ARTÍCULO DE INVESTIGACIÓN
Assessing the Sensitivity of EEG-Based
Frequency-Tagging as a Metric for
Statistical Learning
un acceso abierto
diario
Danna Pinto1
, Anat Prior2
, and Elana Zion Golumbic1
1The Leslie and Susan Gonda Multidisciplinary Brain Research Center, Bar Ilan University, Ramat Gan, Israel
2Department of Learning Disabilities, University of Haifa, Haifa, Israel
Palabras clave: auditory statistical learning, frequency-tagging, EEG
ABSTRACTO
Statistical learning (SL) is hypothesized to play an important role in language development.
Sin embargo, the measures typically used to assess SL, particularly at the level of individual
Participantes, are largely indirect and have low sensitivity. Recientemente, a neural metric based on
frequency-tagging has been proposed as an alternative measure for studying SL. We tested the
sensitivity of frequency-tagging measures for studying SL in individual participants in an
artificial language paradigm, using non-invasive electroencephalograph (EEG) recordings of
neural activity in humans. En tono rimbombante, we used carefully constructed controls to address
potential acoustic confounds of the frequency-tagging approach, and compared the sensitivity
of EEG-based metrics to both explicit and implicit behavioral tests of SL. Group-level results
confirm that frequency-tagging can provide a robust indication of SL for an artificial language,
above and beyond potential acoustic confounds. Sin embargo, this metric had very low sensitivity
a nivel de participantes individuales, with significant effects found only in 30% of participants.
Comparison of the neural metric to previously established behavioral measures for assessing
SL showed a significant yet weak correspondence with performance on an implicit task, cual
was above-chance in 70% of participants, but no correspondence with the more common
explicit 2-alternative forced-choice task, where performance did not exceed chance-level.
Given the proposed ubiquitous nature of SL, our results highlight some of the operational and
methodological challenges of obtaining robust metrics for assessing SL, as well as the potential
confounds that should be taken into account when using the frequency-tagging approach in
EEG studies.
INTRODUCCIÓN
Statistical learning (SL) refers to the remarkable ability to implicitly learn the rules and relation-
ship between different stimuli and events in the environment. The capacity for SL has been
studied in both humans and non-human species (Kang et al., 2021; Santolin et al., 2016;
Santolin & Saffran, 2018), and has been demonstrated across different sensory domains,
emerging relatively early in infancy (Gómez & Gerken, 2000; Graf Estes et al., 2007; Pelucchi
et al., 2009; Saffran et al., 1996, 1997). SL has been hypothesized to play an important role in
the development of many key cognitive abilities such as communication skills, object recog-
nition, and sensory-motor learning (Arciuli & von Koss Torkildsen, 2012; Emberson et al.,
2011; Erickson & Thiessen, 2015; Evans et al., 2009; Hsu et al., 2014; Kent & Leer, 2002;
Citación: Pinto, D., Previo, A., & Zion
Golumbic, mi. (2022). Assessing the
sensitivity of EEG-based frequency-
tagging as a metric for statistical
aprendiendo. Neurobiology of Language,
3(2), 214–234. https://doi.org/10.1162
/nol_a_00061
DOI:
https://doi.org/10.1162/nol_a_00061
Supporting Information:
https://doi.org/10.1162/nol_a_00061
Recibió: 11 Junio 2021
Aceptado: 10 Noviembre 2021
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Elana Zion Golumbic
elana.zion-golumbic@biu.ac.il
Editor de manejo:
Sonja A. Kotz
Derechos de autor: © 2022
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Transitional probability:
The statistical relationship between
two consecutive events.
Pseudowords:
Combinations of syllables that are
not lexical entities and do not carry
semantic meaning in a particular
idioma.
Frequency-tagging:
A method by which specific features
of a stimulus are presented at a
particular frequency, which allows
discerning a neural-signature of this
feature in the spectrum of the neural
datos.
Kidd, 2012; Kidd & Arciuli, 2016; Misyak & Christiansen, 2012; Siegelman et al., 2017;
Spencer et al., 2015; Thiessen & Saffran, 2003, 2007). Y todavía, despite the potentially pivotal
role of SL for cognition, current empirical metrics used to assess SL, particularly at the level
of individuals, are largely indirect, and often have low sensitivity.
In typical SL experiments a sequence of stimuli is presented in which the transitional prob-
abilities between consecutive stimuli are manipulated so that some items carry predictive
information about which stimulus will follow. One prominent example is the artificial
language paradigm, where participants hear sequences of syllable triplets that are always pre-
sented consecutively (transitional probability = 1) and thus form words in an artificial language
(which we refer to throughout this paper as pseudowords). Participants are exposed to these
stimuli for a short period of time (exposure phase), which can range between 2 y 24 mín.
(Batterink et al., 2015; Franco, Gaillard, et al., 2015; Karuza et al., 2013; Saffran et al., 1997),
and then perform a test to assess whether the statistical regularities within the sequence have
been picked up by the listener. A variety of explicit and implicit tests can be applied to eval-
uate SL following an exposure phase, such as a 2-alternative forced-choice test (2AFC) o un
target-detection task (Batterink, 2017; Batterink & Paller, 2017; Batterink et al., 2015). Behav-
ioral results on these tests usually show moderate yet above-chance performance when ana-
lyzed at the group-level. Por ejemplo, performance on 2AFC tasks ranges between 54% y
68% across studies, which constitutes a significant yet fairly weak demonstration of learning
(Batterink & Paller, 2017; Batterink et al., 2015; Buiatti et al., 2009; de Diego-Balaguer et al.,
2015; Fernandes et al., 2010; Franco et al., 2011; Franco, Gaillard, et al., 2015; Frost et al.,
2015; Kim y cols., 2009; Olson & Chun, 2001; Saffran et al., 1997; Siegelman & Frost, 2015;
Toro et al., 2005; Turk-Browne et al., 2005; tyler & Cutler, 2009). Sin embargo, success rates of
individual participants are rarely reported, and the few studies that do include this data find that
al menos 30% of the participants show no evidence for SL at all and in many individuals behav-
ioral effects are quite small (Cunillera et al., 2008; Franco, Gaillard, et al., 2015; Romberg &
Saffran, 2013). It is also worth noting that the within-subject correlation between different
behavioral tasks (p.ej., explicit vs. implicit tests) is often low, raising questions about the optimal
experimental operationalization for capturing and assessing SL (Batterink et al., 2015; Franco,
Eberlen, et al., 2015; Misyak et al., 2010). Given the hypothesized fundamental role of SL for a
variety of cognitive processes (Arciuli, 2017; Erickson & Thiessen, 2015), it seems pertinent to
develop a more robust empirical measure of SL, that can reliably assess whether or not SL has
occurred at the level of individual subjects.
Rather than relying on post-exposure behavioral testing for assessing SL, an alternative
approach is to analyze participants’ neural activity during the exposure phase and look for
evidence that statistical regularities within the stimulus are being picked up. Along these lines,
an EEG-based frequency-tagging approach has recently been proposed using a variation of the
artificial language paradigm (Batterink, 2020; Batterink & Paller, 2017, 2019; Buiatti et al.,
2009; Choi et al., 2020; Elmer et al., 2021; Getz et al., 2018; Henin et al., 2021; Kiai &
Melloni, 2021; Lukics & Lukács, 2021). In this version, syllables are presented at a constant
tasa (p.ej., X Hz), and consequently the tri-syllabic pseudowords also occur at a fixed rate
(X/3 Hz). These two levels of information are thus distinguishable in frequency, which can
potentially be observed in the spectrum of the EEG neural recording. This frequency-tagging
approach has been successfully employed for studying real speech processing, demonstrating
that a peak at the word-level frequency emerges in the spectrum of the neural response when
syllables make up words that participants know, but not if they are in a foreign language or do
not form recognizable words or phrases (Ding et al., 2016; Lu et al., 2021; luo & Ding, 2020;
Makov et al., 2017; Sheng et al., 2019). Applying this approach to a SL paradigm, Batterink
Neurobiology of Language
215
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Obligatory contour principle (OCP):
A hypothesis in phonology which
states that certain phonetic features
may not occur consecutively.
and Paller (2017) demonstrated that the ratio between the power at the syllable vs. pseudo-
word frequency during the exposure-phase was positively correlated with behavioral perfor-
mance on an implicit (but not an explicit) behavioral task for assessing SL. This was taken as an
indication for the adeptness (and perhaps advantage) of using frequency-tagging to assess SL
experimentally, circumventing the need for overt post-exposure behavioral testing.
Sin embargo, despite the promise held by this approach as providing a more direct and objec-
tive measure of SL, some of the previous findings raise questions regarding the sensitivity of
this measure, particularly at the level of individual subjects. Por ejemplo, the individual-level
data presented by Batterink and Paller (2017) indicate that SL effects were limited only to a
subset of participants, with others showing effects in the opposite direction. Además, in that
study significant effects were also reported when participants listened to random sequences of
syllables, where there should not be any SL. As suggested by recent studies, these results may
have been somewhat confounded by acoustic contributions to the neural response at the pseu-
doword frequency that occur naturally for this type of stimuli (luo & Ding, 2020; van der
Wulp, 2021). En particular, in a recent re-analysis of the EEG data originally reported by
Batterink and Paller (2017), van der Wulp (2021) demonstrated that at least some of the
reported effects can be explained by variations in place of articulation of different syllables
(known as the obligatory contour principle; OCP), rather than by SL of transitional probabilities
between syllables. Como consecuencia, without proper controls, the magnitude of the neural
response at the pseudoword frequency might be over-interpreted as only reflecting SL, mientras
the acoustic contribution to this peak is discounted or ignored.
Por lo tanto, it seems that further validation of the frequency-tagging approach is required,
and adequate controls implemented, before adopting it as a demonstrably preferable measure
of SL. This is an important endeavor not only for furthering our understanding of the potential
para, and possible limitations of, frequency-tagging for studying SL in humans, but also for asses-
sing its potential sensitivity for use in clinical conditions (p.ej., non-consciousness states; Gui
et al., 2020; Sokoliuk et al., 2021) as well as in non-human species, where data analysis typ-
ically relies on within-subject effects and not on group-effects.
MATERIALES Y MÉTODOS
Participantes
Los participantes fueron 40 adultos (25 femenino, 35 right-handed), ages 20–38 (mean = 24.78, DE =
3.96). Due to technical issues, EEG data from one participant and behavioral data on the
implicit test from 13 participants were lost. All participants reported normal hearing and
had no history of psychiatric or neurological disorders and were native Hebrew speakers. Ellos
were paid or received course credit for participation. The study was approved by the IRB com-
mittee at Bar Ilan University and participants read and signed an informed consent form prior
to starting the experiment.
EEG Recording and Apparatus
EEG was recorded using a 64 Active-Two system (BioSemi) with Ag-AgCl electrodes, placed
according to the 10–20 system, at a sampling rate of 1024 Hz. Additional external electrodes
were used to record from the mastoids bilaterally, and both vertical and horizontal electrooc-
ulography electrodes were used to monitor eye movements. The experiment was conducted in
a dimly lit, acoustically and electrically shielded booth. Participants were seated on a com-
fortable chair and were instructed to keep as still as possible and breathe and blink naturally.
Neurobiology of Language
216
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Modulation spectrum:
The frequency content of the
envelope of a particular signal (para
ejemplo, of the speech-envelope).
Experiments were programmed and presented to participants using PsychoPy (https://
www.psychopy.org; Peirce et al., 2019). Visual instructions were presented on a computer
monitor, and auditory stimuli were delivered through in-ear earphones (Etymotic ER-1).
Button-press responses were recorded using a serial response-box (Cedrus RB).
Estímulos
The stimuli consisted of 18 CV syllables recorded in a male voice. Individual syllables were
recorded in random order to avoid effects of coarticulation, and only recordings with a flat
intonation were used. The recordings were edited offline so that each syllable was precisely
250 ms long (silence periods were added if necessary), and their loudness was equated
(Audacity software; https://www.audacityteam.org/). Additional audio-editing and concatena-
tion of syllables into longer streams were performed in Matlab (Matemáticas; https://www
.mathworks.com/). The artificial language consisted of six tri-syllabic pseudowords (PaShuDi,
SoGuMa, NoMuBe, TuBiPo, GeRoVa, KaLeVi), with each syllable appearing in only one
pseudoword. Respectivamente, the within-word transitional probability was 1 and the between-
words transitional probability was 0.2. Given that the modulation spectra of this type of stim-
uli naturally contains acoustic-driven peaks and frequencies besides the syllable rate itself
(luo & Ding, 2020; van der Wulp, 2021), we tested the modulation spectrum of several
syllable-triplet combinations and selected the combination that yielded the smallest peaks at
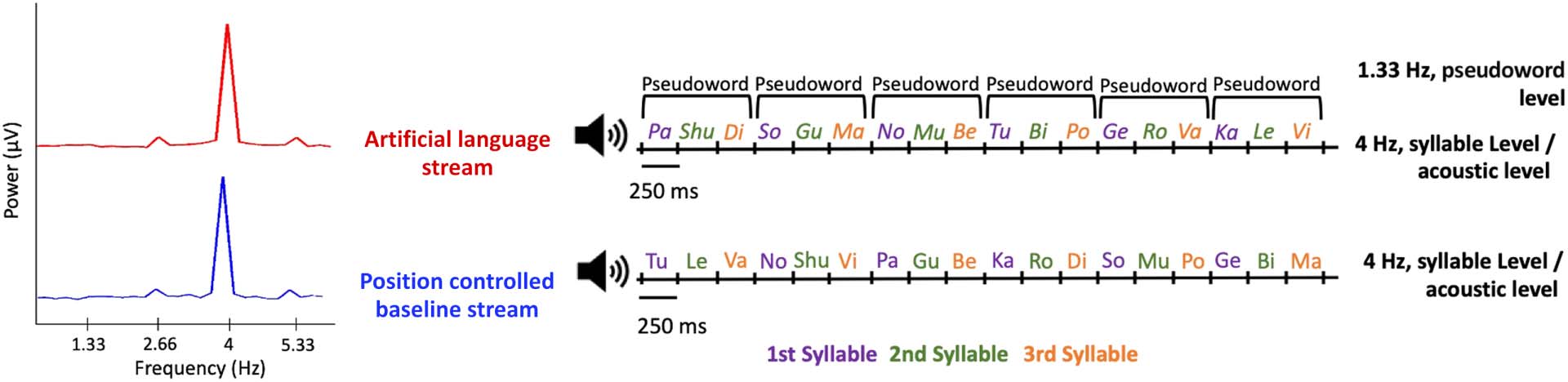
the pseudoword rate and/or its harmonics as the pseudowords in this experiment (Cifra 1).
We also confirmed that the pseudowords do not sound similar to known Hebrew or English
palabras.
Since the acoustic-driven contributions to the modulations spectrum at the triplet-related
frequencies could not be fully eliminated from the artificial language stimulus, we constructed
a position-controlled baseline stimulus to estimate the extent of these acoustic contributions to
the neural signal. The baseline stimulus consisted of syllable triplets constructed from the same
18 syllables, but with less consistent transitional probabilities between them. Similar to the
approach used by Makov et al. (2017), in these position-controlled syllable triplets each syl-
lable maintained the position it held in the original pseudowords; sin embargo, all possible com-
binations were allowed (Cifra 2, bien). This yielded a constant transitional probability of 0.2
both within-triplet and between-triplets in the baseline stimulus.
Both the pseudowords and the position-controlled triplets were concatenated to create
three 3.22-min-long streams of the artificial language and baseline conditions. The order of
pseudowords and position-controlled triplets in each stream was pseudorandomized to avoid
immediate repetitions of the same triplet and ensure their equal distribution over time. Com-
parison of the modulation spectra confirmed that this approach resulted in similar peaks at
1.33 Hz for both the artificial language and baseline streams, making them highly comparable
acoustically and allowing us to gauge the effect of within-word transitional probabilities on the
1.33 Hz peak in the neural response, above and beyond any potential acoustic contributions
from the stimulus itself (Cifra 2, izquierda). Both the artificial language and baseline streams
included a 5 sec ramping up/down period, to avoid inadvertent cues about syllable positions
or pseudoword boundaries.
Experimental Procedure
Exposure phase
The experiment consisted of several stages. It started with a baseline exposure stage during
which participants listened to the baseline condition streams of concatenated syllables
Neurobiology of Language
217
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Cifra 1. Modulation spectrum for four different versions of the artificial language stimuli, all com-
posed of similar syllables but combined to form different pseudowords. As expected, all stimuli con-
tain a peak at the 4-Hz syllable rate. Sin embargo, as shown here, additional peaks are observed at the
pseudoword rate (1.3 Hz) and its harmonics, and the magnitude of these peaks varies for the different
combinations. As shown in similar studies (Har-shai Yahav & Zion Golumbic, 2021; luo & Ding,
2020; van der Wulp, 2021), these peaks stem from the fact that the same subset of syllables is present
in constant positions within the stimulus streams. The artificial language stimuli chosen in the current
experiment was a combination of syllables that generated relatively small peaks at pseudoword rate
frequencies and its harmonics in the modulation spectrum; sin embargo, these were nonetheless still
present (blue line). This motivated the use of position-controlled stimuli as a means to control for
these inherent acoustic peaks, which has a modulation spectrum similar to the artificial language
estímulos. This allowed us to attribute significant differences in the neural response between these
two stimuli to effects of statistical learning, rather than trivial differences in their acoustic structure.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. Diagram illustrating structure of the artificial language and position-controlled baseline streams used in the current experiment. Left:
The modulation spectrum of the artificial language stream (rojo) and the position-controlled baseline stream (azul). Both show a prominent
peak at the syllable rate (4 Hz), as well as more modest peaks at 2.66 Hz and 5.33 Hz, which are the first and third harmonics of the triplet rate.
Right: Examples of the auditory streams. Both stimuli were composed of the same syllables, presented at a constant rate of 4 Hz. Each stream
consisted of syllable triplets, with each syllable consistently either at the 1st (azul), 2nd (verde), or 3rd (naranja) posición. In the artificial lan-
guage stream, fixed syllable triplets were used in forming pseudowords (within-pseudoword transitional probability = 1; between-pseudoword
transitional probability = 0.2), whereas in the position-controlled baseline stream all possible triplet combinations were used, resulting in a
consistent transitional probability of 0.2 between all syllables.
Neurobiology of Language
218
Frequency-tagging in EEG-based statistical learning research
described above. These consisted of hearing three 3.22-min-long streams (separate blocks,
with breaks between them; total exposure time: ∼10 min). Participants were instructed to listen
passively to the stimuli with their eyes open and fixated on a point on the screen. In this stage
no additional instructions were given. After a brief break they were exposed to the three blocks
of the artificial language streams. Here participants were explicitly told that the streams are
made up of words in an artificial language, which they are requested to learn for subsequent
pruebas. Sin embargo, participants were not told the length or number of the pseudowords. El
order between exposure phases was held constant to avoid carryover learning effects in the
baseline condition after exposure to the artificial language.
During the break between the baseline and artificial language conditions, participants per-
formed an English vocabulary task. This task was chosen as a way to clear their verbal working
memory and also in order to test the hypothesis that statistical learning abilities are correlated
with second language learning abilities (since all our participants learned English as a second
language in school). Desafortunadamente, the results of the vocabulary test from almost half of the
participants were lost due to technical difficulties, which did not allow us to further explore
this research question in the current study.
Testing phase
The testing phase consisted of two behavioral tests:
2-Alternative forced choice task (2AFC). The explicit 2AFC discrimination task was designed to
follow the commonly used procedure for explicit testing of statistical learning (Batterink &
Paller, 2017; Batterink et al., 2015; Buiatti et al., 2009; Fernandes et al., 2010; Franco
et al., 2011; Franco, Gaillard, et al., 2015; Saffran et al., 1997; Toro et al., 2005, 2011; tyler
& Cutler, 2009; Wang & Saffran, 2014). In addition to the six pseudowords that made up the
artificial language, six additional part-words were created consisting either of the last two syl-
lables of one pseudoword combined with the first syllable of another, or the last syllable of one
pseudoword combined with the first two syllables of another. Tal como, these are combinations
that participants could have heard occasionally during the learning phase, but not as fre-
quently as the actual pseudowords. En cada prueba, one pseudoword and one part-word were
played (random order), and participants were required to indicate via button-press which
one was familiar to them from the artificial language learning phase. This test consisted of a
total of 36 ensayos (all possible pairs of pseudowords and part-words).
Group-level statistical analysis of performance on the 2AFC task consisted of a single-
sample t test testing whether accuracy rates were significantly higher than chance (50%),
as commonly done in similar studies (Batterink & Paller, 2017; Batterink et al., 2015; Buiatti
et al., 2009; Fernandes et al., 2010; Franco et al., 2011; Franco, Gaillard, et al., 2015; Saffran
et al., 1997; Toro et al., 2005, 2011; tyler & Cutler, 2009; Wang & Saffran, 2014). Sin embargo,
since the 2AFC task consists of only 36 trials and does not necessarily meet the assumptions
required for a t test, we further simulated the null distribution of our specific experiment
using a permutation test. We simulated a random 2AFC guessing pattern for 36 trials and
calculated the “random hit rate” of that simulation. This procedure was repeated 1,000
veces, producing a null distribution reflecting the probability of achieving a particular hit
rate “by chance” (Figura 3A, shown in gray). Además, we assessed the significance of
performance in individual participants by comparing their accuracy rates to a binomial dis-
tribution for 36 2AFC trials (Franco, Gaillard, et al., 2015; Siegelman et al., 2017), permitiendo
us to establish which participants showed evidence for statistical learning according to the
2AFC test.
Neurobiology of Language
219
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Cifra 3. Behavioral results. (A) 2AFC results: Histogram of accuracy rates on the 2AFC task across all participants (negro), overlaid on the
background of the a priori binomial distribution of chance-level results in the current design (gray). Top: Interquartile range and group median
of 2AFC results. Dashed gray line: the p = 0.05 cutoff for determining whether individual level performance was significantly above chance
(relative to the null distribution). (B&C) Target detection results: Interquartile range and group-level median for hit rates and reaction times (RTs)
in response to target syllables that occurred in the 3rd position of pseudowords vs. targets that occurred in non-words. For both metrics,
performance was improved for targets within pseudowords, as indicated by the asterisks ( pag < 0.001) between the conditions. Outliers are
indicated by a gray plus (+) symbol. (D&E) Scatterplots depicting the within-subject relationship between performance on the 2AFC task
and the target detection task. No significant correlation was found between accuracy on the 2AFC task and the magnitude of the behavioral
effects in the target detection task (difference in hit rate / RTs for targets in pseudowords vs. targets in non-words).
Target detection task. The explicit test was followed by an implicit target detection task,
designed based on previous studies using a similar approach (Batterink et al., 2015; Batterink
& Paller, 2017). In each trial one syllable was designated as the target and was played twice for
participants to familiarize themselves with the sound (e.g., Va). Then a sequence of syllables
was played, and participants were required to press a button when they heard the target syl-
lable. The sequences contained pseudowords from the artificial language as well as other
triplet-syllable combinations (non-words). The target syllable in each trial (e.g., Va) was placed
strategically within the sequences and could occur either as the 3rd syllable of a pseudoword
presented in the exposure phase (e.g., GeRoVa) or as the 1st or 3rd syllable in a non-word (e.g.,
VaShuPo or PaMuVa). In this task, enhanced target detection performance for targets presented
as the 3rd syllable of a previously learned pseudoword (vs. syllables in a non-word) would
serve as an indication that participants had successfully learned the structure of the artificial
language because they are able to anticipate the target syllable.
For this task, syllables were presented at a constant rate of 2 Hz with each trial lasting
22.5 sec and including 4–8 targets. The entire task consisted of 24 trials (4 trials per target
syllable). A button-press was considered a hit if it fell within 1 sec after the presentation of
a target syllable. Otherwise, it was considered a false alarm. The order of the explicit 2AFC
task and the implicit target detection task was kept constant and not randomized across par-
ticipants. Since the 2AFC task is the more common test for SL, we felt it was important to
administer it immediately after the exposure phase, and to avoid its potential contamination
by exposure to additional syllable sequences in the implicit task.
Neurobiology of Language
220
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Group-level statistical analysis consisted of paired t tests of hit rates and reaction times (RTs)
for targets occurring within pseudowords vs. non-words (responses to targets occurring in 1st
and 3rd position of non-words were grouped together, since we found no differences between
them). Statistical analysis at the level of individual participants was conducted using permu-
tation tests. The permutation test consisted of random relabeling of all the responses of a
particular participant into two random conditions, regardless of their original status as
pseudoword/non-word targets, and taking the difference between the means of the two ran-
dom conditions. This procedure was repeated 1,000 times, and the differences of the means
extracted from each permutation were used to form a null distribution for each participant. We
then took the real difference between the pseudoword and non-word targets in the original
data and compared it to the null distribution. The difference between conditions was consid-
ered significant if the real value fell in the top fifth percentile of the null distribution (one-
tailed). This procedure was performed for both accuracy and RT data.
We further tested whether performance on the two behavioral tasks was correlated, by
calculating the Pearson correlations between explicit 2AFC accuracy rates and the implicit
target detection task (differences in hit rates / RTs for targets occurring within pseudowords
vs. non-words).
EEG Data Analysis
Preprocessing and spectral analysis
EEG data were measured only during the exposure phase of the experiment, and were not
measured during the testing phase. Data from three blocks (∼11 min) of both conditions were
preprocessed and cleaned together. All EEG preprocessing and analysis were performed in
Matlab (The Mathworks) using the toolbox FieldTrip (Oostenveld et al., 2011) as well as cus-
tom written scripts. Raw data were first visually inspected and gross artifacts that exceeded
±50 μV (and were not eye movements) were removed. Then, independent component analysis
was performed to identify and remove components associated with horizonal or vertical eye
movements as well as heartbeats. Any additional noisy electrodes / segments of the data that
remained after this procedure, and that exhibited either extreme high-frequency activity (>40 Hz)
or low-frequency activity/drifts (<1 Hz), were either replaced with the weighted average of
their neighbors using an interpolation procedure, or (if that was not possible) removed.
The clean data analyzed separately for baseline and artificial language exposure
blocks. The continuous segmented into 4.5 sec epochs, which correspond to 6 syl-
lable triplets. Critically, these segments perfectly aligned such they all started with
the onset of a triplet. Inter-trial phase coherence (ITPC) used analyze neural
response at specific frequencies. ITPC calculated as follows: fast Fourier transform
was each individual segment between 0.3 Hz Hanning window.
The component frequency calculate ITPC, is sum
(absolute value) phases across segments, follows:
ITPC ¼
(cid:2)
(cid:2)
(cid:2)
(cid:2)
(cid:2)
1
N
XN
k¼1
(cid:2)
(cid:2)
(cid:2)
(cid:2)
(cid:2)
ei(cid:2)(cid:1)
k
ITPC analysis performed exposure
blocks blocks well three per condition.
Inter-trial (ITPC):
A measure consistency
across trials.
Neurobiology Language
221
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
> base), but neither the effects of block nor the interaction between condition and block were significant at any of the
frecuencias.
Neurobiology of Language
224
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
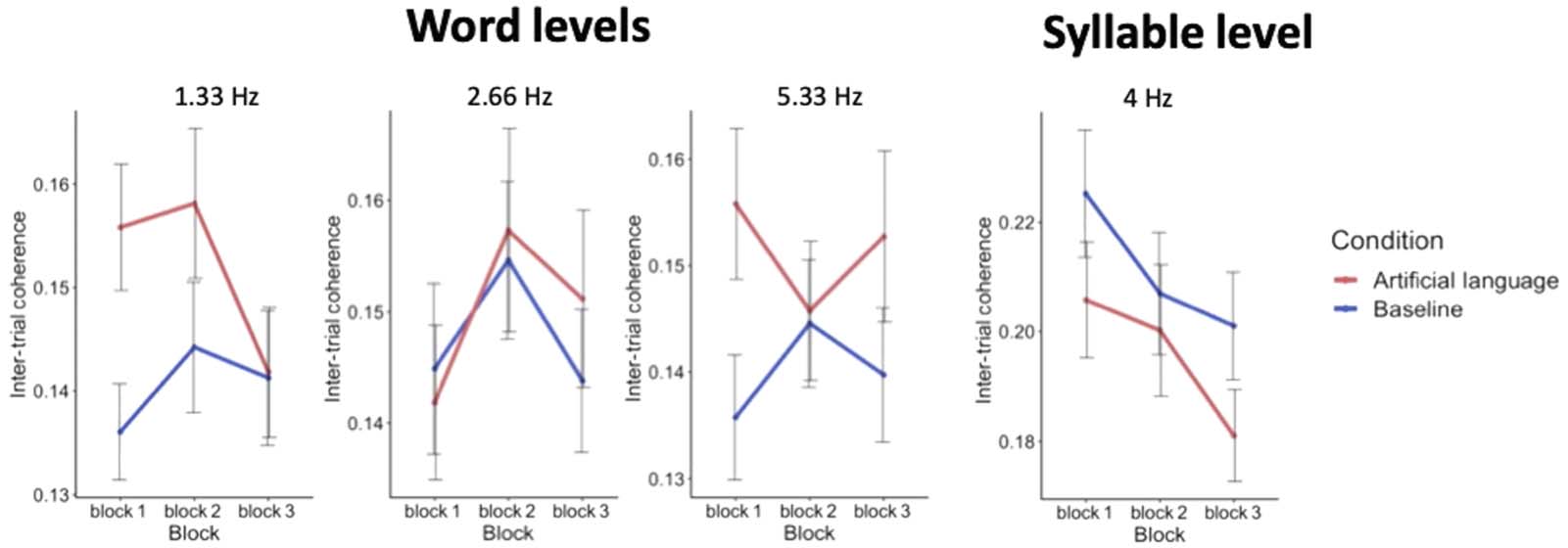
Mesa 1.
Summary of statistical results of block analysis. Significant results are indicated in bold and with an asterisk
Block number (1 vs. 2)
Contrast
Block number (1 & 2 vs. 3)
Condition (artificial language vs. base)
Interaction Block number (1 vs. 2) × Condition
Interaction Block number (1 & 2 vs. 3) × Condition
Frecuencia
Word
Syllable
1st Harmonic
3rd Harmonic
Word
Syllable
1st Harmonic
3rd Harmonic
Word
Syllable
1st Harmonic
3rd Harmonic
Word
Syllable
1st Harmonic
3rd Harmonic
Word
Syllable
1st Harmonic
3rd Harmonic
b
−0,007
0.02
−0,004
−0,006
0.003
0.006
0.01
0.005
0.11
−0,02
0.002
0.01
0.01
−0,006
0.008
0.01
0.01
0.01
−0.005
−0.01
t
−0.88
1.98
−0.49
−0.78
0.34
0.48
1.06
0.51
2.25
−2.15
0.40
2.10
1.163
−0.40
−0.64
1.12
1.07
0.76
−0.32
−0.88
pag
0.38
0.05
0.63
0.43
0.74
0.63
0.29
0.61
0.03*
0.03*
0.69
0.04*
0.25
0.69
0.52
0.26
0.28
0.45
0.75
0.38
1.33 Hz and 5.33 Hz, were confirmed in this analysis as well [F(190) = 2.25, pag = 0.03, y
F(228) = 2.12, pag = 0.03 respectivamente]. Sin embargo, the effect of block and interactions between
condition and block were not significant at any of the frequencies of interest (ver tabla 1 para
full statistical results). Bastante, all the effects of condition seem to be present already in the first
exposure block and were not further enhanced over time.
Individual participant analysis
Assessment of SL effects from the neural response at the level of individual participants was
conducted using permutation tests. Since SL effects could potentially manifest either at the
pseudoword rate itself or at any of its harmonics, this analysis was performed at all the frequen-
cies of interest. When performing the statistical analysis on the average ITPC across all elec-
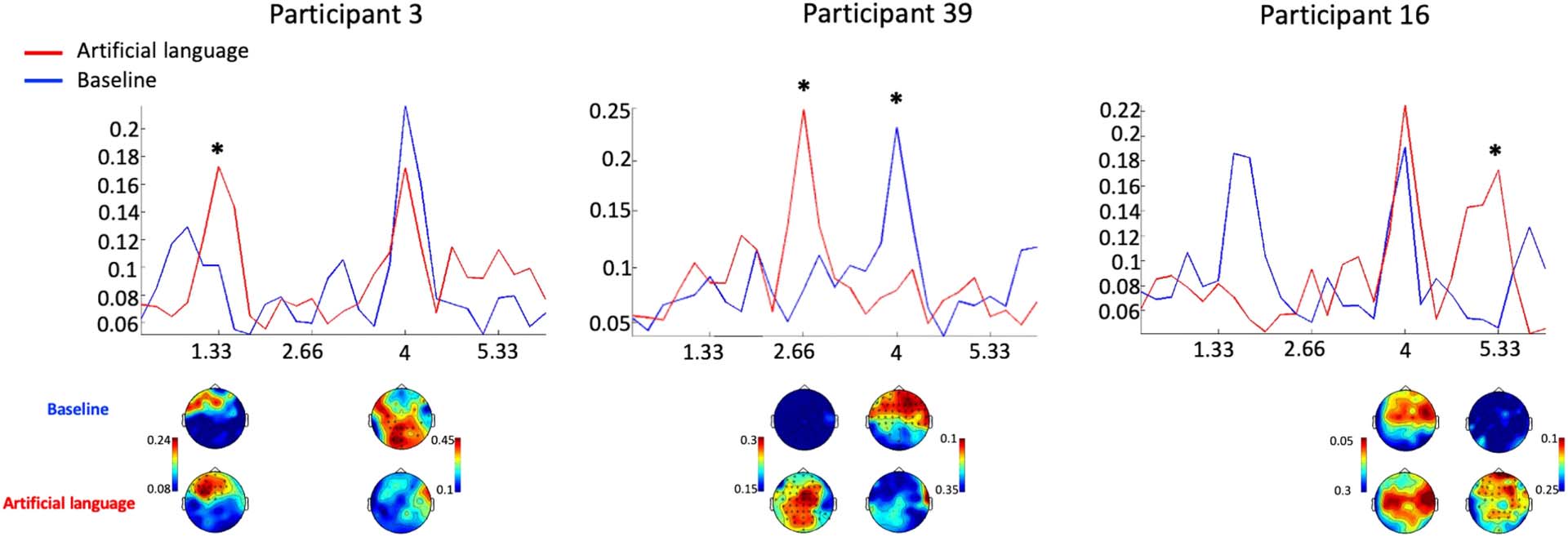
trodes, we found significant effects of condition in 12/39 Participantes (31%), with larger
responses in the artificial language condition vs. the baseline. Of these participants, in n = 5
the effects were at 1.33 Hz, norte = 3 en 2.66 Hz, and in n = 4 en 5.33 Hz. Only one participant had
significant effects at more than one frequency. Además, the reduced response in the artificial
Neurobiology of Language
225
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Cifra 6. Examples of ITPC spectral of individual participants. ITPC spectra in the artificial language (rojo) and baseline (azul) conditions from
three participants who showed significant differences between the artificial language and baseline conditions, albeit at different pseudoword-
related frequencies (indicated by an asterisk). Spectra from each participant are shown from the average across all electrodes.
language condition at the syllable rate (4 Hz) that was observed at the group level was found to
be significant for n = 6 Participantes. For specific examples of the ITPC spectrum of individual
participants who showed significant results see Figure 6. When repeating the analysis using
only the electrodes that had significant SL effects at the group level (como se muestra en la figura 4), nosotros
found similar results: 13/39 (33%) participants had a significant effect of condition, with larger
responses for the artificial language condition than the baseline. Of them, in n = 9 the effects
were at 1.33 Hz, in n = 3 en 2.66 Hz, and in n = 5 en 5.33 Hz. Three participants had significant
effects at more than one frequency. También, the reduced response in the artificial language con-
dition at 4 Hz was found to be significant in n = 7 Participantes. We note that the latter analysis is
a little circular (selection of electrodes based on a previous group-level result). Sin embargo, el
convergence of results in these two analyses (one more conservative, one more permissive)
supports the overall conclusion of a low prevalence of SL effects in individual participants.
Correspondence between ITPC effects and behavior
We next tested whether the ITPC response to pseudoword rates in the artificial language con-
dition corresponds to performance on behavioral tasks administered post-exposure. Desde el
individual level analysis revealed inconsistencies in the specific pseudoword-related frequen-
cies where significant differences were found across participants (es decir., at the pseudoword fre-
quency itself or one of the harmonics), this prevented us from performing a simple correlation
analysis between the ITPC at a particular frequency and behavioral measures. To overcome
this between-participant variability, we used two different approaches.
Primero, we took the average ITPC in the artificial language condition across the three
pseudoword-related frequencies (1.33 Hz, 2.66 Hz, y 5.33 Hz), and calculated the Pearson
correlations with each behavioral measure. This did not, sin embargo, yield any significant results
(correlation with 2AFC accuracy: r2 = 0.16, pag = 0.33; correlation with target detection hit rate:
r2 = 0.13, pag = 0.50; correlation with target detection RTs: r2 = −0.05, pag = 0.81).
Segundo, we separated the participants into two groups based on whether there was evi-
dence for SL from their neural data (regardless of the frequency where this effect was observed)
and compared the behavioral results between the two groups. We used the Welch’s test for
unequal variance to account for the different sample sizes in the two groups. In this analysis
we found that the group in which significant pseudoword EEG responses were observed also
Neurobiology of Language
226
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
had significantly larger behavioral effects in the target detection task (Cifra 7, bottom panel).
Específicamente, this group had larger differences in RTs between targets occurring in the 3rd
position of pseudowords vs. targets within non-words [t(14) = 2.15, pag = 0.03; BF10 = 4.18
(moderate support)]. Sin embargo, this effect was not significant for hit rates in the target detection
tarea [t (10) = −1.02, pag = 0.17] (Cifra 7, middle panel) or for performance on the 2AFC task
[t (21) = −0.19, pag = 0.85] (Cifra 7, top panel).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
norte
oh
_
a
_
0
0
0
6
1
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 7. Correspondence between and pseudoword EEG response during the exposure period and post-exposure behavioral tasks. Partic-
ipants were divided into two groups based on whether a significant response to pseudowords was found in their neural response during the
exposure period (gray) or not (negro). The left-hand panels show the results of all participants on each of the three behavioral measures: 2AFC
exactitud, effects size on hit rate, and RTs in the target detection task (sorted by effect size and color-coded by group). The right-hand panels
show the means and SEM of these behavioral measures, in each group. A significant difference between-groups was found for the target
detection RT effect (bottom panel, pag < 0.05), but not for the other behavioral measures.
Neurobiology of Language
227
Frequency-tagging in EEG-based statistical learning research
DISCUSSION
In this study, we tested the sensitivity of the EEG frequency-tagging approach as an online
measure for assessing auditory statistical learning of an artificial language, at both the group
level and within individual participants. We found that, even after controlling for potential
acoustic contributions to the pseudoword frequency, there is still a significant difference
between the artificial language and the baseline conditions at the group level. This effect man-
ifested most robustly at the 3rd pseudoword-level harmonic (5.33 Hz), and less reliably at the
pseudoword-level rate itself (1.33 Hz). The previously reported decrease at the 4-Hz syllable
level for artificial language stimuli was also observed here, but again with low statistical reli-
ability. Effects were observed already during the first exposure block, (3.22 min) and did not
change significantly with additional exposure. These results help validate the use of the
frequency-tagging approach for assessing SL at the group level, while highlighting important
considerations for implementing this technique in future studies.
However, at the level of individual participants, only 30% showed significant effects of SL
in their neural response, and among them the effects did not occur consistently at the same
frequencies/harmonics. Conversely, performance on the implicit target detection task admin-
istered post-exposure demonstrates that SL occurred in a substantially larger proportion of
individuals (70%). Hence, the current results suggest that the EEG-based metric has a lower
sensitivity than some implicit behavioral metrics and likely underestimates the prevalence of
SL in individual participants.
Strengths and Weaknesses of the Frequency-Tagging Approach for Assessing SL
The EEG frequency-tagging approach has been proposed as a more direct means for assessing
SL, circumventing the need for behavioral post-exposure testing. Among its strengths is its
online nature, which allows researchers to track the formation of a neural representation for
pseudowords over time, without introducing a dual task. This approach has been successfully
applied for studying neural processing of familiar and unfamiliar languages, and how the rep-
resentation of different linguistic levels of speech is modulated by factors such as attention,
state of arousal, and consciousness (Chen et al., 2020; Ding et al., 2016; Getz et al., 2018;
Har-shai Yahav & Zion Golumbic, 2021; Luo & Ding, 2020; Makov et al., 2017; Niesen et al.,
2019). The frequency-tagging approach has also brought great excitement to the field of sta-
tistical learning, since it offers a way to dissociate between the acoustic-representation of indi-
vidual elements in a stream (e.g., syllable rate; 4 Hz in the current study) and its parsing into
larger units (e.g., pseudoword rate; 1.33 Hz and its harmonics in the current study) that reflects
higher-level generalization and learning (Batterink & Paller, 2017, 2019; Buiatti et al., 2009;
Elmer et al., 2021; Getz et al., 2018; Henin et al., 2021).
However, here it is crucial to note an important methodological caveat: The interpretation
that peaks in the neural response at the pseudoword rate reflect detection and parsing of pseu-
dowords relies on the assumption that these peaks cannot be derived from the acoustics of the
stimulus alone. Unfortunately, this assumption does not seem to hold for the type of stimuli
typically used in the triplet-based artificial language SL paradigm. As shown in the modulation
spectra when testing several different combinations of triplet syllables (Figure 1), a prominent
peak can be seen at the triplet rate in addition to the syllable rate. This peak is generated due to
subtle yet systematic differences in the envelope shape of different syllables, which are pre-
sented consistently at the same position—an inherent feature of pseudowords. These caveats
of the frequency-tagging approach have recently been pointed out when using bisyllabic
words in real languages (Har-shai Yahav & Zion Golumbic, 2021; Luo & Ding, 2020).
Neurobiology of Language
228
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Similarly, for artificial languages, the elegant re-analysis of the data in Batterink and Paller
(2017) showed that at least part of the neural response at the triplet-rate frequency can be
attributed to differences in the OCP of different syllables rather than SL per se (van der Wulp,
2021). Therefore, in order to avoid overinterpretation of these peaks, adequate controls must
be implemented in all studies.
Here we addressed this concern by introducing a position-controlled baseline stimulus,
which shared the same modulation spectrum as the artificial language stimulus. As expected,
in addition to the neural response at the syllable rate, the response to this position-controlled
stimulus contained a prominent peak at the triplet rate and its harmonics, even though it
contained no statistical regularities. This demonstrates the methodological caveat of
frequency-tagging mentioned above—that the mere existence of a triplet-rate peak is not, in
and of itself, an indication of statistical learning. Nonetheless, when comparing the neural
response to the two stimuli at the group level, the triplet-rate peak (and its harmonics)
was significantly larger in response to the artificial language stream relative to its position-
controlled baseline stimulus. This pattern suggests that the neural response at the triplet rate
and its harmonics reflects a combination of acoustic responses as well as responses reflecting
detection of the underlying statistical structure and/or pseudoword boundaries.
Interestingly, the strongest effect was not found at 1.33 Hz, which is the triplet rate itself, but
rather at its 3rd harmonic (5.33 Hz). This is similar to the pattern reported by a recent electro-
corticography (ECoG) study, where the most prominent effects of SL were also found at har-
monics of the triplet rate (Henin et al., 2021). Moreover, as detailed below, when inspecting
the individual-level spectra, we found great variability in which frequencies showed the most
prominent SL effects. The manifestation of effects at harmonic frequencies is a natural conse-
quence of presenting rhythmic stimuli, and should not necessarily be interpreted as carrying
nuanced information regarding the nature of neural encoding for these stimuli (Zhou et al.,
2016). However, this variability does present another potential caveat for the utility of the
frequency-tagging approach.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Assessing SL in Individual Participants
One of the main goals of the current study was to investigate the sensitivity of different mea-
sures of SL at the level of individual participants. Due to the proposed ubiquitous nature of SL
and its proposed importance for language acquisition, we expected to find evidence for SL in
most participants. However, this was not case. Rather, the pattern emerging from comparing
the three independent measures used here—the explicit 2AFC, the implicit target detection
task, and the frequency-tagged EEG spectrum—illustrates the operational challenge of empir-
ical assessment of SL. The 2AFC test failed to show a significant effect at the group level, and
at the individual level only 3 participants (7.5%) showed significant effects. These poor per-
formance levels are in line with previous studies where reported group-level detection rates
range between 54% and 74%, and individual-level significance rates are low (fewer than
50% of participants; Franco, Gaillard, et al., 2015). This task also has been shown to have a
medium-low test-retest reliability (Erickson et al., 2016; Siegelman & Frost, 2015), and several
methodological factors have been proposed explaining the low sensitivity of the 2AFC
approach (Siegelman et al., 2017). There also seems to be a lack of correlation among various
auditory SL tasks themselves. A study comparing several auditory SL paradigms using the
explicit 2AFC task on the same participants reported a lack of correlations between these very
similar paradigms that only differed in the language that was used (Erickson et al., 2016). The
authors therefore concluded that these low correlations were most likely the result of the poor
Neurobiology of Language
229
Frequency-tagging in EEG-based statistical learning research
psychometric properties of the 2AFC measure and that using a composite score of all these
measures combined gives the clearest picture of the situation. Given these low performance
rates, which do not coincide with other measures, it seems that the 2AFC metric is
not sufficiently reliable for determining whether SL has or has not occurred in individual
participants.
The weakness of explicit 2AFC testing has led to the development of more implicit mea-
sures for assessing statistical learning. Some examples of implicit tasks include the target detec-
tion task (Batterink & Paller, 2017; Batterink et al., 2015) adapted in the current study, as well
as rapid serial auditory presentation (Franco, Eberlen, et al., 2015), statistically induced chunk-
ing recall (Isbilen et al., 2017, 2020), and the click detection task (Franco, Gaillard, et al.,
2015; Gómez et al., 2011). These tasks all rely on a similar principle: If pseudowords in the
stream are learned, this will produce a faster implicit response to targets that are associated
with that pseudoword.
In the current study, the implicit target detection test showed evidence for SL in the largest
proportion of participants, with 18/27 participants (70%) showing a significant effect on either
hit rate or RT. Indeed, of all the measures tested here, the implicit task seemed to be the most
sensitive to SL at the individual level. At the same time, this measure is also not ideal. Since only
3 participants showed significant effects in both RT and hit rate, perhaps due to speed-accuracy
tradeoffs, this dilutes the group-level effect of both measures and maintains the operational
ambiguity as to which is the “best” measure to use. This ambiguity is mirrored when looking
at previous studies that employed implicit behavioral tasks and report a highly variable propor-
tion of effects in individual participants. For example, Batterink et al. (2015) report SL effects in
43% of participants using a task similar to the one used here, Gómez et al. (2011) reported SL
effects in 85% of participants, whereas Franco, Eberlen, et al. (2015) found these in only 35% of
participants, with many participants actually showing reverse effects. Moreover, as has been
pointed out previously, the implicit nature of the task makes it difficult to ascertain whether
significant effects truly reflect lexical detection of newly learned words, or if effects are driven
by lower level perceptual familiarity with syllable combinations (Batterink et al., 2015; Franco,
Eberlen, et al., 2015; Isbilen et al., 2017). Moreover, in the current study, the implicit task
was always administered after the explicit 2AFC task, which may have reinforced previous
learning due to the additional exposure to the pseudoword syllable combinations (although
in the 2AFC task participants were also exposed to part-words and were not given feedback
regarding their performance). Taken together, although in the current study the implicit target
detection task seemed to be in line with the proposed ubiquitous nature of SL, the large
variability across behavioral studies (in methods and results) makes it difficult to wholeheart-
edly accept these implicit measures as a reliable benchmark for assessing SL. Further, the
cross-study discrepancies make it extremely difficult to determine the true extent of SL in indi-
vidual participants.
The diverse and inconclusive nature of indirect behavioral measures was one of the primary
motivators for looking to neural measures as more direct signatures of SL. The current study is the
first to assess the robustness of neural SL measures in individual participants using the frequency-
tagging approach. In contrast to the expected ubiquity, we found that only 12/39 participants
(30%) showed significant effects of SL in their EEG spectra. One reason for this might be the poor
SNR in individual-level scalp level EEG, which might be improved upon using other neurophys-
iological measures. For example, a recent ECoG study, which by its nature is based on individual
participants, was able to demonstrate robust neural response at pseudoword-related frequencies,
suggesting that improving the SNR might lead to more robust results (Henin et al., 2021). How-
ever, another factor that exacerbates the complexity of interpreting the frequency-tagging results
Neurobiology of Language
230
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
is that the effects of SL were not observed consistently at the same frequencies, but rather were
seen at different harmonics of the pseudoword rate across participants. This was also the case in
the ECoG data reported by Henin et al. (2021), which leaves many questions open regarding the
underlying mechanism driving these spectral modulations. We can hope that future methodolog-
ical advances will improve the SNR of frequency-tagging measures, which in turn might reveal
more extensive evidence for SL. However, at present, the current results leave us wondering
whether the low prevalence of neural effects corresponding to SL are merely a result of poor
SNR or if they challenge the assumption of the ubiquitous nature of SL. Our results are of partic-
ular importance for endeavors to assess the “cognitive state” of unresponsive patients, using scalp
EEG (Gui et al., 2020; Sokoliuk et al., 2021).
In the absence of a gold standard indication for SL, we turn to look for evidence of con-
verging operations among the multitude of tests that all supposedly measure whether SL has
taken place. Unfortunately, results from the different behavioral and neural measures do not
seem to converge as one might expect if they truly capture the same cognitive operation. In
testing whether neural results corresponded in any way with the behavioral responses, we
found that the subgroup of participants who showed neural evidence for SL also had slightly
faster RTs in the implicit target detection task than those who did not. However, no correspon-
dence was found when examining the within-participant correlation, nor were there any cor-
relations with other behavioral measures. The current results align with previous studies that
also reported no correlation between results on explicit and implicit methods of testing for SL
(Batterink et al., 2015; Franco, Eberlen, et al., 2015; Isbilen et al., 2020; Misyak et al., 2010). In
the few studies where there were significant correlations between explicit and implicit mea-
sures, these were not consistent across different modalities (Isbilen et al., 2020), or differences
in the explicit task (Batterink & Paller, 2017). Some have opted to interpret the lack of a reli-
able cross-measure correlation as an indication that each measure picks up on a different cog-
nitive aspect of SL, for example, suggesting a dissociation between explicit recall and implicit
learning (Batterink et al., 2015; Franco, Eberlen, et al., 2015; Isbilen et al., 2017). This debate
in the literature is ongoing and there does not seem to be a consensus about whether these
measures reflect the same processes. The results of the current study do not attempt to answer
this question, but rather address the possibility that we cannot rule out that all of these
measures—behavioral and neural alike—are simply too crude or too indirect for assessing
the formation of internal memory representations arising from SL. Consequently, it seems that
we still lack a “ground truth” indication for SL, which (at the moment) severely limits the extent
to which this ability can be studied at the level of individual participants.
Conclusions
The current study highlights the utility and the limitations of the EEG frequency-tagging
approach as a research tool for studying SL. At the group level, our results indicate that even
after controlling for possible acoustic confounds, peaks in the neural signal at the pseudoword
frequency (and its harmonics) likely reflect the implicit detection of underlying transitional
probabilities between syllable triplets. However, our data also suggest that the frequency-
tagging approach might not be as useful for studying SL in individual participants. The
frequency-tagged EEG data were less sensitive to SL than the implicit behavioral test, with
effects manifesting at different frequencies across participants. Moreover, the overall low cor-
respondence between the different behavioral and neural metrics, which supposedly all test
for SL, leaves much to be desired in our quest to identify the best operationalization for study-
ing SL. Whether the low-reliability of the EEG results is due to the low SNR of this tool or
whether it is indicative of a deeper flaw in the frequency-tagging approach, is beyond the
Neurobiology of Language
231
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
scope of this paper. Therefore, while some researchers may find this experimental approach
suitable for their needs, the limitations and potential confounds highlighted here should be
taken into consideration when interpreting and comparing results across studies, particularly
regarding individual differences.
FUNDING INFORMATION
Elana Zion Golumbic, German-Israeli Foundation for Scientific Research and Development
(https://dx.doi.org/10.13039/501100001736), Award ID: 1422.
AUTHOR CONTRIBUTIONS
Danna Pinto: Data curation: Lead; Formal analysis: Equal; Investigation: Equal; Methodol-
ogy: Equal; Writing – original draft: Equal; Writing – review & editing: Equal. Anat Prior:
Conceptualization: Equal; Methodology: Supporting; Writing – review & editing: Equal.
Elana Zion Golumbic: Conceptualization: Equal; Data curation: Equal; Formal analysis: Equal;
Funding acquisition: Lead; Investigation: Equal; Methodology: Equal; Project administration:
Lead; Resources: Lead; Supervision: Lead; Validation: Lead; Writing – original draft: Equal;
Writing – review & editing: Equal.
REFERENCES
Arciuli, J. (2017). The multi-component nature of statistical learn-
ing. Philosophical Transactions of the Royal Society of London.
Series B, Biological Sciences, 372(1711), Article 58. https://doi
.org/10.1098/rstb.2016.0058, PubMed: 27872376
Arciuli, J., & von Koss Torkildsen, J. (2012). Advancing our under-
standing of the link between statistical learning and language
acquisition: The need for longitudinal data. Frontiers in Psychol-
ogy, 3, Article 324. https://doi.org/10.3389/fpsyg.2012.00324,
PubMed: 22969746
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical
Software, 67(1), 1–48. https://doi.org/10.18637/jss.v067.i01
Batterink, L. J. (2017). Rapid statistical learning supporting word
extraction from continuous speech. Psychological Science,
28(7), 921–928. https://doi.org/10.1177/0956797617698226,
PubMed: 28493810
Batterink, L. J. (2020). Syllables in sync form a link: Neural
phase-locking reflects word knowledge during language learn-
ing. Journal of Cognitive Neuroscience, 32(9), 1735–1748.
https://doi.org/10.1162/jocn_a_01581, PubMed: 32427066
Batterink, L. J., & Paller, K. A. (2017). Online neural monitoring of
statistical learning. Cortex, 90, 31–45. https://doi.org/10.1016/j
.cortex.2017.02.004, PubMed: 28324696
Batterink, L. J., & Paller, K. A. (2019). Statistical learning of speech
regularities can occur outside the focus of attention. Cortex, 115,
56–71. https://doi.org/10.1016/j.cortex.2019.01.013, PubMed:
30771622
Batterink, L. J., Reber, P. J., Neville, H. J., & Paller, K. A. (2015).
Implicit and explicit contributions to statistical learning. Journal
of Memory and Language, 83, 62–78. https://doi.org/10.1016/j
.jml.2015.04.004, PubMed: 26034344
Buiatti, M., Peña, M., & Dehaene-Lambertz, G. (2009). Investigat-
ing the neural correlates of continuous speech computation with
frequency-tagged neuroelectric responses. NeuroImage, 44(2),
509–519. https://doi.org/10.1016/j.neuroimage.2008.09.015,
PubMed: 18929668
Chen, Y., Jin, P., & Ding, N. (2020). The influence of linguistic infor-
mation on cortical tracking of words. Neuropsychologia, 148,
Article 107640. https://doi.org/10.1016/j.neuropsychologia
.2020.107640, PubMed: 33011188
Choi, D., Batterink, L. J., Black, A. K., Paller, K. A., & Werker, J. F.
(2020). Preverbal infants discover statistical word patterns at
similar rates as adults: Evidence from neural entrainment. Psy-
chological Science, 31(9), 1161–1173. https://doi.org/10.1177
/0956797620933237, PubMed: 32865487
Cunillera, T., Gomila, A., & Rodríguez-Fornells, A. (2008). Benefi-
cial effects of word final stress in segmenting a new language:
Evidence from ERPs. BMC Neuroscience, 9, Article 23. https://
doi.org/10.1186/1471-2202-9-23, PubMed: 18282274
de Diego-Balaguer, R., Rodríguez-Fornells, A., & Bachoud-Lévi, A.-C.
(2015). Prosodic cues enhance rule learning by changing speech
segmentation mechanisms. Frontiers in Psychology, 6, Article 1478.
https://doi.org/10.3389/fpsyg.2015.01478, PubMed: 26483731
Ding, N., Melloni, L., Zhang, H., Tian, X., & Poeppel, D. (2016).
Cortical tracking of hierarchical linguistic structures in connected
speech. Nature Neuroscience, 19(1), 158–164. https://doi.org/10
.1038/nn.4186, PubMed: 26642090
Elmer, S., Valizadeh, S. A., Cunillera, T., & Rodriguez-Fornells, A.
(2021). Statistical learning and prosodic bootstrapping differen-
tially affect neural synchronization during speech segmentation.
NeuroImage, 235, Article 118051. https://doi.org/10.1016/j
.neuroimage.2021.118051, PubMed: 33848624
Emberson, L. L., Conway, C. M., & Christiansen, M. H. (2011).
Timing is everything: Changes in presentation rate have opposite
effects on auditory and visual implicit statistical learning. Quar-
terly Journal of Experimental Psychology, 64(5), 1021–1040.
https://doi.org/10.1080/17470218.2010.538972, PubMed:
21347988
Erickson, L. C., Kaschak, M. P., Thiessen, E. D., & Berry, C. A. S.
(2016). Individual differences in statistical learning: Conceptual
and measurement issues. Collabra: Psychology, 2(14), 1–17.
https://doi.org/10.1525/collabra.41
Neurobiology of Language
232
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Erickson, L. C., & Thiessen, E. D. (2015). Statistical learning of lan-
guage: Theory, validity, and predictions of a statistical learning
account of language acquisition. Developmental Review, 37,
66–108. https://doi.org/10.1016/j.dr.2015.05.002
Evans, J. L., Saffran, J. R., & Robe-Torres, K. (2009). Statistical learn-
ing in children with specific language impairment. Journal of
Speech, Language, and Hearing Research, 52(2), 321–335.
https://doi.org/10.1044/1092-4388(2009/07-0189), PubMed:
19339700
Fernandes, T., Kolinsky, R., & Ventura, P. (2010). The impact
of attentional load on the use of statistical information and coar-
ticulation as speech segmentation cues. Attention, Perception, &
Psychophysics, 72, 1522–1532. https://doi.org/10.3758/APP.72.6
.1522, PubMed: 20675798
Franco, A., Cleeremans, A., & Destrebecqz, A. (2011). Statistical
learning of two artificial languages presented successively:
How conscious? Frontiers in Psychology, 2, Article 229. https://
doi.org/10.3389/fpsyg.2011.00229, PubMed: 21960981
Franco, A., Eberlen, J., Destrebecqz, A., Cleeremans, A., & Bertels,
J. (2015). Rapid serial auditory presentation: A new measure of
statistical learning in speech segmentation. Experimental Psy-
chology, 62(5), 346–351. https://doi.org/10.1027/1618-3169
/a000295, PubMed: 26592534
Franco, A., Gaillard, V., Cleeremans, A., & Destrebecqz, A. (2015).
Assessing segmentation processes by click detection: Online
measure of statistical learning, or simple interference? Behavior
Research Methods, 47, 1393–1403. https://doi.org/10.3758
/s13428-014-0548-x, PubMed: 25515838
Frost, R., Armstrong, B. C., Siegelman, N., & Christiansen, M. H.
(2015). Domain generality versus modality specificity: The para-
dox of statistical learning. Trends in Cognitive Science, 19(3),
117–125. https://doi.org/10.1016/j.tics.2014.12.010, PubMed:
25631249
Getz, H., Ding, N., Newport, E. L., & Poeppel, D. (2018). Cortical
tracking of constituent structure in language acquisition. Cogni-
tion, 181, 135–140. https://doi.org/10.1016/j.cognition.2018.08
.019, PubMed: 30195135
Gómez, D. M., Bion, R. A. H., & Mehler, J. (2011). The word seg-
mentation process as revealed by click detection. Language and
Cognitive Processes, 26(2), 212–223. https://doi.org/10.1080
/01690965.2010.482451
Gómez, R., & Gerken, L. (2000). Infant artificial language learning
and language acquisition. Trends in Cognitive Science, 4(5),
178–186. https://doi.org/10.1016/S1364-6613(00)01467-4,
PubMed: 10782103
Graf Estes, K., Evans, J. L., Alibali, M. W., & Saffran, J. R. (2007).
Can infants map meaning to newly segmented words? Statistical
segmentation and word learning. Psychological Science, 18(3),
254–260. https://doi.org/10.1111/j.1467-9280.2007.01885.x,
PubMed: 17444923
Gui, P., Jiang, Y., Zang, D., Qi, Z., Tan, J., Tanigawa, H., Jiang, J.,
Wen, Y., Xu, L., Zhao, J., Mao, Y., Poo, M., Ding, N., Dehaene, S.,
Wu, X., & Wang, L. (2020). Assessing the depth of language
processing in patients with disorders of consciousness. Nature
Neuroscience, 23, 761–770. https://doi.org/10.1038/s41593
-020-0639-1, PubMed: 32451482
Har-shai Yahav, P., & Zion Golumbic, E. (2021). Linguistic processing
of task-irrelevant speech at a cocktail party. eLife, 10, Article
e65096. https://doi.org/10.7554/eLife.65096, PubMed: 33942722
Henin, S., Turk-Browne, N. B., Friedman, D., Liu, A., Dugan, P.,
Flinker, A., Doyle, W., Devinsky, O., & Melloni, L. (2021).
Learning hierarchical sequence representations across human
cortex and hippocampus. Science Advances, 7(8), Article
eabc4530. https://doi.org/10.1126/sciadv.abc4530, PubMed:
33608265
Hsu, H. J., Tomblin, J. B., & Christiansen, M. H. (2014). Impaired
statistical learning of non-adjacent dependencies in adolescents
with specific language impairment. Frontiers in Psychology, 5,
Article 175. https://doi.org/10.3389/fpsyg.2014.00175,
PubMed: 24639661
Isbilen, E. S., McCauley, S. M., Kidd, E., & Christiansen, M. H.
(2017). Testing statistical learning implicitly: A novel chunk-
based measure of statistical learning. In G. Gunzelmann, A.
Howes, T. Tenbrink, & E. Davelaar (Eds.), Proceedings of the
39th annual conference of the Cognitive Science Society (CogSci
2017) (pp. 564–569). Cognitive Science Society.
Isbilen, E. S., McCauley, S. M., Kidd, E., & Christiansen, M. H.
(2020). Statistically induced chunking recall: A memory-based
approach to statistical learning. Cognitive Science, 44(7), Article
e12848. https://doi.org/10.1111/cogs.12848, PubMed: 32608077
JASP Team. (2019). JASP ( Version 0.14) [Computer software].
https://jasp-stats.org/download/
Kang, H., Auksztulewicz, R., An, H., Abi Chacra, N., Sutter, M. L.,
& Schnupp, J. (2021). Neural correlates of auditory pattern learn-
ing in the auditory cortex. Frontiers in Neuroscience, 15, Article
610978. https://doi.org/10.3389/fnins.2021.610978, PubMed:
33790730
Karuza, E. A., Newport, E. L., Aslin, R. N., Starling, S. J., Tivarus,
M. E., & Bavelier, D. (2013). The neural correlates of statistical
learning in a word segmentation task: An fMRI study. Brain and
Language, 127(1), 46–54. https://doi.org/10.1016/j.bandl.2012
.11.007, PubMed: 23312790
Kent, R. D., & Read, C. (2002). The acoustic analysis of speech
(2nd ed.). Singular/Thomson Learning.
Kiai, A., & Melloni, L. (2021). What canonical online and offline
measures of statistical learning can and cannot tell us. BioRxiv.
https://doi.org/10.1101/2021.04.19.440449
Kidd, E. (2012). Implicit statistical learning is directly associated
with the acquisition of syntax. Developmental Psychology,
48(1), 171–184. https://doi.org/10.1037/a0025405, PubMed:
21967562
Kidd, E., & Arciuli, J. (2016). Individual differences in statistical
learning predict children’s comprehension of syntax. Child
Development, 87(1), 184–193. https://doi.org/10.1111/cdev
.12461, PubMed: 26510168
Kim, R., Seitz, A., Feenstra, H., & Shams, L. (2009). Testing assump-
tions of statistical learning: Is it long-term and implicit? Neurosci-
ence Letters, 461, 145–149. https://doi.org/10.1016/j.neulet
.2009.06.030, PubMed: 19539701
Lu, L., Sheng, J., Liu, Z., & Gao, J. H. (2021). Neural representations
of imagined speech revealed by frequency-tagged magnetoen-
cephalography responses. NeuroImage, 229, Article 117724.
https://doi.org/10.1016/j.neuroimage.2021.117724, PubMed:
33421593
Lukics, K. S., & Lukács, Á. (2021). Tracking statistical learning
online: Word segmentation in a target detection task. Acta Psy-
chologica, 215, Article 103271. https://doi.org/10.1016/j.actpsy
.2021.103271, PubMed: 33765521
Luo, C., & Ding, N. (2020). Cortical encoding of acoustic and lin-
guistic rhythms in spoken narratives. eLife, 9, 1–25. https://doi
.org/10.7554/eLife.60433, PubMed: 33345775
Makov, S., Sharon, O., Ding, N., Ben-Shachar, M., Nir, Y., & Zion
Golumbic, E. (2017). Sleep disrupts high-level speech parsing
despite significant basic auditory processing. Journal of Neurosci-
ence, 37(32), 7772–7781. https://doi.org/10.1523/JNEUROSCI
.0168-17.2017, PubMed: 28626013
Neurobiology of Language
233
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Frequency-tagging in EEG-based statistical learning research
Misyak, J. B., & Christiansen, M. H. (2012). Statistical learning and
language: An individual differences study. Language Learning, 62(1),
302–331. https://doi.org/10.1111/j.1467-9922.2010.00626.x
Misyak, J. B., Christiansen, M. H., & Bruce Tomblin, J. (2010). On-
line individual differences in statistical learning predict language
processing. Frontiers in Psychology, 1, Article 21. https://doi.org
/10.3389/fpsyg.2010.00031, PubMed: 21833201
Niesen, M., Vander Ghinst, M., Bourguignon, M., Wens, V., Bertels,
J., Goldman, S., Choufani, G., Hassid, S., & De Tiège, X. (2019).
Tracking the effects of top–down attention on word discrimina-
tion using frequency-tagged neuromagnetic responses. Journal of
Cognitive Neuroscience, 32(5), 877–888. https://doi.org/10.1162
/jocn_a_01522, PubMed: 31933439
Olson, I. R., & Chun, M. M. (2001). Temporal contextual cuing of
visual attention. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 27(5), 1299–1313. https://doi.org/10
.1037/0278-7393.27.5.1299, PubMed: 11550756
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011). Field-
Trip: Open source software for advanced analysis of MEG, EEG,
and invasive electrophysiological data. Computational Intelli-
gence and Neuroscience, 2011, Article 156869. https://doi.org
/10.1155/2011/156869, PubMed: 21253357
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M., Höchenberger, R.,
Sogo, H., Kastman, E., & Lindeløv, J. K. (2019). PsychoPy2:
Experiments in behavior made easy. Behavior Research Methods,
51, 195–203. https://doi.org/10.3758/s13428-018-01193-y,
PubMed: 30734206
Pelucchi, B., Hay, J., & Saffran, J. (2009). Statistical learning in a
natural language by 8-month-old. Child Development, 80(3),
674–685. https://doi.org/10.1111/j.1467-8624.2009.01290.x,
PubMed: 19489896
Romberg, A. R., & Saffran, J. R. (2013). All together now: Concur-
rent learning of multiple structures in an artificial language.
Cognitive Science, 37(7), 1290–1320. https://doi.org/10.1111
/cogs.12050, PubMed: 23772795
Saffran, J. R., Aslin, R. N., & Newport, E. L. (1996). Statistical learn-
ing by 8-month-old infants. Science, 274(5294), 1926–1928.
https://doi.org/10.1126/science.274.5294.1926, PubMed:
8943209
Saffran, J. R., Newport, E. L., Aslin, R. N., Tunick, R. A., & Barrueco,
S. (1997). Incidental language learning: Listening (and learning)
out of the corner of your ear. Psychological Science, 8(2),
101–105. https://doi.org/10.1111/j.1467-9280.1997.tb00690.x
Santolin, C., Rosa-Salva, O., Vallortigara, G., & Regolin, L. (2016).
Unsupervised statistical learning in newly hatched chicks.
Current Biology, 26(23), R1218–R1220. https://doi.org/10.1016
/j.cub.2016.10.011, PubMed: 27923125
Santolin, C., & Saffran, J. R. (2018). Constraints on statistical learn-
ing across species. Trends in Cognitive Science, 22(1), 52–63.
https://doi.org/10.1016/j.tics.2017.10.003, PubMed: 29150414
Sheng, J., Zheng, L., Lyu, B., Cen, Z., Qin, L., Tan, L. H., Huang,
M.-X., Ding, N., & Gao, J.-H. (2019). The cortical maps of hier-
archical linguistic structures during speech perception. Cerebral
Cortex, 29(8), 3232–3240. https://doi.org/10.1093/cercor
/bhy191, PubMed: 30137249
Siegelman, N., Bogaerts, L., & Frost, R. (2017). Measuring individ-
ual differences in statistical learning: Current pitfalls and possible
solutions. Behavior Research Methods, 49(2), 418–432. https://
doi.org/10.3758/s13428-016-0719-z, PubMed: 26944577
Siegelman, N., & Frost, R. (2015). Statistical learning as an individ-
ual ability: Theoretical perspectives and empirical evidence.
Journal of Memory and Language, 81, 105–120. https://doi.org
/10.1016/j.jml.2015.02.001, PubMed: 25821343
Sokoliuk, R., Degano, G., Banellis, L., Melloni, L., Hayton, T.,
Sturman, S., Veenith, T., Yakoub, K. M., Belli, A., Noppeney,
U., & Cruse, D. (2021). Covert speech comprehension predicts
recovery from acute unresponsive states. Annals of Neurology,
89(4), 646–656. https://doi.org/10.1002/ana.25995, PubMed:
33368496
Spencer, M., Kaschak, M. P., Jones, J. L., & Lonigan, C. J. (2015).
Statistical learning is related to early literacy-related skills. Read-
ing and Writing, 28(4), 467–490. https://doi.org/10.1007/s11145
-014-9533-0, PubMed: 26478658
Thiessen, E. D., & Saffran, J. R. (2003). When cues collide: Use
of stress and statistical cues to word boundaries by 7- to
9-month-old infants. Developmental Psychology, 39(4),
706–716. https://doi.org/10.1037/0012-1649.39.4.706,
PubMed: 12859124
Thiessen, E. D., & Saffran, J. R. (2007). Learning to learn: Infants’
acquisition of stress-based strategies for word segmentation.
Language Learning and Development, 3(1), 73–100. https://doi
.org/10.1080/15475440709337001
Toro, J. M., Sinnett, S., & Soto-Faraco, S. (2005). Speech segmenta-
tion by statistical learning depends on attention. Cognition,
97(2), B25–B34. https://doi.org/10.1016/j.cognition.2005.01
.006, PubMed: 16226557
Toro, J. M., Sinnett, S., & Soto-Faraco, S. (2011). Generalizing
linguistic structures under high attention demands. Journal of
Experimental Psychology: Learning, Memory, and Cognition, 37(2),
493–501. https://doi.org/10.1037/a0022056, PubMed: 21261426
Turk-Browne, N. B., Jungé, J. A., & Scholl, B. J. (2005). The auto-
maticity of visual statistical learning. Journal of Experimental
Psychology: General, 134(4), 552–564. https://doi.org/10.1037
/0096-3445.134.4.552, PubMed: 16316291
Tyler, M. D., & Cutler, A. (2009). Cross-language differences in cue
use for speech segmentation. Journal of the Acoustical Society of
America, 126, 367–376. https://doi.org/10.1121/1.3129127,
PubMed: 19603893
van der Wulp, I. M. (2021). Word segmentation: TP or OCP? A
re-analysis of Batterink and Paller (2017) [Unpublished Master’s
Thesis]. Utrecht University.
Wang, T., & Saffran, J. R. (2014). Statistical learning of a tonal lan-
guage: The influence of bilingualism and previous linguistic
experience. Frontiers in Psychology, 5, Article 953. https://doi
.org/10.3389/fpsyg.2014.00953, PubMed: 25232344
Zhou, H., Melloni, L., Poeppel, D., & Ding, N. (2016). Interpreta-
tions of frequency domain analyses of neural entrainment: Peri-
odicity, fundamental frequency, and harmonics. Frontiers in
Human Neuroscience, 10, Article 274. https://doi.org/10.3389
/fnhum.2016.00274, PubMed: 27375465
Neurobiology of Language
234
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
1
4
1
9
8
9
3
1
6
n
o
_
a
_
0
0
0
6
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3