Informe
The Neural Representational
Space of Social Memory
Sarah L. Dziura1 and James C. Thompson1
1Department of Psychology, George Mason University, Fairfax, Virginia, 22030 EE.UU
Palabras clave: social networks, social cognition, aprendiendo, representational similarity analysis, resonancia magnética funcional
un acceso abierto
diario
ABSTRACTO
Social functioning involves learning about the social networks in which we live and interact;
knowing not just our friends, but also who is friends with our friends. This study utilized an
incidental learning paradigm and representational similarity analysis (RSA), a functional MRI
multivariate pattern analysis technique, to examine the relationship between learning social
networks and the brain’s response to the faces within the networks. We found that accuracy
of learning face pair relationships through observation is correlated with neural similarity
patterns to those pairs in the left temporoparietal junction (TPJ), the left fusiform gyrus, y
the subcallosal ventromedial prefrontal cortex (vmPFC), all areas previously implicated in
social cognition. This model was also significant in portions of the cerebellum and thalamus.
These results show that the similarity of neural patterns represent how accurately we
understand the closeness of any two faces within a network. Our findings indicate that these
areas of the brain not only process knowledge and understanding of others, but also support
learning relations between individuals in groups.
INTRODUCCIÓN
Social relationships guide and support much of human behavior. Not only do we form
strong lifelong family bonds, we also interact with others in work, education, and leisure
settings and create lasting non-kin relationships. For many species, including humans, non-
kin–based social networks can have important consequences for health and fitness (Cheney
2011; Tung et al.,2015). Benefits of social relationships can come both from direct connections
(our friends) as well as second-order or indirect connections (friends of our friends) (Brent
2015; Seyfarth & Cheney, 2015). Knowledge of social ties involves a network of brain regions
including bilateral posterior superior temporal sulcus (pSTS) and temporoparietal junction
(TPJ) (Bault, Pelloux, Fahrenfort, Ridderinkhof, & van Winden,2015; Satpute, malo, & Ochsner,
2014). Learning and representing information about social hierarchies also recruits amyg-
dala, hippocampus, and ventral mPFC (Kumaran, Melo, & Duzel, 2012; Zink et al., 2008).
Recientemente, the position of individuals in one’s own social network were found to be represented
in lateral temporal, inferior parietal, and medial and lateral prefrontal cortices as participants
viewed videos of network members (Parkinson, Kleinbaum, & Wheatley,2017). These findings
suggest that information about personal social connections is represented in patterns of fMRI
responses elicited by viewing the individuals from one’s real-world network. Sin embargo, menos
is known about how we learn, remember, and represent information about new and indi-
rect social connections. Understanding this process is important, as how we interact with our
extended social network is often influenced by our knowledge of established relationships.
Citación: Dziura, S. l., & Thompson,
j. C. (2018). The Neural
Representational Space of Social
Memoria. Mente abierta: Descubrimientos
en Ciencias Cognitivas, 3, 1–12.
https://doi.org/10.1162/opmi_a_00021
DOI:
https://doi.org/10.1162/opmi_a_00021
Materiales suplementarios:
https://doi.org/10.1162/opmi_a_00021
Recibió: 18 Enero 2018
Aceptado: 7 Noviembre 2018
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Sarah L. Dziura
sdziura@gmu.edu
Derechos de autor: © 2018
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
oh
pag
metro
_
a
_
0
0
0
2
1
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
One of the ways that we acquire this knowledge is by observing patterns of associations
between individuals (De Soto, 1960). The frequency of interaction between individuals can
provide an important index of their relationship strength (Hombre libre, Hombre libre, & Michaelson,
1988; lin, Dayton, & Greenwald,1978). In nonhuman primates, observation of the frequency of
association is suggested to be an essential cue to learning information about social connections
(Seyfarth & Cheney, 2012; Tiddi, Sorrentino, pescador, & Schino, 2017). Studies of the cognitive
representation of social networks in humans have employed artificial social networks learned
through paired associates learning-type paradigms (De Soto, 1960; De Soto & Bosley, 1962;
Janicik & Larrick,2005; Zitek & Tiedens,2012). Artificial networks give the ability to experimen-
tally control network statistics, for instance by varying the centralization (the extent to which
a network is grouped around a particular point), which can affect the efficiency of information
passed among members and consequently group performance (Leavitt, 1951). They also allow
us to assess the role of the memory (en)accuracy for relationship characteristics in participants’
representations of social networks (Brashears,2013; Zitek & Tiedens,2012). Humans can learn
quite complex statistical relationships between stimuli through simple associations, even in the
absence of explicit knowledge of these relationships (Turk-Browne, Scholl, Johnson, & Chun,
2010). We expect that such associative learning plays an important role in acquiring knowl-
edge about social connections. We also expect that the accuracy with which participants recall
these connections plays a role in how these associations are represented neurally.
We examined the memory and neural representation of connections between members
of two novel social networks of varying centralization, using fMRI and representational similar-
ity analysis (RSA). We examined if the pattern similarity of fMRI responses to any two faces from
a learned social network reflected the tie strength (closeness) of those two individuals within
the network: eso es, does the similarity of the pattern of response to two network members
increase as a function of the closeness of those members? We also examined if the memory for
tie strength between network members was related to the similarity of the fMRI voxel pattern
response to the faces of members.
MATERIALES Y MÉTODOS
Participantes
Twenty-two healthy individuals (10 hembras; age range = 18–34; edad media = 23; ethnicity =
64% Blanco, 18% Hispanic/Latino, 18% asiático) participated in a 1.5-hour learning session
immediately followed by a 1.5-hour fMRI scanning session. Behavioral data from a total of
31~individuals were collected, pero 7 subjects did not meet the learning criteria from the behav-
ioral task, one subject was unable to be scanned, and one subject’s fMRI data were incomplete.
All participants were right-handed (self-reported) with normal or corrected-to-normal vision.
Participants provided written informed consent in accordance with the Declaration of Helsinki
and the Human Subjects Review Board at George Mason University and were compensated
for their time.
Experimental Design and Statistical Analysis
Task stimuli consisted of 24 faces of varying ethnicities, equally divided by gender.
Estímulos.
Faces were all in color and facial expressions were all smiling. These stimuli were downloaded
from the Park Aging Mind Laboratory Database at UT Dallas (Minear & Parque, 2004) and were
chosen to be as realistic to a college campus as possible, ensuring the perception of real people
who might interact and be friends with each other.
Participants completed a two-alternative forced-choice task to become famil-
Task Design.
iar with the structure of two 6-person social networks (Cifra 1). Pairs of faces represented
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
2
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
.
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
oh
pag
metro
_
a
_
0
0
0
2
1
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
oh
pag
metro
_
a
_
0
0
0
2
1
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
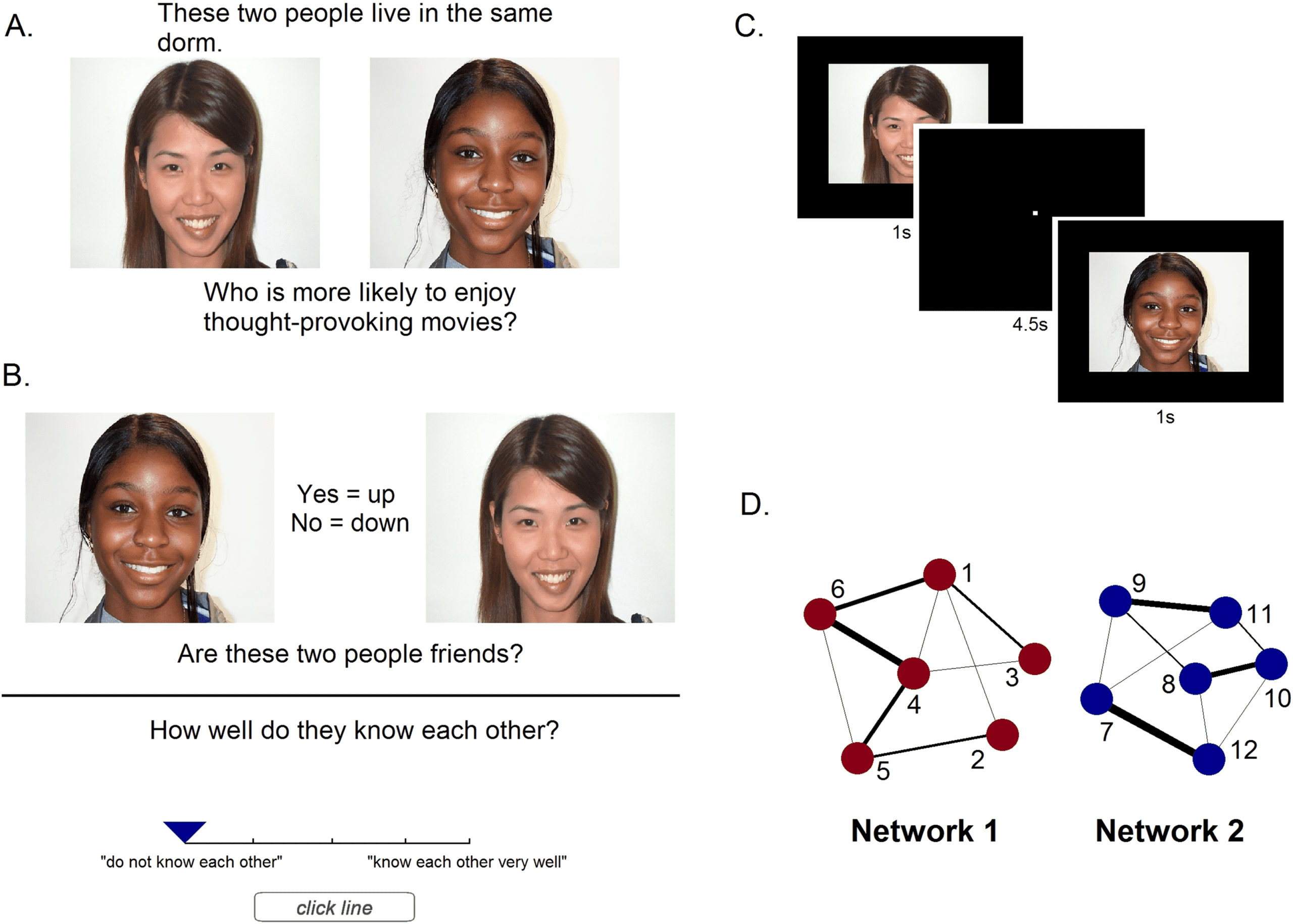
Cifra 1. Details of the experimental design. A. Example trial of the paired presentation of a social network tie, where subjects were asked to
judge between the two faces on an unrelated characteristic. B. Example trial of the recall task, where subjects were asked to report whether
a pair of faces was connected, and how well they know each other (0–4 scale). C. fMRI task, where each face was presented individually for
1 segundo (4.5-second inter-stimulus interval). D. Structure of the two social networks. Each node represents a different face and line thickness
represents connection strength. All ties are nondirected (reciprocal).
connections within each network, with the frequency of pairing indicating relationship strength.
Each network had an equal number of male and female faces of varying ethnicities. Red
properties differed between the two in that although each network had an equal number of con-
nections of each strength level, there were differences among the individual members (faces) en
each network. The faces in network 1 had varying numbers of connections and therefore each
had a different average closeness to the rest of the network, whereas the faces in network 2
had an equal number of connections and an equal average closeness to all other faces in the
network. This meant that in network 1 the centrality of members was varied (variable-centrality
network), while in network 2 centrality was equated across members (fixed-centrality network).
This also meant that the frequency of presentation of each individual face differed in network 1,
but was equivalent in network 2. Each trial consisted of a face pair presented for 4 artículos de segunda clase
accompanied by a question, and participants were asked to make a comparison between the
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
3
Neural Representation of Social Memory Dziura and Thompson
faces and decide which person better fit the question. Questions consisted of behavioral and
personality characteristics taken from various personality surveys included in the International
Personality Item Pool (http://ipip.ori.org/). Half of the questions asked which person was more
likely to exhibit a characteristic, and half asked which person was less likely (ejemplo: “Who
is more likely to be easily intimidated?"). Network learning took place in alternating blocks,
where the subjects viewed 36 randomly presented trials of one network followed by 36 ensayos
of the second network. Participants completed 720 trials in total (360 per network), con el
weakest network connections being presented a total of 20 times and the strongest a total of
80 veces.
After completing the paired face viewing portion, participants were explicitly tested on
their knowledge of the network connections. They were told that the faces represented col-
lege students living in a dorm together, the faces that they saw paired together previously
represented friend connections, and the more often they were presented together, the closer
in friendship the pair was. They were asked to group all faces into two separate halls to ensure
that they could distinguish between the faces in different networks. They were then presented
with all possible within-network face pairs twice and asked to rate their relationship on a scale
de 0 (do not know each other) a 4 (know each other very well). They were not asked about
cross-network face relationships. This explicit testing period was included to ensure that par-
ticipants learned the structure of the networks to an appropriate level before being scanned.
Participants who were within 2 standard deviations of pilot data (hit rate = .85, DE = 0.14;
false alarm rate = .35, DE = 0.15) were included in further analysis. Both parts of the behav-
ioral task (learning and recall) were presented to the participant using PsychoPy version 1.842
software (Peirce, 2007).
The fMRI task stimuli included the same 12 faces from the behavioral task as well as
12 novel faces as a control. Faces were presented one at a time for one second on a black
background with a 4.5-second inter-stimulus interval (black screen with a white fixation dot),
and participants completed a 1-back task to ensure attention. The task consisted of four runs
de 9.6 minutes each, resulting in each face being presented a total of 16 veces (not count-
ing repeats, which were included in analysis as a separate regressor). Following the face task,
participants underwent a dynamic localizer session. Localizer stimuli consisted of 18-s blocks
each of moving faces, body parts, outdoor scenes, objects, and scrambled objects. The fMRI
experiment was presented to the participant using Presentation software (Versión 16.3, Neuro-
behavioral Systems, Cª, berkeley, California, www.neurobs.com). Analysis of social network recall
data was conducted in Microsoft Excel (versión 2016) and R Version 3.3.2 (R Core Team, 2017).
RSA is a form of multivariate pattern analysis that com-
Representational Similarity Analysis.
pares the distance between stimuli in neural representational space (Kriegeskorte, Mur, &
Bandettini, 2008), and correlates these neural patterns of information with external patterns of
información. In this way it can be utilized to assess different models or patterns of cognition above
and beyond univariate analysis, or even more traditional multivariate pattern classification
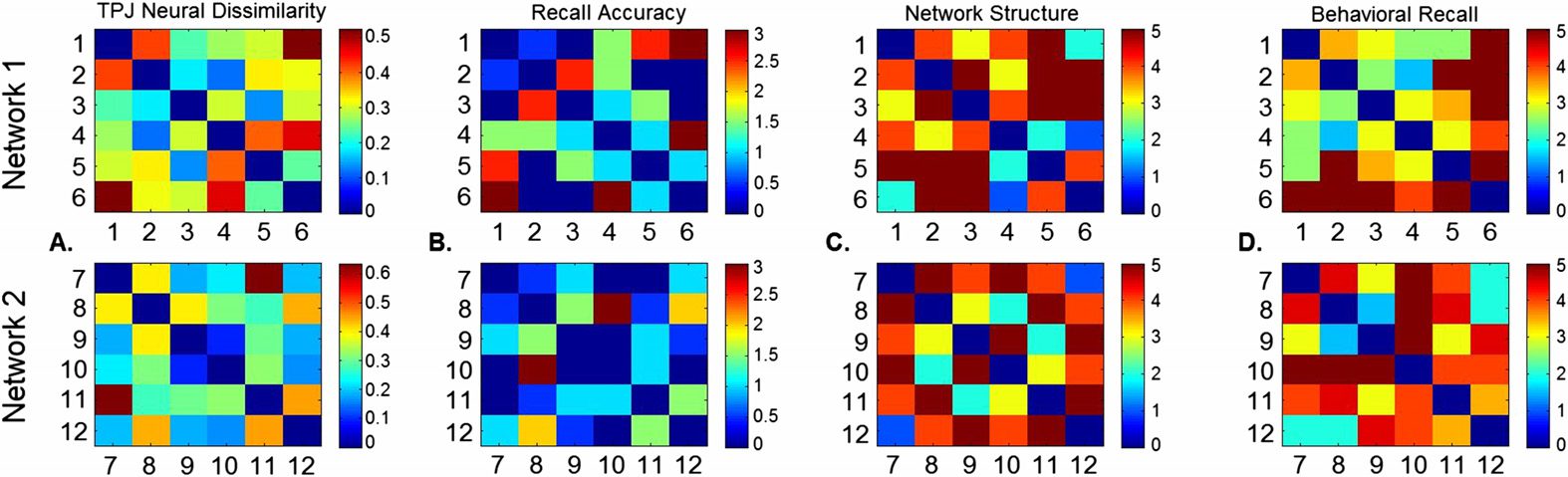
técnicas (Haxby, Connolly, & Guntupalli, 2014). Four separate dissimilarity matrices (DMs)
were created for each network (Cifra 2): true network structure, perception of network
estructura, group average of perceived structure, and recall accuracy. To contrast within-network
and cross-network pairs, a DM was created with all 12 faces included. This treated all con-
nected faces the same, then coded within-network unconnected faces as more dissimilar,
followed by cross-network unconnected faces. For more detail on this analysis, as well as fMRI
preprocessing and univariate analysis information, ver los materiales complementarios (Dziura &
Thompson, 2018).
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
oh
pag
metro
_
a
_
0
0
0
2
1
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
Cifra 2. Dissimilarity matrices between face pairs for a sample subject. A. Neural dissimilarity
in an example region in the temporoparietal junction. B. Recall accuracy DM (0 = perfectly accurate
recordar). C. True network structure DM (5 = unconnected). D. Behavioral recall of face pair strength
(5 = unconnected).
RESULTADOS
Behavioral task
Subjects correctly identified social network relationship ties significantly greater than chance
across both networks [(t(21) = 8.08, pag < .0005]. Table 1 shows the average hit rate, false alarm
rate, sensitivity (d’), and the correlation between true and reported perceived strength for ties
and relationship strength across subjects. Paired sample two-tailed t tests revealed no signifi-
cant differences between recall measures for the two networks. There were also no significant
age or gender effects for any of the measures. When averaged together across subjects, group
perceived relationship strength was highly correlated with the true network structure (r = .896,
p < .0005). In order to assess whether our behavioral task was comparable to previous forms of
social network learning and recall, we calculated performance measures used by Brashears
(2013). Accuracy refers to the number of ties correctly recalled divided by the number of to-
tal ties reported, coverage refers to the number of ties correctly recalled divided by the total
tie number in the network, and performance refers to the product of accuracy and coverage.
T tests revealed no significant differences between accuracy or performance measures in our
task and those of Brashears [accuracy: t(21) = 0.98, p = .34; performance: t(21) = 0.58, p = .56],
and we actually saw an increase in coverage [t(21) = 3.58, p < .005], although our networks
Table 1. Accuracy of recalling network relationships after incidental learning.
Network 1
Network 2
Both Networks
t value ( p)
Hit Rate
SD
False Alarm Rate
SD
d’
SD
Strength Correlation (r)
SD
.82
0.14
.39
0.21
1.3
0.71
.58
0.25
.83
0.11
.47
0.24
1.2
0.86
.53
0.21
.83
0.09
.43
0.19
1.2
0.6
.54
0.21
−0.13 (.89)
−1.23 (.23)
0.70 (.49)
0.82 (.42)
Notes. Hit rate, false alarm rate, and d’ represent the accuracy of recalling the true connections
within the networks. Strength correlation refers to the correlation between the matrix of true
relationship strength of the faces in the networks and the behavioral judgments of strength, and
is therefore a measure of accuracy of recalling relationship strength. T values and p values for
paired sample two-tailed t tests between the two networks are reported at the right of the table.
Bold indicates primary data, and italics indicate the standard deviation of the data.
OPEN MIND: Discoveries in Cognitive Science
5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
/
.
/
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
/
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
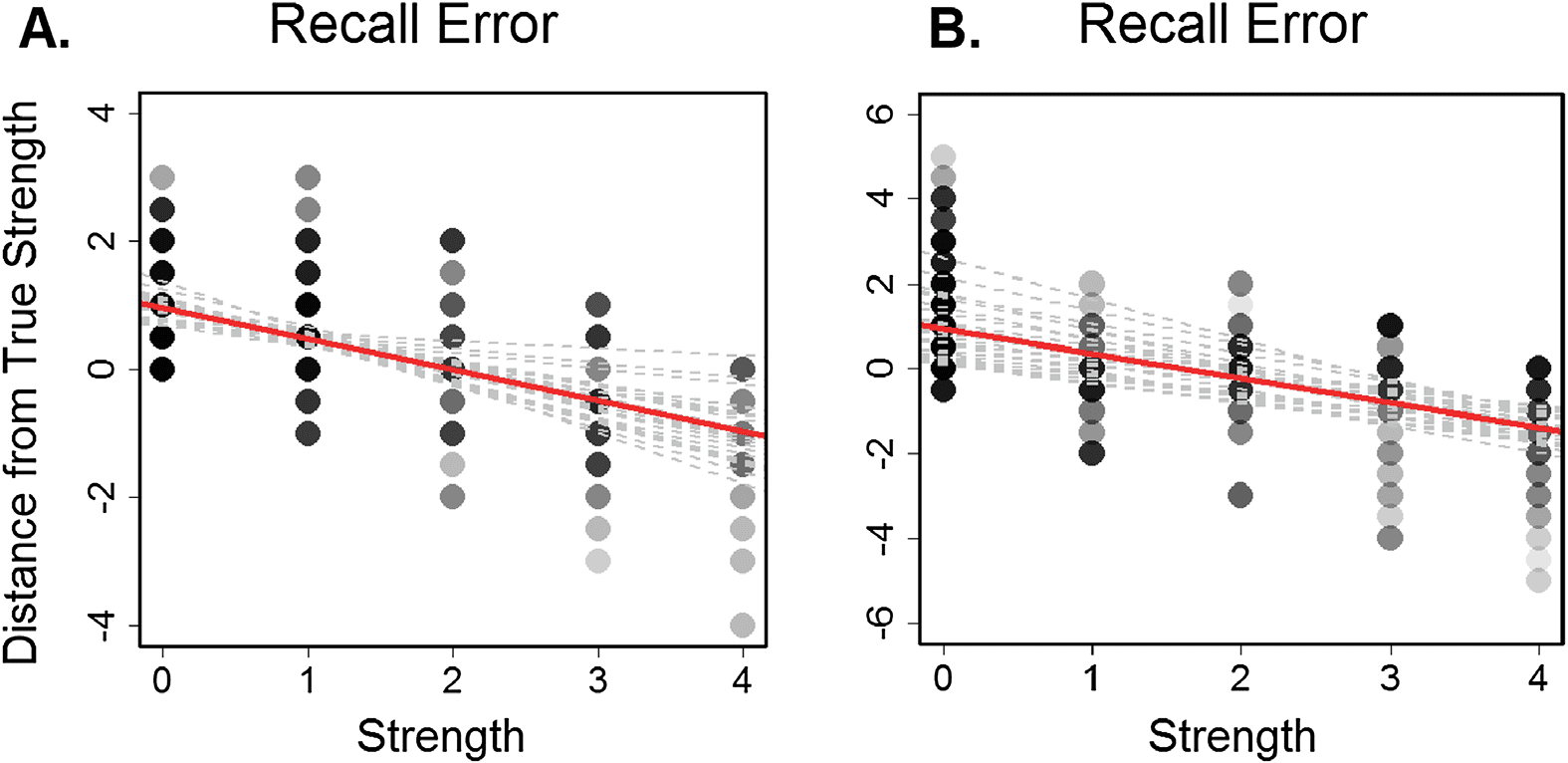
Figure 3. Subject recall accuracy by relationship strength. A. Each subject’s error by pair strength
level (0 = unconnected, 4 = close friends) from the primary dataset. B. Subject error by pair strength
level (0 = unconnected, 6 = close friends) from the secondary dataset with a larger social network
(N = 9). Positive values = overestimation of strength and negative values = underestimation of
strength. Darker colors indicate more data overlap at that point. Gray lines show individual subject
regression lines. The red line shows the group regression line.
were smaller, so participants did not need to remember as many ties. In order to thoroughly
explore network recall, we not only looked at the correctly identified ties, but also at any sys-
tematic biases that could be predicted by the level of relationship strength of the friend pairs.
We assessed recall by relationship strength by the relative direction of the errors made (i.e.,
how much subjects overestimated or underestimated the strength of the connection). A linear
mixed-effects regression model (fixed effect = strength; random effects = subject, residual) re-
vealed that relationship strength affected recall error compared to a null model [χ2(1) = 226.9,
p < .0005]. This pattern shows that overall, weak ties were reported to be stronger than they
actually were whereas strong ties were reported to be less strong (Figure 3a). This reflects a
general tendency to assume a mid-level relationship between observed people when the rela-
tionship is unable to be recalled. This central tendency effect seems to be robust, as it was also
observed in a separate subject sample (N = 23, 17 females, mean age = 19.6 [SD = 2.4])
learning a larger social network (N = 9) and a larger possible range of relationship tie strengths
to choose from (0-6) (χ2(1) = 362.84, p < .0005) (Figure 3b). To compare network memory
performance to the neural patterns in response to each face in the network, we converted the
relative error for each subject to absolute error, which gives a measure of distance from the true
network structure, regardless of the direction. The absolute error measure for each subject for
each network was then used as a dissimilarity model for RSA to elucidate what neural patterns
underlie these memory patterns.
Representational Similarity Analysis
We carried out RSA searchlight analysis on several DMs representing different types of infor-
mation about the networks. The first compared neural pattern similarity to social tie strength,
with more similar neural responses to any pair of faces representing a closer relationship be-
tween those faces. Neural pattern similarity that reflects this network structure would indicate
that the brain carries information about the true relationship between individuals, regardless of
whether people recall those relationships accurately. We did not find a significant correlation
OPEN MIND: Discoveries in Cognitive Science
6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
.
/
/
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
.
/
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
between these measures in our analyses. As the network properties differed between networks
1 and 2 (see Methods section for details), we compared the two networks and found no signif-
icant differences.
The second RSA searchlight compared neural pattern similarity to the subjects’ memory
for network tie strength. We assessed this by measuring each subject’s absolute distance from
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
.
/
/
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
.
/
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
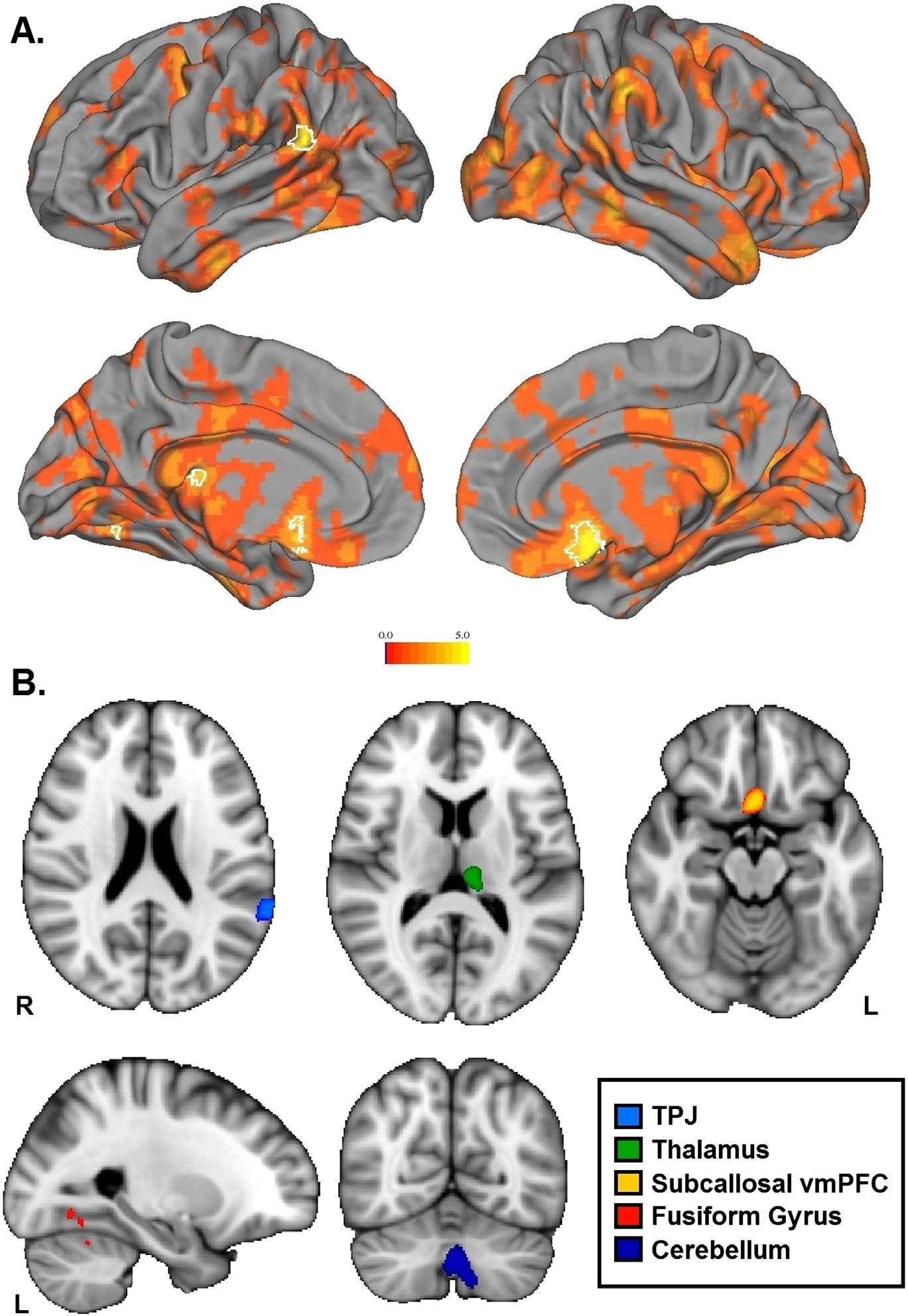
Figure 4. Results from group-level nonparametric 1-sample t test on the correlation maps from
RSA searchlight on the recall accuracy model. A. t-statistic map of all positive t values projected
onto the surface, where white borders delineate significant clusters from the group analysis (p <
.05, familywise error-corrected with threshold-free cluster enhancement). B. The same significant
clusters projected in the volume.
OPEN MIND: Discoveries in Cognitive Science
7
Neural Representation of Social Memory Dziura and Thompson
Table 2. Coordinates, cluster size, and peak activity for the group-level significant clusters from
the recall accuracy model.
Cluster
Peak Value (t)
Voxels
Cerebellum
Subcallosal vmPFC
Thalamus
TPJ
Fusiform Gyrus
Fusiform Gyrus
Fusiform Gyrus
Lateral Occipital
Lateral Occipital
3.92
5.6
4.14
4.31
3.28
3.5
3.37
3.65
3.66
730
274
132
117
11
5
5
3
3
x
0
2
−10
−64
−26
−24
−26
−52
−56
y
−64
14
−28
−48
−60
−66
−56
−56
−70
z
−38
−16
10
18
−12
−8
26
2
−2
each true network structure and the 1-correlation distance between the neural response to
each face viewed in the scanner. An association between these two measures would indicate
that the more accurately a subject perceives the true relationship strength between a pair of
faces, the more similar their neural pattern response is to those two faces. In other words, this
association does not rely on the actual connection strength of the relationships themselves,
but the subject’s memory of that connection, reflecting a second-order knowledge or under-
standing of a social relationship. Neural pattern similarity in the left TPJ, the left fusiform gyrus,
the subcallosal cingulate cortex, the cerebellum, the left thalamus, and a small portion of the
left lateral occipital lobe was significantly correlated with the recall accuracy model, suggest-
ing that neural populations within these areas are important for accurate perception of social
relationship strength (Figure 4). Table 2 reports MNI coordinates, cluster size, and peak voxel
activity of results. As with tie strength similarity, we compared the two networks to each other
separately and found no significant differences.
We also conducted RSA searchlights using two other dissimilarity matrix models: re-
called structure as measured by behavioral judgments, and the group average of those be-
havioral judgments (Figure 4). Neural pattern similarity that reflects behavioral recall would
indicate that the brain carries information about an individual’s perception of relationships, re-
gardless of how accurate those perceptions are. However, neither model reached significance.
Finally, we utilized a separate functional localizer to create regions of interest selective for face
processing in the bilateral superior temporal sulcus (STS) and fusiform gyrus. We also created
bilateral hippocampus regions of interest (ROI) using anatomical subcortical segmentation. We
conducted RSA correlations across each ROI for every subject (see the Supplemental Materials
[Dziura & Thompson, 2018]). No regions yielded significant results.
DISCUSSION
In this study, we used fMRI and RSA to examine the neural representational space of social
connections, learned from observing how frequently individuals were paired together. We ex-
amined if either (1) the tie strength between pairs of network members or (2) the memory
for the relationship between pairs of network members was represented in the human brain
via the similarity of fMRI responses associated with viewing the faces of those members. We
found support for the second proposal, but not the first. The accuracy of relationship recall
was represented in several brain regions, including the TPJ, subcallosal vmPFC, fusiform gyrus,
cerebellum, and thalamus. Our results indicate that these regions represent memory or knowl-
edge of the association between pairs of faces. That is, these areas code memory for association
OPEN MIND: Discoveries in Cognitive Science
8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
/
.
/
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
.
/
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
strength regardless of what that strength is. The more accurately a participant recalled the tie
strength for a pair of faces, the more similar the pattern of fMRI responses was to viewing those
two faces.
The correlation between neural pattern similarity and knowledge of a relationship be-
tween network members is a second-order association. Previous studies have shown that this
is not the sole mechanism for representing knowledge about relationships between people;
neural pattern similarity also codes direct social distance in other brain areas (Parkinson et al.,
2017). It is possible that the incidental learning task did not allow for these social network
relationships to be embedded enough to see this relationship in all our subjects, unlike real-
world or personal relationships that hold more importance to individuals. However, it may be
useful to represent this second-order knowledge about social information as well, especially
during the process of learning new relationships, or when gathering information about the
social world. Confidence in our knowledge about others can affect how we interact with
them. In simple terms, we have shown that the brain not only codes what we know, but also
how well we know it.
Both the TPJ and vmPFC have been consistently linked with knowledge and recall of
complex social information, such as tracking the popularity of real-world social network mem-
bers (Zerubavel et al., 2015). The TPJ, dorsomedial PFC, and ventrolateral PFC are engaged
when participants recall different facets of socially relevant knowledge (Satpute et al., 2014).
The left TPJ is selectively modulated by vasopressin, a neuropeptide linked to complex so-
cial behaviors, during social recognition (Zink et al., 2011) and lesions to the left TPJ lead to
specific deficits in social reasoning (Samson, Apperly, Chiavarino, & Humphreys, 2004). The
vmPFC shows increased activation when thinking about friends compared to kin (Wlodarski
& Dunbar, 2016), and the subgenual cingulate cortex is involved in tracking individual differ-
ences in perceptions of cohesiveness in kin groups (Rüsch et al.,2014). Our findings are in line
with this previous literature showing the importance of these areas in forming and maintaining
social relationships. The fusiform gyrus is also involved in social perception, particularly in
response to face stimuli (Atkinson & Adolphs, 2011; Kanwisher, McDermott, & Chun, 1997).
Our data indicate that patches of the fusiform gyrus do not simply perceive and distinguish
facial features, but are also involved in learning more abstract relationships between faces.
Areas of the cerebellum are also consistently activated in several features of social cognition,
with increases in activity occurring with increasing social abstraction levels in the cognitive
tasks (Overwalle, Baetens, Marien, & Vandekerckhove,2014). Our finding that the cerebellum
is involved in accurate knowledge of abstract learned relationships between others is consis-
tent with this. Furthermore, we found that the thalamus is also involved in this process. The
thalamus has a large number of connections to other areas of the brain, and has been shown
to have specific emotional and socially relevant associations (Christoffel et al., 2015; Feng
et al., 2016; Ioannidis et al., 2013). It also has high functional connectivity to the hippocam-
pus (Stein et al., 2000), and may be a critical link in the formation of episodic memories
(Aggleton et al., 2010).
While most of our subjects were able to accurately report relationship ties, there were
individual differences between ability to recall relationship strength, consistent with prior
literature (Casciaro, 1998). People also show differences in the ability to perceive and remem-
ber nonsocial patterns, but evidence suggests that learning, remembering, and storing social
information might be distinct from traditional learning and memory systems (Meyer, Taylor, &
Lieberman, 2015; Okuyama, Kitamura, Roy, Itohara, & Tonegawa, 2016; Tendler & Wagner,
2015). In this study we examined the representation of social connections acquired incidentally
OPEN MIND: Discoveries in Cognitive Science
9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
.
/
/
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
/
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
from the frequency of co-occurrence of different face pairs. While such paired associates-type
learning has its limitations in modeling the rich and complex nature of social connections
acquired in real life, it is likely that we learn the statistics of connections between people in
part through repeated observation of their association. This study does not directly examine
whether these associative neural patterns are exclusively social, as all learned pairs were faces
within a larger network. However, unlike previous studies of the representation of statistical
regularities of abstract, nonsocial stimuli, we did not observe representation in medial temporal
regions (Garvert, Dolan, & Behrens, 2017; Schapiro, Kustner, & Turk-Browne, 2012; Schapiro,
Turk-Browne, Norman, & Botvinick, 2016). Future work might seek to determine the extent to
which memory for social connections reflects the involvement of a general purpose system or
if there are distinct mechanisms for representing this kind of social information.
The way in which people learn and remember associations between individuals in groups
has a considerable impact on everyday life. We are not only able to perceive and understand
the social signals of other individuals, but we can also perceive and understand information
about social connections or relationships in which we are not directly involved. Our results
show that the memory for these indirect connections is coded in the pattern of neural responses
associated with viewing related individuals. This is an important skill because the accuracy
with which we see and remember subtle associations from our surroundings helps us move
more freely and easily in our highly social world.
FUNDING INFORMATION
JCT, Office of Naval Research (http://dx.doi.org/10.13039/100000006), Award ID: N00014-
10-1-0198.
AUTHOR CONTRIBUTIONS
SLD: Data curation: Lead; Formal analysis: Lead; Investigation: Lead; Methodology: Equal;
Validation: Lead; Visualization: Lead; Writing – original draft: Lead; Writing – review & editing:
Equal. JCT: Conceptualization: Lead; Formal analysis: Supporting; Funding acquisition: Lead;
Methodology: Equal; Project administration: Lead; Supervision: Lead; Writing – original draft:
Supporting; Writing – review & editing: Equal.
REFERENCES
Aggleton, J. P., O’Mara, S. M., Vann, S. D., Wright, N. F., Tsanov,
M., & Erichsen, J. T. (2010). Hippocampal-anterior thalamic
pathways for memory: Uncovering a network of direct and
indirect actions. The European Journal of Neuroscience, 31,
2292–2307. doi:10.1111/j.1460-9568.2010.07251.x
Atkinson, A. P., & Adolphs, R. (2011). The neuropsychology of
face perception: Beyond simple dissociations and functional
selectivity. Philosophical Transactions of the Royal Society B,
366, 1726–1738. doi:10.1098/rstb.2010.0349
Bault, N., Pelloux, B., Fahrenfort, J. J., Ridderinkhof, K. R., & van
Winden, F. (2015). Neural dynamics of social tie formation
in economic decision-making. Social Cognitive and Affective
Neuroscience, 10, 877–884. doi:10.1093/scan/nsu138
Brashears, M. E. (2013). Humans use compression heuristics to
improve the recall of social networks. Scientific Reports, 3, 1–7.
doi:10.1038/srep01513
Brent, L. J. (2015). Friends of friends: Are indirect connections in so-
cial networks important to animal behavior? Animal Behaviour,
103, 211–222. doi:10.1016/j.anbehav.2015.01.020
Casciaro, T. (1998). Seeing things clearly: Social structure, personal-
ity, and accuracy in social network perception. Social Networks,
20, 331–351. doi:10.1016/S0378-8733(98)00008-2
Cheney, D. L. (2011). Extent and limits of cooperation in ani-
mals. Proceedings of the National Academy of Sciences, 108,
10902–10909. doi:10.1073/pnas.1100291108
Christoffel, D. J., Golden, S. A., Walsh, J. J., Guise, K. G., Heshmati,
M., Friedman, A. K., . . . Russo, S. J. (2015). Excitatory transmission at
thalamo-striatal synapses mediates susceptibility to social stress.
Nature Neuroscience, 18, 962–964. doi:10.1038/nn.4034
De Soto, C. B.
(1960). Learning a social structure.
Journal of
Abnormal and Social Psychology, 60, 417–421. doi:10.1037/
h0047511
OPEN MIND: Discoveries in Cognitive Science
10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
/
/
.
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
/
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
De Soto, C. B., & Bosley, J. J. (1962). The cognitive structure of a
social structure. Journal of Abnormal and Social Psychology, 64,
303–307.
Dziura, S. L., & Thompson, J. C. (2018). Supplemental material for
“the neural representational space of social memory.” Open
Mind: Discoveries in Cognitive Science, 3, 1–12. doi:10.1162/
opmi_a_00021
Feng, C., Li, Z., Feng, X., Wang, L., Tian, T., & Luo, Y. J. (2016).
Social hierarchy modulates neural responses of empathy for
pain. Social Cognitive and Affective Neuroscience, 11, 485–495.
doi:10.1093/scan/nsv135
Freeman, L. C., Freeman, S. C., & Michaelson, A. G. (1988). On
Journal of Social and Biological
intelligence.
human social
Structures, 11, 415–425.
Garvert, M. M., Dolan, R. J., & Behrens, T. E. J. (2017). A map
of abstract relational knowledge in the human hippocampal-
entorhinal cortex. eLife, 6, e17086. doi:10.7554/eLife.17086
Haxby, J. V., Connolly, A. C., & Guntupalli, J. S. (2014). Decoding
neural representational spaces using multivariate pattern anal-
ysis. Annual Reviews of Neuroscience, 37, 435–456. doi:10.
1146/annurev-neuro-062012-170325
Ioannidis, A. E., Kimiskidis, V. K., Loukopoulou, E., Geroukis, T.,
Vlaikidis, N., & Kosmidis, M. H. (2013). Apathy, cognitive dys-
function and impaired social cognition in a patient with bilat-
eral thalamic infarction. Neurocase, 19, 513–520. doi:10.1080/
13554794.2012.701645
Janicik, G. A., & Larrick, R. P. (2005). Social network schemas and
the learning of incomplete networks. Journal of Personality and
Social Psychology, 88, 348–364.
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The fusiform
face area: A module in human extrastriate cortex specialized for
face perception. The Journal of Neuroscience, 17, 4302–4311.
doi:10.3410/f.717989828.793472998
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008). Representational
similarity analysis—Connecting the branches of systems neuro-
science. Frontiers in Systems Neuroscience, 2, 1–28. doi:10.3389/
neuro.06.004.2008
Kumaran, D., Melo, H. L., & Duzel, E. (2012). The emergence and
representation of knowledge about social and nonsocial hierar-
chies. Neuron, 76, 653–666. doi:10.1016/j.neuron.2012.09.035
Leavitt, H. J. (1951). Some effects of certain communication pat-
terns on group performance. The Journal of Abnormal and Social
Psychology, 46, 38–50.
Lin, N., Dayton, P. W., & Greenwald, P. (1978). Analyzing the in-
strumental use of relations in the context of social structure.
Sociological Methods & Research, 7, 149–166. doi:10.1177/
004912417800700203
Meyer, M. L., Taylor, S. E., & Lieberman, M. D. (2015). Social work-
ing memory and its distinctive link to social cognitive ability:
An fMRI study. Social Cognitive and Affective Neuroscience,
10, 1338–1347. doi:10.1093/scan/nsv065
Minear, M., & Park, D. C. (2004). A lifespan database of adult facial
stimuli. Behavior Research Methods, Instruments, & Computers,
36, 630–633. doi:10.3758/BF03206543
Okuyama, T., Kitamura, T., Roy, D. S., Itohara, S., & Tonegawa, S.
(2016). Ventral CA1 neurons store social memory. Science, 353,
1536–1541. doi:10.1126/science.aaf7003
Overwalle, F. V., Baetens, K., Marien, P., & Vandekerckhove, M.
(2014). Social cognition and the cerebellum: A meta-analysis of
over 350 fMRI studies. NeuroImage, 86, 554–572. doi:10.1016/
j.neuroimage.2013.09.033
Parkinson, C., Kleinbaum, A. M., & Wheatley, T.
(2017). Sponta-
neous neural encoding of social network position. Nature Hu-
man Behaviour, 1, 1–7.
Peirce, J. W.
(2007). PsychoPy-Psychophysics software in Python.
Journal of Neuroscience Methods, 162, 8–13.
Rüsch, N., Bado, P., Zahn, R., Bramati, I. E., De Oliveira-Souza, R.,
& Moll, J. (2014). You and your kin: Neural signatures of family-
based group perception in the subgenual cortex. Social Neuro-
science, 9, 326–331. doi:10.1080/17470919.2014.912676
Samson, D., Apperly, I. A., Chiavarino, C., & Humphreys, G. W.
(2004). Left temporoparietal junction is necessary for represent-
ing someone else’s belief. Nature Neuroscience, 7, 499–500.
doi:10.1038/nn1223
Satpute, A. B., Badre, D., & Ochsner, K. N. (2014). Distinct regions of
prefrontal cortex are associated with the controlled retrieval and
selection of social information. Cerebral Cortex, 24, 1269–1277.
doi:10.1093/cercor/bhs408
Schapiro, A. C., Kustner, L. V., & Turk-Browne, N. B. (2012). Shap-
ing of object representations in the human medial temporal lobe
based on temporal regularities. Current Biology, 22, 1622–1627.
Schapiro, A. C., Turk-Browne, N. B., Norman, K. A., & Botvinick,
M. M. (2016). Statistical learning of temporal community struc-
ture in the hippocampus. Hippocampus, 26, 3–8.
Seyfarth, R. M., & Cheney, D. L. (2012). The evolutionary origins of
friendship. Annual Review of Psychology, 63, 153–177.
Seyfarth, R. M., & Cheney, D. L. (2015). Social cognition. Animal
Behaviour, 103, 191–202. doi:10.1016/j.anbehav.2015.01.030
Stein, T., Moritz, C., Quigley, M., Cordes, D., Haughton, V., &
Meyerand, E. (2000). Functional connectivity in the thalamus and
hippocampus studied with functional MR imaging. American
Journal of Neuroradiology, 21, 1397–1401.
Tendler, A., & Wagner, S. (2015). Different types of theta rhythmicity
are induced by social and fearful stimuli in a network associated
with social memory. eLife, 4, e03614. doi:10.7554/eLife.03614
Tiddi, B., Sorrentino, E. P. D., Fischer, J., & Schino, G. (2017).
Acquisition and functional consequences of social knowledge
in macaques. Royal Society Open Science, 4, 160639. doi:10.
1098/rsos.160639
Tung, J., Barreiro, L. B., Burns, M. B., Grenier, J. C., Lynch, J.,
Greineisen, L. E., . . . Archie, E. A. (2015). Social networks predict
gut microbiome composition in wild baboons. eLife, 4, e05224.
doi:10.7554/eLife.05224
Turk-Browne, N. B., Scholl, B. J., Johnson, M. K., & Chun, M. M.
(2010). Implicit perceptual anticipation triggered by statistical
learning. The Journal of Neuroscience, 30, 11177–11187.
Wlodarski, R., & Dunbar, R. I. M. (2016). When BOLD is thicker than
water: Processing social information about kin and friends at dif-
ferent levels of the social network. Social Cognitive and Affective
Neuroscience, 11, 1952–1960. doi:10.1093/scan/nsw101
Zerubavel, N., Bearman, P. S., Weber, J., & Ochsner, K. N. (2015).
Neural mechanisms tracking popularity in real-world social
networks. Proceedings of the National Academy of Sciences,
112, 15072–15077. doi:10.1073/pnas.1511477112
OPEN MIND: Discoveries in Cognitive Science
11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
/
.
/
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
/
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural Representation of Social Memory Dziura and Thompson
Zink, C. F., Kempf, L., Hakimi, S., Rainey, C. A., Stein, J. L., & Meyer-
Lindenberg, A. (2011). Vasopressin modulates social recognition-
related activity in the left temporoparietal junction in humans.
Translational Psychiatry, 1, e3. doi:10.1038/tp.2011.2
Zink, C. F., Tong, Y., Chen, Q., Bassett, D. S., Stein, J. L., & Meyer-
Lindenberg, A. (2008). Know your place: Neural processing of
social hierarchy in humans. Neuron, 58, 273–283. doi:10.1016/
j.neuron.2008.01.025
Zitek, E. M., & Tiedens, L. Z. (2012). The fluency of social hierarchy:
The ease with which hierarchical relationships are seen, re-
membered, learned, and liked. Journal of Personality and Social
Psychology, 102, 98–115. doi:10.1037/a0025345
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
i
/
/
.
1
0
1
1
6
2
o
p
m
_
a
_
0
0
0
2
1
1
8
6
8
3
7
7
o
p
m
_
a
_
0
0
0
2
1
p
d
/
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
OPEN MIND: Discoveries in Cognitive Science
12