cortex, areas involved in
speech articulation. More interestingly, they found robust activity in bilateral auditory cortex
when motor articulators were engaged, but not when speech was imagined, and this activation
was present even though there was no external auditory stimulation. The authors suggest that
these activations reflect stronger forward predictions generated in the articulation condition
compared to the imagining condition (Levelt, 1989).
The main goal of the present research is to examine a speech production mechanism that
has been elusive thus far: neural evidence of internal error correction. Con ese fin, we leverage
a tongue twister paradigm that has been previously used to generate suggestive evidence of
internal error correction during speech in the pMTG. Using the same stimulus set and design
as this prior work, we attempt to replicate evidence for internal error correction in a much
larger sample of participants. críticamente, we also improve our odds of finding evidence of inter-
nal error correction by introducing additional stimuli designed to tax the error correction
mechanism: tongue twisters that elicit taboo word errors. We also attempt to replicate the for-
ward predictive signal effect reported by this previous work. Replication is particularly impor-
tant since the aforementioned study was the first fMRI experiment to show evidence of forward
predictive signals involving auditory cortex, a sensory region that plays an important role in
speech production.
MATERIALES Y MÉTODOS
Participantes
Forty participants (25 hembras) entre 18 y 40 years of age were recruited from the Uni-
versity of California, Irvine (UCI) comunidad. Participants received monetary compensation
por su participación. The volunteers were right-handed, native English speakers with normal
Neurobiology of Language
83
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
Taboo effect:
Stronger neural response when
recitation is biased to increase the
potential for errors to be taboo words
than non-taboo words.
or corrected-to-normal vision, no known history of neurological disease, and no contraindica-
tions for MRI. Informed consent was obtained from each participant prior to participation in the
study in accordance with guidelines from the local ethics committee at UCI that approved this
estudiar. A handful of participants were excluded from analysis for excessive head motion (norte = 2)
and unanticipated scanning termination for a variety of reasons (norte = 4; es decir., claustrophobia,
equipment malfunction, excessive tardiness to the point of being unable to collect more than
a single fMRI session), leaving a total of 34 participants to contribute to the results. Four par-
ticipants from the group were not able to complete all nine scanning sessions but completed
89% (norte = 2; 8 sessions), 77% (norte = 1; 7), y 66% (norte = 1; 6) of the fMRI sessions.
The number of participants recruited for this study was anticipated based on a power anal-
ysis of pilot data collected for four participants. The power analysis was carried out over two
regiones de interés (ROI) using the fMRIpower toolbox (Mumford & Nichols, 2008). Anatom-
ical ROIs for this analysis were selected from the Harvard–Oxford cortical atlas (Desikan et al.,
2006), and adjustment for Type 1 error was made by applying a Bonferroni correction based
on the number of ROIs being compared ( pag < 0.005). The power analysis indicated that
approximately 40 participants were necessary to achieve 80% power for detecting a forward
prediction effect within Heschl’s gyrus (effect size of 1.0623; effect sizes expressed in standard
deviation units, which is analogous to Cohen’s d ) and internal error correction effects in tem-
porooccipital middle temporal gyrus (toMTG; effect size of 0.3815 for a lexicality effect and
0.3985 for taboo effect). We note that a smaller sample size of 34 was indicated to achieve
72% power for detecting these effects. See the Data Analysis section for more information
about how these effects were measured. Temporooccipital middle temporal gyrus was used
for the power analysis instead of pMTG because this anatomical area aligned more closely
with the foci reported by the study we sought to replicate (Okada et al., 2018).
Stimuli and Task
Scanning took place at the Facility for Imaging and Brain Research at UCI. Participants were
scanned while they recited a set of four words (e.g., lean reed reef leach) in sync with a visual
metronome. Thirty-two sets of tongue twisters used in previous experiments were employed in
the current study (Oppenheim & Dell, 2010). These tongue twisters are known to behaviorally
elicit a lexical bias effect. Lexical bias refers to the tendency for word errors to create a real
word instead of a nonword (e.g., target word is “reef” but slips to “leaf,” is more likely than if
target word is “wreath” and slips to “leath” because leath is a nonword). These stimuli were
designed so that if an error occurred on the third or fourth word of each sequence, the out-
come would yield either a real word error (e.g., “leaf”) or a nonword error (e.g., “leath”). In
addition to these tongue twisters, we included 32 taboo tongue twisters (see Table 1 for exam-
ple of stimuli). In a behavioral pilot involving 28 participants (15 females), we found that these
taboo tongue twisters were effective in eliciting speech errors (non-taboo error rate = 23%,
taboo word error rate = 20%).
The present study followed the experimental procedure outlined in Okada et al. (2018). On
each trial, a tongue twister phrase was visually presented on screen for 3 s, and then subjects
were cued to silently articulate the sequence or imagine saying the sequence without mouth
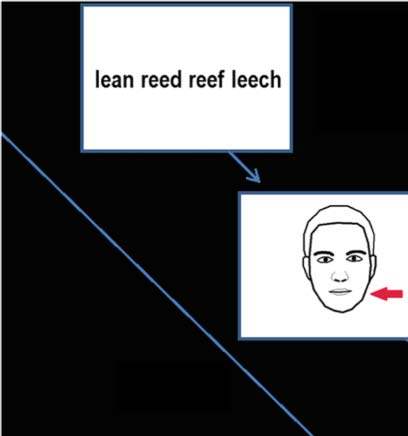
movements (see Figure 1). The presented cue was a cartoon face that remained on screen for
500 ms and contained a red arrow pointing either to the head or to the lips. An arrow pointing
to the head cued the participants to imagine saying the word, and an arrow pointing to the lips
cued the participants to silently articulate the words. A red fixation appeared on screen 500 ms
after cue offset and served as the visual metronome, flashing at a rate of 2/s. Participants
Neurobiology of Language
84
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Predictive coding and error correction
Table 1.
Stimulus set
Non-taboo tongue twister
nod mod mock knob
mine bikes bit mice
bike wild wise bile
bane gave gan bait
name make mail nag
wing bib bit whip
six finch fill sin
jail cheek cheap jean
lean reed reef leech
yore wan wok yawn
gun bulb buck gull
singe fib fish sip
sing hitch his sick
zinc niece need zest
jog mod mock job
job rob rock jot
van match mat verve
lull nudge buck love
zing bib bit zip
hinge fib fit hip
sing that them zed
daft gab gas dam
than bunk nuzz there
chicks fich fizz chin
king hitch his kcik
nab match mat nerve
gun bulb but gull
pen bunk nus pair
zing that then zed
gore wan watt gone
goon nab nap gar
nun bulb but null
Neurobiology of Language
Taboo tongue twister
cod mod mock cob
dine bite bike dice
dial bile bike dies
shave bane bit shank
fake name nag fail
shine bib bit ship
ding sin sick dill
dale chip chick dean
queen reed reef queer
core mud mum caught
fun bulb buck full
nib singe sip knit
ditch sing sick diss
pink niece need pest
cot mod mock cob
call rob rock cot
shack match mat shave
full bud buck fun
ting bib bit tip
shin fib fit ship
shing them that shed
dab laugh lamb dan
jan bid bizz jar
jicks finch fizz gin
ping hitch hiss pick
shab volt vat shot
con grub grunt cup
fend bus buck fair
shang then that shed
whole gone gore hat
food tab tuck far
pun mull miss pull
85
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Predictive coding and error correction
Example of a single trial. Participants were presented with a tongue twister sequence,
Figure 1.
which remained on screen for 3 s, followed by a cue to either articulate the sequence or imagine the
sequence. They recited each word in sync with the visual metronome.
recited one word per fixation in sync with the metronome. The interstimulus interval was
500 ms. After recitation, participants indicated with a button press if they were correct or incor-
rect on the sequence using their left hand. Recall failure was treated as an incorrect response
to ensure that only trials where rehearsal was successful (i.e., error free) would be analyzed.
Participants were also instructed to indicate an incorrect response if overt production occurred
accidentally. Prior to scanning, participants spent roughly 5 min (more when necessary) prac-
ticing silently reciting words in a way that minimized but did not eliminate articulatory move-
ments. Continuous feedback was provided by lab staff during this practice period.
A single trial in the experiment was 8 s in length and there were approximately 42 trials in
each session. Each session consisted of an equal number of tongue twister phrases biased to
produce word errors or nonword errors and taboo errors. There were eight experimental ses-
sions and each session consisted of approximately 14 trials of each type, which were ran-
domly presented along with six rest trials (fixation). The study started with a high-resolution
structural scan. This was followed by a short practice session of the experiment using approx-
imately 10 trials to further familiarize subjects with the task. Scanning was conducted during
the practice session to acclimatize subjects to the fMRI environment, as well as to monitor
head movement and provide feedback prior to the start of the experiment. Participants also
received feedback about head movement in between sessions based on qualitative assessment
of the images that had been collected. The study lasted approximately 1.5 hr. Stimulus pre-
sentation and timing was controlled using PsychToolbox (Borgo et al., 2012) implemented in
MATLAB (Mathworks, 2022).
Imaging
Imaging data were collected on a 3T Siemens Prisma scanner (Siemens Medical Solutions,
2022) equipped with a 32-channel RF receiver head coil. A single T1-weighted MPRAGE
sequence was acquired (matrix = 256 × 256 mm, TR = 2.3 s, TE = 2.32 ms, flip angle = 8°,
size = 0.937 × 0.937 × 0.9 mm). An echo-planar imaging (EPI) pulse sequence was collected
for each of the eight experimental sessions and the practice session (matrix = 100 × 100 mm,
TR = 2 s, TE = 35 ms, flip angle = 90°, size = 2.4 × 2.4 × 2.4 mm, 56 slices).
Data Analysis
Data were preprocessed and analyzed using the FMRIB’s Software Library (FSL; Jenkinson
et al., 2012). First, rigid-body motion correction was performed with FSL’s intramodal motion
Neurobiology of Language
86
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Predictive coding and error correction
correction tool (MCFLIRT) using the normalized correlation cost function and the middle vol-
ume as the initial template (Grabner et al., 2006). Participants with excessive head movement
(>0.3 mean framewise displacement, as defined by Power et al., 2012, across sessions) eran
eliminated from further analysis. Mean framewise displacement among the remaining partic-
ipants (norte = 34) was relatively low (m = 0.15, DE = 0.07). Echo planar images were high pass
filtered by calculating the minimal period that retains 90% of the variance in the design matrix
regressors (this amounts to a roughly 0.01 hz cutoff ). These images were then spatially
smoothed using an isotropic 8-mm full width half maximum (FWHM) gaussian, and the ana-
tomical image for each subject was coregistered to their middle EPI volume. Data analysis was
performed with FSL’s fMRI Expert Analysis Tool (Jenkinson et al., 2012) and proceeded in three
steps: (i) modeling within-session parameter estimates for events of interest using fixed effects,
(ii) using these parameter estimates in a between-session analysis to model participant mean
respuesta, y (iii) using mean participant response in a between-subjects analysis to model
group response using mixed effects (FSL’s FLAME1+2). Parameter estimates for each partici-
pant were transformed into standardized space using the MNI152 template (Jenkinson et al.,
2002). All trials on which participants indicated making speech errors were excluded from
análisis. De término medio, participants reported errors on 14% of trials (DE = 8%). Error rates were
comparable across different types of tongue twister (biased toward word errors: m = 15.3%,
DE = 10%; biased toward nonword errors: m = 15.6%, DE = 9.1%; biased toward taboo
errores: m = 12.4%, DE = 7.8%).
Regressors for events of interest were created by convolving the predictor variables repre-
senting the time course of stimulus presentation with a gamma variate function. As in the
previous experiment, regressors modeled the following experimental trial types: “Articulation:
Nonword Errors,” “Articulation: Word Errors,” “Articulation: Taboo Errors,” “Imagining: No-
word Errors,” “Imagining: Word Errors,” “Imagining: Taboo Errors.” All trials on which partic-
ipants reported making an incorrect response, the visual presentation of words on all trials,
and the six motion parameter estimates determined during the realignment stage of prepro-
cessing were included in the model as nuisance regressors. Regressors were used to generate
parameter estimates for each condition of speech task (es decir., imagined and silently articulated)
and error type (es decir., tongue twisters biased toward: nonword, palabra, and taboo word errors).
Parameter estimates for each speech task modeled tongue twisters of all error types and
parameter estimates for each error type modeled both silently articulated and imagined
tongue twisters. Multiple contrasts were set up using these parameter estimates. As the pri-
mary goal of the present research was to seek evidence for internal error correction, we first
tested for the lexicality effect described by Okada et al. (2018), followed by a taboo effect
based on the new stimuli introduced in this study, and finally the forward prediction effect
also described by Okada et al. (2018). Identical to prior work, the lexicality effect was based
on the contrast between parameter estimates for the nonword and word tongue twister con-
ditions. The same logic was extended to the novel taboo stimuli and the taboo effect was
based on the contrast between taboo and word tongue twister conditions. The forward pre-
diction effect was based on the contrast between the two speech task conditions (es decir., silently
articulated vs. imagined). For each of these three contrasts an additional analysis was per-
formed contrasting each condition in the pair against baseline fixation. The contrasts between
each condition and baseline fixation were used to mask the contrast maps, allowing us to
distinguish brain areas that show a significant difference in BOLD response between condi-
tions but overall show below-baseline response.
Consistent with previous research, we expected to find taboo and lexicality effects in
pMTG, and a forward predictive signal effect in auditory cortex. These hypotheses target
Neurobiology of Language
87
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
specific regions and as such we complemented whole-brain analyses with a ROI approach
that compared mean condition-level parameter estimates used in the whole brain contrast
analyses within predefined anatomical areas. The same ROI analysis was carried out across
and within participants, the latter of which allowed us to characterize the consistency of the
effects we sought. Although the ROI analyses directly tested our hypotheses and provided
complementary information to the whole-brain contrast analysis approach, we elected to place
relatively greater focus on the whole-brain analysis as it allowed us to more comprehensively
test which brain areas are involved in the effects we were interested in (es decir., including those
areas not implicated in prior work), as well as to determine more precisely where effects within
anatomical ROIs were observed. Left and right hemisphere anatomical ROIs were extracted
from the Harvard–Oxford atlas. The anatomically defined pMTG area was used to test for
the lexicality and taboo effects, and Heschl’s gyrus was used to test for the forward prediction
efecto. The ROI from the power analysis was substituted based on the results of the whole brain
analiza. To foreshadow our findings, these analyses revealed a taboo effect consistent with
the location of the effect reported by Okada et al. (2018), but more clearly concentrated within
anatomical pMTG than toMTG. Group effects within ROIs were tested with paired t tests car-
ried out across participants. Tests were additionally carried out across voxels for each partic-
ipant to characterize the consistency of the effects. Minimum significance in the ROI analyses
was based on a Bonferroni corrected p-value threshold of 0.05, and minimum significance for
whole brain voxelwise testing was based on a voxelwise p-value threshold of 0.01 (es decir., grupo-
forming Z-threshold of 2.6) and a GRF-based cluster size p-value threshold of 0.05.
In addition to the core analyses detailed above, we present post hoc analyses that give fur-
ther context for some of our findings. We emphasize that these secondary analyses are inci-
dental by separating them from the main results section. The aim of the post hoc analyses is to
better functionally characterize the network of regions associated with the internal error cor-
rection effects (es decir., lexicality and taboo effects), particularly the wider network capturing
regions where neural response increased during the nonword or taboo conditions relative to
the word condition but overall remained below-baseline. Functions associated with this net-
work and with each of its constituent regions were probed using the Neurosynth meta-analytic
database, which associates word frequencies in the abstracts of studies with their activation
foci, enabling meta-analysis of groups of studies that frequently use particular terms (Yarkoni
et al., 2011). Network-level decoding was performed by computing the Pearson correlation
coefficient between contrast activation maps from the current study and each of the meta-
analyses that were generated for the 3,228 terms frequently used in the neuroimaging literature
and embedded in Neurosynth. Brevemente, performing a meta-analysis for each term involved sep-
arating all studies in Neurosynth into two groups: those that used a particular term frequently
(minimum rate of 1/1,000 words which has been shown to control for incidental word usage;
Yarkoni et al., 2011) and those that did not. Próximo, a search was performed for voxels where
activity was more consistently reported in the set of studies that do frequently use the term
relative to those that do not. This was accomplished by extracting the activation tables from
these two groups of studies, creating contingency tables at each voxel that described whether
activity was present and whether a phrase was used, and then performing a chi-square test.
Due to their smaller size, regions were decoded in a slightly different way—by computing the
mean posterior probability that a phrase was used within a study if activity was observed in
each of a regions’ voxels. Posterior probability estimates assumed a uniform prior (es decir., todo
terms are equally likely to appear) and were generated for each term in Neurosynth. Más
complete details about how posterior probability was computed can be found in prior work
(Yarkoni et al., 2011).
Neurobiology of Language
88
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
RESULTADOS
Core Findings
Internal error correction effects
Our first analysis aimed to replicate the lexicality effect described in previous work (Okada
et al., 2018). críticamente, all analyses we present were restricted to error-free trials. A group-level
contrast between the nonword and word conditions of the experiment revealed no significant
differences in brain response at our minimal significance thresholds (Z > 2.6 or p < 0.01; clus-
ter corrected at p < 0.05). We tested for effects on the cusp of significance by avoiding cluster
correction but did not find any significant differences in brain response. Lowering the cluster-
forming threshold further (Z > 2.3) did not reveal any areas where response to nonwords was
greater than response to words. Además, we tested whether the mean parameter estimates
within the pMTG ROI differed between these two conditions but found no significant differ-
ence across participants in either the left hemisphere (parameter estimates for words: m = 9.59,
DE = 41.84; parameter estimates for nonwords: m = 8.37, DE = 39.4; t(33) = 0.56, pag = 0.58) o
the right hemisphere (parameter estimates for words: M = −4.3, DE = 32.18; parameter esti-
mates for nonwords: M = −7.5, DE = 28.26; t(33) = −1.35, pag = 0.19). Because we failed to
replicate the lexicality effect reported in previous work (es decir., no effect was observed at the
group-level), we did not investigate how consistently the effect appeared within participants.
We next evaluated whether the novel taboo stimuli generated evidence for internal error
correction by contrasting the whole-brain parameter estimates for the taboo and word condi-
ciones. As we anticipated, the taboo condition appears to have successfully increased the load
on internal error detection and correction. Although no areas of the brain showed significantly
greater response for the word condition than the taboo condition, relatively higher response for
the taboo condition was found in a wide network that included the pMTG bilaterally (Z > 2.6
or p < 0.01, cluster corrected at p < 0.05 with a minimum significant cluster size of 803 voxels;
see Figure 2A). A more comprehensive description of brain regions in this network was pro-
vided by registering the contrast map to anatomical areas of the Harvard–Oxford cortical atlas
(Table 3). Incidental overlap between the contrast map and anatomical areas as a result of
activity spilling over an anatomical boundary in a way that is inappreciable was deempha-
sized by focusing only on those areas in which more than 5% of voxels showed a significant
difference between conditions. For visualization, both the atlas and the contrast map were
projected onto the fsaverage inflated cortical surface using a recently developed procedure
that implements registration fusion with advanced normalization tools to improve projection
accuracy (Wu et al., 2018; Figure 2B). The network of regions that responded more strongly to
the taboo than the word condition spanned the bilateral frontal poles (FPs), bilateral frontal
medial cortex (FMC), bilateral superior frontal gyrus (SFG), right anterior cingulate gyrus
(aCG), bilateral posterior cingulate gyrus (pCG), bilateral paracingulate gyrus (paraCG), right
subcallosal cortex, bilateral precuneous cortex (preCC), bilateral anterior middle temporal
gyrus (aMTG), bilateral pMTG, left posterior inferior temporal gyrus (pITG), bilateral angular
gyrus (AG), and bilateral superior lateral occipital cortex (sLOC; Figure 2).
The regions that responded more strongly to the taboo than the word condition were then
evaluated based on whether they showed above-baseline response during the taboo condi-
tion. Our first approach was to mask the contrast map between taboo and word conditions
by the contrast map for the taboo condition (i.e., taboo > word AND taboo > baseline; ambos
contrasts set to Z > 2.6 or p < 0.01, cluster corrected at p < 0.05). The resulting map produced
a single large cluster and a handful of implausibly small and less meaningful clusters. A 20-
voxel cluster-extent threshold was applied to the resulting map to emphasize the largest and
Neurobiology of Language
89
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Predictive coding and error correction
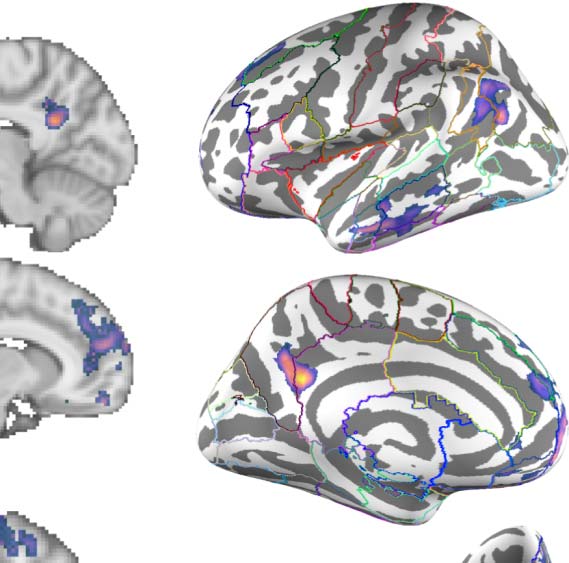
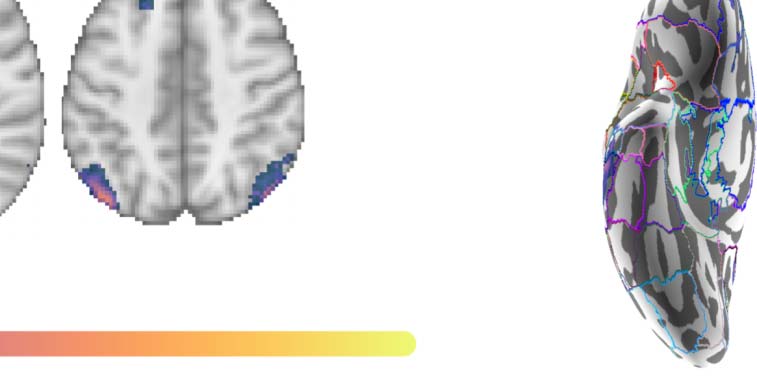
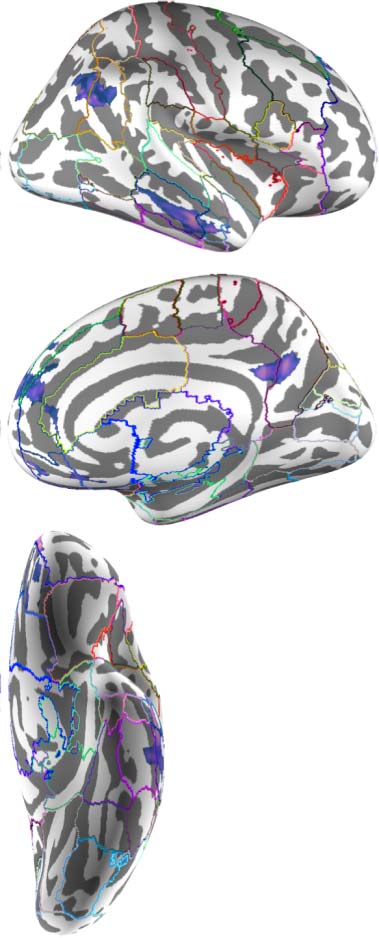
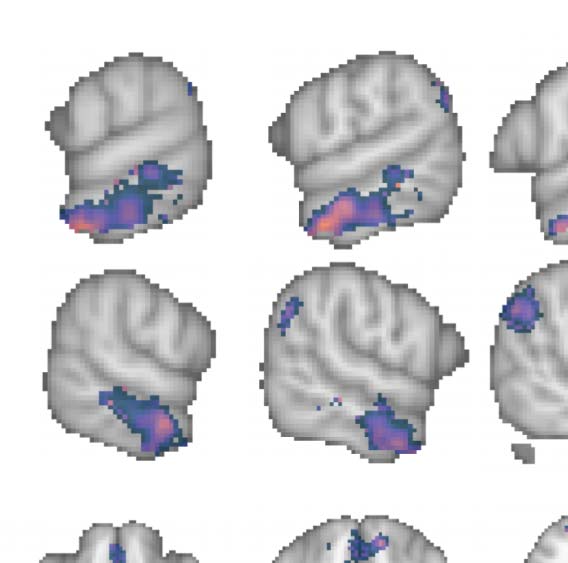
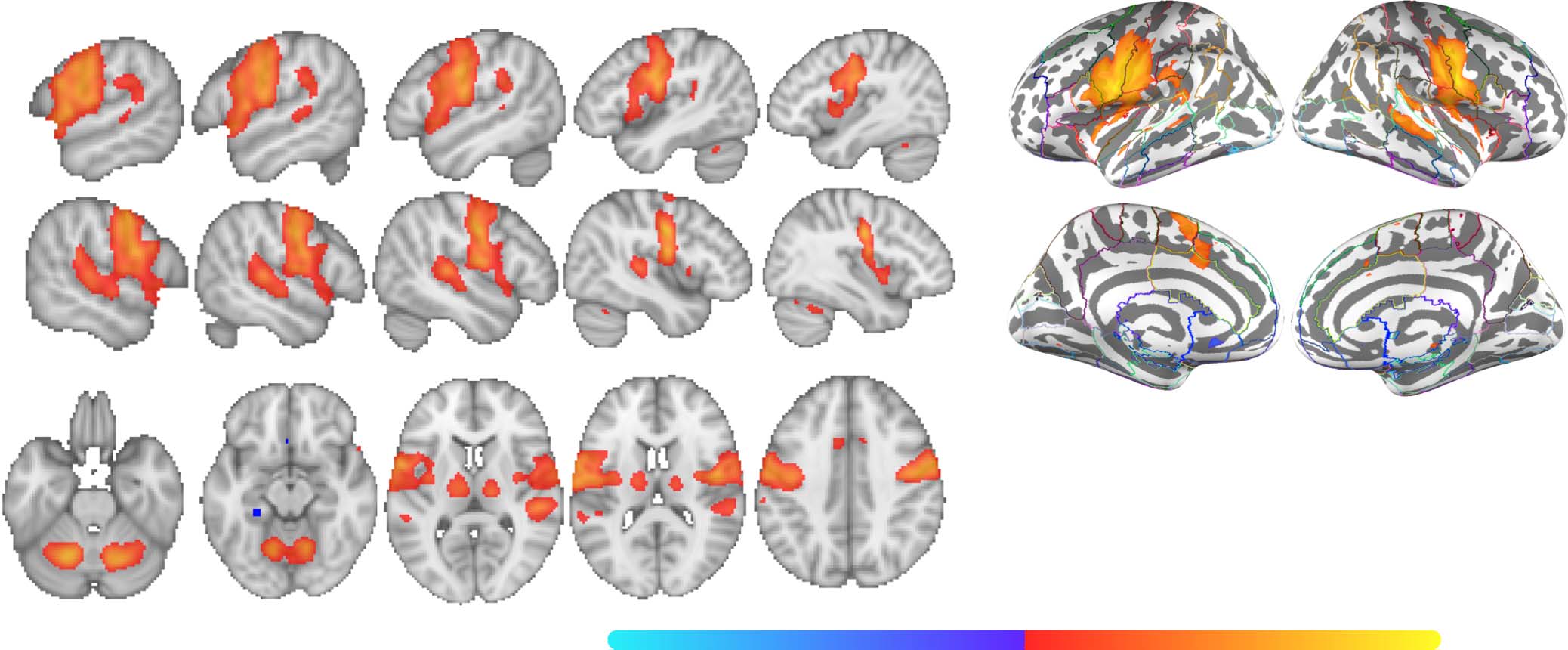
Figure 2. Group contrast between taboo and (neutral) word conditions. (A) select slices from the volume-based group activation map. Sagittal
slices cut through peak activation observed in anterior middle temporal gyrus (x = ±64) and three separate peaks in posterior middle temporal
gyrus (x = ±60, ±56). They also show the emergence of activity in superior lateral occipital cortex (x = ±56) and its splitting into more anterior
and posterior foci, with the anterior activity intruding into angular gyrus (x = ±42), as well as more medial activations (x = ±10; i.e., posterior
cingulate gyrus and frontal medial cortex). Axial slices show the same patterns: peak of activation in anterior middle temporal gyrus (z = −26),
two peaks of activation in posterior middle temporal gyrus (z = −14), a third peak of activation in posterior middle temporal gyrus (z = −4), and
two peaks of activation in superior lateral occipital cortex (z = 22, 44). (B) Volume-based results projected onto the fsaverage surface along
with the Harvard–Oxford cortical atlas to better visualize overlap between activations and anatomical regions. Bilateral anatomical regions are
shown as colored outlines and any region overlapping with activation is highlighted by an opaque superimposed number in one hemisphere
that corresponds to the region’s index within the atlas. The labels for these indices are provided in Figure 4.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
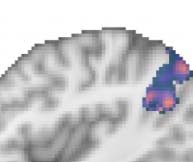
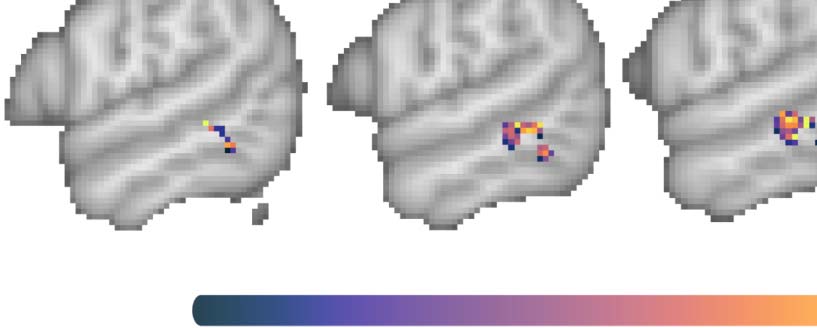
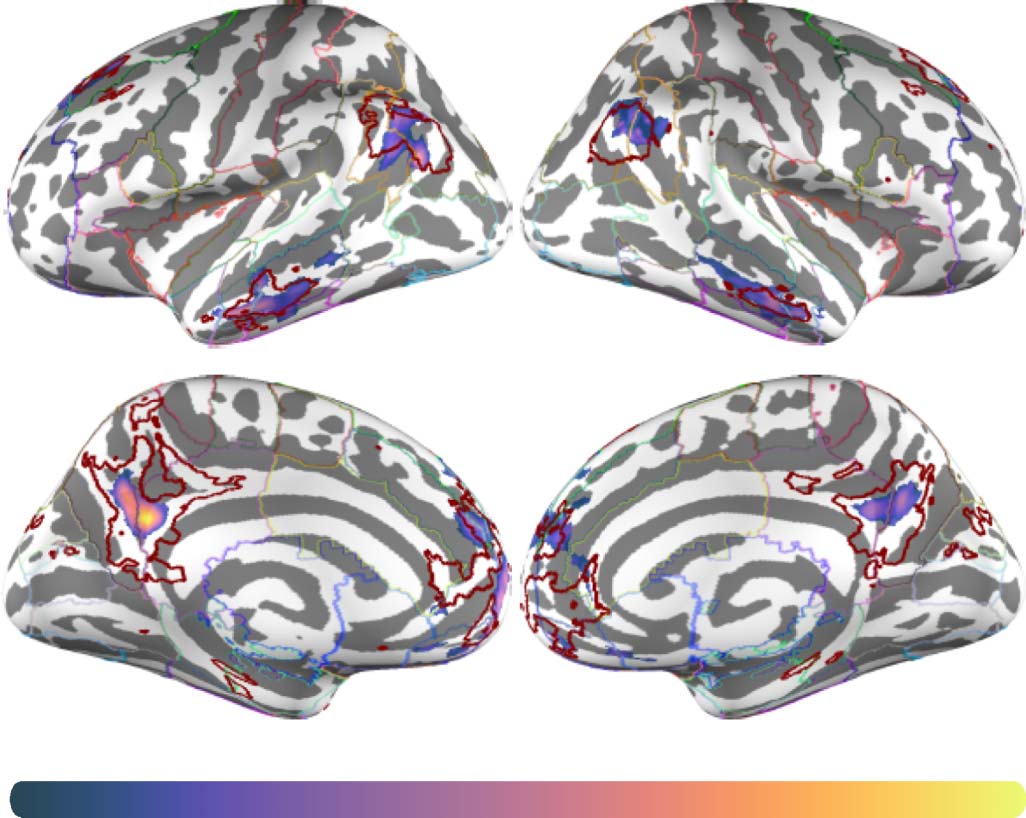
most readily interpretable cluster in our visualization of this result in Figure 3. We emphasize
that the individual maps submitted to the masking procedure were all cluster corrected them-
selves and the exceedingly small clusters in the masked map may reflect uninteresting differ-
ences between contrasts (e.g., noise caused by co-registration). In general, this analysis
revealed that the left pMTG was by far the largest area to show significantly greater response
for taboo than word conditions while also responding significantly above-baseline to the taboo
condition (see Figure 3). The cluster of activity that centered on left pMTG crossed only super-
ficially over the boundary between this area and posterior superior temporal gyrus (STG),
toMTG and temporooccipital inferior temporal gyrus (ITG). We found additional clusters that
peaked in pMTG below the cluster-extent threshold but note that we also found clusters below
this threshold which peaked in right AG and left sLOC (Table 2).
In an additional analysis, we averaged the parameter estimates for the taboo and word con-
ditions (i.e., taboo > base, word > baseline) within the portion of each anatomical area that
exhibited a significant difference between these two conditions (es decir., the contrast map from
Cifra 2). This areal analysis was used to estimate taboo effect size and provided an additional
glimpse into which areas differentially responded to the taboo and word conditions while
simultaneously showing a stronger preference for baseline fixation than the task. En general, este
analysis highlighted the same group of areas as having both greater response to the taboo than
Neurobiology of Language
90
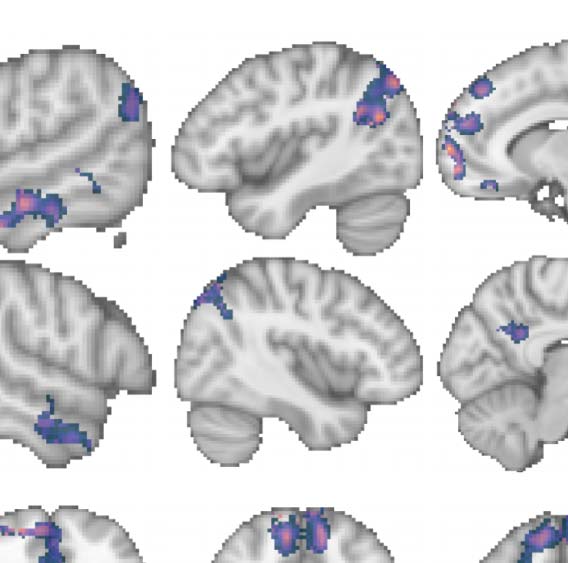
Predictive coding and error correction
Cifra 3. Group contrast between taboo and (neutral) word conditions: masked by significant
activity during the taboo condition (es decir., taboo > baseline) y 20 voxel cluster-extent threshold
is applied, revealing a large area in pMTG.
Mesa 2.
Clusters from the taboo versus word conditions contrast masked by taboo > baseline contrast
Cluster
tamaño
(2 mm
vóxeles)
140
Cohen’s
d
1.78
Peak
z-
valor
3.88
Peak x-
coordinate
−60
Peak y-
coordinate
−38
Peak z-
coordinate
−8
Anatomical atlas areas overlapping with
grupo (% of anatomical area covered)
Left middle temporal gyrus, posterior
división (7%); left superior temporal
gyrus (2%); left middle temporal gyrus,
temporooccipital part (2%); left inferior
temporal gyrus, temporooccipital part
(<1%)
Right angular gyrus (>1%); right lateral
occipital cortex, superior division (>1%)
Left lateral occipital cortex, superior
división (>1%); left angular gyrus (>1%)
19
15
2.13
3.06
52
3.56
3.67
−46
−56
−60
46
46
Left lateral occipital cortex, superior
10
0.81
3.5
−34
−70
56
división (>1%)
Left middle temporal gyrus, posterior
división (>1%); left inferior temporal
gyrus, posterior division (>1%)
4
3.9
2.84
−54
−30
−17
Anatomical
región
overlapping
with peak
Left middle
temporal
gyrus,
posterior
división
Right angular
gyrus
Left lateral
occipital
corteza,
superior
división
Left lateral
occipital
corteza,
superior
división
Left middle
temporal
gyrus,
posterior
división
Neurobiology of Language
91
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4. Group contrast between taboo and (neutral) word conditions: parameter estimates, or beta values for the individual conditions
being contrasted, presented as an average within each anatomical region. Only voxels showing a significant effect in the contrast were
included in the averages. Averages are presented separately for left hemisphere (navy and violet colored bars) and right hemisphere regions
(salmon and yellow colored bars). Error bars represent standard error of the mean. Regions are organized on the x-axis based on descending
mean difference between taboo and word parameter estimates. The magnitude of this difference is shown as a gray line that corresponds to the
secondary y-axis.
word condition and above-baseline response to the taboo condition: bilateral pMTG, left AG,
left pITG, and left aMTG (ver figura 4). Más, we report medium to large effect sizes (es decir.,
taboo vs. palabras) in all areas, with the pITG showing the largest effect size in the left hemi-
sphere, and preCC showing the largest effect size in the right hemisphere (Mesa 3). Areas that
additionally responded above baseline to the taboo condition all showed large effect sizes. No
hemispheric differences were found for effect size (izquierda: m = 1.17, DE = 0.6; bien: m = 0.88,
DE = 0.33; t(9) = 1.04, pag = 0.32) or the spatial extent of activity within areas (izquierda: m = 22%,
DE = 16.18%; bien: m = 19.77%, DE = 16.13%; t(9) = 1.22, pag = 0.25), but activation maxima
inside areas was typically higher in the left hemisphere (izquierda: m = 3.93, DE = 0.37; bien: m =
3.6, DE = 0.26; t(9) = 3.96, pag < 0.01). Finally, we report that only a single area tended to show
above-baseline response to both taboo and word conditions: the left pMTG (Figure 4). This
area overlapped exclusively with the largest cluster identified when masking the taboo versus
words contrast by the taboo > baseline contrast (cf. Figure 2B and Figure 3).
In the preceding whole brain analyses we showed evidence for internal error correction
(es decir., taboo effect) within a portion of the pMTG that our hypotheses targeted. We next carried
out a more specific test for the taboo effect across all voxels of anatomically defined pMTG.
Mean parameter estimates for the taboo and word conditions were extracted from pMTG
voxels for each participant and a paired t test between conditions was performed over
Neurobiology of Language
92
Predictive coding and error correction
Mesa 3.
Areal effect sizes for taboo versus word condition contrast
ROI size Cohen’s d
Atlas ROI
(1) Frontal pole
(3) Superior frontal gyrus
(11) Middle temporal gyrus, anterior division
6997
3347
512
(12) Middle temporal gyrus, posterior division
1400
(15) Inferior temporal gyrus, posterior division
1175
(21) Angular gyrus
(22) Lateral occipital cortex, superior division
(25) Frontal medial cortex
(27) Subcallosal cortex
(28) Paracingulate gyrus
(29) Cingulate gyrus, anterior division
(30) Cingulate gyrus, posterior division
(31) Precuneous cortex
1197
5251
561
782
1720
1385
1332
3047
Atlas ROI
(1) Frontal pole
(3) Superior frontal gyrus
(11) Middle temporal gyrus, anterior division
8195
3005
472
(12) Middle temporal gyrus, posterior division
1374
(15) Inferior temporal gyrus, posterior division
1056
(21) Angular gyrus
(22) Lateral occipital cortex, superior division
(25) Frontal medial cortex
(27) Subcallosal cortex
(28) Paracingulate gyrus
(29) Cingulate gyrus, anterior division
(30) Cingulate gyrus, posterior division
(31) Precuneous cortex
1658
5127
594
749
1650
1526
1393
3141
Left hemisphere
Peak z-value
4.2
Peak x
−12
Peak y
66
3.68
3.99
3.94
3.3
4.25
4.41
3.44
–
3.73
–
4.36
4
−10
64
−60
−60
−50
−36
−6
–
−10
–
−10
−6
54
−6
−26
−32
−56
−68
54
–
54
–
−50
−54
Peak z % ROI active
8
26
−24
−8
−16
42
60
−10
–
24
–
26
32
14
14
45
49
14
28
15
51
5
18
4
18
11
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
0.6
0.61
1.62
0.83
2.38
1.53
0.78
0.6
–
0.71
–
1.6
1.62
ROI size Cohen’s d
Peak z-value
Peak x
Peak y
Peak z % ROI active
Right hemisphere
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
0.53
0.61
0.79
1.49
–
1.01
0.75
0.63
0.7
0.71
0.79
1.12
1.53
3.98
3.6
3.45
3.88
–
3.69
3.91
3.36
3.15
3.59
3.26
3.55
3.74
12
10
58
70
–
48
44
−2
2
10
−2
−2
0
68
54
−2
−28
–
−54
−68
54
26
52
42
−52
−60
22
20
−26
−4
–
34
44
−20
−20
20
−4
32
38
9
9
46
51
2
18
10
41
14
26
8
14
9
Participantes (Figure 5A). The taboo condition elicited higher parameter estimates than the
word condition in the left pMTG (taboo: m = 18.53, DE = 36.77; palabra: m = 9.59, DE =
41.84; t(33) = 3.12, Bonferroni corrected p < 0.05). The same effect was on the cusp of
significance in the right hemisphere (taboo: M = 2.84, SD = 25.66; word: M = −4.23, SD =
Neurobiology of Language
93
Predictive coding and error correction
Figure 5. Taboo effect in posterior middle temporal gyrus (pMTG) within-participants. Anatomical ROI-based analysis of the internal error
correction effect in pMTG. (A) Parameter estimates, or beta values for taboo and word conditions are averaged across all voxels of left pMTG
(left) and right pMTG (right) in each subject. The shaded portions of the line plot for each subject represents standard error of the mean. Results
of a paired t test across participants is presented below, showing significantly higher response in left pMTG to the taboo condition, and a
difference between the two conditions that is on the cusp of significance for right pMTG. (B) T values showing magnitude and significance
( p < 0.001; Bonferroni corrected) of parameter estimate differences between the two conditions are shown for each subject as a bar plot. Note
the asterisk marks insignificant results. Each subject is assigned a color that is consistent between panels. Bars are presented in pairs such that
the first and darker shaded bar of any pair represents the result of the t test performed on the left hemisphere ROI, and the second and lighter
shaded bar represents the result of the t test performed over the right hemisphere ROI. Overall, 26/34 participants show a significant effect in at
least one hemisphere (22 show an effect in both hemispheres).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
32.61; t(33) = 2.58, Bonferroni corrected p = 0.05). In addition, we ensured that the left
hemisphere internal error correction effect was present independently during imagined
(taboo: M = 10.24, SD = 19.49; word: M = 4.75, SD = 24.53; t(33) = 2.05, Bonferroni cor-
rected p < 0.05) and silently articulated tongue twister trials (taboo: M = 2.84, SD = 25.66;
word: M = −4.23, SD = 32.61; t(33) = 2.99, Bonferroni corrected p < 0.05).
Finally, we characterized the consistency of the taboo effect within anatomical pMTG by
applying paired t tests between the taboo and word conditions within participants (Figure 5B).
This analysis yielded a significant taboo effect in roughly 76% of participants (N = 26/34;
Bonferroni-corrected p < 0.001). The effect occurred most often bilaterally (N = 22/26) and
showed no hemispheric preference when it occasionally occurred in a single hemisphere
(left: N = 2/26; right: N = 2/26). In four participants the effect was present, but response
in pMTG was higher during baseline fixation than during the taboo condition. That is,
response in pMTG was greater for the taboo than word conditions but highest for baseline
fixation. In all cases where the effect was not present, we saw a significant reverse effect (N = 9).
We considered whether portions of pMTG still showed a taboo effect in those participants who did
not exhibit a mean effect across the entire pMTG by inspecting significant voxel-level differences
between taboo and word conditions. No participant who showed a nonsignificant mean taboo
effect presented with significant differences between taboo and word condition within portions
of pMTG.
Neurobiology of Language
94
Predictive coding and error correction
Forward prediction effect
One aim of the current study was to replicate the forward predictive signal effect described in
prior work (Okada et al., 2018) by identifying brain regions that respond more strongly during
silently articulated than imagined tongue twisters. To that end, we confirmed that silently artic-
ulated speech activates portions of auditory cortex more strongly than imagined speech, even
though both conditions lack auditory input and do not involve overt production ( p < 0.01 clus-
ter corrected at p < 0.05; see Figure 6A–B). Overall, silently articulated tongue twisters pro-
duced greater activity than imagined tongue twisters in and around Heschl’s gyrus (HG), but
also in a broad network of speech-related regions that span STG, planum temporale (PT),
pMTG, toMTG, precentral gyrus, postcentral gyrus (postCG), insula, IFG, aCG, and the cere-
bellum (Figure 6A–B). We also found that silently articulated tongue twisters yielded greater
activity in other brain regions, most of which have also been observed to activate during speech
processing (Figure 6A–B). These regions were found in temporal cortex (i.e., bilateral planum
polare, bilateral temporal pole), inferior temporal and neighboring portions of occipital cortex
(i.e., left temporooccipital ITG, bilateral lingual gyrus, bilateral temporal occipital fusiform cor-
tex, bilateral occipital fusiform gyrus), parietal cortex (i.e., bilateral parietal operculum, bilateral
anterior and posterior supramarginal gyrus, bilateral superior parietal lobule), and frontal cortex
(i.e., bilateral supplementary motor cortex, bilateral middle frontal gyrus, bilateral frontal and
central operculum, bilateral SFG, left frontal orbital cortex; Figure 6A–B).
One point of difference between our results here and prior work (Okada et al., 2018) is that
we also report regions that express greater activity for imagined than silently articulated word
lists in several areas including bilateral anterior parahippocampal gyrus (paraHG) and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
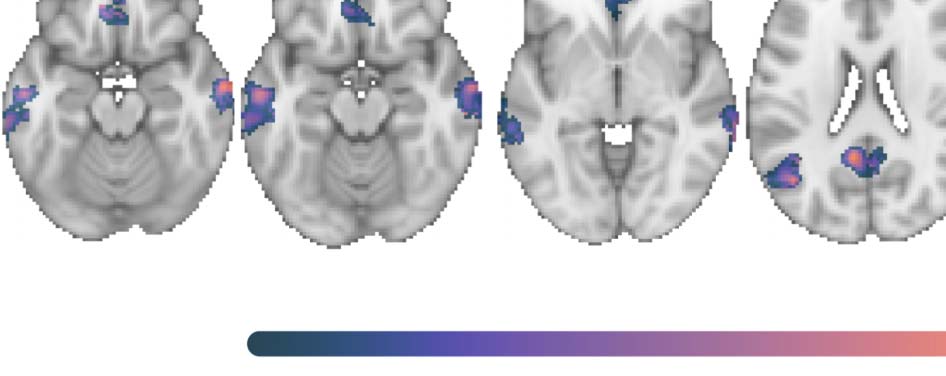
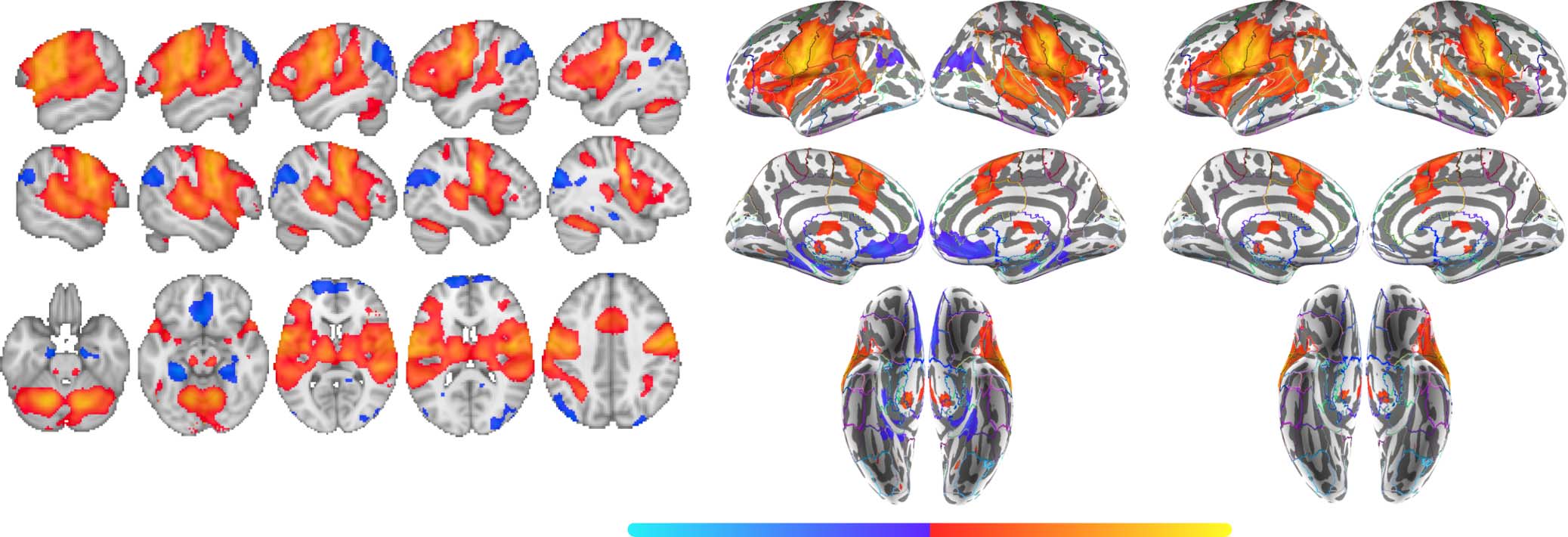
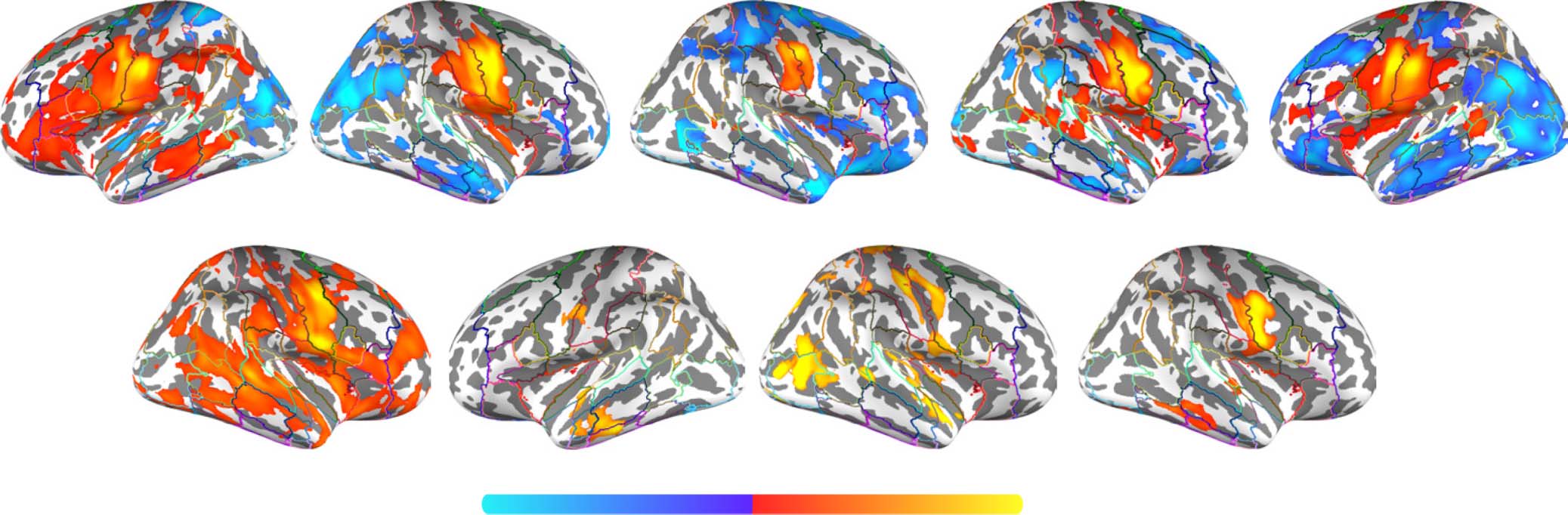
Figure 6. Group contrast between silently articulated and imagined speech. (A) Select slices from the volume-based group activation map.
Sagittal slices cut through peak activation for the contrast in precentral and postcentral gyri (x = ±58) and follow Heschl’s gyrus (x = ±50, ±44,
and ±38). Axial slices show the most ventral activations in the contrast, including the cerebellum and neighboring mesial structures (e.g.,
anterior paraHG and brainstem; z = −26), more dorsal activations in inferior temporal cortex (z = −14), activations along middle and superior
temporal cortex (including Heschl’s gyrus; z = 8 and 14), and the most posterior activation in the contrast within posterior supramarginal gyrus
(z = 40). (B) Volume-based results projected onto the fsaverage surface along with the Harvard–Oxford cortical atlas to better visualize overlap
between activations and anatomical regions. Bilateral anatomical regions are shown as colored outlines and any region overlapping with
activation is highlighted by an opaque superimposed number in one hemisphere that corresponds to the region’s index within the atlas.
The labels for these indices are provided in Figure 7. (C) Masked by significant activity during either condition (i.e., silent articulation > base-
line OR imagined > baseline), revealing that areas which showed relatively stronger response to imagined speech were deactivated during the
imagined speech task.
Neurobiology of Language
95
Predictive coding and error correction
posterior paraHG, bilateral posterior temporal fusiform cortex, bilateral AG, bilateral sLOC,
bilateral FP, bilateral FMC, bilateral subcallosal cortex, bilateral paraCG, right preCC, y correcto
occipital pole (OccP; Figure 6A–B).
Just as for the taboo effect, the contrast between silently articulated and imagined tongue
twisters was masked by areas that showed significant above-baseline response to each of these
two conditions, revealing below-baseline response almost exclusively in areas that produced
significantly higher response to imagined than silently articulated tongue twisters (Figure 6C).
En efecto, all areas that showed greater response to imagined tongue twisters also showed
below-baseline response during the imagined condition, while only a few small areas in infe-
rior temporal and occipital cortex that showed greater response to silently articulated tongue
twisters exhibited below-baseline response during the silent articulation condition (cf.
Figure 6B with 6C). The latter areas included bilateral lingual gyrus, bilateral occipital fusiform
gyrus, and right OccP (cf. Figure 6B with 6C).
Areal parameter estimates for silently articulated and imagined tongue twister conditions
confirmed that areas with stronger response to the silently articulated condition all showed
above-baseline response (Cifra 7). Sin embargo, this analysis also revealed areas with stronger
response to the imagined condition that showed above-baseline response, mainly left AG, izquierda
FP, and left anterior paraHG (Cifra 7). Notablemente, parameter estimates for these three areas were
relatively low for the imagined condition. The contrast between silently articulated and imag-
ined conditions that was masked by above-baseline imagined response did not pick out voxels
in these areas due to the more stringent statistical significance criteria in that analysis.
Areal effect sizes for the contrast between silently articulated and imagined conditions were
large for left anterior paraHG, but small for left AG, and negligible but significant for the left FP
(Mesa 4). For other areas which showed greater response during the imagined than silently
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 7. Group contrast between silently articulated and imagined speech: parameter estimates, or beta values for the individual conditions
being contrasted, presented as an average within each anatomical region. Only voxels showing a significant effect in the contrast were
included in the averages. Averages are presented separately for left hemisphere (blue and cyan colored bars) and right hemisphere regions
(red and yellow bars). Error bars represent standard error of the mean. Regions are organized on the x-axis based on descending mean
difference between articulated and imagined betas. The magnitude of this difference is shown as a gray line that corresponds to the sec-
ondary y-axis.
Neurobiology of Language
96
ROI
tamaño
6997
1463
3347
Cohen’s
d
−0.1
0.95
0.36
Peak
z-value
4.82
5.28
5.3
3280
0.52
4.97
836
1.12
5.33
Peak
x-coord
−44
−36
−10
−48
−48
915
0.91
6.02
−62
4981
2465
340
1.22
0.72
1.37
6.59
5.56
5.17
−62
−56
−66
−66
48
−8
0
30
28
12
0
14
0
−8
(10) Superior
1019
1.93
4.72
Mesa 4.
Areal effect sizes for articulated versus imagined word lists
Left hemisphere
Right hemisphere
Peak
y-coord
Peak
z-coord
Peak
z-value
Peak
x-coord
Peak
y-coord
Peak
z-coord
% ROI
active
% ROI
active
19
68
23
27
62
ROI
tamaño
8195
1465
3005
Cohen’s
d
−0.46
1.09
0.79
4.36
5.37
4.79
3141
0.65
5.65
745
0.56
4.4
20
18
76
34
12
40
34
0
44
46
36
−6
28
10
34
10
18
52
48
8
7
58
13
10
28
4
96
847
0.68
5.19
56
14
24
94
24
−2
6
50
13
47
4822
2490
348
1.45
0.83
1.47
7.01
4.68
5.02
40
58
64
4
94
1155
2.2
5.84
48
−6
12
0
−28
36
2
8
8
1400
1.39
4.06
−54
−40
−2
16
1374
2.02
4.67
50
−20
−4
1019
1.71
3.96
−54
−42
8
29
1287
1.93
5.28
60
−48
12
15
808
1.35
4.83
−52
−58
−12
19
907
–
–
–
–
–
(17) Postcentral gyrus
4046
1.55
(18) Superior
parietal lobule
1731
0.9
6.38
4.72
−60
−44
−4
−38
26
46
36
15
3627
1669
2.22
0.93
6.46
3.58
52
30
−6
−50
40
46
Atlas ROI
(1) Frontal pole
(2) Insular cortex
(3) Superior
frontal gyrus
(4) Middle
frontal gyrus
(5) Inferior
frontal gyrus,
pars triangularis
(6) Inferior
frontal gyrus,
pars opercularis
(7) Precentral gyrus
(8) Temporal Pole
(9) Superior
temporal gyrus,
anterior division
temporal gyrus,
posterior division
(12) Middle
temporal gyrus,
posterior division
(13) Middle
temporal gyrus,
temporooccipital
part
(16) Inferior
temporal gyrus,
temporooccipital
part
norte
mi
tu
r
oh
b
oh
oh
gramo
y
i
yo
oh
F
l
a
norte
gramo
tu
a
gramo
mi
9
7
PAG
r
mi
d
i
C
t
i
v
mi
C
oh
d
i
norte
gramo
a
norte
d
mi
r
r
oh
r
C
oh
r
r
mi
C
t
i
oh
norte
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
48
14
47
92
25
<1
35
7
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
Atlas ROI
(19) Supramarginal
gyrus, anterior
division
(20) posterior
division
(21) Angular gyrus
(22) Lateral occipital
cortex, superior
division
Table 4.
(continued )
Left hemisphere
Right hemisphere
ROI
size
1128
Cohen’s
d
Peak
z-value
0.77
4.93
Peak
x-coord
−66
Peak
y-coord
−30
Peak
z-coord
% ROI
active
ROI
size
Cohen’s
d
Peak
z-value
Peak
x-coord
26
82
959
1.64
4.85
48
Peak
y-coord
−32
Peak
z-coord
% ROI
active
54
45
1357
0.8
4.87
−52
−40
24
46
1516
1.04
4.91
58
−42
16
34
1197
5251
−0.35
−0.24
3.83
4.54
(25) Frontal medial
561
−0.93
−3.72
874
0.79
5.28
cortex
(26) Juxtapositional
lobule cortex
(formerly s
upplementary
motor cortex)
(27) Subcallosal
cortex
gyrus
(29) Cingulate gyrus,
anterior division
(31) Precuneous
cortex
782
−2.52
−4.53
−4
(28) Paracingulate
1720
0.01
4.9
1385
0.34
4.95
3047
–
–
–
−38
−28
−2
−8
−12
−12
−50
−64
34
2
22
14
12
–
20
44
54
−14
64
−6
38
38
–
−6
(33) orbital
1943
0.76
4.42
−50
cortex
(34) Parahippocampal
709
−1.41
−4.19
−16
−10
−22
gyrus, anterior
division
(35) Parahippocampal
gyrus, posterior
division
(36) Lingual gyrus
(38) Temporal
fusiform cortex,
posterior division
9
8
475
−0.8
−4.51
−28
−32
−12
1934
1001
0.67
5.93
−0.69
−3.92
−18
−28
−62
−34
−12
−16
12
12
46
20
14
17
20
25
68
48
41
42
1658
5127
−0.29
−0.48
4.33
−4.24
594
−1.18
−3.93
834
0.9
4.76
749
−2.56
−4.45
1650
−0.01
4.41
1526
0.3
4.37
0
3141
1681
–
–
–
–
42
28
2
2
−2
10
2
–
–
−46
−86
34
6
22
16
18
–
–
52
34
−12
74
−4
40
38
–
–
766
−1.59
−4.25
16
−10
−22
378
−1.07
−3.72
26
−32
−10
2037
0.43
5.35
812
−0.99
−3.53
14
34
−66
−20
−12
−18
20
27
42
57
44
38
40
<1
4
11
39
26
8
P
r
e
d
i
c
t
i
v
e
c
o
d
i
n
g
a
n
d
e
r
r
o
r
c
o
r
r
e
c
t
i
o
n
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
> 4.2; Figure 8A–B). críticamente, the resulting independent cluster within temporal
cortex was just posteroventral to HG, remaining in the vicinity of auditory cortex. At this
límite, the frontal cluster peaking in left precentral gyrus extended into left supramarginal
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 8. Group contrast between silently articulated and imagined speech. Minimal significance threshold from Figure 13 was increased by
increments of 0.1 until the frontal-temporal cluster splintered into multiple independent clusters. (A) The results of the same contrast thresh-
olded at Z > 4.2 (cluster corr., pag < 0.05) are shown in volume-space. (B) The same maps from Figure 6A projected onto the fsaverage surface.
Neurobiology of Language
100
Predictive coding and error correction
Forward prediction effects in Heschl’s gyrus within-participants. Atlas region of interest (ROI) based analysis of the forward pre-
Figure 9.
diction effect in Heschl’s gyrus is shown. (A) Beta values for silently articulated word lists and imagined word lists are averaged across all
voxels of left and right Heschl’s gyri in each participant. The shaded portions of the line plot for each participant represents standard error of
the mean. The bottom portion of the panel also reports the result of a t test for a group-level difference between conditions (i.e., paired t test
across participants), which shows significantly greater response during silently articulated speech in Heschl’s gyri. (B) T values showing mag-
nitude and significance ( p < 0.001; Bonferroni corrected) of beta differences between the two conditions are shown for each participant as a
bar plot. Note the asterisk marks insignificant results. Each participant is assigned a color that is consistent between panels (as well as with
Figure 5). Bars representing the results of t tests performed on both hemisphere ROIs are presented in pairs: left (darker shaded bar) and right
(lighter shaded bar). Overall, 26/34 participants show a significant effect in at least one hemisphere (20 show an effect in both hemispheres).
gyrus and posterior PT. The same cluster also covered left insular cortex, operculum, IFG (pars
opercularis), and postCG. Activity from left frontal cortex spilled over into anterior temporal
regions as well as anterior PT. Independent clusters were also found in left supplementary
motor area (SMA), extending into preCC, and in left cerebellum. Anatomical overlap was
remarkably similar for right hemisphere activity, but the frontal-temporal cluster remained
intact, the right SMA was absent, and temporal activity was approximately closer to HG.
A test for the forward prediction effect was also carried out across all voxels of HG. Mean
parameter estimates for the silently articulated and imagined conditions were extracted from
HG voxels for each participant and a paired t test between conditions was performed over
participants (Figure 9A). A forward prediction effect for mean response in HG was present
at the group level, with silently articulated tongue twisters producing higher mean parameter
estimates across participants than imagined tongue twisters in both the left hemisphere (artic-
ulated: M = 67.4, SD = 79.84; imagined: M = 36.39, SD = 58.1; t(33) = 3.62, Bonferroni-
corrected p < 0.01) and the right hemisphere (articulated: M = 36.6, SD = 74.46; imagined:
M = 4.15, SD = 59; t(33) = 4.52, Bonferroni-corrected p < 0.01).
The forward prediction effect based on mean response in HG was present in roughly 76%
(N = 26/34) of participants (Bonferroni-corrected p < 0.001; Figure 9B). Most of these par-
ticipants (N = 20/26) presented with a bilateral effect, but in one participant the effect was
present only in the left hemisphere, and in five participants it was present only in the right
hemisphere. Four participants that showed the effect also showed a stronger response to base-
line than silent articulation. Most participants that showed no forward prediction effect in
Neurobiology of Language
101
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Predictive coding and error correction
Figure 10. Contrast maps for example participants who show no mean forward prediction effect in Heschl’s gyrus. Maximum and minimum
z-scores vary among participants. Participants 2, 13, 14, 15, 22, and 32 show higher response for imagined than silently articulated speech
across Heschl’s gyrus bilaterally. Participant 11 shows higher response to imagined speech in left Heschl’s gyrus and no significant difference
in the right hemisphere. Note, there is still higher response to silently articulated speech within Heschl’s gyrus for all participants, except
Participant 13, who still shows higher response to silently articulated speech in the posterior superior temporal sulcus. Participants 16 and
23 all show higher response during silently articulated speech than imagined speech in Heschl’s gyrus bilaterally, but also present with
response during silently articulated speech that is below baseline fixation. These participants’ contrast maps are masked by activity that is
above baseline during silent articulation. Each participant shows higher activity for silently articulated than imagined speech in areas bordering
Heschl’s gyrus.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
either hemisphere presented with a significant effect in the reverse direction (i.e., imagined >
silently articulated across HG; norte = 7/8), and a few participants with an established right hemi-
sphere forward prediction effect presented with a reverse effect in the contralateral hemisphere
(norte = 4). Inspecting significant voxel-level differences between speech task conditions within
participants revealed that the majority of participants that failed to show a mean effect in HG
bilaterally still had voxels within that area that showed greater response to silently articulated
than imagined speech (pag < 0.05, cluster corrected p < 0.05; N = 6/8). Notably, however, this
activity often peaked or at least covered areas bordering HG, including the PT, the posterior
STG, the anterior STG, and the planum polare (N = 4/6; see participants 2, 14, 22, and 32 in
Figure 10). In at least one additional participant we found evidence of stronger response to
silently articulated speech, but in the posterior superior temporal sulcus (N = 1/8; see partic-
ipant 13). By masking participant-level contrast maps (i.e., silently articulated vs. imagined) by
condition-level activation maps (i.e., silently articulated vs. baseline), we also found that half
of the participants that presented with a mean effect in HG but showed below-baseline
response during articulated speech (bilaterally) showed some voxels in HG that responded
more strongly to articulated speech and produced above-baseline response (N = 2/4; see par-
ticipants 16 and 23 in Figure 10).
In general, we found at least one of the two effects that we sought (i.e., internal error
correction and forward prediction) in 94% of participants (N = 32/34) and both effects in
59% of participants (N = 20). No significant relationship was documented between mean
framewise displacement and participant-level t scores for contrasts between conditions
(silently articulated vs. imagined speech in left HG: r(33) = 0.21, p = 0.21; silently articulated
versus imagined speech in right HG: r(33) = 0.02, p = 0.93; taboo errors versus word errors in
left pMTG: r(33) = −0.14, p = 0.43; taboo errors versus word errors in right pMTG: r(33) =
−0.11, p = 0.53).
Neurobiology of Language
102
Predictive coding and error correction
Exploratory Findings
Comparing the default mode and taboo networks
The wider network that responded more strongly to taboo than word conditions while some-
times showing below-baseline response (referred to as TN throughout) bears a strong resem-
blance to the default mode network (DMN). Despite the stronger response to baseline fixation
in some portions of this network, the fact that the class of the error word—the main difference
between conditions—appears to be driving changes in activity entirely across the DMN is
intriguing. Areas of the DMN commonly activate during mentalization and theory of mind
tasks (Mars et al., 2012), suggesting that taboo words may be tapping into social cognitive
systems. Here, we sought to describe the functional relationship between areas of the TN
and the DMN.
First, we demonstrated that areas of the TN are in fact recruited by the DMN. To define the
boundaries of the DMN, we performed a meta-analysis for the phrase “default mode” in Neu-
rosynth (FDR-corrected p < 0.01). The resulting meta-analytic map was found to cover canon-
ical areas of the DMN (Mars et al., 2012): the anterior medial prefrontal cortex, pCG, AG,
MTG, and paraHG. We refer to this map as the DMN throughout the results. The boundaries
of the DMN were superimposed over the TN in Figure 11A, qualitatively showing that roughly
the same regions activated across the two networks, but that activity was more focal in the TN
and often formed a subset of each of the regions that was active in the DMN (Figure 11A). The
major exception to this pattern was the paraHG, which appeared in the DMN but not in the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
n
o
_
a
_
0
0
0
8
8
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 11. Relationship between taboo and default mode networks. Decoding the relationship between taboo and default mode networks.
(A) Spatial overlap between cortical areas significantly associated with the taboo network (TN) from Figure 2 and Figure 3 and cortical areas
significantly associated with the default mode network (DMN). The TN is represented by purple to yellow colors on the cortical surface and the
DMN is represented by a maroon outline. The DMN is defined using the Neurosynth meta-analytic database, by analyzing activity associated
with studies frequently using the term “default mode.” The resulting map represents areas significantly likely to activate across the set of studies
frequently using this term (FDR corrected p < 0.01). (B) Percentage of TN voxels that overlap with the DMN, plotted as a function of network
threshold. Overlap in the left hemisphere is plotted as a peach line and overlap in the right hemisphere is plotted as a teal line.
Neurobiology of Language
103
Predictive coding and error correction
TN. Although we saw particularly strong overlap between the two networks in the MTG, the
TN contained large portions of the pMTG that did not appear in the DMN. We calculated that
45% of the voxels in the left TN and 36% of the voxels in the right TN overlapped with the
DMN (Figure 11B). The finding of greater overlap in the left TN than the right was invariant to
network significance thresholds (Figure 11B).
To better understand the relationship between the TN and the DMN, we used Neurosynth
to probe for the main functional associations of the TN and then compared the extent to
which the same functions were associated with the DMN. That is, we executed the same
meta-analysis procedure that we used to define the DMN over the entire set of 3,228 terms
frequently used in the neuroimaging literature that are embedded in Neurosynth. We then
estimated the Pearson correlation coefficient between each of these meta-analytic networks
and the TN. Networks were not thresholded prior to correlation because our goal was to deter-
mine purely whether the likelihood of seeing activity in a group of neuroimaging studies
related to the likelihood of observing activity in the TN. However, the resulting correlation
coefficients were corrected for multiple comparisons, and we focused only on the top 100
out of 2,952 significant correlations (FDR-corrected p < 0.001). Finally, each of the 100 net-
works most strongly correlated to the TN were in turn correlated to the DMN (Figure 12A). The
DMN (i.e., the term “default mode”) marked the strongest relationship to the TN, explaining
roughly 20% of activity in the TN and validating our early assessment of the areas that form the
TN. The set of unique terms that explained the greatest amount of variance in TN activity (r >
0.35, r2 > 0.12) additionally included “theory of mind,” “mentalizing,” “social” [Procesando],
“social cognition,” “autobiographical memory,” “self-referential,” and “beliefs.” In this set,
associations were higher to the DMN only for the following terms: “default mode,” “self-
referential.” and “autobiographical memory.” For the other terms in the set, association to
the TN was on average modestly higher (mean difference in r = 0.07, standard error of the
mean = 0.01), except for the terms “social” [Procesando] (DMN r = 0.27, TN r = 0.42) y
“social cognition” (DMN r = 0.23, TN r = 0.36), which showed a difference that was about
twice that of the mean.
A host of other terms were overall significantly associated with the TN as well. Broadly,
these covered other memory systems (p.ej., “episodic,” “semantic memory”), generic memory
procesos (p.ej., “retrieval,” “recall,” “remembering,” “recollection”), emotional processes and
estímulos (p.ej., “emotions,” “affective,” “valence,” “emotional regulation”), references to resting
estado (p.ej., “intrinsic,” “resting”), other social cognitive processes (p.ej., “social interaction,"
“inference,” “intentions,” “morals,” “personality,” “future”), Toma de decisiones (p.ej., “judg-
mentos,” “value”), idioma (p.ej., “semantic,” “sentence, “comprehension”; note semantic
memory does not show a strong difference in association strength to the TN and the DMN),
and disorders (p.ej., “depression,” “spectrum”).
Across all terms that more strongly associated with the TN than the DMN, the language-
related category of terms (p.ej., semantics and sentence comprehension) showed the largest
difference in association strength (es decir., between TN and DMN), followed by emotion pro-
cesses and social cognition (see the bilateral plots in Figure 12B, which show the terms
from Figure 12A organized by association difference). Of the terms with stronger associa-
tion to the DMN, resting state terms and self-reference showed some of the largest differ-
ences with the TN. When we decomposed differences in association to the DMN and TN
by hemisphere, we found that the high association difference for language-related terms
was being entirely driven by activity in the left hemisphere, whereas the association difference
for resting state terms and self-reference was moderately higher in the right hemisphere
(Figure 12B).
Neurobiology of Language
104
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 12. Groups of studies associated with the TN are found by correlating TN z-scores across the whole brain with the likelihood of seeing
activity for studies frequently using each of the other terms contained in the Neurosynth database. (A) The top 100 significant correlations (FDR
corrected p < 0.001; r > 0.18) are presented as purple dots and organized by descending association strength. For each of these terms, el
correlation between the likelihood of seeing activity for that term and the likelihood of seeing activity for the “default mode” term (es decir., DMN)
is also shown by red dots. A line is drawn between the purple and red dots to emphasize the magnitude of difference between a term and its
association to the TN and DMN. The color of the lines shows which of the two networks showed the strongest association to the term. (B)
Terms associated with the TN are sorted by descending difference in their association strength to the TN minus their association strength to the
DMN. Associations between each term and the two networks and the differences between them are computed separately for the left hemi-
sphere (pale purple dots), hemisferio derecho (deep violet dots), and bilaterally (es decir., matching the data shown in panel (A); medium violet dots).
Hemisphere-dependent variability is highlighted by connecting each set of dots by a line. The color of the line shows whether the term was
overall more strongly associated with the TN (violet line) or DMN (maroon line).
Neurobiology of Language
105
Predictive coding and error correction
Decoding regions of the taboo network
The finding that activity in the left TN more closely resembled language-related terms than
activity in the left DMN was remarkable because spatial overlap between the TN and DMN
was higher in the left hemisphere. This suggested that what small differences existed in the
distribution of activity between the left TN and DMN were functionally relevant for language.
We tested whether the largest of such differences, the segment of the left pMTG that does not
show overlap with the DMN was driving the taboo network-level association with language.
Para hacer esto, we separated the left MTG region in the TN into two segments based on its overlap
with the DMN (see top panel of Figure 13B) and then decoded the functions of both segments
using Neurosynth. We note that the portion of the pMTG in the TN that did not overlap with
the DMN was nearly identical to the largest region from the contrast between taboo and word
conditions that also showed above-baseline response (cf. Figure 9B with Figure 3). In the inter-
est of better characterizing the functional contributions of each region in the TN, we decoded
the other regions of the TN as well, including the unsegmented left MTG (see top panel of
Figure 13A). We decoded regions by generating a map for each term in Neurosynth that cap-
tures the posterior probability that a study uses a term if activity was observed in a particular
voxel, assuming a uniform prior (es decir., all terms are equally likely to appear). Mean probabilities
were calculated across voxels of each region in the TN, and we presented all terms that
showed at least a 65% mean likelihood of being used when activity was observed in one
of the regions in the TN.
Decoding each region of the TN (Figure 13A) broadly showed that different functional cat-
egories were relatively more likely to be referenced in the context of different portions of the
TN. The majority of language-related terms, including sentence comprehension and lexical-
semantics were more likely to be used when there was activity in the left MTG compared to
other regions of the TN (p.ej., see “language network,” “reading,” “syntactic,” “phonological,"
“lexical,” “word,” “words,” “sentence,” “sentences,” “concepts,” “semantic,” “meaning,"
“aphasia” in Figure 13A). Auditory comprehension terms were most likely to be used when
the right MTG was active (p.ej., see “voice,” “speaker,” “speech,” “acoustic,” “comprehen-
sión,” “listening,” “linguistic” in Figure 9A). The DMN and social cognition, including theory
of mind and self-reference were most likely to be referenced when activity was in bilateral
pCG (p.ej., see “thoughts,” “moral,” “personal,” “theory of mind,” “episodic memory,” “auto-
biographical memory,” “self-referential,” “social interactions,” “default mode” in Figure 9A).
Beliefs were most likely to be referenced when either of parietal regions in the TN were active,
but the right parietal region was more strongly associated with multiple beliefs. The term
“mentalization,” but curiously also “read,” “readers,” and “written” were most likely to be
used when the right parietal region was active as well. “Word form,” “retrieval,” “recollec-
ción,” “reappraisal,” “semantics,” “judgment tasks” and “native English” are other terms that
were more likely to be used when activity was observed in the left parietal region. No terms
were more likely to be used when activity was found in the bilateral frontal cortex region
instead of anywhere else in the TN.
Despite the relative differences in term use probabilities between regions of the TN in dif-
ferent functional domains, it was overwhelmingly social cognitive and memory terms that
were most likely to be used when activity was found in any region of the TN. Por ejemplo,
the three terms that were most likely to appear when activity was found in each of the regions
of the TN (Figure 9A) were largely the same: “autobiographical” (left MTG, right MTG, izquierda
parietal, bilateral pCG, bilateral frontal cortex), “autobiographical memory” (right MTG, bilat-
eral pCG), “theory of mind” (right MTG, left parietal, bilateral frontal cortex), “mind” (izquierda
MTG, bilateral frontal cortex), “mental states” (left MTG), “mentalizing” (right parietal),
Neurobiology of Language
106
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 13. Decoding clusters of the TN. (A) The association between clusters in the TN from Figure 2A and groups of different studies in
Neurosynth is found by computing the probability of studies in the database using a specific term when activity is reported in each of the
voxels belonging to a cluster, then taking the mean of these probabilities across voxels of the cluster. (Top) Clusters in the TN by color; grupos
are defined in volume space but projected onto the cortical surface to facilitate visualization. (Bottom) Mean probabilities for each cluster in a
set of terms that has a greater than 65% mean probability of activating voxels in at least one of the six TN clusters from this panel, or two
additional TN clusters from panel (B). The terms are presented in groups (by background color) based on which cluster was associated with the
highest mean probability of using a term. Terms are further organized within-group by the difference in probability for the most likely cluster
associated with a term, and the mean probability of the remaining clusters. Bars around each point represent standard error of the mean across
vóxeles. Clusters with ambiguous mean probabilities are not shown (<50%). (B) Two segments of the left MTG cluster from panel (A). (Top) Left
MTG cluster from panel A segmented into a posterior area in left anatomical MTG that has no spatial overlap with the DMN, and an anterior
area that has some spatial overlap with the DMN. (Bottom) Mean probabilities for the same set of terms in panel (A).
Neurobiology of Language
107
Predictive coding and error correction
“beliefs” (right parietal), and “default network” (right parietal, bilateral pCG), and “default
mode” (left parietal). However, in segmenting the MTG region we found that language-related
terms were more likely to be used than social cognitive terms when the posterior segment of
the left MTG region was active (Figure 13B; e.g., “sentences,” “sentence,” “language net-
work,” “language comprehension,” “comprehension,” “linguistic,” “speaker,” “voice”). In
the context of activating the posterior segment of the left MTG, the highest likelihood of using
a term from the social cognitive category was 67.6% for the term “autobiographical.” This
likelihood was 2.4% lower than the likelihood of using the term “semantic” (70%), 3.7% lower
than the likelihood of using the term “lexical” (71.3%), 7.5% lower than using the term “com-
prehension” (75.1%), and 10.1% lower than using the term “sentences” (77.7%). These par-
ticular language-related terms were highlighted here on the basis that they aligned with the
same terms associated with activity across the entire TN (i.e., sentence comprehension and
semantics), and that they additionally implicated the posterior segment of the left MTG region
in lexical processes.
Language-related terms were also more likely to be used when activity was observed in the
posterior segment of the left MTG than the anterior segment of the left MTG. When the pos-
terior segment of the left MTG was active, the likelihood of using the term “semantic” was
1.9% higher (posterior left MTG: 70%; anterior left MTG: 68.1%), the likelihood of using
the term “lexical” was 8.5% higher (posterior left MTG: 71.3%; anterior left MTG: 62.8%),
the likelihood of using the term “comprehension” was 13.2% higher (posterior left MTG:
75.1%; anterior left MTG: 61.9%), and the likelihood of using the term “sentences” was
19% higher (posterior left MTG: 77.7%; anterior left MTG: 58.7%). In general, the likelihood
that language-related terms were used when the posterior segment of the MTG was active was
strikingly high. The top percentile (>75.4%) of all probabilities that a term was used when
activity was found in any region of the TN was comprised of four terms for the pCG (es decir., “auto-
biographical,” “autobiographical memory,” “default network”) and five terms for the posterior
segment of the MTG (“linguistic,” “speaker,” “sentence,” “sentences,” “language network”).
Mientras tanto, social cognitive and memory terms were still most likely to be used when the
anterior segment of the MTG that overlapped with the DMN was active (p.ej., see “autobio-
graphical,” “mind,” “mental states,” “theory of mind,” “default mode” in Figure 13B). Splitting
the left MTG region into anterior and posterior portions typically increased the likelihood that
social cognitive terms were used in the anterior portion and decreases the likelihood that they
were used in the posterior portion (p.ej., see “autobiographical,” “mind,” “mental states,” “the-
ory of mind,” “default mode,” “social cognitive” in Figure 13B).
Finalmente, the range of language-related terms associated with the posterior segment of the left
MTG was much broader than the range of terms associated with the whole left MTG. Este
included terms related to auditory comprehension that were more strongly associated with
the right MTG than the left MTG region, but not more strongly associated with the right
MTG than the posterior segment of the left MTG. Other language-related terms that followed
this pattern included “visual word form,” “English,” “semantics,” and judgment task” (previ-
ously more strongly associated with the left parietal region than other regions), and “read,"
“readers,” and “written” (previously more strongly associated with the right parietal region
than other regions). Además, several language-related terms were only likely to be used
when activity was found in the posterior segment of the left MTG, including the terms “ortho-
graphic,” “visual word,” “audiovisual,” and “speech perception.”
Despite the strong association between language-related terms and activity in the posterior
segment of the left MTG, we did document a couple of language-related terms that were more
likely to be used when activity was found in the anterior segment of the left MTG than the
Neurobiology of Language
108
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
posterior segment. These were “concepts” (posterior left MTG: 59.6%; anterior left MTG: 67%)
and “semantic memory” (posterior left MTG: 64.4%; anterior left MTG: 66.3%). Notablemente, otro
regions were likely to activate these terms as well. The term “semantic memory” was about as
likely to be used when the bilateral pCG was active (66.2%), and only slightly less likely to be
used when the right MTG (62.7%) or the left parietal region of the TN (62.6%) were active. El
term “concepts” also had a relatively moderate likelihood of being used when the left parietal
region of the TN was active (61%).
DISCUSIÓN
Main Effect of Internal Error Correction in Posterior Middle Temporal Gyrus
The main goal of the present research was to evaluate neural evidence for internal error detec-
tion and correction in speech production. Con ese fin, we have leveraged a tongue twister
paradigm that has generated suggestive evidence of internal error correction during speech
production in the pMTG. Eso es, prior research (Okada et al., 2018) had suggested that tongue
twisters designed such that slips would result in nonword errors generate greater activity in
pMTG than tongue twisters designed such that slips would result in word errors, even though
no overt errors occurred. This is intriguing because both conditions involved successfully recit-
ing the same real words, and the only difference between them was an increased potential to
produce a nonword speech error. The one caveat to this finding was that the lexicality effect
ultimately failed to survive cluster correction. Aquí, we have tested for the same lexicality
efecto, using the same stimuli, but in a much larger group of participants. Además, tenemos
introduced a new class of stimuli designed to put greater demands on internal error correction
mechanisms by biasing potential speech errors toward taboo words.
The present study found no evidence of a lexicality effect. This could be because it is a
subtle effect not easily detectible with fMRI. Alternativamente, it could be that the lexicality effect
is caused not by a word versus nonword difference in salience to the error detection system,
but by the nature of the lexical activation system itself, which has been argued to be biased
toward activating real words over nonwords in the first place (Nozari & Dell, 2009). On this
vista, we observed no lexicality effect of internal error processing because lexicality is not a
feature that this system naturally monitors. We submit that our null result is most consistent
with this latter possibility.
A diferencia de, we observed strong taboo word effect on pMTG activation (our a priori ROI).
Biasing potential speech errors toward taboo words during word list recitation generated a
significantly stronger response in the pMTG relative to biasing potential speech errors toward
(neutral) palabras. En tono rimbombante, this effect was observed on correctly produced trials where no
errors occurred, indicating that at some level the system detected taboo words even though
they were not present either in the stimulus or the ultimately expressed utterance. Basado en el
psycholinguistic literature on taboo word slips, which occur at a lower frequency than non-
taboo word slips, we reason that during recitation of the taboo-biased word lists, slips occurred
yielding the lexical activation of a taboo word, cual, due to its salience, had a higher chance
of being corrected than a non-taboo word slip (see Gabitov et al., 2020, for evidence of an
important relation between error detection and the brain’s salience network). This implies that
among the correct trials, internal error correction occurred more frequently for taboo-biased
lists than non-taboo biased lists leading to greater activation of the pMTG, an area implicated
in lexical access (p.ej., Gow, 2012; Indefrey, 2011; Lau & Namyst, 2019) and lexical-semantics
(p.ej., Baldo et al., 2013; Devereux et al., 2013).
Neurobiology of Language
109
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
In a whole brain contrast between taboo and non-taboo word conditions, we demon-
strated that portions of pMTG exhibited the taboo effect bilaterally. A more targeted ROI anal-
ysis using anatomically defined pMTG showed a stronger effect in the left hemisphere. El
taboo effect held across all voxels of left pMTG but was on the cusp of significance in right
pMTG ( pag = 0.05). Within-participant testing for the effect showed that it was quite consistent,
presenting in at least one hemisphere in 76% (norte = 26/34) of participants. In the majority of
cases the effect was bilateral (norte = 22/34), suggesting that a significant group-level effect may
be difficult to map in right pMTG due to individual variability in functional response.
Why should error correction result in greater activation of the pMTG? The hierarchical state
feedback control model proposed by Hickok (2012) at the phonological level of speech pro-
duction can be extended to the lexical level and provides a mechanistic explanation. Hickok
(2012) proposes that phonological encoding is implemented in a sensorimotor-like architec-
ture with a sensory-related system in the posterior temporal lobe that codes the phonological
targets of speech acts and a motor-related system in the inferior frontal lobe that codes
phonological-level motor planning codes (a “syllabary” in psycholinguistic terms) that aim
to reproduce the target. Error detection is achieved by comparing the activated “sensory-
phonological” target with the “motor-phonological” plan: Específicamente, the activated motor-
phonological plan generates a forward prediction signal (ultimately an inhibitory input) a
the sensory-phonological system where a match or mismatch is detected (via a cancellation
mechanism for a match or failure of cancellation for a mismatch). Note that this does not imply
that axons projection from motor to sensory systems are inhibitory signals. Long-range excit-
atory signals are likely to activate local inhibitory interneurons that generate the inhibitory
efecto (Isaacson & Scanziani, 2011). In the case of a mismatch, the sensory-phonological target
remains active (it is not cancelled) and sends a corrective activation to its corresponding
motor-phonological plan. De este modo, error correction results in greater action of the target
sensory-related network. A simple computational simulation presented by Hickok (2012) estafa-
firmed the feasibility of mechanism. Although Hickok (2012) did not propose that the lexical
level was organized with a similar sensorimotor-like architecture, it has recently been
extended to the lexical-syntactic level by Matchin and Hickok (2020; see also Isaacson &
Scanziani, 2011).
Main Effect of Forward Predictive Signals in Auditory Cortex
A secondary aim of the current research was to replicate evidence for forward predictive sig-
nals in auditory cortex (Okada et al., 2018). We successfully replicated this effect, mostrando que
auditory cortex does seem to receive predictive signals during speech production. Eso es, nosotros
found that engaging motor-phonological and lexical-level processes together during silently
articulated speech generated significantly greater activity in and around auditory cortex than
engaging only lexical-level processes during imagined speech. críticamente, the activity we
observed in auditory cortex cannot reflect auditory stimulation because no speech output
was generated in either of the speech task conditions. This forward predictive signal effect
was apparent at the group-level based on a whole brain contrast between speech tasks, y
a ROI analysis that showed that the effect held across all voxels of anatomically defined HG in
both hemispheres. Within-participant testing revealed that the forward predictive signal effect
was about as consistent as the taboo effect and presented in the same proportion of participants
(norte = 26/34). Sin embargo, we also found that the majority of participants who did not exhibit a
mean forward predictive signal effect in HG still presented higher brain response to silently
articulated speech either in portions of HG or in bordering auditory association areas, demostración
that there can be substantial individual variability in the precise locus of the effect.
Neurobiology of Language
110
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
Response around auditory cortex can be explained in the context of a mismatch error
between the auditory consequence of a planned articulatory sequence that is expected by
the system, and the absence of any such signal as a result of silent articulation. Mismatch
error induced by altered auditory feedback has produced a similar pattern of response in
auditory cortex (Tourville et al., 2008). Speech induced suppression of auditory response
as documented by electrocorticographic, electroencephalographic, and magnetoencephalo-
graphic recordings has also been interpreted in the context of a similar kind of mismatch error
between a forward prediction of auditory consequences of speech and actual auditory input
(Ford et al., 2010; Greenlee et al., 2011; Guenther & Hickok, 2015; Ventura et al., 2009). En
models of motor control, mismatch is often extended not only to the content of speech, pero
also to its fundamental acoustic properties (p.ej., pitch; Guenther & Hickok, 2015). Este
explains why FMRI studies have demonstrated a similar decrease in auditory response during
silently articulated speech when it is coupled with hearing another person produce the same
speech stimuli (Agnew et al., 2013).
Network Associated With the Taboo Effect
Comparison between the taboo and word condition revealed preferential response to the
taboo condition in areas outside pMTG and commonly associated with the DMN, incluido
in AG and adjacent portions of sLOC, aMTG, pCG, paraCG, SFG, and several medial frontal
areas. An additional comparison between the taboo condition and baseline fixation demon-
strated that most areas that showed a taboo effect were deactivated during the taboo condition.
The strongest evidence for above-baseline response was found in left pMTG, although we note
that an areal analysis we performed also implied above-baseline response could be found
within left AG and left pITG at significance thresholds lower than those used in the present
study for direct contrast analyses.
The finding that left pMTG showed a taboo effect and had the clearest above-baseline
response during the taboo condition implicated this area in internal error correction above
other areas that exhibited a taboo effect. It also implied that internal error correction processes
are likely left lateralized. This is further supported by the aforementioned ROI analysis of
pMTG that demonstrated a stronger effect in the left hemisphere. The broader network of areas
that exhibited a taboo effect also showed evidence for left lateralization—maximum activation
within areas was significantly higher in the left hemisphere.
Recent research has also implicated left pMTG in word–picture interference using taboo
distractor words (Hansen et al., 2019). This work has also demonstrated increased activity
for taboo distractors in a thalamocortical network of areas that do not appear in the taboo
effect we map using tongue twisters. The functional interpretation of this thalamocortical net-
work is consistent with our results here—left pMTG is interpreted to reflect lexical processes,
whereas other areas of the thalamocortical network (p.ej., aCG, thalamus, IFG) support pro-
cessing the arousing properties of taboo words and deploying attention mechanisms to sup-
press them. That our results did not reveal areas strongly associated with attention suggests that
internal correction of speech errors, including taboo speech errors, may not require interven-
tion from attentional mechanisms.
Although we predicted a robust response in left pMTG for taboo tongue twisters, we did not
expect them to generate increased, albeit below-baseline, activity across areas of the DMN.
We suggest that the taboo effect tapped into a broader lexical-semantic network and that activ-
ity in the DMN was driven by the social features of taboo words. Areas of the DMN appear to
show a preference for social concepts embedded in narrative stories (Huth et al., 2016) y
Neurobiology of Language
111
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
respond more strongly to words that connect to interpersonal interactions compared to those
that do not (Lin et al., 2015, 2020). Observations like these have been used to argue that the
brain areas responsible for theory of mind encode intentionality in semantic space (Binder
et al., 2016). In this context, that taboo words generally elicited stronger response across
the DMN is less surprising. Taboo words tend to be highly versatile and convey very different
intentions (positivo, negative, and otherwise) depending on the context in which they are used
(Vingerhoets et al., 2013). Por ejemplo, work in natural language processing has demonstrated
that detection of hate speech is considerably less successful, suffering from high false positive
tarifas, when the pragmatic functions of taboo words are not explicitly modeled (Davidson
et al., 2017).
Networks Associated With Speech Tasks
Previous research (Okada et al., 2018) has demonstrated that the contrast between silently
articulated and imagined speech produces activity that is right lateralized in certain regions
(p.ej., sensorimotor cortex). In the present study we broke this contrast apart into anatomical
areas, which showed that although generally the same areas were recruited bilaterally and
maximum response within areas did not differ between hemispheres, larger portions of areas
in the left hemisphere were sensitive to the difference between articulated and imagined
speech, while the smaller portions of areas in the right hemisphere that were sensitive to this
difference presented larger effect sizes. En general, this describes a left hemisphere network that
was more sensitive to differences between silently articulated and imagined speech and a right
hemisphere network with higher specificity. De hecho, the largest two effect sizes we reported
were in the right PT and HG, underlining the robustness of auditory response during silently
articulated speech. It is worth noting too that HG was one of the few areas that broke away
from the trend we described across the contrast—response in HG was substantially higher in
the right hemisphere, a much larger portion of the right HG was sensitive to the contrast, y
right HG also showed a larger effect size. This pattern echoed our findings at the participant
nivel, where a forward prediction effect was typically present in HG for both hemispheres (norte =
20/26), but when the effect was occasionally unilateral it was most often found in the right
hemisferio (norte = 5). De este modo, it is possible that right auditory cortex plays a larger role in process-
ing forward predictive models in speech.
A nivel de grupo, greater response during silently articulated than imagined speech was
found in a network that closely followed recent pooled analysis of speech production exper-
elementos (Tourville et al., 2019). The core of this network was highly consistent with the prior
study we sought to replicate and primarily covered areas connected to phonological and artic-
ulatory processes, including the primary sensorimotor and somatosensory cortices (es decir., pre-
central and postcentral gyri), aCG, SMA, the cerebellum, the IFG, the PT, the supramarginal
gyrus, the insula, and the adjacent central, frontal, and parietal operculum (p.ej., Hickok, 2012,
2014; Kearney & Guenther, 2019; Precio, 2012; Tourville et al., 2019). As alluded to earlier, el
silent articulation network also contained areas involved in auditory processing, which were
HG and the planum polare, but also associative auditory areas in anterior and posterior seg-
ments of the STG (Binder et al., 1996; Kearney & Guenther, 2019). We note that the anatom-
ical STG we used for reference included portions of the posterior superior temporal sulcus
putatively involved in speech perception (Venezia et al., 2017). Stronger activity in these areas
is consistent with the behavioral research this study builds on, which has demonstrated that
increasing the amount of articulation in speech imagery induces higher rates of phonological
speech errors that involve similarly articulated phonemes (Oppenheim & Dell, 2010). El
silent articulation network also recruited areas not as closely related to speech production.
Neurobiology of Language
112
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
Por ejemplo, some of these areas are commonly associated with executive function (es decir., el
intraparietal sulcus, which crosses anatomically defined superior parietal lobule, posterior
supramarginal gyrus, and sLOC; Velenosi et al., 2020), orthographic processing (es decir., tempor-
ooccipital portion of ITG and neighboring fusiform areas; Precio, 2012), and lexical and/or
semantic processing (es decir., MTG, temporal pole, middle frontal gyrus, SFG, and an area of ana-
tomically defined orbitofrontal cortex that maps onto the pars orbitalis portion of the IFG in
atlases that make the distinction between pars orbitalis and pars triangularis; p.ej., Binder et al.,
2009; Precio, 2012). Sin embargo, all of these areas play a role during speech production
(Precio, 2012; Tourville et al., 2019), and it is possible that the phonological-level processes
that distinguished silently articulated speech from imagined speech had the effect of producing
greater activity in areas that interface with phonological features.
Curiously, some areas responded more strongly to imagined than silently articulated speech,
including paraHG, AG, sLOC, temporal fusiform cortex, OccP, and several medial frontal areas.
These are many of the same areas that exhibited stronger response to taboo tongue twisters rel-
ative to word tongue twisters, and just as in the case of the taboo effect, they were all deactivated
during the imagined speech condition. Sin embargo, areal effect analysis hinted that left AG, FP, y
anterior paraHG may contain above-baseline response at lower significance thresholds than
those used in the current work for contrast analyses. Although prior research did not observe
any areas associated with imagined speech (Okada et al., 2018), a similar network has been
documented in auditory and visual imagery tasks (p.ej., Daselaar et al., 2010; Pearson, 2019;
Tian et al., 2016), and functional connectivity between the AG and the other areas of this net-
work has been shown to increase during imagined musical performance (instrumental or vocal;
Tanaka & Kirino, 2019). The AG itself has been routinely implicated in semantic, episodic, y
memoria autobiográfica, and response in this area has been related to subjective vividness
during episodic memory retrieval and encoding (Tibon et al., 2019). Similarity between areas
that show up in the DMN and areas involved in various mental imagery tasks (including auditory
imagery) has implied a close relationship between mental imagery and the kinds of internally
directed processes that have been connected to DMN areas (Daselaar et al., 2010; Pearson,
2019). De este modo, the pattern of activity that we found could simply reflect the fact that imagined
speech is relatively more internally oriented than articulated speech, or perhaps that imagined
articulation taps into a domain general imagery network.
Given the association between many of the areas in the imagined speech network and
memory, another possibility is that imagined speech recruits memory systems to facilitate
imaging the auditory consequences of simulated speech under conditions where precise pre-
diction is more difficult. Although we have shown here that forward predictions were stronger
when articulatory features were salient in speech imagery (es decir., silently articulated speech),
they still appear to be generated in a weaker form when such features are impoverished or
absent (es decir., imagined speech; Tian et al., 2016). Más, if the quality of forward predictions
produced in this memory-driven process (Tian et al., 2016) can be substantially different, él
could also explain our observation that different portions of auditory cortex can simulta-
neously show greater activity for silently articulated and imagined speech in a minority of par-
ticipants. At least one other explanation is that the areas associated with imagined speech
reflect a different process for maintaining speech images in memory. Por ejemplo, alguno
research has related activity in areas of the imagined speech network to the detail of ongoing
thoughts during working memory maintenance (Sormaz et al., 2018), and other studies have
implicated the paraHG in the maintenance of novel information during working memory
(Schon et al., 2016). Because verbal memory taps into areas involved in speech production
(Buchsbaum & D'Esposito, 2019), it may be the case that weaker access to articulatory features
Neurobiology of Language
113
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
in imagined speech requires additional engagement in nonverbal memory systems for main-
taining speech imagery. More research is necessary to adjudicate between these possibilities
and we emphasize that no areas that generated greater response to imagined than articulated
speech showed clear above-baseline response during the imagined speech condition.
Decoding the Taboo Network
We have suggested that the richness of social features in taboo words—particularly
intentionality—has driven activity to areas of the DMN in the taboo contrast. We have also
suggested that activity in the left pMTG does not reflect these social features, in part because
this area alone shows clear above-baseline response during the recitation of taboo tongue
twisters. A functional distinction between pMTG and other areas of the taboo network was
further supported by a meta-analysis of studies that frequently mentioned the DMN, cual
showed strong overlap in most areas, but not pMTG.
Our explanation for the response pattern in the TN predicted that pMTG should be asso-
ciated with language-specific processes, especially at the word level, while other areas of the
TN should be associated with DMN functions relevant to processing intentionality. probamos
these predictions by decoding the taboo network and its constituent regions in Neurosynth
(Yarkoni et al., 2011). Decoding the TN involved correlating activity that we mapped as part
of the taboo effect (es decir., taboo > words) with the likelihood of finding activity in different
groups of neuroimaging studies that frequently mentioned different neuroimaging terms
(es decir., meta-analyses for different terms in Neurosynth). This unsurprisingly revealed that
DMN studies were most strongly associated with TN activity, followed by studies on func-
tions commonly ascribed to areas of the DMN (p.ej., “theory of mind,” “mentalization,” “self-
reference,” “autobiographical memory,” “social cognition”). Sin embargo, activity in the TN was
also related to comprehension (p.ej., “comprehension,” “semantic,” “sentence”). When we
correlated the same term-based meta-analyses with the likelihood of observing activity in the
DMN (es decir., the earlier DMN meta-analysis), we found that comprehension terms showed by
far the largest difference between association strength to the TN and the DMN. Además,
this difference in association was primarily driven by activity in the left hemisphere, cual
also overlapped more extensively with the DMN. This pattern suggested that left hemisphere
areas of the TN that fall outside the DMN play a more substantial role in comprehension, y
therefore that left pMTG may be driving the network-level association between the TN and
comprensión.
We directly tested whether the posterior portions of the left MTG cluster in the TN was more
closely connected to word-level processing by decoding the TN at a regional level, decoding
both the entire MTG cluster as well as the posterior and anterior portions of it that sit outside
and inside of the DMN, respectivamente. Decoding was made to be more sensitive to the spatial
distribution of activity within smaller regions by capturing the mean likelihood that a study
uses a particular term (that is frequently used in neuroimaging studies) if there is activity in
each voxel of a TN cluster. We found that although virtually all language-related terms were
more likely to be used when there was activity in the MTG compared to other regions of the
TN, seeing activity in any cluster of the TN, including the MTG, was most likely to signal
engagement in “autobiographical memory,” “theory of mind,” “mentalization,” or “default
mode network.” This is exactly what might have been expected based on the results of the
network-level decoding. Sin embargo, separating the left MTG cluster based on overlap with
the DMN produced a pattern that supported word processing in the posterior segment of this
región. Primero, we found that activity in the anterior segment of MTG was still most likely to
Neurobiology of Language
114
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
signal DMN-related functions, whereas activity in the posterior segment was most likely to
signal engagement in language processes. Más, relative to the whole MTG cluster, the like-
lihood of using language-related terms generally increased and the likelihood of using
DMN-related terms generally decreased when the posterior segment of MTG was active.
The reverse of this trend was observed in the anterior segment of MTG. Closer inspection of
the terms that were most likely to be used when the posterior segment of the MTG was active
revealed an association with comprehension that was broad, covering both sentence and word
estímulos. Terms that were likely to be used such as “speaker,” “acoustic,” “lexical,” “meaning,"
“comprehension,” “word,” “linguistic,” and “phonological” were all consistent with a role for
the pMTG in lexical processing. Other terms, such as “semantic,” “sentences,” “language
comprensión,” “syntactic,” and “reading” may reflect the activity of lexical processes
engaged during semantic tasks. In any case, these patterns clearly showed that one specific
portion of the TN—the left pMTG—was much more strongly associated with language, and at
least played a relatively more significant role in lexical processing.
Regional decoding of the TN also indicated that the posterior segment of MTG was
involved in semantic processes, but not necessarily semantic memory. Activity in the anterior
segment of MTG was more strongly associated with concepts and semantic memory than
activity in the posterior segment of MTG. Más, semantic memory was just as strongly asso-
ciated with the pCG and had a remarkably weak association with the posterior segment of
MTG (es decir., less than 50% likelihood of term use). The stronger association to both social cog-
nitive terms and semantic memory in areas of the TN that happened to show stronger overlap
with the DMN was consistent with our interpretation that taboo words drive response in areas
of the DMN as a result of their semantic features, mainly intentionality. This interpretation
aligns with a recent proposal for a componential model of semantic representation that sug-
gests areas involved in theory of mind and mentalization are used to encode and understand
words that place stronger emphasis on intentionality (Binder et al., 2016). It also aligns with a
growing body of work that shows words rich in social semantic features tap into areas that
overlap with the DMN (p.ej., Huth et al., 2016; Lin et al., 2015, 2018, 2020; Vingerhoets
et al., 2013).
Conclusions and Limitations
En resumen, the present study has provided evidence that forward predictive signals are pres-
ent in auditory cortex during speech production, and that error detection and correction in
speech involves left pMTG. We have shown that silently articulated speech produces greater
activity than imagined speech within early auditory areas, even though no auditory input is
present during silently articulated speech. We have also shown that biasing potential word
errors toward taboo words rather than neutral words during word list recitation generates
greater activity in left pMTG, even though no overt speech errors are produced in either con-
condición. Although we found similar response in areas that demonstrably overlapped with the
DMN as well as right pMTG, these areas appeared to respond more robustly to baseline fix-
ation than word list recitation. Más, we have provided evidence from meta-analysis that
activity in left pMTG is most likely to reflect word-level processes and by extension word-level
error correction. We have hypothesized, with some support from meta-analysis, that the other
areas of the taboo network are involved in processing the social semantic features of taboo
palabras, particularly intentionality.
Sin embargo, there are several limitations to the present research. Primero, overt speech typ-
ically increases the amount of motion observed during scanning. Although here we used
Neurobiology of Language
115
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
mouthed speech, it is difficult to assess the full impact motion had on the results. En general,
we reported what can be considered as relatively low motion in our group of participants
(p.ej., Eichenbaum et al., 2021; Yang et al., 2019). We also tried to mitigate the possibility that
motion had a substantial impact on our results by showing that our primary effects of interest
(es decir., the ROI analyses for forward prediction and internal error correction) are not correlated
with mean framewise displacement. One other concern is that overt speech has been shown
to produce susceptibility artifacts. It is uncertain the extent to which such artifacts may have
impinged on our results, but we note that past studies have found these artifacts to dispro-
portionately affect insular and opercular areas (Kemeny et al., 2005), whereas the current
study focuses on lateral temporal cortex. Finalmente, our failure to find a lexicality effect in
pMTG for tongue twisters suggests that evidence for such an effect trending toward signifi-
cance in prior research may reflect a false positive result. Sin embargo, we suspect this is not the
case and that a larger amount of data is necessary to achieve significance for such a small
effect size, especially considering that our manipulation of tongue twisters to elicit a greater
potential for taboo word errors did successfully produce stronger activity in pMTG, as we
had hypothesized.
EXPRESIONES DE GRATITUD
The authors wish to thank the individuals that participated in this research. Discretionary
research funding was provided by University of California, Irvine.
INFORMACIÓN DE FINANCIACIÓN
Gregory Hickok, National Institute on Deafness and Other Communication Disorders (https://
dx.doi.org/10.13039/100000055), Award ID: R01 DC009659. Alex Teghipco, National Sci-
ence Foundation (https://dx.doi.org/10.13039/100000001), Award ID: DGE-1321846.
CONTRIBUCIONES DE AUTOR
Alex Teghipco: Curación de datos: Lead; Análisis formal: Lead; Investigación: Lead; Metodología:
Secundario; Software: Lead; Visualización: Lead; Escritura – borrador original: Lead; Escritura – revisión
& edición: Equal. Kayoko Okada: Conceptualización: Equal; Análisis formal: Secundario; Fund-
ing acquisition: Secundario; Investigación: Secundario; Metodología: Equal; Recursos: Equal;
Supervisión: Equal; Escritura – borrador original: Equal; Escritura – revisión & edición: Equal. Emma
Murphy: Curación de datos: Secundario; Análisis formal: Secundario; Investigación: Secundario;
Recursos: Secundario. Gregory Hickok: Conceptualización: Lead; Adquisición de financiación: Lead;
Metodología: Equal; Supervisión: Equal; Escritura – borrador original: Secundario; Escritura – revisión
& edición: Equal.
DATA AND CODE AVAILABILITY STATEMENT
Group-level contrast maps for this study can be found on neurovault (https://identifiers.org
/neurovault.collection:12982). Raw data is available to download on data dryad (https://doi
.org/10.7280/D1BH77). Example code for replicating meta-analyses can be found on our
GitHub repository (https://github.com/alexteghipco/MetaAnalysisResources). FMRI analyses
can be replicated with FMRIB’s Software Library ( Versión 6.0.5.1; Jenkinson et al., 2012),
using parameters provided in the methods (see FSL course preparatory material for guides
on how to replicate purely using GUIs). Additional parameter information that may be useful
is provided along with the neurovault collection linked above as metadata. Surface visualiza-
tions can be replicated using the brainSurfer toolbox for MATLAB (https://doi.org/10.5281
Neurobiology of Language
116
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
/zenodo.7271544). A saved GUI state that can be loaded into brainSurfer (Teghipco, 2022) a
show interactive and manipulable surface renderings identical to those shown in our figures
(es decir., including all thresholds, colormaps, and other visualization settings) can be found on fig-
compartir (10.6084/m9.figshare.21357525).
REFERENCIAS
Agnew, z. K., McGettigan, C., Banks, B., & Scott, S. k. (2013).
Articulatory movements modulate auditory responses to speech.
NeuroImagen, 73, 191–199. https://doi.org/10.1016/j.neuroimage
.2012.08.020, PubMed: 22982103
Baars, B. J., Motley, METRO. T., & MacKay, D. GRAMO. (1975). Output editing
for lexical status in artificially elicited slips of the tongue. Diario
of Verbal Learning and Verbal Behavior, 14(4), 382–391. https://
doi.org/10.1016/S0022-5371(75)80017-X
Baldo, j. v., Arévalo, A., Patterson, j. PAG., & Dronkers, norte. F. (2013).
Grey and white matter correlates of picture naming: Evidencia
from a voxel-based lesion analysis of the Boston Naming Test.
Corteza, 49(3), 658–667. https://doi.org/10.1016/j.cortex.2012
.03.001, PubMed: 22482693
Binder, j. r., Conant, l. l., Humphries, C. J., Fernandino, l.,
simons, S. B., Aguilar, METRO., & Desai, R. h. (2016). Toward a
brain-based componential semantic representation. Cognitivo
Neuropsychology, 33(3–4), 130–174. https://doi.org/10.1080
/02643294.2016.1147426, PubMed: 27310469
Binder, j. r., Desai, R. h., Tumbas, W.. w., & Conant, l. l. (2009).
Where is the semantic system? A critical review and
meta-analysis of 120 functional neuroimaging studies. Cerebral
Corteza, 19(12), 2767–2796. https://doi.org/10.1093/cercor
/bhp055, PubMed: 19329570
Binder, j. r., Frost, j. A., Hammeke, t. A., Rao, S. METRO., & Cox, R. W..
(1996). Function of the left planum temporale in auditory and
linguistic processing. Cerebro, 119(4), 1239–1247. https://doi.org
/10.1093/brain/119.4.1239, PubMed: 8813286
Borgo, METRO., Soranzo, A., & Grassi, METRO. (2012). Psychtoolbox: Sound,
keyboard and mouse. In MATLAB for psychologists (páginas. 249–273).
Saltador. https://doi.org/10.1007/978-1-4614-2197-9_10
Buchsbaum, B. r., & D'Esposito, METRO. (2019). A sensorimotor view of
verbal working memory. Corteza, 112, 134–148. https://doi.org/10
.1016/j.cortex.2018.11.010, PubMed: 30639088
Corley, METRO., Brocklehurst, PAG. h., & Moat, h. S. (2011). Error biases
in inner and overt speech: Evidence from tongue twisters. Diario
of Experimental Psychology: Aprendiendo, Memoria, and Cognition,
37(1), 162–175. https://doi.org/10.1037/a0021321, PubMed:
21244112
Daselaar, S. METRO., Porat, y., Huijbers, w., & Pennartz, C. METRO. (2010).
Modality-specific and modality-independent components of the
human imagery system. NeuroImagen, 52(2), 677–685. https://doi
.org/10.1016/j.neuroimage.2010.04.239, PubMed: 20420931
Davidson, T., Warmsley, D., Macy, METRO., & Weber, I. (2017). Auto-
mated hate speech detection and the problem of offensive lan-
guage. Proceedings of the International AAAI Conference on
Web and Social Media, 11(1), 512–515. https://doi.org/10.1609
/icwsm.v11i1.14955
Dell, GRAMO. S. (1986). A spreading-activation theory of retrieval in sen-
tence production. Revisión psicológica, 93(3), 283–321. https://
doi.org/10.1037/0033-295X.93.3.283, PubMed: 3749399
Desikan, R. S., Ségonne, F., pescado, B., quinn, B. T., Dickerson,
B. C., Blacker, D., Buckner, R. l., Valle, A. METRO., Maguire, R. PAG.,
Hyman, B. r., Alberto, METRO. S., & Killiany, R. j. (2006). An auto-
mated labeling system for subdividing the human cerebral cortex
on MRI scans into gyral based regions of interest. NeuroImagen,
31(3), 968–980. https://doi.org/10.1016/j.neuroimage.2006.01
.021, PubMed: 16530430
Devereux, B. J., Clarke, A., Marouchos, A., & tyler, l. k. (2013).
Representational similarity analysis reveals commonalities and
differences in the semantic processing of words and objects. Jour-
nal of Neuroscience, 33(48), 18906–18916. https://doi.org/10
.1523/JNEUROSCI.3809-13.2013, PubMed: 24285896
Eichenbaum, A., Pappas, I., Lurie, D., cohen, j. r., & D'Esposito,
METRO. (2021). Differential contributions of static and time-varying
functional connectivity to human behavior. Network Neurosci-
ence, 5(1), 145–165. https://doi.org/10.1162/netn_a_00172,
PubMed: 33688610
Vado, j. METRO., Roach, B. J., & Mathalon, D. h. (2010). Assessing cor-
ollary discharge in humans using noninvasive neurophysiologi-
cal methods. Nature Protocols, 5(6), 1160–1168. https://doi.org
/10.1038/nprot.2010.67, PubMed: 20539291
Gabitov, MI., Lungu, o., Albouy, GRAMO., & Doyon, j. (2020). Movement
errors during skilled motor performance engage distinct prediction
error mechanisms. Communications Biology, 3(1), Article 763.
https://doi.org/10.1038/s42003-020-01465-4, PubMed: 33311566
Garnham, A., Shillcock, R. C., Marrón, GRAMO. D., Mill, A. I. D., & Cutler,
A. (1982). Slips of the tongue in the London-Lund corpus of spon-
taneous conversation. In A. Cutler (Ed.), Slips of the tongue and
language production (páginas. 251–263). Walter de Gruyter. https://
doi.org/10.1515/9783110828306.251
Gow, D. w., Jr. (2012). The cortical organization of lexical knowl-
borde: A dual lexicon model of spoken language processing.
Brain and Language, 121(3), 273–288. https://doi.org/10.1016/j
.bandl.2012.03.005, PubMed: 22498237
Grabner, GRAMO., Janke, A. l., Budge, METRO. METRO., Herrero, D., Pruessner, J., &
collins, D. l. (2006). Symmetric atlasing and model based seg-
mentation: An application to the hippocampus in older adults. En
International Conference on Medical Image Computing and
Computer-Assisted Intervention (páginas. 58–66). Saltador. https://
doi.org/10.1007/11866763_8, PubMed: 17354756
verdelee, j. D. w., Jackson, A. w., Chen, F., larson, C. r., Oya, h.,
Kawasaki, h., Chen, h., & Howard, METRO. A., III. (2011). Humano
auditory cortical activation during self-vocalization. PLOS One,
6(3), Article e14744. https://doi.org/10.1371/journal.pone
.0014744, PubMed: 21390228
Guenther, F. h., & Hickok, GRAMO. (2015). Role of the auditory system in
speech production. Handbook of Clinical Neurology, 129, 161–
175. https://doi.org/10.1016/ B978-0-444-62630-1.00009-3,
PubMed: 25726268
Hansen, S. J., McMahon, k. l., & de Zubicaray, GRAMO. I. (2019). El
neurobiology of taboo language processing: fMRI evidence dur-
ing spoken word production. Social Cognitive and Affective Neu-
roscience, 14(3), 271–279. https://doi.org/10.1093/scan/nsz009,
PubMed: 30715549
Neurobiology of Language
117
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
Hickok, GRAMO. (2012). Computational neuroanatomy of speech pro-
ducción. Naturaleza Reseñas Neurociencia, 13(2), 135–145. https://
doi.org/10.1038/nrn3158, PubMed: 22218206
Hickok, GRAMO. (2014). The architecture of speech production and the
role of the phoneme in speech processing. Language and Cogni-
tive Processes, 29(1), 2–20. https://doi.org/10.1080/01690965
.2013.834370, PubMed: 24489420
Huth, A. GRAMO., de Heer, W.. A., Griffiths, t. l., Theunissen, F. MI., &
Gallant, j. l. (2016). Natural speech reveals the semantic maps
that tile human cerebral cortex. Naturaleza, 532(7600), 453–458.
https://doi.org/10.1038/nature17637, PubMed: 27121839
Indefrey, PAG. (2011). The spatial and temporal signatures of word pro-
duction components: A critical update. Fronteras en psicología,
2, 255. https://doi.org/10.3389/fpsyg.2011.00255, PubMed:
22016740
Isaacson, j. S., & Scanziani, METRO. (2011). How inhibition shapes cor-
tical activity. Neurona, 72(2), 231–243. https://doi.org/10.1016/j
.neuron.2011.09.027, PubMed: 22017986
Jacewicz, MI., Fox, R. A., & Wei, l. (2010). Between-speaker and
within-speaker variation in speech tempo of American English. El
Journal of the Acoustical Society of America, 128(2), 839–850.
https://doi.org/10.1121/1.3459842, PubMed: 20707453
Jenkinson, METRO., Bannister, PAG., Brady, METRO., & Herrero, S. (2002).
Improved optimization for the robust and accurate linear registra-
tion and motion correction of brain images. NeuroImagen, 17(2),
825–841. https://doi.org/10.1006/nimg.2002.1132, PubMed:
12377157
Jenkinson, METRO., beckman, C. F., Behrens, t. mi. J., lana rica, METRO. w.,
& Herrero, S. METRO. (2012). FSL. NeuroImagen, 62(2), 782–790. https://
doi.org/10.1016/j.neuroimage.2011.09.015, PubMed: 21979382
Loco, METRO. (1999). Internal models for motor control and trajectory
planificación. Opinión actual en neurobiología, 9(6), 718–727.
https://doi.org/10.1016/S0959-4388(99)00028-8, PubMed:
10607637
Kearney, MI., & Guenther, F. h. (2019). Articulating: el neural
mechanisms of speech production. Idioma, Cognition and
Neurociencia, 34(9), 1214–1229. https://doi.org/10.1080
/23273798.2019.1589541, PubMed: 31777753
Kemeny, S., S.M, F. P., Hijo, r., & Braun, A. R. (2005). Comparison
of continuous overt speech fMRI using BOLD and arterial spin
labeling. Mapeo del cerebro humano, 24(3), 173–183. https://doi
.org/10.1002/hbm.20078, PubMed: 15486986
Lau, mi. F., & Namyst, A. (2019). fMRI evidence that left posterior
temporal cortex contributes to N400 effects of predictability
independent of congruity. Brain and Language, 199, 104697.
https://doi.org/10.1016/j.bandl.2019.104697, PubMed:
31585316
Levelt, W.. j. (1983). Monitoring and self-repair in speech. Cogni-
ción, 14(1), 41–104. https://doi.org/10.1016/0010-0277(83)
90026-4, PubMed: 6685011
Levelt, W.. j. (1989). Speaking: From intention to articulation. CON
Prensa.
Levelt, W.. j. (1992). Accessing words in speech production: Stages,
processes and representations. Cognición, 42(1–3), 1–22. https://
doi.org/10.1016/0010-0277(92)90038-j, PubMed: 1582153
Levelt, W.. J., Roelofs, A., & Meyer, A. S. (1999). A theory of lexical
access in speech production. Ciencias del comportamiento y del cerebro, 22(1),
1–38. https://doi.org/10.1017/S0140525X99001776, PubMed:
11301520
lin, NORTE., Bi, y., zhao, y., luo, C., & li, X. (2015). The theory-of-mind
network in support of action verb comprehension: Evidencia de
an fMRI study. Brain and Language, 141, 1–10. https://doi.org/10
.1016/j.bandl.2014.11.004, PubMed: 25498409
lin, NORTE., Wang, X., Xu, y., Wang, X., Hua, h., zhao, y., & li, X.
(2018). Fine subdivisions of the semantic network supporting
social and sensory–motor semantic processing. Corteza cerebral,
28(8), 2699–2710. https://doi.org/10.1093/cercor/ bhx148,
PubMed: 28633369
lin, NORTE., Xu, y., Cual, h., zhang, GRAMO., zhang, METRO., Wang, S., Hua, h.,
& li, X. (2020). Dissociating the neural correlates of the sociality
and plausibility effects in simple conceptual combination. Cerebro
Structure and Function, 225(3), 995–1008. https://doi.org/10
.1007/s00429-020-02052-3, PubMed: 32140848
Mars, R. B., Neubert, F. X., Noonan, METRO. PAG., Sallet, J., Toni, I., &
Rushworth, METRO. F. (2012). On the relationship between the
“default mode network” and the “social brain.” Frontiers in
Human Neuroscience, 6, 189. https://doi.org/10.3389/fnhum
.2012.00189, PubMed: 22737119
Matchin, w., & Hickok, GRAMO. (2020). The cortical organization of syn-
tax. Corteza cerebral, 30(3), 1481–1498. https://doi.org/10.1093
/cercor/bhz180, PubMed: 31670779
MathWorks. (2022). MATLAB. https://www.mathworks.com
/products/matlab.html
moller, J., Jansma, B. METRO., Rodriguez-Fornells, A., & monedas, t. F.
(2007). What the brain does before the tongue slips. Cerebral
Corteza, 17(5), 1173–1178. https://doi.org/10.1093/cercor
/bhl028, PubMed: 16831855
Motley, METRO. T., Camden, C. T., & Baars, B. j. (1982). Covert formu-
lation and editing of anomalies in speech production: Evidencia
from experimentally elicited slips of the tongue. Journal of Verbal
Learning and Verbal Behavior, 21(5), 578–594. https://doi.org/10
.1016/S0022-5371(82)90791-5
Mumford, j. A., & Nichols, t. mi. (2008). Power calculation for group
fMRI studies accounting for arbitrary design and temporal auto-
correlation. NeuroImagen, 39(1), 261–268. https://doi.org/10
.1016/j.neuroimage.2007.07.061, PubMed: 17919925
Nooteboom, S. GRAMO. (2005). Lexical bias revisited: Detecting,
rejecting and repairing speech errors in inner speech. Discurso
Comunicación, 47(1–2), 43–58. https://doi.org/10.1016/j
.specom.2005.02.003
Nozari, NORTE., & Dell, GRAMO. S. (2009). More on lexical bias: How effi-
cient can a “lexical editor” be? Journal of Memory and Language,
60(2), 291–307. https://doi.org/10.1016/j.jml.2008.09.006,
PubMed: 20126302
Nozari, NORTE., Dell, GRAMO. S., & Schwartz, METRO. F. (2011). Is comprehension
necessary for error detection? A conflict-based account of monitor-
ing in speech production. Psicología cognitiva, 63(1), 1–33. https://
doi.org/10.1016/j.cogpsych.2011.05.001, PubMed: 21652015
Okada, K., Matchin, w., & Hickok, GRAMO. (2018). Neural evidence for
predictive coding in auditory cortex during speech production.
Boletín psiconómico & Revisar, 25(1), 423–430. https://doi.org
/10.3758/s13423-017-1284-x, PubMed: 28397076
Oppenheim, GRAMO. METRO., & Dell, GRAMO. S. (2008). Inner speech slips exhibit
lexical bias, but not the phonemic similarity effect. Cognición,
106(1), 528–537. https://doi.org/10.1016/j.cognition.2007.02
.006, PubMed: 17407776
Oppenheim, GRAMO. METRO., & Dell, GRAMO. S. (2010). Motor movement matters:
The flexible abstractness of inner speech. Memoria & Cognición,
38(8), 1147–1160. https://doi.org/10.3758/ MC.38.8.1147,
PubMed: 21156877
Pearson, j. (2019). The human imagination: The cognitive neuro-
science of visual mental imagery. Naturaleza Reseñas Neurociencia,
20(10), 624–634. https://doi.org/10.1038/s41583-019-0202-9,
PubMed: 31384033
Fuerza, j. D., Barnes, k. A., Snyder, A. Z., Schlaggar, B. l., &
Petersen, S. mi. (2012). Spurious but systematic correlations in
Neurobiology of Language
118
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Predictive coding and error correction
functional connectivity MRI networks arise from subject motion.
NeuroImagen, 59(3), 2142–2154. https://doi.org/10.1016/j
.neuroimage.2011.10.018, PubMed: 22019881
Precio, C. j. (2012). A review and synthesis of the first 20 years of PET
and fMRI studies of heard speech, spoken language and reading.
NeuroImagen, 62(2), 816–847. https://doi.org/10.1016/j
.neuroimage.2012.04.062, PubMed: 22584224
Runnqvist, MI., Chanoine, v., Strijkers, K., Patamadilok, C., Bonnard,
METRO., Nazarian, B., Sein, J., Anton, J.-L., Belin, PAG., & Alario, F.-X.
(2020). The cerebellum is involved in internal and external
speech error monitoring. bioRxiv. https://doi.org/10.1101/2020
.11.26.400317
Schon, K., Newmark, R. MI., ross, R. S., & Stern, C. mi. (2016). A work-
ing memory buffer in parahippocampal regions: Evidence from a
load effect during the delay period. Corteza cerebral, 26(5),
1965–1974. https://doi.org/10.1093/cercor/ bhv013, PubMed:
25662713
Shadmehr, r., & Krakauer, j. W.. (2008). A computational neuroanatomy
for motor control. Experimental Brain Research, 185(3), 359–381.
https://doi.org/10.1007/s00221-008-1280-5, PubMed: 18251019
Siemens Medical Solutions. (2022). 3T Prisma scanner [Apparatus].
https://www.siemens-healthineers.com/en-us/magnetic
-resonance-imaging/3t-mri-scanner/magnetom-prisma
Sormaz, METRO., Murphy, C., Wang, h. T., Hymers, METRO., Karapanagiotidis,
T., Poerio, GRAMO., Margulies, D. S., Jeffereis, MI., Smallwood, j. (2018).
Default mode network can support the level of detail in experi-
ence during active task states. Proceedings of the National Acad-
emy of Sciences, 115(37), 9318–9323. https://doi.org/10.1073
/pnas.1721259115, PubMed: 30150393
Tanaka, S., & Kirino, mi. (2019). Increased functional connectivity of
the angular gyrus during imagined music performance. Frontiers
in Human Neuroscience, 13, 92. https://doi.org/10.3389/fnhum
.2019.00092, PubMed: 30936827
Teghipco, A. (2022). brainSurfer ( Versión 2.0.2) [Computer soft-
mercancía]. https://doi.org/10.5281/zenodo.7271544
tian, X., Zarate, j. METRO., & Poeppel, D. (2016). Mental imagery of
speech implicates two mechanisms of perceptual reactivation.
Corteza, 77, 1–12. https://doi.org/10.1016/j.cortex.2016.01.002,
PubMed: 26889603
Tibon, r., Fuhrmann, D., Exacción, D. A., simons, j. S., & Henson, R. norte.
(2019). Multimodal integration and vividness in the angular gyrus
during episodic encoding and retrieval. Revista de neurociencia,
39(22), 4365–4374. https://doi.org/10.1523/ JNEUROSCI.2102
-18.2018, PubMed: 30902869
Tourville, j. A., Nieto-Castañón, A., Heyne, METRO., & Guenther, F. h.
(2019). Functional parcellation of the speech production cortex.
Journal of Speech, Idioma, and Hearing Research, 62(8S),
3055–3070. https://doi.org/10.1044/2019_JSLHR-S-CSMC7-18
-0442, PubMed: 31465713
Tourville, j. A., Reilly, k. J., & Guenther, F. h. (2008). Neural mech-
anisms underlying auditory feedback control of speech. Neuro-
Image, 39(3), 1429–1443. https://doi.org/10.1016/j.neuroimage
.2007.09.054, PubMed: 18035557
Velenosi, l. A., Wu, Y. h., Schmidt, t. T., & Blankenburg, F. (2020).
Intraparietal sulcus maintains working memory representations of
somatosensory categories in an adaptive, context-dependent
manner. NeuroImagen, 221, Article 117146. https://doi.org/10
.1016/j.neuroimage.2020.117146, PubMed: 32659356
Venezia, j. h., Vaden, k. I.,
Jr., Rong, F., Maddox, D., Saberi, K., &
Hickok, GRAMO. (2017). Auditory, visual and audiovisual speech pro-
cessing streams in superior temporal sulcus. Fronteras en lo humano
Neurociencia, 11, 174. https://doi.org/10.3389/fnhum.2017
.00174, PubMed: 28439236
ventura, METRO. I., Nagarajan, S. S., & Houde, j. F. (2009). Speech target
modulates speaking induced suppression in auditory cortex.
BMC Neuroscience, 10(1), Article 58. https://doi.org/10.1186
/1471-2202-10-58, PubMed: 19523234
Vingerhoets, A. J., Bylsma, l. METRO., & de Vlam, C. (2013). Swearing:
A biopsychosocial perspective. Psihologijske Teme, 22(2),
287–304.
Wolpert, D. METRO., Ghahramani, Z., & Jordán, METRO. I. (1995). un inter-
nal model for sensorimotor integration. Ciencia, 269(5232),
1880–1882. https://doi.org/10.1126/science.7569931, PubMed:
7569931
Wu, J., Ngo, GRAMO. h., Greve, D., li, J., Él, T., pescado, B., Eickoff, S. B.,
& yo, B. t. t. (2018). Accurate nonlinear mapping between MNI
volumetric and FreeSurfer surface coordinate systems. Humano
Mapeo cerebral, 39(9), 3793–3808. https://doi.org/10.1002/hbm
.24213, PubMed: 29770530
Cual, Z., Zhuang, X., Sreenivasan, K., Mishra, v., Cordes, D., &
Alzheimer’s Disease Neuroimaging Initiative. (2019). Robusto
motion regression of resting-state data using a convolutional neu-
ral network model. Frontiers in Neuroscience, 13, 169. https://doi
.org/10.3389/fnins.2019.00169, PubMed: 31057348
Yarkoni, T., Poldrack, R. A., Nichols, t. MI., VanEssen, D. C., &
Apostar, t. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8(8), 665–670.
https://doi.org/10.1038/nmeth.1635, PubMed: 21706013
Neurobiology of Language
119
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
8
1
2
0
7
4
4
9
4
norte
oh
_
a
_
0
0
0
8
8
pag
d
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3