INVESTIGACIÓN
Morphometric features of drug-resistant essential
tremor and recovery after stereotactic
radiosurgical thalamotomy
Thomas A. W.. Bolton1,2
, Dimitri Van De Ville3,4, Jean Régis5, Tatiana Witjas6,
Nadine Girard7, Marc Levivier1,8, and Constantin Tuleasca1,8,9
1Department of Clinical Neurosciences, Neurosurgery Service and Gamma Knife Center,
Centre Hospitalier Universitaire Vaudois (CHUV ), Lausanne, Suiza
2Connectomics Laboratory, Department of Radiology, Centre Hospitalier Universitaire Vaudois (CHUV ), Lausanne, Suiza
3Institute of Bioengineering, Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Suiza
4Department of Radiology and Medical Informatics, University of Geneva (UNIGE), Geneva, Suiza
5Stereotactic and Functional Neurosurgery Service and Gamma Knife Unit, Assistance Publique-Hôpitaux de Marseille,
Centre Hospitalier Universitaire de la Timone, Marsella, Francia
6Neurology Department, Assistance Publique-Hôpitaux de Marseille, Centre Hospitalier Universitaire de la Timone,
Marsella, Francia
7Department of Diagnostic and Interventional Neuroradiology, Centre de Résonance Magnétique Biologique et Médicale,
Assistance Publique-Hôpitaux de Marseille, Centre Hospitalier Universitaire de la Timone, Marsella, Francia
8Faculty of Biology and Medicine (FBM), University of Lausanne (UNIL), Lausanne, Suiza
9Signal Processing Laboratory (LTS 5), Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Suiza
Palabras clave: Essential tremor, Radiosurgery, Thalamotomy, Ventro-intermediate nucleus, Structural
covariance analysis, Morphometry, Cortical thickness, Surface area, Mean curvature
ABSTRACTO
Essential tremor (ET) is the most common movement disorder. Its neural underpinnings remain
unclear. Aquí, we quantified structural covariance between cortical thickness (CT), surface
área (SA), and mean curvature (MC) estimates in patients with ET before and 1 year after
ventro-intermediate nucleus stereotactic radiosurgical thalamotomy, and contrasted the
observed patterns with those from matched healthy controls. For SA, complex rearrangements
within a network of motion-related brain areas characterized patients with ET. This was
complemented by MC alterations revolving around the left middle temporal cortex and the
disappearance of positive-valued covariance across both modalities in the right fusiform gyrus.
Recovery following thalamotomy involved MC readjustments in frontal brain centers, el
amygdala, and the insula, capturing nonmotor characteristics of the disease. The appearance
of negative-valued CT covariance between the left parahippocampal gyrus and hippocampus
was another recovery mechanism involving high-level visual areas. This was complemented by
the appearance of negative-valued CT/MC covariance, and positive-valued SA/MC covariance,
in the right inferior temporal cortex and bilateral fusiform gyrus. Our results demonstrate that
different morphometric properties provide complementary information to understand ET, y
that their statistical cross-dependences are also valuable. They pinpoint several anatomical
features of the disease and highlight routes of recovery following thalamotomy.
RESUMEN DEL AUTOR
Doubts remain regarding the anatomical alterations underlying essential tremor, partly owing
to heterogeneity in symptoms’ severity and response to medication. Aquí, we studied drug-
resistant patients clinically assessed and imaged before as well as 1 year after stereotactic
un acceso abierto
diario
Citación: Bolton, t. A. w., Van De Ville,
D., Régis, J., Witjas, T., Girard, NORTE.,
Levivier, METRO., & Tuleasca, C. (2022).
Morphometric features of drug-
resistant essential tremor and recovery
after stereotactic radiosurgical
thalamotomy. Neurociencia en red,
6(3), 850–869. https://doi.org/10.1162
/netn_a_00253
DOI:
https://doi.org/10.1162/netn_a_00253
Supporting Information:
https://doi.org/10.1162/netn_a_00253
Recibió: 16 Febrero 2022
Aceptado: 2 Puede 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Thomas A. W.. Bolton
thomas.bolton@chuv.ch
Editor de manejo:
Andrew Zalesky
Derechos de autor: © 2022
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
norte
mi
norte
_
a
_
0
0
2
5
3
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
radiosurgical thalamotomy, which significantly lowered tremor intensity. We extracted
morphometric estimates of volume (subcortex and cerebellum), cortical thickness, surface
área, and mean curvature (corteza), and quantified cross-regional statistical dependences across
subjects (es decir., structural covariance or SC) for each measure, as well as cross-measure
relationships for each region. Compared to matched healthy controls, patients showed altered
surface area structural covariance within motion-related areas. Thalamotomy modulated mean
curvature SC in frontal and subcortical centers. In both comparisons, SC and cross-measure
relationship differences were also observed in visual areas.
INTRODUCCIÓN
Essential tremor (ET) is the most common movement disorder, affecting up to 1% of the pop-
ulation and 5% of individuals above 65 years of age (luis & Ferreira, 2010). The main diag-
nosis is clinical. Patients present with postural and kinetic tremor of the upper limbs (manos
y brazos), sometimes completed by further head, legs, or voice tremor (Chunling & Zheng,
2016). In addition to motor symptoms, it has become increasingly clear that there are other
possible concomitant manifestations of the disease, including cognitive deficits, psychiatric
disorders, or sensory deficiencies (Chandran & Pal, 2012; Jhunjhunwala & Pal, 2014; luis,
2016).
The exact etiology of ET remains largely elusive. While a genetic origin is unequivocal, como
más que 50% of individuals have a positive family history, no ET-specific gene mutation has
so far been reliably identified, and genome-wide association studies have only enabled
researchers to pinpoint few disease-causing genetically complex variants (Kuhlenbäumer
et al., 2014; Siokas et al., 2020; Tio & Broncearse, 2016). Candidate genes are involved in a wide
array of functions, as they may code for ion channels, partake in neuroregeneration, or con-
tribute to axonal myelination (Deng et al., 2019). Además, environmental and epigenetic
influences are also hypothesized (Hopfner & Helmich, 2018).
This complex etiology translates to a pathophysiology that remains to be fully elucidated.
While the key culprit brain regions are acknowledged to lie within the cerebello-dentato-
thalamo-cortical network (commonly known as the tremor network), including the
cerebellum—believed to be the cornerstone of ET by some (Benito-León & Labiano-
Fontcuberta, 2016; Ibrahim et al., 2021)—thalamus, motor thalamus (ventro-intermediate
núcleo, Vim), and motor cortex (Hallett, 2014; Sharifi et al., 2014), the exact underlying
mechanisms remain debated. Some authors have suggested that ET may result from abnormal
neuronal oscillations within the tremor network (Deuschl & Elble, 2009), while others consider
it as a neurodegenerative disorder with progressive cell loss (Benito-León, 2014). A GABAergic
dysfunction has also been evoked (Gironell, 2014). These theories are not mutually exclusive
and can all be justified by convincing evidence, but their diversity shows that gathering a com-
plete picture of ET remains an ongoing endeavor.
Many have used magnetic resonance imaging (resonancia magnética) to try and pinpoint the alterations in
brain anatomy caused by ET. With voxel-based brain morphometry ( VBM), decreases in gray
asunto (GM) density were observed in the cerebellum, frontal and parietal cortices, and insula
(Benito-León et al., 2009). Atrophy of the cerebellum and the frontal, occipital, middle tem-
poral, and superior parietal cortices was also put forward (Bagepally et al., 2012), while other
works pinpointed atrophy of the cerebellum (Quattrone et al., 2008) and of the left temporal
pole and occipital cortex (Tuleasca et al., 2017). The use of automated segmentation labeling
Essential tremor (ET):
A brain disorder whose clinical
symptoms include tremor of the
upper limbs and subtle cognitive
deficits.
Tremor network:
A set of interconnected brain areas
believed to be involved in the onset
and progression of ET.
Brain morphometry:
The quantitative analysis of the shape
and dimensions of brain structures,
including local volume, thickness,
surface area, and curvature.
Neurociencia en red
851
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
norte
mi
norte
_
a
_
0
0
2
5
3
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
instead of VBM confirmed cerebellar atrophy in ET (Cerasa et al., 2009), while also revealing
lower thalamus, hippocampus,and ventral diencephalon volume, and larger right caudate,
pallidum, amygdala, and bilateral putamen and nucleus accumbens volume (Pietracupa
et al., 2019; Prasad et al., 2019).
At the level of cortical morphometric properties, Benito-León and colleagues reported that
more severe tremor correlated with atrophy in medial orbitofrontal and lingual cortices, el
supramarginal gyrus, paracentral lobule, and in reduced thalamus and amygdala areas
(Benito-León et al., 2019). In another study, patients who did not respond to propranolol
(a widely prescribed antitremor drug) showed diminished left orbitofrontal and right temporal
CT (cortical thickness) compared to responders (Chung et al., 2013). Serrano et al. (2017)
also found that the standard deviation of CT in the right inferior parietal and fusiform areas
plays a key role in distinguishing between ET patients and healthy controls.
For such patients with ET who do not respond to commonly prescribed medication, several
surgical options are available (Elble et al., 2018; Picillo & Fasano, 2016). While the standard
intervention remains deep-brain stimulation (Benabid et al., 1996), during the past 20 años,
minimally invasive stereotactic radiosurgery (SRS, particularly Gamma Knife, GK) of the Vim
of the thalamus has also been proofed as safe and effective, in particular for patients with
contraindication for open surgery or as a patient’s choice (Elaimy et al., 2010; Tuleasca,
Najdenovska, et al., 2018). Hasta la fecha, the impacts of such an intervention (or existing alterna-
tives) have primarily been studied through resting-state functional MRI (Jang et al., 2016; Parque
et al., 2017; Tuleasca, Regis, et al., 2018), but structural investigations at the level of anatom-
ical brain properties remain lacking.
Aquí, we focus on drug-resistant patients who underwent unilateral GK radiosurgery of the
left Vim for right dominant ET. Our primary aims were to (1) refine our understanding of ET
before thalamotomy, by comparing our cohort of ET patients in the absence of medication to
matched healthy controls (HCs), y (2) gain insight into potential brain plasticity mechanisms
triggered by GK thalamotomy, by comparing the pre- and posttherapeutic (1 year after inter-
vention) states to account for delayed clinical and radiological effects.
In order to achieve this, we quantified morphometric brain attributes. In addition to cerebel-
lar and subcortical volume, we also extracted three regional cortical morphometric features:
CT, SA (surface area) and mean curvature (MC). Several reasons motivated this multimeasure
análisis: primero, SA and MC have not yet been investigated in the context of ET. Segundo, cada
measure encodes, to an extent, unique information: por ejemplo, in healthy subjects, they yield
distinct cortical asymmetry maps (Chiarello et al., 2016). Tercero, they are genetically and phe-
notypically complementary (Sanabria-Diaz et al., 2010; Yang et al., 2016), which renders a
joint analysis interesting as their interactions may be altered in ET. Cuatro, each measure is mod-
ulated differently by environmental factors, such as urban upbringing (Besteher et al., 2017) o
maltreatment (Kelly y col., 2013); de este modo, it is reasonable to expect that ET may also exert distinct
impacts on them.
More specifically, we sought to complement previous morphometric works by leveraging
structural covariance analysis (SCA) to study cross-regional dependences. In SCA—see
Alexander-Bloch et al. (2013) and Evans (2013) for reviews—one is interested in the extent
to which a measure of interest (p.ej., CT) correlates across subjects in a pair of regions. Semejante
patterns of covariance are a characteristic attribute of the human cortex (Mechelli et al., 2005),
are genetically regulated (Morgan et al., 2019; Romero-Garcia et al., 2018; Schmitt et al.,
2008), and partly reflect structural connectivity (Yee et al., 2018). Their relevance is further
corroborated by the fact that the foci of atrophy in many brain disorders largely overlap with
Structural covariance analysis:
An analytical approach in which the
cross-regional covariance in a
morphometric measure of interest is
quantified across subjects.
Neurociencia en red
852
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
norte
mi
norte
_
a
_
0
0
2
5
3
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
covariance patterns found when using the most atrophied locations as seeds in healthy sub-
jects (Seeley et al., 2009). SCA has been a fruitful analytical approach to better comprehend
the healthy and diseased brain (Bassett y cols., 2008; Bernhardt et al., 2011; Bethlehem et al.,
2017; Chen et al., 2008; He et al., 2008; Khundrakpam et al., 2017). To our knowledge, solo
one recent study has considered SCA in the context of ET (Yang et al., 2021), in which the
authors quantified the similarity of GM profiles across regions to assess covariance and studied
a cohort of drug-naïve ET patients. With this work, we seek to instead leverage SCA to study
drug-resistant patients, and how they may improve following GK thalamotomy.
MATERIALES Y MÉTODOS
Participantes
We considered uniform structural MRI data from 34 ET patients (both prethalamotomy and
1 year later) scanned on the same 3T MR machine, y 29 HCs. All patients were right-handed
and presented with drug-resistant right-dominant tremor. All underwent left Vim thalamotomy
by GK.
The Timone University Hospital Ethical Committee (ID-RCB: 2017-A01249-44) granted for-
mal approval for this study (including by the Ethics Committee at national level, CNIL-MR-03),
and individual consent was also obtained from all subjects. Patients were neurologically eval-
uated and referred by T.W., a neurologist specialized in movement disorders. All patients had a
clear diagnosis of ET and showed no other structural abnormalities on pretherapeutic 3T MRI.
Demographic characteristics of the ET patients and HC subjects can be found in Table 1,
where it can be seen that the groups were matched for both age and gender.
Several measures were used to clinically evaluate ET patients, and their potential recovery
after thalamotomy: Activities of Daily Living from the survey designed by Bain and colleagues
(Bain et al., 1993), Tremor Score on Treated Hand (TSTH) from the Fahn-Tolosa-Marín rating
escala (Fahn et al., 1993), head tremor (Tremor Research Group Essential Tremor Rating Assess-
mento, de 0 a 3), and Quality of Life in Essential Tremor (QUEST; Tröster et al., 2005). Clinical
data is summarized in Table 1, where a significant improvement in clinical tremor scores can be
observed across all the quantified measures upon thalamotomy. In what follows, we will focus
on TSTH values before the intervention (to quantify the extent of tremor) and on the percentage
change in TSTH from before to after the intervention (to address the extent of recovery).
En tono rimbombante, SCA is not compatible with the subject-wise investigation of these scores: en efecto,
only one measure of covariance is generated per group (p.ej., posttherapeutic ET patients). De este modo,
one cannot conduct classical correlation analyses between morphometric features and clinical
puntuaciones. In order to nonetheless account for the fact that different patients recovered to distinct
extents, we devised a strategy to gauge a possible link between the extracted SC features and
tremor symptoms or postinterventional recovery (see Link to Clinical Scores section).
Imaging
The imaging data was acquired on a head-only 3T machine (SIEMENS SKYRA, Munich,
Alemania, 32-channel receive-only phase-array head coil). Native T1-weighted images were
acquired before and 1 year after thalamotomy with the following parameters: TR/TE = 2
300/2.98 EM, isotropic voxels of 1 mm3, 160 slices.
As medication was frequently ineffective, most patients no longer received treatment at the
time of thalamotomy. Scanning was performed in a drug-naïve state (drugs having been
stopped at least 3 days beforehand).
Tremor Score on Treated
Hand (TSTH):
A clinical score that quantifies the
extent of tremor in a patient, con
higher values denoting greater motor
impairment.
Neurociencia en red
853
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
norte
mi
norte
_
a
_
0
0
2
5
3
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
t
norte
mi
w
oh
r
k
norte
mi
tu
r
oh
s
C
mi
norte
C
mi
i
Variable
norte
Age (años)
Gender (METRO:F)
ADL
HEAD
QUEST
TSTH
Lesion volume (ml)
Time to tremor
arrest (días)
Symptoms duration
(años)
Mesa 1. Demographic and clinical details of the subjects
HC
29
ETpre
34
69.93 ± 7.14
[59, 69, 83]
70.06 ± 9.12
[49, 72, 83]
ETpost
34
n.a.
17:17
Drop (puntos)
n.a.
Drop (%)
n.a.
Nmissing
n.a.
P value
n.a.
17:17
29.59 ± 11.39
[13, 28.5, 49]
1 ± 0.85
[0, 1, 2]
45.46 ± 16.4
[12, 41.5, 80]
20.41 ± 5.53
[8, 20.5, 30]
n.a.
n.a.
n.a.
n.a.
6.03 ± 11.26
[0, 1, 41]
0.56 ± 0.75
[0, 0, 3]
23.16 ± 16.57
[1, 26, 57]
−23.56 ± 12.35
[−48, −24.5, 2]
82.83 ± 29.64
[0, 96.75, 100]
−0.39 ± 0.83
[−2, 0, 1]
−24.79 ± 13.21
[−47, −25, −2]
n.a.
n.a.
6.26 ± 7.71
[0, 3, 27]
−14.15 ± 6.6
[−26, −14.5, 1]
72.73 ± 29.19
[0, 86.05, 100]
0.12 ± 0.13
[0.002, 0.076, 0.6]
n.a.
n.a.
35.53 ± 18.28
[5, 33, 61]
127.56 ± 81.38
[15, 120, 300]
n.a.
n.a.
n.a.
n.a.
n.a.
n.a.
n.a.
n.a.
n.a.
0/0
0/1
8/9
0/0
n.a.
2
n.a.
t66 = −0.06,
pag = 0.95
n.a.
t66 = 8.57,
pag = 2.48 · 10
−12
t65 = 2.16,
pag = 0.035
t43 = 15.37,
pag = 4.47 · 10
−19
t66 = 8.69,
pag = 1.52 · 10
−12
n.a.
n.a.
n.a.
12:17
n.a.
n.a.
n.a.
n.a.
n.a.
n.a.
n.a.
Nota. For healthy controls (HCs), patients before (ETpre) and after thalamotomy (ETpost), values are reported as mean ± standard deviation, with minimum, median, and maximum into
squared brackets. Some clinical scores could not be collected in a few occasions (Nmissing), in which case the associated subjects were excluded from statistical computations. Significant
statistical comparisons are highlighted in bold. METRO, masculino; F, femenino.
8
5
4
METRO
oh
r
pag
h
oh
metro
mi
t
r
y
i
norte
mi
s
s
mi
norte
t
i
a
yo
t
r
mi
metro
oh
r
a
norte
d
r
mi
C
oh
v
mi
r
y
a
F
t
mi
r
t
h
a
yo
a
metro
oh
t
oh
metro
y
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
norte
mi
norte
_
a
_
0
0
2
5
3
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
FreeSurfer:
An open source brain imaging
package enabling the extraction of
morphometric estimates from
structural neuroimaging data.
Stereotactic Radiosurgical Procedure
GK thalamotomy was performed using Leksell Gamma Knife (Elekta Instruments, AB, Suecia)
between September 2014 and April 2016, always at the Centre Hospitalier Universitaire de la
Timone in Marseille, by the same neurosurgeon ( J.R.). To avoid artifacts, diffusion tensor
imaging data was first acquired without the frame, and then co-registered with the therapeutic
stereotactic images. The Leksell coordinate G frame (Elekta Instruments, AB, Suecia) era
always applied under local anesthesia on the day of the GK thalamotomy. After positioning
the frame, patients underwent both stereotactic CT and MRI.
Landmarks of interest, including the anterior and posterior commissures, were identified on
an MR scan (particularly on T2 CISS/FIESTA sequence, Siemens). Uniform indirect targeting
was performed using the Guiot diagram (Tuleasca, Regis, et al., 2018), placed 2.5 mm above
the anterior-posterior commissure line, y 11 mm lateral to the wall of the third ventricle. A
single 4-mm isocenter was always used, and a maximum prescription dose of 130 Gy at the
100% isodose line was uniformly prescribed (Tuleasca, Regis, et al., 2018).
Computation of Morphometric Properties
The FreeSurfer software (pescado, 2012) was used to extract three morphometric measures of
interest from structural MR images for a set of Pcort = 68 cortical regions: CT, SA, and MC.
We selected these because they have been widely used and acknowledged as relevant indi-
cators of brain geometry; ver, Por ejemplo, Harnett et al. (2020), Raznahan et al. (2011), y
Yang y otros. (2013). We hoped to find possibly distinct relationships with ET and, in addition,
considered to also explicitly quantify the interplay across properties (see Cross-Measure Anal-
ysis section).
Brevemente, after linear registration to MNI space and bias field removal, the image at hand is
skull-stripped (Ségonne et al., 2004), and voxels are classified as belonging to white matter or
to another tissue category based on their intensity and direct neighborhood. Hemispheres are
separated, the cerebellum and subcortex are removed, and the interface between the white
and gray matter is located. From there, the pial surface is also tiled, and local estimates of
CT, SA, and MC can be extracted (pescado & Valle, 2000). Further details about these steps of
the methodology can be found in Dale et al. (1999) and Fischl et al. (1999). Entonces, local voxel-
wise measurements are converted into Pcort regional values per morphometric measure, usando
the Desikan-Killiany atlas (Desikan et al., 2006).
In addition to the Pcort = 68 cortical brain regions, we also extracted regional volume for
Pnoncort = 19 noncortical areas, including the cerebellum and subcortical nuclei. Secundario
Information Table S1 summarizes the considered regions.
Of note, noncortical measures quantify regional volume instead of the three morphometric
properties at hand. Sin embargo, because we only assess cross-regional correlations, the distinct
ranges of values do not impact our analyses. De este modo, similarly for CT, SA, and MC, the Pnoncort
noncortical volume values were appended to the Pcort cortical estimates, resulting in a
P-dimensional vector for each subject, with P = 87. Note that there is no redundancy in our
analiza, since the correlational relationships between noncortical volume and a given mor-
phometric measure may differ from those with another.
These measurements were eventually linearly regressed out for age, género, and total gray
matter volume, separately within each group. The obtained residuals were used for all subse-
quent analyses.
Neurociencia en red
855
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
norte
mi
norte
_
a
_
0
0
2
5
3
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
Edge-Wise Analysis
The process described below was identically conducted for each morphometric measure of
interés. In total, there are P(P − 1)/2 = 3,741 cross-regional edges available. Separately for
the HC, prethalamotomy (abbreviated ETpre from there onwards) and postthalamotomy (abbre-
viated ETpost) datos, Pearson’s correlation coefficient R was computed for a given edge. A
positive/negative value means that when the measure in the first region is larger in one subject,
it tends to be larger/lower in the second region. The coefficient of determination R2 denotes
the associated percentage of explained variance.
We first attempted to exclude the edges that only reflect noise (es decir., for which the explained
variance is minimal). Noisy edges were defined as those for which, across the HC, ETpre, y
ETpost cases, the coefficient of determination was always lower than 0.2 (es decir., less than 20%
explained variance). Formalmente, this amounts to R2 < 0.2 or |R| < 0.4472. We selected this
threshold value as it is regarded as a moderate-to-high correlation across scientific subfields
(Akoglu, 2018). In the specific context of SCA, recent work has also considered lower/larger
values as low/high correlations and shown that fluctuations due to acquisition or preprocess-
ing settings span a lower range of R = 0.01 to 0.1 (Carmon et al., 2020).
Our thresholding step excluded 2,206 edges for CT (amounting to 41.03% of edges kept),
2,990 for SA (20.07%) and 1,408 for MC (62.36%). Note that in line with the above cri-
terion, when interpreting our results, we also consider a structural covariance negligible if
R2 < 0.2.
On the remaining edges, to assess the differences between HC subjects and ET patients, we
computed the difference HC-ETpre for each edge. Similarly, to investigate the effects of thala-
motomy, we computed the difference ETpost-ETpre.
For statistical assessment, these differences must be compared to an appropriate null distri-
bution. To do so, we resorted to nonparametric permutation testing, by recomputing covari-
ance relationships after having randomly shuffled subjects across groups. In total, 300,000 null
realizations were generated in each case, and false discovery rate (FDR)-corrected p values
were obtained and analyzed.
Link to Clinical Scores
SC estimates are obtained on full populations (e.g., the whole ETpre set of subjects) and thus do
not enable direct correlational analysis with the severity of clinical symptoms or the extent of
recovery upon intervention. It is thus impossible, with these results alone, to know whether the
most impaired subjects (or the ones who recovered the most) more strongly drove a given
group difference.
In order to circumvent this limitation, we devised an analytical strategy in which all the
significant group differences are recomputed upon the exclusion of a selected number of data
points ranging from 1 to 10. For the HC-ETpre case, we excluded either the least impaired sub-
jects in terms of baseline TSTH (yielding an estimate SC1(r ), with r the number of removed
subjects), or the most impaired ones (SC2(r )). If the most impaired subjects cause the HC-ETpre
group difference, we would expect the first case to only slightly alter the results; conversely,
the second case should yield a greater extent of change.
We computed the difference |SC2(r )| − |SC1(r )|, and took its average μSC, and the regression
coefficient of a linear model βSC (y = βSC r + c, with y = |SC2(r )| − |SC1(r )|, r the number of
subjects removed and c as constant) as summarizing metrics. A positive μSC value means that
on the whole, the assessed group difference is larger when focusing on the most impaired as
Network Neuroscience
856
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
opposed to the least impaired subjects. A positive βSC value means that as more subjects are
removed in both cases, the group difference increases. We considered the connections for
which both quantities were positive valued as showing an association with the extent of
tremor, as quantified by baseline TSTH.
The same strategy was also applied to the ETpost-ETpre case, where the percentage of TSTH
improvement was used as clinical score instead. In this case, positive-valued μSC and βSC mean
that a focus on the subjects that recover the most following the intervention (by excluding
the 1 to 10 worst recoverers), as opposed to those that recover the least (by excluding the
1 to 10 best recoverers), magnifies the group difference. Such connections were considered
as showing an association with the extent of postinterventional recovery, as quantified by the
percentage of TSTH improvement.
Cross-Measure Analysis
We were also interested in studying the covariance across structural properties within a region
(for example, if an area is thicker in a subject, does it also tend to exhibit a greater surface
area?). For this purpose, for each pair of morphometric measures and each cortical brain
region, we computed the covariance across subjects. We then submitted the outputs to the
same statistical pipeline as above for both the HC-ETpre and the ETpost-ETpre cases to yield
FDR-corrected p values.
Note that in the case of this analysis, we only perform Pcort = 68 assessments (one per
cortical area); as we do not consider cross-regional (but rather cross-measure) relationships,
the thresholding strategy described in the Edge-Wise Analysis section is not required.
Availability of the Data and Scripts
All the analytical steps described above were performed with custom scripts and
MATLAB2014b (MathWorks, Natick, USA). Color maps for plotting were generated with the
cbrewer toolbox (https://www.mathworks.com/matlabcentral/fileexchange/34087-cbrewer
-colorbrewer-schemes-for-matlab). All the scripts used in this work are freely available at
https://github.com/TiBiUan/SCA_EdgeWise.git.
Data sharing is not applicable to this article as no new data were created or analyzed in
this study.
RESULTS
Edge-Wise Analysis
Structural covariance matrices and SC value distributions for all three groups, as well as asso-
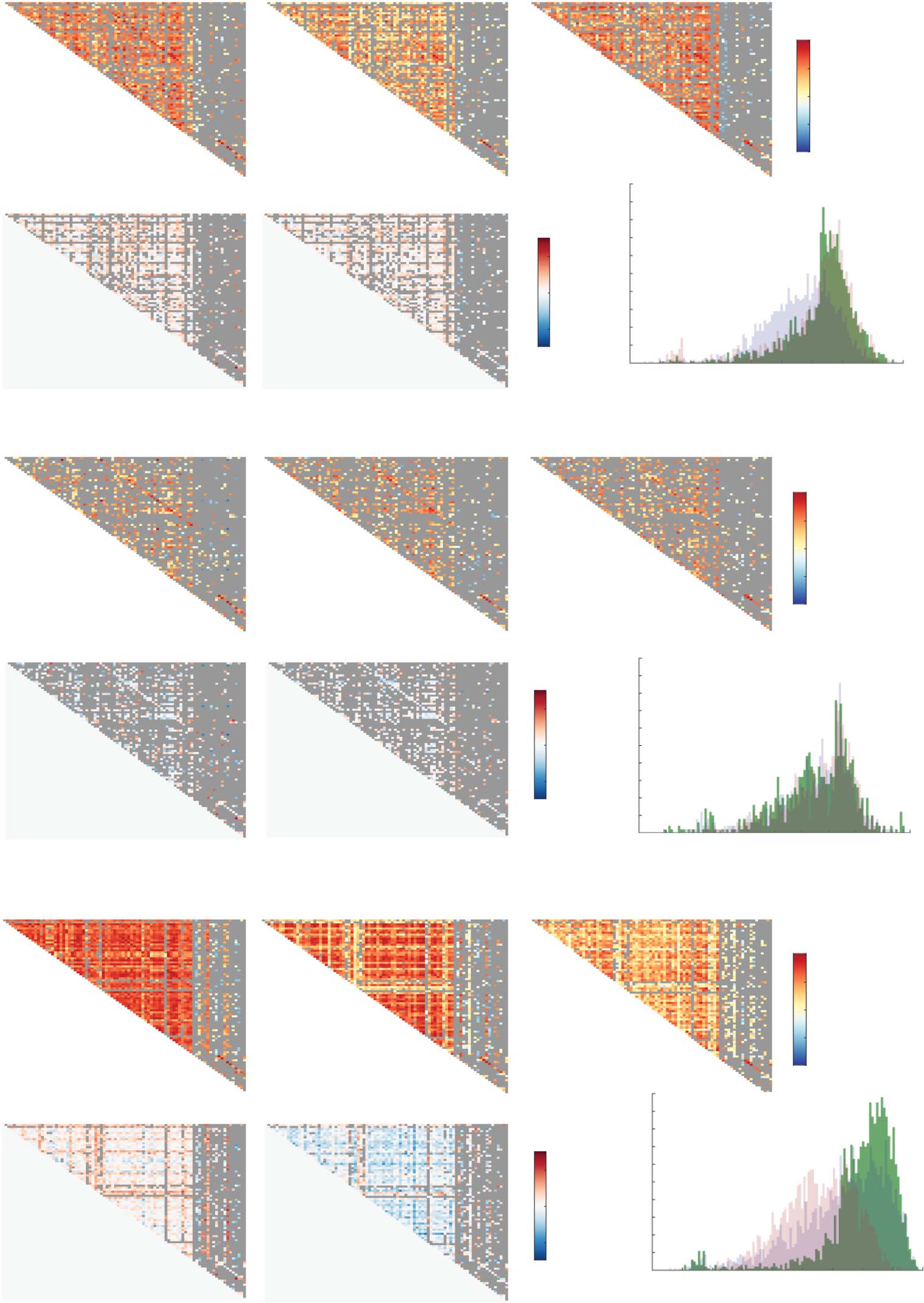
ciated HC-ETpre and ETpost-ETpre group differences, are shown in Figure 1. For CT (Figure 1A),
structural covariance was overall lower in ETpre subjects compared to the other two groups, as
reflected by more SC values between −0.2 and 0.4 in the histogram. A few connections from
the ETpost group also showcased particularly negative structural covariance (between −0.6 and
−0.4). This was quantitatively confirmed by respective average HC, ETpre, and ETpost SC values
of 0.25 ± 0.28, 0.15 ± 0.26, and 0.19 ± 0.32, and significant associated ETpre-HC ( Wilcoxon
rank sum test’s z = −15.56, p < 0.001), ETpost-ETpre (z = 4.8, p < 0.001) and ETpost-HC (z =
−8.61, p < 0.001) group differences.
The one significant edge-wise SC difference between the ETpre and ETpost groups involved
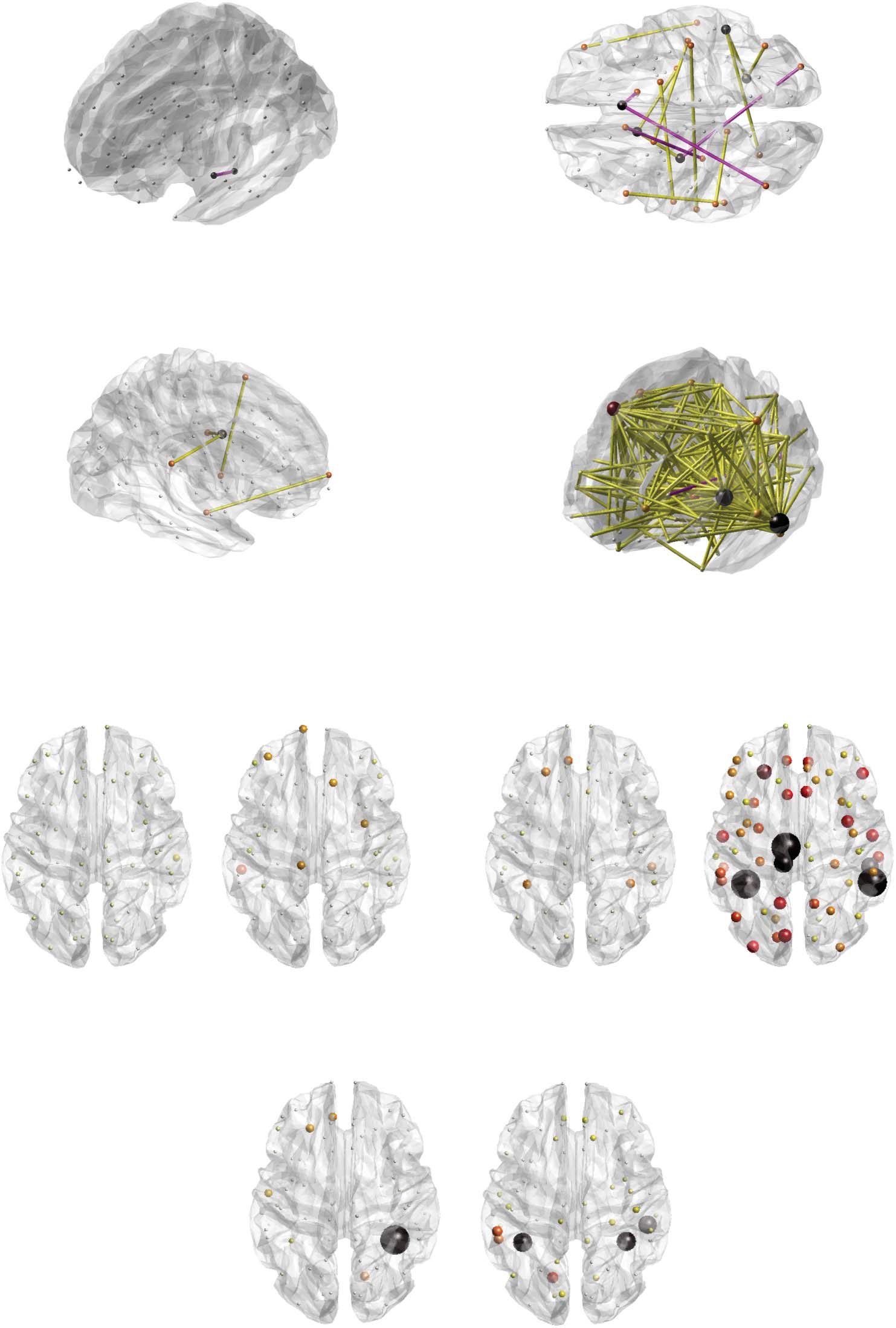
the left parahippocampal gyrus and the left hippocampus (Figure 2A). Structural covariance
was negligible at baseline (SCpre = 0.25), and became negative valued following intervention
Network Neuroscience
857
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural covariance patterns across groups and modalities. For cortical thickness (A), surface area (B), and mean curvature (C),
Figure 1.
structural covariance matrices for the HC, ETpre, and ETpost groups (top row, from left to right), associated HC-ETpre and ETpost-ETpre group differ-
ences (bottom row, left and middle panels), and summarizing histogram (bottom row, right panel). Discarded connections are depicted in gray.
(SCpost = −0.65, ΔSC = −0.91). This group difference was also related to the extent of tremor
recovery (βSC = 0.0069, μSC = 0.024).
For SA (Figure 1B), no difference across groups transpired at the global level. Fifteen con-
nections reached significance between the HC and ETpre groups (as summarized in Supporting
Network Neuroscience
858
Morphometry in essential tremor and recovery after thalamotomy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Graphical overview of significant findings. Significant structural covariance edges for cortical thickness and the ETpost-ETpre contrast
(A), surface area and the HC-ETpre contrast (B), and mean curvature and the ETpost-ETpre (C), and HC-ETpre (D) contrasts. Positive-valued group
differences are represented by yellow connections, and negative-values ones by pink connections. Nodal size and color are proportional to the
number of emanating significant connections. For the cortical thickness/surface area (E), cortical thickness/mean curvature (F), and surface
area/mean curvature (G) cross-measure analyses, overview of significance for the HC-ETpre (left) and ETpost-ETpre (right) group differences.
Nodal size and color are proportional to −log( p value), with black nodes the ones found significant in our analyses. P, posterior; R, right;
A, anterior; L, left.
Network Neuroscience
859
Morphometry in essential tremor and recovery after thalamotomy
Information Table S2 and displayed in Figure 2B), seven of which were related to the extent of
tremor (βSC, μSC > 0). SC between the left supramarginal and left isthmus cingulate cortices,
between the right pars orbitalis and right middle temporal cortex, and between the right post-
central gyrus and left middle temporal cortex was strongly positive valued in the HC group but
close to zero in patients with ET. SC between the right caudate and right caudal anterior cingu-
late cortex was largely negative in HCs, but close to zero in the ETpre group. SC between the left
parahippocampal gyrus and left caudal anterior cingulate cortex, and between the right caudal
anterior cingulate and the left inferior parietal cortices was conversely only present (and positive
valued) in patients with ET. Finalmente, SC between the left pars opercularis and the left Banks supe-
rior temporal sulcus showed opposite sign in the HC (positivo) and ETpre (negative) grupos.
For MC (Figura 1C), SC was consistently greater in magnitude in HC subjects compared to
patients with ET both before and after the intervention (mean SC of 0.41 ± 0.35, 0.3 ± 0.35,
y 0.22 ± 0.24, respectivamente; ETpre-HC z = −14.15, pag < 0.001; ETpost-HC z = −28.5107, p <
0.001). In addition, the ETpost group also exhibited overall lower SC values compared to the
ETpre one (ETpost-ETpre z = −11.2123, p < 0.001).
This resulted in four connections reaching significance for the ETpost-ETpre contrast (Figure 2C),
between the left insula and left superior temporal cortex (SCpre = 0.19, SCpost = 0.77, ΔSC = 0.58,
p = 0.023, βSC = 0.0002, μSC = −0.0062), right supramarginal gyrus and left insula (SCpre = 0.13,
SCpost = 0.86, ΔSC = 0.73, p = 0.043, βSC = −0.003, μSC = −0.0055), left amygdala and left
caudal middle frontal cortex (SCpre = −0.33, SCpost = 0.57, ΔSC = 0.9, p = 0.023, βSC =
0.0032, μSC = −0.011), and right hippocampus and left frontal pole (SCpre = −0.38, SCpost =
0.51, ΔSC = 0.89, p = 0.043, βSC = −0.025, μSC = −0.12). In all cases, structural covariance
became significant following the intervention, but there was no association with the extent of
tremor recovery.
In the HC-ETpre case, there was a broad pattern of 195 significant connections, summarized
in Supporting Information Table S3 and Figure 2D. Fifty-three of them showed an association
with the extent of tremor, with the most implicated regions including the left inferior temporal
cortex (showcased five times), left middle temporal cortex (17) and right caudal middle frontal
cortex (11).
Cross-Modality Analysis
The investigation of cross-modality relationships revealed, for the interactions between CT and
SA, respective average covariance values across regions of −0.37 ± 0.25, −0.3 ± 0.2, and −0.4 ±
0.2 in the HC, ETpre, and ETpost groups. The ETpost-ETpre group difference was significant
( Wilcoxon rank sum test’s z = −2.81, p = 0.045), but no individual regional interaction sur-
vived multiple testing correction.
For the interplay between CT and MC, covariance was on the whole negligible in the HC
(−0.1 ± 0.27) and ETpre (0.03 ± 0.26) groups, but became negative valued following the inter-
vention (−0.42 ± 0.2). This was confirmed by significant ETpost-HC (z = −6.59, p < 0.001) and
ETpost-ETpre (z = −8.14, p < 0.001) group differences. Five regions reached significance for the
ETpost-ETpre contrast (Figure 3A and Figure 2F): the left fusiform gyrus (Cpre = 0.44, Cpost =
−0.68, ΔC = −1.12, p = 0), the left paracentral gyrus (Cpre = −0.075, Cpost = −0.68, ΔC =
−0.61, p = 0.0034), the left posterior cingulate cortex (Cpre = 0.17, Cpost = −0.69, ΔC = −0.86,
p = 0), the right Banks superior temporal sulcus (Cpre = 0.24, Cpost = −0.67, ΔC = −0.91, p = 0),
and the right inferior temporal cortex (Cpre = 0.11, Cpost = −0.65, ΔC = −0.76, p = 0.0054). In
all cases, the cross-modality covariance expectedly became strongly negative valued following
the intervention.
Network Neuroscience
860
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Cross-property covariance relationships. For the relationship between cortical thickness and mean curvature for the ETpost-ETpre
contrast (A), and for the relationship between surface area and mean curvature for the HC-ETpre (B), and ETpost-ETpre (C) group differences,
cross-property covariance differences between groups (black squares) and associated null distributions (box plots) for all 68 cortical regions
from the Desikan-Killiany atlas. Color coding of the box plots denotes brain lobes. Significant cases are highlighted by a light gray box in the
background.
When probing the relationships between SA and MC, cross-measure covariance was mildly
positive in the HC group (0.18 ± 0.22), close to zero in the ETpre group (0.06 ± 0.24), and
increased after the intervention (0.25 ± 0.21). Accordingly, significant group differences were
found in the HC-ETpre (z = 3.04, p = 0.022) and ETpost-ETpre (z = 4.49, p < 0.001) cases
Network Neuroscience
861
Morphometry in essential tremor and recovery after thalamotomy
(Figure 2G). At the level of individual regions, for the HC-ETpre contrast (Figure 3B), the right
fusiform gyrus showed a positive covariance between modalities in HC subjects, but only a
mild negative one in patients with ET (CHC = 0.66, Cpre = −0.34, ΔC = 0.99, p = 0). For the
ETpost-ETpre contrast (Figure 3C), there was a transition toward positive-valued covariance
upon intervention in the left fusiform gyrus (Cpre = −0.33, Cpost = 0.44, ΔC = 0.77, p =
0.02), the right fusiform gyrus (Cpre = −0.34, Cpost = 0.55, ΔC = 0.88, p = 0.02), and the right
inferior temporal cortex (Cpre = −0.18, Cpost = 0.57, ΔC = 0.75, p = 0.027).
DISCUSSION
Motor and Nonmotor Characteristics of ET Are Captured by SCA
We examined structural covariance patterns in patients with ET before and after thalamotomy,
as well as preintervention ET cases compared to age-matched HCs. In doing so, we considered
three popular and complementary morphometric properties (Harnett et al., 2020; Lerch et al.,
2006; Raznahan et al., 2011; Sanabria-Diaz et al., 2010; Yang et al., 2013; Yang et al., 2016):
CT, SA, and MC.
At the whole-brain level, we observed very different structural covariance patterns across
them: for CT, SC was overall lower in ETpre subjects compared to HCs. It then reverted follow-
ing the intervention, denoting large-scale recovery. This did, however, not translate into sig-
nificant individual SC edges (likely owing to the multiple testing approach and too small
changes to be captured with our statistical power), as only the structural covariance between
the left hippocampus and left parahippocampal cortex was found to differ between the ETpre
and ETpost groups, in a way that was also associated with the extent of tremor recovery. The
appearance of a negative-valued SC, while it was strongly positive in the HC group (SCHC =
0.5406), hints at an alternative mechanism of tremor recovery in which the geometry of both
structures becomes jointly regulated (i.e., if one is thicker/larger in a subject, then the other
tends to be thinner/smaller).
For SA, there was no clear difference in whole-brain SC patterns across groups. However,
spatially subtle differences could be captured upon edge-wise analysis between the HC and
ETpre groups. In some cases, structural covariance disappeared in patients with ET, while in
others it was only seen then. The supramarginal and postcentral cortices, but also the caudal
anterior cingulate cortex, at the interface between frontal executive and parietal sensorimotor
systems (Taylor et al., 2007), and the caudate, canonically linked to movement planning
(Villablanca, 2010), were all involved areas, highlighting that ET induces complex structural
covariance rearrangements within a broad network of motion-related brain areas.
The pattern observed for MC was particularly intriguing, as SC was lower in ETpre subjects
compared to HCs, and then continued to globally decrease following the intervention. Thus,
the observed MC changes could relate to aspects of ET that are not purely motor, such as asso-
ciated cognitive deficits and concomitant depression. Lending support to this theory, at the
level of the ETpost-ETpre group differences, the four unraveled connections did not show an
association to the extent of motor recovery, and involved frontal brain centers, implicated
in executive functions (Braun et al., 2015; Stuss, 2011), as well as the amygdala and the insula,
respectively most well-known for their role in emotional processes (Phelps, 2006) and salience
monitoring (Menon & Uddin, 2010). In previous studies, ET patients with depression showed
larger superior frontal nodal efficiency (Li et al., 2021), and regional homogeneity was
increased in the middle prefrontal cortex (Duan et al., 2021), demonstrating that frontal alter-
ations should be expected in ET patients with depression. While it could thus be that our
862
Nodal efficiency:
A graph theoretical measure that
quantifies how fast information can
spread from a given node to all
others in the network.
Regional homogeneity:
A metric that quantifies the temporal
coherence between a set of a given
voxel’s spatial neighbors.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
results relate to nonmotor characteristics of ET, the lack of dedicated ratings in our data pre-
cludes further investigation at this stage.
To summarize, our edge-wise analysis enabled us to pinpoint a set of structural covariance
relationships altered upon the presence of ET, or that have been modified following Vim thal-
amotomy, across a set of three morphometric properties. Many of the implicated areas relate
to movement planning or execution, as could be expected in the context of ET; in these cases,
we also observed an association with the extent of motor symptoms (for the HC-ETpre compar-
ison) or motor recovery (for the ETpost-ETpre one). Furthermore, some of the unraveled connec-
tions also involved regions modulating nonmotor functions, such as emotional control or
attention. As no association to motor symptoms was found in these cases, these features are
likely indicative of nonmotor attributes of ET.
Evidence for a Role of Visual Cortical Areas in ET
In addition to the above observations, our results also converged on a picture in which brain
areas linked to visual functions could be related to the occurrence of ET, or to plasticity fol-
lowing Vim thalamotomy. Recall that for CT, the only significant edge (in the ETpost-ETpre case)
linked the left parahippocampal gyrus and hippocampus; the parahippocampal gyrus has been
implicated in episodic memory, but also in visuospatial processing (Aminoff et al., 2013),
while the hippocampus relates to memory, spatial navigation, and cognition (Lisman et al.,
2017). Fittingly, a previous VBM study of ours also related the left parahippocampal place area
to TSTH improvement following intervention (Tuleasca et al., 2017).
For MC, between the HC and ETpre groups, the most involved area in the 53 significant
connections showing an association with tremor severity was the left middle temporal cortex,
which includes the famous area MT that encodes the perception of movement within the
visual field (Born & Bradley, 2005). The right caudal middle frontal cortex was the second most
implicated region, and may relate to alterations at the level of the frontal eye fields, which
control eye movements (Robinson & Fuchs, 1969) and contribute to visual selection (Muggleton
et al., 2003).
Our second analysis, in which we considered the interplay across morphometric properties,
further corroborates the importance of visual areas in ET. Only one feature contrasted HC and
ETpre subjects: the covariance between SA and MC in the right fusiform gyrus, an area involved
in higher order visual functions (Weiner & Zilles, 2016). The strong positive-valued covariance
across modalities seen in HCs disappeared in patients with ET, and was accompanied by
broadly lower cross-property covariance in the diseased group.
Interestingly, this diffuse weakening renormalized following the intervention, and interac-
tions between the other pairs of properties also changed: CT and SA regained stronger anti-
correlation akin to that seen in HCs, and there was a marked appearance of negative-valued
CT/MC covariance. Thus, in addition to the renormalization of CT/SA and SA/MC interactions,
the development of a more antagonistic relationship between CT and MC appears to be an
alternative mechanism of recovery.
At the regional level, the paracentral gyrus, the right inferior temporal cortex—important for
visual perception (Miyashita, 1993)—and the bilateral fusiform gyrus were involved in these
changes following stereotactic radiosurgical thalamotomy. Such findings are supported by a
previous resting-state functional MRI work from our group, in which a component reminiscent
of a salience network showed altered interconnectivity with the right fusiform gyrus and mid-
dle temporal visual area (Tuleasca, Regis, et al., 2018). Further highlighting the importance of
863
Stereotactic radiosurgical
thalamotomy:
A surgical intervention in which the
thalamus is locally lesioned with
ionizing radiation to alleviate tremor
in drug-resistant patients.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
visual regions in postinterventional tremor recovery, another recent study on Parkinson’s dis-
ease patients evidenced a correlation between tremor improvement following MRI-guided
focused ultrasound thalamotomy and signal intensity in the left occipital cortex (Xiong
et al., 2021).
Taken together, these results suggest that cross-regional or cross-property dependences
involving brain regions mediating high-level visual functions are tied to ET. This extends a past
morphometric report in which a direct relationship between posttherapeutic improvement in
tremor of the treated hand and pretherapeutic GM density of the right visual association area
was evidenced (Tuleasca, Witjas, et al., 2018). It also raises the question of potentially target-
ing visual networks in the near future.
Limitations and Future Perspectives
It is important to acknowledge the limitations of our study: in particular, the number of subjects
at our disposal remains quite low, likely precluding the unraveling of finer ET-related structural
covariance patterns. However, it should be remembered that the patients analyzed therein
belong to a subgroup of drug-resistant individuals only; in addition, morphometric data could
be collected at two time points before and 1 year after thalamotomy. Of course, it would be
interesting to monitor how recovery continues to evolve over the course of time, with an even
more longitudinal design. One might hypothesize that additional structural covariance adjust-
ments would then start developing, although empirically, clinical and radiological changes
remain minimal.
Another potential limitation is the dependence of our results on the parcellation that was
used for the analyses; indeed, a finer-grained atlas could perhaps provide yet more spatially
accurate results. However, the number of statistical tests conducted in parallel would then also
dramatically increase, and so would the extent of dependences between spatially neighboring
areas. More advanced statistical correction methods would then have to be deployed for
appropriate analysis; see, for example, Meskaldji et al. (2015).
In addition, care should be taken in the mechanistic interpretation of structural covariance
findings: positive-valued edges are only sometimes paralleled by physical wiring (perhaps due
to the impact of mutually trophic effects), while factors such as the (anti)coherence of neural
activity (leading to similar or opposite plastic changes depending on the sign of the interaction)
may be at play in other cases (Gong et al., 2012).
Covariance between different morphometric properties should also be contemplated with
caution; indeed, the interactions across measures are largely reshaped throughout the lifespan.
For example, while CT and SA evolve with different timings and characteristic regional pat-
terns (Raznahan et al., 2011; Wierenga et al., 2014), they do so in a tightly intertwined and
nontrivial manner (Schnack et al., 2015). While our morphometric estimates were regressed
out for age and only spanned a relatively narrow age range, further nonlinear effects could still
have remained present in the data.
Our cross-measure results on HCs were in line with previous research: cortical gyrification
has been positively/negatively associated with SA/CT, and a negative relationship between CT
and SA has also been evidenced throughout the lifespan, including in older adults as assessed
here (Hogstrom et al., 2013). Mechanistically speaking, a larger SA is believed to facilitate
resource allocation, computational capacity, and functional specificity, while cortical thinning
may be associated to pruning and the potentiation of information flow within cortical columns
(Tadayon et al., 2020). We observed this antagonism to be restored to a level comparable to
Network Neuroscience
864
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
HCs in patients with ET following intervention, and the same held true for the positive rela-
tionship between SA and MC. The negative relationship between CT and MC was, however,
strengthened much beyond its level in HCs. Cortical thinning and concomitant greater gyral
complexity have been observed in anxiety disorder and 22q11.2 deletion syndrome (Bearden
et al., 2009; Molent et al., 2018); in future work, it will be interesting to determine what are
the neurobiological mechanisms at play, and whether this antagonistic coupling in ET may
relate to subtle anxiety- or cognition-related deficits known to occur in the disease (Bermejo-
Pareja, 2011).
Methodologically speaking, it will also be interesting to translate the proposed analytical
approach to the network-level spatial scale: to achieve this, instead of focusing on individual
cross-regional edges, graph theoretical metrics (Rubinov & Sporns, 2010) could be computed
from whole-brain structural covariance patterns and compared between groups. Given the fact
that ET is widely regarded as a network-level disorder (Raethjen & Deuschl, 2012), such an
analysis can be expected to further illuminate our understanding of the disease, and of brain
plasticity following intervention.
ACKNOWLEDGMENTS
Dr. Tuleasca gratefully acknowledges the receipt of a Young Researcher in Clinical Research
Grant (“Jeune Chercheur en Recherche Clinique”) from the University of Lausanne (UNIL),
Faculty of Biology and Medicine (FBM), and the Lausanne University Hospital (CHUV). The
authors also wish to thank Andrea Sullca for her help in compiling the presented results.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00253.
AUTHOR CONTRIBUTIONS
Thomas Bolton: Formal analysis; Investigation; Methodology; Software; Validation; Visualization;
Writing – original draft; Writing – review & editing. Dimitri Van De Ville: Supervision; Writing –
review & editing. Jean Régis: Conceptualization; Data curation; Funding acquisition; Supervision.
Tatiana Witjas: Data curation; Project administration. Nadine Girard: Data curation; Project
administration. Marc Levivier: Conceptualization; Data curation; Funding acquisition; Project
administration; Resources; Supervision. Constantin Tuleasca: Conceptualization; Data curation;
Funding acquisition; Investigation; Resources; Supervision; Validation; Writing – review & editing.
FUNDING INFORMATION
Constantin Tuleasca, Université de Lausanne (https://dx.doi.org/10.13039/501100006390).
REFERENCES
Akoglu, H. (2018). User’s guide to correlation coefficients. Turkish
Journal of Emergency Medicine, 18(3), 91–93. https://doi.org/10
.1016/j.tjem.2018.08.001, PubMed: 30191186
Alexander-Bloch, A., Giedd, J. N., & Bullmore, E. (2013). Imaging
structural co-variance between human brain regions. Nature
Reviews Neuroscience, 14(5), 322–336. https://doi.org/10.1038
/nrn3465, PubMed: 23531697
Aminoff, E. M., Kveraga, K., & Bar, M. (2013). The role of the para-
hippocampal cortex in cognition. Trends in Cognitive Sciences,
17(8), 379–390. https://doi.org/10.1016/j.tics.2013.06.009,
PubMed: 23850264
Bagepally, B. S., Bhatt, M. D., Chandran, V., et al. (2012). Decrease
in cerebral and cerebellar gray matter in essential tremor: A
voxel-based morphometric analysis under 3T MRI. Journal of
Network Neuroscience
865
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
NeuroImaging, 22(3), 275–278. https://doi.org/10.1111/j.1552
-6569.2011.00598.x, PubMed: 21447032
Bain, P. G., Findley, L. J., Atchison, P., et al. (1993). Assessing
tremor severity. Journal of Neurology, Neurosurgery & Psychiatry,
56(8), 868–873. https://doi.org/10.1136/jnnp.56.8.868, PubMed:
8350102
Bassett, D. S., Bullmore, E., Verchinski, B. A., et al. (2008). Hierarchi-
cal organization of human cortical networks in health and schizo-
phrenia. Journal of Neuroscience, 28(37), 9239–9248. https://doi
.org/10.1523/JNEUROSCI.1929-08.2008, PubMed: 18784304
Bearden, C. E., Van Erp, T. G. M., Dutton, R. A., et al. (2009). Alter-
ations in midline cortical thickness and gyrification patterns
mapped in children with 22q11.2 deletions. Cerebral Cortex,
19(1), 115–126. https://doi.org/10.1093/cercor/ bhn064,
PubMed: 18483006
Benabid, A. L., Pollak, P., Gao, D., et al. (1996). Chronic electrical
stimulation of the ventralis intermedius nucleus of the thalamus
as a treatment of movement disorders. Journal of Neurosurgery,
84(2), 203–214. https://doi.org/10.3171/jns.1996.84.2.0203,
PubMed: 8592222
Benito-León, J. (2014). Essential tremor: A neurodegenerative disease?
Tremor and Other Hyperkinetic Movements, 4, 252. https://doi
.org/10.5334/tohm.227, PubMed: 25120943
Benito-León, J., Alvarez-Linera, J., Hernández-Tamames, J. A., et al.
(2009). Brain structural changes in essential tremor: Voxel-based
morphometry at 3-Tesla. Journal of the Neurological Sciences,
287(1–2), 138–142. https://doi.org/10.1016/j.jns.2009.08.037,
PubMed: 19717167
Benito-León, J., & Labiano-Fontcuberta, A. (2016). Linking essential
tremor to the cerebellum: Clinical evidence. The Cerebellum, 15(3),
253–262. https://doi.org/10.1007/s12311-015-0741-1, PubMed:
26521074
Benito-León, J., Serrano, J. I., Louis, E. D., et al. (2019). Essential
tremor severity and anatomical changes in brain areas control-
ling movement sequencing. Annals of Clinical and Translational
Neurology, 6(1), 83–97. https://doi.org/10.1002/acn3.681,
PubMed: 30656186
Bermejo-Pareja, F. (2011). Essential tremor—A neurodegenerative
disorder associated with cognitive defects? Nature Reviews
Neurology, 7(5), 273–282. https://doi.org/10.1038/nrneurol
.2011.44, PubMed: 21487422
Bernhardt, B. C., Chen, Z., He, Y., et al. (2011). Graph-theoretical
analysis reveals disrupted small-world organization of cortical
thickness correlation networks in temporal lobe epilepsy. Cere-
bral Cortex, 21(9), 2147–2157. https://doi.org/10.1093/cercor
/bhq291, PubMed: 21330467
Besteher, B., Gaser, C., Spalthoff, R., et al. (2017). Associations
between urban upbringing and cortical thickness and gyrifica-
tion. Journal of Psychiatric Research, 95, 114–120. https://doi
.org/10.1016/j.jpsychires.2017.08.012, PubMed: 28841461
Bethlehem, R. A., Romero-Garcia, R., Mak, E., et al. (2017). Struc-
tural covariance networks in children with autism or ADHD.
Cerebral Cortex, 27(8), 4267–4276. https://doi.org/10.1093
/cercor/bhx135, PubMed: 28633299
Born, R. T., & Bradley, D. C. (2005). Structure and function of visual
area MT. Annual Review of Neuroscience, 28, 157–189. https://
doi.org/10.1146/annurev.neuro.26.041002.131052, PubMed:
16022593
Braun, U., Schäfer, A., Walter, H. et al. (2015). Dynamic reconfig-
uration of frontal brain networks during executive cognition in
humans. Proceedings of the National Academy of Sciences,
11 2 ( 3 7 ) , 11 6 7 8 – 11 6 8 3 . h t t p s : / / d o i . o rg / 1 0 . 1 0 7 3 / p n a s
.1422487112, PubMed: 26324898
Carmon, J., Heege, J., Necus, J. H., et al. (2020). Reliability and
comparability of human brain structural covariance networks.
N e u ro I m a g e , 2 2 0 , 11 7 1 0 4 . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 6 / j
.neuroimage.2020.117104, PubMed: 32621973
Cerasa, A., Messina, D., Nicolleti, G., et al. (2009). Cerebellar atro-
phy in essential tremor using an automated segmentation method.
American Journal of Neuroradiology, 30(6), 1240–1243. https://
doi.org/10.3174/ajnr.A1544, PubMed: 19342539
Chandran, V., & Pal, P. K. (2012). Essential tremor: Beyond the motor
features. Parkinsonism & Related Disorders, 18(5), 407–413.
https://doi.org/10.1016/j.parkreldis.2011.12.003, PubMed:
22217558
Chen, Z. J., He, Y., Rosa-Neto, P., et al. (2008). Revealing modular
architecture of human brain structural networks by using cortical
thickness from MRI. Cerebral Cortex, 18(10), 2374–2381. https://
doi.org/10.1093/cercor/bhn003, PubMed: 18267952
Chiarello, C., Vazquez, D., Felton, A., et al. (2016). Structural asym-
t h e h u m a n c e r e b r a l c o r t e x : R e g i o n a l a n d
m e t r y o f
between-subject variability of surface area, cortical thickness
and local gyrification. Neuropsychologia, 93, 365–379. https://
doi.org/10.1016/j.neuropsychologia.2016.01.012, PubMed:
26792368
Chunling, W., & Zheng, X. (2016). Review on clinical update of
essential tremor. Neurological Sciences, 37(4), 495–502. https://
doi.org/10.1007/s10072-015-2380-1, PubMed: 26749268
Chung, S. J., Kwon, H., Lee, D. K., et al. (2013). Neuroanatomical
heterogeneity of essential tremor according to propanolol
response. PLoS One, 8(12), e84054. https://doi.org/10.1371
/journal.pone.0084054, PubMed: 24358327
Dale, A. M., Fischl, B., & Sereno, M. I. (1999). Cortical surface-
based analysis: I. Segmentation and surface reconstruction.
NeuroImage, 9(2), 179–194. https://doi.org/10.1006/nimg.1998
.0395, PubMed: 9931268
Deng, H., Wu, S., & Jankovic, J. (2019). Essential tremor: Genetic
update. Expert Reviews in Molecular Medicine, 21, e8. https://doi
.org/10.1017/erm.2019.7, PubMed: 31818346
Desikan, R. S., Ségonne, F., Fischl, B., et al. (2006). An automated
labeling system for subdividing the human cerebral cortex on
MRI scans into gyral based regions of interest. NeuroImage, 31(3),
968–980. https://doi.org/10.1016/j.neuroimage.2006.01.021,
PubMed: 16530430
Deuschl, G., & Elble, R. (2009). Essential tremor—Neurodegenerative
or nondegenerative disease towards a working definition of ET.
Movement Disorders, 24(14), 2033–2041. https://doi.org/10.1002
/mds.22755, PubMed: 19750493
Duan, X., Fang, Z., Tao, L., et al. (2021). Altered local and matrix
functional connectivity in depressed essential tremor patients.
BMC Neurology, 21(1), 1–9. https://doi.org/10.1186/s12883
-021-02100-3, PubMed: 33573615
Elaimy, A. L., Demakas, J. J., Arthurs, B. J., et al. (2010). Gamma
knife radiosurgery for essential tremor: A case report and review
of the literature. World Journal of Surgical Oncology, 8(1), 1–7.
https://doi.org/10.1186/1477-7819-8-20, PubMed: 20307307
Network Neuroscience
866
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
Elble, R. J., Shih, L., & Cozzens, J. W. (2018). Surgical treatments
for essential tremor. Expert Review of Neurotherapeutics, 18(4),
303–321. https://doi.org/10.1080/14737175.2018.1445526,
PubMed: 29475371
Evans, A.C. (2013). Networks of anatomical covariance. Neuro-
Image, 80, 489–504. https://doi.org/10.1016/j.neuroimage.2013
.05.054, PubMed: 23711536
Fahn, S., Tolosa, E., & Marín, C. (1993). Clinical rating scale for tremor.
Parkinson’s Disease and Movement Disorders, 2, 271–280.
Fischl, B. (2012). FreeSurfer. NeuroImage, 62(2), 774–781. https://
doi.org/10.1016/j.neuroimage.2012.01.021, PubMed: 22248573
Fischl, B., & Dale, A. M. (2000). Measuring the thickness of the human
cerebral cortex from magnetic resonance images. Proceedings of
the National Academy of Sciences, 97(20), 11050–11055. https://
doi.org/10.1073/pnas.200033797, PubMed: 10984517
Fischl, B., Sereno, M. I., & Dale, A. M. (1999). Cortical surface-based
analysis: II: Inflation, flattening, and a surface-based coordinate
system. NeuroImage, 9(2), 195–207. https://doi.org/10.1006
/nimg.1998.0396, PubMed: 9931269
Gironell, A. (2014). The GABA hypothesis in essential tremor:
Lights and shadows. Tremor and Other Hyperkinetic Movements,
4, 254. https://doi.org/10.5334/tohm.229, PubMed: 25120944
Gong, G., He, Y., Chen, Z. J., et al. (2012). Convergence and diver-
gence of thickness correlations with diffusion connections across
the human cerebral cortex. NeuroImage, 59(2), 1239–1248.
https://doi.org/10.1016/j.neuroimage.2011.08.017, PubMed:
21884805
Hallett, M. (2014). Tremor: Pathophysiology. Parkinsonism & Related
Disorders, 20, S118–S122. https://doi.org/10.1016/S1353-8020(13)
70029-4
Harnett, N. G., Stevens, J. S., Fani, N., et al. (2020). Acute posttrau-
matic symptoms are associated with multimodal neuroimaging
structural covariance patterns: A possible role for the neural
substrates of visual processing in posttraumatic stress disorder.
Biological Psychiatry: Cognitive Neuroscience and NeuroImaging,
7(2), 129–138. https://doi.org/10.1016/j.bpsc.2020.07.019,
PubMed: 33012681
He, Y., Chen, Z., & Evans, A. (2008). Structural insights into aberrant
topological patterns of large-scale cortical networks in Alzheimer’s
disease. Journal of Neuroscience, 28(18), 4756–4766. https://doi
.org/10.1523/JNEUROSCI.0141-08.2008, PubMed: 18448652
Hogstrom, L. J., Westlye, L. T., Walhovd, K. B., et al. (2013). The
structure of the cerebral cortex across adult life: Age-related
patterns of surface area, thickness, and gyrification. Cerebral
Cortex, 23(11), 2521–2530. https://doi.org/10.1093/cercor
/bhs231, PubMed: 22892423
Hopfner, F., & Helmich, R. C. (2018). The etiology of essential
tremor: Genes versus environment. Parkinsonism & Related
Disorders, 46, S92–S96. https://doi.org/10.1016/j.parkreldis
.2017.07.014, PubMed: 28735798
Ibrahim, M. F., Beevis, J. C., & Empson, R. M. (2021). Essential
tremor—A cerebellar driven disorder? Neuroscience, 462,
262–273. https://doi.org/10.1016/j.neuroscience.2020.11.002,
PubMed: 33212218
Jang, C., Park, H. J., Chang, W. S., et al. (2016). Immediate and
longitudinal alterations of functional networks after thalamotomy
in essential tremor. Frontiers in Neurology, 7, 184. https://doi.org
/10.3389/fneur.2016.00184, PubMed: 27822200
Jhunjhunwala, K., & Pal, P. K. (2014). The non-motor features of
essential tremor: A primary disease feature or just a secondary
phenomenon? Tremor and Other Hyperkinetic Movements, 4,
255. https://doi.org/10.5334/tohm.230, PubMed: 25120945
Kelly, P. A., Viding, E., Wallace, G. L., et al. (2013). Cortical
thickness, surface area, and gyrification abnormalities in children
exposed to maltreatment: Neural markers of vulnerability?
Biological Psychiatry, 74(11), 845–852. https://doi.org/10.1016/j
.biopsych.2013.06.020, PubMed: 23954109
Khundrakpam, B. S., Lewis, J. D., Reid, A., et al. (2017). Imaging
structural covariance in the development of intelligence. Neuro-
Image, 144, 227–240. https://doi.org/10.1016/j.neuroimage
.2016.08.041, PubMed: 27554529
Kuhlenbäumer, G., Hopfner, F., & Deuschl, G. (2014). Genetics of
essential tremor: Meta-analysis and review. Neurology, 82(11),
1000–1007. https://doi.org/10.1212/ WNL.0000000000000211,
PubMed: 24532269
Lerch, J. P., Worsley, K., Shaw, W. P., et al. (2006). Mapping
anatomical correlations across cerebral cortex (MACACC) using
cortical thickness from MRI. NeuroImage, 31(3), 993–1003.
https://doi.org/10.1016/j.neuroimage.2006.01.042, PubMed:
16624590
Li, J. Y., Suo, X. L., Li, N. N., et al. (2021). Disrupted brain network
topology in drug-naïve essential tremor patients with and without
depression. Clinical Neuroradiology, 31, 981–992. https://doi.org
/10.1007/s00062-021-01002-8, PubMed: 33687483
Lisman, J., Buzsáki, G., Eichenbaum, H., et al. (2017). Viewpoints:
How the hippocampus contributes to memory, navigation and
cognition. Nature Neuroscience, 20(11), 1434–1447. https://doi
.org/10.1038/nn.4661, PubMed: 29073641
Louis, E. D. (2016). Non-motor symptoms in essential tremor: A
review of the current data and state of the field. Parkinsonism
& Related Disorders, 22, S115–S118. https://doi.org/10.1016/j
.parkreldis.2015.08.034, PubMed: 26343494
Louis, E. D., & Ferreira, J. J. (2010). How common is the most com-
mon adult movement disorder? Update on the worldwide prev-
alence of essential tremor. Movement Disorders, 25(5), 534–541.
https://doi.org/10.1002/mds.22838, PubMed: 20175185
Mechelli, A., Friston, K. J., Frackowiak, R. S., et al. (2005). Struc-
tural covariance in the human cortex. Journal of Neuroscience,
25(36), 8303–8310. https://doi.org/10.1523/JNEUROSCI.0357
-05.2005, PubMed: 16148238
Menon, V., & Uddin, L. Q. (2010). Saliency, switching, attention
and control: A network model of insula function. Brain Structure
and Function, 214(5–6), 655–667. https://doi.org/10.1007
/s00429-010-0262-0, PubMed: 20512370
Meskaldji, D. E., Vasung, L., Romascano, D., et al. (2015).
Improved statistical evaluation of group differences in connec-
tomes by screening—Filtering strategy with application to study
maturation of brain connections between childhood and adoles-
cence. NeuroImage, 108, 251–264. https://doi.org/10.1016/j
.neuroimage.2014.11.059, PubMed: 25498390
Miyashita, Y. (1993). Inferior temporal cortex: Where visual percep-
tion meets memory. Annual Review of Neuroscience, 16(1),
245–263. https://doi.org/10.1146/annurev.ne.16.030193
.001333, PubMed: 8460893
Molent, C., Maggioni, E., Cecchetto, F. et al. (2018). Reduced cor-
tical thickness and increased gyrification in generalized anxiety
Network Neuroscience
867
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
disorder: A 3 T MRI study. Psychological Medicine, 48(12),
2001–2010. https://doi.org/10.1017/S003329171700352X,
PubMed: 29239286
Morgan, S. E., Seidlitz, J., Whitaker, K. J., et al. (2019). Cortical
patterning of abnormal morphometric similarity in psychosis is
associated with brain expression of schizophrenia-related genes.
Proceedings of the National Academy of Sciences, 116(19),
9604–9609. https://doi.org/10.1073/pnas.1820754116,
PubMed: 31004051
Muggleton, N. G., Juan, C. H., Cowey, A., et al. (2003). Human
frontal eye fields and visual search. Journal of Neurophysiology,
89(6), 3340–3343. https://doi.org/10.1152/jn.01086.2002,
PubMed: 12783960
Park, H. J., Pae, C., Friston, K., et al. (2017). Hierarchical dynamic
causal modeling of resting-state fMRI reveals longitudinal
changes in effective connectivity in the motor system after thala-
motomy for essential tremor. Frontiers in Neurology, 8, 346.
https://doi.org/10.3389/fneur.2017.00346, PubMed: 28775707
Phelps, E. A. (2006). Emotion and cognition: Insights from studies of
the human amygdala. Annual Review in Psychology, 57, 27–53.
https://doi.org/10.1146/annurev.psych.56.091103.070234,
PubMed: 16318588
Picillo, M., & Fasano, A. (2016). Recent advances in essential
tremor: Surgical treatment. Parkinsonism & Related Disorders,
22, S171–S175. https://doi.org/10.1016/j.parkreldis.2015.09
.012, PubMed: 26382842
Pietracupa, S., Bologna, M., Bharti, K., et al. (2019). White matter
rather than gray matter damage characterizes essential tremor.
European Radiology, 29(12), 6634–6642. https://doi.org/10
.1007/s00330-019-06267-9, PubMed: 31139970
Prasad, S. Pandey, U., Saini, J., et al. (2019). Atrophy of cerebellar
peduncles in essential tremor: A machine learning-based volumet-
ric analysis. European Radiology, 29(12), 7037–7046. https://doi
.org/10.1007/s00330-019-06269-7, PubMed: 31161314
Quattrone, A., Cerasa, A., Messina, D., et al. (2008). Essential head
tremor is associated with cerebellar vermis atrophy: A volumetric
and voxel-based morphometry MR imaging study. American
Journal of Neuroradiology, 29(9), 1692–1697. https://doi.org/10
.3174/ajnr.A1190, PubMed: 18653686
Raethjen, J., & Deuschl, G. (2012). The oscillating central network of
essential tremor. Clinical Neurophysiology, 123(1), 61–64. https://
doi.org/10.1016/j.clinph.2011.09.024, PubMed: 22055842
Raznahan, A., Shaw, P. Lalonde, F., et al. (2011). How does your cor-
tex grow? Journal of Neuroscience, 31(19), 7174–7177. https://doi
.org/10.1523/JNEUROSCI.0054-11.2011, PubMed: 21562281
Robinson, D. A., & Fuchs, A. F. (1969). Eye movements evoked by
stimulation of frontal eye fields. Journal of Neurophysiology, 32(5),
637–648. https://doi.org/10.1152/jn.1969.32.5.637, PubMed:
4980022
Romero-Garcia, R., Whitaker, K. J., Váša, F., et al. (2018). Structural
covariance networks are coupled to expression of genes
enriched in supragranular layers of the human cortex. Neuro-
Image, 171, 256–267. https://doi.org/10.1016/j.neuroimage
.2017.12.060, PubMed: 29274746
Rubinov, M., & Sporns, O. (2010) Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Sanabria-Diaz, G., Melie-García, L., Iturria-Medina, Y., et al.
(2010). Surface area and cortical thickness descriptors reveal
different attributes of the structural human brain networks. Neuro-
Image, 50(4), 1497–1510. https://doi.org/10.1016/j.neuroimage
.2010.01.028, PubMed: 20083210
Schmitt, J. E., Lenroot, R. K., Wallace, G. L., et al. (2008). Identifi-
cation of genetically mediated cortical networks: A multivariate
study of pediatric twins and siblings. Cerebral Cortex, 18(8),
1737–1747. https://doi.org/10.1093/cercor/ bhm211, PubMed:
18234689
Schnack, H. G., Van Haren, N. E., Brouwer, R. M., et al. (2015).
Changes in thickness and surface area of the human cortex
and their relationship with intelligence. Cerebral Cortex, 25(6),
1608–1617. https://doi.org/10.1093/cercor/ bht357, PubMed:
24408955
Seeley, W. W., Crawford, R. K., Zhou, J., et al. (2009). Neurodegen-
erative diseases target large-scale human brain networks. Neuron,
62(1), 42–52. https://doi.org/10.1016/j.neuron.2009.03.024,
PubMed: 19376066
Ségonne, F., Dale, A. M., Busa, E., et al. (2004). A hybrid approach
to the skull stripping problem in MRI. NeuroImage, 22(3),
1060–1075. https://doi.org/10.1016/j.neuroimage.2004.03.032,
PubMed: 15219578
Serrano, J. I., Romero, J. P., Castillo, M. D. D., et al. (2017). A data
mining approach using cortical thickness for diagnosis and char-
acterization of essential tremor. Scientific Reports, 7(1), 1–16.
https://doi.org/10.1038/s41598-017-02122-3, PubMed:
28526878
Sharifi, S., Nederveen, A. J., Booij, J., et al. (2014). Neuroimaging
essentials in essential tremor: A systematic review. NeuroImage:
Clinical, 5, 217–231. https://doi.org/10.1016/j.nicl.2014.05.003,
PubMed: 25068111
Siokas, V., Aloizou, A. M., Tsouris, Z., et al. (2020). Genetic risk
factors for essential tremor: A review. Tremor and Other Hyper-
kinetic Movements, 10, 4. https://doi.org/10.5334/tohm.67,
PubMed: 32775018
Stuss, D. T. (2011). Functions of the frontal lobes: Relation to
executive functions. Journal of the International Neuropsycho-
logical Society, 17(5), 759–765. https://doi.org/10.1017
/S1355617711000695, PubMed: 21729406
Tadayon, E., Pascual-Leone, A., & Santamecchi, E. (2020). Differen-
tial contribution of cortical thickness, surface area, and gyrifica-
tion to fluid and crystallized intelligence. Cerebral Cortex, 30(1),
215–225. https://doi.org/10.1093/cercor/ bhz082, PubMed:
31329833
Taylor, S. F., Stern, E. R., & Gehring, W. J. (2007). Neural systems for
error monitoring: Recent findings and theoretical perspectives.
The Neuroscientist, 13(2), 160–167. https://doi.org/10.1177
/1073858406298184, PubMed: 17404376
Tio, M., & Tan, E. K. (2016). Genetics of essential tremor. Parkinsonism
& Related Disorders, 22, S176–S178. https://doi.org/10.1016/j
.parkreldis.2015.09.022, PubMed: 26411503
Tröster, A. I., Pahwa, R., Fields, J. A., et al. (2005). Quality of life in
Essential Tremor Questionnaire (QUEST): Development and initial
validation. Parkinsonism & Related Disorders, 11(6), 367–373.
https://doi.org/10.1016/j.parkreldis.2005.05.009, PubMed: 16103000
Tuleasca, C., Najdenovska, E., Régis, J., et al. (2018). Clinical
response to Vim’s thalamic stereotactic radiosurgery for essential
Network Neuroscience
868
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morphometry in essential tremor and recovery after thalamotomy
tremor is associated with distinctive functional connectivity
patterns. Acta Neurochirurgica, 160(3), 611–624. https://doi.org
/10.1007/s00701-017-3456-x, PubMed: 29335882
Tuleasca, C., Regis, J., & Levivier, M. (2018). Essential tremor. The
New England Journal of Medicine, 379(6), 595–596. https://doi
.org/10.1056/NEJMc1807690, PubMed: 30091353
Tuleasca, C., Witjas, T., Najdenovska, E., et al. (2017). Assessing
the clinical outcome of Vim radiosurgery with voxel-based
morphometry: Visual areas are linked with tremor arrest! Acta
Neurochirurgica, 159(11), 2139–2144. https://doi.org/10.1007
/s00701-017-3317-7, PubMed: 28942466
Tuleasca, C., Witjas, T., Van De Ville, D., et al. (2018). Right
Brodmann area 18 predicts tremor arrest after Vim radiosurgery:
A voxel-based morphometry study. Acta Neurochirurgica, 160,
603–609. https://doi.org/10.1007/s00701-017-3391-x, PubMed:
29128955
Villablanca, J. R. (2010). Why do we have a caudate nucleus? Acta
Neurobiologiae Experimentalis, 70(1), 95–105. PubMed: 20407491
Weiner, K. S., & Zilles, K. (2016). The anatomical and functional
specialization of the fusiform gyrus. Neuropsychologia, 83, 48–62.
https://doi.org/10.1016/j.neuropsychologia.2015.06.033, PubMed:
26119921
Wierenga, L. M., Langen, M., Oranje, B., et al. (2014). Unique
developmental trajectories of cortical thickness and surface area.
NeuroImage, 87, 120–126. https://doi.org/10.1016/j.neuroimage
.2013.11.010, PubMed: 24246495
Xiong, Y., Han, D., He, J. et al. (2021). Correlation of visual area
with tremor improvement after MRgFUS thalamotomy in Parkin-
son’s disease. Journal of Neurosurgery, 136(3), 681–688. https://
doi.org/10.3171/2021.3.JNS204329, PubMed: 34479209
Yang, J. J., Kwon, H., & Lee, J. M. (2016). Complementary charac-
teristics of correlation patterns in morphometric correlation
networks of cortical thickness, surface area, and gray matter
volume. Scientific Reports, 6(1), 1–9. https://doi.org/10.1038
/srep26682, PubMed: 27226000
Yang, J., Lei, D., Peng, J., et al. (2021). Disrupted brain gray matter
networks in drug-naïve participants with essential tremor. Neuro-
radiology, 63, 1501–1510. https://doi.org/10.1007/s00234-021
-02653-7, PubMed: 33782719
Yang, J.-J., Yoon, U., Im, K., et al. (2013). Prediction of human intelli-
gence using morphometric characteristics of cortical surface: Partial
least squares analysis. Neuroscience, 246, 351–361. https://doi.org
/10.1016/j.neuroscience.2013.04.051, PubMed: 23643979
Yee, Y., Fernandes, D. J., French, L., et al. (2018). Structural covari-
ance of brain region volumes is associated with both structural
connectivity and transcriptomic similarity. NeuroImage, 179,
357–372. https://doi.org/10.1016/j.neuroimage.2018.05.028,
PubMed: 29782994
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
5
0
2
0
4
6
4
6
1
n
e
n
_
a
_
0
0
2
5
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
869