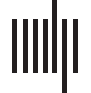INVESTIGACIÓN
Detecting modular brain states in rest and task
Aya Kabbara
1,2,3
Youssof El Traboulsi
, Mohamad Khalil
7
, Georges O’Neill
2
, Fabrice Wendling
, and Mahmoud Hassan
4
, Kathy Dujardin
5,6
,
2
1,3
1Azm Center for Research in Biotechnology and Its Applications, EDST, Lebanese University, Beirut, Líbano
2University of Rennes, LTSI – U1099, Rennes, Francia
3CRSI Lab, Engineering Faculty, Lebanese University, Beirut, Líbano
4Sir Peter Mansfield Imaging Centre, Escuela de Física y Astronomía, University of Nottingham,
University Park, Nottingham, Reino Unido
5INSERM, U1171, Lille, Francia
6CHU Lille, Neurology and Movement Disorders Department, Lille, Francia
7LaMA-Liban, Lebanese University, Tripoli, Líbano
un acceso abierto
diario
Palabras clave: Brain network dynamics, Conectividad funcional, M/EEG source-space networks
ABSTRACTO
The human brain is a dynamic networked system that continually reconfigures its functional
connectivity patterns over time. De este modo, developing approaches able to adequately detect fast
brain dynamics is critical. Of particular interest are the methods that analyze the modular
structure of brain networks, eso es, the presence of clusters of regions that are densely
interconnected. en este documento, we propose a novel framework to identify fast modular states
that dynamically fluctuate over time during rest and task. We started by demonstrating the
feasibility and relevance of this framework using simulated data. Compared with other
methods, our algorithm was able to identify the simulated networks with high temporal and
spatial accuracies. We further applied the proposed framework on MEG data recorded during
a finger movement task, identifying modular states linking somatosensory and primary motor
regiones. The algorithm was also performed on dense-EEG data recorded during a picture
naming task, revealing the subsecond transition between several modular states that relate to
visual processing, semantic processing, and language. Próximo, we tested our method on a
dataset of resting-state dense-EEG signals recorded from 124 patients with Parkinson’s
enfermedad. Results disclosed brain modular states that differentiate cognitively intact patients,
patients with moderate cognitive deficits, and patients with severe cognitive deficits. Nuestro
new approach complements classical methods, offering a new way to track the brain
modular states, in healthy subjects and patients, on an adequate task-specific timescale.
RESUMEN DEL AUTOR
The brain is a dynamic modular network. De este modo, exploring the dynamic behavior of the brain
can reveal insights about its characteristics during rest and task, in healthy and pathological
condiciones. en este documento, we propose a new framework that aims to track the dynamic
changes of the modular brain organization. The method presents two algorithms that can be
applied during task-free or task-related paradigms. Using simulations, we demonstrated the
advantages of the proposed algorithm over existing methods. The proposed algorithm was
also tested on three datasets recorded from real EEG and MEG acquisitions. En general, resultados
showed the capacity of the proposed algorithm to track brain network dynamics with good
spatial and temporal accuracy.
Citación: Kabbara, A., Khalil, METRO.,
O’Neill, G.,Dujardin, K., El Traboulsi, y.,
Wendling, F. & Hassan, METRO. (2019).
Detecting modular brain states in rest
and task. Neurociencia en red, 3(3),
878–901. https://doi.org/10.1162/
netn_a_00090
DOI:
https://doi.org/10.1162/netn_a_00090
Supporting Information:
https://doi.org/10.1162/netn_a_00090
https://github.com/librteam/
Modularity_algorithm_NN
Recibió: 3 Enero 2019
Aceptado: 18 Abril 2019
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Aya Kabbara
aya.kabbara7@gmail.com
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2019
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Detecting modular brain states in rest and task
INTRODUCCIÓN
The human brain is a modular dynamic system. Following fast neuronal activity (Pfurtscheller &
Aranibar, 1977; Pfurtscheller & Lopes Da Silva, 1999), the spatiotemporal organization of rest-
En g (Baker et al., 2014; Damaraju et al., 2014; de Pasquale et al., 2012; de Pasquale, Della Penna,
despreciar, Romani, & Corbetta, 2015; Kabbara, El Falou, Khalil, Wendling, & Hassan, 2017) y
task-evoked connectivity (Bola & Sabel, 2015; Hassan, Benquet, et al., 2015; Hutchison et al.,
2013; O’Neill, Tewarie, Colclough, et al., 2017) is in constant flux.
Por eso, an appropriate description of time-varying connectivity is of utmost importance
to understand how cognitive and behavioral functions are supported by networks. En esto
contexto, several frameworks have been suggested to explore the time-varying nature of func-
tional brain connectivity. entre ellos, hidden Markov model approaches (Baker et al., 2014;
Vidaurre et al., 2017), K-means clustering (allen, Damaraju, Eichele, Wu, & Calhoun, 2017;
Allen et al., 2014; Damaraju et al., 2014), independent component analysis (ICA; O’Neill,
Tewarie, Colclough, et al., 2017), principal component analysis (Preti & Van De Ville, 2016),
or orthogonal connectivity factorization (Hyvärinen, Hirayama, Kiviniemi, & Kawanabe, 2016)
have been applied to identify the main brain networks shaping dynamic functional connec-
actividad. These methods consist of grouping the temporal networks into states, where each state
reflects a unique spatial connectivity pattern. It is important to note that in these frameworks,
states are identified without looking at the modular organization of the networks.
Sin embargo, because of the modular organization of the human brain network (despreciar &
Betzel, 2016) methods for detecting network communities (or modules) are of particular inter-
est (despreciar & Betzel, 2016). These methods decompose the network into building blocks or
modules that are internally strongly connected, often corresponding to specialized functions.
En tono rimbombante, during a learning task, Basset et al. showed that the flexibility (defined as how
often a given node changes its modular affiliation over time) of the networks facilitates the
prediction of individual future performances in next learning sessions (Bassett y cols., 2011),
remembering, atención, and integrated reasoning (Gallen et al., 2016).
To compute modularity in networks collected across multiple slices (time points), one could
simply decompose each network independently. Another algorithm proposed to link corre-
sponding nodes across slices before generating communities. This algorithm is known as
multislice modularity (also called multilayer modularity; Mucha, Richardson, Macon, Portero,
& Onnela, 2010), which has been used to follow changes of the modular architecture in differ-
ent applications (Bassett y cols., 2011; bassett, Cual, Wymbs, & Grafton, 2015). Sin embargo, el
problem with solely applying these techniques is that they may not lead to easily interpretable
results because of the particularly high number of modular structures generated. More specifi-
cally, the dynamic modular structures obtained are technically difficult to visualize since their
number is equal to the number of slices. Alternativamente, one could use the multislice modularity
algorithm to generate a community structure consistent over time. But such a strategy will con-
strain the dynamic analysis. Por eso, it would be very useful to find the main modular structures
driving the dynamics of neural activity. Hasta la fecha, there is no automatic framework that is able
to decipher modular brain states, eso es, subsets of brain modules implicated in a given brain
function at a given time period.
Aquí, we propose a novel framework aiming to elucidate the main modular brain struc-
turas, called modular states (MS), that fluctuate over time during rest and task. We attempt
to find the modular structures that share the same topology by quantifying the similarity be-
tween all the computed temporal partitions (Cifra 1). The proposed framework simply takes
Dynamic functional connectivity:
Measurement of transient changes in
the functional brain network across
tiempo.
Module:
A subnetwork composed of strongly
connected nodes that are weakly
connected to the rest of the network.
Modular structure:
The partition of the network into
nonoverlapping modules.
Brain state:
A network pattern representing time
points that share some similar
topological properties.
Neurociencia en red
879
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Detecting modular brain states in rest and task
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. The algorithm procedure. (A) Computation of modules for each temporal network (Wt).
(B) Assessment of the similarity between the dynamic modular structures. (C) Clustering the similarity
matrix into “categorical modules.”
as input a set of connectivity matrices, without making any constraint on how these matrices
are computed. This means that it is independent of the choice of the connectivity method, el
frequency range of interest, the temporal resolution of dynamic connectivity, and the regions
de interés (ROI). Connectivity matrices could be also extracted from different individuals, o
different experimental conditions. The main purpose of the framework is to find the common
modular states between the connectivity patterns interpreted. In the paper, we used simulated
data to show the advantages of our method over other existing methods in terms of spatial and
temporal accuracy. Entonces, using three different MEG/EEG datasets, recorded in rest and task
from healthy subjects and/or patients, we demonstrate the feasibility of the method to reveal
both time-varying functional connectivity and the corresponding spatial patterns, as compared
with the state-of-the art findings. En general, findings show that the algorithm was able to track
the dynamics of brain networks in adapted timescale, offering a new way to explore the spa-
tiotemporal organization of brain function.
880
Conectividad funcional:
Statistical dependency between
spatially separated brain regional
time series.
Neurociencia en red
Detecting modular brain states in rest and task
MATERIALES Y MÉTODOS
We have developed two versions of the algorithm: (a) “categorical,” where we aim to find the
main modular structures over time, with no interest in their sequential order; y (b) “con-
secutive,” where the objective is to find the modular structures in a successive way. The two
versions are described hereafter.
Categorical Version
The categorical version includes three main steps:
(1) Decompose each temporal network into modules (es decir., clusters of nodes that are inter-
nally strongly connected, but externally weakly connected; Figura 1A). To do that, diferente
modularity algorithms were proposed in the literature (rubia, Guillaume, Lambiotte,
& Lefebvre, 2008; Duch & Arenas, 2005; girvan & Hombre nuevo, 2002; Guimerà & Amaral,
2005).
In our study, we adopted the consensus modularity approach that was previously used in
many studies (Bassett y cols., 2013; Kabbara et al., 2017): Given an ensemble of partitions
acquired from the Newman algorithm (girvan & Hombre nuevo, 2002) and Louvain algorithm
(Blondel et al., 2008) repeated for 200 carreras, an association matrix is obtained. This results
in an N × N matrix (N is the number of nodes), and an element Ai,j represents the
number of times the nodes i and j are assigned to the same module across all runs
and algorithms. The association matrix is then compared with a null model association
matrix generated from a permutation of the original partitions, and only the significant
values are retained (Bassett y cols., 2013). To ultimately obtain consensus communities,
we reclustered the association matrix using the Louvain algorithm.
(2) Assess the similarity between the temporal modular structures (Figura 1B). In this context,
several methods have been suggested to compare community structures (Traud, Kelsic,
Mucha, & Portero, 2008). Aquí, we focused on the pair-counting method, which defines
a similarity score by counting each pair of nodes drawn from the N nodes of a network
according to whether the pair falls in the same or in different groups in each partition
(Traud et al., 2008). We considered the z-score of Rand coefficient, bounded between
0 (no similar pair placements) y 1 (identical partitions). This yields a T × T similarity
matrix, where T is the number of time windows.
(3) Cluster the similarity matrix into “categorical” modular states (MS) using the consensus
modularity method (Figura 1C). This step combines similar temporal modular structures
in the same community. Por eso, the association matrix of each “categorical” community
is computed using the modular affiliations of its corresponding networks.
Consecutive Version
The difference between the two versions of the algorithm is essentially in the third step, en
which the final communities were defined. Particularly, the similarity matrix is segmented in a
sequential way using the following steps:
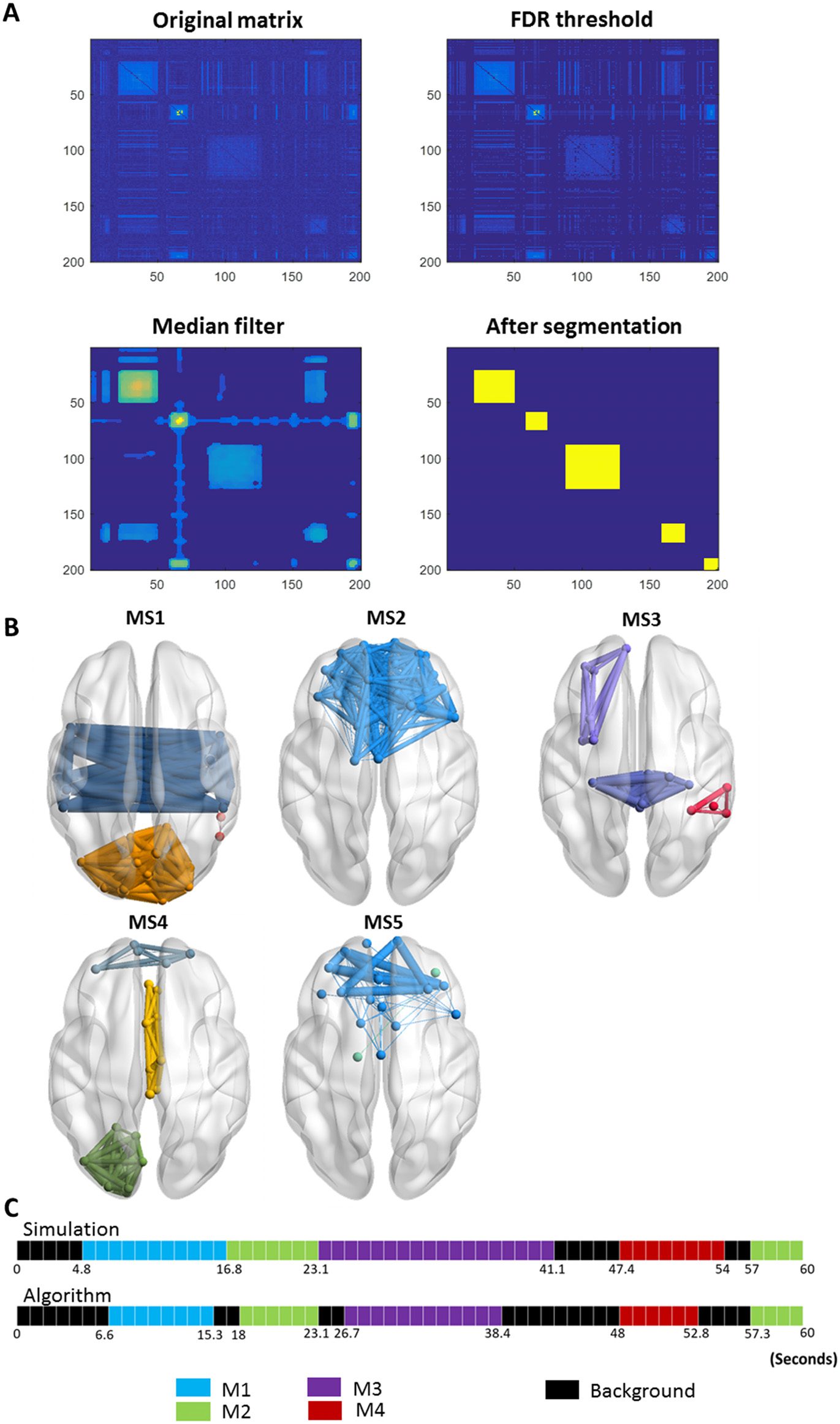
(1) Threshold the similarity matrix using an automatic thresholding algorithm described in
Genovese, Lázaro, and Nichols (2002). Briefly the matrix was converted into a p value
map that is then thresholded based on the false discovery rate (FDR) controlling.
(2) Apply a median filter in order to get a smoother presentation of the similarity matrix.
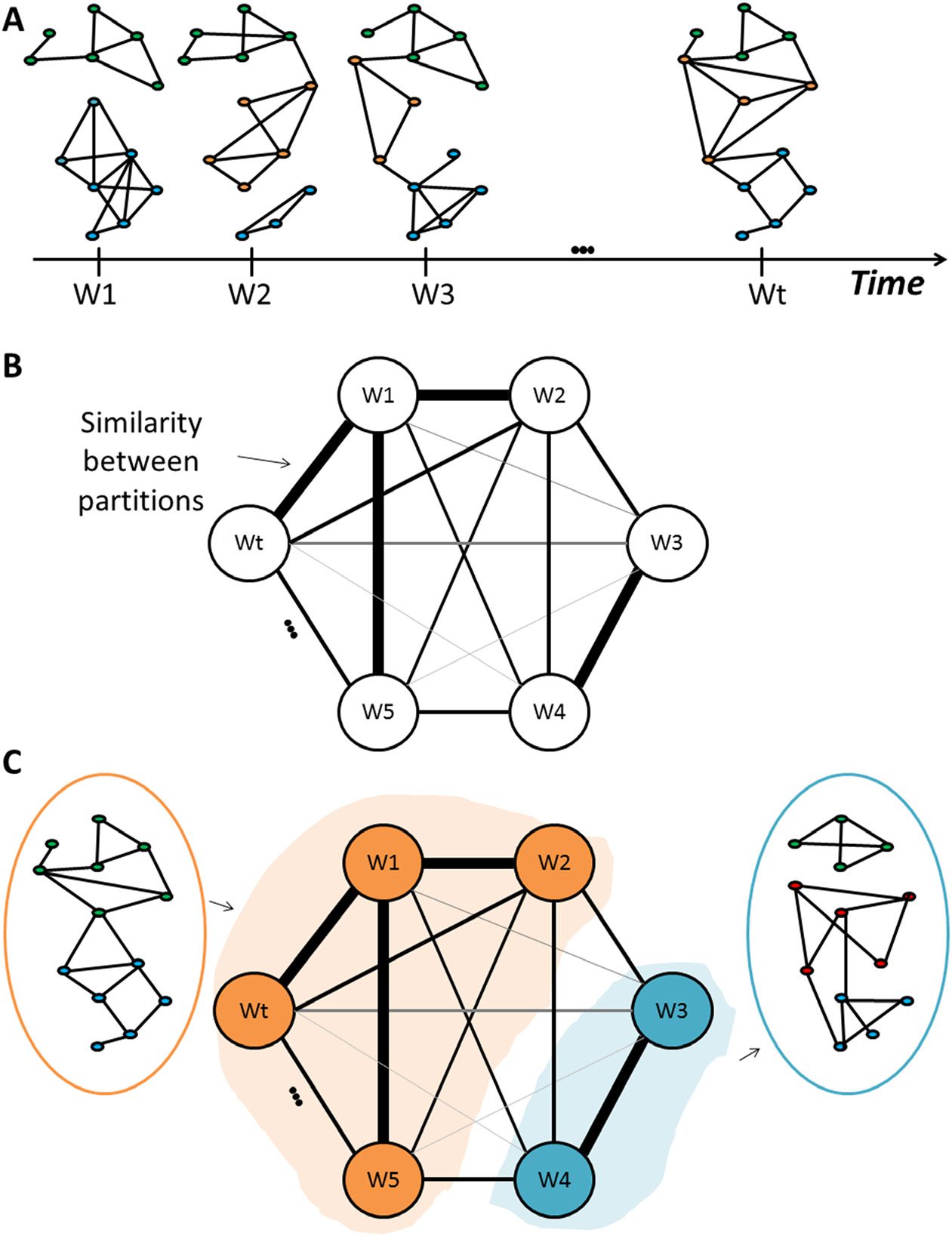
(3) Segment the matrix in a sequential way following the algorithm illustrated in the flowchart
of Figure 2. In brief, the method groups similar consecutive modular structures. As these
Neurociencia en red
881
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Detecting modular brain states in rest and task
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2.
Flowchart of the segmentation algorithm.
modular structures show high similarity values with each other, the algorithm detects the
squares located around the diagonal of the similarity matrix. As presented in Figure 2,
the condition for which two consecutive structures are associated with the same state is
the following:
Y,j > a,
where Si,j = ∑
j
k=0 ai+k,i+j − ai+j,i+j ; i, j ∈ [1, t], where al,m denotes the
similarity value between the modular structure corresponding to the time window l and that
corresponding to the time window m.
j
k=0 ai+j,i+k + ∑
a is the “accuracy parameter,” strictly bounded between 0 y 1. It regulates the temporal-
spatial accuracy of detected modular states. We recommend choosing an adaptive value of
a. en este documento, a equals the average of the similarity matrix. A segment is considered to be
relevant if the number of included time windows is greater than jmin (the minimal size allowed
for a segment).
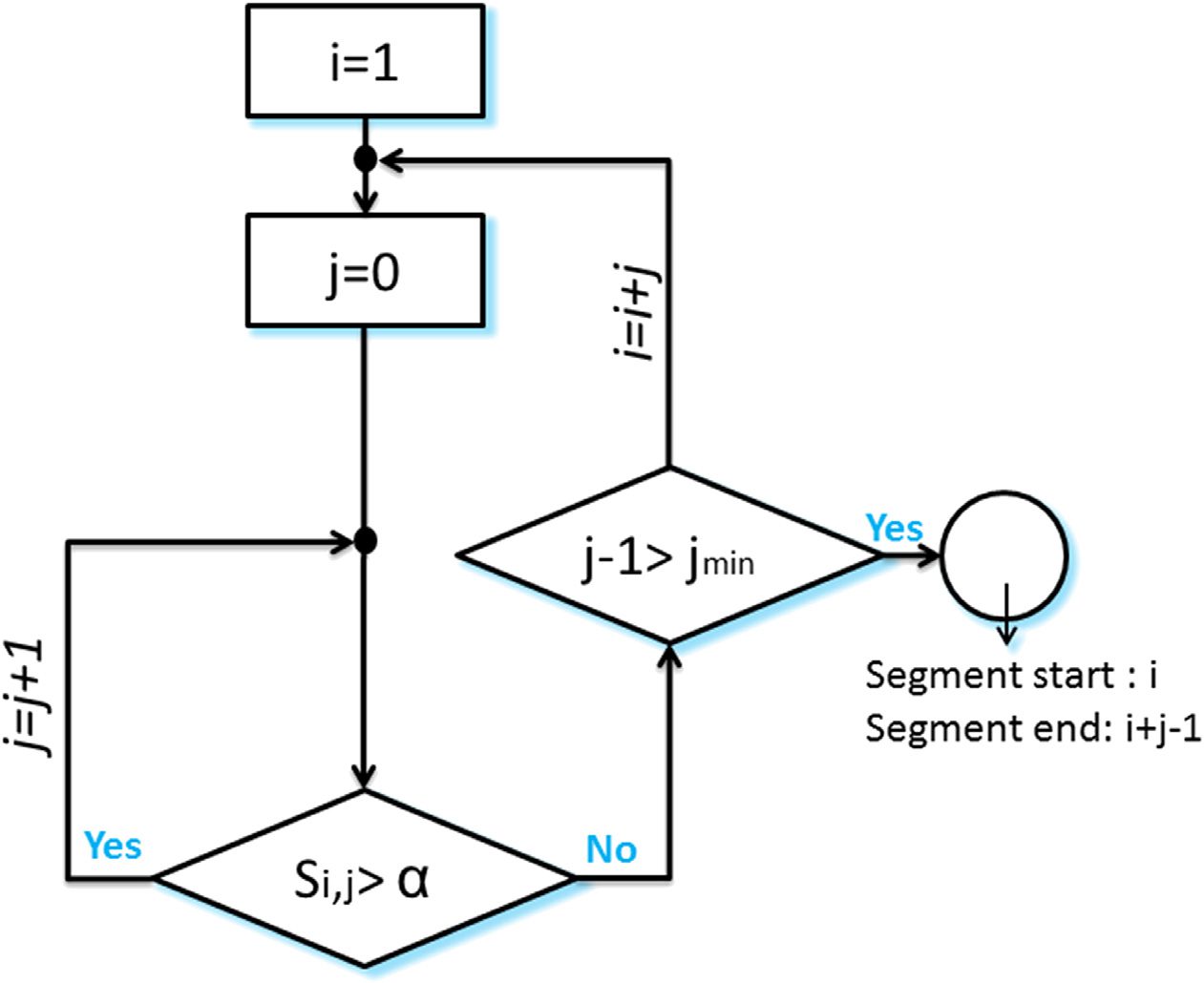
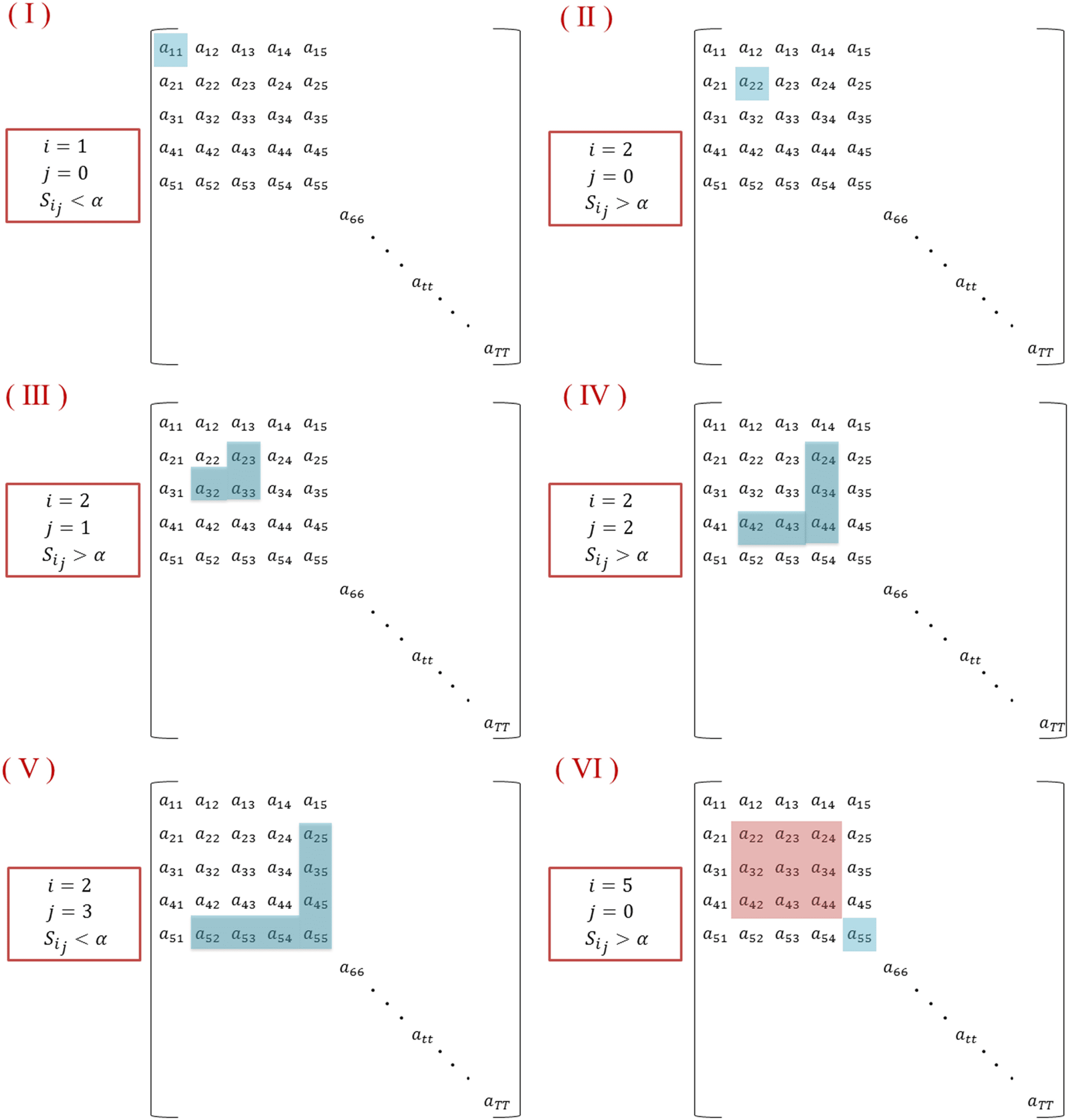
The algorithm is illustrated in Figure 3. (I): Starting with i = 1, j = 0; and considering that a11
is lower than a, we obtained S1,0 = a11 < a, and the algorithm moves to the next time window
i = 2. (II): As S2,0 = a22 is greater than a, j is incremented by 1. (III): Having S2,1 = a23+a23+a33
>
a, the second and the third time windows are associated with the segment. (IV): The algorithm
succeeds to add also the fourth time window as S2,2 = a24+a42+a34+a43+a44
> a. (V): Entonces, para
j = 3, we obtain S2,3 = a25+a35+a45+a55+a52+a53+a54
< a. This means that the fifth time window
differs from the previous windows in its modular structure. Afterwards, the algorithm moves
toward finding another segment by incrementing i and repeating the process (IV).
3
5
7
For each detected segment, the modular structure is obtained after computing the associa-
tion matrix of the corresponding time windows modular affiliations.
Network Neuroscience
882
Detecting modular brain states in rest and task
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. An illustrative example describing the segmentation algorithm.
Network Neuroscience
883
Detecting modular brain states in rest and task
The Matlab code developed to apply the algorithm in the two versions is publicly available
(Kabbara et al., 2019).
Simulated Data
We simulated the adjacency tensor data following the methodology applied in O’Neill, Tewarie,
Colclough, et al. (2017). Briefly, four N × N adjacency matrices Pj were constructed, where
j ∈ [1, 4] and N is the number of ROIs. We used an anatomical atlas of 221 ROIs with the
mean of the Desikan-Killiany atlas (Desikan et al., 2006) subdivided by Hagmann et al. (2008),
yielding to N = 221. The adjacency matrices and the 3-D visualization of the networks are
presented in Figure 4A. Following this step, the time evolution of dynamic connectivity in each
network is given by the following:
Mj(t) = a. f1j(t) + b. f2j(t);
f1j(t) is the modulation function, which was represented by the Hanning window of unit
amplitude; f2j(t) represents uncorrelated Gaussian noise added to the simulated time courses;
and a and b are scalar values set to 0.45 and 0.15 as in O’Neill, Tewarie, Colclough, et al.
(2017).
In our study, Mj(t) is sampled at 3.3 Hz (to obtain a sliding window of 0.3 s as in real
data). The onset as well as the duration of each module structure is illustrated in Figure 4B.
We then combined the four network matrices in order to generate a single adjacency matrix
at each time point t over a time course spanning 60 s. As a final step, we added a random
Gaussian noise to the adjacency tensor, and the standard deviation of the noise was allowed
to vary between 0.2 and 0.5.
Validation
On simulated data, we evaluated the performance of the algorithm by computing the simi-
larity between the reconstructed and the simulated (reference) networks, taking into account
both spatial and temporal similarities. The spatial similarity is given by the z-score of Rand
coefficient between the simulated and the constructed modular structures, while the temporal
similarity represents the rate of the correct affiliation of time windows.
In addition, we have compared our algorithm with three methods originally developed
to identify the brain states without looking at their modular structures. The three methods
are K-means clustering as applied by Allen et al. (2014), independent component analysis as
suggested by O’Neill, Tewarie, Colclough, et al. (2017), and the consensus clustering algorithm
proposed by Rasero et al. (2017). For each method, we followed the same pipeline adapted
by the authors. To ultimately obtain the modular brain states, we preceded the analysis by
decomposing each connectivity state to modules using the consensus modularity approach as
detailed in the Categorial Version section.
Real Data
In order to track the brain network dynamics effectively, we particularly tested our method
using electro- and magnetoencephalography (EEG/MEG), which allow the tracking of brain
dynamics on a subsecond timescale, a resolution not reachable using other techniques such
as functional magnetic resonance imaging (fMRI; Cohen, 1972; Nunez & Srinivasan, 2007;
Penfield & Jasper, 1954).
Network Neuroscience
884
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. The simulation scenario. (A) Left: The adjacency matrix Pj of the constructed networks.
Right: The 3-D cortical presentation of the modular structures of the simulated networks. (B) The time
axis showing the beginning and the end of each network Mj (which was generated by combining
Pj to uncorrelated noise).
Thus, three M/EEG datasets were analyzed. Before applying the algorithm, we reconstucted
the functional brain networks at the level of the cortex using an emerging technique called EEG
source connectivity (Chavez et al., 2013; De Vico Fallani et al., 2008; De Vico Fallani, Latora,
& Chavez, 2017; De Vico Fallani, Richiardi, Chavez, & Achard, 2014; Hassan, Shamas, Khalil,
Network Neuroscience
885
Detecting modular brain states in rest and task
El Falou, & Wendling, 2015; Hassan & Wendling, 2018; Kabbara et al., 2017; Lai, Demuru,
Hillebrand, & Fraschini, 2018; Schoffelen & Gross, 2009). In order to remove any bias from
our analysis, we have generated the dynamic functional connectivity matrices according
to how they were computed in the three previous studies that interpreted the data (Hassan,
Benquet, et al., 2015; Hassan et al., 2017; O’Neill, Tewarie, Colclough, et al., 2017) The aim
of this work is to test whether the modular states generated by our algorithm match the findings
obtained in the literature.
Previously used in O’Neill
Dataset 1: Self-paced motor task for healthy participants (MEG data).
et al. (2015); O’Neill, Tewarie, Colclough, et al. (2017), and Vidaurre et al. (2016), this dataset
includes 15 participants (9 male, 6 female) asked to press a button using the index finger of their
dominant hand, once every 30 s. Using a 275-channel CTF MEG system (MISL; Coquitlam,
BC, Canada), MEG data were recorded at a sampling rate of 600 Hz. MEG data were coreg-
istered with a template MRI. The cortex was parcellated using the Desikan-Killiany atlas (68
regions; Desikan et al., 2006). The preprocessing, the source reconstruction, and the dynamic
functional connectivity computations were performed similarly to those in O’Neill, Tewarie,
Colclough, et al. (2017). Briefly, the preprocessing comprises the exclusion of trials (t =
−12s → 12s) contaminated by noise. Then, source time courses were reconstructed using
a beamforming approach (please refer to O’Neill, Tewarie, Colclough, et al., 2017, for more
details). Afterwards, the regional time series were symmetrically orthogonalized following the
method proposed in Colclough, Brookes, Smith, & Woolrich (2015) to remove the effects of
“signal leakage.” The amplitude envelopes of the time courses were obtained using a Hilbert
transform. Finally, the dynamic connectivity was estimated by the Pearson correlation measure
using a sliding window approach of 6 s of length. The sliding window was shifted by 0.5 s over
time. The number of connectivity matrices obtained for each trial was then 49. A statistical
threshold (FDR) approach was used to threshold the matrices (Genovese, Lazar, & Nichols,
2002). This yields to a set of connectivity matrices that are thresholded and weighted. Then,
the consecutive scheme of the proposed method was tested.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Twenty-one right-handed
Dataset 2: Picture naming task for healthy participants (dense-EEG data).
healthy subjects (11 women and 10 men) with no neurological disease participated in this
study. In a session of about 8 min, each participant was asked to name 148 displayed pictures
on a screen using E-Prime 2.0 software (Psychology Software Tools, Pittsburgh, PA; Schneider,
Eschman, & Zuccolotto, 2002). Oral responses were recorded to set the voice onset time. This
study was approved by the National Ethics Committee for the Protection of Persons (CPP),
ConneXion study, agreement number 2012-A01227-36, and promoter, Rennes University Hos-
pital. All participants provide their written informed consent to participate in this study. A typi-
cal trial started with the appearance of an image during 3 s followed by a jittered interstimulus
interval of 2 or 3 s randomly. Errors in naming were discarded from the analysis. A total of
2,926 on 3,108 events were considered.
Dense-EEG data were recorded using a system of 256 electrodes (EGI, Electrical Geodesic,
Inc.). EEGs were collected at 1 kHz sampling frequency and bandpass filtered between 3 and
45 Hz. The preprocessing and the computation of the functional connectivity followed the
same pipeline applied in Hassan, Benquet, et al. (2015). In brief, each trial (t = 0 → 600 ms)
was visually inspected, and epochs contaminated by eye blinks, muscle movements, or other
noise sources were rejected. As described in the previous study (Hassan, Benquet, et al., 2015),
the source connectivity method was performed using the wMNE/PLV (phase locking value)
combination, and the dynamic functional connectivity was computed at each millisecond.
886
Phase locking value:
A measure used to quantify the
statistical coupling, in term of phase
synchrony, between two time series.
Network Neuroscience
Detecting modular brain states in rest and task
Resting state:
An experimental paradigm in which
the subject is asked to relax and to
do nothing.
Authors also used the Destrieux atlas subdivided into 959 regions (Hassan, Benquet, et al.,
2015). To remove spurious connections from the dynamic connectivity matrices, we have
adopted a statistical threshold based on FDR (Genovese et al., 2002). Finally, a weighted and
thresholded connectivity tensor of dimension 959 × 959 × 600 was obtained and analyzed
using the consecutive scheme of our algorithm.
This dataset includes
Dataset 3: Resting state in Parkinson’s disease patients (dense-EEG data).
124 patients with idiopathic Parkinson’s disease defined according to the UK Brain Bank cri-
teria for idiopathic Parkinson’s disease (Gibb & Lees, 1988). These patients were separated
into three groups: (G1) cognitively intact patients (N = 63); (G2) patients with mild cognitive
deficits (N = 46); and (G3) patients with severe cognitive impairment (N = 15). All partic-
ipants gave their informed consent to participate in the study, which had been approved by
the local institutional review boards (CPP Nord-Ouest IV, 2012-A 01317-36, ClinicalTrials.gov
Identifier: NCT01792843). Dense-EEG were recorded with a cap (Waveguard, ANT software
BV, Enschede, Netherlands) with 122 scalp electrodes distributed according to the interna-
tional system 10-05 (Oostenveld & Praamstra, 2001). Electrode impedance was kept below
10 kΩ. Patients were asked to relax without performing any task. Signals were sampled at
512 Hz and bandpass filtered between 0.1 and 45 Hz.
The data were preprocessed according to Hassan et al. (2017) dealing with the same dataset.
Briefly, EOG artifact detection and correction was applied following the method developed
in Gratton, Coles, & Donchin (1983). Afterwards, epochs with voltage fluctuation between
90 μV and −90 μV were kept. For each participant, two artifact-free epochs of 40-s lengths
were selected. This epoch length was used previously and considered as a good compromise
between the needed temporal resolution and the reproducibility of the results in resting state
(Kabbara et al., 2017).
To compute the dynamic functional connectivity, the steps adopted here are the same used
in many previous studies (Hassan et al., 2017; Kabbara et al., 2018, 2017). First, EEG data were
coregistered with a template MRI through identification of the same anatomical landmarks (left
and right preauricular points and nasion). Second, the lead field matrix was computed for a
cortical mesh with 15,000 vertices using the OpenMEEG package (Gramfort, Papadopoulo,
Olivi, & Clerc, 2010) available in Brainstorm. The noise covariance was estimated using a
1-min resting segment. After that, the time series of EEG sources were estimated using the
wMNE algorithm where the regularization parameter was set according to the signal to noise
ratio (λ = 0.1 in our analysis). An atlas-based segmentation approach was used to project
EEGs onto an anatomical framework consisting of 68 cortical regions identified by means of
the Desikan-Killiany atlas (Desikan et al., 2006). The dynamic functional connectivity was
then computed using a sliding window over which PLV was calculated (Lachaux, Rodriguez,
Martinerie, & Varela, 1999). In the previous study (Hassan et al., 2017), the disruptions of the
functional connectivity were found in the alpha 2 band (10–13 Hz). For this reason, we con-
sidered the same frequency band in our analysis. To obtain a sufficient number of cycles at the
6
given frequency band, we chose the smallest window length that is equal to
central frequency ,
as recommended in Lachaux et al. (1999). This yields a sliding window of 0.52 s. We then
adopted a proportional threshold of 10% to maintain only the top 10% of correlation values
of the connectivity matrix. These steps produce, for each epoch, a weighted thresholded con-
nectivity tensor of dimension N × N × T, where N is the number of ROIs (68 regions) and
T is the number of time windows (77 time windows). This tensor is formally equivalent to
dynamic functional connectivity matrices, and it was analyzed using the categorical version
of the proposed algorithm.
Network Neuroscience
887
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
RESULTS
Having the set of connectivity matrices, we first applied a community detection algorithm
(Louvain method) to decompose each network into modules (i.e., clusters of nodes that are
internally strongly connected, but externally weakly connected). The similarity between the
temporal modular structures was calculated (Figure 1B). Finally, the modular states (MS) were
obtained by applying a community detection algorithm to the similarity matrix (Figure 1C).
We propose two different frameworks: (a) “categorical,” where the objective is to find the
main modular structures, without any interest in their sequential order; and (b) “consecutive,”
where the objective is to find the modular structures in a successive way.
Validation on Simulated Data
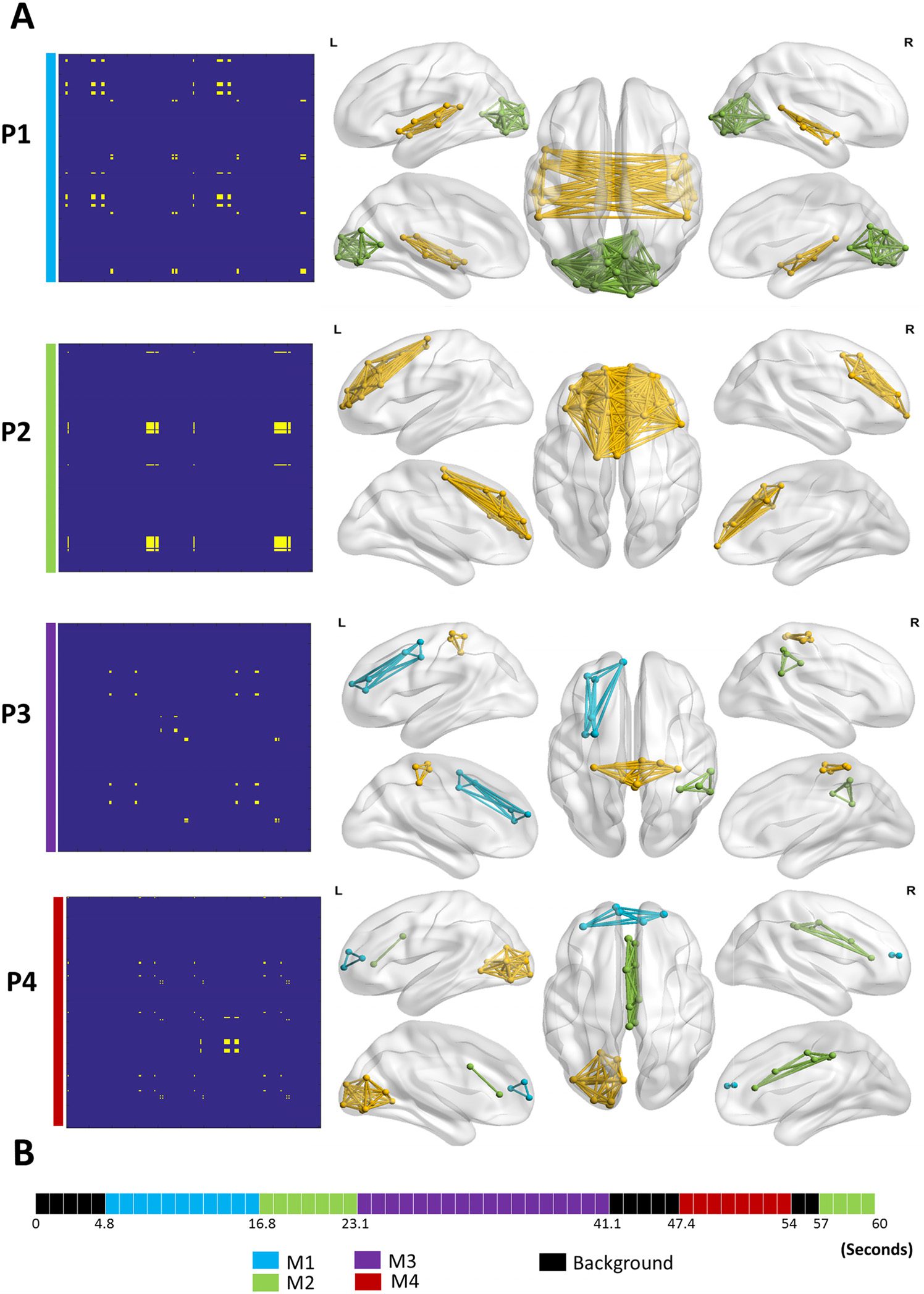
Figure 5 shows the results of the categorical method applied on the dynamic networks gen-
erated by the simulation scenario (STDnoise = 0.2). Four modular states were obtained: MS1,
MS2, MS3, and MS4. Figure 5A illustrates the modular states’ time courses, showing the most
likely state at each time window, while Figure 5B shows the 3-D representation of the MS. Qual-
itatively, the four simulated modular structures have been successfully reconstructed. However,
one time window that actually belongs to the background (i.e., random) has been wrongly
affiliated with MS2. Moreover, the MS3 state time course presented two false time window
detections: one belongs to MS4, and the other belongs to the background. To quantitatively
validate the obtained results, we compared the simulated structures (M1, M2, M3, and M4)
with the reconstructed structures (MS1, MS2, MS3, and MS4) in terms of spatial and temporal
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Results of the categorical method applied on simulated data. (A) The time course of the
four modular structures reconstructed. The gray square indicates false time window detection. (B)
The 3-D representation of the four modular structures’ states.
Network Neuroscience
888
Detecting modular brain states in rest and task
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Results of the consecutive method applied on simulated data. (A) The results of the
segmentation algorithm used to derive the consecutive modular structures from the similarity matrix
by (1) thresholding the matrix using FDR, (2) applying a median filter on the thresholded matrix,
and (3) extracting the most significant segments. (See the Methods section for more details about the
consecutive algorithm steps.) (B) The 3-D representation of the five consecutive modular structures
obtained. (C) The difference between the simulated time axis and the obtained time axis.
Network Neuroscience
889
Detecting modular brain states in rest and task
similarities. The spatial similarities between the simulated and the reconstructed data are 0.99,
0.98, 0.99, and 0.98 for MS1, MS2, MS3, and MS4, respectively. The temporal similarities are
0.79, 0.83, 0.9, and 0.71 for MS1, MS2, MS3, and MS4, respectively. On average, the cate-
gorical method achieved spatial accuracy of 98.5% and a temporal accuracy of 80.8%.
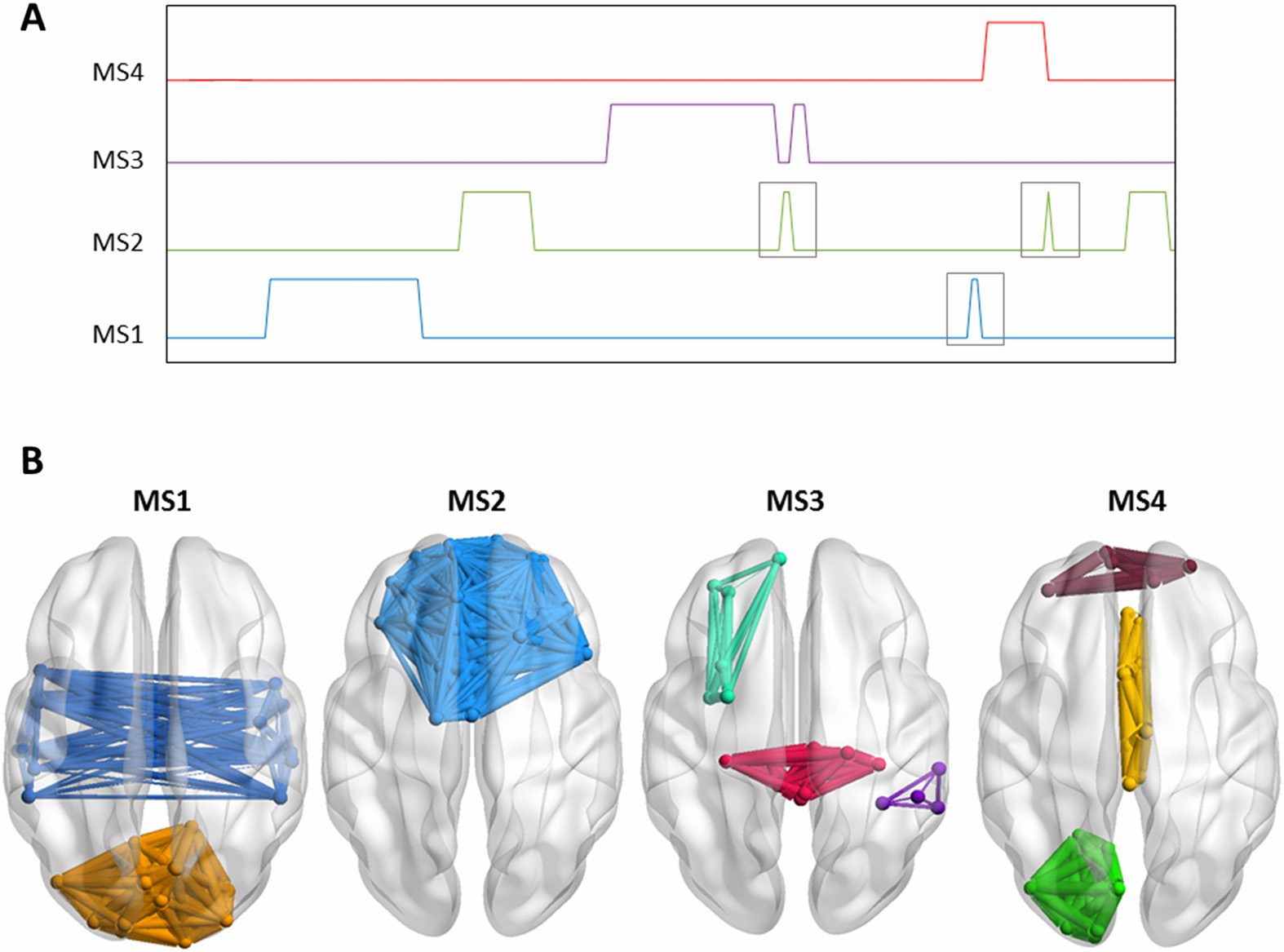
Using the consecutive method, the algorithm has segmented the similarity matrix yielding
to the detection of five modular states (Figure 6A). Their 3-D representations are shown in
Figure 6B. One can remark that MS1 (spatial similarity = 0.94; temporal similarity = 0.88), MS2
(spatial similarity = 0.99; temporal similarity = 0.94), MS3 (spatial similarity = 0.97; temporal
similarity = 0.77), MS4 (spatial similarity = 1; temporal similarity = 0.88), and MS5 (spatial
similarity = 0.95; temporal similarity = 1) matched, temporally and spatially, the simulated
networks generated at the corresponding time windows. In addition, one can remark that the
algorithm hasn’t produced false positive results. Globally, the assessed spatial and temporal
accuracies are 94% and 83%, respectively.
Results corresponding to STDnoise = 0.35; STDnoise = 0.5 are illustrated in the Supporting
Information. In brief, results show that using the categorical algorithm, the spatial characteriza-
tions of the four modular states were successfully detected. However, the state time course of
MS3 failed to detect the second corresponding segment (Figures 1 and 2 in the Supporting In-
formation). Using the consecutive algorithm, the five MSs were temporally detected (Figures 3
and 4 in the Supporting Information).
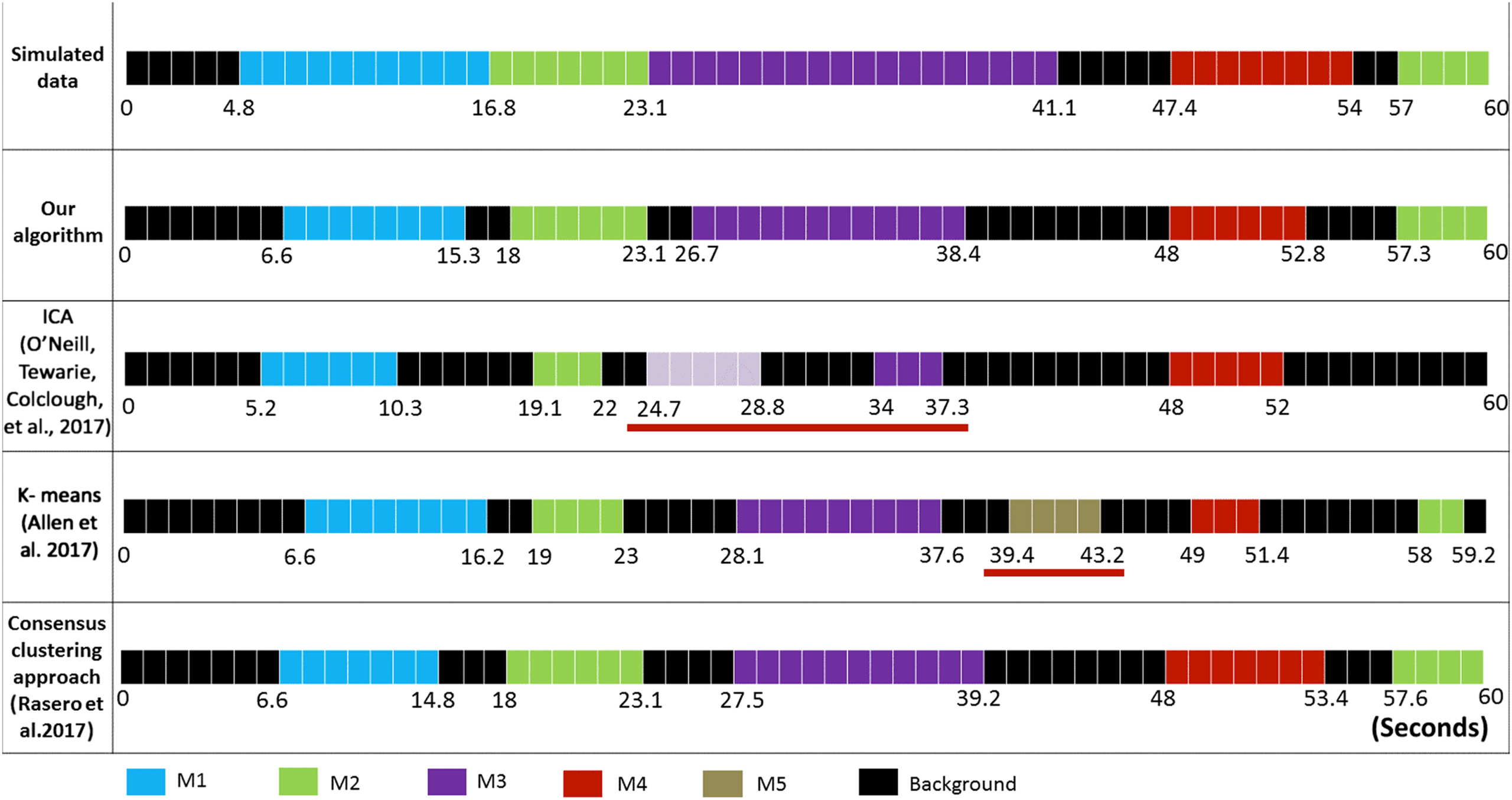
The difference between the simulated time axis and the obtained time axis using other differ-
ent algorithms is presented in Figure 7. Using ICA (O’Neill, Tewarie, Colclough, et al., 2017),
five independent components (ICs) were detected: IC1, IC2, and IC5 correspond, respectively,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. The difference between the simulated time axis and the obtained time axis using the different algorithms.
Network Neuroscience
890
Detecting modular brain states in rest and task
Table 1. Temporal similarities between the simulated MSs and those obtained using the different algorithms
Algorithm
Our algorithm
ICA (O’Neill, Tewarie, Colclough,
et al., 2017)
K-means
(Allen et al., 2017)
Consensus clustering approach
(Rasero et al., 2017)
MS1
0.73
0.43
MS2
0.98
0.6
MS3
0.67
0.41
MS4
0.86
0.71
MS5
0.93
0
0.8
0.77
0.53
0.61
0.55
0.68
0.98
0.65
0.79
0.85
False positive rate
0
0
0.07
0
Average temporal similarity
0.83
0.43
0.58
0.79
to M1 (spatial similarity = 0.96; temporal similarity = 0.43), M2 (spatial similarity = 0.85;
temporal similarity = 0.60), and M4 (spatial similarity = 0.86; temporal similarity = 0.71).
However, M3 (spatial similarity = 0.94; temporal similarity = 0.42) was reflected by two sep-
arated components: IC3 and IC4. One can realize also that the ICA algorithm failed to detect
M2 spanning between 57 ms and 60 ms. Using K-means clustering (Allen et al., 2014), six
brain states (BSs) were generated. BS1, BS2, BS3, BS5, and BS6 correspond, respectively, to
M1 (spatial similarity = 0.96; temporal similarity = 0.8), M2 (spatial similarity = 0.94; tem-
poral similarity = 0.77), M3 (spatial similarity = 0.97; temporal similarity = 0.53), M4 (spatial
similarity = 1; temporal similarity = 0.61), and M5 (spatial similarity = 0.99; temporal similar-
ity = 0.55). While the five simulated modular states were reconstructed, a false time win-
dow detection was captured (BS4). Regarding the consensus clustering algorithm proposed by
Rasero et al. (2017), the algorithm succeeded to detect the five simulated states. Tables 1 and 2
report the temporal and spatial similarities between the simulated MSs and those obtained us-
ing the different algorithms. Overall, results show that our algorithm reached the highest tem-
poral detection accuracy (0.83) compared with ICA (0.43), K-means clustering (0.58), and the
consensus clustering (0.79). Spatially, our algorithm procured a good spatial similarity (0.97),
outperforming ICA (0.72) and the consensus clustering algorithms (0.91).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
Task-related paradigm:
An experimental paradigm in which
the subject is asked to focus on a
specific function.
Real Data
As the considered dataset
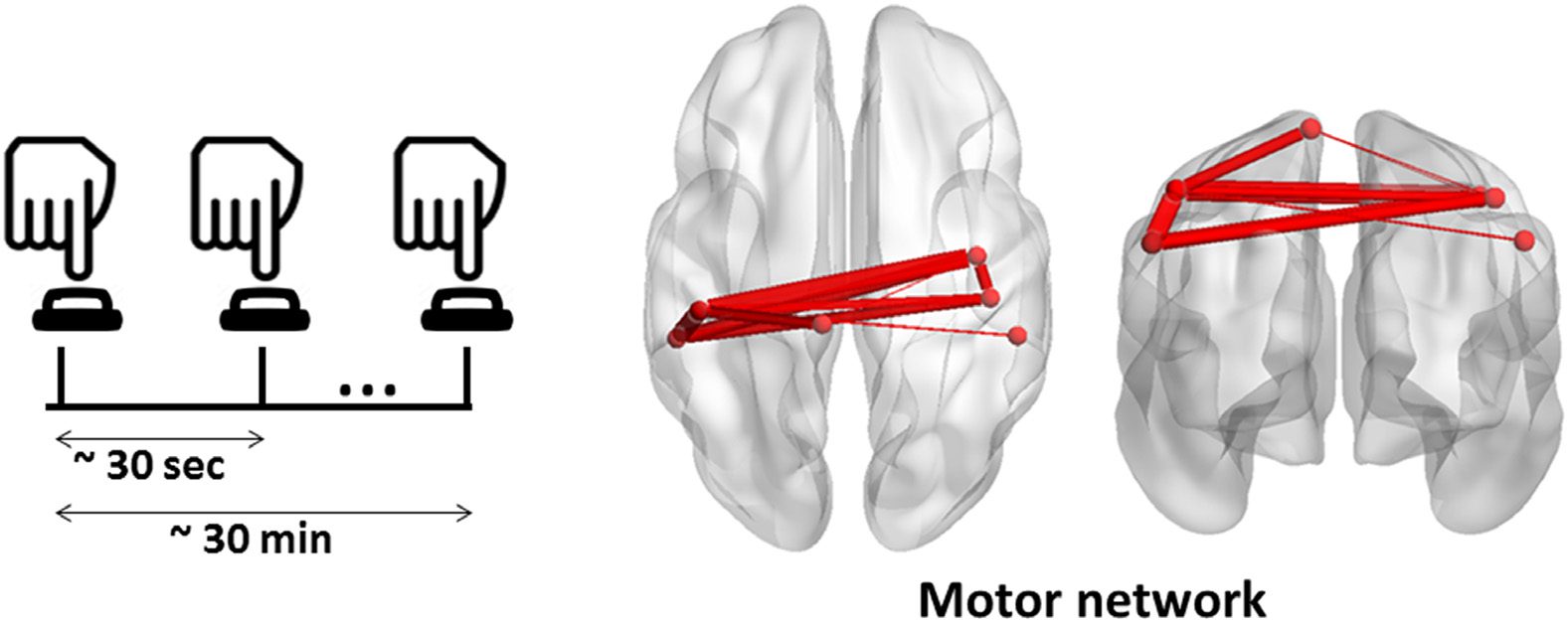
Dataset 1: Self-paced motor task for healthy participants (MEG data).
is collected during a task-related paradigm, our objective was to follow the spatiotemporal
organization of the dynamic brain networks during time (from the stimulus onset to the reaction
time). Thus, we applied the consecutive algorithm to track the MSs in a successive way. The
input of the algorithm is the tensor of dynamic connectivity matrices averaged over all trials
and subjects. The same dataset and methods were previously used in O’Neill et al. (2015),
O’Neill, Tewarie, Colclough, et al. (2017), and Vidaurre et al. (2016). The algorithm results in
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Spatial similarities between the simulated MSs and those obtained using the different algorithms
Algorithm
Our algorithm
ICA (O’Neill, Tewarie, Colclough,
et al., 2017)
K-means
(Allen et al., 2017)
Consensus clustering approach
(Rasero et al., 2017)
MS1
0.94
0.96
0.96
0.92
MS2
0.99
0.85
0.94
0.96
MS3
0.97
0.94
0.97
0.89
MS4
1
0.86
1
0.91
MS5
0.95
0
0.99
0.89
Average spatial similarity
0.97
0.72
0.99
0.91
Network Neuroscience
891
Detecting modular brain states in rest and task
Figure 8. The MS of the MEG motor task obtained using the consecutive method.
one significant MS found between −0.5 s and 1.5 s (Figure 8). As illustrated, this module impli-
cates the sensory motor area, the post- and precentral regions of both hemispheres.
The objective of using
Dataset 2: Picture naming task for healthy participants (dense-EEG data).
this dataset was to track the fast space/time dynamics of functional brain networks at subsecond
timescale from the onset (presentation of the visual stimuli) to the reaction time (articulation).
Hence, the consecutive version was applied on the dynamic connectivity matrices averaged
over subjects.
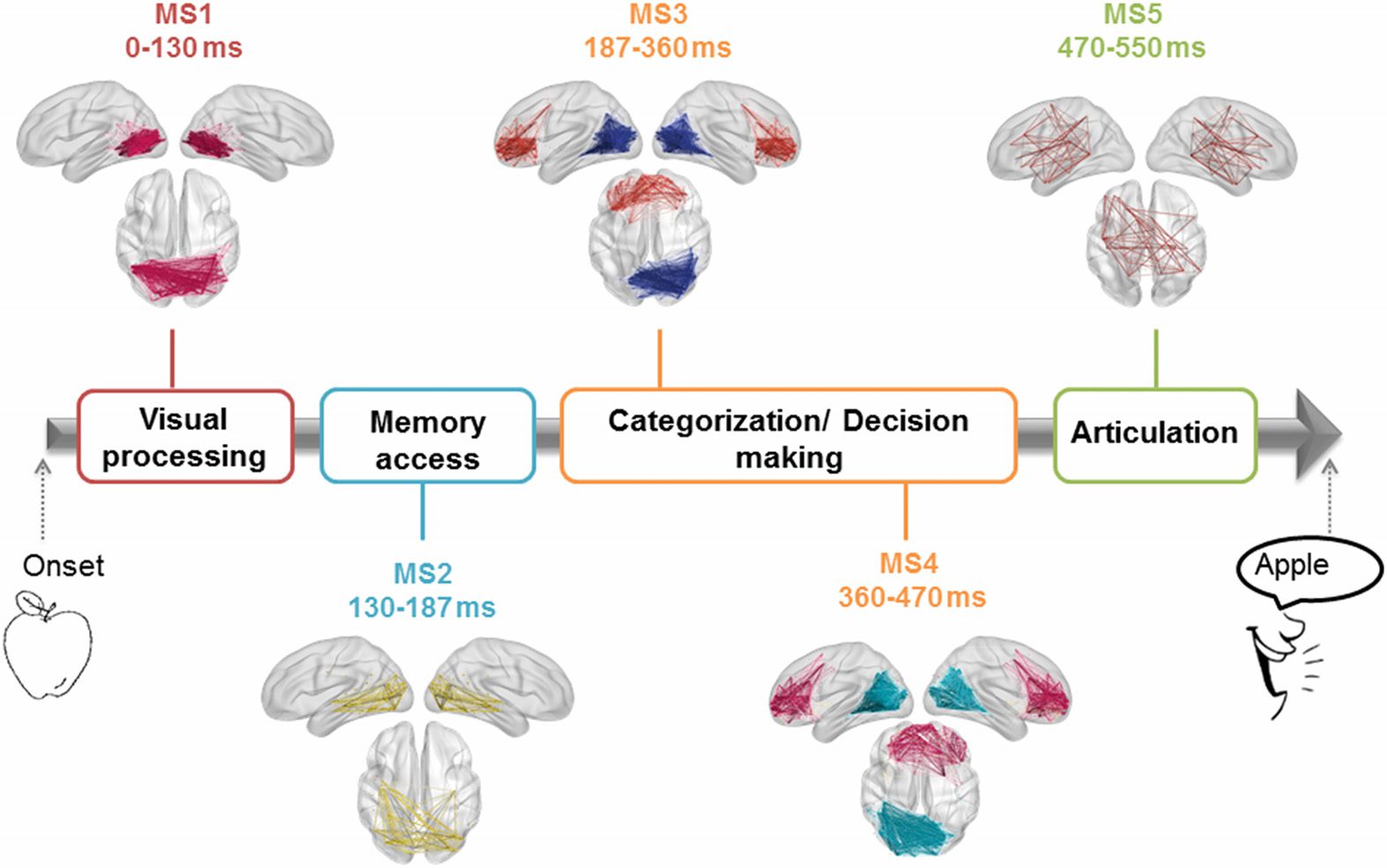
Figure 9 shows the results, revealing that the cognitive process can be divided into five
modular structures: The first MS corresponds to the time period ranging from the stimulus onset
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 9. The sequential MSs of the EEG picture naming task obtained using the consecutive
method and their corresponding cognitive functions.
Network Neuroscience
892
Detecting modular brain states in rest and task
to 130 ms and presents one module located mainly in the occipital region. The second MS
is observed between 131 and 187 ms, and involves one module showing occipitotemporal
connections. The third MS is identified between 188 and 360 ms, and illustrates a module
located in the occipitotemporal region, and another module located in the frontocentral region.
This structure was then followed by a fourth MS, found over the period 361–470 ms. MS4 was
very similar to the previous MS but with additional frontocentral connections. The last MS is
observed between 471 and 500 ms, and it shows a module connecting the frontal, central,
and temporal regions. It is worth noting that these MSs denote the transitions from the visual
processing and recognition to the semantic processing and categorization to the preparation
of the articulation process (Hassan, Benquet, et al., 2015; VanRullen & Thorpe, 2001).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
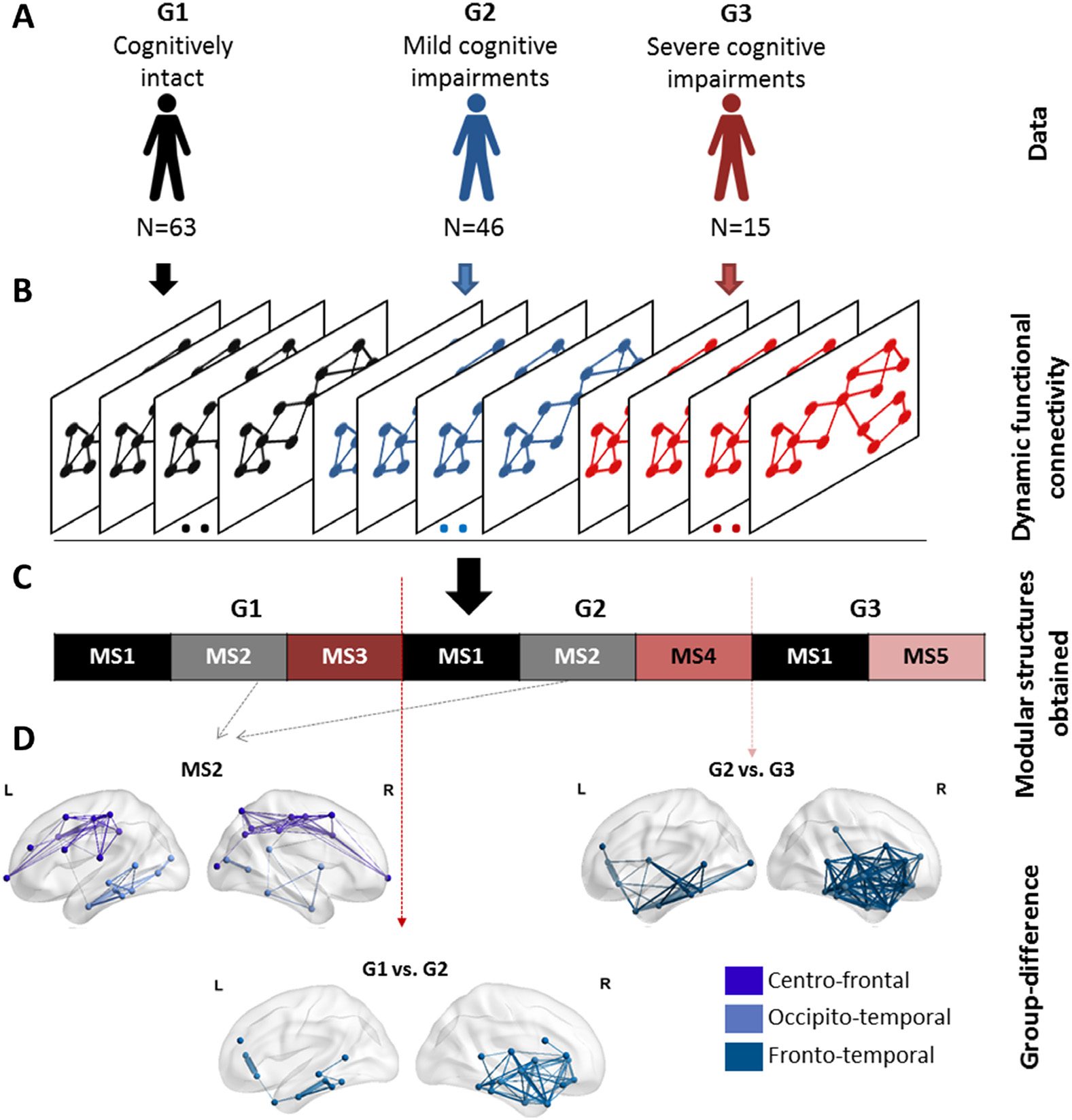
Figure 10. The analysis pipeline and the results of the categorical method applied on the Parkinson’s
disease EEG dataset. (A) The dataset composed of 124 patients partitioned into three groups: (G1)
cognitively intact patients (N = 63), (G2) patients with mild cognitive deficits (N = 46), and (G3) pa-
tients with severe cognitive impairment (N = 15). (B) The functional dynamic connectivity matrices
of the three groups concatenated over time. (C) The five modular structures obtained after applying
the categorical algorithm on the concatenated tensor. (D) The modular differences between G1 and
G2, G1 and G3, and G2 and G3.
Network Neuroscience
893
Detecting modular brain states in rest and task
Dataset 3: Resting state in Parkinson’s disease patients (dense-EEG data). Our objective here was
to identify the modular structures that are common between G1, G2, and G4, and those that
are specific to each group. As the sequential order of MSs is not intended, we applied the cat-
egorical version of the proposed algorithm. The latter takes as input the concatenation of the
dynamic connectivity matrices of the three groups. This will form a single data tensor of dimen-
sion N × N × T, where N is the number of ROIs and T is equal to the number of time windows ×
the number of patients (Figure 10). The algorithm was then applied to validate the usefulness
of the categorical version in detecting the modular alterations between G1, G2, and G3.
Results are illustrated in Figure 10. Five modular structures were identified (MS1, MS2,
MS3, MS4, and MS5). Three MSs were found for G1 and G2. However, the number of MSs
decreased from three to two MSs in G3. Results revealed that MS1 was found to be present in
the three groups, while MS2 was present only in G1 and G2. The modular structure MS2 (absent
in G3) is illustrated in Figure 10 and includes two modules involving mainly frontocentral
and occipitotemporal connections. The difference between G1 and G2 was reflected by the
absence of the structure MS3 replaced by the structure MS4 in G2. Results in Figure 10 show
that the difference concerns mainly the frontotemporal connections. The difference between
G2 and G3 was reflected by the absence of the structure MS4 from G2 and the presence of
the structure MS5 in G3. Figure 10 shows that the functional disruptions between G2 and G3
are mainly frontotemporal connections. It is worth noting that the frontotemporal disruptions
were widely reported in mild cognitive impairments (Beyer, Janvin, Larsen, & Aarsland, 2007;
de Haan et al., 2012; Song et al., 2011; Zhang et al., 2009), while the central disruptions
are widely observed in severe cognitive impairments (Hassan et al., 2017) and dementia (Song
et al., 2011).
DISCUSSION
In this paper, we have developed a novel framework to explore the fast reconfiguration of
the functional brain networks during rest and task. This new method can be used to track
the sequential evolution of brain modules during a task-directed paradigm or to identify the
modular brain states that arise at rest. The simulation-based analysis showed the ability of
the method to “reestimate” the modular network structures over time. Compared with other
methods, our proposed algorithm reached the highest temporal accuracy detection and a very
good spatial accuracy.
The new framework was validated on three different EEG/MEG datasets: (a) MEG data
recorded from 15 healthy subjects during a self-paced motor task, (b) dense-EEG data recorded
from 21 healthy subjects during a picture naming task, and (c) dense-EEG data recorded at rest
from 124 Parkinson’s disease patients with different cognitive phenotypes. Results show that
our method can track the fast modular states of the human brain network at a subsecond
timescale, and also highlight its potential clinical applications, such as the detection of the
cognitive decline in Parkinson’s disease.
“Categorical” and “Consecutive” Processing Schemes
The two processing schemes proposed here are both derived from the similarity matrix between
the temporal modules (Steps 1 to 3 in the Methods section). However, each version highlights
a specific characterization of the modular structures, which can then be exploited depend-
ing on the application (time/condition dependent). In particular, the results of the categorical
algorithm on the simulated data reveal high spatial resolution and relatively low temporal
resolution compared with those obtained using the consecutive algorithm. The low temporal
Network Neuroscience
894
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
resolution of the categorical version is reflected by the false (Figure 5) as well as the missed
time windows detection (Figures 1 and Figure 2 in the Supporting Information). In contrast,
these time windows were correctly detected by the consecutive version despite their short
length (Figure 6, and Supporting Information Figures 3 and 4). Yet, the low spatial resolution
of the consecutive version can be illustrated by MS5 (Figure 6, and Supporting Information
Figures 3 and 4) that should represent M2 (Figure 4). This is probably due to the categorical
version using the maximum number of available data points to generate their corresponding
MS, while the consecutive version treats each temporal segment solely.
We suggest using the consecutive version where sequential order of MSs is interesting to in-
vestigate, such as the tracking of cognitive tasks. When the temporal aspects are not necessary,
we would recommend the categorical version.
Tracking of Fast Cognitive Functions
The brain dynamically reconfigures its functional network structure on subsecond temporal scales
to guarantee efficient cognitive and behavioral functions (O’Neill, Tewarie, Vidaurre, et al.,
2017). Tracking the spatiotemporal dynamics of large-scale networks over this short time dura-
tion is a challenging issue (Allen et al., 2014; Hutchison et al., 2013). In this paper, we aimed
to examine how fast changes in the modular architecture shape information processing and
distribution in (a) motor tasks and (b) picture naming tasks.
Concerning the self-paced motor task, it is a simple task where only motor areas are ex-
pected to be involved over time. Our results indeed showed that the motor module is clearly
elucidated related to the tactile movement of the button press. The spatial and temporal fea-
tures of the obtained module are very close to the significant component obtained by O’Neill,
Tewarie, Colclough, et al. (2017) using the temporal ICA method.
The different MSs obtained in the EEG picture naming task are temporally and spatially anal-
ogous to the network brain states detected using other approaches such as K-means clustering
by Hassan, Benquet, et al. (2015). In particular, the first MS representing the visual network
is probably modulated by the visual processing and recognition processes (Thorpe, Fize, &
Marlot, 1996; VanRullen & Thorpe, 2001). The second MS reflects the memory access re-
flected by the presence of the occipital-temporal connections (Martin & Chao, 2001). In other
words, the brain tries to retrieve the information related to the picture illustrated from memory
(Martin & Chao, 2001). In the third and the fourth MSs, we notice the implication of a sep-
arated frontal module. This module may be related to the object category recognition (tools
vs. animals) and the decision-making process (Andersen & Cui, 2009; Clark & Manes, 2004;
Rushworth, Noonan, Boorman, Walton, & Behrens, 2011). After making the decision, speech
articulation and the naming process is prepared and started (Dronkers, 1996). This is reflected
by the MS5 that combines the frontal, the motor, and the temporal brain areas.
Modular Brain States and Cognitive Phenotypes in Parkinson’s Disease
Emerging evidence shows that Parkinson’s disease (PD) is associated with alteration in struc-
tural and functional brain networks (Fornito, Zalesky, & Breakspear, 2015). Hence, from a
clinical perspective, the demand is high for a network-based technique to identify the patho-
logical networks and to detect early cognitive decline in PD. Here, we used a dataset with a
large number (N = 124) of PD patients categorized into three groups in terms of their cogni-
tive performance: G1, cognitively intact patients; G2, patients with mild to moderate cognitive
Network Neuroscience
895
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
deficits; and G3, patients with severe cognitive deficits in all cognitive domains. See Dujardin
et al. (2015), Hassan et al. (2017), and Lopes et al. (2017) for more information about this
database.
The obtained MSs presented in Figure 10 show that while some MSs remain unchangeable
during cognitive decline from G1 to G3, others are altered and replaced by new MSs. More
specifically, the number of MSs detected in G3 has decreased compared with G1 and G2;
MS3 in G1 was replaced by MS4 in G2, while MS4 in G2 was replaced by MS5 in G3. In
addition, the alterations in G3 involve more distributed modules (central, frontotemporal) than
the alterations occurring between G1 and G2 (frontotemporal modules) where the impairment
still moderate.
Interestingly, the underlying modular differences between the MSs of groups are con-
sistent with the previously reported studies that explored the network changes in PD (Beyer
et al., 2007; Bosboom, Stoffers, Wolters, Stam, & Berendse, 2009; de Haan & Pijnenburg,
2009; Hassan et al., 2017; Song et al., 2011; Zhang et al., 2009). Particularly, the loss of fron-
totemporal connections in PD is supported by several EEG and MEG studies (Bosboom et al.,
2009; Hassan et al., 2017; Zhang et al., 2009). Similarly, results of structural MRI studies reveal
frontal and temporal atrophies in PD with mild cognitive impairment (Beyer et al., 2007; Song
et al., 2011). Other functional (de Haan et al., 2012) and structural (Zhang et al., 2009) studies
showed that Alzheimer’s disease networks are characterized by frontotemporal alterations. In
addition, the brain regions involved in the modular alterations in G3 found in our study are in
line with findings obtained by EEG edgewise analysis (Hassan et al., 2017), and by structural
MRI studies showing widespread atrophy associated with PD patients’ related dementia (Song
et al., 2011).
Methodological Considerations
First, we used a template anatomical image generated from MRIs of healthy controls for EEG/
MEG source functional connectivity analysis. The template-based method is common practice
in the absence of individual anatomical images and was previously employed by multiple
EEG and MEG source reconstruction studies, because of nonavailability of native MRIs (Hassan
et al., 2017; Kabbara et al., 2018; Lopez et al., 2014). Furthermore, a recent study showed that
there are few potential biases introduced during the use of a template MRI compared with
individual MRI coregistration.
Second, the connectivity matrices in Dataset 3 (Parkinson’s disease analysis) were thresh-
olded by keeping the highest 10% of the edge’s weights. According to previous studies, the
10% threshold provides an optimal trade-off between retaining the true connections and reduc-
ing spurious connections (Lord, Horn, Breakspear, & Walter, 2012). However, the consistency
of results across a range of proportional thresholds would be interesting to consider. In con-
trast, a statistical threshold (FDR) approach was used in Dataset 1 (self-paced motor tasks) and
Dataset 2 (picture naming); see Methods. The reason is that in Dataset 3, three groups were
compared. The proportional threshold approach, compared with other threshold approaches,
ensures equal density between the analyzed groups (van den Heuvel et al., 2017). More-
over, studies suggest that FDR controlling procedures are effective for the analysis of neuro-
imaging data in the absence of intergroup comparison (Bassett et al., 2013; Genovese et al.,
2002; Patel & Bullmore, 2016). One should also note that other strategies and criteria could
be used to objectively threshold the connectivity matrices, such as the minimum spanning tree
metric (Tewarie, van Dellen, Hillebrand, & Stam, 2015) and the efficiency cost optimization
(De Vico Fallani et al., 2017).
EEG/MEG source connectivity:
A method used to estimate functional
brain networks at the cortical level
using EEG/MEG sensors.
Network Neuroscience
896
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
Third, according to the consecutive algorithm, it is noteworthy that two parameters should
be adequately tuned to obtain relevant results: (a) the accuracy parameter a, and (b) the min-
imal length allowed for a state jmin. In particular, the accuracy parameter a will dramatically
affect the number and the temporal accuracy of detected states. Thus, it should be appropriately
selected. A high value may eliminate strong and significant similarity values in a low-average
matrix, whereas a low value may overemphasize weak similarity values in high-average con-
nectivity networks. Thus, we recommend choosing an adaptive value determined based on the
distribution of the similarity matrix. For this reason, the use of the average value is proposed.
In order to evidence our choice, we illustrate, based on simulated data with different noise
levels, the number of detected MSs as well the temporal accuracy as a function of a (Figures 5,
6, and 7 in the Supporting Information). In all cases, results show that the average value of the
similarity matrix provides the exact number of MSs and a good temporal accuracy. Concerning
the second parameter, jmin, it will depend on the way in which the connectivity matrices are
computed. In the case where the connectivity is calculated over a time window that guarantees
a sufficient number of data points, there is no need to set a minimal length for a state (jmin = 1).
In contrast, when a connectivity matrix is computed at each millisecond, one may restrict the
length of a state to be less than a specific number of milliseconds. As an example, Mheich,
Hassan, Khalil, Berrou, & Wendling (2015) suggested to reject a brain state that has a time
interval less than 30 ms. In our study, jmin was set to 1 time window in simulated data, Dataset
1 and Dataset 3, since an optimal sliding window was used. In Dataset 2, where a connectivity
matrix based on the PLV metric was computed each millisecond, jmin was set equal to 30 ms,
as proposed by Mheich et al. (2015).
Fourth, we are aware that spurious correlations caused by the problem of “source leakage”
should be carefully considered. Here, we have adopted in each dataset the same pipeline
(from data processing to network construction) used by the previous studies dealing with the
same dataset. Thus, for the MEG dataset, the correction for source leakage was performed by
the symmetric orthogonalization method proposed by Colclough et al. (2015). Using the same
pipeline also helps to avoid influencing factors caused by changing the source connectivity
method, the number of ROIs, the connectivity measure, or the sliding window length. By rely-
ing on previous studies (Hassan, Benquet, et al., 2015; Hassan et al., 2017; O’Neill, Tewarie,
Colclough, et al., 2017), we provide appropriate input—already tested and validated—to the
algorithm, regardless of how it was obtained. The source leakage issue was extensively dis-
cussed in a very recent review about M/EEG source-space networks (Hassan & Wendling,
2018).
In addition, a quantitative comparison presented in the paper was performed between our
algorithm, K-means clustering as proposed by Allen et al. (2014), independent component
analysis as proposed by O’Neill, Tewarie, Vidaurre, et al. (2017), and the consensus cluster-
ing algorithm as proposed by Rasero et al. (2017). Nevertheless, other strategies that showed
accurate results in previous studies could be also investigated and compared, such as the use
of a hidden Markov model (HMM), which models the cortical time series using a probabilis-
tic generative model (Vidaurre et al., 2017), or the use of a multivariate autoregressive model
(De Vico Fallani et al., 2008).
ACKNOWLEDGMENTS
This work has received a French government support granted to the CominLabs excellence
laboratory and managed by the National Research Agency in the “Investing for the Future”
program under reference ANR-10-LABX-07-01. It was also financed by the Rennes University
Network Neuroscience
897
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
Hospital (COREC Project named conneXion, 2012-14). This work was financed by the Azm
Center for Research in Biotechnology and Its Applications. GCO is funded by a Medical
Research Council New Investigator Research Grant (MR/M006301/1). This study was also sup-
ported by the Future Emerging Technologies (H2020-FETOPEN-2014-2015-RIA under agree-
ment No. 686764) as part of the European Union’s Horizon 2020 research and training program
2014–18. The study was also funded by the National Council for Scientific Research (CNRS)
in Lebanon.
AUTHOR CONTRIBUTIONS
Aya Kabbara: Conceptualization; Formal analysis; Methodology; Software; Validation; Visu-
alization; Writing - Original Draft. Mohamad Khalil: Funding acquisition. Georges O’Neill:
Data curation. Kathy Dujardin: Data curation. Youssof El Traboulsi: Methodology. Fabrice
Wendling: Funding acquisition. Mahmoud Hassan: Conceptualization; Formal analysis; Super-
vision; Writing - Original Draft.
FUNDING INFORMATION
Fabrice Wendling, Rennes University Hospital (http://dx.doi.org/10.13039/501100001665),
Award ID: ANR-10-LABX-07-01. Mohamad Khalil, National Council for Scientific Research.
Georges O’Neill, Medical Research Council, Award ID: MR/M006301/1. Fabrice Wendling,
Hôpital de rennes1, Award ID: COREC Project named conneXion, 2012–14.
REFERENCES
Allen, E. A., Damaraju, E., Eichele, T., Wu, L., & Calhoun, V. D.
(2017). EEG signatures of dynamic functional network connec-
tivity states. Brain Topography, 1–16. https://doi.org/10.1007/
s10548-017-0546-2
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T.,
& Calhoun, V. D. (2014). Tracking whole-brain connectivity dy-
namics in the resting state. Cerebral Cortex, 24, 663–676. https://
doi.org/10.1093/cercor/bhs352
Andersen, R. A., & Cui, H. (2009). Intention, action planning, and
decision making in parietal-frontal circuits. Neuron. https://doi.
org/10.1016/j.neuron.2009.08.028
Baker, A. P., Brookes, M. J., Rezek, I. A., Smith, S. M., Behrens, T.,
Smith, P. J. P., & Woolrich, M. (2014). Fast transient networks in
spontaneous human brain activity. eLife. https://doi.org/10.7554/
eLife.01867
Bassett, D. S., Porter, M. A., Wymbs, N. F., Grafton, S. T., Carlson,
J. M., & Mucha, P. J. (2013). Robust detection of dynamic com-
munity structure in networks. Chaos, 23. https://doi.org/10.1063/
1.4790830
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of hu-
man brain networks during learning. Proceedings of the National
Academy of Sciences, 108, 7641–7646. https://doi.org/10.1073/
pnas.1018985108
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T. (2015).
Learning-induced autonomy of sensorimotor systems. Nature
Neuroscience, 18, 744–751. https://doi.org/10.1038/nn.3993
Beyer, M. K., Janvin, C. C., Larsen, J. P., & Aarsland, D. (2007).
A magnetic resonance imaging study of patients with parkin-
son’s disease with mild cognitive impairment and dementia
using voxel-based morphometry. Journal of Neurology, Neuro-
surgery, and Psychiatry, 78, 254–259. https://doi.org/jnnp.2006.
093849[pii]\r10.1136/jnnp.2006.093849
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks. Jour-
nal of Statistical Mechanics: Theory and Experiment, 10008, 6.
https://doi.org/10.1088/1742-5468/2008/10/P10008
Bola, M., & Sabel, B. A. (2015). Dynamic reorganization of brain func-
tional networks during cognition. NeuroImage, 114, 398–413.
https://doi.org/10.1016/j.neuroimage.2015.03.057
Bosboom, J. L. W., Stoffers, D., Wolters, E. C., Stam, C. J., &
Berendse, H. W.
(2009). MEG resting state functional con-
Journal of
nectivity in Parkinson’s disease related dementia.
Neural Transmission,116,193–202. https://doi.org/10.1007/s00702-
008-0132-6
Chavez, M., De Vico Fallani, F., Valencia, M., Artieda, J., Mattia,
D., Latora, V., & Babiloni, F. (2013). Node accessibility in cor-
tical networks during motor tasks. Neuroinformatics. https://doi.
org/10.1007/s12021-013-9185-2
Clark, L., & Manes, F. (2004). Social and emotional decision-making
following frontal lobe injury. Neurocase, 10, 398–403. https://
doi.org/10.1080/13554790490882799
Cohen, D.
(1972). Magnetoencephalography: Detection of
the
brain’s electrical activity with a superconducting magnetome-
ter. Science, 175, 664–666. https://doi.org/10.1126/science.175.
4022.664
Colclough, G. L., Brookes, M. J., Smith, S. M., & Woolrich, M. W.
(2015). A symmetric multivariate leakage correction for MEG
connectomes. NeuroImage, 117, 439–448. https://doi.org/10.
1016/j.neuroimage.2015.03.071
Network Neuroscience
898
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
Damaraju, E., Allen, E. A., Belger, A., Ford, J. M., McEwen, S.,
Mathalon, D. H., . . . Calhoun, V. D. (2014). Dynamic functional
connectivity analysis reveals transient states of dysconnectivity
in schizophrenia. NeuroImage: Clinical, 5, 298–308. https://doi.
org/10.1016/j.nicl.2014.07.003
de Haan, W., & Pijnenburg, Y. (2009). Functional neural network
analysis in frontotemporal dementia and Alzheimer’s disease us-
ing EEG and graph theory. BMC Neuroscience, 10, 101. https://
doi.org/10.1186/1471-2202-10-101
de Haan, W., Van der Flier, W. M., Koene, T., Smits, L. L., Scheltens,
P., & Stam, C. J. (2012). Disrupted modular brain dynamics reflect
cognitive dysfunction in Alzheimer’s disease. NeuroImage, 59,
3085–3093. https://doi.org/10.1016/j.neuroimage.2011.11.055
de Pasquale, F., Della Penna, S., Snyder, A. Z., Marzetti, L., Pizzella,
V., Romani, G. L., & Corbetta, M. (2012). A cortical core for dy-
namic integration of functional networks in the resting human
brain. Neuron, 74, 753–764. https://doi.org/10.1016/j.neuron.
2012.03.031
de Pasquale, F., Della Penna, S., Sporns, O., Romani, G. L., &
Corbetta, M. (2015). A dynamic core network and global ef-
ficiency in the resting human brain. Cerebral Cortex, bhv185.
https://doi.org/10.1093/cercor/bhv185
De Vico Fallani, F., Latora, V., Astolfi, L., Cincotti, F., Mattia, D.,
Marciani, M. G., . . . Babiloni, F. (2008). Persistent patterns of in-
terconnection in time-varying cortical networks estimated from
high-resolution EEG recordings in humans during a simple motor
act. Journal of Physics A: Mathematical and Theoretical. https://
doi.org/10.1088/1751-8113/41/22/224014
De Vico Fallani, F., Latora, V., & Chavez, M. (2017). A topo-
logical criterion for filtering information in complex brain
networks. PLoS Computational Biology. https://doi.org/10.1371/
journal.pcbi.1005305
De Vico Fallani, F., Richiardi, J., Chavez, M., & Achard, S. (2014).
Graph analysis of functional brain networks: Practical issues
in translational neuroscience. Philosophical Transactions of the
Royal Society B: Biological Sciences. https://doi.org/10.1098/
rstb.2013.0521
Desikan, R. S., Sugonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C.,
Blacker, D., . . . Killiany, R. J. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI scans
into gyral based regions of interest. NeuroImage, 31, 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021
Dronkers, N. F.
(1996). A new brain region for coordinating
speech articulation. Nature, 384, 159–161. https://doi.org/10.
1038/384159a0
Duch, J., & Arenas, A. (2005). Community detection in complex
networks using extremal optimization. Physical Review E, 72,
027104. https://doi.org/10.1103/PhysRevE.72.027104
Dujardin, K., Moonen, A. J. H., Behal, H., Defebvre, L., Duhamel,
A., Duits, A. A., . . . Leentjens, A. F. G. (2015). Cognitive disor-
ders in Parkinson’s disease: Confirmation of a spectrum of sever-
ity. Parkinsonism and Related Disorders, 21, 1299–1305. https://
doi.org/10.1016/j.parkreldis.2015.08.032
Fornito, A., Zalesky, A., & Breakspear, M. (2015). The connectomics
of brain disorders. Nature Reviews Neuroscience. https://doi.org/
10.1038/nrn3901
Gallen, C. L., Baniqued, P. L., Chapman, S. B., Aslan, S.,
Keebler, M., Didehbani, N., & D’Esposito, M. (2016). Modu-
lar brain network organization predicts response to cognitive
training in older adults. PLoS ONE, 11. https://doi.org/10.1371/
journal.pone.0169015
Genovese, C. R., Lazar, N. A., & Nichols, T. (2002). Thresholding
of statistical maps in functional neuroimaging using the false dis-
covery rate. NeuroImage, 15, 870–878. https://doi.org/10.1006/
nimg.2001.1037\rS1053811901910377[pii]
Gibb, W. R. G., & Lees, A. J. (1988). The relevance of the Lewy
body to the pathogenesis of idiopathic Parkinson’s disease. Jour-
nal of Neurology, Neurosurgery, and Psychiatry. https://doi.org/
10.1136/jnnp.51.6.745
Girvan, M., & Newman, M. E. J. (2002). Community structure
in social and biological networks. Proceedings of the National
Academy of Sciences, 99, 7821–7826. https://doi.org/10.1073/
pnas.122653799
Gramfort, A., Papadopoulo, T., Olivi, E., & Clerc, M. (2010). Open-
MEEG: Opensource software for quasistatic bioelectromagnetics.
Biomedical Engineering Online, 9, 45. https://doi.org/10.1186/
1475-925X-9-45
Gratton, G., Coles, M. G. H., & Donchin, E. (1983). A new method
for off-line removal of ocular artifact. Electroencephalography
and Clinical Neurophysiology, 55, 468–484. https://doi.org/10.
1016/0013-4694(83)90135-9
Guimerà, R., & Amaral, L. A. N. (2005). Functional cartography of
complex metabolic networks. Nature, 433, 895–900. https://doi.
org/10.1038/nature03288
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Van Wedeen, J., & Sporns, O. (2008). Mapping the structural core
of human cerebral cortex. PLoS Biology, 6, 1479–1493. https://
doi.org/10.1371/journal.pbio.0060159
Hassan, M., Benquet, P., Biraben, A., Berrou, C., Dufor, O., &
Wendling, F. (2015). Dynamic reorganization of functional brain
networks during picture naming. Cortex, 73, 276–288. https://
doi.org/10.1016/j.cortex.2015.08.019
Hassan, M., Chaton, L., Benquet, P., Delval, A., Leroy, C.,
Plomhause, L., . . . Dujardin, K. (2017). Functional connectiv-
ity disruptions correlate with cognitive phenotypes in Parkinson’s
disease. NeuroImage: Clinical, 14, 591–601. https://doi.org/10.
1016/j.nicl.2017.03.002
Hassan, M., Shamas, M., Khalil, M., El Falou, W., & Wendling, F.
(2015). EEGNET: An open source tool for analyzing and visualiz-
ing M/EEG connectome. PLoS ONE, 10. https://doi.org/10.1371/
journal.pone.0138297
Hassan, M., & Wendling, F. (2018). Electroencephalography source
connectivity: Toward high time/space resolution brain networks.
IEEE Signal Processing Magazine, 1–25.
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M., . . . Chang, C. (2013). Dynamic
functional connectivity: Promise,
issues, and interpretations.
NeuroImage, 80, 360–378. https://doi.org/10.1016/j.neuroimage.
2013.05.079
Hyvärinen, A., Hirayama, J. I., Kiviniemi, V., & Kawanabe, M.
(2016). Orthogonal connectivity factorization: Interpretable de-
composition of variability in correlation matrices. Neural Com-
putation. https://doi.org/10.1162/NECO_a_00810
Network Neuroscience
899
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
n
e
n
_
a
_
0
0
0
9
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Detecting modular brain states in rest and task
Kabbara, A., Eid, H., El Falou, W., Khalil, M., Wendling, F., &
Hassan, M. (2018). Reduced integration and improved segrega-
tion of functional brain networks in Alzheimer’s disease. Journal
of Neural Engineering, 15. https://doi.org/10.1088/1741-2552/
aaaa76
Kabbara, A., El Falou, W., Khalil, M., Wendling, F., & Hassan,
M. (2017). The dynamic functional core network of the human
brain at rest. Scentific Reports, 7, 2936. https://doi.org/10.1038/
s41598-017-03420-6
Kabbara, A., Khalil, M., O’Neill, G., Dujardin, K., El Traboulsi, Y.,
Wendling, F., & Hassan, M. (2019). Modularity_algorithm_NN,
GitHub, https://github.com/librteam/Modularity_algorithm_NN
J.
(1999). Measuring phase synchrony in brain signals. Human
Brain Mapping, 8, 194–208. https://doi.org/10.1002/(SICI)1097-
0193(1999)8:4<194::AID-HBM4>3.0.CO;2-C
j. PAG., Rodriguez, MI., Martinerie,
J., & Varela, F.
Lachaux,
Dejar, METRO., Demuru, METRO., Hillebrand, A., & Fraschini, METRO. (2018). A
comparison between scalp- and source-reconstructed EEG net-
obras. Scientific Reports. https://doi.org/10.1038/s41598-018-
30869-w
Lopes, r., Delmaire, C., Defebvre, l., Moonen, A. J., Duits, A. A.,
Hofman, PAG., . . . Dujardin, k. (2017). Cognitive phenotypes in
Parkinson’s disease differ in terms of brain-network organiza-
tion and connectivity. Mapeo del cerebro humano, 38, 1604–1621.
https://doi.org/10.1002/hbm.23474
López, METRO. MI., Bruna, r., Aurtenetxe, S., Pineda-Pardo,
j. A.,
Marcos, A., Arrazola, J., . . . Maestu, F. (2014). Alpha-band hy-
persynchronization in progressive mild cognitive impairment: A
magnetoencephalography study. Revista de neurociencia, 34,
14551–14559.https://doi.org/10.1523/JNEUROSCI.0964-14.2014
Lord, A., Horn, D., romper la lanza, METRO., & walter, METRO. (2012). Changes
in community structure of resting state functional connectivity
in unipolar depression. PLoS ONE, 7(8). https://doi.org/10.1371/
diario.pone.0041282
Martín, A., & chao, l. l. (2001). Semantic memory and the brain:
Structure and processes. Opinión actual en neurobiología, 11,
194–201. https://doi.org/10.1016/S0959-4388(00)00196-3
Mheich, A., Hassan, METRO., Khalil, METRO., Berrou, C., & Wendling, F.
(2015). A new algorithm for spatiotemporal analysis of brain
conectividad funcional. Journal of Neuroscience Methods. https://
doi.org/10.1016/j.jneumeth.2015.01.002
Mucha, PAG. J., Richardson, T., Macon, K., Portero, METRO. A., & Onnela,
j. PAG. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Ciencia, 328, 876–878. https://doi.org/
10.1126/science.1184819
Núñez, PAG. l., & Srinivasan, R. (2007). Electroencephalogram. Schol-
arpedia, 2, 1348. https://doi.org/10.4249/scholarpedia.1348
O’Neill, GRAMO. C., Bauer, METRO., lana rica, METRO. w., morris, PAG. GRAMO., Barnes,
GRAMO. r., & Brookes, METRO. j. (2015). Dynamic recruitment of resting
state sub-networks. NeuroImagen, 115, 85–95. https://doi.org/10.
1016/j.neuroimage.2015.04.030
O’Neill, GRAMO. C., Tewarie, PAG. K., Colclough, GRAMO. l., Gascoyne, l. MI.,
Hunt, B. A. MI., morris, PAG. GRAMO.,
(2017).
Measurement of dynamic task related functional networks us-
ing MEG. Neuroimagen, 146, 667–678. https://doi.org/10.1016/
j.neuroimage.2016.08.061
. Brookes, METRO.
j.
.
.
O’Neill, GRAMO. C., Tewarie, PAG. K., Vidaurre, D., Liuzzi, l., lana rica,
METRO. w., & Brookes, METRO. j. (2017). Dynamics of large-scale electro-
physiological networks: A technical review. NeuroImagen. https://
doi.org/10.1016/j.neuroimage.2017.10.003
Oostenveld, r., & Praamstra, PAG. (2001). The five percent electrode
system for high-resolution EEG and ERP measurements. Clin-
ical Neurophysiology, 112, 713–719. https://doi.org/10.1016/
S1388-2457(00)00527-7
patel, A. X., & bullmore, mi. t. (2016). A wavelet-based estima-
tor of the degrees of freedom in denoised fMRI time series for
probabilistic testing of functional connectivity and brain graphs.
NeuroImagen, 142, 14–26. https://doi.org/10.1016/j.neuroimage.
2015.04.052
Penfield, w., & Jaspe, h. (1954). Epilepsy and the functional
anatomy of the human brain. JAMA, 155, 86. https://doi.org/10.
1001/jama.1954.03690190092039
Pfurtscheller, GRAMO., & Aranibar, A.
(1977). Event-related cortical
desynchronization detected by power measurements of scalp
EEG. Electroencephalography and Clinical Neurophysiology, 42,
817–826. https://doi.org/10.1016/0013-4694(77)90235-8
Pfurtscheller, GRAMO., & Lopes Da Silva, F. h. (1999). Event-related
EEG/MEG synchronization and desynchronization: Basic prin-
ciples. Clinical Neurophysiology. https://doi.org/10.1016/S1388-
2457(99)00141-8
Preti, METRO. GRAMO., & Van De Ville, D.
(2016). Eigenmaps of dy-
namic functional connectivity: Voxel-level dominant patterns
through eigenvector centrality. 2016 IEEE 13th International Sym-
posium on Biomedical
Imaging. https://doi.org/10.1109/ISBI.
2016.7493431
Rasero, J., Pellicoro, METRO., Angelini, l., Cortes, j. METRO., Marinazzo, D.,
& Stramaglia, S. (2017). Consensus clustering approach to group
brain connectivity matrices. Neurociencia en red, 1, 242–253.
https://doi.org/10.1162/NETN_a_00017
Rushworth, METRO. F. S., Noonan, METRO. A. PAG., Boorman, mi. D., Walton,
METRO. MI., & Behrens, t. mi. (2011). Frontal cortex and reward-guided
learning and decision-making. Neurona. https://doi.org/10.1016/
j.neuron.2011.05.014
Schneider, w., Eschman, A., & Zuccolotto, A. (2002). E-Prime ref-
erence guide. Psychology Software Tools, 3, 1.
Schoffelen, J., & Bruto, j. (2009). Source connectivity analysis with
MEG and EEG. Mapeo del cerebro humano. https://doi.org/10.1002/
hbm.20745
Song, S. K., Sotavento, j. MI., Parque, h. J., Sohn, Y. h., Sotavento, j. D., & Sotavento,
PAG. h. (2011). The pattern of cortical atrophy in patients with
Parkinson’s disease according to cognitive status. Movment Dis-
orders, 26, 289–296. https://doi.org/10.1002/mds.23477
despreciar, o., & Betzel, R. F. (2016). Modular brain networks. Annual
Review of Psychology, 67, 613–640. https://doi.org/10.1146/
annurev-psych-122414-033634
Tewarie, PAG., van Dellen, MI., Hillebrand, A., & estampar, C.
j.
(2015). The minimum spanning tree: An unbiased method for
brain network analysis. NeuroImagen. https://doi.org/10.1016/
j.neuroimage.2014.10.015
Thorpe, S., Fize, D., & Marlot, C. (1996). Speed of processing
in the human visual system. Naturaleza. https://doi.org/10.1038/
381520a0
Neurociencia en red
900
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Detecting modular brain states in rest and task
Traud, A. l., Kelsic, mi. D., Mucha, PAG. J., & Portero, METRO. A. (2008). Com-
paring community structure to characteristics in online collegiate
social networks. SIAM Review, 53, 526–543. https://doi.org/10.
1137/080734315
van den heuvel, METRO. PAG., el largo, S. C., Brilla, A., Seguin, C.,
yo, B. t. T., & Schmidt, R. (2017). Proportional thresholding in
resting-state fMRI functional connectivity networks and conse-
quences for patient-control connectome studies: Issues and rec-
ommendations. NeuroImagen, 152, 437–449. https://doi.org/10.
1016/j.neuroimage.2017.02.005
VanRullen, r., & Thorpe, S. j. (2001). The time course of visual
Procesando: From early perception to decision-making. Diario de
Neurociencia Cognitiva, 13, 454–461. https://doi.org/10.1162/
08989290152001880
Vidaurre, D., Abeysuriya, r., Becker, r., quinn, A. J., Alfaro-
Almagro, F., Herrero, S. METRO., & lana rica, METRO. W.. (2017). Discovering
dynamic brain networks from big data in rest and task. Neuro-
Image. https://doi.org/10.1016/j.neuroimage.2017.06.077
Vidaurre, D., quinn, A. J., Panadero, A. PAG., Dupret, D., Tejero-Cantero,
A., & lana rica, METRO. W.. (2016). Spectrally resolved fast transient
brain states in electrophysiological data. NeuroImagen, 126,81–95.
https://doi.org/10.1016/j.neuroimage.2015.11.047
zhang, y., Schuff, NORTE., Du, A.-T., rosa, h. J., Kramer, j. h., Gorno-
Tempini, METRO. l., . . . Weiner, METRO. W.. (2009). White matter damage
in frontotemporal dementia and Alzheimer’s disease measured
by diffusion MRI. Cerebro, 132, 2579–92. https://doi.org/10.1093/
brain/awp071
zhang, y., Wang, S. J., Xing, J., Liu, B., Mamá, z. l., Cual, METRO., . . .
Teng, GRAMO. j. (2009). Detection of PCC functional connectivity
characteristics in resting-state fMRI in mild Alzheimer’s disease.
Behavioural Brain Research, 197, 103–108. https://doi.org/10.
1016/j.bbr.2008.08.012
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
3
8
7
8
1
0
9
2
5
1
8
norte
mi
norte
_
a
_
0
0
0
9
0
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurociencia en red
901