INVESTIGACIÓN
Time-evolving dynamics in brain networks
forecast responses to health messaging
Nicole Cooper
1,2
, Javier O. Garcia
, Steven H. Tompson
1
, and Jean M. Vettel
Emily B. Falk
2,3
, Matthew B. O'Donnell
1
,
2,3
2,3,4
1Annenberg School for Communication, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
2A NOSOTROS. Army Research Laboratory, Aberdeen Proving Ground, Aberdeen, Maryland, EE.UU
3Department of Bioengineering, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
4Department of Psychological and Brain Sciences, Universidad de California, Santa Bárbara, California, EE.UU
un acceso abierto
diario
Palabras clave: Functional MRI (resonancia magnética funcional), Neuroimaging, Conectividad funcional, Behavior change,
Smoking
ABSTRACTO
Neuroimaging measures have been used to forecast complex behaviors, including how
individuals change decisions about their health in response to persuasive communications,
but have rarely incorporated metrics of brain network dynamics. How do functional
dynamics within and between brain networks relate to the processes of persuasion and
behavior change? Para abordar esta pregunta, we scanned 45 adult smokers by using functional
magnetic resonance imaging while they viewed anti-smoking images. Participants reported
their smoking behavior and intentions to quit smoking before the scan and 1 month later. Nosotros
focused on regions within four atlas-defined networks and examined whether they formed
consistent network communities during this task (measured as allegiance). Smokers who
showed reduced allegiance among regions within the default mode and fronto-parietal
networks also demonstrated larger increases in their intentions to quit smoking 1 month later.
We further examined dynamics of the ventromedial prefrontal cortex (vmPFC), as activation
in this region has been frequently related to behavior change. The degree to which vmPFC
changed its community assignment over time (measured as flexibility) was positively
associated with smoking reduction. These data highlight the value in considering brain
network dynamics for understanding message effectiveness and social processes more
broadly.
RESUMEN DEL AUTOR
How do functional dynamics within and between brain networks relate to the processes
of persuasion and behavior change? In this report, we assess brain network dynamics by
using fMRI while smokers view antismoking messages, and relate these metrics to smoking
behavior and intentions to quit smoking 1 month following the scan. Smokers who showed
reduced allegiance (less consistent network communities) among regions within the default
mode and fronto-parietal networks also demonstrated larger increases in their intentions to
quit smoking. Además, the degree to which the ventromedial prefrontal cortex flexibly
changed its community assignment over time was positively associated with later smoking
reducción. These data show that metrics of functional network dynamics can provide new
information about individual differences in responsiveness to anti-smoking messaging.
Citación: Cooper, NORTE., Garcia, j. o.,
Tompson, S., O'Donnell, METRO. B., Falk,
mi. B., & Vettel, j. METRO. (2019).
Time-evolving dynamics in brain
networks forecast responses to health
messaging. Neurociencia en red,
3 (1), 138–156. https://doi.org/
10.1162/netn_a_00058
DOI:
https://doi.org/10.1162/netn_a_00058
Supporting Information:
https://doi.org/10.1162/netn_a_00058
Recibió: 4 December 2017
Aceptado: 9 Puede 2018
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Nicole Cooper
ncooper@asc.upenn.edu
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2018
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
Health behaviors:
An individual’s actions and habits
that relate to the maintenance
and improvement of health and
well-being or prevention of
enfermedad.
Persuasive messaging:
In the context of public health
campaigns, stimuli designed to
motivate an audience to engage
in health-promoting behaviors or
refrain from unhealthy behaviors.
Large-scale brain networks:
Neural systems that are widespread
and distributed across the brain,
and are commonly recruited as a
coordinated unit during performance
of a task or at rest.
Dynamic functional connectivity:
Analytic measures that calculate
synchronized patterns of activity
between regional time series data.
Network communities:
Clusters of brain regions that are
identified as having similar functional
connectivity patterns, defined using
a data-driven technique.
Intentions:
An individual’s readiness to perform
a given behavior, assessed through
self-reported surveys.
INTRODUCCIÓN
Neural measures have forecasted future changes in behavior across a number of domains
(Berkman & Falk, 2013; Gabrieli, Ghosh, & Whitfield-Gabrieli, 2015). This has included
clinical treatment outcomes and health (Costafreda, Khanna, Mourao-Miranda, & Fu, 2009;
Doehrmann et al., 2013; Feldstein Ewing et al., 2017; Lopez et al., 2017; Wilcox et al., 2017;
Yang et al., 2016) as well as changes in individuals’ health behaviors in response to per-
suasive messaging. Neural activity during health messaging has been associated with re-
ductions in smoking (Chua et al., 2011; Cooper, Tompson, O'Donnell, & Falk, 2015; Falk,
Berkman, Whalen, & Lieberman, 2011; Falk et al., 2015; Riddle, Newman-Norlund, Baer, &
Thrasher, 2016; Zelle, Gates, Fiez, Sayette, & wilson, 2017), decreases in sedentary behavior
(Cooper, bassett, & Falk, 2017; Falk et al., 2015), and increased sunscreen use (Falk, Berkman,
Mann, harrison, & Lieberman, 2010; Vezich, Katzman, Ames, Falk, & Lieberman, 2017). Estos
studies have largely related future health behaviors to neural activity in a small number of brain
regiones. Sin embargo, these individual regions are also actively communicating with one another
by forming dynamic networks to integrate activity across disparate brain regions (Bressler &
menón, 2010; despreciar, Chialvo, Kaiser, & Hilgetag, 2004; despreciar, Tononi, & Edelman, 2000).
Como consecuencia, a host of recent research has developed new approaches to studying global
patterns in large-scale brain networks and has demonstrated that analyses of networks can
provide new insight into brain function and behavior (bullmore & despreciar, 2009; Friston, 2009;
Medalla, Lynall, & bassett, 2015; menón, 2011).
We examined dynamic functional connectivity among network communities while a group
of smokers were exposed to anti-smoking health messaging, and we hypothesized that individ-
ual differences in network interactions during messaging would precede subsequent changes
in intentions to quit smoking and actual smoking behavior. We focused on four a priori
networks which were defined based on resting-state data (Power et al., 2011). Large-scale
brain networks can be identified through the analysis of correlated neural activity during
rest or during relevant cognitive tasks (Bressler & menón, 2010; Friston, 1994; Raichle et al.,
2001). Regional interactions when the brain is at rest capture its intrinsic architecture (Fox &
Raichle, 2007; Greicius, Krasnow, Reiss, & menón, 2003), and as such, the resulting network
communities are thought to impose strong constraints on information processing in the brain
(Fox et al., 2005; Power et al., 2011; Shirer, Ryali, Rykhlevskaia, menón, & Greicius, 2012).
Network communities identified at rest are relevant for behavior and performance and can
be mapped on to broad categories of cognitive processes (Smith et al., 2009); Por ejemplo,
dynamic changes in interactions among these network communities can account for perfor-
mance variability (bassett, Wymbs, et al., 2013; bassett, Cual, Wymbs, & Grafton, 2015; Braun
et al., 2015; Deng, Chandrasekaran, Wang, & Wong, 2016; Gerraty et al., 2018; Liang, Zou,
Él, & Cual, 2016; Wang, Ong, Patanaik, zhou, & che, 2016). De este modo, we argue that networks
defined during the resting state identify fundamentally related systems of regions, cuales son
functionally relevant and predictive of task performance. We examine dynamics in these net-
works during task performance to demonstrate a link between individual differences in health
message processing and later smoking-related outcomes.
Behavior change:
In public health, any effort
(spontaneous or in response to an
intervention or persuasive messaging)
to change an individual’s actions to
promote health and prevent disease.
More specifically, we focused on four a priori networks of interest whose regions have
been associated with processes relevant for behavior change in previous research:
the de-
fault mode, fronto-parietal control, prominencia, and subcortical networks (Falk & Scholz, 2017;
Kaye, Blanco, & Luis, 2017). The default mode network is thought to form a system for self-
related cognitive processing, including social processing, memory, and prospection (Bressler
& menón, 2010; Buckner et al., 2009; Laird et al., 2011). The salience network is critical for
Neurociencia en red
139
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
selecting and responding to behaviorally relevant stimuli (Barrett & Satpute, 2013; menón,
2011; Seeley et al., 2007). A growing body of previous work relating health-related outcomes
to brain activity has implicated individual brain regions that are part of the default mode and
salience networks (Chua et al., 2011; Cooper et al., 2017; Dinh-Williams, Mendrek, Dumais,
Bourque, & Potvin, 2014; Falk et al., 2015; Ramsay, Yzer, Luciana, Vohs, & macdonald,
2013; Riddle et al., 2016; Vezich et al., 2017; Wang y cols., 2013; Weber, Huskey, Mangus,
Westcott-Baker, & Tornero, 2015; Zelle et al., 2017); future behavior has also been related
to task activation in the striatum (Berns & moore, 2012; Genevsky & Knutson, 2015; Kühn,
Strelow, & Gallinat, 2016; Venkatraman et al., 2015). Finalmente, changes in the fronto-parietal
control network, thought to support task-switching, have been linked to learning and decision-
haciendo (Bassett y cols., 2011; Braun et al., 2015; Gerraty et al., 2018), processes which are
likely to be relevant to belief updating when receiving new information. Based on the criti-
cal role of these cognitive systems in support of behavior change, we hypothesized that bet-
ter understanding the interactions among the regions in these a priori, atlas-defined networks
would uncover an important and yet unstudied component of brain dynamics that can forecast
critical health outcomes, changes in intentions to perform a behavior and actual changes in
ese comportamiento. We note that individual differences in network dynamics during the task could
be due to a trait-like intrinsic difference in network dynamics, or context-dependent differences
in how individual smokers process the experience of a smoker viewing anti-smoking messages
(which may stem in part from properties of the messages themselves); effects observed here
could be due to one or a combination of these possibilities.
Previous research has found that although mean activation in ventromedial prefrontal cor-
tex (vmPFC) is associated with subsequent behavior change, this same brain activity is often
uncorrelated with participants’ self-reported intentions (Cooper et al., 2015; Falk et al., 2010,
2011). Several theories of health behavior posit that intentions to perform a behavior are an
important precursor to behavior change, but that other factors also influence whether behavior
change occurs (Ajzen, 1985, 1991; Armitage & Conner, 2001; Fishbein, 1979; Fishbein &
Ajzen, 2011; Webb & Sheeran, 2006). En breve, although related, intentions to change and actu-
ally changing behavior may be associated with partially differing neural precursors. To further
explore this possibility, we compare both outcomes (changes in intentions and behavior) a
network dynamics.
Although previous research has identified the regions in the default mode, fronto-parietal
control, prominencia, and subcortical networks as key components of successful behavior change,
little work has examined how they work in concert. To assess the variable interactions between
brain regions in these networks, we utilized two complementary metrics recently developed
in network science to quantify regional dynamics, allegiance and flexibility. We first tested
whether sustained coordinated processing within regions in the default mode, fronto-parietal,
prominencia, and subcortical networks results in lasting changes in message-consistent outcomes.
The extent to which regions form a cohesive community and demonstrate the same pattern
of activity across time can be quantified by allegiance, where higher allegiance in a network
would indicate more sustained coordination of activity and processing within nodes in that
network and decreased allegiance would indicate greater diversity in processing across nodes.
We thus compare individuals’ changes in smoking-relevant outcomes to the allegiance in four
key brain networks during messaging.
We further examined the vmPFC specifically, which is the region most consistently asso-
ciated with future behavior change in previous work (Chua et al., 2011; Cooper et al., 2015;
Falk et al., 2010, 2015; Falk & Scholz, 2017; Riddle et al., 2016; Vezich et al., 2017). Given the
140
Allegiance:
A metric that quantifies the cohesion
among regions in a network
community across time.
Flexibility:
A metric that quantifies how
frequently a single region changes
its assignment across communities
con el tiempo.
Nodos:
The smallest unit of a network
análisis, representing a cohesive
brain region. Nodes can be defined
based on brain anatomy or patterns
of functional activation.
Neurociencia en red
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
vmPFC’s role in integrating multiple sources of information to compute a value signal (Bartra,
McGuire, & Kable, 2013), we propose that successful change in behavior requires dynamic
connections between the vmPFC and other relevant cognitive systems, which will be indexed
by increased vmPFC flexibility. This measure focuses on the activity of single brain regions,
revealing whether a region remains a member of the same community over time or if it fre-
frecuentemente (and flexibly) changes its assignment across communities between time points. De este modo,
we test the importance of both consistent interactions among regions and dynamic changes
between networks during messaging about smoking cessation.
MÉTODOS
Participantes
The study sample consisted of 45 Participantes (28 hombres; edad media = 32 años, DE = 13;
30 Blanco). All participants gave written, informed consent in accordance with the procedures
of the Institutional Review Board at the University of Michigan. Of the original 50 Participantes,
2 participants were excluded for missing data (1 due to an error at the scanner, and another for
not participating in the final session). Three participants were excluded for data quality issues
(1 for neurological abnormalities, 1 for excessive head motion, and a third for both vision
problems and excessive head motion).
Participants were recruited from the general population by using Craigslist and a university
website. Initial eligibility was assessed through a phone call. To be eligible, potential partic-
ipants must have been between the ages of 18 y 65, have smoked at least 5 cigarettes per
day for the past month, and have been a smoker for at least 12 meses. Además, participar-
pants had to meet standard fMRI eligibility criteria, including having no metal in their body,
no history of psychiatric or neurological disorders, and currently not taking any psychiatric or
illicit drugs.
Study Timeline and Measures
Following a screening for eligibility via telephone, participants completed three study ses-
siones. The first session (Session 1) provided baseline measures of self-reported smoking be-
havior and intentions to quit or reduce smoking, which were reported again at each following
session. The fMRI scan (Session 2) took place an average of 6 days later. The follow-up session
(Session 3) was conducted via telephone, an average of 39 days after Session 2.
Smoking Outcomes
We assessed two smoking outcomes and their relation to neural dynamics. We first examined
changes in intentions to quit smoking. At each of the three sessions, participants were asked
three questions about their intentions to quit, reduce, or refrain from smoking in the next
three months. The intention ratings were made on a four-point scale (anchors: 1 = definitely
no lo hará, 2 = probably will not, 3 = probably will, y 4 = definitely will). Responses to
these questions were averaged for each time point. Intention change for each individual was
measured as the difference between the average of all intention questions at Session 1 y el
average at Session 3. Intentions were also measured immediately after the scan (Session 2),
but intention change from Session 1 to Session 2 was not associated with network measures
or behavior change.
We also examined changes in self-reported smoking behavior. Participants were asked to
report the number of cigarettes they smoked per day at each of the three study sessions. Como un
reference, they were told that a pack contains 20 cigarettes. We related neural dynamics to
Neurociencia en red
141
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
the percent change in cigarettes smoked per day from Session 1 to Session 3 in each individual.
We started with self-reports at Session 1 to match the time point of the intention measure; el
reports of daily smoking at Session 1 and Session 2 were very consistent (r= 0.94). Self-report
measures are commonly used to track smoking behavior change (Chua et al., 2011; Jasinska
et al., 2012) and have been shown to have a moderate to high correlation with physiological
metrics such as expired CO (Falk et al., 2011; jarvis, Tunstall-Pedoe, Feyerabend, Vesey, &
Saloojee, 1987; Middleton & Morice, 2000) and saliva and serum cotinine (Etter, Vu Duc, &
Perneger, 2000; Patrick et al., 1994; Pokorski, Chen, & Bertholf, 1994; Vartiainen, Seppälä,
Lillsunde, & Puska, 2002).
fMRI Task
Participants completed four tasks in the scanner, but this analysis focused on the main task
de interés, a persuasive messaging task that promoted smoking cessation. Participants saw
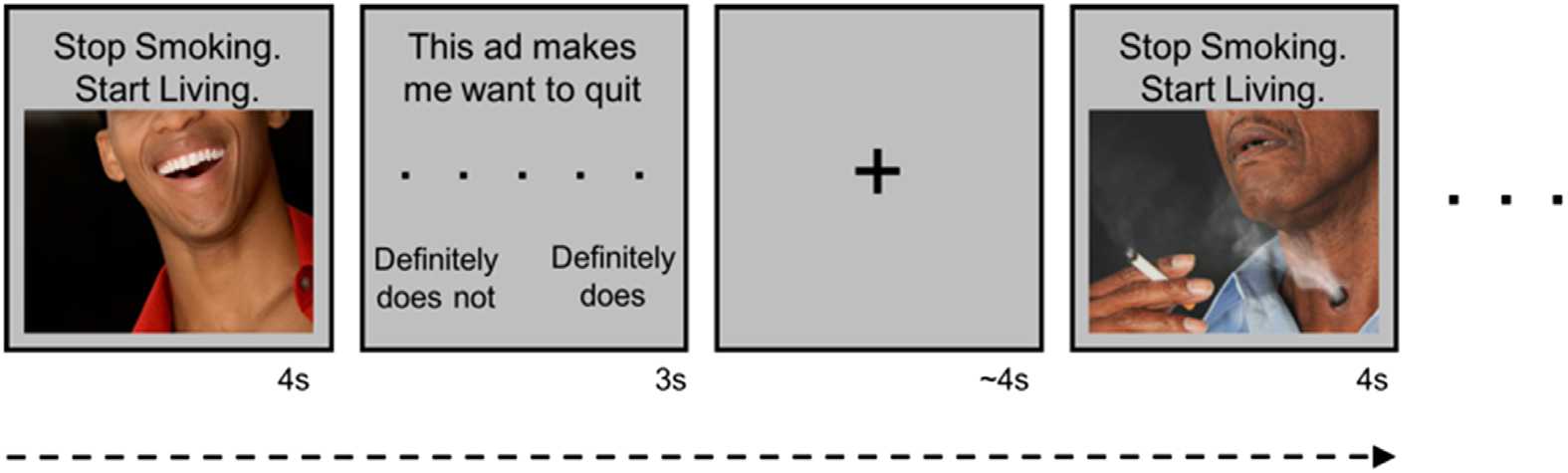
80 images with the tagline “Stop Smoking. Start Living.” Each trial consisted of 4 s of image
presentación, followed by a 3-s response screen with the statement “This makes me want to
quit” and a five-point rating scale (1 = definitely does not, 5 = definitely does); ver figura 1.
The response period was followed by a jittered intertrial interval, consisting of a screen with
only a fixation cross (3–7.5 s, mean = 4.10 s, median = 3.32 s, DE = 1.01 s).
Participants viewed 30 negative anti-smoking images, based on the FDA’s proposed graphic
warning labels. De estos, 12 portrayed social consequences of smoking (p.ej., exclusion from
a group) y 18 portrayed nonsocial and health-related consequences of smoking (p.ej., a
tracheotomy). Además, participants viewed 30 neutral control images (11 social, 19 non-
social). The negative and neutral images were qualitatively matched in pairs, by overall com-
position of the content (p.ej., X-ray image of a diseased lung and X-ray image of a healthy
lung), focal point, and number of people in the image. El restante 20 face images were a
between-subject manipulation of personalization, where one set of participants saw images of
their Facebook friends (norte = 19 Participantes) and the other (norte = 26 Participantes) saw unknown
faces from a public database known as NimStim (Tottenham et al., 2009). We controlled for
this between-subject manipulation in the regression analyses below, confirming that it was
not significantly related to outcomes of interest. Each image was presented once, y el
order of image presentation was randomized across individuals.
MRI Data Acquisition
Neuroimaging data were acquired using a 3 Tesla GE Signa MRI scanner. Two functional runs
of one task (454 volumes total) are analyzed here. Functional images were recorded using a
Cifra 1. Task design. While undergoing fMRI, participants viewed images paired with the tagline
“Stop Smoking. Start Living.”
Neurociencia en red
142
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
reverse spiral sequence (TR = 2,000 EM, TE = 30 EM, flip angle = 90◦
, 43 axial slices, FOV =
220 mm, slice thickness = 3 mm; voxel size = 3.44 × 3.44 × 3.0 mm). We also acquired
in-plane T1-weighted images (43 slices; slice thickness = 3 mm; voxel size = .86 × .86 ×
3.0 mm) and high-resolution T1-weighted images (SPGR; 124 slices; slice thickness = 1.02 ×
1.02 × 1.2 mm) for use in coregistration and normalization.
fMRI Preprocessing
Functional data were preprocessed and analyzed using Statistical Parametric Mapping (SPM8,
Wellcome Department of Cognitive Neurology, Institute of Neurology, Londres, Reino Unido). To al-
low for the stabilization of the BOLD signal, the first five volumes (10 s) of each run were
not recorded by the scanner. Functional images were despiked using the 3dDespike program
(AFNI; Cox, 1996). Próximo, data were corrected for differences in the time of slice acquisition
by using sinc interpolation, where the first slice served as the reference slice. Data were then
spatially realigned to the first functional image. We then coregistered the functional and struc-
tural images by using a two-stage procedure. Primero, in-plane T1 images were registered to the
mean functional image. Próximo, high-resolution T1 images were registered to the in-plane T1 im-
edad. After coregistration, high-resolution structural images were segmented to produce a gray
matter mask, and then normalized to the skull-stripped MNI template provided by FSL. Finalmente,
functional images were smoothed using a Gaussian kernel (8-mm full width at half maximum).
Based on preliminary recent evidence suggesting the possible sensitivity of network results
to spatial smoothing (Alakörkkö, Saarimäki, Glerean, Saramäki, & Korhonen, 2017; Chen &
Calhoun, 2018), we conducted comparative analyses with unsmoothed data and confirmed
that both the regional time course dynamics and a region’s temporally evolving community
affiliation were highly similar across smoothed and unsmoothed data in this study. We repeated
the main analyses below with unsmoothed data, and present these results in the Supporting
Información (Cooper, Garcia, Tompson, O'Donnell, Falk, & Vettel, 2019).
Functional Connectivity Analysis
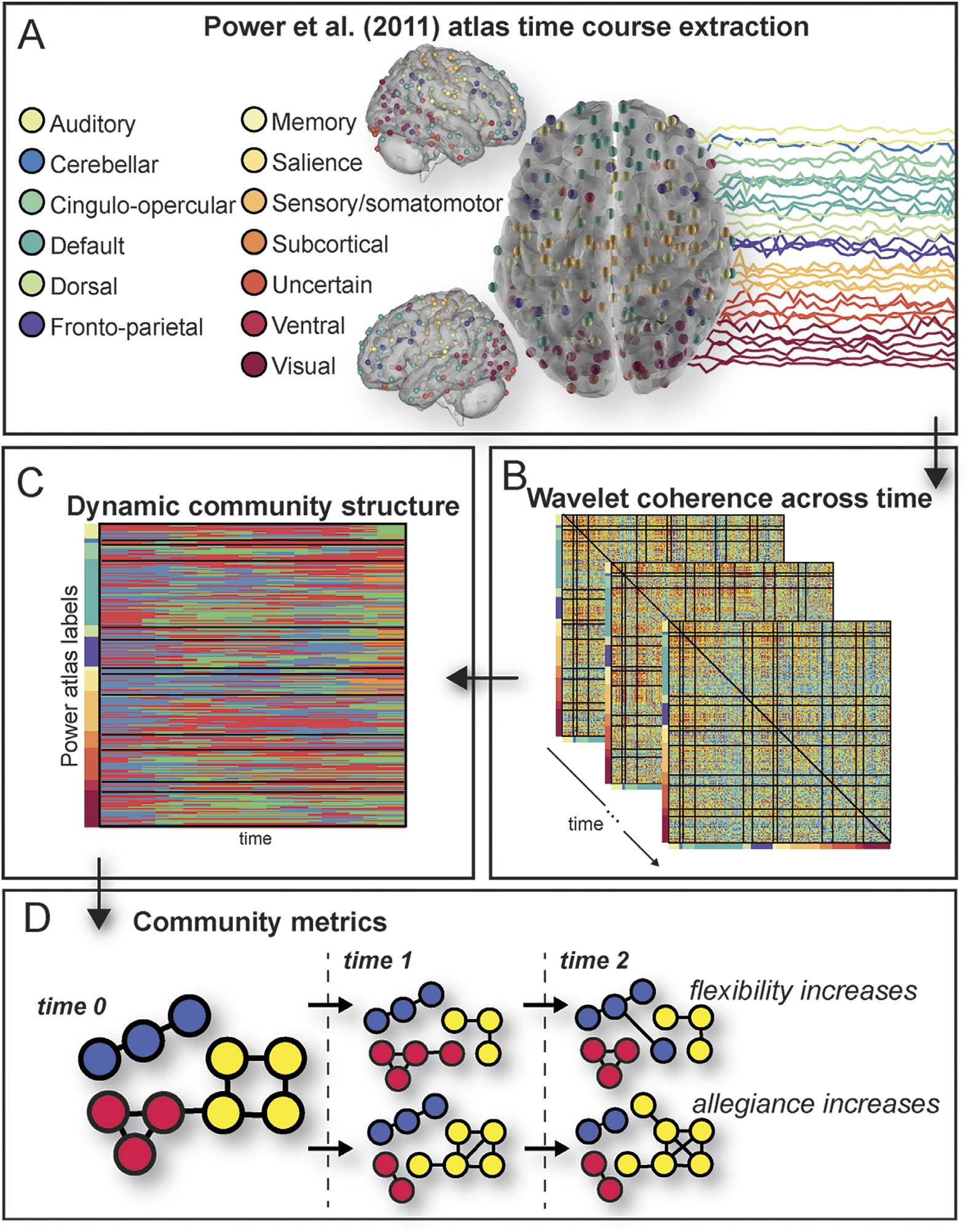
Following preprocessing, the mean signal was extracted from 264 atlas-defined regions of
interés (ROI) by using the MarsBar package for SPM. These ROIs were spherical regions with
an 8-mm radius, centered on the 264 coordinates defined by Power et al. (2011). The de-
trended time courses from these regions were divided into 22 nonoverlapping bins of 20 TRs
(dónde 20 TRs = 40 s); this bin size was chosen to optimize the detection of individual dif-
ferences in dynamics during the task (Telesford et al., 2016). Given the short event–related
design of this task, and relatively small number of images in each task condition, we did
not compare dynamics of connectivity across the task separately by task condition. Wavelet
coherence was estimated in each bin for each pair of regions, and was averaged across fre-
quency bands between 0.06 y 0.12 Hz, a task-relevant frequency range of coherence (Sol,
Molinero, & D'Esposito, 2004). This resulted, for each bin, en un 264 × 264 matrix of coherence
values for each pair of regions (Figura 2B). Estos 264 regions are identified by Power et al.
(2011) as composing 13 redes, depicted in Figure 2A. Based on previous research, nuestro
analysis focused on four networks from the Power atlas: the default mode, fronto-parietal,
prominencia, and subcortical networks. These networks have been associated with processing in-
dicative of persuasion and successful behavior change (Bassett y cols., 2011; Braun et al., 2015;
Chua et al., 2011; Cooper et al., 2017; Dinh-Williams et al., 2014; Falk et al., 2015; Gerraty
et al., 2018; Ramsay et al., 2013; Riddle et al., 2016; Vezich et al., 2017; Wang y cols., 2013;
Weber et al., 2015; Zelle et al., 2017).
Neurociencia en red
143
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. Analysis design: overview of analysis scheme. We extracted the time series of activation
in all nodes of the Power atlas brain parcellation during the task (A). Using wavelet coherence as
a measure of functional connectivity (B) and input to a dynamic community detection algorithm
(C), we explored community affiliations across the time course of the task using two metrics, flex-
ibility and allegiance, which are explained in a hypothetical network (D). From an initial network
configuration at time 0, regions reconfigure over time. In the top row, a node changes its affiliation
from the yellow community at time 0 to the red community at time 1, then to the blue community
at time 2, indicating increased flexibility relative to nodes remaining in the same community at all
time points. In the bottom row, the yellow community gains more nodes and more connections
between nodes across time, indicating increased allegiance.
Neurociencia en red
144
Time-evolving brain dynamics forecast responses to health messaging
Multilayer community detection
análisis:
The process of assigning brain
regiones (or nodes) to network
communities, when the network has
“multilayer” observations, como
multiple measurements over time.
Community Detection and Network Metrics
We employed recent advancements from network science to examine whether the synchrony
within a network community (allegiance among the brain regions in the same community) o
interactions between network communities (flexibility of brain regions to coordinate across
communities) accounted for lasting changes in smoking outcomes. To capture changes in net-
work communities over the course of the task, we utilized a multilayer community detection
análisis (Bassett y cols., 2011; Mucha, Richardson, Macon, Portero, & Onnela, 2010). This allows
for the investigation of changes in network structure over time by coupling nodes between
adjacent time slices, and results in a community partition for each time window (Figura 2C).
The algorithm utilized a generalized Louvain algorithm to optimize modularity (bassett, Portero,
et al., 2013; Telesford et al., 2016). We repeated this optimization 100 veces, since the
algorithm is nondeterministic and susceptible to near degeneracies (Bien, de Montjoye, &
cláusula, 2010), and we averaged the iterations to compute the community metrics.
The resulting community structures were used to estimate flexibility and allegiance
(Ashourvan, Gu, Mattar, Vettel, & bassett, 2017). Allegiance is defined as the proportion of time
windows during which each pair of nodes were assigned to the same community. Flexibility
is defined as the proportion of time windows during which each node changes community
asignación. As shown in Figure 2D, the central region shows high flexibility as it changes
assignment from the yellow community to the red community at time 1 and the blue com-
munity at time 2.
A diferencia de, allegiance identifies regions that are strongly connected over
tiempo, as demonstrated by the yellow community in Figure 2D. We employed these two met-
rics to examine the relationship between brain activity and health outcomes.
Relating Network Allegiance Metrics and Smoking-Related Outcomes
In our first set of analyses, we examined the relationship between network allegiance measures
and changes in smoking-related outcomes. We tested these relationships in the four atlas-
defined networks of interest (default mode, fronto-parietal, prominencia, and subcortical networks).
For analyses in the a priori networks, allegiance of all node pairs was averaged to obtain
a composite measure of allegiance within the atlas-defined network. In separate models for
each a priori network, we used robust regression to predict changes in smoking intentions and
percent changes in daily smoking. We examined average allegiance both as a linear metric and
binned into quartiles to identify robust trends in the community dynamics (Lange, Oostenveld,
& Fries, 2013; van Dijk, Schoffelen, Oostenveld, & Jensen, 2008), where quartile labels were
entered as a categorical variable in the robust regression model.
We used the robust regression (RLM) function in R’s (versión 3.2.4) MASS library. The Wald
test was used to assess significance of RLM coefficients (robtest, R’s sfsmisc package). All mod-
els controlled for personalization condition (Facebook vs. NimStim faces), género, edad, y
etnicidad (White vs. otro); models predicting intention change also controlled for Session 1
(base) intentions. Robust linear models are less sensitive to outliers and high leverage
data points, allowing the inclusion of all data points. Personalization condition, a between-
participants variable, did not significantly relate to the main outcomes discussed in this inves-
tigation (metrics of network allegiance and flexibility or smoking-related outcomes).
Relating Network Flexibility Metrics and Smoking-Related Outcomes
In our final analysis, we examined the relationship between vmPFC flexibility and changes in
smoking-related outcomes. We examined vmPFC flexibility both binned into quartiles to test
Neurociencia en red
145
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
norte
mi
norte
_
a
_
0
0
0
5
8
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
Brain network dynamics:
Time-evolving interactions between
brain regions thought to underlie
cognición y acción.
categorical differences (categorical predictor in regression) and as a linear metric. We used
robust regression to relate vmPFC flexibility to changes in smoking intentions and percent
changes in daily smoking. As above, these models controlled for task condition (Facebook vs.
NimStim faces), género, edad, and ethnicity (White vs. otro); models predicting intention
change also controlled for Session 1 (base) intentions.
RESULTADOS
en este estudio, we examine how the dynamics of brain networks during exposure to anti-smoking
messaging relate to smoking-related outcomes in the following month. Nosotros planteamos la hipótesis de que
individual differences in metrics of brain network dynamics during a behaviorally relevant
tarea, rating anti-smoking messages, would precede changes in smoking-related intentions and
comportamiento. Smokers participated in an fMRI scanning session, during which they viewed anti-
smoking messages. Before the fMRI scan and 1 month later, participants self-reported their
intentions to quit smoking and the number of cigarettes they smoked per day. We first exam-
ined brain network dynamics during exposure to anti-smoking messaging in nodes belonging
to four a priori networks based on the Power et al. (2011) atlas: the default mode network
(DMN), fronto-parietal network (FPN), salience network, and subcortical network. Activation
in regions that comprise these networks has been previously linked to persuasion and health
behavior change, but their community dynamics have not been investigated. We assessed the
functional connectivity between all pairs of regions in 22 consecutive time windows across
the course of the task. We then used a dynamic community detection algorithm to study
the relationship between brain network dynamics and smoking-related outcomes in two com-
plementary analyses: the first investigated allegiance in our a priori networks of interest, y
the second studied flexibility in the vmPFC based on its consistent association with future
behavior change in previous work.
Changes in Smoking Intentions and Behavior
Average intentions to reduce or quit smoking significantly increased from the intake session to
the follow-up session (paired t(44) = 4.59, pag < 3.6 × 10−5
). At Session 1, intentions to quit
averaged 2.41 (SD = 0.81); at Session 3, intentions to quit averaged 2.97 (SD = 0.76). We also
examined a second smoking outcome, changes in smoking behavior. At Session 1, participants
reported smoking an average of 13.3 (SD = 6.5) cigarettes per day. At Session 3, which took
place an average of 45 days later, participants smoked an average of 10.2 (SD = 7.7) cigarettes
per day. This represented a significant decline in the number of cigarettes participants smoked
per day from the intake to follow-up session (paired t(44) = 3.22, p < 0.0024). In the following
sections, we examine the relationships between changes in smoking intentions and behavior,
and dynamics in neural network measures during exposure to anti-smoking messaging.
Allegiance in Subnetworks Relates to Changes in Intentions
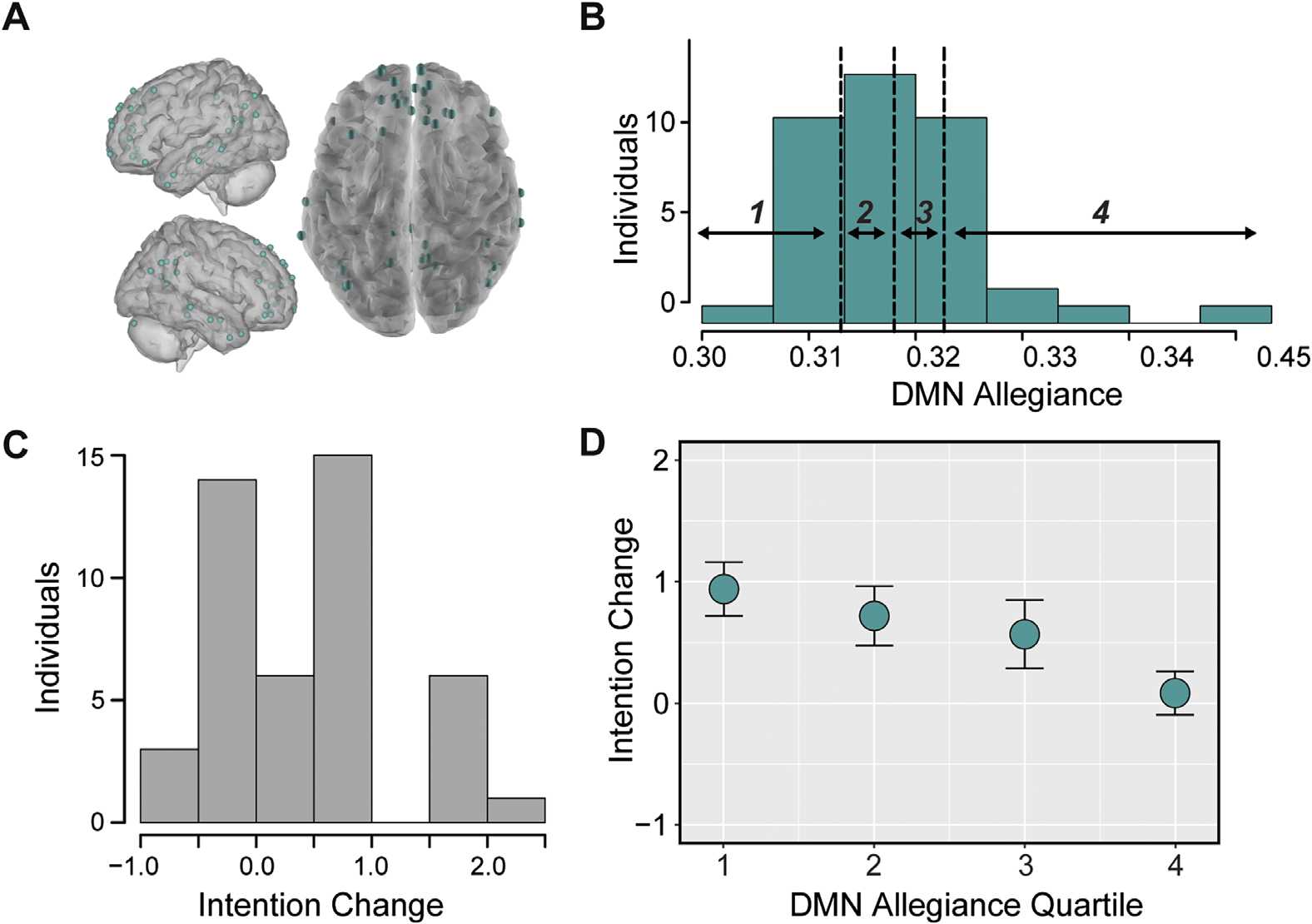
We first tested whether individual differences in allegiance between nodes within the atlas-
defined DMN, a network associated with social processing, self-relevance, valuation, memory,
and prospection, were related to message-consistent outcomes after the scanning session. We
averaged allegiance between all node pairs in the atlas-defined DMN (Figure 3A) and divided
individuals into quartiles based on this distribution (Figure 3B). We then related allegiance in
these quartiles to changes in participants’ intentions to quit smoking. A histogram of intention
change can be found in Figure 3C. We found that reduced allegiance between nodes within
the DMN predicted a greater increase in intentions to quit smoking (quartile robust regression,
Network Neuroscience
146
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 3. Reduced allegiance within the DMN precedes increased intention change. (A) Nodes
in the atlas-defined DMN. (B) Histogram of allegiance between pairs of nodes in the atlas-defined
DMN, averaged within individuals. These averages are divided into quartiles, with bin borders
noted as vertical dotted lines. (C) Histogram of changes in intentions from Session 1 to Session
3 for each individual, where positive value indicate an increased intention to change over time.
(D) Relationship between allegiance of nodes within the atlas-defined DMN and intention change,
where intention change was averaged in DMN allegiance quartiles. Error bars represent standard
error of the mean.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
t(38) = −2.86, p < 0.007; continuous variable robust regression t(38) = −1.99, p < 0.049;
see Figure 3D and Supporting Information Figure S1, Cooper et al., 2019), controlling for in-
tentions at baseline and demographic covariates.
In a parallel analysis, we examined the
relationship between allegiance in the atlas-defined DMN and behavior change. In our main
analysis, DMN allegiance was not significantly related to reductions in daily smoking (con-
tinuous robust regression t(39) = 1.41, p < 0.167), but these results became significant when
examining unsmoothed data (see Supporting Information).
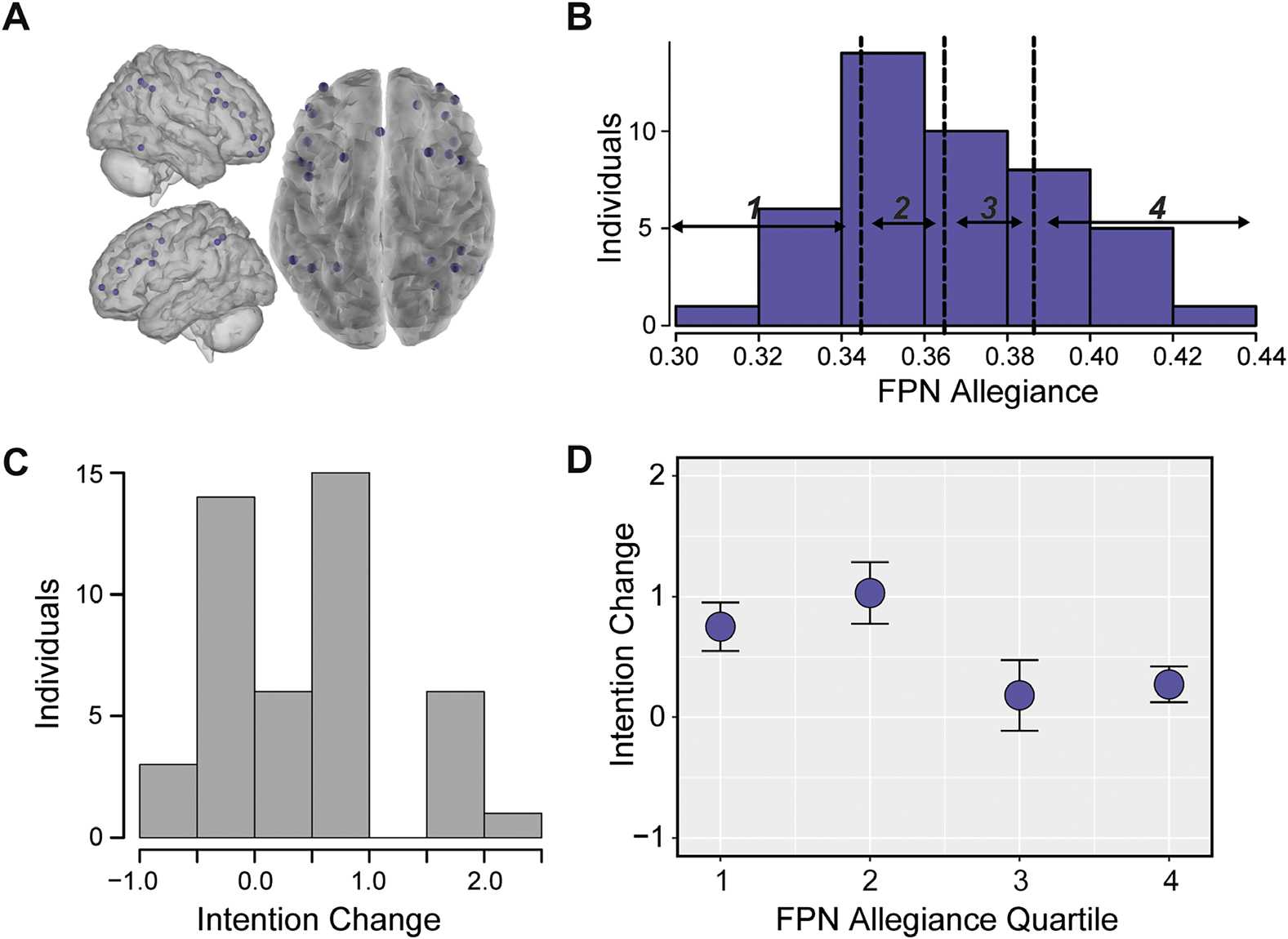
We repeated this analysis for the atlas-defined FPN, a network that has been associated with
decision-making and may play a critical role in belief updating. Following the same process
as DMN, we averaged allegiance between nodes in the FPN (Figure 4A) and divided this dis-
tribution into quartiles (Figure 4B). We then related allegiance in these quartiles to changes in
participants’ intentions to quit smoking, and our primary results identified that reduced alle-
giance in the FPN was also related to increased intentions (quartile, t(38) = −2.37, p < 0.021;
continuous variable robust regression t(38) = −2.10, p < 0.038; see Figure 4D and Support-
ing Information, Figure S2, Cooper et al., 2019). However, this relationship was trending in the
same direction but not significant using unsmoothed data (see Supporting Information). Alle-
giance in the FPN was not related to reductions in daily smoking (continuous robust regression
t(39) = 1.19, p < 0.238).
Network Neuroscience
147
Time-evolving brain dynamics forecast responses to health messaging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 4. Reduced allegiance within the FPN precedes increased intention change. (A) Nodes in
the atlas-defined FPN. (B) Histogram of allegiance between pairs of nodes in the atlas-defined FPN,
averaged within individuals. These averages are divided into quartiles, with bin borders noted as
dotted vertical lines. (C) Histogram of changes in intentions from Session 1 to Session 3 for each
individual. (D) Relationship between allegiance of nodes within the atlas-defined FPN and intention
change, where intention change was averaged within FPN allegiance quartiles. Error bars represent
standard error of the mean.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
We performed parallel analyses in the final two networks of interest, the salience and sub-
cortical networks, for a total of eight tests each of the relationship between network allegiance
and intention change, and network allegiance and behavior change for each subnetwork
(including the supplemental analyses using unsmoothed data). We found no significant rela-
tionships between allegiance and intentions (salience: continuous robust regression, t(38) =
− 0.27, p < 0.78; subcortical: continuous robust regression, t(38) = −0.11, p < 0.91) or
reductions in daily smoking (salience: continuous robust regression, t(39) = 1.14, p < 0.265;
subcortical: continuous robust regression, t(39) = 1.74, p < 0.097).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
vmPFC Flexibility Relates to Later Changes in Behavior
In our final analysis, given its particularly robust presence in the literature on behavior change,
we examined whether vmPFC demonstrated coordinated, but flexible, dynamics across mul-
tiple network communities. The vmPFC has been posited to be a hub of information process-
ing, integrating inputs about the self-relevance and valuation of information and influencing
decision-making, and localized activation in vmPFC has been frequently reported to predict
behavior changes following persuasive messaging. To complement these previous activation
findings and investigate the possible role of vmPFC in integrating information between multiple
network communities, we selected the node in the Power parcellation that was closest to the
center of mass of the vmPFC region identified as predictive of behavior change in a sunscreen
Network Neuroscience
148
Time-evolving brain dynamics forecast responses to health messaging
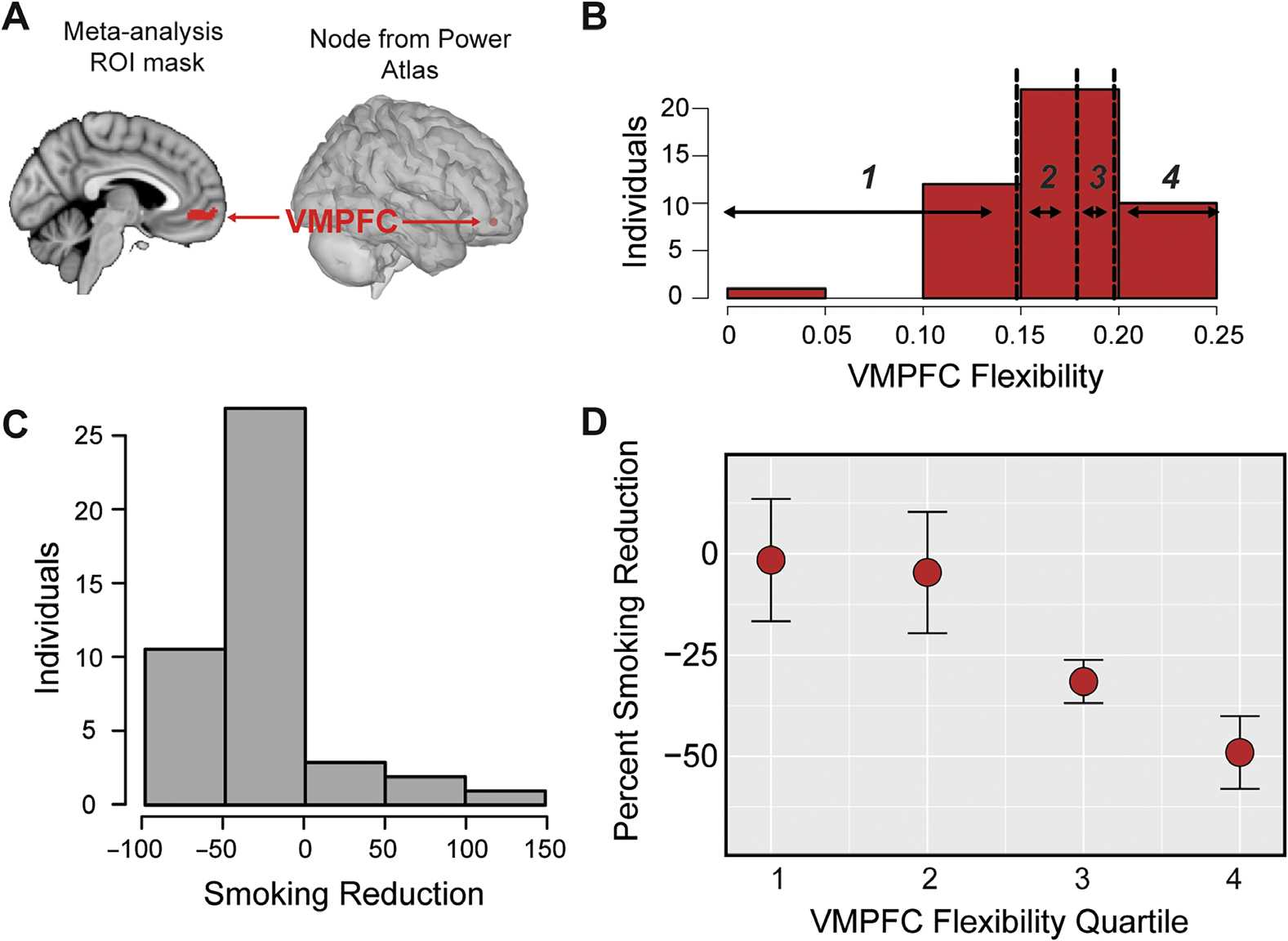
use study (Falk et al., 2010), shown in Figure 5A; this region has now been used to predict
behavior change in several contexts (Cooper et al., 2015; Falk et al., 2011, 2015; Riddle et al.,
2016). Of note, the same node is closest to the center of mass of the vmPFC region identified
as responding to subjective value in a value-based decision-making meta-analysis by Bartra
et al. (2013). This vmPFC node is classified as belonging to the DMN; however, we examined
vmPFC separately, as the default mode as defined in the Power atlas is a large network com-
posed of 45 nodes, and thus the behavior of the vmPFC node may not be representative of the
entire network (e.g., in past research on behavior change, vmPFC is robustly associated with
behavior change, but several regions of the DMN are not).
Our analysis evaluated the flexibility of vmPFC to quantify how often it changed com-
munity affiliations over time. We tested whether individuals who demonstrated differential
levels of flexibility in the vmPFC region showed corresponding variation in their intentions
to quit smoking or smoking behavior in the month following the scanning session (for a total
of 4 tests, including the Supporting Information using unsmoothed data, Cooper et al., 2019).
vmPFC flexibility (displayed in Figure 5B) was not significantly related to changes in intentions
using smoothed (continuous robust regression: t(38) = 1.55, p < 0.120) or unsmoothed data
(Supporting Information). We next examined the relationship between vmPFC flexibility and
behavior change (displayed in Figure 5C). vmPFC flexibility was significantly related to indi-
vidual differences in smoking reductions 1 month after the scan, such that individuals with
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. vmPFC flexibility relates to behavior change. (A) Region of vmPFC identified by Falk
et al. (2010) (left) and the closest Power parcellation node (right). (B) Histogram of vmPFC flexi-
bility in each individual, with the vertical dotted black line denoting the border for the quartile
bins. (C) Histogram of the percent change in cigarettes smoked per day in each individual, where
negative values indicate a reduction in cigarettes smoked per day. (D) Relationship between vmPFC
flexibility and behavior change, where behavior change was averaged within vmPFC allegiance
quartiles. Error bars represent standard error of the mean.
Network Neuroscience
149
Time-evolving brain dynamics forecast responses to health messaging
more flexible vmPFC network activity demonstrated larger reductions in their smoking behav-
ior using smoothed (quartile robust regression, t(39) = −2.93, p < 0.005; continuous measure
robust regression, t(39) = −2.85, p < 0.006) and unsmoothed data (Supporting Information);
see Figure 5D and Supporting Information Figure S3. This suggests that the network interactions
of vmPFC also capture an important component of its role in forecasting health outcomes.
DISCUSSION
Previous research has identified the critical role of regions in several brain networks for per-
suasion and successful behavior change, but to date research has not examined whether inter-
actions among these networks can account for individual differences in smoking outcomes.
Interactions between pairs of regions have been related to message effectiveness and behavior
change (Cooper et al., 2017, 2019; Dinh-Williams et al., 2014; Ramsay et al., 2013; Zelle et al.,
2017), and we extend this work by utilizing a large-scale network approach. We employed
recent advancements from network science to examine whether the synchrony within a net-
work community (allegiance among the brain regions in the community) or between-network
community interactions (flexibility of brain regions to coordinate across communities) ac-
counted for lasting changes in smoking outcomes. We find that dynamics in two networks, the
dafault mode (DMN) and fronto-parietal control (FPN) networks, may be relevant to smoking-
related outcomes. We also find that more frequent network changes in a key node of the DMN
consistently linked to predictions of behavior change, the ventromedial prefrontal cortex
(vmPFC), are associated with reductions in smoking behavior.
Relationship Between Network Allegiance and Changes in Smoking Intentions
Larger increases in intentions to quit smoking were related to reduced allegiance between
nodes belonging to the atlas-defined DMN and FPN, particularly in analyses using smoothed
data. In other words, there was lower consistent functional connectivity across the time course
of the task within regions in each of these networks for those individuals who showed an in-
crease in intentions to quit smoking. This reduction in network allegiance over the duration of
the task may reflect differential recruitment of nodes in each of these networks to interactions
with outside-network nodes, and it is plausible that this diversification of communication could
support long-term intention change. A point of interest in future investigations will be identi-
fying internetwork interactions that precede intention change, and examining whether these
interactions involve entire functional network communities or subsets of these atlas-defined
networks.
The finding of reduced allegiance within the atlas-defined FPN and DMN may be related
to the possible division of these networks into smaller modules dependent on context and
task demands. If these a priori networks are fractionated into modules that are more strongly
connected to other networks than to each other, this could result in reduced intranetwork
allegiance. Several studies identify meaningful subnetworks of both FPN and DMN; for
example, Spreng et al. (2013) and Dixon et al. (2018) find separate types of nodes within
the fronto-parietal control network, based on their interactions with other networks (Dixon
et al., 2018; Spreng, Sepulcre, Turner, Stevens, & Schacter, 2013). The DMN has also been
shown to be separable into subnetworks based on task-related functional connectivity (Dixon
et al., 2017; Fornito, Harrison, Zalesky, & Simons, 2012), and both DMN flexibility (Stanley,
Dagenbach, Lyday, Burdette, & Laurienti, 2014; Vatansever, Menon, Manktelow, Sahakian, &
Stamatakis, 2015) and its connectivity with other networks (Finc et al., 2017) can change with
task demands.
Network Neuroscience
150
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
The FPN has been posited to change its connectivity patterns in response to changes in
task demands to a greater extent than other functional networks (Cole et al., 2013), and such
changes in FPN connectivity have been previously reported to correlate with greater changes
in behavior. For example, reduced allegiance in hubs of the FPN predicted individual differ-
ences in learning (Bassett et al., 2015; Gerraty et al., 2018), as well as better performance on
working memory and executive cognition tasks (Braun et al., 2015). Although these previous
findings relating FPN connectivity changes to behavior come from other task domains, it is
possible that the core process—learning—is similar to what participants are experiencing dur-
ing exposure to persuasive messaging; in particular, the updating of beliefs during exposure
to self-relevant information from the messages could be akin to learning. The results linking
FPN to intention change, however, should be interpreted with caution, given that these results
were less robust using unsmoothed data (see Supporting Information, Cooper et al., 2019).
Relevance of vmPFC Flexibility for Smoking Behavior
We also find evidence for the importance of vmPFC flexibility.
Individuals who displayed
higher vmPFC flexibility, or switching of community affiliations, across the duration of the
task also reported larger reductions in their daily smoking levels 1 month later. Activation
in vmPFC during exposure to messaging has been repeatedly linked to long-term behavior
change (Chua et al., 2011; Cooper et al., 2015; Falk et al., 2011, 2015; Riddle et al., 2016;
Vezich et al., 2017; Wang et al., 2013), and it is possible that frequent community changes,
corresponding to high flexibility, relate to the activation levels detected in prior work. The
vmPFC has structural and functional connectivity with an array of regions in networks involving
memory, affective regulation, and higher-order cognition (Amodio & Frith, 2006; Buckner
et al., 2009; Price & Drevets, 2012; Roy, Shohamy, & Wager, 2012; Tomasi & Volkow, 2011).
Our result suggests that the time-varying strength of these connections may influence long-term
behavior. These results are also consistent with the possible broader relationship between re-
duced default mode allegiance and behavior change observed in our supplemental analyses
(see Supporting Information, Cooper et al., 2019) using unsmoothed data; vmPFC is one key
node in the default mode network, and greater flexibility in key nodes of the DMN would
correspondingly be related to lower allegiance.
We also find that changes in smoking behavior and intentions are related to partially di-
vergent metrics of neural dynamics. Several theories of health behavior posit that intentions
to perform a behavior are an important precursor to behavior change, but that other factors
also influence whether behavior change occurs (Ajzen, 1985, 1991; Armitage & Conner, 2001;
Fishbein, 1979; Fishbein & Ajzen, 2011; Webb & Sheeran, 2006). This partial dissociation be-
tween intentions and behavior seems to be reflected in the brain in this study, where we find
that some neural metrics related to changes in intentions are not related to changes in be-
havior, and vice versa. The present findings complement and extend previous neuroimaging
studies of behavior change in which intention changes do not mediate the relationship be-
tween vmPFC activation and behavior change (Cooper et al., 2015; Falk et al., 2010, 2011),
and also suggest that dynamics between subportions of the DMN may be worth exploring
to bridge the neural underpinnings of intentions and behaviors. Together, these reports sug-
gest that different neurocognitive processes during initial exposure may support the evaluation
of intentions to perform a behavior, and the additional cognitions and actions that result in
longitudinal behavior change. These results highlight promise in additional research to build a
more a complete model of the relationship between immediate brain responses to persuasive
messaging, and later outcomes such as self-reported intentions and behavior.
Network Neuroscience
151
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
Future Directions and Limitations
The underlying origin of individual differences in functional connectivity dynamics is an open
question and an intriguing avenue for future research. Here, we expect that some individuals
are more susceptible to persuasion through health messaging than others, and that we can
detect this propensity by assessing network dynamics during the task. Furthermore, this could
be due to differences in intrinsic dynamics of the networks of interest (i.e., a person-level fac-
tor), differences in the dynamics associated with processing the anti-smoking images (i.e., a
message-level factor), or both (i.e., an interaction between the two). Although individual differ-
ences in networks may suggest that regional activity differences have a trait-like component,
our inclination is to avoid a hard split between context dependence and traits, and instead
consider the importance of varying timescales when considering context versus trait effects.
That is, individual differences may appear trait-like when brain activity is observed in a single
session or narrow time frame, but these between-subject differences may show more context-
based effects when examined over longitudinal timescales and in response to different types
of stimuli. We expect that network dynamics in the same individual might vary depending on
the task presented (e.g., different message frames), although we do not directly investigate task
conditions in this analysis; that is, we expect that network dynamics during this anti-smoking
task may be related to changes in future smoking behavior, but not necessarily to other be-
havior domains, or even to different types of message approaches. Whether the effects we
observe here are more strongly related to task-related processing or intrinsic dynamics could
have differing implications for the design of more effective health messaging campaigns and
broader questions about persuasion and influence, and hence provide valuable directions for
future research; for example, the former would implicate the need for changes in the design
of messages, and the latter might suggest participant-level interventions to improve receptivity
to messaging (such as self-affirmation, as in Epton & Harris, 2008; Epton, Harris, Kane, van
Koningsbruggen, & Sheeran, 2015; MCQueen & Klein, 2006; Taber, Klein, Ferrer, Augustson,
& Patrick, 2016). Thus, the extent to which network dynamics vary across different timescales
and in response to different task domains is an important question for future work.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
The difference in functional dynamics between task conditions, such as the negative and
neutral anti-smoking messages presented here, might also provide further insight into the
mechanism of the effects we identify in this report. Specifically, this could aid in understanding
what message characteristics are important for changes in brain response and later behavior,
and whether the effects we report in the current manuscript stem from stable trait-like neural
tendencies, context-dependent shifts in brain dynamics, or an interaction between the two.
However, constraints of the task design in this experiment (namely, short stimulus presentation
times and a relatively small number of stimuli presented) prevented the estimation of func-
tional connectivity dynamics separately between task conditions. Future work incorporating
slower and longer task designs will provide insightful extensions of our results, advancing our
understanding about message characteristics important for behavior change.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Conclusions
Here we investigated the relationship between the time-varying nature of brain activity during
exposure to anti-smoking messages and future changes in smoking behavior and intentions to
quit. We found robust evidence that reduced allegiance within the atlas-defined DMN related
to changes in intentions to quit smoking and that flexibility in the vmPFC related to changes
in smoking behavior, and suggestive evidence that reduced allegiance in the FPN related to
intention change. There is increasing recognition that consideration of brain networks and
Network Neuroscience
152
Time-evolving brain dynamics forecast responses to health messaging
their dynamics, and not just activation in individual regions, is necessary for understanding
human cognition and behavior; here, we show that metrics of functional dynamics can provide
new information about individual differences in responsiveness to anti-smoking messaging.
These results highlight the value in considering brain network dynamics for understanding
message effectiveness and social processes more broadly.
ACKNOWLEDGMENTS
We acknowledge Richard Gonzalez, Sonya Dal Cin, Victor Strecher, and Lawrence An for
collaboration on a larger study relevant to this work; and Francis Tinney Jr., Kristin Shumaker,
Li Chen, Nicolette Gregor, Becky Lau, Larissa S. Svintsitski, and Cole Schaffer for assistance
with data collection.
AUTHOR CONTRIBUTIONS
Nicole Cooper: Conceptualization; Data curation; Formal analysis; Funding acquisition;
Methodology; Project administration; Software; Validation; Visualization; Writing – original
draft; Writing – review & editing.
Javier O. Garcia: Conceptualization; Formal analysis;
Methodology; Resources; Software; Validation; Visualization; Writing – review & editing.
Steven Tompson: Conceptualization; Data curation; Investigation; Software; Writing – review
& editing. Matthew B. O’Donnell: Conceptualization; Data curation; Investigation; Resources;
Software; Writing – review & editing. Emily B. Falk: Conceptualization; Funding acquisition;
Project administration; Resources; Supervision; Writing – original draft; Writing – review
Jean M. Vettel: Conceptualization; Funding acquisition; Resources; Supervision;
& editing.
Writing – original draft; Writing – review & editing.
FUNDING INFORMATION
Emily B Falk and Jean M Vettel, Army Research Laboratory (http://dx.doi.org/10.13039/
100006754), Award ID: W911NF-10-2-0022. PI Victor Strecher and Co-I Emily B Falk, Na-
tional Institutes of Health (http://dx.doi.org/10.13039/100000002), Award ID: P50 CA101451.
Emily B Falk, National Institutes of Health (http://dx.doi.org/10.13039/100000002), Award ID:
1DP2DA03515601. PI Emily B Falk and Co-I Nicole Cooper, National Institutes of Health/
National Cancer Institute and FDA Center for Tobacco Products (http://dx.doi.org/10.13039/
100000002), Award ID: P50CA179546. The content is solely the responsibility of the authors
and does not necessarily represent the official views of the NIH, the Food and Drug Adminis-
tration (FDA), the Army Research Office, or the U.S. Government.
REFERENCES
Ajzen, I. (1985). From intentions to actions: A theory of planned
behavior. In Action control (pp. 11–39). Springer: Berlin.
Ajzen, I. (1991). The theory of planned behavior. Organizational
Behavior and Human Decision Processes, 50(2), 179–211.
Alakörkkö,T.,Saarimäki,H.,Glerean,E.,Saramäki,J.,&Korhonen,O.
(2017). Effects of spatial smoothing on functional brain networks.
The European Journal of Neuroscience, 46(9), 2471–2480.
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: The medial
frontal cortex and social cognition. Nature Reviews. Neuro-
science, 7(4), 268–277.
Armitage, C. J., & Conner, M.
(2001). Efficacy of the theory of
planned behaviour: A meta-analytic review. British Journal of
Social Psychology, 40(Pt 4), 471–499.
Ashourvan, A., Gu, S., Mattar, M. G., Vettel, J. M., & Bassett, D. S.
(2017). The energy landscape underpinning module dynamics
in the human brain connectome. NeuroImage, 157, 364–380.
Barrett, L. F., & Satpute, A. B. (2013). Large-scale brain networks in
affective and social neuroscience: Towards an integrative func-
tional architecture of the brain. Current Opinion in Neurobiol-
ogy, 23(3), 361–372.
Bartra, O., McGuire, J. T., & Kable, J. W.
(2013). The valuation
system: A coordinate-based meta-analysis of BOLD fMRI exper-
iments examining neural correlates of subjective value. Neuro-
Image, 76, 412–427.
Bassett, D. S., Porter, M. A., Wymbs, N. F., Grafton, S. T., Carlson,
(2013). Robust detection of dynamic
J. M., & Mucha, P. J.
Network Neuroscience
153
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
community structure in networks. Chaos: An Interdisciplinary
Journal of Nonlinear Science, 23(1), 013142.
online antismoking messages predicts behavior change. Journal
of Media Psychology, 27(3), 93–108.
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
(2011). Dynamic reconfiguration of hu-
J. M., & Grafton, S. T.
man brain networks during learning. Proceedings of the National
Academy of Sciences of the United States of America. https://doi.
org/10.1073/pnas.1018985108
Bassett, D. S., Wymbs, N. F., Rombach, M. P., Porter, M. A., Mucha,
P. J., & Grafton, S. T. (2013). Task-based core-periphery organ-
ization of human brain dynamics. PLoS Computational Biology,
9(9), e1003171.
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T. (2015).
Learning-induced autonomy of sensorimotor systems. Nature
Neuroscience, 18(5), 744–751.
Berkman, E. T., & Falk, E. B. (2013). Beyond brain mapping: Using
neural measures to predict real-world outcomes. Current Direc-
tions in Psychological Science, 22(1), 45–50.
Berns, G. S., & Moore, S. E. (2012). A neural predictor of cultural
popularity. Journal of Consumer Psychology, 22(1), 154–160.
Braun, U., Schäfer, A., Walter, H., Erk, S., Romanczuk-Seiferth, N.,
Haddad, L., . . . Bassett, D. S. (2015). Dynamic reconfiguration
of frontal brain networks during executive cognition in humans.
Proceedings of the National Academy of Sciences of the United
States of America, 112(37), 11678–11683.
Bressler, S. L., & Menon, V. (2010). Large-scale brain networks in
cognition: emerging methods and principles. Trends in Cognitive
Sciences, 14(6), 277–290.
Buckner, R. L., Sepulcre, J., Talukdar, T., Krienen, F. M., Liu, H.,
Hedden, T., . . . Johnson, K. A. (2009). Cortical hubs revealed
by intrinsic functional connectivity: Mapping, assessment of sta-
bility, and relation to Alzheimer’s disease. The Journal of Neuro-
science, 29(6), 1860–1873.
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews. Neuroscience, 10(3), 186–198.
Chen, Z., & Calhoun, V. (2018). Effect of spatial smoothing on task
fMRI ICA and functional connectivity. Frontiers in Neuroscience,
12, 15.
Chua, H. F., Ho, S. S., Jasinska, A. J., Polk, T. A., Welsh, R. C.,
(2011). Self-related neural re-
Liberzon, I., & Stretcher, V. J.
sponse to tailored smoking-cessation messages predicts quitting.
Nature Neuroscience, 14(4), 426–427.
Cole, M. W., Reynolds, J. R., Power, J. D., Repovs, G., Anticevic,
A., & Braver, T. S. (2013). Multi-task connectivity reveals flexi-
ble hubs for adaptive task control. Nature Neuroscience, 16(9),
1348–1355.
Cooper, N., Bassett, D. S., & Falk, E. B. (2017). Coherent activity
between brain regions that code for value is linked to the mal-
leability of human behavior. Scientific Reports, 7, 43250.
Cooper, N., Garcia, J. O., Tompson, S., O’Donnell, M. B., Falk,
E. B., & Vettel, J. M. (2019). Supporting Information for “Time-
evolving dynamics in brain networks forecast responses to health
messaging.” Network Neuroscience, 3(1), 138–156. https://doi.
org/10.1162/netn_a_00058
Cooper, N., Tompson, S., O’Donnell, M. B., & Falk, E. B.
(2015).
Brain activity in self- and value-related regions in response to
Cooper, N., Tompson, S., O’Donnell, M. B., Vettel, J. M., Bassett,
D. S., & Falk, E. B. (2018). Associations between coherent
neural activity in the brain’s value system during antismoking
messages and reductions in smoking. Health Psychology, 37(4),
375–384.
Costafreda, S. G., Khanna, A., Mourao-Miranda, J., & Fu, C. H. Y.
(2009). Neural correlates of sad faces predict clinical remission to
cognitive behavioural therapy in depression. Neuroreport, 20(7),
637–641.
Cox, R. W.
(1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29(3), 162–173.
Deng, Z., Chandrasekaran, B., Wang, S., & Wong, P. C. M. (2016).
Resting-state low-frequency fluctuations reflect individual differ-
ences in spoken language learning. Cortex, 76, 63–78.
Dinh-Williams, L., Mendrek, A., Dumais, A., Bourque, J., & Potvin,
S.
(2014). Executive-affective connectivity in smokers viewing
anti-smoking images: An fmri study. Psychiatry Research, 224(3),
262–268.
Dixon, M. L., Andrews-Hanna, J. R., Spreng, R. N., Irving, C., Mills,
C., Girn, M., & Christoff, K.
Interactions between the
default network and dorsal attention network vary across de-
fault subsystems, time, and cognitive states. NeuroImage, 147,
632–639.
(2017).
Dixon, M. L., De La Vega, A., Mills, C., Andrews-Hanna, J., Spreng,
R. N., Cole, M. W., & Christoff, K. (2018). Heterogeneity within
the frontoparietal control network and its relationship to the de-
fault and dorsal attention networks. Proceedings of the National
Academy of Sciences of the United States of America. https://
doi.org/10.1073/pnas.1715766115
Doehrmann, O., Ghosh, S. S., Polli, F. E., Reynolds, G. O., Horn,
F., Keshavan, A.,
(2013). Predicting
. Gabrieli,
treatment response in social anxiety disorder from functional
magnetic resonance imaging. JAMA Psychiatry, 70(1), 87–97.
Epton, T., & Harris, P. R. (2008). Self-affirmation promotes health
J. D.
.
.
behavior change. Health Psychology, 27(6), 746–752.
Epton, T., Harris, P. R., Kane, R., van Koningsbruggen, G. M., &
(2015). The impact of self-affirmation on health-
Sheeran, P.
behavior change: A meta-analysis. Health Psychology, 34(3),
187–196.
Etter, J. F., Vu Duc, T., & Perneger, T. V. (2000). Saliva cotinine levels
in smokers and nonsmokers. American Journal of Epidemiology,
151(3), 251–258.
Falk, E. B., Berkman, E. T., Mann, T., Harrison, B., & Lieberman.
(2010). Predicting persuasion-induced behavior change from
the brain. Journal of Neuroscience, 30(25), 8421–8424.
Falk, E. B., Berkman, E. T., Whalen, D., & Lieberman. (2011). Neural
activity during health messaging predicts reductions in smoking
above and beyond self-report. Health Psychology, 30(2), 177–185.
Falk, E. B., O’Donnell, M. B., Cascio, C. N., Tinney, F., Kang,
Y., Lieberman, . . . Stretcher, V. J. (2015). Self-affirmation alters
the brain’s response to health messages and subsequent behav-
ior change. Proceedings of the National Academy of Sciences
of the United States of America, 112(7), 1977–1982.
Network Neuroscience
154
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
Falk, E. B., & Scholz, C.
influence, and
value: perspectives from communication and social neuro-
science. Annual Review of Psychology. https://doi.org/10.1146/
annurev-psych-122216-011821
(2017). Persuasion,
Feldstein Ewing, S. W., Chung, T., Caouette, J. D., Ketcherside,
A., Hudson, K. A., & Filbey, F. M. (2017). Orbitofrontal cortex
connectivity as a mechanism of adolescent behavior change.
NeuroImage, 151, 14–23.
Finc, K., Bonna, K., Lewandowska, M., Wolak, T., Nikadon, J.,
Dreszer, J., . . . Kühn, S. (2017). Transition of the functional brain
network related to increasing cognitive demands. Human Brain
Mapping. https://doi.org/10.1002/hbm.23621
Fishbein, M.
(1979). A theory of reasoned action: Some appli-
cations and implications. Nebraska Symposium on Motivation.
Nebraska Symposium on Motivation, 27, 65–116.
Fishbein, M., & Ajzen, I. (2011). Predicting and Changing Behavior:
The Reasoned Action Approach. Hoboken, NJ: Taylor & Francis.
Fornito, A., Harrison, B. J., Zalesky, A., & Simons, J. S. (2012). Com-
petitive and cooperative dynamics of large-scale brain functional
networks supporting recollection. Proceedings of the National
Academy of Sciences of the United States of America, 109(31),
12788–12793.
Fox, M. D., & Raichle, M. E. (2007). Spontaneous fluctuations
in brain activity observed with functional magnetic resonance
imaging. Nature Reviews. Neuroscience, 8(9), 700–711.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences of the United
States of America, 102(27), 9673–9678.
Friston, K. J. (1994). Functional and effective connectivity in neu-
roimaging: A synthesis. Human Brain Mapping, 2(1-2), 56–78.
Friston, K. J. (2009). Modalities, Modes, and models in functional
neuroimaging. Science, 326(5951), 399–403.
Gabrieli, J. D. E., Ghosh, S. S., & Whitfield-Gabrieli, S.
(2015).
Prediction as a humanitarian and pragmatic contribution from
human cognitive neuroscience. Neuron, 85(1), 11–26.
Genevsky, A., & Knutson, B. (2015). Neural affective mechanisms
predict market-level microlending. Psychological Science, 26(9),
1411–1422.
Gerraty, R. T., Davidow, J. Y., Foerde, K., Galvan, A., Bassett, D. S.,
& Shohamy, D. (2018). Dynamic flexibility in striatal-cortical cir-
cuits supports reinforcement learning. Journal of Neuroscience,
094383.
Good, B. H., de Montjoye, Y.-A., & Clauset, A. (2010). Performance
of modularity maximization in practical contexts. Physical Re-
view. E, Statistical, Nonlinear, and Soft Matter Physics, 81(4 Pt 2),
046106.
Greicius, M. D., Krasnow, B., Reiss, A. L., & Menon, V. (2003).
Functional connectivity in the resting brain: A network analysis
of the default mode hypothesis. Proceedings of the National Acad-
emy of Sciences of the United States of America, 100(1), 253–258.
Jarvis, M. J., Tunstall-Pedoe, H., Feyerabend, C., Vesey, C., &
Saloojee, Y. (1987). Comparison of tests used to distinguish
smokers from nonsmokers. American Journal of Public Health,
77(11), 1435–1438.
Jasinska, A. J., Chua, H. F., Ho, S. S., Polk, T. A., Rozek, L. S., &
Strecher, V. J. (2012). Amygdala response to smoking-cessation
messages mediates the effects of serotonin transporter gene vari-
ation on quitting. NeuroImage, 60(1), 766–773.
Kaye, S.-A., White, M. J., & Lewis, I. (2017). The use of neuro-
cognitive methods in assessing health communication mes-
sages: A systematic review. Journal of Health Psychology, 22(12),
1534–1551.
Kühn, S., Strelow, E., & Gallinat, J. (2016). Multiple “buy buttons”
in the brain: Forecasting chocolate sales at point-of-sale based
on functional brain activation using fMRI. NeuroImage, 136,
122–128.
Laird, A. R., Fox, P. M., Eickhoff, S. B., Turner, J. A., Ray, K. L.,
McKay, D. R., . . . Fox, P. T. (2011). Behavioral interpretations
Journal of Cognitive Neuro-
of intrinsic connectivity networks.
science, 23(12), 4022–4037.
Lange, J., Oostenveld, R., & Fries, P. (2013). Reduced occipital
alpha power indexes enhanced excitability rather than im-
Journal of Neuroscience, 33(7),
proved visual perception.
3212–3220.
Liang, X., Zou, Q., He, Y., & Yang, Y.
(2016). Topologically re-
organized connectivity architecture of default-mode, executive-
control, and salience networks across working memory task
loads. Cerebral Cortex, 26(4), 1501–1511.
Lopez, R. B., Chen, P.-H. A., Huckins, J. F., Hofmann, W., Kelley,
W. M., & Heatherton, T. F. (2017). A balance of activity in
brain control and reward systems predicts
self-regulatory
outcomes. Social Cognitive and Affective Neuroscience, 12(5),
832–838.
MCQueen, A., & Klein, W. M. P. (2006). Experimental manipula-
tions of self-affirmation: A systematic review. Self and Identity,
5(4), 289–354.
Medaglia, J. D., Lynall, M.-E., & Bassett, D. S.
(2015). Cognitive
network neuroscience. Journal of Cognitive Neuroscience, 27(8),
1471–1491.
Menon, V. (2011). Large-scale brain networks and psychopathology:
A unifying triple network model. Trends in Cognitive Sciences,
15(10), 483–506.
Middleton, E. T., & Morice, A. H. (2000). Breath carbon monoxide
as an indication of smoking habit. Chest, 117(3), 758–763.
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J.-P. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Science, 328(5980), 876–878.
Patrick, D. L., Cheadle, A., Thompson, D. C., Diehr, P., Koepsell,
T., & Kinne, S. (1994). The validity of self-reported smoking: a
review and meta-analysis. American Journal of Public Health,
84(7), 1086–1093.
Pokorski, T. L., Chen, W. W., & Bertholf, R. L. (1994). Use of urine
cotinine to validate smoking self-reports in U.S. Navy recruits.
Addictive Behaviors, 19(4), 451–454.
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., . . . Petersen, S. E. (2011). Functional network
organization of the human brain. Neuron, 72(4), 665–678.
Price, J. L., & Drevets, W. C. (2012). Neural circuits underlying
the pathophysiology of mood disorders. Trends in Cognitive
Sciences, 16(1), 61–71.
Network Neuroscience
155
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Time-evolving brain dynamics forecast responses to health messaging
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of brain
function. Proceedings of the National Academy of Sciences of the
United States of America, 98(2), 676–682.
Ramsay, I. S., Yzer, M. C., Luciana, M., Vohs, K. D., & MacDonald,
A. W., 3rd. (2013). Affective and executive network processing
associated with persuasive antidrug messages. Journal of Cogni-
tive Neuroscience, 25(7), 1136–1147.
Riddle, P. J., Newman-Norlund, R. D., Baer, J., & Thrasher, J. F.
(2016). Neural response to pictorial health warning labels can
predict smoking behavioral change. Social Cognitive and Affec-
tive Neuroscience, nsw087.
Roy, M., Shohamy, D., & Wager, T. D.
(2012). Ventromedial
prefrontal-subcortical systems and the generation of affective
meaning. Trends in Cognitive Sciences, 16(3), 147–156.
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover, G. H.,
Kenna, H., . . . Greicius, M. D. (2007). Dissociable intrinsic con-
nectivity networks for salience processing and executive control.
Journal of Neuroscience, 27(9), 2349–2356.
Shirer, W. R., Ryali, S., Rykhlevskaia, E., Menon, V., & Greicius, M. D.
(2012). Decoding subject-driven cognitive states with whole-
brain connectivity patterns. Cerebral Cortex, 22(1), 158–165.
Smith, S. M., Fox, P. T., Miller, K. L., Glahn, D. C., Fox, P. M.,
Mackay, C. E., . . . Beckmann, C. F.
(2009). Correspondence
of the brain’s functional architecture during activation and rest.
Proceedings of the National Academy of Sciences of the United
States of America, 106(31), 13040–13045.
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C.
(2004).
Organization, development and function of complex brain net-
works. Trends in Cognitive Sciences, 8(9), 418–425.
Sporns, O., Tononi, G., & Edelman, G. M. (2000). Connectivity and
complexity: the relationship between neuroanatomy and brain
dynamics. Neural Networks, 13(8-9), 909–922.
Spreng, R. N., Sepulcre,
J., Turner, G. R., Stevens, W. D., &
Intrinsic architecture underlying the
Schacter, D. L.
relations among the default, dorsal attention, and frontopari-
Journal of Cognitive
etal control networks of the human brain.
Neuroscience, 25(1), 74–86.
(2013).
Stanley, M. L., Dagenbach, D., Lyday, R. G., Burdette, J. H., &
Laurienti, P. J. (2014). Changes in global and regional modular-
ity associated with increasing working memory load. Frontiers in
Human Neuroscience, 8, 954.
Sun, F. T., Miller, L. M., & D’Esposito, M. (2004). Measuring in-
terregional functional connectivity using coherence and partial
coherence analyses of fMRI data. NeuroImage, 21(2), 647–658.
Taber, J. M., Klein, W. M. P., Ferrer, R. A., Augustson, E., & Patrick,
H.
(2016). A pilot test of self-affirmations to promote smoking
cessation in a national smoking cessation text messaging pro-
gram. JMIR mHealth and uHealth, 4(2), e71.
Telesford, Q. K., Lynall, M.-E., Vettel, J., Miller, M. B., Grafton, S. T.,
& Bassett, D. S.
(2016). Detection of functional brain network
reconfiguration during task-driven cognitive states. NeuroImage,
142, 198–210.
Tomasi, D., & Volkow, N. D. (2011). Functional connectivity hubs
in the human brain. NeuroImage, 57(3), 908–917.
Tottenham, N., Tanaka, J. W., Leon, A. C., McCarry, T., Nurse, M.,
Hare, T. A., . . . Nelson, C. (2009). The NimStim set of facial
expressions:
Judgments from untrained research participants.
Psychiatry Research, 168(3), 242–249.
van Dijk, H., Schoffelen,
J.-M., Oostenveld, R., & Jensen, O.
(2008). Prestimulus oscillatory activity in the alpha band predicts
visual discrimination ability. Journal of Neuroscience, 28(8),
1816–1823.
Vartiainen, E., Seppälä, T., Lillsunde, P., & Puska, P. (2002). Valida-
tion of self reported smoking by serum cotinine measurement
in a community-based study. Journal of Epidemiology and Com-
munity Health, 56(3), 167–170.
Vatansever, D., Menon, D. K., Manktelow, A. E., Sahakian,
B. J., & Stamatakis, E. A. (2015). Default Mode Dynamics For
Global Functional Integration. Journal Of Neuroscience, 35(46),
15254–15262.
Venkatraman, V., Dimoka, A., Pavlou, P. A., Vo, K., Hampton,
W., Bollinger, B., . . . Winer, R. S. (2015). Predicting advertising
success beyond traditional measures: New insights from neuro-
physiological methods and market response modeling. Journal of
Marketing Research, 52(4), 436–452.
Vezich, S., Katzman, P. L., Ames, D. L., Falk, E. B., & Lieberman.
(2017). Modulating the neural bases of persuasion: why/how,
gain/loss, and users/non-users. Social Cognitive and Affective
Neuroscience, nsw113.
Wang, A.-L., Ruparel, K., Loughead, J. W., Strasser, A. A., Blady,
S. J., Lynch, K. G.,
. . . Langleben, D. D. (2013). Content
matters: Neuroimaging investigation of brain and behavioral
impact of televised anti-tobacco public service announcements.
Journal of Neuroscience, 33(17), 7420–7427.
Wang, C., Ong, J. L., Patanaik, A., Zhou, J., & Chee, M. W. L.
(2016). Spontaneous eyelid closures link vigilance fluctuation
with fMRI dynamic connectivity states. Proceedings of
the
National Academy of Sciences of the United States of America,
113(34), 9653–9658.
Webb, T. L., & Sheeran, P. (2006). Does changing behavioral
intentions engender behavior change? A meta-analysis of the
experimental evidence. Psychological Bulletin, 132(2), 249–268.
Weber, R., Huskey, R., Mangus, J. M., Westcott-Baker, A., & Turner,
B. O. (2015). Neural predictors of message effectiveness during
counterarguing in antidrug campaigns. Communication Mono-
graphs, 82(1), 4–30.
Wilcox, C. E., Calhoun, V. D., Rachakonda, S., Claus, E. D.,
. . . Hutchison, K. E. (2017).
J.,
Littlewood, R. A., Mickey,
Functional network connectivity predicts treatment outcome
during treatment of nicotine use disorder. Psychiatry Research,
265, 45–53.
Yang, D., Pelphrey, K. A., Sukhodolsky, D. G., Crowley, M. J.,
Dayan, E., Dvornek, N. C., . . . Ventola, P. (2016). Brain re-
sponses to biological motion predict treatment outcome in young
children with autism. Translational Psychiatry, 6(11), e948.
Zelle, S. L., Gates, K. M., Fiez, J. A., Sayette, M. A., & Wilson, S. J.
(2017). The first day is always the hardest: Functional connectiv-
ity during cue exposure and the ability to resist smoking in the
initial hours of a quit attempt. NeuroImage, 151, 24–32.
Network Neuroscience
156
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
3
8
1
0
9
2
3
4
8
n
e
n
_
a
_
0
0
0
5
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3