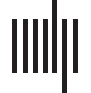ARTÍCULO DE INVESTIGACIÓN
The Neural Basis of Metaphor Comprehension:
Evidence from Left Hemisphere Degeneration
Nathaniel Klooster1,2
, Marguerite McQuire1, Murray Grossman1,2
,
Corey McMillan1,2, Anjan Chatterjee1,3,4, and Eileen Cardillo1,4
1Department of Neurology, Perelman School of Medicine, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
2Penn Frontotemporal Degeneration Center, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
3Moss Rehabilitation Research Institute, Elkins Park, Pensilvania, EE.UU
4Penn Center for Neuroaesthetics, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
Palabras clave: figurative language, neurodegeneration, left hemisphere, LpMTG, LIFG
ABSTRACTO
Despite the ubiquity of metaphor in cognition and communication, it is absent from standard
clinical assessments of language, and the neural systems that support metaphor processing are
debated. Previous research shows that patients with focal brain lesions can display selective
impairments in processing metaphor, suggesting that figurative language abilities may be
disproportionately vulnerable to brain injury. We hypothesized that metaphor processing is
especially vulnerable to neurodegenerative disease, and that the left hemisphere is critical for
normal metaphor processing. To evaluate these hypotheses, we tested metaphor comprehension
in patients with left-hemisphere neurodegeneration, and in demographically matched healthy
comparison participants. Stimuli consisted of moderately familiar metaphors and closely
matched literal sentences sharing the same source term (p.ej., The interview was a painful crawl /
The infant’s motion was a crawl). Written sentences were presented, followed by four modifier-
noun answer choices (one target and three foils). Healthy controls, though reliably better at literal
than metaphor trials, comprehended both sentence conditions well. Por el contrario, participants with
left-hemisphere neurodegeneration performed disproportionately poorly on metaphor
comprensión. Anatomical analyses show relationships between metaphor accuracy and
patient atrophy in the left middle and superior temporal gyri, and the left inferior frontal gyrus,
areas that have been implicated in supporting metaphor comprehension in previous imaging
investigación. The behavioral results also suggest deficits of metaphor comprehension may be a
sensitive measure of cognitive dysfunction in some forms of neurodegenerative disease.
INTRODUCCIÓN
Metaphor, the description of one idea in terms of another, plays an important role in cognition
and communication. The use of metaphor is pervasive, accounting for more than 13% of words in
written discourse and more than 6% of spoken language (Steen et al., 2010). Metaphor strongly
influences people’s thinking in fundamental ways. The metaphors of “shaping” or “framing” are
often used to describe the influence of metaphor on thinking (p.ej., Lakoff & Johnson, 1980). Meta-
analyses show that metaphorical language is more persuasive than comparable literal language
(Sopory & Dillard, 2002; Van Stee, 2018). The presence or absence of metaphor when describing
an issue affects how people think about topics such as cancer (p.ej., Gibbs & Franks, 2002; Hauser
& Schwarz, 2015), crime (Thibodeau & Boroditsky, 2011, 2013), significant personal relation-
buques (Sotavento & Schwarz, 2014; Robins & Mayer, 2000), the acceptability of cognitive enhancement
un acceso abierto
diario
Citación: Klooster, NORTE., McQuire, METRO.,
Grossman, METRO., McMillan, C., Chatterjee, A.,
& Cardillo, mi. (2020). The neural basis of
metaphor comprehension: Evidencia
from left hemisphere degeneration.
Neurobiology of Language, 1(4),
474–491. https://doi.org/10.1162/
nol_a_00022
DOI:
https://doi.org/10.1162/nol_a_00022
Supporting Information:
https://doi.org/10.1162/nol_a_00022
Recibió: 26 Abril 2019
Aceptado: 14 Junio 2020
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Nathaniel Klooster
klooster@pennmedicine.upenn.edu
Editor de manejo:
Steven Small
Derechos de autor: © 2020 Massachusetts
Institute of Technology. Publicado
bajo una atribución Creative Commons
4.0 Internacional (CC POR 4.0) licencia.
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
norte
oh
_
a
_
0
0
0
2
2
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
(Conrad, Humphries, & Chatterjee, 2019), the urgency of climate change (Flusberg, Matlock, &
Thibodeau, 2017; Nerlich & Jaspal, 2012), and the brilliance of an idea (Elmore & Luna-Lucero,
2017).
Beyond influencing how people think, metaphor can affect how people act. Metaphor can
drive attention (Matlock, 2004), affecting what information is attended to in social environments
(Bowes & katz, 2015). The choice of metaphor can affect how communities choose to handle
public health issues (Barry, Brescoll, Brownell, & Schlesinger, 2009) and how patients are routed
through health care facilities (Hilligoss, 2014). Because metaphorical language can be persuasive
and can influence behavior, metaphors are widely used in didactic contexts (see the review in
Saban, 2006), and in the teaching of medicine (Van Rijn-Van Tongeren, 1997), and in science
específicamente (Beger & Jäkel, 2015). Metaphor use is pervasive, it frames thinking, and it affects
human behavior.
The neural bases of metaphor are debated. Early studies of metaphor comprehension focused
on possible hemispheric differences, with some highlighting a privileged role for the right hemi-
sphere (Bottini et al., 1994; Brownell, Simpson, Bihrle, Potter, & jardinero, 1990; Winner &
jardinero, 1977). The right hemisphere hypothesis for metaphor argues that the right hemisphere
plays a specific and necessary role in supporting the ability to understand metaphor. The right
hemisphere is thought to be especially important in the comprehension of novel metaphors
(Bohrn, Altmann, & Jacobs, 2012; Mashal & Faust, 2009; Mashal, Faust, & Hendler, 2005).
Over time, sin embargo, the role of the left hemisphere in metaphor comprehension has begun to
be considered more seriously. Functional imaging studies indicate left hemisphere regions
support metaphor comprehension in addition to right hemisphere regions (Bambini, Gentili,
Ricciardi, Bertinetto, & Pietrini 2011; Cardillo, watson, Schmidt, Kranjec, & Chatterjee, 2012;
Chen, Widick, & Chatterjee, 2008; Sotavento & Dapretto, 2006; Obert et al., 2014; Schmidt &
Seger, 2009; Cual, Batán, Khodaparast, & Krawczyk, 2010), and sometimes perhaps exclusively
(Diaz, Barrett, & Hogstrom, 2011; Rapp, Leube, Erb, Grodd, & Kircher, 2004, 2007). Meta-analyses
of the functional imaging literature also suggest that metaphor comprehension is a bilaterally
mediated process (Bohrn et al., 2012; Rapp, Mutschler, & Erb, 2012; Reyes-Aguilar, Valles-
Capetillo,& Giordano, 2018; Cual, 2014). These studies indicate that the bilateral inferior
frontal gyri and the left posterior middle temporal gyrus (LpMTG) specifically are activated
during metaphor comprehension.
Neuroimaging studies have their inferential limitations. They are limited in making claims of a
causal role for brain regions underlying a cognitive function. Patient studies, while more difficult
to conduct, offer an important constraint on theorizing based solely on imaging studies (Fellows
et al., 2005). Studies with focal lesion patients find evidence of patients with selective metaphor
deficits despite normal literal sentence comprehension following left-sided damage (Cardillo,
McQuire, & Chatterjee, 2018; Gagnon, Goulet, Giroux, & Joanette, 2003; Ianni, Cardillo,
McQuire, & Chatterjee, 2014; Tompkins, 1990). These studies implicate the left frontal and pos-
terior temporal cortices in metaphor comprehension (Cardillo et al., 2018; Zaidel, Kasher,
Soroker, & Batori, 2002). The traditional view that the right hemisphere plays a privileged role
in metaphor comprehension is increasingly difficult to justify, though it remains a staple of clinical
and basic neuroscience teaching.
The neural bases for metaphor comprehension are relevant to models of language processing,
but also for adequately addressing the therapeutic needs of clinical populations. Studies of
diverse patient populations indicate metaphor processing is frequently impacted by brain injury
or disease. Following a traumatic brain injury, patients displayed disruptions with metaphor pro-
cesando (Yang et al., 2010). Studies also report impairments in metaphor comprehension in
Neurobiology of Language
475
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
norte
oh
_
a
_
0
0
0
2
2
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
Parkinson’s disease (Fernandino et al., 2013; Monetta & Pell, 2007) and Alzheimer’s disease
(Amanzio, Geminiani, Leotta, & Cappa, 2008; Papagno, 2001; Roncero & de Almeida, 2014;
Winner & jardinero, 1977). Figurative language abilities more broadly are impaired in mild cog-
nitive impairment (Cardoso, silva, Maroco, de Mendonça, & Guerreiro, 2014) and in Alzheimer’s
enfermedad (Papagno, Lucchelli, Muggia, & Rizzo, 2003). These studies raise the possibility that met-
aphor comprehension is an especially fragile linguistic ability. In contrast to stable, focal lesions,
neurodegenerative diseases have neural consequences and cognitive impairments that change
gradually over time. Metaphor processing is a complex cognitive ability that requires contribu-
tions from many subdomains of cognition. Degenerative damage in distinct neural systems may
lead to a common cognitive impairment. Por ejemplo, the effects of small decrements in multiple
aspects of cognition could aggregate to a failure in metaphor comprehension. The complexity of
metaphor processing and its resulting fragility in the face of distributed brain damage raise
the possibility that metaphor comprehension is a sensitive measure of cognitive dysfunction in
neurodegenerative disease.
The Current Study
We hypothesized that metaphor processing relies on an intact left hemisphere and that met-
aphor comprehension is vulnerable to neurodegenerative disease. To evaluate these hypoth-
eses, we tested metaphor comprehension with a rigorously controlled task in a group of
patients with left hemisphere neurodegeneration (LHND), and in demographically matched
healthy control (HC) Participantes. While many patient studies of metaphor processing are
strictly behavioral, the current study related behavior to structural MRI. Patients first under-
went structural MRI scans and later completed the metaphor task. Task performance was re-
lated to cortical thickness values. We predicted that patients would be especially impaired on
metaphor trials (compared to HCs and compared to their own performance on literal trials) y
that these impairments would be associated with patterns of left temporal lobe atrophy in the
patient group. Based on results of functional imaging studies of metaphor processing using
stimuli from the same set as used here (Cardillo et al., 2012), we assessed whether patterns
of atrophy in three regions of interest (ROI) frequently engaged by metaphor tasks—the left
inferior frontal gyrus (LIFG) , the right inferior frontal gyrus (RIFG), and the LpMTG—would
relate to metaphor task performance.
MATERIALES Y MÉTODOS
Metaphor Task
Target sentences
Stimuli included 32 matched metaphor–literal sentence pairs taken from the published stimuli
sets of Cardillo, Schmidt, Kranjec, & Chatterjee, 2010; Cardillo, watson, & Chatterjee, 2016).
Todo 64 sentences were in the form “The X was a Y,” where Y was the shared word or phrase in
the literal–metaphor match (lo sucesivo, source term). The source term of the sentence, Y, was either
an entity noun (p.ej., The relay was a sprint race / The math test was an intelligence race) o un
event noun (The interruption was a loud knock / His emails were an insistent knock). Source terms
also always included sensorimotor features—half auditory (p.ej., knock) and half motion (p.ej.,
carrera). See Table 1 for examples.
Stochastic Optimization of Stimuli software (Armstrong, watson, & Plaut, 2012) fue usado
to ensure that metaphor and literal sentences were matched on many of the published norms
for the items: the number of characters, palabras, and content words; and the average frequency
and concreteness of their content words; as well as their valence (% positivo), an online
Neurobiology of Language
476
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
norte
oh
_
a
_
0
0
0
2
2
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
Mesa 1.
Stimuli examples
Oración
Metaphor
Tipo
Ejemplo
Entity-Auditory Her chores were a sad tune.
Target
gloomy routine
Foil 1
funeral hymn
Foil 2
playful exercise
Foil 3
prison system
Entity-Motion
The summer romance was
dizzying delight
amusement
serious
brick fireplace
a merry-go-round.
ride
punishment
Event-Auditory
The cool breeze was
calming weather
a lullaby.
reassuring
melody
uncomfortable
temperatura
cracked basin
Event-Motion
The prize money was a lift.
financial
small elevator
economic burden
delicious apple
asistencia
Literal
Entity-Auditory
The jingle was a happy tune.
catchy song
loose change
radio static
flower vase
Entity-Motion
The construction was a new
colorful carousel
real estate
moldy dungeon
grocery cart
merry-go-round.
Event-Auditory
The child’s favorite was
soothing song
a lullaby.
biological
offspring
screaming match
cracked
knuckles
Event-Motion
The bed was a heavy lift.
weighty mattress
striped sofa
lightweight frame
full trashcan
measure of semantic processing difficulty (valence judgment reaction time), and familiarity (todo
p’s > 0.09). Values for these variables were taken from published norms (Cardillo et al., 2010;
Cardillo et al., 2016). En general, metaphors were moderately familiar (4.6 / 7; 1 = very unfamiliar,
7 = very familiar) and were well-understood (Interpretability M = 0.91, DE = 0.09). Metaphors
were less imageable than their literal counterparts ( pag < 0.01) and, as intended, more figurative
( p < 0.01). These differences and equivalences held true whether collapsing across variables
of noninterest (modality, source term), or whether distinguishing items further by these char-
acteristics. For item properties, see Supplementary Table 2 in the online supporting informa-
tion located at https://www.mitpressjournals.org/doi/suppl/10.1162/nol_a_00022.
Answer choices
Each target sentence was accompanied by four possible answers, a correct target and three foils.
Each answer choice consisted of a modifier (adjective or noun) and a noun. Foils for the metaphor
stimuli (Table 1) were (1) the literal meaning of the sentence, (2) the opposite of the metaphorical
meaning of the sentence, and (3) an unrelated answer. Foils for the literal sentences were (1) a
category associate of the agent of the sentence not implied by the sentence, (2) the opposite of the
literal meaning of the sentence, and (3) an unrelated answer. In this way, answer choices were
designed to be informative of the nature of comprehension difficulty on incorrect trials. Answer
choices were matched on average frequency, concreteness, and valence. For full materials used
in this study see Supplementary Table 1 in the online supporting information.
Task procedure
For both groups, stimuli were presented visually on a laptop using E-Prime 2.0 software. On
every trial, a sentence was presented at the top of the screen. When the participant indicated
they were done reading the target sentence, the sentence remained on the screen and answer
choices were presented below it, randomized to one of four quadrants in the lower half of the
screen. Participants were instructed to choose the answer that best matched the meaning of the
sentence and to guess if unsure.
Neurobiology of Language
477
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
Patients read the sentences and indicated to a researcher (orally or by pointing) which answer
they thought best matched the meaning of the sentence. To limit demands on working memory
for patients, the research assistant recorded the patient’s answer and advanced the trial. HCs
controlled the testing laptop and made their responses without assistance.
Piloting Process
Ten adults free of neurological disease or brain injury and with overall cognition and verbal
intelligence within the normal range (Mean = 27.9 ± 2.0, Mini-Mental State Examination
[MMSE; Folstein, Folstein, & McHugh, 1975]; Mean = 113.6 ± 10.1, American National
Adult Reading Test [AMNART; Nelson & O’Connell, 1978]) were recruited from a database
of healthy older adults to serve as pilot participants for the metaphor and literal multiple-
choice stimuli. Participants were paid $15/hr and gave informed consent for their participation in accordance with procedures of the University of Pennsylvania Institutional Review Board (protocol #806447). Pilot participants were native English speakers and were matched to the patient group on age (M = 69.5 ± 7.5) and education (M = 15.3 ± 3.0). Pilot participants completed the task as described above. Item accuracy analysis revealed seven of the 64 items for which the correct answer was selected ≤60% of the time. Based on the foils most commonly chosen for these items, target answers and/or foils were revised to disambiguate the answer choices. Target sentences were not altered. Patients and a new group of HCs were tested on these revised stimuli. l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u n o / l / l a r t i c e - p d f / / / / 1 4 4 7 4 1 8 6 7 7 7 6 n o _ a _ 0 0 0 2 2 p d . / l f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Participants Healthy comparison participants Nineteen HCs were recruited from a database of healthy older adults to participate in the study. HCs were paid $15/hr and gave informed consent in accordance with procedures of
the University of Pennsylvania Institutional Review Board (protocol #806447). HCs were native
English speakers, matched to the patient group on age (M = 65.2 ± 10.2) and education (M =
15.4 ± 2.6). Neuropsychological testing confirmed they scored within the normal range on the
MMSE (M = 28.8 ± 1.6), indicating normal cognition, and on the AMNART (M = 117.83 ± 8.20),
indicating normal verbal intelligence.
Patients
Thirteen patients were recruited from a Frontotemporal Degeneration Center to participate in
the study. All patients were diagnosed with logopenic-variant primary progressive aphasia
(lvPPA) at the time of testing, according to established criteria (Gorno-Tempini et al., 2011),
and confirmed through a consensus process. Since this initial diagnosis, on reassessment three
were diagnosed with progressive supranuclear palsy, one with Alzheimer’s disease, one with
behavioral-variant fronto-temporal dementia, and one with nonfluent PPA. These were clinical
diagnoses based only on phenotype. The remaining patients retained a lvPPA diagnosis at the
time of publication. Patients and HC participants were matched on age and education. Patients
displayed mild impairment on the MMSE (M = 24.62, Tables 2 and 3) and as a group performed
significantly worse than HCs (Table 2). Patients were paid $15/hr and gave informed consent
in accordance with procedures of the University of Pennsylvania Institutional Review Board
(protocol #806447).
Neurobiology of Language
478
Impaired metaphor comprehension with left hemisphere degeneration
Table 2. Demographics
Group
LHND (n =13)
Age
63.31 (± 6.9)
Chronicity
3.69 (± 1.49)
Education
15.62 (± 2.9)
MMSE
24.62 (± 4.5)**
HC (n = 19)
65.21 (± 10.2)
15.39 (± 2.6)
28.58 (± 1.6)**
Note. ** HC MMSE > LHND ( pag < 0.01). LHND = left-hemisphere neurodegeneration, HC = healthy control,
MMSE = Mini-Mental State Examination.
Anatomical Methods
T1 Whole-brain imaging
High-resolution T1-weighted MPRAGE structural scans were acquired for all but one patient
(excluded due to claustrophobia and anxiety even following sedation) and 35 healthy controls
comparable to the patient group (Age: M = 67.37, p < 0.15; Education: M = 16.03, p < 0.65;
10 males). MRI data was acquired on a 3T Siemens Tim Trio scanner with an 8-channel head
coil, with T = 1,620 ms, T = 3.09 ms, flip angle = 15°, 192 × 256 matrix, and 1 mm3 voxels.
T1-weighted MRI images were then preprocessed to compute cortical thickness using Advanced
Normalization Tools (ANTs; Tustison et al., 2014). Briefly, each individual dataset was deformed
using a symmetric and diffeomorphic registration routine using ANTs to register each volume to a
standard local template space in a canonical stereotactic coordinate system. ANTs provide a
highly accurate registration routine using symmetric and topology-preserving diffeomorphic
deformations to minimize bias toward the reference space and to capture the deformation
necessary to aggregate images in a common space. Then, we used N4 bias correction to min-
imize heterogeneity (Tustison et al., 2010) and the ANTs Atropos tool to segment images into
six tissue classes (cortex, white matter, cerebrospinal fluid, subcortical grey structures, brain-
stem, and cerebellum) using template-based priors, and to generate probability maps of each
tissue. Voxel-wise cortical thickness was measured in millimeters (mm) from the pial surface
and then transformed into Montreal Neurological Institute (MNI) space, smoothed using a 2
sigma full-width half-maximum Gaussian kernel, and downsampled to 2 mm isotropic voxels.
We conducted several types of anatomic analyses. The first targeted specific ROIs based on
activation patterns in previous fMRI studies of metaphor comprehension. The second exam-
ined single-subject atrophy patterns in a subgroup of patients who show metaphor impairment.
Exploratory supplemental analyses probed brain-behavior relationships of task performance
within areas of degeneration as identified by a group mask.
To characterize brain areas where the patients as a group displayed significant neural degen-
eration relative to matched controls, a patient atrophy mask (Figure 1 and Supplementary Table 3
in the online supporting information) was calculated through nonparametric permutation-based
analyses with threshold-free cluster enhancement (TFCE; Smith & Nichols, 2009) with the
randomize tool in FSL (http://fsl.fmrib.ox.ac.uk/fsl/fslwiki). Cortical thickness was compared
across groups. To depict atrophic areas that show maximal overlap in the group, clusters that meet
a conservative threshold of p < 0.005 (family-wise error corrected with TFCE), and contain a
minimum of 200 adjacent voxels are reported.
When a more liberal threshold of p < 0.05 is applied, areas of variability driven by smaller
subsets of participants are evident (Supplementary Figure 1 in the online supporting information).
Patients varied widely in the extent of atrophy present in regions outside the left temporal lobe.
To specifically probe ROIs implicated as critical nodes in the metaphor comprehension network,
individual variability in patient performance was related to variability in cortical thickness in three
ROIs: the LIFG, the RIFG, and the LpTMG temporal gyrus. ROIs were generated by creating 10 mm
Neurobiology of Language
479
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 3.
Patient neuropsychological profile
LHND1 LHND2 LHND3 LHND4 LHND5 LHND6 LHND7 LHND8 LHND9 LHND10 LHND11 LHND12 LHND13 Mean
4.77
3
3
9
8
4
0
0
5
4
8
2
8
8
Subject
PVLT Recall
PPT Words
PPT Pictures
24
26
25
26
Naming
93.33
86.67
Animal Fluency
“f” fluency
Trails A Time
Trails B Time
PVLT1
Digit Span
Forward
Digit Span
Backward
19
13
31
80
5
3
5
19
15
35
72
4
3
5
25
24
90
10
7
46
25
25
40
11
7
45
300
130
1
5
4
1
5
3
21
23
60
1
3
–
–
3
5
0
16
19
25
26
23
22
23
24
25
26
13
24
22
24
36.67
81.25
18.75
93.33
93.75
96.88
78.13
6
10
89
11
7
37
282
211
1
5
6
1
1
2
5
5
30
166
0
2
2
13
4
48
228
4
2
3
10
17
130
300
4
4
4
16
9
34
182
4
4
4
24
24
–
6
4
66
–
0
2
1
22.38
24.10
57.9
10.31
8.08
126.23
193.82
2.54
3.92
3.54
30
24.61
7
4
54
181
5
10
7
29
MMSE
29
27
23
28
25
19
21
26
28
20
15
Note. LHND = left-hemisphere neurodegeneration, PVLT = Philadelphia Verbal Learning Test (Libon, Mattson, Glosser, & Kaplan, 1996), PPT = Pyramids and Palm Trees (Howard &
Patterson, 1992), MMSE = Mini-Mental State Exam.
I
m
p
a
i
r
e
d
m
e
t
a
p
h
o
r
c
o
m
p
r
e
h
e
n
s
i
o
n
w
i
t
h
l
e
f
t
h
e
m
i
s
p
h
e
r
e
d
e
g
e
n
e
r
a
t
i
o
n
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
4
8
0
Impaired metaphor comprehension with left hemisphere degeneration
Figure 1. Patient whole brain atrophy. Pattern of cortical thinning in patients compared to healthy
comparisons (significant at p < 0.005, family-wise error corrected with threshold-free cluster
enhancement).
radius spheres around the peak coordinates in MNI space reported for each of these clusters in an
fMRI study of metaphor comprehension using similar stimuli (LIFG = −50, 29, −1; RIFG = 50, 26, 5;
LpMTG = −62, −50, −9; Cardillo et al., 2012). Performance across literal and metaphor conditions
was related to cortical thickness in these ROIs while controlling for age, sex, and years of education.
To characterize individual subject patterns of reduced cortical thickness in three individuals
from the metaphor-impaired subgroup (see Neuroanatomical Results), individualized heatmaps
of Z-transformed cortical thickness relative to 156 demographically comparable healthy adults
with a self-reported negative psychiatric and neurological history were generated. These HCs were
additionally screened as cognitively normal using a >27 (out of 30) score on the MMSE. To gen-
erate these heatmaps, the mean and SD for each voxel in template space for the control cohort
were calculated. Entonces, Z scores of each patient’s whole brain cortical thickness map relative to the
HC means and SDs were generated. Individual heatmaps were then masked by the patient atrophy
mask and can be interpreted as an LHND-specific Z map of age, sexo, and education appropriate
patterns of reduced cortical thickness.
Supplemental exploratory analyses relate task performance to cortical atrophy in the patient group
as a whole. The randomize tool in the FMRIB Software Library was used to run regression analyses
between the behavior of interest and patient cortical thickness values. Nonparametric permutations
(norte = 10,000) were run for each score of interest. Clusters that met a height threshold of p < 0.05 uncor-
rected with TFCE and a minimum of 25 adjacent voxels are reported. A 25-voxel threshold was chosen.
Statistical Methods
Linear mixed-effects models (LMEMs) were used to analyze the influence of variables of interest
on metaphor task performance. The lme4 package (Bates, Maechler, Bolker, & Walker, 2015) in R
(Version 3.3.1) was used for the following analyses. A theoretically motivated model was built for
each analysis.
Neurobiology of Language
481
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
RESULTS
Behavioral Results
Analysis of HC performance revealed 1 of the 19 participants performed more than 2.5 SDs
below the group mean (overall accuracy = 42.2% correct; 46.9% literal, 37.5% metaphor).
This participant’s data was removed from further analysis.
HC participants responded to two of the items with low accuracy, indicating that these
items were statistical outliers (for both items, only 8/18 participants answered correctly placing
these items more than 2.5 SDs below the other items’ mean accuracy). These two items were
removed from further analysis. The inclusion of these items, or of the outlier participant de-
scribed above, did not affect the patterns of results described below.
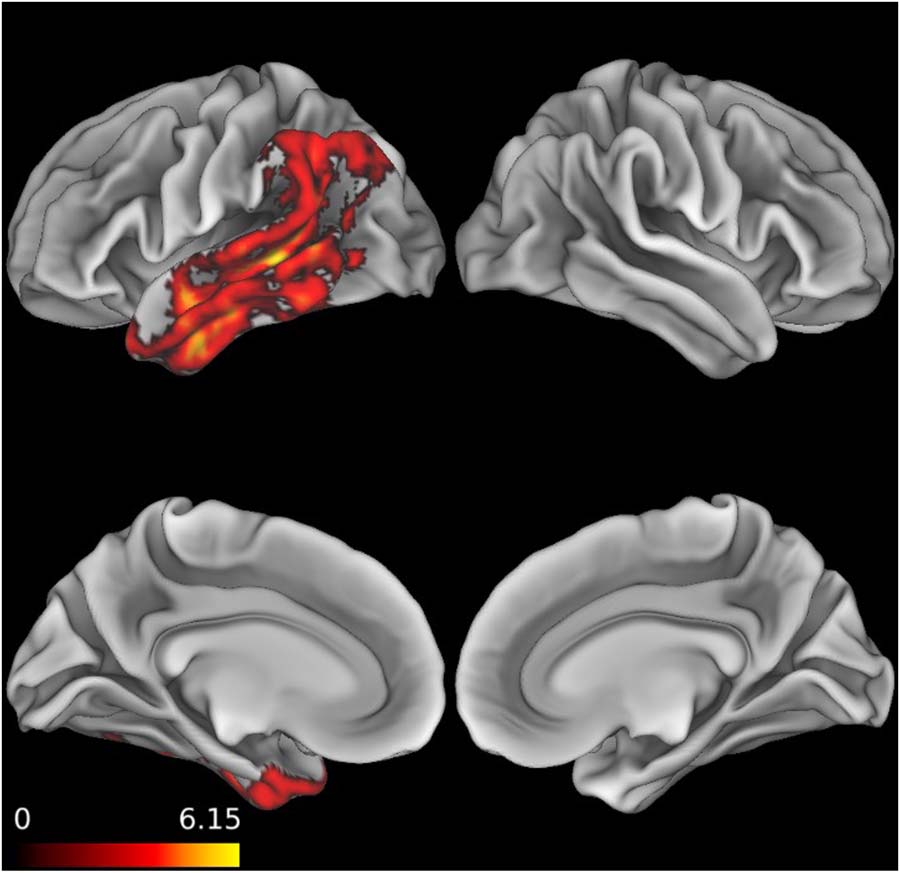
To examine group differences, LMEMs were used to analyze the relationships between
group (patients, HC), figurativeness (literal, metaphor), and accuracy. As random effects, par-
ticipant and item were included as intercepts, with a by-subject random slope included for the
effect of figurativeness and a by-group random slope included for item. These analyses
(Tables 4 and 5 and Figure 2) revealed a fixed effect of figurativeness, with literal sentences
receiving more accurate responses than metaphors ( p < 0.05, Cohen’s d = 0.23), a fixed effect
of group, with HCs outperforming LHND patients ( p < 0.01, d = 0.48), and an interaction
between group and figurativeness with the LHND group performing especially poorly on
the metaphor stimuli ( p < 0.05, d = 0.34). The difference between auditory (75.96%) and
motion (79.55%) accuracy was not significant. There were no interactions of modality with
group or figurativeness. There was no significant difference between event (80.00%) and entity
(76.88%) accuracy, and there were no interactions of source with group or figurativeness.
Error analysis
To illuminate the nature of comprehension failures, the proportion of each foil type selected
on incorrect trials was calculated. For metaphor trials, HCs (87%) and LHND (78%) showed a
strong literal bias, most often choosing the foil that provided the literal meaning to the target
metaphor rather than the correct metaphorical meaning. For literal trials, HCs (48%, 48%) and
LHND (46% vs. 38%) showed an even split between the semantic associate and the opposite
meaning foils.
Patients
Single case analyses
To examine different patterns of impaired comprehension, individual patient performance was
compared to the HC group using LMEMs. Subgroups based on behavior were then examined
for different patterns of atrophy. Accuracy was modeled as a function of group identity (patient
or HC), figurativeness (literal vs. metaphor trials), and education, and an interaction between
Table 4.
Percent accuracy by group and figurativeness
LHND (n = 13)
HC (n = 18)
Mean
Literal
75.0
93.58
85.79
Metaphor
50.51
84.26
70.75
Note. LHND = left-hemisphere neurodegeneration, HC = healthy control.
Mean
63.90
89.10
482
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
Table 5.
Fixed effects
Parameter
Intercept
Group (LHND)
Education
Figurativeness (Metaphor)
Group (LHND) × Figurativeness (Metaphor)
Estimate
0.533194
Standard Error
0.125328
t value
4.254***
−0.189489
0.054223
−3.495**
0.026019
0.007747
3.359**
−0.093171
−0.136316
0.040388
0.057948
−2.307*
−2.352*
Note. * p < 0.05; ** p < 0.01; *** p < 0.001. LHND = left-hemisphere neurodegeneration.
figurativeness and group, with the random effects of subject and item. The Holm method was
used to correct for multiple comparisons.
These analyses revealed that four patients (LHND2, LHND3, LHND4, LHND11) performed
indistinguishably from HCs overall and comprehended the literal and metaphor stimuli alike
( p’s > 0.05). Five patients displayed a general sentence-comprehension impairment (LHND5,
LHND6, LHND8, LHND10, LHND13), with impaired performance on the task overall and on
the literal stimuli (all p’s < 0.0001). For four patients, there was an interaction between group
(patient or HC) and figurativeness (all p’s < 0.05), showing evidence for a disproportionate
metaphor impairment, with worse metaphor comprehension performance than predicted by
their literal sentence performance (LHND1, LHND7, LHND9, LHND12; see Table 6). LHND7
could not complete scanning due to anxiety. While all were diagnosed with lvPPA at the time
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Metaphor task performance. Accuracy on literal (Lit) and metaphor (Met) trials by group
showing significant effects of group (HC > LHND, pag < 0.01), figurativeness (literal accuracy > metaphor
exactitud, pag < 0.05), and their interaction (with LHND especially impaired on metaphor accuracy,
p < 0.05). LHND = left-hemisphere neurodegeneration, HC = healthy control.
Neurobiology of Language
483
Impaired metaphor comprehension with left hemisphere degeneration
Table 6. Single cases with metaphor impairment
Patient
LHND1
LHND7
LHND9
LHND12
Literal
(32 max)
30
30
26
28
Literal
t value
0.03
0.03
−2.14
−1.05
Literal
p value
0.488
0.488
0.024
0.154
Metaphor
(30 max)
17
17
13
16
Metaphor
t value
−1.77
−1.77
−2.62
−1.98
Metaphor
p value
0.048
0.048
0.009
0.032
Cohen’s d
0.894
0.894
0.909
0.794
Note. p-values are one-tailed and compared to the control group’s performance using the Crawford-Howell test (Crawford & Howell, 1998) for case-control
comparisons. They provide a point estimate of the abnormality of each patient’s score. Cohen’s d values are the effect sizes for the differences between con-
ditions for each patient. LHND = left-hemisphere neurodegeneration.
of testing, LHND7 was subsequently diagnosed with Alzheimer’s disease, while the other
patients retained a diagnosis of lvPPA at the time of publication.
Neuropsychological measures and performance
In the patient group, task performance was related to performance on standard neuropsycholog-
ical assessments (Table 3). Item accuracy was modeled as a function of the fixed effects of figura-
tiveness and education, with random intercepts of subject and item and a by-subject random slope
for figurativeness. As the current study is not powered to consider all the neuropsychological tests’
influence on task performance in the same model, the neuropsychological tests were examined
one at a time for their ability to predict task performance by evaluating whether their inclusion
improved model fit.
These analyses revealed that while many tests were related to performance on the literal sen-
tences, including MMSE ( p < 0.05, d = 0.0695), reverse Digit Span ( p < 0.05, d = 0.1505), Naming
( p < 0.05, d = 0.2314), Animals ( p < 0.005, d = 0.0790), Philadelphia Verbal Learning Test (Libon,
Mattson, Glosser, & Kaplan, 1996) recall ( p < 0.05, d = 0.1005), Pyramids and Palm Trees (PPT;
Howard & Patterson, 1992) picture ( p < 0.05, d = 0.1408), PPT word ( p < 0.001, d = 0.1165),
and Complex Figure Test copy ( p < 0.01, d = 0.1141), only lexical fluency was related to metaphor
accuracy ( p < 0.01, d = 0.0570).
Neuroanatomical Results
ROI analyses
Relationships between cortical thickness and literal and metaphor performance were investigated
in three ROIs (LIFG, RIFG, LpMTG) motivated by previous fMRI results indicating that these
regions are engaged in comprehension of metaphors from the same stimulus set as used in this
study. Patient cortical thickness did not differ from that of HCs in the RIFG ( p > 0.91, re = 0.036),
while the LIFG showed a trend and a much larger effect size ( pag < 0.11, d = 0.591). Cortical thick-
ness in HCs did not differ between the RIFG and the LIFG ( p < 0.37), while there was a significant
difference in these areas in the patient group (t = 6.48, p < 0.0001, d = 0.322), with the LIFG
showing reduced thickness compared to the RIFG. No relationship was seen between overall
performance or literal performance and integrity of these areas in the patients ( p’s > 0.7). On
metaphor comprehension, LIFG thickness was significantly associated with accuracy ( pag <
0.05, d = 0.368), while RIFG thickness showed a trend ( p > 0.18).
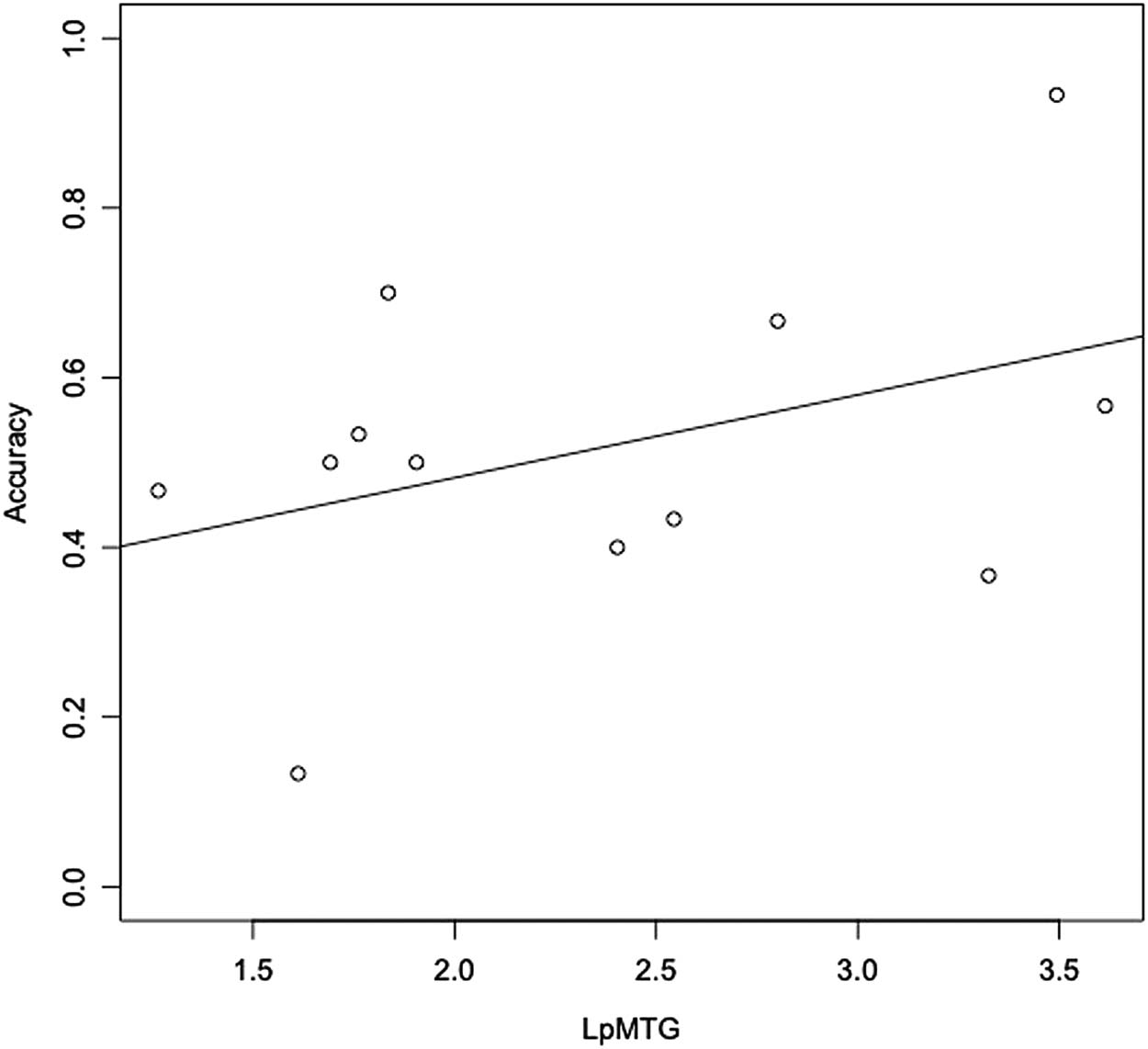
Patients showed significant atrophy relative to HCs in the LpMTG ( pag < 0.01, d = 1.174). For
overall task performance, the LpMTG showed a trend toward being associated with accuracy
Neurobiology of Language
484
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
( p < 0.098). Atrophy in the LpMTG did not relate to literal accuracy ( p > 0.3). On metaphor trials
(Cifra 3), LpMTG thickness was significantly related to accuracy ( pag < 0.01, d = 0.730).
Single subject analyses
Figure 4 and Supplementary Table 3 in the online supporting information depict single subject
atrophy heatmaps for the three participants with a disproportionate metaphor deficit and avail-
able neuroimaging.
Supplemental analyses: VBM within areas of degeneration
Exploratory voxel-based morphometry regressions were run relating patient performance on
literal and metaphor trials to the cortical thickness values within their atrophy mask (Figure 1).
For literal stimuli (Supplementary Figure 2 and Supplementary Table 4), atrophy in the superior
temporal gyrus and the angular gyrus related to poorer accuracy. For metaphor stimuli
(Supplementary Figure 3 and Supplementary Table 4), atrophy in the fusiform gyrus and the
middle temporal gyrus related to poorer accuracy.
DISCUSSION
Metaphor is used pervasively in communication and in cognition, shaping thought and influencing
behavior. How the brain mediates comprehension of metaphor is debated. To investigate the neural
bases of metaphor comprehension, we tested patients with neurodegenerative disease affecting the
left hemisphere and a group of demographically matched HC participants on their ability to resolve
the meaning of metaphoric and matched literal sentences. To summarize our results before discuss-
ing them in detail, our patients performed poorly on metaphor trials compared to HCs and compared
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Metaphor trial accuracy in patients as a function of LpMTG cortical thickness (mm).
LpMTG = left posterior middle temporal gyrus.
Neurobiology of Language
485
Impaired metaphor comprehension with left hemisphere degeneration
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
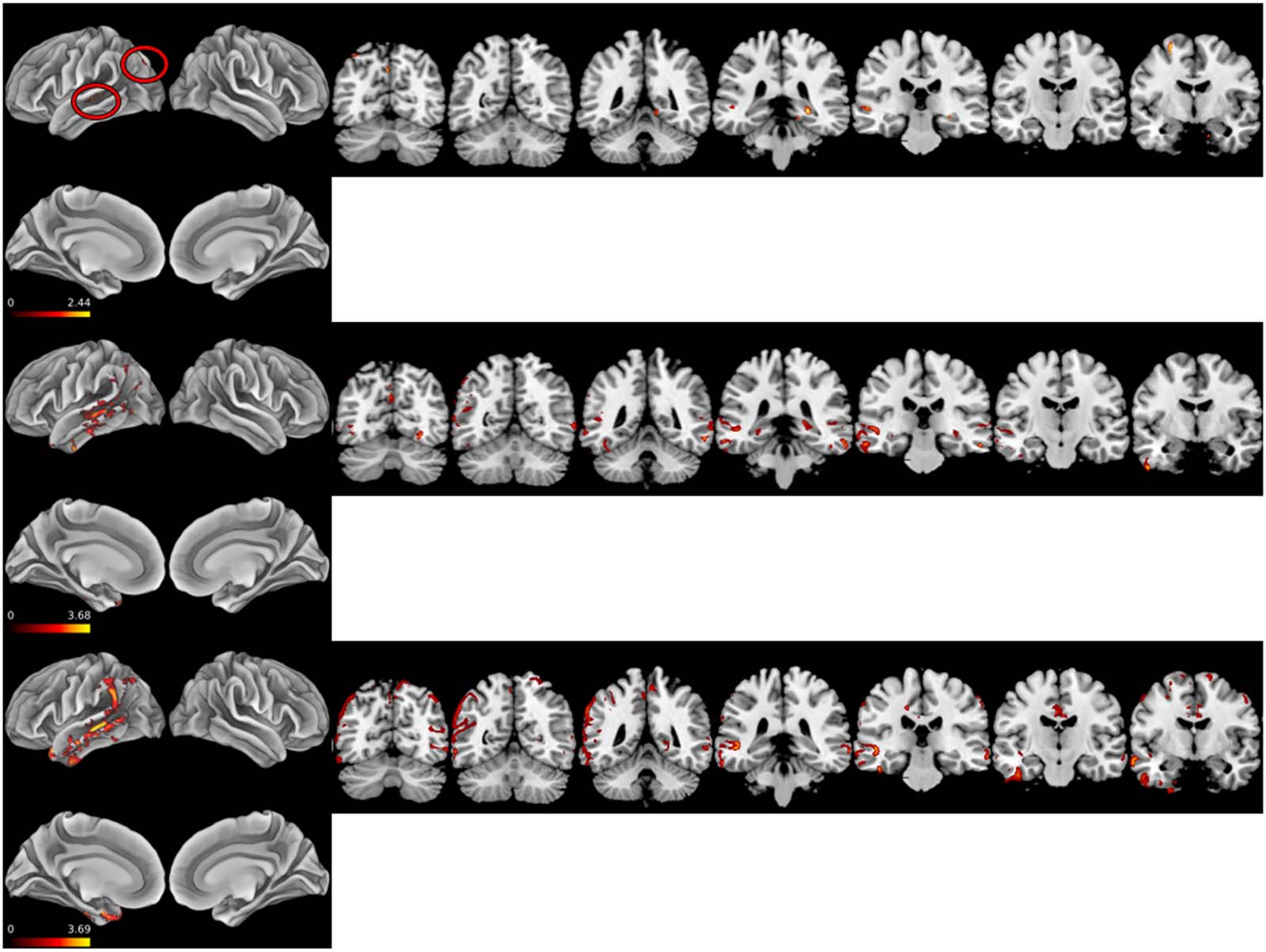
Figure 4. Z score heat maps for three patients with disproportionate metaphor impairment. Top: LHND1 Middle: LHND9. Bottom: LHND12.
LHND = left-hemisphere neurodegeneration.
to their own performance on literal trials. These results point to the important role that the left hemi-
sphere plays in understanding metaphor. Thinning of LIFG, left middle temporal gyrus, and left su-
perior temporal gyrus, areas implicated in previous work as supporting metaphor comprehension,
was associated with poor metaphor performance in some of our patients here. The results also show
that metaphor processing can be disproportionately impaired compared to literal sentence compre-
hension, suggesting that metaphor comprehension deficits could be a sensitive measure of cognitive
change in neurodegenerative disease, revealing impairments before literal language is impacted.
The variability in previously proposed neural substrates for metaphor comprehension present
in the literature may have risen from relative lack of control of psycholinguistic properties of
sentences used to test brain-behavior relationships (Cardillo et al., 2010; Citron & Goldberg,
2014; Schmidt, Kranjec, Cardillo, & Chatterjee, 2010). By measuring and balancing literal and
metaphor stimuli on various properties impacting comprehension difficulty—number of charac-
ters, words, and content words, average frequency and concreteness of content words, and
familiarity, valence, and a measure of semantic processing speed of sentences—the current study
attempted to avoid these difficulties and provide a more balanced contrast between literal and
metaphor sentences.
The patients’ impaired metaphor task performance cannot be explained as following from
general cognitive impairment. While analyses relating patient task performance to their neuro-
psychological test performance are exploratory given the relatively small sample size, patients’
neuropsychological profiles were mostly not associated with metaphor deficits. The patients’
performance on literal trials related to many neuropsychological test scores, including tests of
Neurobiology of Language
486
Impaired metaphor comprehension with left hemisphere degeneration
overall cognition, working memory, episodic memory, and semantic memory. It makes sense that
patients struggling to understand simple literal sentences would display impairments in multiple
cognitive domains. The lack of clear relationships observed here between performance on
metaphor trials and other aspects of cognition as measured through psychometric testing has
been noted in other patient studies of metaphor (Amanzio et al., 2008). This observation affirms
that traditional neuropsychological tests do not adequately capture metaphoric language deficits.
We agree with the suggestion (Rapp & Wild, 2011) that nonliteral language should be included in
standard neuropsychological assessment batteries.
The observation raises the question of why the patients struggle on metaphor trials. One
possibility is that the inhibitory demands of metaphor comprehension are difficult for patients.
To resolve the meaning of a metaphor, the literal sense of the sentence or features of the source
term must be inhibited in order to identify the correct figurative sense of the word and sentence
(Gernsbacher & Robertson, 1999; Papagno, 2001). The error analysis suggests that this inhibition
is difficult for healthy participants. On incorrect trials, the literal sense was most often endorsed by
HCs. For patients too, the literal foil was most often chosen on incorrect trials. The patients failed
more often here than HCs, suggesting greater difficulty with the inhibitory demands of metaphor
trials. A failure of inhibitory control also accords with the observation that lexical fluency—an
index of executive function—was correlated in this population with metaphor comprehension
accuracy. Successfully resolving the meaning of a metaphorical sentence also requires greater
flexibility as features and properties of one domain are applied and compared to another.
Cognitive flexibility is often diminished with general atrophy (Eslinger, Moore, Anderson, &
Grossman, 2011; Kehagia, Barker, & Robbins, 2010; Swartz, Stuss, Gao, & Black, 2008) and this
association may be contributing to the patients’ deficits.
Our results provide important evidence for left-hemisphere mediation of metaphor. In this
study, patients with neurodegeneration restricted to the left hemisphere displayed deficits in
understanding moderately familiar metaphors. Functional imaging evidence also implicates
areas in the left hemisphere used by the healthy brain in metaphor comprehension (Bohrn
et al., 2012; Rapp et al., 2012; Yang, 2014). The current study and other patient studies
(Cardillo et al., 2018; Gagnon et al., 2003; Ianni et al., 2014; Tompkins, 1990) provide evidence
for the importance of left-hemisphere regions for normal metaphor comprehension. Our ana-
tomic analyses included a targeted ROI analysis, an analysis within areas of group degenera-
tion, and a subgroup brain-behavior analysis of patients based on their patterns of performance.
For the ROI analysis, metaphor comprehension related to thickness of LpMTG and LIFG.
The left temporal lobe and LIFG are linked to semantic ambiguity resolution in literal language
(Zempleni, Renken, Hoeks, Hoogduin, & Stowe, 2007; Davis et al., 2007). When words have
multiple senses, or when multiple concepts can resolve the meaning of an ambiguous sentence,
these left hemisphere structures are implicated in successful meaning resolution. This ability
shares similarities to the challenge of resolving the meaning of a metaphor, a sentence with
multiple possible meanings to be resolved, and indeed similar brain regions are implicated in
supporting these abilities.
The LpMTG and the LIFG were related to metaphor comprehension deficits, consistent with
previous neuroimaging studies of metaphor (Cardillo et al., 2012; Chen et al., 2008; Lee &
Dapretto, 2006; Schmidt & Seger, 2009; Yang et al., 2010) and suggesting an important role for
these regions. While such a relationship was absent in RIFG, this may be because the current pa-
tient sample was not suitable for testing its role, as the patients show atrophy in LIFG but not RIFG.
The LpMTG has been linked to demands of semantic processing more generally (e.g., Noonan,
Jefferies, Visser, & Lambon Ralph, 2013).
Neurobiology of Language
487
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
The single-subject atrophy maps document the relationship between the left temporal lobe and
metaphor trial accuracy broadly. All three show relationships between LpMTG integrity and metaphor
trial performance. Interestingly, all three display a relationship between medial temporal lobe atrophy
and metaphor performance. Metaphor task performance may be a sensitive measure of cognitive de-
cline in neurodegenerative diseases that affect the medial temporal lobe such as Alzheimer’s disease.
When considering anatomy and neuropsychological profile and their relation to successful
metaphor comprehension, there may be “many routes to failure.” As the network of brain regions
that support metaphor processing is delineated, it is likely that disruptions to any nodes of this
network, or the connections between them, could lead to impairment. Metaphor processing is
a complex cognitive ability that depends on many subdomains of cognition including semantic
memory, working memory, (semantic) executive demands, inhibition, abstract thinking, and
cognitive flexibility. It is possible that subtle disruptions to any of these subdomains can lead
to impaired metaphor comprehension.
Limitations of the current study include the sample sizes of the participant groups. The current
results should be replicated with larger groups of participants. A priori power analyses were not
completed. We tested as many patients as possible. Given the relatively small sample size in the
current study and the difficulty of recruiting such patients, we elected to limit our ROI analyses to
a few strongly motivated areas. Analyses relating patient task performance to neuropsychological
assessment should be examined in larger populations. HC participants were closely matched to
the patients demographically, and their behavioral data was normally distributed, but a larger
comparison group would strengthen the confidence in our findings. Despite these limitations,
robust group differences were observed, and strong relationships were detailed between anatomy
and metaphor task performance.
The current study provides evidence that metaphor processing can be disproportionately
impaired compared to literal sentence comprehension. The evaluation of metaphor processing
may provide a more sensitive assessment of the earliest cognitive changes in neurodegenerative
disease. Tests of metaphor may reveal impairments before literal language is impacted. While the
cognitive consequences of focal lesions are observed rapidly, neurodegenerative disease
processes are progressive and are often quite subtle in the earliest stages. More sensitive measures
of cognitive change are needed as screening instruments and as outcome measures in the study of
neurodegenerative disease. Metaphor processing, with its distributed neural support and cogni-
tive complexity may be sensitive to early anatomical and cognitive changes.
In conclusion, we show that patients can display a disproportionate deficit in metaphor com-
prehension, compared to their own performance on literal sentence comprehension and to HC
performance. The left hemisphere plays an important role in metaphor processing.
FUNDING INFORMATION
Anjan Chatterjee, NIH, Award ID: R01DC012511. Nathaniel Bloem Klooster, NIH, Award ID:
T32 HD071844.
AUTHOR CONTRIBUTIONS
Nathaniel Klooster analyzed the data, created figures, and wrote the manuscript. Marguerite
McQuire designed the study, created stimuli, and collected data. Murray Grossman oversaw
experimental and neuropsychological data collection and contributed to manuscript writing.
Neurobiology of Language
488
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
Corey McMillan created figures and contributed to manuscript writing. Anjan Chatterjee
designed the study and contributed to manuscript writing. Eileen Cardillo designed the study,
created stimuli, and contributed to manuscript writing.
REFERENCES
Amanzio, M., Geminiani, G., Leotta, D., & Cappa, S. (2008).
Metaphor comprehension in Alzheimer’s disease: Novelty matters.
Brain and Language, 107(1), 1–10. DOI: https://doi.org/10.1016/j.
bandl.2007.08.003, PMID: 17897706
Armstrong, B. C., Watson, C. E., & Plaut, D. C. (2012). SOS! An
algorithm and software for the stochastic optimization of stimuli.
Behavior Research Methods, 44(3), 675–705. DOI: https://doi.
org/10.3758/s13428-011-0182-9, PMID: 22351612
Bambini, V., Gentili, C., Ricciardi, E., Bertinetto, P. M., & Pietrini,
P. (2011). Decomposing metaphor processing at the cognitive
and neural level through functional magnetic resonance imaging.
Brain Research Bulletin, 86(3–4), 203–216. DOI: https://doi.org/
10.1016/j.brainresbull.2011.07.015, PMID: 21803125
Barry, C. L., Brescoll, V. L., Brownell, K. D., & Schlesinger, M.
(2009). Obesity metaphors: How beliefs about the causes of
obesity affect support for public policy. The Milbank Quarterly,
87(1), 7–47. DOI: https://doi.org/10.1111/j.1468-0009.2009.
00546.x, PMID: 19298414, PMCID: PMC2879183
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical
Software, 67(1), 1–48. DOI: https://doi.org/10.18637/jss.v067.i01
Beger, A., & Jäkel, O. (2015). The cognitive role of metaphor in
teaching science: Examples from physics, chemistry, biology,
psychology and philosophy. Philosophical Inquiries, 3(1), 89–112.
DOI: https://doi.org/10.4454/philinq.v3i1.116
Bohrn, I. C., Altmann, U., & Jacobs, A. M. (2012). Looking at the
brains behind figurative language—A quantitative meta-analysis
of neuroimaging studies on metaphor, idiom, and irony processing.
Neuropsychologia, 50(11), 2669–2683. DOI: https://doi.org/
10.1016/j.neuropsychologia.2012.07.021, PMID: 22824234
Bottini, G., Corcoran, R., Sterzi, R., Paulesu, E., Schenone, P.,
Scarpa, P., … Frith, D. (1994). The role of the right hemisphere
in the interpretation of figurative aspects of language: A positron
emission tomography activation study. Brain, 117(6), 1241–1253.
DOI: https://doi.org/10.1093/brain/117.6.1241, PMID: 7820563
Bowes, A., & Katz, A. (2015). Metaphor creates intimacy and tem-
porarily enhances theory of mind. Memory & Cognition, 43(6),
953–963. DOI: https://doi.org/10.3758/s13421-015-0508-4,
PMID: 25762491
Brownell, H. H., Simpson, T. L., Bihrle, A. M., Potter, H. H., &
Gardner, H. (1990). Appreciation of metaphoric alternative word
meanings by left and right brain-damaged patients. Neuropsychologia,
28(4), 375–383. DOI: https://doi.org/10.1016/0028-3932(90)90063-T,
PMID: 1692980
Cardillo, E. R., McQuire, M., & Chatterjee, A. (2018). Selective meta-
phor impairments after left, not right, hemisphere injury. Frontiers in
Psychology, 9, 2308. DOI: https://doi.org/10.3389/fpsyg.2018.
02308, PMID: 30559690, PMCID: PMC6286990
Cardillo, E. R., Schmidt, G. L., Kranjec, A., & Chatterjee, A. (2010).
Stimulus design is an obstacle course: 560 matched literal and
metaphorical sentences for testing neural hypotheses about
metaphor. Behavior Research Methods, 42(3), 651–664. DOI:
https://doi.org/10.3758/BRM.42.3.651, PMID: 20805587,
PMCID: PMC2952404
Cardillo, E. R., Watson, C., & Chatterjee, A. (2016). Stimulus needs
are a moving target: 240 additional matched literal and
metaphorical sentences for testing neural hypotheses about meta-
phor. Behavior Research Methods, 49(2), 471–483. DOI: https://
doi.org/10.3758/s13428-016-0717-1, PMID: 26956680
Cardillo, E. R., Watson, C. E., Schmidt, G. L., Kranjec, A., &
Chatterjee, A. (2012). From novel to familiar: Tuning the brain
for metaphors. NeuroImage, 59(4), 3212–3221. DOI: https://
doi.org/10.1016/j.neuroimage.2011.11.079, PMID: 22155328,
PMCID: PMC3288556
Cardoso, S., Silva, D., Maroco, J., de Mendonça, A., & Guerreiro,
M. (2014). Non-literal language deficits in mild cognitive impair-
ment. Psychogeriatrics, 14(4), 222–228. DOI: https://doi.org/
10.1111/psyg.12101, PMID: 25495083
Chen, E., Widick, P., & Chatterjee, A. (2008). Functional–anatomical
organization of predicate metaphor processing. Brain and Language,
107(3), 194–202. DOI: https://doi.org/10.1016/j.bandl.2008.06.007,
PMID: 18692890, PMCID: PMC2638768
Citron, F. M., & Goldberg, A. E. (2014). Metaphorical sentences are
more emotionally engaging than their literal counterparts. Journal
of Cognitive Neuroscience, 26(11), 2585–2595. DOI: https://doi.
org/10.1162/jocn_a_00654, PMID: 24800628
Conrad, E. C., Humphries, H., & Chatterjee, A. (2019). Attitudes toward
cognitive enhancement: The role of metaphor and context. AJOB
Neuroscience, 10(1), 35–47. DOI: https://doi.org/10.1080/
21507740.2019.1595771, PMID: 31070552
Crawford, J. R., & Howell, D. C. (1998). Comparing an individual’s
test score against norms derived from small samples. The Clinical
Neuropsychologist, 12(4), 482–486. DOI: https://doi.org/
10.1076/clin.12.4.482.7241
Davis, M. H., Coleman, M. R., Absalom, A. R., Rodd, J. M.,
Johnsrude, I. S., Matta, B. F., … Menon, D. K. (2007).
Dissociating speech perception and comprehension at reduced
levels of awareness. Proceedings of the National Academy of
Sciences, 104(41), 16032–16037. DOI: https://doi.org/10.1073/
pnas.0701309104, PMID: 17938125, PMCID: PMC2042157
Diaz, M. T., Barrett, K. T., & Hogstrom, L. J. (2011). The influence of sen-
tence novelty and figurativeness on brain activity. Neuropsychologia,
49, 320–330. DOI: https://doi.org/10.1016/j.neuropsychologia.
2010.12.004, PMID: 21146553, PMCID: PMC3034783
Elmore, K. C., & Luna-Lucero, M. (2017). Light bulbs or seeds? How
metaphors for ideas influence judgments about genius. Social
Psychological and Personality Science, 8(2), 200–208. DOI: https://
doi.org/10.1177/1948550616667611
Eslinger, P. J., Moore, P., Anderson, C., & Grossman, M. (2011).
Social cognition, executive functioning, and neuroimaging corre-
lates of empathic deficits in frontotemporal dementia. The Journal
of Neuropsychiatry and Clinical Neurosciences, 23(1), 74–82.
DOI: https://doi.org/10.1176/appi.neuropsych.23.1.74, PMID:
21304142, PMCID: PMC3641646
Fellows, L. K., Heberlein, A. S., Morales, D. A., Shivde, G., Waller, S.,
& Wu, D. H. (2005). Method matters: An empirical study of impact
in cognitive neuroscience. Journal of Cognitive Neuroscience, 17(6),
850–858. DOI: https://doi.org/10.1162/0898929054021139,
PMID: 15969904
Fernandino, L., Conant, L. L., Binder, J. R., Blindauer, K., Hiner, B.,
Spangler, K., … Desai, R. H. (2013). Where is the action? Action
sentence processing in Parkinson's disease. Neuropsychologia,
Neurobiology of Language
489
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
51(8), 1510–1517. DOI: https://doi.org/10.1016/j.neuropsychologia.
2013.04.008
Flusberg, S. J., Matlock, T., & Thibodeau, P. H. (2017). Metaphors for the
war (or race) against climate change. Environmental Communication,
11(6), 769–783. DOI: https://doi.org/10.1080/17524032.2017.1289111
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). “Mini-mental
state”: A practical method for grading the cognitive state of patients
for the clinician. Journal of Psychiatric Research, 12(3), 189–198.
DOI: https://doi.org/10.1016/0022-3956(75)90026-6, PMID:
1202204
Gagnon, L., Goulet, P., Giroux, F., & Joanette, Y. (2003). Processing of
metaphoric and non-metaphoric alternative meanings of words
after right- and left-hemispheric lesion. Brain and Language, 87(2),
217–226. DOI: https://doi.org/10.1016/S0093-934X(03)00057-9,
PMID: 14585291
Gernsbacher, M. A., & Robertson, R. R. (1999). The role of suppression
in figurative language comprehension. Journal of Pragmatics, 31(12),
1619–1630. DOI: https://doi.org/10.1016/S0378-2166(99)00007-7,
PMID: 25520540, PMCID: PMC4266403
Gibbs, R. W., Jr., & Franks, H. (2002). Embodied metaphor in
women’s narratives about their experiences with cancer. Health
Communication, 14(2), 139–165. DOI: https://doi.org/10.1207/
S15327027HC1402_1, PMID: 12046796
Gorno-Tempini, M. L., Hillis, A. E., Weintraub, S., Kertesz, A.,
Mendez, M., Cappa, S. F., … Manes, F. (2011). Classification of
primary progressive aphasia and its variants. Neurology, 76(11),
1006–1014. DOI: https://doi.org/10.1212/WNL.0b013e31821103e6,
PMID: 21325651, PMCID: PMC3059138
Hauser, D. J., & Schwarz, N. (2015). The war on prevention:
Bellicose cancer metaphors hurt (some) prevention intentions.
Personality and Social Psychology Bulletin, 41(1), 66–77. DOI:
https://doi.org/10.1177/0146167214557006, PMID: 25352114
Hilligoss, B. (2014). Selling patients and other metaphors: A discourse
analysis of the interpretive frames that shape emergency department
admission handoffs. Social Science & Medicine, 102, 119–128.
DOI: https://doi.org/10.1016/j.socscimed.2013.11.034, PMID:
24565149
Howard, D., & Patterson, K. (1992). Pyramid and palm trees: A test
of semantic access from pictures and words. Bury St. Edmunds,
UK: Thames Valley Test Company.
Ianni, G. R., Cardillo, E. R., McQuire, M., & Chatterjee, A. (2014).
Flying under the radar: Figurative language impairments in focal
lesion patients. Frontiers in Human Neuroscience, 8, 871. DOI: https://
doi.org/10.3389/fnhum.2014.00871, PMID: 25404906, PMCID:
PMC4217389
Kehagia, A. A., Barker, R. A., & Robbins, T. W. (2010).
Neuropsychological and clinical heterogeneity of cognitive impair-
ment and dementia in patients with Parkinson’s disease. The Lancet
Neurology, 9(12), 1200–1213. DOI: https://doi.org/10.1016/S1474-
4422(10)70212-X, PMID: 20880750
Lakoff, G., & Johnson, M. (1980). The metaphorical structure of the
human conceptual system. Cognitive Science, 4(2), 195–208. DOI:
https://doi.org/10.1207/s15516709cog0402_4
Lee, S. S., & Dapretto, M. (2006). Metaphorical vs. literal word
meanings: fMRI evidence against a selective role of the right
hemisphere. NeuroImage, 29, 536–544. DOI: https://doi.org/
10.1016/j.neuroimage.2005.08.003, PMID: 16165371
Lee, S. W., & Schwarz, N. (2014). Framing love: When it hurts to think
we were made for each other. Journal of Experimental Social
Psychology, 54, 61–67. DOI: https://doi.org/10.1016/j.jesp.2014.
04.007
Libon, D. J., Mattson, R. E., Glosser, G., & Kaplan, E. 1996. A nine-
word dementia version of the California Verbal Learning Test.
The Clinical Neuropsychologist, 10, 237–244. DOI: https://doi.
org/10.1080/13854049608406686
Mashal, N., & Faust, M. (2009). Conventionalisation of novel meta-
phors: A shift in hemispheric asymmetry. Laterality, 14(6), 573–589.
DOI: https://doi.org/10.1080/13576500902734645, PMID:
19253086
Mashal, N., Faust, M., & Hendler, T. (2005). The role of the right
hemisphere in processing nonsalient metaphorical meanings:
Application of principal components analysis to fMRI data.
Neuropsychologia, 43(14), 2084–2100. DOI: https://doi.org/
10.1016/j.neuropsychologia.2005.03.019, PMID: 16243053
Matlock, T. (2004). Fictive motion as cognitive simulation. Memory &
Cognition, 32(8), 1389–1400. DOI: https://doi.org/10.3758/
BF03206329, PMID: 15900932
Monetta, L., & Pell, M. D. (2007). Effects of verbal working memory
deficits on metaphor comprehension in patients with Parkinson’s
disease. Brain and Language, 101(1), 80–89. DOI: https://doi.org/
10.1016/j.bandl.2006.06.007, PMID: 16875726
Nelson, H. E., & O’Connell, A. (1978). Dementia: The estimation of
premorbid intelligence levels using the New Adult Reading Test.
Cortex, 14(2), 234–244. DOI: https://doi.org/10.1016/S0010-
9452(78)80049-5, PMID: 679704
Nerlich, B., & Jaspal, R. (2012). Metaphors we die by? Geoengineering,
metaphors, and the argument from catastrophe. Metaphor and
Symbol, 27(2), 131–147. DOI: https://doi.org/10.1080/10926488.
2012.665795
Noonan, K. A., Jefferies, E., Visser, M., & Lambon Ralph, M. A.
(2013). Going beyond inferior prefrontal involvement in seman-
tic control: Evidence for the additional contribution of dorsal
angular gyrus and posterior middle temporal cortex. Journal of
Cognitive Neuroscience, 25(11), 1824–1850. DOI: https://doi.
org/10.1162/jocn_a_00442, PMID: 23859646
Obert, A., Gierski, F., Calmus, A., Portefaix, C., Declercq, C.,
Pierot, L., & Caillies, S. (2014). Differential bilateral involvement
of the parietal gyrus during predicative metaphor processing: An
auditory fMRI study. Brain and language, 137, 112–119. DOI:
https://doi.org/10.1016/j.bandl.2014.08.002, PMID: 25193417
Papagno, C. (2001). Comprehension of metaphors and idioms in
patients with Alzheimer’s disease: A longitudinal study. Brain,
124(7), 1450–1460. DOI: https://doi.org/10.1093/brain/124.7.1450,
PMID: 11408339
Papagno, C., Lucchelli, F., Muggia, S., & Rizzo, S. (2003). Idiom
comprehension in Alzheimer’s disease: The role of the central
executive. Brain, 126(11), 2419–2430. DOI: https://doi.org/10.1093/
brain/awg243, PMID: 12902312
Rapp, A. M., Leube, D. T., Erb, M., Grodd, W., & Kircher, T. T. (2004).
Neural correlates of metaphor processing. Brain Research, 20,
395–402. DOI: https://doi.org/10.1016/j.cogbrainres.2004.03.017,
PMID: 15268917
Rapp, A. M., Leube, D. T., Erb, M., Grodd, W., & Kircher, T. T.
(2007). Laterality in metaphor processing: Lack of evidence from
functional magnetic resonance imaging for the right hemisphere
theory. Brain and Language, 100, 142–149. DOI: https://doi.org/
10.1016/j.bandl.2006.04.004, PMID: 16677700
Rapp, A. M., Mutschler, D. E., & Erb, M. (2012). Where in the brain
is nonliteral language? A coordinate-based meta-analysis of func-
tional magnetic resonance imaging studies. NeuroImage, 63(1),
600–610. DOI: https://doi.org/10.1016/j.neuroimage.2012.06.022,
PMID: 22759997
Rapp, A. M., & Wild, B. (2011). Nonliteral language in Alzheimer
dementia: A review. Journal of the International Neuropsy-
chological Society, 17(2), 207–218. DOI: https://doi.org/10.1017/
S1355617710001682, PMID: 21241530
Neurobiology of Language
490
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Impaired metaphor comprehension with left hemisphere degeneration
Reyes-Aguilar, A., Valles-Capetillo, E., & Giordano, M. (2018). A
quantitative meta-analysis of neuroimaging studies of pragmatic
language comprehension: In search of a universal neural sub-
strate. Neuroscience, 395, 60–88. DOI: https://doi.org/10.1016/
j.neuroscience.2018.10.043, PMID: 30414881
Robins, S., & Mayer, R.E. (2000). The metaphor framing effect:
Metaphorical reasoning about text-based dilemmas. Discourse
P r o c e s s e s , 3 0 , 5 7 – 8 6 . D O I : h t t p s : / / d o i . o r g / 1 0 . 1 2 0 7 /
S15326950dp3001_03
Roncero, C., & de Almeida, R. G. (2014). The importance of being apt:
Metaphor comprehension in Alzheimer’s disease. Frontiers in
Human Neuroscience, 8, 973. DOI: https://doi.org/10.3389/fnhum.
2014.00973, PMID: 25520642, PMCID: PMC4251318
Saban, A. (2006). Functions of metaphor in teaching and teacher
education: A review essay. Teaching Education, 17(4), 299–315.
DOI: https://doi.org/10.1080/10476210601017386
Schmidt, G. L., Kranjec, A., Cardillo, E. R., & Chatterjee, A. (2010).
Beyond laterality: A critical assessment of research on the neural
basis of metaphor. Journal of the International Neuropsychological
S o c i e t y , 1 6 ( 1 ) , 1 – 5 . D O I : h t t p s : / / d o i . o r g / 1 0 . 1 0 1 7 /
S1355617709990543, PMID: 19765354, PMCID: PMC2926935
Schmidt, G. L., & Seger, C. A. (2009). Neural correlates of metaphor
processing: The roles of figurativeness, familiarity and difficulty.
Brain and Cognition, 71(3), 375–386. DOI: https://doi.org/10.1016/
j.bandc.2009.06.001, PMID: 19586700, PMCID: PMC2783884
Smith, S. M., & Nichols, T. E. (2009). Threshold-free cluster enhance-
ment: Addressing problems of smoothing, threshold dependence
and localisation in cluster inference. NeuroImage, 44(1), 83–98.
DOI: https://doi.org/10.1016/j.neuroimage.2008.03.061, PMID:
18501637
Sopory, P., & Dillard, J. P. (2002). The persuasive effects of metaphor: A
meta-analysis. Human Communication Research, 28(3), 382–419.
DOI: https://doi.org/10.1111/j.1468-2958.2002.tb00813.x
Steen, G. J., Dorst, L., Herrmann, B., Kaal, A., Krennmayr, T., &
Pasma, T. (2010). A method for linguistic metaphor identification:
From MIP to MIPVU. Amsterdam/Philadelphia: John Benjamins.
DOI: https://doi.org/10.1075/celcr.14
Swartz, R. H., Stuss, D. T., Gao, F., & Black, S. E. (2008).
Independent cognitive effects of atrophy and diffuse subcortical
and thalamico-cortical cerebrovascular disease in dementia. Stroke,
39(3), 822–830. DOI: https://doi.org/10.1161/STROKEAHA.107.
491936, PMID: 18258840
Thibodeau, P. H., & Boroditsky, L. (2011). Metaphors we think
with: The role of metaphor in reasoning. PLOS One, 6(2), e16782.
DOI: https://doi.org/10.1371/journal.pone.0016782, PMID:
21373643, PMCID: PMC3044156
Thibodeau, P. H., & Boroditsky, L. (2013). Natural language meta-
phors covertly influence reasoning. PLOS One, 8(1),
e52961. DOI: https://doi.org/10.1371/journal.pone.0052961,
PMID: 23301009, PMCID: PMC3534638
Tompkins, C. (1990). Knowledge and strategies for processing lexical
metaphor after right or left hemisphere brain damage. Journal of
Speech, Language, and Hearing Research, 33(2), 307–316. DOI:
https://doi.org/10.1044/jshr.3302.307
Tustison, N. J., Avants, B. B., Cook, P. A., Zheng, Y., Egan, A.,
Yushkevich, P. A., & Gee, J. C. (2010). N4ITK: Improved N3 bias
correction. IEEE Transactions on Medical Imaging, 29(6), 1310–1320.
DOI: https://doi.org/10.1109/TMI.2010.2046908, PMID: 20378467,
PMCID: PMC3071855
Tustison, N. J., Cook, P. A., Klein, A., Song, G., Das, S. R.,
Duda, J. T., … Avants, B. B. (2014). Large-scale evaluation of ANTs
and FreeSurfer cortical thickness measurements. NeuroImage,
99, 166–179. DOI: https://doi.org/10.1016/j.neuroimage.2014.
05.044, PMID: 24879923
Van Rijn-van Tongeren, G. W. (1997). Metaphors in medical texts
(Vol. 8). Amsterdam: Rodopi.
Van Stee, S. K. (2018). Meta-analysis of the persuasive effects of
metaphorical vs. literal messages. Communication Studies, 69(5),
1–22. DOI: https://doi.org/10.1080/10510974.2018.1457553
Winner, E., & Gardner, H. (1977). The comprehension of metaphor
in brain-damaged patients. Brain, 100(4), 717–729. DOI: https://
doi.org/10.1093/brain/100.4.717, PMID: 608117
Yang, F. G., Fuller, J., Khodaparast, N., & Krawczyk, D. C. (2010).
Figurative language processing after traumatic brain injury in adults:
A preliminary study. Neuropsychologia, 48(7), 1923–1929. DOI:
https://doi.org/10.1016/j.neuropsychologia.2010.03.011, PMID:
20230844
Yang, J. (2014). The role of the right hemisphere in metaphor com-
prehension: A meta-analysis of functional magnetic resonance
imaging studies. Human Brain Mapping, 35(1), 107–122. DOI:
https://doi.org/10.1002/hbm.22160, PMID: 22936560, PMCID:
PMC6868953
Zaidel, E., Kasher, A., Soroker, N., & Batori, G. (2002). Effects of
right and left hemisphere damage on performance of the “Right
Hemisphere Communication Battery.” Brain and Language, 80(3),
510–535. DOI: https://doi.org/10.1006/brln.2001.2612, PMID:
11896655
Zempleni, M. Z., Renken, R., Hoeks, J. C., Hoogduin, J. M., &
Stowe, L. A. (2007). Semantic ambiguity processing in sentence
context: Evidence from event-related fMRI. NeuroImage, 34(3),
1270–1279. DOI: https://doi.org/10.1016/j.neuroimage.2006.
09.048, PMID: 17142061
Neurobiology of Language
491
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
7
4
1
8
6
7
7
7
6
n
o
_
a
_
0
0
0
2
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3