INVESTIGACIÓN
MRI network progression in mesial temporal lobe
epilepsy related to healthy brain architecture
Victoria L. Morgan 1∗
, Graham W. Johnson2, Leon Y. Cai2, Bennett A. Landman3,
Kurt G. Schilling1, Dario J. Englot4, Baxter P. Rogers1, and Catie Chang3
1Institute of Imaging Science, Department of Radiology and Radiological Sciences, Vanderbilt University Medical Center,
Nashville, TN, EE.UU
2Department of Biomedical Engineering, Vanderbilt University, Nashville, TN, EE.UU
3Department of Electrical Engineering and Computer Science, Vanderbilt University, Nashville, TN, EE.UU
4Department of Neurological Surgery, Vanderbilt University Medical Center, Nashville, TN, EE.UU
un acceso abierto
diario
Palabras clave: Network connectivity, Focal epilepsy, Functional MRI, Diffusion MRI
ABSTRACTO
We measured MRI network progression in mesial temporal lobe epilepsy (mTLE) patients as a
function of healthy brain architecture. Resting-state functional MRI and diffusion-weighted
MRI were acquired in 40 unilateral mTLE patients and 70 healthy controls. Data were used
to construct region-to-region functional connectivity, structural connectivity, and streamline
length connectomes per subject. Three models of distance from the presumed seizure focus
in the anterior hippocampus in the healthy brain were computed using the average
connectome across controls. A fourth model was defined using regions of transmodal (más alto
cognitive function) to unimodal (perceptual) networks across a published functional gradient
in the healthy brain. These models were used to test whether network progression in patients
increased when distance from the anterior hippocampus or along a functional gradient in the
healthy brain decreases. Results showed that alterations of structural and functional networks
in mTLE occur in greater magnitude in regions of the brain closer to the seizure focus based
on healthy brain topology, and decrease as distance from the focus increases over duration of
enfermedad. En general, this work provides evidence that changes across the brain in focal epilepsy
occur along healthy brain architecture.
RESUMEN DEL AUTOR
In patients with focal epilepsy, seizures originate in the focus and propagate across the brain.
Over years of duration of disease, these repeated seizures lead to network reorganization and
ruptura. We hypothesized that these changes occur along a framework that could be
identified through healthy brain architecture, with the greatest changes occurring closest to
the seizure focus and decreasing as distance from the focus increases. In this work we
detected this pattern of change in functional and structural networks when distance to the
focus was measured by functional and structural connectivity, respectivamente. En general, this work
presents a framework of spatiotemporal network progression over duration of disease related
to the seizure focus and healthy brain architecture that may predict individual network
evolution in focal epilepsy.
Citación: morgan, V. l., Johnson, GRAMO. w.,
Cai, l. y., Landman, B. A., Schilling,
k. GRAMO., Englot, D. J., Rogers, B. PAG., &
Chang, C. (2021). MRI network
progression in mesial temporal lobe
epilepsy related to healthy brain
architecture. Neurociencia en red,
5(2), 434–450. https://doi.org/10.1162
/netn_a_00184
DOI:
https://doi.org/10.1162/netn_a_00184
Recibió: 1 Noviembre 2020
Aceptado: 11 Enero 2021
Autor correspondiente:
Victoria L. morgan
victoria.morgan@vanderbilt.edu
Editor de manejo:
Álex Amueblado
Derechos de autor: © 2021
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
Focal epilepsy:
When seizures originate in one
region of the brain.
Structural connectivity:
Brain region interaction measured by
integrity of diffusion of water across
white matter using MRI.
Conectividad funcional:
Temporal correlations of spontaneous
low-frequency blood oxygenation
signal measured by MRI.
INTRODUCCIÓN
Approximately 60%–70% of focal epilepsy patients are drug-refractory (Laxer et al., 2014;
ángel, 2016). Many of these patients experience seizures for decades, prior to surgical or
other intervention (ángel, 2016). Evidence suggests that epilepsy is not a static condition, y
that this progression can have significant clinical implications. A recent review concluded
that patients with shorter epilepsy duration are more likely to be seizure free after surgical
resection (Bjellvi et al., 2019). Similarmente, shorter duration of disease predicts better seizure
outcome according to several prediction algorithms (Gracia et al., 2015; Jehi et al., 2015;
Dugan et al., 2017). This implies that our current surgical treatment strategies do not success-
fully consider the issue of temporal progression in epilepsy. Focal epilepsy also has spatially
extensive effects on the brain evidenced by widespread functional (Englot et al., 2016) y
structural (Bonilha et al., 2012; Besson et al., 2014) network connectivity changes measured
by MRI. Network connectivity has been shown to be related to seizure outcome after surgery
(Morgan et al., 2017; Gleichgerrcht et al., 2018; Taylor et al., 2018; Morgan et al., 2019) y
correlated with duration of disease (Morgan et al., 2011; Haneef et al., 2015; Wang y cols., 2017;
Owen et al., 2020). De este modo, the progression of focal epilepsy includes a complex spatiotemporal
interaction of widespread brain networks, and this evolution of network architecture may play
a significant role in the efficacy of treatment.
To be able to utilize the spatiotemporal network information as a biomarker of treatment
outcome prediction, it is important to model these network changes in the context of a large-
scale overarching organization of the brain. This would (a) allow the ability to predict changes
in an individual patient to facilitate more accurate outcome predictions and (b) improve the
understanding of how focal epilepsy progresses over time and space across the brain. Varios
topological structures have been proposed to explain variability in brain network connectivity
in healthy controls (Suárez et al., 2020). Por ejemplo, some relate functional relationships
across the brain in terms of structural connectivity (van den Heuvel et al., 2009), commu-
nication measures (Goñi et al., 2014; Fukushima et al., 2018), microstructural characteris-
tics (Vázquez-Rodríguez et al., 2020), or geometric properties (Betzel et al., 2016;
investigated the func-
Oligschläger et al., 2017).
tional connectivity differences in patients as they relate to geodesic distance between cortical
vertices (Larivière et al., 2020). En general, they found increases in short-range and decreases in
long-range functional connectivity in the temporal and frontal lobes mediated by white mat-
ter microstructural changes measured by diffusion MRI. Además, reductions in short-range
connectivity were related to better surgical outcomes. These findings support that pathological
changes in focal epilepsy are not random, but rather progress along an identifiable framework
and may have clinical significance.
In temporal lobe epilepsy, Larivière et al.
In this work we investigated brain organization models specifically related to unilateral
mesial temporal lobe epilepsy (mTLE) as a framework on which MRI functional and structural
connectivity changes occur over time and space in these patients. In mTLE, seizures originate
within one hippocampus and propagate across the brain (ángel, 2001). Por lo tanto, our pro-
posed models were based on the general assumption that greater network changes will occur
“closer” to the hippocampus based on some topology, and then spread outward. We presented
two separate investigations. Primero, we limited our network to regional connections to the hip-
pocampi (presumed seizure focus). Segundo, we investigated whole-brain connectomes with
each region connected to all other regions in the brain. If functional and structural network
evolution in mTLE can be modeled using a framework defined in the healthy brain, a similar
approach may be used to predict progression in other focal epilepsies.
Neurociencia en red
435
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
Mesa 1. Participant characteristics.
Right mTLE
(norte = 29)
Left mTLE
(norte = 11)
Control S
(norte = 70)
M/F
Age (años: mean ± SD, mín., máximo)
Epilepsy duration (años: mean ± SD, mín., máximo)
MRI HS or hippocampal gliosis on pathology (norte, %)
Lateralizing PET hypometabolism (norte, %)
Localizing video scalpEEG of ictal events (norte, %)
Nota. M = male; F = female; HS = hippocampal sclerosis including T2 hyperintensity.
14/15
40.1 ± 10.7, 23, 62
21.2 ± 14.4, 2, 50
27, 93%
24, 83%
25, 86%
8, 73%
10, 91%
8/3
36.6 ± 16.4, 18, 68
20.6 ± 16.2, 3, 46
13, 100%
37/33
38.0 ± 13.7, 18, 71
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Electroencephalography (EEG):
The measurement of electrical
activity in your brain using
noninvasive electrodes attached to
your scalp, but can also use invasive
probes.
Positron emission tomography (PET):
Noninvasive functional imaging
measure of metabolic processes in
the body using radioactive tracers.
Hippocampal sclerosis:
Anatomic hallmark of mesial
temporal lobe epilepsy characterized
by severe neuronal cell loss and
gliosis in the hippocampus.
MATERIALES Y MÉTODOS
Participantes
Los participantes fueron 40 drug-refractory unilateral mTLE patients (29 bien, 11 izquierda) prior to surgical
intervención (Mesa 1). Diagnoses were based on long-term video EEG of ictal events local-
izing to anterior/mesial temporal regions, unilateral mesial temporal lobe hypometabolism
on positron emission tomography (PET), hippocampal sclerosis on standard MRI, and seizure
semiology consistent with mTLE. Two patients did not have hippocampal sclerosis on clinical
MRI or pathology-confirmed gliosis in the resected tissue. One of these had resection with
an Engel I-d outcome 1 year postsurgery, while one did not have resection. Exclusion criteria
Además, 70 saludable
included structural abnormalities outside the mesial temporal lobe.
control participants were enrolled. The protocol was approved by the Vanderbilt University
Institutional Review Board. All participants gave informed consent.
Imaging
Imaging was identical for all participants and was acquired on a 3T MRI scanner using a 32-
channel head coil. Cardiac and respiratory fluctuations were recorded at 500 Hz using the
scanner-integrated pulse oximeter and respiratory belt. The following images were acquired:
(a) high-resolution T1-weighted MRI for intersubject normalization and regional and tissue
segmentation (1 mm × 1 mm × 1 mm), (b) T1-weighted MRI acquired in the same slice
∗
orientation as the functional images (1 mm × 1 mm × 3.5 mm with 0.5-mm gap), (C) T2
–
weighted functional MRI (resonancia magnética funcional) BOLD MRI at rest with eyes closed (34 axial slices, echo time =
35 EM, repetition time = 2 segundo, 3 mm × 3 mm × 3.5 mm with a 0.5-mm gap, 10 minutos), y
(d) diffusion-weighted MRI (DWI) for structural connectivity (50 slices, 2.5 mm × 2.5 mm ×
2.5 mm, 92 directions, b = 0, 1, 600 s/mm2).
Connectomes
Un total de 109 regions of interest were identified on the 1-mm3 T1-weighted images, en-
cluding 54 in each hemisphere and the bilateral brainstem. Primero, the Multi-Atlas algorithm
(Asman & Landman, 2013; Huo et al., 2016) was used to identify cortical and subcortical re-
gions across the brain. Since this atlas did not parcellate subregions of the hippocampus, nosotros
then used the FreeSurfer 6 suite (pescado, 2012) to identify hippocampal subfields. These were
used to form composite anterior and posterior hippocampal regions according to McHugo et al.
(Plassard et al., 2017; McHugo et al., 2018). En general, this parcellation is relatively coarse to
reduce dimensions in our analyses and to account for lower spatial sampling of functional MRI
data relative to anatomic imaging.
Neurociencia en red
436
Progression of temporal lobe epilepsy networks
The fMRI images were preprocessed using SPM12 software (http://www.fil.ion.ucl.ac.uk
/spm/software/spm12/) and MATLAB 2019a (MathWorks, Natick, MAMÁ). Primero, physiological
noise correction using the retrospective correction of physiological motion effects in functional
resonancia magnética (RETROICOR) protocol (Glover et al., 2000) was implemented using the pulse oximeter
and respiratory belt data. Próximo, SPM12 was used for slice timing correction, motion correc-
ción, spatial normalization to the Montreal Neurological Institute template via the T1-weighted
conjuntos de datos, and spatial smoothing (6 mm × 6 mm × 6 mm FWHM Gaussian kernel). Then the
fMRI time series were temporally band-pass filtered at 0.0067 a 0.1 Hz (Cordes et al., 2001).
The preprocessed functional MRI time series were averaged across all voxels in each region
de interés. Then a partial Pearson correlation between each pair of averaged time series was
computed using six motion and one mean white matter time series as confounds. The cor-
relation coefficients were then normalized using the Fisher Z transform (Pescador, 1915). Este
resulted in a matrix of pairwise functional connectivity values, which in this work will be gen-
erally referred to as a functional connectivity (FC) conectoma. To account for the effects of
age in this connectome, a linear relationship was assumed. Using only the healthy controls,
the linear fit and the root mean squared error of the fit were computed for each pair of re-
gions (borde). This edge-wise fit was then used to correct the FC in each subject, Resultando en
a connectome of the FC residuals after linear age regression (FCres, Z values) and in standard
deviation from age-matched control when further divided by root mean squared error of the
linear fit for that edge (FCcorr). This correction was done in native left and right hemispheres,
but patients were then transformed into ipsilateral and contralateral with respect to seizure
focus for some analyses, where specified.
The DWI images were preprocessed using MRtrix3 (Tournier et al., 2019) incluido
denoising (Cordero-Grande et al., 2019), eddy current and motion correction (andersson &
Sotiropoulos, 2016), and bias correction of B1 field inhomogeneity (Tournier et al., 2019).
Then the response function was estimated for spherical deconvolution for estimation of fiber
orientation distribution (Tournier et al., 2007). Próximo, SPM12 and MATLAB 2019a were used
to generate the gray matter–white matter interface using the high-resolution T1-weighted im-
age and the mean B0 image. Using MRtrix3, 2 × 107 anatomically constrained probabilistic
streamlines were generated through the white matter from this interface (Smith et al., 2012).
The streamlines were then reduced to 1 × 107 using spherical convolution–informed filtering
to match the fiber orientation density integrals (SIFT2) (Smith et al., 2015). El 109 regiones
of interest were then used to create a connectome matrix of the streamline count between
each pair of regions scaled by the inverse of the two region volumes as a measure of structural
conectividad (CAROLINA DEL SUR). Además, similar to the FC connectome, the SC connectomes were cor-
rected for age by using linear fits of the healthy control data. Sin embargo, to convert these data
to a Gaussian distribution, a log transform was used prior to the fitting. This resulted in SC
connectomes of residuals and standard deviation from age-matched control, SCres, and SCcorr
for each subject. These were also computed in left and right hemispheres, but were then con-
verted to ipsilateral and contralateral to seizure focus in specified analyses. Similarmente, usando
MRtrix3, a connectome matrix of mean streamline length between each pair of regions was also
creado (LEN).
Models of Distance in the Healthy Brain
We developed four models, each based on a different topology (t), in the healthy brain to
which MRI network progression in patients can be compared. Three topologies studied were
based on distance to the presumed seizure focus and constructed using the connectomes
Neurociencia en red
437
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
defined above: streamline length (TLEN), structural connectivity (TSC), and functional connec-
actividad (TFC). For each, the designated connectomes were averaged across all healthy controls
to yield the topology matrix of the healthy brain.
Rather than distance between regions reflected in the first three topologies, the fourth
topology was created based on membership into functional connectivity resting-state network
(TRSN). Distance here refers to the hierarchy of transmodal functional networks (default mode
network, other higher cognitive networks) to unimodal functional networks (perceptual, pri-
mary sensory/motor networks) defined by a principal gradient of cortical organization in the
human functional connectome by Margulies et al. (2016). Using this idea, four resting-state
redes (Buckner et al., 2008; Uddin et al., 2019), each consisting of five bilateral regions,
were identified and ordered as follows: (a) default mode network, (b) attention network, (C)
primary visual network, y (d) motor/sensorimotor network. This topology matrix was de-
signed by repeating rows where each position represents one region of interest with the value
(0–4) indicating which of the four networks in which it is included (1–4, respectivamente, o 0 si
ninguno).
Entonces, similar to the work by Betzel et al. (2016), we developed models of distance in
the healthy brain with respect to a given seed region and topology. Específicamente, let R =
(cid:2)
be a binary vector encoding the selected seed region, con R(norte) =
R(1) . . . R(norte)
(cid:3)
(cid:4)
0, region n is not seed #
1, region n is seed
, and let T be a topology matrix corresponding to one of TLEN,
TSC, TFC, or TRSN as defined above, with elements
modeled distance from region n to each other region, under topology, t, is given by the vector:
R x T = [t(norte, 1), t(norte, 2) . . . t(norte, norte)]
(1)
⎡
⎢
⎢
⎣
t(1, 1)
…
t(norte, 1)
t(1, norte)
…
· · ·
. . .
· · · T(norte, norte)
⎤
⎥
⎥
⎦. Then the
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
t
.
For the FC, CAROLINA DEL SUR, and LEN topologies, norte = 55, which represents the regions across one hemi-
sphere and the brainstem. Cross-hemisphere connections in many subjects, especially for the
CAROLINA DEL SUR, were nonexistent and so were not included. For resting-state networks (RSN) topología,
norte = 109 to represent all the regions of interest. We used these models to test whether net-
work progression in patients increased when distance from the anterior hippocampus or along
a functional gradient in the healthy brain decreased.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Hippocampal Network Change in Patients
Two types of MRI network progression were investigated and compared to the four models
arriba. The first type of network was defined as the connection between the anterior hip-
pocampus (presumed seizure focus) and each other region. To compare these hippocampal
connections to a model, the following procedure was implemented (Figura 1A). The model
vector from (Ecuación 1) was sorted into bins of regions based on distance defined by each
topología. For the LEN model, the “closest” bin (bin 1) had the shortest length. For the FC
modelo, bin 1 had the highest absolute value of the functional connectivity to the anterior hip-
pocampus. For the SC model, bin 1 had the highest structural connectivity to the anterior
hippocampus. For the RSN model, the edges were binned from their respective assignment
to resting-state network based on transmodal to unimodal gradient (1–4). De este modo, each bin con-
sisted of approximately 10 edges between anterior hippocampus and other regions. Entonces, a
Neurociencia en red
438
Progression of temporal lobe epilepsy networks
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Hippocampal network change in patients. (A) Diagram representing analysis process
using streamline length as the topology, t, and the anterior hippocampus (seizure focus) como el
region of interest, R, to develop an example model of distance from R in the healthy brain. El
model vector of LEN values is sorted and binned (in this example). The Mahalanobis distance is
then computed for edges in each bin to quantify the patient’s change from the population of healthy
controls. The hypothesis is that the Mahalanobis distance for the “closest” bin (bin 1, shortest LEN)
will be highest, with decreasing values for bins of increasing LEN. (B) The Mahalanobis distance
is a multivariate measure of change that accounts for the covariance between the variables, dónde
the Euclidean distance does not. An example of 100 samples of two-variable data with a Pearson
correlation of r = 0.74 and a mean at [0 0] is illustrated. The contours of Euclidean distance (ED)
de 1, 2, y 3 are shown in blue (izquierda), while the contours of the Mahalanobis distance (METRO) de 1, 2,
y 3 are shown in red (bien). An example point ([2, −1], shown in green) has an ED = 2.2 y
m = 21.5; ant hip = anterior hippocampus; LEN = connectome based on streamline length across
healthy controls.
Mahalanobis distance:
The measurement of distance
between objects that takes into
account the correlation in the data
by calculating the inverse of the
variance-covariance matrix of the
data of interest.
multivariate z-score, Mahalanobis distance (De Maesschalck et al., 2000; Taylor et al., 2020)
was used to compute network connectivity change between each patient and the control group
across the edges in the given bin. The Mahalanobis distance, METRO, is a multivariate quantifica-
tion of distance that, unlike Euclidean distance, considers the correlation between the variables
(Figura 1A). As described in Taylor et al. (2020), we computed the Mahalanobis distance of
the edges in a bin for a patient, METRO, using the following:
(cid:11)
m =
(s − μ)T · C−1 · (s − μ)
(2)
where s is the vector of length k of FCres or SCres values (after linear regression to account for
edad) of the edges in the bin in the patient, k is the number of edges in a bin, μ = the mean of
Neurociencia en red
439
Progression of temporal lobe epilepsy networks
the same FCres or SCres edges across a set of controls, and C is the covariance matrix between
those measures in the set of controls computed using shrinkage estimators. To compare M
across bins and patients, the number of edges (k) must be held constant across computations.
Because some bins had 10 edges and some had 11, and because there were a few subjects in
which some edges were not existent (mostly for SC), we performed 500 permutations with a
random subset of 7 (out of 10 o 11) bordes. Any sample in which the patient did not have seven
detected edges was discarded. Y, nested within this loop, we performed a second set of 500
permutations of a subset of 50 random control subjects (out of 70). Any loop with less than
45 controls having all seven edges was discarded. The mean of these permutations was used
as the final M. These calculations are carried out in regions with respect to left and right in
the brain in both patients and controls, and then converted to ipsilateral and contralateral with
respect to the seizure focus to compare across patients. The analyses were performed with
functional (MFC) and structural connectivity (MSC) across the edges separately to determine
whether each type of connectivity is altered in patients as a function of distance to the anterior
hippocampus or along a functional gradient in the healthy brain.
To test our hypothesis that M of bin 1 > bin 2 > bin 3 > bin 4 > bin 5, we used three
Estadísticas. Primero, there must be difference in M between bins across all patients using repeated
measures ANOVA. Segundo, the means across bins had to decrease from bin 1 to bin 5. Tercero,
(uncorrected) paired t tests had to confirm that no higher number bins were significantly greater
than lower number bins. Models were developed with the anterior hippocampus ipsilateral
and contralateral to the seizure focus, separately. Próximo, to investigate temporal progression, en
any findings in which network change from control was related to distance along a topology,
statistics were repeated after patients were grouped into three categories based on duration of
enfermedad (described in results).
Whole-Brain Network Change in Patients
Próximo, we developed a method to compare whole-brain network progression in patients as
a function of distance in the healthy brain. Para hacer esto, a hybrid functional and structural
connectome independent component analysis (ICA) method proposed by Amico and Goñi
(Amico & Goñi, 2018) with fastICA (Hyvarinen, 1999) was implemented (Figura 2A). Joint
functional-structural connectomes from each patient were created by concatenating the FCcorr
and SCcorr connectomes (in units of SD from healthy age-matched controls) after transforming
from left and right hemispheres to ipsilateral and contralateral to group all patients. Principal
component analysis was used to reduce dimensionality of the data through decomposition and
reconstruction of 90% of variance. Entonces, due to the nondeterministic behavior of the fastICA
algoritmo, it was run 40 veces. The output connectome components for each run were com-
pared, and any two component connectomes with a Pearson correlation coefficient greater
than 0.8 were combined through averaging. Their associated patient weights were also aver-
aged. This resulted in a set of combined component connectomes and patient weight vectors
across all runs (Amico & Goñi, 2018). The weights for the final components were compared to
duration of disease across patients to identify any FCSC components that exhibit a monotonic
relationship (Spearman correlation) to increasing duration of disease. In such components, el
values in the connectome reflect connections that increase (or decrease) in connectivity with
increase in duration of disease. Próximo, to compare the connectome components to the model
vectors of distance (Ecuación 1), they were converted to vectors representing full brain con-
nectivity to each region in two steps (Figura 2B). Primero, the FC and SC connectome components
were each thresholded at +/ − 0.55 y +/ − 0.175, respectivamente, to remove the effect of the
edges with lowest relationship to duration of disease and create a matrix of approximately 20%
Independent component analysis:
A statistical method for identifying
sources (componentes) that are
linearly mixed in a measured signal.
Neurociencia en red
440
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. Whole-brain network change in patients. (A) A hybrid FC SC independent component
análisis (ICA) was performed using age corrected functional (FCcorr) and structural connectomes
(SCcorr) across all patients. Resulting connectome component weights were each correlated with
duration of disease across patients to identify the ICA components related to duration of disease.
(B) The FC and SC component connectomes related to duration of disease were thresholded, y un
weighted degree to each region was computed to create vectors of region to whole-brain connec-
tivity related to duration of disease. These vectors were then compared to the models of distance
to the anterior hippocampus or along a functional gradient in the healthy brain; ant hip = anterior
hippocampus; FC = functional connectivity; SC = structural connectivity.
density. Segundo, a weighted degree (Rubinov & despreciar, 2010) was computed for each region
to create a single value of net connectivity from each region to the whole brain. This vector
of connectivity from each region to whole brain was then correlated to the models of distance
in the healthy brain. Note that the ICA component regions are identified in relation to ipsilat-
eral and contralateral to the seizure focus, whereas the model distances were computed from
controls as left and right regions. Por lo tanto, the distance to each region used in the model was
the average of the distances in the left and right hemispheres. Significant correlation would
suggest that a region’s connectivity change across the brain with increasing duration of disease
is associated with its distance from the anterior hippocampus or across a functional gradient
in the healthy brain.
RESULTADOS
Hippocampal Network Change in Patients Related to Healthy Brain Topology
The Mahalanobis distance (METRO) was used to quantify hippocampal network change in the pa-
tients compared to controls in bins that were ranked by distance to the focus or by functional
Neurociencia en red
441
Progression of temporal lobe epilepsy networks
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3. Hippocampal network change in patients related to healthy brain topology. (A) The four topologies were used to create 4 o 5
bins of regions based on their distance to the anterior hippocampus (ant hip) in the same hemisphere or along a functional network gradient.
Papelera 1 has the lowest streamline length (LEN), highest absolute value of functional connectivity (FC), or highest structural connectivity (CAROLINA DEL SUR)
to the anterior hippocampus in the same hemisphere. The resting-state networks (RSN) were ranked based on a gradient of transmodal to
unimodal function. (B) Ipsilateral hippocampal functional network change across all patients, MFC, decreases as a function of the gradient of
transmodal to unimodal resting-state network, TRSN. (C) Ipsilateral hippocampal structural network change across all patients, MSC, decreases
with increasing distance based on structural connectivity to the anterior hippocampus, TSC. (D) Hippocampal functional network change,
MFC, as a function of the gradient of transmodal to unimodal functional resting-state network, TRSN, in patients grouped by duration of
(mi) Hippocampal structural network change in patients, MSC, with increasing distance from the anterior hippocampus based on
enfermedad.
structural connectivity, TSC, in patients grouped by duration of disease. The mixed ANOVAs for parts D and E did not reveal statistical
difference between the duration groups. Individual group comparisons are shown for illustration. In B–E, the p value listed refers to repeated
measures ANOVA between bins. Dashed lines represent uncorrected paired t-test difference between bins with p = 0.05. Solid lines represent
uncorrected paired t-test difference between bins with p < 0.01. The color of the p value listed and the lines of significance reflect the data
to which they refer; ipsi = anterior hippocampus ipsilateral to seizure focus; contra = anterior hippocampus contralateral to seizure focus;
DMN = default mode network; ATT = attention network; VIS = primary visual network; MOT = motor/sensorimotor network.
network gradient based on topology in the healthy brain (Figure 3A). Considering MFC, where
FC denotes functional connectivity hippocampal network change in patients, four healthy brain
topologies were investigated (TLEN, TSC, TFC, TRSN). Therefore, the threshold for statistical
significance of the repeated measures ANOVA was 0.0125. Post hoc t tests between bins of
significant models are not corrected for multiple comparisons to illustrate all relationships. For
the ipsilateral anterior hippocampus, the repeated measures ANOVA using TLEN, F(4, 156) =
2.62, p > 0.0125, and TSC, F(3, 117) = 1.53, p > 0.0125, were not significant. Using TFC,
there were differences between bins, repeated measures ANOVA, F(4, 156) = 3.63, pag = 0.011,
but the means did not decrease across bins. Using TRSN, there were differences between bins,
repeated measures ANOVA, F(3, 117) = 5.12, pag = 0.003, with means decreasing across bins:
default mode network > attention network > primary visual network > motor/sensorimotor
network. The paired t test showed that MFC was higher in the default mode network than the
Neurociencia en red
442
Progression of temporal lobe epilepsy networks
Functional resting-state network:
Regions of the brain with correlated
spontaneous oxygenation
fluctuations measured by functional
resonancia magnética; usually denotes groups of
regions forming networks for
integrated task performance.
atención (pag = 0.05), the primary visual (pag = 0.002), and motor/sensorimotor network (pag =
0.001) (Figura 3B). These results suggest that functional connectivity changes from the seizure
focus in mTLE occur as a function of the gradient of transmodal to unimodal functional resting-
state network in the healthy brain. For the contralateral anterior hippocampus, there were no
significant differences between bins for any topology (repeated measures ANOVA, p > 0.0125).
Considering MSC, where SC denotes structural connectivity hippocampal network change,
three healthy brain topologies were investigated (TLEN, TSC, TFC). The TRSN bins included
cross-hemispheric edges for which structural connectivity was not reliably detected. Allá-
delantero, the threshold for significance of the repeated measures ANOVA was 0.0167. As above,
posthoc t tests between bins of significant models are not corrected for multiple comparisons
to illustrate all relationships. For the ipsilateral anterior hippocampus, using TLEN there was
no decreasing trend across bins, only that the bin 1 (shortest streamline length) had greater
MSC than the others, repeated measures ANOVA, F(6, 152) = 6.33, pag < 0.001; paired t test,
bin 2: p = 0.02, bin 3: p = 0.003, bin 4: p = 0.008, bin 5: p = 0.004. Similarly, using
TFC the same pattern of greater MSC in bin 1 (highest absolute functional connectivity) than
the other bins, repeated measures ANOVA, F(4, 152) = 4.11, p = 0.003; paired t test, bin 2:
p = 0.013, bin 3: p = 0.004, bin 4: p = 0.012, bin 5: p = 0.039. Using TSC, there were dif-
ferences in MSC between bins, repeated measures ANOVA, F(3, 114) = 9.81, p < 0.001; and
the means decreased across the bins, with significant differences to support the trend, paired
t test, bin 1 > bin 3: pag = 0.016, bin 1 > bin 4: pag < 0.001, bin 2 > bin 4: pag = 0.001, bin
3 > bin 4: pag = 0.003 (Figura 3C). These results suggest that structural connectivity changes
from the seizure focus in mTLE occur as a function of structural connectivity distance to the
anterior hippocampus in the healthy brain. For the contralateral anterior hippocampus, allá
were no significant differences between bins (repeated measures ANOVA, p > 0.0167).
To investigate the temporal changes, the patients were divided into three groups based on
duration of disease (≤ 10 años, ‘short,' [norte = 15]; 11–30 years, ‘medium,' [norte = 14]; >30 years,
‘long,' [norte = 11]), and the statistics for the best model for MFC and MSC were recomputed using
a mixed ANOVA by adding the group identification as a between-subject factor. For MFC
using the resting-state network topology, TRSN, and for MSC using the structural connectivity
network topology, TSC, the mixed ANOVAs did not detect a significant effect of bin by group
interacción, MFC: F(3, 6) = 0.33, p > 0.05; MSC: F(3, 6) = 0.65, p > 0.05. Sin embargo, to begin
to investigate potential relationships of interest across the duration groups, individual repeated
measures ANOVA were performed are reported.
For MFC the resting-state network topology, TRSN, fue usado. For the short-duration group,
there was no difference between the bins, repeated measures ANOVA, F(3, 42) = 0.28, p >
0.05. For the medium-duration group, the means were different between groups, repeated
measures ANOVA, F(3, 39) = 3.17, pag = 0.047, with the default mode network > primary
visual (paired t test, pag = 4 0.014) and motor/sensorimotor network (paired t test, pag = 0.011).
For the long-duration group, there was difference between bins, repeated measures ANOVA,
F(3, 30) = 3.9, pag = 0.033, with the mean of the default mode network > the attention (paired
t test, pag = 0.012), primary visual (paired t test, pag = 0.027) and motor/sensorimotor network
(paired t test, pag = 0.008) (Figura 3D). These findings imply that the progression of functional
connectivity alterations in patients tends to occur over time with the highest changes in regions
of transmodal resting-state networks occurring in the medium- to long-durations groups.
For MSC the structural connectivity topology, TSC, fue usado. For the short-duration group,
there was no difference between the bins, repeated measures ANOVA, F(3, 39) = 1.99, p >
Neurociencia en red
443
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
Cifra 4. Hybrid independent component analysis (ICA) component related to duration of dis-
ease.
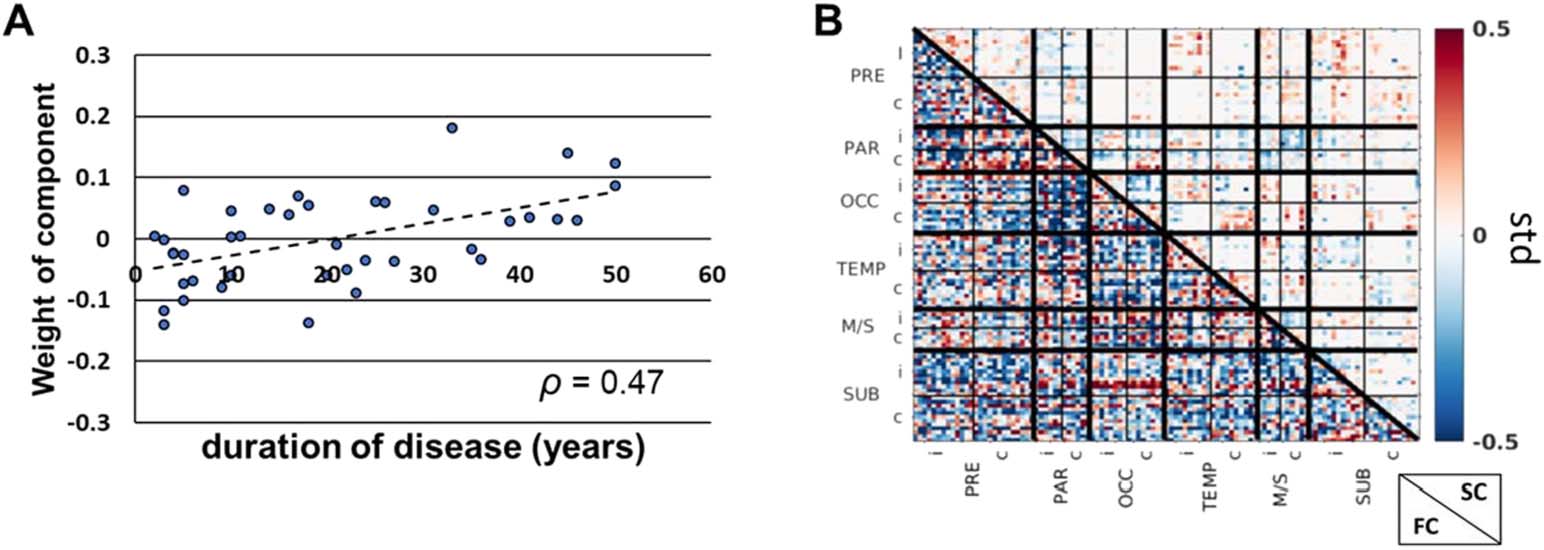
(A) One ICA component set of weights was correlated with duration of disease across all
patients (Spearman ρ = 0.47, pag < 0.05). (B) The FC and SC connectome of the component that
was positively correlated with duration of disease in A. Positive (negative) values indicate increased
(decreased) connectivity with increased duration of disease. FC = functional connectivity; SC =
structural connectivity; PRE = prefrontal; PAR = parietal; OCC = occipital; TEMP = temporal;
M/S = somatosensory/motor; SUB = subcortical regions; i = ipsilateral to seizure focus; c = con-
tralateral to seizure focus.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
0.05. For the medium-duration group, there was difference between bins, repeated measures
ANOVA, F(3, 39) = 4.44, p = 0.009, with MSC in bin 1 higher than bin 3 and bin 4 (paired
t test, p = 0.048 and p = 0.006, respectively). For the long-duration group, again there was
difference between bins, repeated measures ANOVA, F(3, 30) = 4.63, p = 0.022, with MSC
decreasing across bins. In this case, bin 4 was less than the first and second bin (paired t test,
bin 1: p = 0.008 and p = 0.001, respectively) (Figure 3E). Taken together, these results suggest
that changes in regions most structurally connected to the presumed focus in the healthy brain
tend to occur in the second and third decade of disease.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
n
e
n
_
a
_
0
0
1
8
4
p
d
.
t
Whole-Brain Network Change in Patients As a Function of Distance in the Healthy Brain
One component of the hybrid ICA analysis had weights that were positively correlated with
duration of disease (Spearman ρ = 0.47, p < 0.05) (Figure 4A and B). This component was
identified in 50% of the ICA iterations. The component FC and SC connectomes were then
transformed to a vector capturing region to whole-brain connectivity (Figure 5A and B) and
correlated with the four models of distance in the healthy brain. Therefore, the threshold for
significance for the correlation was 0.0125.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The FC component connectome related to duration of disease correlated with the model
using TFC (Pearson r = 0.22, p = 0.02) (Figure 5C). This is not statically significant when
correcting for multiple comparisons, but is shown for illustration as a promising relationship.
When using models created with TLEN, TSC, and TRSN there was no significant correlation
with the connectome component (p > 0.05). This suggests that there is a trend that a region’s
whole-brain functional connectivity change with increasing duration of disease in mTLE is a
function of the region’s functional connectivity distance from the anterior hippocampus in the
healthy brain. The SC component connectome related to duration of disease correlated with
the model using TLEN (Pearson r = −0.26, pag = 0.009) (Figure 5D). No other models correlated
with this connectome component (p > 0.05). These results imply that a region’s whole-brain
structural connectivity change with duration of disease is inversely proportional to the region’s
mean streamline length to the anterior hippocampus in the healthy brain.
Neurociencia en red
444
Progression of temporal lobe epilepsy networks
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
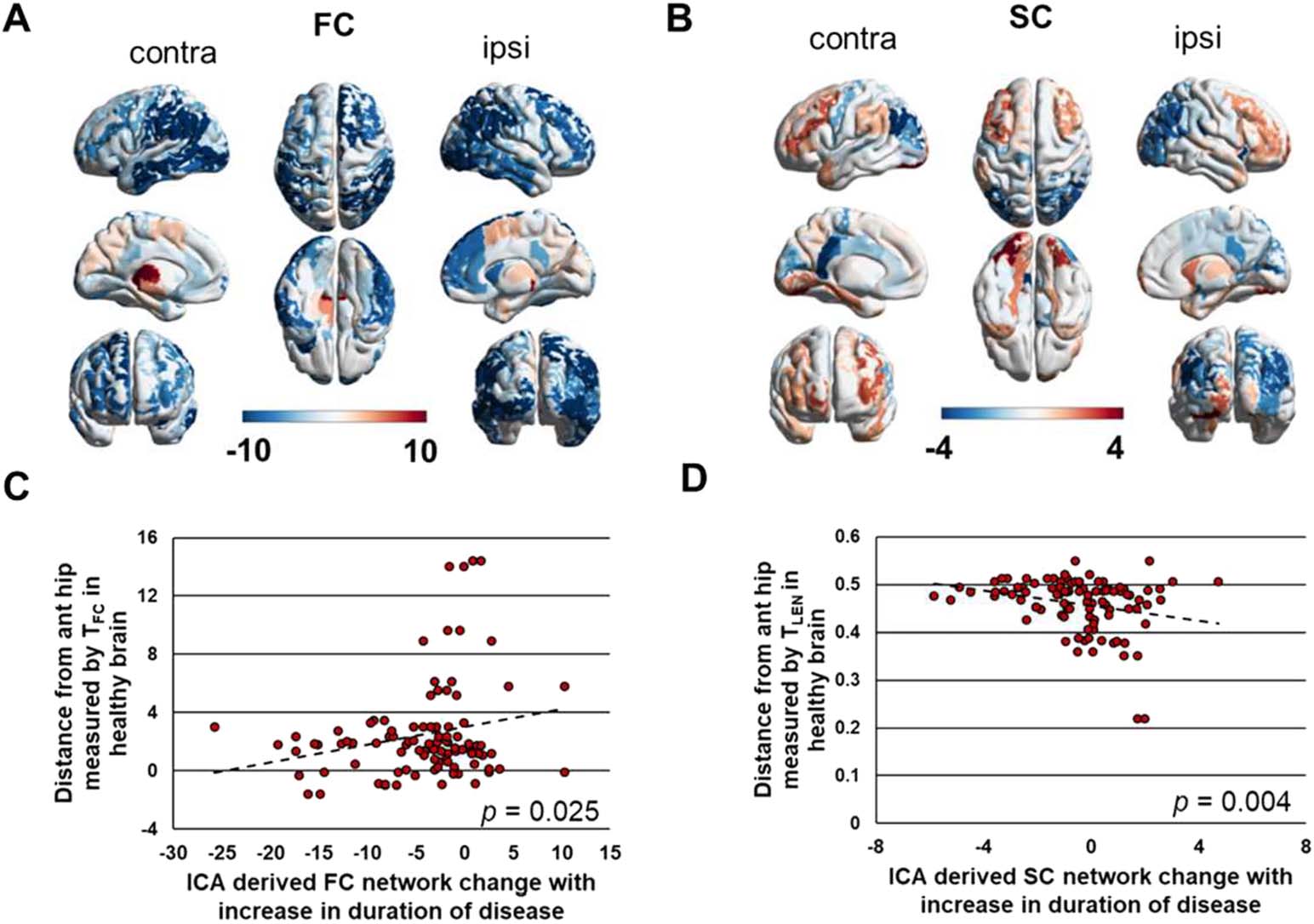
Cifra 5. Whole-brain connectivity changes with disease duration as a function of distance in
healthy brain. (A) Vector of ICA-derived regional average functional connectivity change with in-
(B) Vector of ICA-derived regional aver-
creasing duration of disease illustrated across the brain.
age structural connectivity change with increasing duration of disease illustrated across the brain.
(C) ICA-derived functional connectivity change with increasing duration of disease shows a trend
toward positive correlation with distance from the anterior hippocampus in the healthy brain mea-
sured by functional connectivity, TFC (Pearson r = 0.22, pag = 0.02).
(D) ICA-derived structural
connectivity change with increasing duration of disease is negatively correlated with distance from
the anterior hippocampus in the healthy brain measured by streamline length, TLEN (Pearson r =
−0.26, pag = 0.009). The colors of regions in A and B are represented by the x-axis in C and D, respetar-
activamente. FC = functional connectivity; SC = structural connectivity; ipsi = ipsilateral to seizure focus;
ICA = independent
contra = contralateral to seizure focus; ant hip = anterior hippocampus;
component analysis.
DISCUSIÓN
Focal epilepsy manifests widespread functional and structural alterations across the brain that
have been well characterized (Tavakol et al., 2019). What remains unclear, sin embargo, are why
are some regions more affected than others, and what is the timeline of this progression? En esto
work we showed that both networks connected to the seizure focus (anterior hippocampus)
and global, whole-brain networks were altered in mTLE along a predictable framework defined
by a given topology in the healthy brain.
Además, these changes evolved over several
decades of disease duration.
The hippocampal functional network alterations in mTLE occurred across an organization
of the brain related to inclusion in functional resting-state network ranked by transmodal to
unimodal gradients (Margulies et al., 2016). This suggests that functional reorganization occurs
with the most pronounced changes in the regions within the default mode network and those
serving transmodal functions supporting higher cognitive functions, with decreasing effects
into unimodal networks supporting perceptual functions. One explanation for this is that the
seizure focus in the anterior hippocampus is highly connected to and sometimes considered
Neurociencia en red
445
Progression of temporal lobe epilepsy networks
a node in the default mode network (Buckner et al., 2008), thus reflecting more within func-
tional network changes. Alternativamente, hippocampal structural connectivity alterations in mTLE
occurred as a function of distance to the anterior hippocampus in the healthy brain measured
by structural connectivity. This implies that the greatest structural network changes occur in
patients in regions more structurally connected to the region of the seizure focus and decrease
as structural connectivity to the focus decreases in healthy controls.
When investigating the temporal characteristics of these changes in hippocampal networks,
we found no difference between groups based on duration of disease. Closer examination
of patterns of change in each group, sin embargo, revealed that the trends in both functional
and structural networks suggest that the patterns related to the focus may develop later in the
enfermedad. This is interesting because it is counter to our presumption that changes occur close to
the focus first and then spread outward later. Pero, this question will most likely require larger
datasets to confirm.
The global whole-brain network to each region was investigated through the identifica-
tion of a joint whole-brain functional and structural connection pattern related to duration of
disease that was then compared to models of healthy brain architecture. Results showed a
preliminary trend that functional network changes related to duration of disease increased as
functional connectivity to the anterior hippocampus in the healthy brain increased. Structural
network changes related to duration of disease decreased as a function of increasing distance
from the anterior hippocampus measured by streamline length in the healthy brain. This means
that regions with the greatest changes over the duration of disease had the highest functional
connections or shortest streamline length to the presumed seizure focus in the anterior hip-
pocampus in the same hemisphere.
En general, it is not completely surprising that hippocampal functional network changes are
related to a functional architecture, while structural network changes are related to a structural
network architecture in the healthy brain. The potential consequences of this difference in
architectural framework are interesting. Primero, these results may imply that interventions that
target the seizure focus will have structural effects on those regions directly connected to the
focus by white matter tracts, while functional effects may be more widespread across the brain
in regions not physically connected to the focus. Segundo, structural (físico) changes in white
matter may take longer to occur than functional changes, and so functional network changes
may be detected prior to structural network connectivity. Although we did not detect this
difference in this study, the magnitude of the functional network changes were approximately
three times those of the structural changes when measured in standard deviations from age-
matched controls, which is consistent with this idea.
There are potential clinical implications of these findings. Network information has emerged
as a potential predictor of treatment outcome (Morgan et al., 2017; Gleichgerrcht et al., 2018;
Taylor et al., 2018), and understanding how and when networks evolve could identify win-
dows of time when specific treatments may be most effective.
In a similar way, it may be
possible to predict widespread longitudinal postsurgical network changes to the resected area
to predict long-term postsurgical outcomes (Morgan et al., 2019). Además, cognitive and
behavioral impairments in mTLE and their changes over time (Helmstaedter & Kockelmann,
2006) may be better understood by their relation to network reorganization (Park et al., 2017).
Finalmente, this work may provide a framework for investigations to predict progression in other
focal epilepsies.
Neurociencia en red
446
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
This study could be improved in several ways. Primero, a longitudinal study would provide
better temporal data than this cross-sectional study. Segundo, these analyses did not control
for age of onset of mTLE. In our cohort, age of onset was significantly negatively correlated
with duration of disease (Pearson r = −0.59, pag < 0.001), but the medium- and long-duration
groups were not different, ANOVA F(2, 37) = 8.2, p = 0.001; t-test medium vs. long duration
p > 0.05; all other t tests p < 0.05. Thus, duration is similar to, but not an exact proxy, for
age of onset changes. Third, we also did not control for variations in seizure type or frequency.
This analysis would require a much a larger cohort in which the complete analyses could
be performed on sets of patients with more homogeneous seizure frequencies within the set.
Similarly, we did not control for variations in medication use. While all patients were on
medication, to address this thoroughly would require analyses of subsets of patients on the
same or similar medications. In addition, a larger, more balanced cohort of right and left mTLE
patients would allow for separate investigations rather than the ipsilateral/contralateral pooling
performed here. These are important potential confounds that would need a larger or more
homogeneous sample size to address. Finally, the investigations were based on associations,
while causation and other effects for these changes require further studies.
In conclusion, these findings support the idea that widespread network changes in mTLE
occur along specific pathways that can be predicted by healthy brain architecture over the du-
ration of disease. The greatest changes in hippocampal functional networks in mTLE occurred
in regions of transmodal functional resting-state networks, with decreasing changes in regions
involved in unimodal functional networks. Other networks studied had greatest change in
regions with shortest distance to the seizure focus, with decreasing changes as the distance
from the focus increased. Distance was quantified by functional connectivity, structural con-
nectivity, or streamline length to the anterior hippocampus (presumed seizure focus) in the
healthy brain. Overall, this work presents a framework of spatiotemporal network progression
over duration of disease related to the seizure focus and healthy brain architecture that may
be used to predict individual network evolution in focal epilepsy.
AUTHOR CONTRIBUTIONS
Victoria L. Morgan: Conceptualization; Data curation; Formal analysis; Funding acquisition;
Investigation; Methodology; Project administration; Resources; Supervision; Validation; Visu-
alization; Writing – original draft; Writing – review & editing. Graham W. Johnson: Formal anal-
ysis; Methodology. Leon Y. Cai: Formal analysis; Methodology. Bennett A. Landman: Formal
analysis; Methodology; Software. Kurt G. Schilling: Methodology. Dario J. Englot: Concep-
tualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology;
Writing – review & editing. Baxter P. Rogers: Formal analysis; Investigation; Methodology;
Writing – review & editing. Catie Chang: Conceptualization; Formal analysis; Funding acqui-
sition; Investigation; Methodology; Writing – review & editing.
FUNDING INFORMATION
Victoria L. Morgan, National Institute of Neurological Disorders and Stroke (http://dx.doi.org
/10.13039/100000065), Award ID: NS075270. Victoria L. Morgan, National Institute of Neu-
rological Disorders and Stroke (http://dx.doi.org/10.13039/100000065), Award ID: NS108445.
Victoria L. Morgan, National Institute of Neurological Disorders and Stroke (http://dx.doi.org
/10.13039/100000065), Award ID: NS110130. Dario J. Englot, National Institute of Neuro-
logical Disorders and Stroke (http://dx.doi.org/10.13039/100000065), Award ID: NS097618.
Dario J. Englot, National Institute of Neurological Disorders and Stroke (http://dx.doi.org/10
.13039/100000065), Award ID: NS112252.
Network Neuroscience
447
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
n
e
n
_
a
_
0
0
1
8
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Progression of temporal lobe epilepsy networks
REFERENCES
Amico, E., & Goñi, J. (2018). Mapping hybrid functional-structural
connectivity traits in the human connectome. Network Neu-
roscience, 2(3), 306–322. DOI: https://doi.org/10.1162/netn_a
_00049, PMID: 30259007, PMCID: PMC6145853
Andersson, J. L. R., & Sotiropoulos, S. N. (2016). An integrated ap-
proach to correction for off-resonance effects and subject move-
ment in diffusion MR imaging. NeuroImage, 125, 1063–1078.
DOI: https://doi.org/10.1016/j.neuroimage.2015.10.019, PMID:
26481672, PMCID: PMC4692656
Asman, A. J., & Landman, B. A. (2013). Non-local statistical label
fusion for multi-atlas segmentation. Medical
Image Analysis,
17(2), 194–208. DOI: https://doi.org/10.1016/j.media.2012.10
.002, PMID: 23265798, PMCID: PMC3648421
Besson, P., Dinkelacker, V., Valabregue, R., Thivard, L., Leclerc, X.,
Baulac, M., . . . Dupont, S. (2014). Structural connectivity diff-
erences in left and right temporal lobe epilepsy. NeuroImage,
100, 135–144. DOI: https://doi.org/10.1016/j.neuroimage.2014
.04.071, PMID: 24814212
Betzel, R. F., Avena-Koenigsberger, A., Goñi, J., He, Y., de Reus,
M. A., Griffa, A., . . . Sporns, O. (2016). Generative models of
the human connectome. NeuroImage, 124(Pt A), 1054–1064.
DOI: https://doi.org/10.1016/j.neuroimage.2015.09.041, PMID:
26427642, PMCID: PMC4655950
Bjellvi, J., Olsson, I., Malmgren, K., & Wilbe Ramsay, K.
(2019).
Epilepsy duration and seizure outcome in epilepsy surgery.
Neurology, 93(2), e159. DOI: https://doi.org/10.1212/WNL
.0000000000007753, PMID: 31182508, PMCID: PMC6656653
Bonilha, L., Nesland, T., Martz, G. U., Joseph, J. E., Spampinato,
M. V., Edwards,
(2012). Medial tem-
poral lobe epilepsy is associated with neuronal fibre loss and
paradoxical increase in structural connectivity of limbic struc-
tures. Journal of Neurology. Neurosurgery, & Psychiatry, 83(9),
903–909. DOI: https://doi.org/10.1136/jnnp-2012-302476, PMID:
22764263, PMCID: PMC3415309
J. C., & Tabesh, A.
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L.
(2008).
The brain’s default network: Anatomy, function, and relevance
to disease. Annals of the New York Academy of Sciences, 1124,
1–38. DOI: https://doi.org/10.1196/annals.1440.011, PMID:
18400922
Cordero-Grande, L., Christiaens, D., Hutter, J., Price, A. N., &
Hajnal, J. V. (2019). Complex diffusion-weighted image estima-
tion via matrix recovery under general noise models. NeuroIm-
age, 200, 391–404. DOI: https://doi.org/https://doi.org/10.1016
/j.neuroimage.2019.06.039,
PMCID:
PMC6711461
31226495,
PMID:
Cordes, D., Haughton, V. M., Arfanakis, K., Carew, J. D., Turski,
P. A., Moritz, C. H., . . . Meyerand, M. E.
(2001). Frequencies
contributing to functional connectivity in the cerebral cortex in
“resting-state” data. American Journal of Neuroradiology, 22(7),
1326–1333.
De Maesschalck, R., Jouan-Rimbaud, D., & Massart, D. L. (2000).
The Mahalanobis distance. Chemometrics and Intelligent Labor-
atory Systems, 50(1), 1–18. DOI: https://doi.org/https://doi.org
/10.1016/S0169-7439(99)00047-7
Dugan, P., Carlson, C., Jette, N., Wiebe, S., Bunch, M., Kuzniecky,
R., & French, J. (2017). Derivation and initial validation of a sur-
gical grading scale for the preliminary evaluation of adult patients
with drug-resistant focal epilepsy. Epilepsia, 58(5), 792–800.
DOI: https://doi.org/10.1111/epi.13730, PMID: 28378422
Engel, J.
(2001). Mesial temporal lobe epilepsy: What have we
learned? Neuroscientist, 7(4), 340–352. DOI: https://doi.org/10
.1177/107385840100700410, PMID: 11488399
Engel, J. Jr. (2016). What can we do for people with drug-resistant
epilepsy? The 2016 Wartenberg Lecture. Neurology, 87(23),
2483–2489. DOI: https://doi.org/10.1212/wnl.0000000000003407,
PMID: 27920283, PMCID: PMC5177675
Englot, D. J., Konrad, P. E., & Morgan, V. L. (2016). Regional and
global connectivity disturbances in focal epilepsy, related neuro-
cognitive sequelae, and potential mechanistic underpinnings.
Epilepsia, 57(10), 1546–1557. DOI: https://doi.org/10.1111/epi
.13510, PMID: 27554793, PMCID: PMC5056148
Fischl, B. (2012). FreeSurfer. NeuroImage, 62(2), 774–781. DOI:
https://doi.org/10.1016/j.neuroimage.2012.01.021, PMID:
22248573, PMCID: PMC3685476
Fisher, R. A.
(1915). Frequency distribution of the values of the
correlation coefficient
in samples from an indefinitely large
population. Biometrika, 10(4), 507–521. DOI: https://doi.org/10
.2307/2331838
Fukushima, M., Betzel, R. F., He, Y., van den Heuvel, M. P., Zuo,
X. N., & Sporns, O.
(2018). Structure-function relationships
during segregated and integrated network states of human brain
functional connectivity. Brain Structure and Function, 223(3),
1091–1106. DOI: https://doi.org/10.1007/s00429-017-1539-3,
PMID: 29090337, PMCID: PMC5871577
Gleichgerrcht, E., Munsell, B., Bhatia, S., Vandergrift, W. A.,
Rorden, C., McDonald, C., . . . Bonilha, L. (2018). Deep learn-
ing applied to whole-brain connectome to determine seizure
control after epilepsy surgery. Epilepsia, 59(9), 1643–1654.
DOI: https://doi.org/https://doi.org/10.1111/epi.14528, PMID:
30098002
Glover, G. H., Li, T. Q., & Ress, D. (2000). Image-based method
for retrospective correction of physiological motion effects in
fMRI: RETROICOR. Magnetic Resonance in Medicine, 44(1),
162–167. DOI: https://doi.org/10.1002/1522-2594(200007)44:1
<162::AID-MRM23>3.0.CO;2-mi
Goñi,
J., van den heuvel, METRO. PAG., Avena-Koenigsberger, A.,
Velez de Mendizabal, NORTE., Betzel, R. F., Griffa, A., . . . despreciar,
oh. (2014). Resting-brain functional connectivity predicted by
analytic measures of network communication. Actas de
the National Academy of Sciences, 111(2), 833–838. DOI:
https://doi.org/10.1073/pnas.1315529111, PMID: 24379387,
PMCID: PMC3896172
Gracia, C. GRAMO., Yardi, r., Kattan, METRO. w., Nair, D., Gupta, A., Najm, I.,
. . . Jehi, l. (2015). Seizure freedom score: A new simple method
to predict success of epilepsy surgery. Epilepsia, 56(3), 359–365.
DOI: https://doi.org/10.1111/epi.12892, PMID: 25530458
Haneef, Z., Chiang, S., Yeh, h. J., ángel, J., & Stern, j. METRO. (2015).
Functional connectivity homogeneity correlates with duration of
Neurociencia en red
448
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
temporal lobe epilepsy. Epilepsy & Comportamiento, 46, 227–233. DOI:
https://doi.org/10.1016/j.yebeh.2015.01.025, PMID: 25873437,
PMCID: PMC4458387
Helmstaedter, C., & Kockelmann, mi.
(2006). Cognitive outcomes
in patients with chronic temporal lobe epilepsy. Epilepsia, 47,
96–98. DOI: https://doi.org/10.1111/j.1528-1167.2006.00702.x,
PMID: 17105474
Huo, y., Plassard, A. J., Carass, A., Resnick, S. METRO., Pham, D. l.,
Príncipe, j. l., & Landman, B. A. (2016). Consistent cortical recon-
struction and multi-atlas brain segmentation. NeuroImagen, 138,
197–210. DOI: https://doi.org/10.1016/j.neuroimage.2016.05
.030, PMID: 27184203, PMCID: PMC4927397
Hyvarinen, A. (1999). Fast ICA for noisy data using Gaussian mo-
mentos. ISCAS’99. Actas de la 1999 IEEE International
Symposium on Circuits and Systems VLSI, 5, 57–61. DOI:
https://doi.org/10.1109/ISCAS.1999.777510
.
. Kattan, METRO. W..
Jehi, l., Yardi, r., Chagin, K., Tassi, l., Lo Russo, GRAMO., Worrell, GRAMO.,
.
(2015). Development and validation of
nomograms to provide individualised predictions of seizure
outcomes after epilepsy surgery: A retrospective analysis. Lancet
Neurología, 14(3), 283–290. DOI: https://doi.org/10.1016/s1474
-4422(14)70325-4
Larivière, S., Weng, y., Vos de Wael, r., Royer, J., Frauscher, B.,
Wang, Z., . . . Bernhardt, B. C. (2020). Functional connectome
contractions in temporal lobe epilepsy: Microstructural under-
pinnings and predictors of surgical outcome. Epilepsia. DOI:
https://doi.org/10.1111/epi.16540, PMID: 32452574
Laxer, k. D., Trinka, MI., Hirsch, l. J., Cendes, F., Langfitt, J., Delanty,
NORTE., . . . Benbadis, S. R. (2014). The consequences of refractory
epilepsy and its treatment. Epilepsy & Comportamiento, 37, 59–70. DOI:
https://doi.org/http://dx.doi.org/10.1016/j.yebeh.2014.05.031,
PMID: 24980390
Margulies, D. S., Ghosh, S. S., Goulas, A., Falkiewicz, METRO.,
Huntenburg, j. METRO., Langs, GRAMO., . . . Smallwood, j.
(2016). Situ-
ating the default-mode network along a principal gradient of
macroscale cortical organization. Actas del Nacional
Academy of Sciences of the United States of America, 113(44),
12574–12579. DOI: https://doi.org/10.1073/pnas.1608282113,
PMID: 27791099, PMCID: PMC5098630
McHugo, METRO., Talati, PAG., Woodward, norte. D., Armstrong, K.,
Blackford, j. Ud., & Heckers, S. (2018). Regionally specific volume
deficits along the hippocampal long axis in early and chronic
psychosis. NeuroImage Clinical, 20, 1106–1114. DOI: https://
doi.org/10.1016/j.nicl.2018.10.021, PMID: 30380517, PMCID:
PMC6202690
morgan, V. l., Englot, D. J., Rogers, B. PAG., Landman, B. A., Cakir,
A., Abou-Khalil, B. w., . . . anderson, A. W.. (2017). Magnético
resonance imaging connectivity for the prediction of seizure out-
come in temporal lobe epilepsy. Epilepsia, 58(7), 1251–1260.
DOI: https://doi.org/10.1111/epi.13762, PMID: 28448683,
PMCID: PMC5498250
j.
morgan, V. l., Rogers, B. PAG., anderson, A. w., Landman, B. A.,
(2019). Divergent network properties that
& Englot, D.
failure versus late recurrence in tem-
predict early surgical
Journal of Neurosurgery, 132.
poral
DOI:
https://doi.org/10.3171/2019.1.Jns182875, PMID: 30952126,
PMCID: PMC6778487
lobe epilepsy.
morgan, V. l., Rogers, B. PAG., Sonmezturk, h. h., Sangre, j. C., &
Abou-Khalil, B. (2011). Cross hippocampal influence in mesial
temporal lobe epilepsy measured with high temporal resolu-
tion functional magnetic resonance imaging. Epilepsia, 52(9),
1741–1749. DOI: https://doi.org/10.1111/j.1528-1167.2011
.03196.X, PMID: 21801166, PMCID: PMC4428312
Oligschläger, S., Huntenburg, j. METRO., Golchert, J., Lauckner, METRO. MI.,
Bonnen, T., & Margulies, D. S. (2017). Gradients of connectivity
distance are anchored in primary cortex. Brain Structure and
Function, 222(5), 2173–2182. DOI: https://doi.org/10.1007
/s00429-016-1333-7, PMID: 27807628, PMCID: PMC5504232
Owen, t. w., de Tisi, J., vos, S. B., Winston, GRAMO. PAG., Duncan,
j. S., Wang, y., & taylor, PAG. norte. (2020). Multivariate white matter
alterations are associated with epilepsy duration. arXiv e-prints,
arXiv:2007.07643. Retrieved from https://ui.adsabs.harvard.edu
/abs/2020arXiv200707643O
Parque, C.-H., Yun Seo Choi, Y. S., Jung, A.-R., Chung, H.-K., kim,
h. J., Yoo, j. h., & Sotavento, h. W.. (2017). Seizure control and mem-
ory impairment are related to disrupted brain functional integra-
tion in temporal lobe epilepsy. The Journal of Neuropsychiatry
and Clinical Neurosciences, 29(4), 343–350. DOI: https://doi
.org/10.1176/appi.neuropsych.16100216, PMID: 28449635
Plassard, A. J., McHugo, METRO., Heckers, S., & Landman, B. A. (2017).
Multi-Scale hippocampal parcellation improves atlas-based seg-
mentation accuracy. Proceedings Volume 10133. Medical Imag-
En g 2017: Image Processing, 10133. DOI: https://doi.org/10.1117
/12.2254425, PMID: 28781411, PMCID: PMC5544133
Rubinov, METRO., & despreciar, oh.
(2010). Complex network measures
of brain connectivity: Uses and interpretations. NeuroImagen,
52(3), 1059–1069. DOI: https://doi.org/10.1016/j.neuroimage
.2009.10.003, PMID: 19819337
Herrero, R. MI., Tournier, J.-D., Calamante, F., & connelly, A. (2012).
Anatomically-constrained tractography: Improved diffusion MRI
streamlines tractography through effective use of anatomical
información. NeuroImagen, 62(3), 1924–1938. DOI: https://doi
.org/10.1016/j.neuroimage.2012.06.005, PMID: 22705374
Herrero, R. MI., Tournier, j. D., Calamante, F., & connelly, A. (2015).
SIFT2: Enabling dense quantitative assessment of brain white
matter connectivity using streamlines tractography. NeuroImagen,
119, 338–351. DOI: https://doi.org/10.1016/j.neuroimage.2015
.06.092, PMID: 26163802
Suárez, l. MI., Markello, R. D., Betzel, R. F., & Varios, B.
(2020).
Linking structure and function in macroscale brain networks.
Tendencias en Ciencias Cognitivas. DOI: https://doi.org/https://doi.org
/10.1016/j.tics.2020.01.008, PMID: 32160567
Tavakol, S., Royer, J., Lowe, A. J., Bonilha, l., Tracy, j. I., Jackson,
GRAMO. D., . . . Bernhardt, B. C. (2019). Neuroimaging and connecto-
mics of drug-resistant epilepsy at multiple scales: From focal le-
sions to macroscale networks. Epilepsia, 60(4), 593–604. DOI:
https://doi.org/10.1111/epi.14688, PMID: 30889276, PMCID:
PMC6447443
taylor, PAG. NORTE., Moreira da Silva, NORTE., Blamire, A., Wang, y., &
Forsyth, R. (2020). Early deviation from normal structural con-
necxtivity: A novel intrinsic severity score for mild TBI. Neurol-
ogia, 94(10), 10.1212/WNL.0000000000008902. DOI: https://
doi.org/10.1212/wnl.0000000000008902, PMID: 31937623,
PMCID: PMC7238920
Neurociencia en red
449
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Progression of temporal lobe epilepsy networks
taylor, PAG. NORTE., Sinha, NORTE., Wang, y., vos, S. B., de Tisi, J., Miserocchi,
A., . . . Duncan, j. S.
(2018). The impact of epilepsy surgery
on the structural connectome and its relation to outcome.
NeuroImage Clinical, 18, 202–214. DOI: https://doi.org/10.1016
/j.nicl.2018.01.028, PMID: 29876245, PMCID: PMC5987798
Tournier, j. D., Calamante, F., & connelly, A. (2007). Robust deter-
mination of the fibre orientation distribution in diffusion MRI:
Non-negativity constrained super-resolved spherical deconvolu-
ción. NeuroImagen, 35(4), 1459–1472. DOI: https://doi.org/10
.1016/j.neuroimage.2007.02.016, PMID: 17379540
Tournier, j. D., Herrero, r., Raffelt, D., Tabbara, r., Dhollander, T.,
Pietsch, METRO., . . . connelly, A. (2019). MRtrix3: A fast, flexi-
ble and open software framework for medical image processing
and visualisation. NeuroImagen, 202(116137). DOI: https://
doi.org/10.1016/j.neuroimage.2019.116137, PMID: 31473352
Uddin, l. P., yo, B. t. T., & Spreng, R. norte. (2019). Towards a uni-
versal taxonomy of macro-scale functional human brain net-
obras. Brain Topography. DOI: https://doi.org/10.1007/s10548
-019-00744-6, PMID: 31707621, PMCID: PMC7325607
van den heuvel, METRO. PAG., Mandl, R. C. w., Kahn, R. S., & Pol,
h. mi. h. (2009). Functionally linked resting-state networks re-
flect the underlying structural connectivity architecture of the hu-
man brain. Mapeo del cerebro humano, 30(10), 3127–3141. DOI:
https://doi.org/10.1002/hbm.20737, PMID: 19235882, PMCID:
PMC6870902
Vázquez-Rodríguez, B., Liu, Z.-Q., Hagmann, PAG., & Misiæ, B.
(2020). Signal propagation via cortical hierarchies. bioRxiv,
2020.2002.2015.950907. DOI: https://doi.org/10.1101/2020
.02.15.950907
Wang, h., Huang, y., Coman, D., Munbodh, r., Dhaher, r., Zaveri,
h. PAG., . . . Eid, t.
(2017). Network evolution in mesial tempo-
ral lobe epilepsy revealed by diffusion tensor imaging. Epilepsia,
58(5), 824–834. DOI: https://doi.org/10.1111/epi.13731, PMID:
28378878, PMCID: PMC5429866
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
2
4
3
4
1
9
1
3
5
1
1
norte
mi
norte
_
a
_
0
0
1
8
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurociencia en red
450