Sondeando los sistemas neuronales subyacentes a la flexibilidad
Dimensional Attention
Aaron T. Buss1, Vincent Magnotta2, Eliot Hazeltine2, Kaleb Kinder1, and John P. Spencer3
Abstracto
■ Flexibly shifting attention between stimulus dimensions
(p.ej., shape and color) is a central component of regulating cog-
nition for goal-based behavior. In the present report, we ex-
amine the functional roles of different cortical regions by
manipulating two demands on task switching that have been
confounded in previous studies—shifting attention between
visual dimensions and resolving conflict between stimulus–
response representations. Dimensional shifting was manipulated
by having participants shift attention between dimensions
(either shape or color; dimension shift) or keeping the task-
relevant dimension the same (dimension same). Conflict be-
tween stimulus–response representations was manipulated by
creating conflict between response-driven associations from
the previous set of trials and the stimulus–response mappings
on the current set of trials (p.ej., making a leftward response
to a red stimulus during the previous task, but being required
to make a rightward response to a red stimulus in the current
tarea; stimulus–response conflict), or eliminating conflict by alter-
ing the features of the dimension relevant to the sorting rule
(stimulus–response no-conflict). These manipulations revealed
activation along a network of frontal, temporal, parietal, and oc-
cipital cortices. Específicamente, dimensional shifting selectively acti-
vated frontal and parietal regions. Stimulus–response conflict,
por otro lado, produced decreased activation in temporal
and occipital cortices. Occipital regions demonstrated a complex
pattern of activation that was sensitive to both stimulus–
response conflict and dimensional attention switching. These re-
sults provide novel information regarding the distinct role that
frontal cortex plays in shifting dimensional attention and poste-
rior cortices play in resolving conflict at the stimulus level. ■
INTRODUCCIÓN
Controlled cognition often involves switching attention
between visual dimensions that are relevant in different
behavioral contexts. Por ejemplo, we may attend to the
shapes of objects when trying to find bananas in the gro-
cery store but then attend to the color when trying to
find ones that are ripe. In such situations, different di-
mensions (p.ej., shape and color) of the same object are
associated with different decisions, and attentional pro-
cesses must select the appropriate dimension for pro-
cessing and response selection. Previously attended
visual information must be ignored, and previously ig-
nored visual information must now be attended. En esto
way, dimension switching is unique from other types of
switching. Task or response switching only requires alter-
nating between different stimulus–response mappings or
assigning new responses to the same stimuli (Crone,
Wendelken, Donohue, & Bunge, 2006). De este modo, estos
other forms of switching do not require early selection
at the level of the stimulus dimension. In this report, nosotros
use fMRI with adult participants to examine the neural basis
of this form of task switching using a novel implementation
1University of Tennessee, Knoxville, 2University of Iowa,
3University of East Anglia, Reino Unido
© 2021 Instituto de Tecnología de Massachusetts
of a dimensional attention task that has been extensively
used with children.
The Processes Underlying Task Switching
The ability to update behavior is often studied using a task
switching paradigm (p.ej., Kiesel et al., 2010). In this con-
texto, a task consists of a set of stimulus–response map-
pings indicating the motor actions (p.ej., pressing a
button) that should be made in response to a set of stim-
uli. Switch costs are typically observed in the form of lon-
ger RTs when switching to a new task compared to
repeating the same task. These switch costs are typically
thought to reflect the additional time needed to activate
the new task in working memory. Además, the rela-
tionship between the two tasks can impact the magnitude
of switch costs. Por ejemplo, if the tasks pair different re-
sponses with the same stimuli, then larger switch costs are
observed compared to switching between two tasks that
use different sets of stimuli. Stimuli that have response
mappings for two tasks are bivalent and impose additional
inhibitory demands because of the stimuli being mapped
to multiple response options.
Task switching can also involve shifting attention to vi-
sual features of different dimensions of the same set of
estímulos. Por ejemplo, one task may instruct responses
Revista de neurociencia cognitiva 33:7, páginas. 1365–1380
https://doi.org/10.1162/jocn_a_01720
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
to the colors of a set of objects and the other task may
instruct responses to the shapes of the same set of ob-
jects. En este caso, the stimuli are bivalent because they
are mapped to two sets of responses, but also require
dimensional attention to select the information along
the dimension that is relevant for the current task.
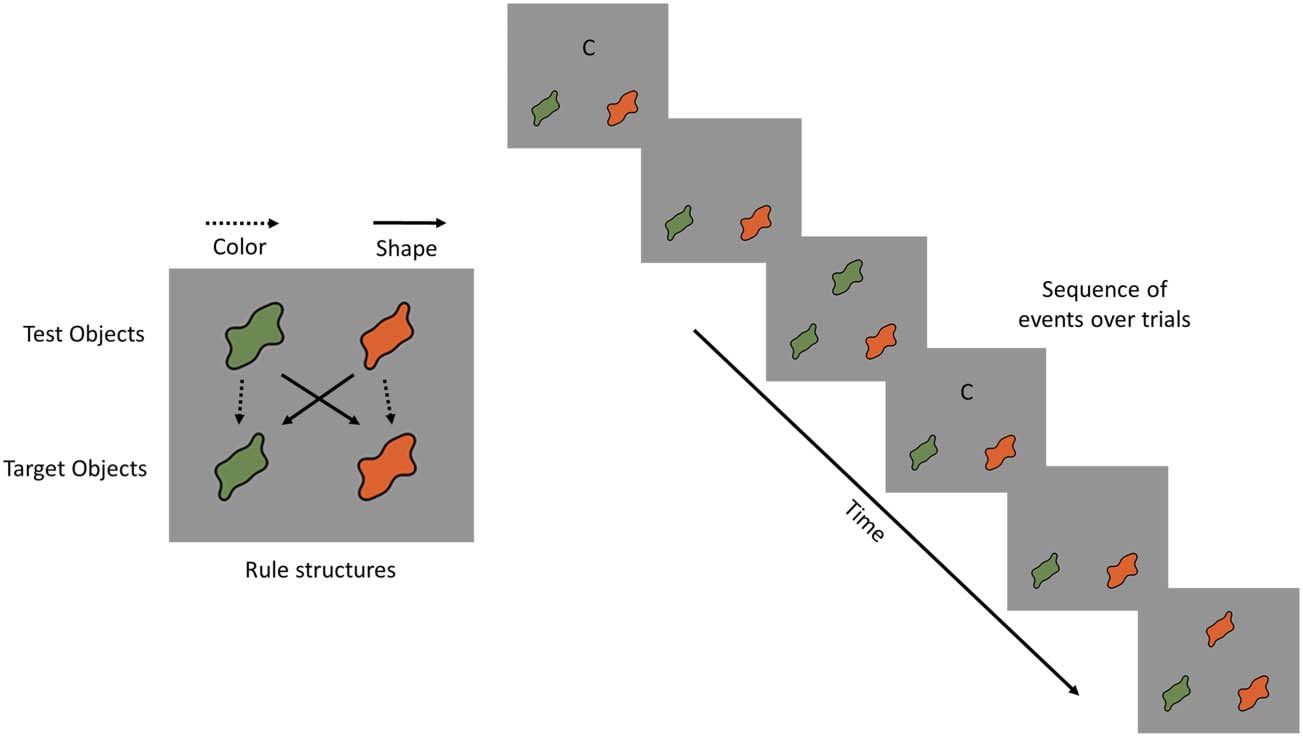
Consider the dimensional change card sort (DCCS) tarea
como se muestra en la figura 1. The DCCS task explicitly cues the
need to shift dimensional attention by instructing partic-
ipants to match objects based on shape or color. Target
objects and arrow types in the left panel of Figure 1 espectáculo
the response mappings for the shape or color rules for
each test object. Por ejemplo, the wavy green test object
in the upper left corner of this panel should be matched
to the left green target object in the color game (see dot-
ted arrow), whereas this same test object should be
matched to the right wavy orange target object in the
shape game (see solid arrow). Note that the to-be-sorted
test objects match either target object along different di-
mensions, creating visual conflict and forcing the selec-
tion of the relevant dimension. The right panel of
Cifra 1 shows how the task unfolds over trials. Primero, a
dimensional cue is presented (see “c” for the color game)
along with the two target objects in the lower portion of
the display. Próximo, the dimensional cue disappears, y un
test object is presented centrally in the upper panel. El
participant must then press a button to indicate the tar-
get object to which the test object should be sorted—the
left green target object in this case. This is followed by
the next trial that starts with the presentation of either
the same or different target cards and/or the same or a
different dimensional cue. Switch costs are typically
observed in the form of longer RTs when shifting dimen-
siones (p.ej., a color sorting trial preceded by a shape sorting
trial) compared to repeating dimensions (p.ej., a color
sorting trial preceded by a color sorting trial; Morton,
Bosma, & Ansari, 2009; Diamante & Kirkham, 2005).
Although the DCCS task has only been used to a lim-
ited degree with adults, literature on executive function
development sheds light on the unique processes that
underlie dimensional attention switching in this task.
The DCCS task is popular in the developmental literature
because it reveals a qualitative improvement in perfor-
mance: 3-year-olds predominantly fail to switch rules,
but most 4-year-olds have little difficulty switching
(Zelazo et al., 2003). De este modo, this task is often used as a
measure of the developmental status of executive func-
ción. The widespread use of this task with children has
resulted in numerous variations, some of which improve
switching in young children. De este modo, the pattern of success
or failure with children indicates which factors of the task
create challenges to dimensional attention switching.
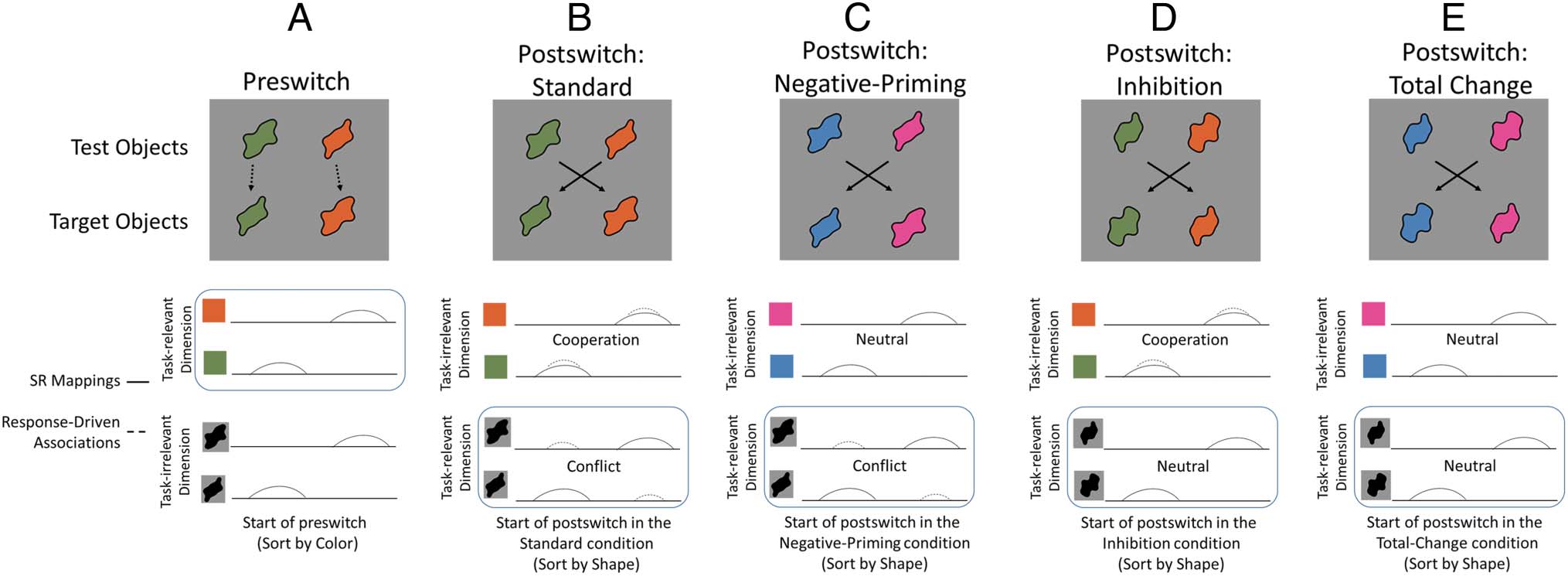
Two characteristics of the stimulus–response represen-
tations that impact dimensional switching in this task are
illustrated in Figure 2. The top shows the configuration of
test objects and target objects with lines showing the
test-to-target mappings as in Figure 1. Notice that the vi-
sual structure of the task primes conflicting responses for
each test object. Por ejemplo, the wide-green test object
matches the leftward target object by color but matches
the rightward target object by shape. This visual conflict
matters: If target objects are not used in the task and
rules are simply provided verbally, then young children
have little difficulty switching (Towse, Redbond,
Houston-Price, & Cocinar, 2000). The primary challenge
of the DCCS is not task switching itself, but the need
to use dimensional attention to map the features of the
test objects to the features on the target objects.
To illustrate the dynamics of neural representations of
the features involved in this task, we use the schematic
representation in the lower panels of Figure 2. The solid
line bumps in the bottom shows the “activation” in
feature-encoding neural populations created by viewing
the target objects in particular spatial positions. De este modo,
the green target object on the left generates a bump of
activation on the left side of the color representation at
the green value, and on the left side of the shape repre-
sentation at the “less wavy” spatial frequency value. Por
Cifra 1. Left: Depiction of the
standard DCCS task. Target
objects show which features
go to which response location
for the shape or color rules.
Test objects match either
target object along different
dimensions. Right: Depiction
of sequence of events over
two trials.
1366
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. Depiction of stimulus–response representations across different variations of the DCCS task (see also Table 1). Lines illustrate the strength
of input being contributed by the stimulus–response (SR) mappings (solid) and response-driven associations (dashed). In this example, color is the
preswitch dimension and shape is the postswitch dimension.
contrast, the orange target object on the right generates a
bump on the right side of the color representation at the
orange value, and on the right side of the shape repre-
sentation at the “more wavy” spatial frequency.
The dashed bumps in the lower panels of Figure 2 gorra-
ture the second important characteristic of stimulus–
response representations in this task—response-driven
associations that build up over trials (dashed lines).
Figure 2A shows a preswitch phase in which the rules
are to sort by color. Note that there are no response-
driven association bumps in the lower stimulus–response
representations because there is no prior history with the
tarea. Over trials (moving from Figure 2A to 2B), stimulus–
response associations form between the responses and
the features of both the relevant and irrelevant dimen-
sions of the test objects. En particular, Figure 2B shows
the status of stimulus–response representations at the
start of the postswitch phase in the standard DCCS task.
Because the “more wavy” green test object was sorted to
the left in the preswitch phase (see Figure 2A), there is a
dashed activation bump on the left at the green feature
value and also a dashed bump on the left at the “more
wavy” spatial frequency. Similarmente, there is a dashed bump
on the right at the orange feature value and a dashed
bump on the right at the “less wavy” spatial frequency.
Notice that this leads to a pattern of “cooperation” in
the color-response representation because the solid and
dashed bumps overlap, and a pattern of “conflict” in the
shape–response representation because the solid and
dashed overlap with opposite response. In the context
of dimensional switching, this means that the preswitch
dimension—color—is primed by the cooperation whereas
the postswitch dimension—shape—experiences inhibitory
competition based on this conflict.
Evidence that these response-driven associations im-
pact performance in the DCCS task comes from multiple
estudios (ver tabla 1 for a summary). In a negative prim-
ing version of the task (Figura 2C), the features that were
relevant during the preswitch phase (p.ej., Figura 2A) son
altered before the start of the postswitch phase. Para en-
postura, in Figure 2C, the features of the color dimension
are changed in both the target and test objects.
Como consecuencia, response-driven associations are no lon-
ger present in the color-response representation (nota
the absence of dashed lines in the “neutral” color repre-
sentation of Figure 2C). In this condition, the majority of
young children still perseverate (Buss & Spencer, 2014;
Müller, Dick, Gela, Overton, & Zelazo, 2006; Zelazo
et al., 2003) because of the conflict that remains within
the shape–response representation (see Figure 2C).
De este modo, even though the stimulus–response mapping that
was relevant during the preswitch phase is no longer rel-
evant because those features are now absent from the
estímulos, young children still fail to switch rules. Sobre el
other hand, children also perseverate in an inhibition ver-
sión (Zelazo et al., 2003) of the task in which the features
of the dimension that is being switched to are altered be-
fore the start of the postswitch phase (see Figure 2D). En
Mesa 1. Summary of Conditions Used with Children in the
DCCS Task and the Task Factors That Have Been Manipulated
Irrelevant
Dimension
Relevant
Dimension
3-year-old
Comportamiento
Neutral
Competition Perseverate
Cooperation Neutral
Perseverate
Negative-
priming
Inhibition
Total change Neutral
Neutral
Switch
No-conflict
Cooperación
Cooperación
Switch
The bold and underlined factors in the table highlight the factors that
drive perseveration in 3-year-olds.
Buss et al.
1367
este caso, the “cooperative” stimulus–response associa-
tions that remain along the features of the preswitch di-
mension (es decir., color in Figure 2D) interfere with the
ability to use the features of the postswitch dimension
(es decir., forma) to make sorting decisions.
En tono rimbombante, the majority of young children can switch
rules in a total-change version (Zelazo et al., 2003) en
which the features of both dimensions are changed before
the start of the postswitch phase (see Figure 2E). Nota
that the dashed bumps reflecting the learned stimulus–
response associations from the preswitch phase are absent
(“neutral”) for the features of both dimensions. In the ab-
sence of any influence of stimulus–response associations
from the previous sorting phase, young children have little
difficulty switching rules. Juntos, these results indicate
that the object features that are irrelevant for the current
sorting rules are nonetheless bound to responses, y el
status of stimulus–response representations across both
the relevant and irrelevant dimensions impacts the ability
to shift dimensional attention.
En resumen, the DCCS presents unique challenges in
the context of dimensional task switching. The task re-
quires dimensional attention, which is an additional level
of visual processing beyond the stimulus identification re-
quired in other response selection tasks. Además, el
binding of features across stimulus dimensions leads to
the formation of stimulus–response associations that also
includes task-irrelevant features. Beyond early childhood,
sin embargo, it is unclear whether such bindings occur or
how they might influence dimensional attention.
Neural Basis of Task Switching
Neuroimaging research has identified a network of fron-
tal and parietal regions that are engaged in response to
demands on task switching. This network includes dorso-
lateral prefrontal cortex, pre-SMA, dorsal premotor cor-
tex, inferior frontal junction, anterior insula cortex, y
posterior parietal cortex (Col & Schneider, 2007).
Además, some regions are sensitive to the relationship
between tasks. SMA and pre-SMA are more strongly acti-
vated for bivalent compared to univalent rules, and ven-
trolateral prefrontal cortex is more strongly activated
when switching to bivalent rules (Crone et al., 2006).
Data from studies using the DCCS task have identified
a similar network composed of inferior and dorsal frontal
corteza, parietal cortex, and thalamus that is more strongly
activated when switching dimensions compared to re-
peating the same dimension (Morton et al., 2009). Este
research also suggests that distinct neural mechanisms
may be involved in dimensional attention shifting.
Específicamente, switch-related activity has been identified in
fusiform cortex, a region not previously implicated in task
switching (Morton et al., 2009). Fusiform cortex is in-
volved in object representation processes and is modu-
lated by attentional signals that prioritize processing of
specific perceptual information (Tallon-Baudry, Bertrand,
Hénaff, Isnard, & pescador, 2005; Liu, Slotnick, serencias,
& Yantis, 2003; Clark et al., 1997). Por último, electrophysiolog-
ical data have demonstrated dissociable electrophysiologi-
cal signatures of processing demands in the DCCS.
Específicamente, central frontal N2 amplitude is greater for bi-
valent compared to univalent test objects, whereas frontal
negativity was associated with the magnitude of costs
when switching dimensions (Waxer & Morton, 2011).
Developmental neuroimaging studies using the DCCS
converge with these patterns of results for task switching
and dimensional attention switching. Children demon-
strate increased activation within frontal, parietal, y
temporal cortex as the ability to switch rules develops
(Buss & Spencer, 2018; Moriguchi & Hiraki, 2009). En
addition, this literature sheds light on the influence of
conflict between learned stimulus–response associations
and stimulus–response mappings for a given task.
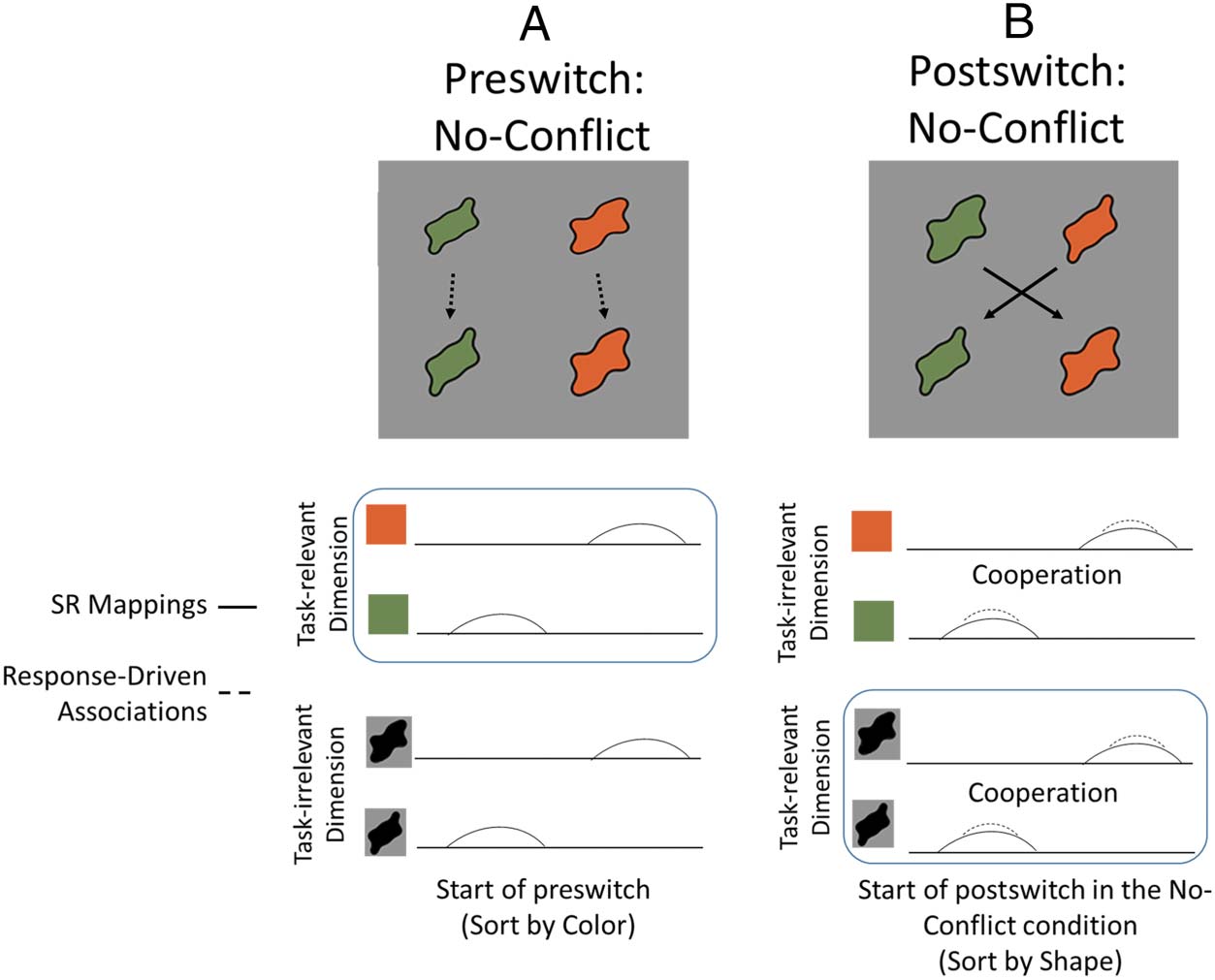
Específicamente, this research examined neural activation dur-
ing a no-conflict negative priming version of the DCCS
illustrated in Figure 3. As shown in Figure 3A, the pre-
switch phase of the task is modified so that the test cards
match the target cards along both dimensions. Eso es, a
“less wavy” green test object is sorted to a “less wavy”
green target object and the “more wavy” orange test ob-
ject is sorted to a “more wavy” orange target object.
Standard test objects that contain visual conflict are used
during the postswitch phase. The primary influence of
this manipulation is to create stimulus–response associa-
tions that cooperate with the stimulus–response map-
pings required for the postswitch phase. In the absence
of conflict, stronger activation is observed in parietal and
temporal regions (Buss & Spencer, 2018) suggesting that
posterior object representation areas are sensitive to the
conflict between response-driven associations from the
previous task and stimulus–response mappings for a cur-
rent task.
Current Study
Developmental studies have revealed that stimulus–
response conflict (CAROLINA DEL SUR) plays a distinctive role in chil-
dren’s ability to switch rules in dimensional attention
tareas. Both the relevant and irrelevant features matter,
suggesting that rule-use is not selectively focused on
the task-relevant dimension but instead integrates across
all object features. In previous fMRI studies that have
used the standard DCCS task, conflict at the level of
dimensional attention created by switching from one
dimensional rule to another is confounded with SC.
Given that recent neuroimaging results with children
demonstrate that switching dimensions and resolving
SC have different influences on the functioning of frontal
and posterior brain regions, our goal in this study was to
clarify the roles of different brain regions in shifting di-
mensional attention.
1368
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3. Depiction of
stimulus–response
representations in the no-
conflict version of the DCCS.
The goal of the current study is to independently
manipulate the status of stimulus–response representa-
tions and dimensional shifting while collecting functional
MRI with adults. As discussed above, stimulus–response
representations in the context of the DCCS task are im-
portant to consider along both the task-relevant and task-
irrelevant dimensions. De este modo, to manipulate the status of
stimulus–response representations, both aspects must be
considered and controlled. en este estudio, we focus on SC
with regard to the features that are relevant for the cur-
rent phase of sorting; stimulus–response representations
involving the features that are irrelevant for the current
phase of sorting was held constant across conditions. En
particular, the stimulus–response associations always
cooperated with the target stimuli features within the
task-irrelevant dimension (es decir., the location of the task-
irrelevant features on the target objects always overlap-
ped with the locations where those features were sorted
during the previous phase as illustrated along the color
dimension in Figures 2B). Note that this is a property
of the standard DCCS task.
To manipulate SC along the task-relevant dimension, nosotros
either kept the same features between the pre- and post-
switch phases or we changed the features along the post-
switch dimension. If the features were not altered, entonces
the current stimulus–response mappings conflicted with
the stimulus–response associations from the previous task.
Eso es, participants had to make responses to the task-
relevant features that were the opposite of the responses
made to those features in the previous phase (CAROLINA DEL SUR), as in
the standard DCCS task (illustrated along the shape dimen-
sion in Figure 2B). If the relevant features changed, entonces
there were no stimulus–response associations from the pre-
vious phase for those features and there was no SC along
the relevant dimension (stimulus–response no-conflict
[SN]), as in the inhibition version (illustrated along the
shape dimension in Figure 2D).
We also manipulated the need to shift attention be-
tween visual dimensions. Específicamente, conditions either
required participants to shift attention to the other di-
mension (dimension change [corriente continua]) or to reverse the
stimulus–response mapping for features within the same
dimension (dimension same [DS]). When shifting dimen-
siones, participants were instructed to shift from sorting
by shape to sorting by color, or vice versa. If participants
were to reverse the stimulus–response mappings, entonces
participants were cued to continue sorting by the same
dimension, but the target objects would swap locations,
indicating a set of stimulus–response mappings. De este modo, en
ambos casos, there was a change that required a new map-
ping of the stimuli to response locations.
These manipulations created four conditions (ver
Mesa 2): dimension change/stimulus–response conflict
(DCSC), dimension change/stimulus–response no-
conflicto (DCSN), dimension same/stimulus–response
conflicto (DSSC), and dimension same/stimulus–response
no-conflict (DSSN). The DCSC condition is the standard
condition from the literature (Morton et al., 2009; Zelazo
Mesa 2. Factors Manipulated across Conditions in the
Behavioral Task
Stimulus-Response
Conflicto
Stimulus-Response
No-Conflict
Dimension Change
Dimension Same
DCSC
DSSC
DCSN
DSSN
Note that switch and repeat trials were included for all conditions.
Buss et al.
1369
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
et al., 2003). The DCSN is the inhibition condition from
the literature (Zelazo et al., 2003). The DSSC and DSSN
are novel conditions that have not been used in previous
estudios. Previous fMRI research has only examined the
DCSC condition.
Our task was administered in an alternating runs de-
sign in which participants were given three trials (TR1,
TR2, and TR3) for each condition before transitioning
to the next condition. Based on previous studies, nosotros
can outline a set of predictions regarding the pattern
of activation across different conditions. Primero, we expect
switch trials (the first of the three trials) to elicit stronger
activation relative to repeat trials (the subsequent two
ensayos) in frontal and parietal regions previously implicated
in dimensional attention as well as object representa-
tion regions in temporal cortex (Buss & Spencer, 2018;
Morton et al., 2009). Segundo, we expect stronger activa-
tion in frontal and parietal regions for DC conditions
compared to DS conditions. Tercero, based on the hemo-
dynamic data reported by Buss and Spencer (2018), nosotros
expect stronger activation in parietal and temporal corti-
ces when SC is absent compared to when conflict is pres-
ent along the task-relevant dimension. En resumen, nosotros
expect manipulations of the demands on shifting atten-
tion to a new dimension to produce activation differ-
ences in frontal and parietal cortices and manipulations
of task-relevant feature conflict to produce activation
differences in object representation areas in temporal
and parietal cortices.
MÉTODOS
Participantes
Twenty adults (Mage = 23.8 años, DE = 3.8 años; nueve
hombres) were enrolled into this study after providing in-
formed written consent in accordance with the local in-
stitutional review board. Monetary compensation was
provided to the subjects for their participation. All partic-
ipants were right-handed and reported normal or
corrected-to-normal vision. The data were collected as
part of a larger six-session fMRI study. The DCCS sessions
presented here were collected over four half-hour ses-
siones. Each session included two runs that were com-
posed of 123 ensayos, which lasted for 12 min each.
Behavioral Procedure
The study used a continuous event-related design with
interleaved phases, similar to other task switching para-
digms that use alternating runs (p.ej., Rogers & monsell,
1995). Participants were not informed about this aspect
of the task. Participants were instructed to match the ob-
ject presented at the top of the screen to responses indi-
cated by the location of target objects at the bottom of
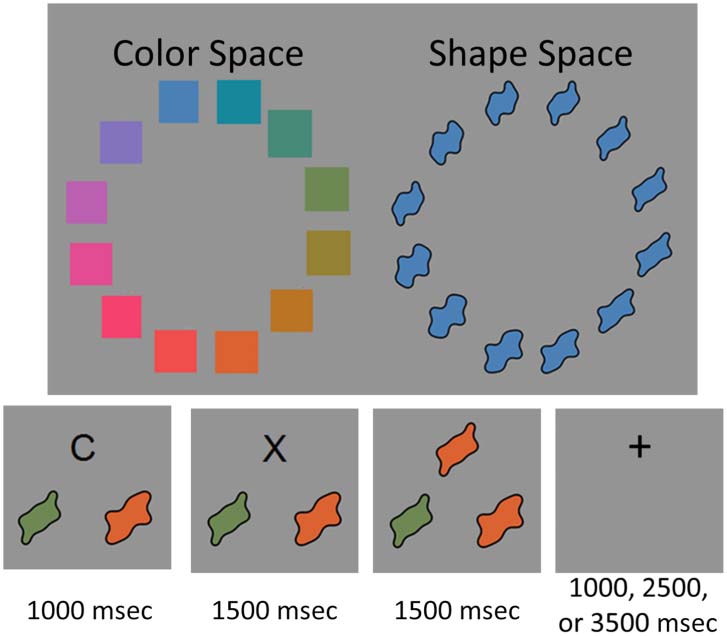
the screen. A dimensional cue of the letter “C” or “S”
was presented at the start of each trial to indicate which
dimension was relevant for the upcoming trial. Colors
were sampled from CIE Lab color space (ver figura 4).
Shape stimuli were generated from a continuous space
defined by the phase angle of Fourier components
(Drucker & Aguirre, 2009; Zahn & Roskies, 1972; ver
Cifra 4). This allows the metric details of shape to be
controlled in a similar fashion as color. The objects used
in the task were approximately 60 × 60 pixels and con-
sisted of a black outline filled with color.
The bottom of Figure 4 shows the sequence of events
on a given trial. Each trial began with an “S” or “C” pre-
sented in the upper center of the screen to indicate whe-
ther to match by shape or color on the upcoming trial, y
the target objects were presented to the left and right on
the lower portion of the screen. Después 1000 mseg, the di-
mensional cue was removed and replaced with an “X.”
Después 1500 mseg, the “X” was replaced with a test object.
Target object and test objects were approximately 2° of vi-
sual angle. Participants were given 1500 msec to press a
button with their right index or middle finger, mapping
onto a leftward or rightward sorting response. If a response
was not produced during this time window, a warning
appeared on the screen telling the participant to respond
more quickly. The duration between the end of a trial and
the beginning of a subsequent trial was jittered at 1500,
3000, o 5000 msec in a 2:1:1 ratio, respectivamente.
Cifra 5 illustrates the different conditions used in the
experiment by showing an example of the stimulus pre-
sentation phase across a sequence of trials (see also
Mesa 2) and the different task factors present for each
condición. In the bottom, task inputs are plotted in solid
lines and correspond to the locations of inputs in the tar-
get images. Memories accumulated from the previous
phase of sorting are plotted in dashed lines. The features
that are relevant for each phase of sorting are outlined in
the box. Each panel in Figure 5 illustrates an example of
stimuli used for a sorting phase, each of which consisted
of three trials. The first panel shows stimuli presented
during the first three-trial phase of sorting. This is called
Cifra 4. Stimuli and sequence of events in a trial used in the
behavioral task.
1370
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
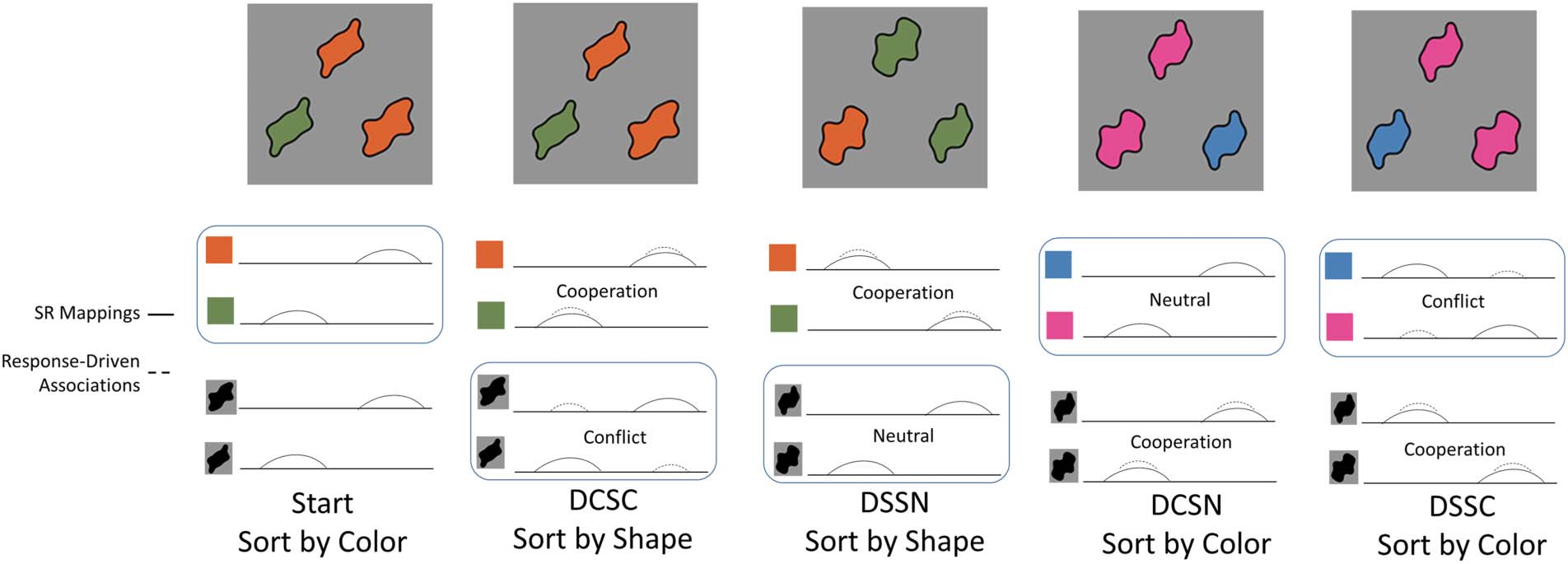
Cifra 5. Example sequence of stimuli to illustrate the conditions used in the behavioral task. See Table 2 for details on factors manipulated across
condiciones. Full sequence was Start (color), DCSC (forma), DSSN (forma), DCSN (color), DSSC (color), DCSN (forma), DSSC (forma), DCSC (color),
DSSN (color), DCSN (forma), DSSC (forma), DCSN (color), DSSC (color), DCSC (forma), Etcétera.
the Start phase. In this phase, there is no previous task
and no memories from a previous sorting phase. El
next panel shows the DCSC condition. Aquí, the relevant
dimension switches from color to shape, the features re-
main the same, and the spatial orientation of the target
cards remains the same. De este modo, the DCSC condition
requires participants to shift attention to a new dimen-
sion and resolve SC between stimulus–response associa-
tions from previous sorting decisions and the current
stimulus–response mapping. The next panel shows the
DSSN condition. Aquí, the relevant dimension stays with
forma, the features for the relevant dimension (shape in
this example) change to new feature values, and the tar-
get objects swap locations. De este modo, in the DSSN condition,
participants need to establish a new stimulus–response
mapping of the task within the dimension that was pre-
viously relevant but do not have to overcome SC. El
next panel shows the DCSN condition. Aquí, the relevant
dimension switches (in this example from shape to color),
the features that are relevant for this phase (color) son
changed to new values, and the spatial orientation of the
target cards remain the same. De este modo, the DCSN condition
requires participant to shift attention between dimensions
but does not require resolving SC. Finalmente, the last panel
illustrates the DSSC condition. In this condition, the rele-
vant dimension stays the same (color in this example), el
features in the task remain the same, and the target images
swap spatial locations. Aquí, participants do not need to
shift attention to a new dimension but must resolve SC
by reversing the stimulus–response mappings that were
used during the previous phase.
Participants completed 10 phases of each condition
(counterbalancing shape and color) in each of four total
carreras, completing a total of 40 phases for each condition.
The order of conditions was the same for every run, pero
the dimensions were the opposite from one run to the
next. Features were randomly selected from the array at
the beginning of each phase. Each set of features used
during any phase were six steps apart. When features
changed, they shifted either three or four steps in the
feature space (this was randomly selected each time
the features changed).
fMRI Data Collection and Analysis
The fMRI study used a 3T Siemens TIM Trio system with
a 12-channel head coil. Anatomical T1-weighted volumes
were collected using and magnetization prepared rapid
gradient echo sequence. Functional BOLD imaging was
acquired using an axial 2-D echo-planar gradient echo
sequence with the following parameters: echo time =
30 mseg, repetition time = 2000 mseg, flip angle = 70°,
campo de visión = 240 × 240 mm, matrix = 64 × 64, slice
thickness/gap = 4.0/1.0 mm, and bandwidth = 1920 Hz/
píxel. Preprocessing and statistical analyses were con-
ducted in AFNI version 17.3.07 (Cox, 1996). Standard pre-
processing was used that included slice timing correction,
outlier removal, motion correction, and spatial smoothing
(Gaussian FWHM = 5 mm). Data were transformed into
Montreal Neurological Institute space using a nonlinear
transform to warp the data to the common coordinate sys-
tema. The T1-weighted images were used to define the
transformation to the common coordinate system. Primero-
level analysis was performed using multiple linear regres-
sión, which included regressors for trial repetition (TR1,
TR2, TR3), dimension (same/change), and SC (conflict/
no-conflict), as well as motion parameters and baseline
parámetros (polort = 6) as nuisance regressors.
A 3 × 2 × 2 ANOVA was conducted to contrast effects
of switching dimensions (Dimension: change/same), el
effects of SC (CAROLINA DEL SUR: conflict/no-conflict), and switching
versus repeat trials (Trial Repetition: TR1, TR2, TR3).
The ANOVA was corrected for multiple comparisons using
3dClustSim and an estimated autocorrelation function to
control for false-positive rates (NN2, voxelwise p = .01,
and alpha =.01; Cox, Chen, Glen, Reynolds, & taylor,
Buss et al.
1371
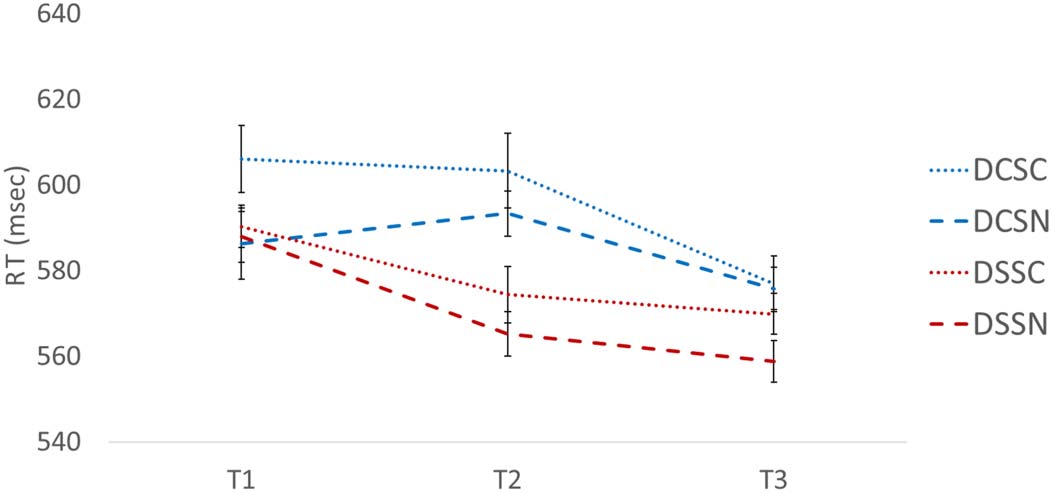
Cifra 6. RTs for the three
trials in a sorting phase. Error
bars represent within-subject
error based on Cousineau
(2005).
2017). The minimum volume for a cluster was 2672 mm3
(334 vóxeles).
RESULTADOS
Behavioral Results
Cifra 6 plots the average RTs across the three trials of
each condition. A 2 (Dimension: cambiar, mismo) × 2
(CAROLINA DEL SUR: conflicto, no-conflict) × 3 (Trial: TR1, TR2, TR3)
ANOVA revealed a main effect of Dimension, F(1, 19) =
20.5, pag < .001, ηp
2 = .519, with participants performing
slower when the dimension changed (M = 590 msec)
compared to when the dimension remained the same
(M = 574 msec). The ANOVA also revealed a main effect
of SC, F(1, 19) = 5.4, p = .032, ηp
2 = .220, with responses
being faster when conflict was absent (M = 578 msec)
compared to when conflict was present (M = 587 msec).
The ANOVA also revealed a main effect of Trial Repetition,
F(2, 18) = 8.8, p = .001, ηp
2 = .316. A simple effects test
revealed that Trial 1 RT (M = 593 msec) was not different
from Trial 2 RT (M = 584 msec; p = .526), but was signif-
icantly slower than Trial 3 RT (M = 570 msec; t(18) =
4.083, p < .001). In addition, Trial 2 RT was significantly
slower than Trial 3 RT, t(18) = 3.130, p = .006. Thus, par-
ticipants performed more quickly over the series of three
trials in each block.
Finally, the ANOVA revealed an interaction between
Dimension and Trial Repetition, F(2, 18) = 3.7, p =
.035, ηp
2 = .161. Simple effects tests on the dimension-
change conditions revealed that Trials 1 (M = 596 msec)
and 2 (M = 598 msec) were not different from one another
( p = 1.00), but Trial 3 (M = 576 msec) was significantly
faster than both Trial 1 ( p = .034) and Trial 2 ( p =
.006). When the dimension remained the same, Trial 1
(M = 589 msec) was marginally slower than Trial 2 (M =
570 msec; p = .056) and significantly slower than Trial 3
(M = 564 msec; p = .001). Trial 2 was not different from
Trial 3 ( p = 1.00). Thus, switch costs persisted for two
trials when the dimension changed, but switch costs were
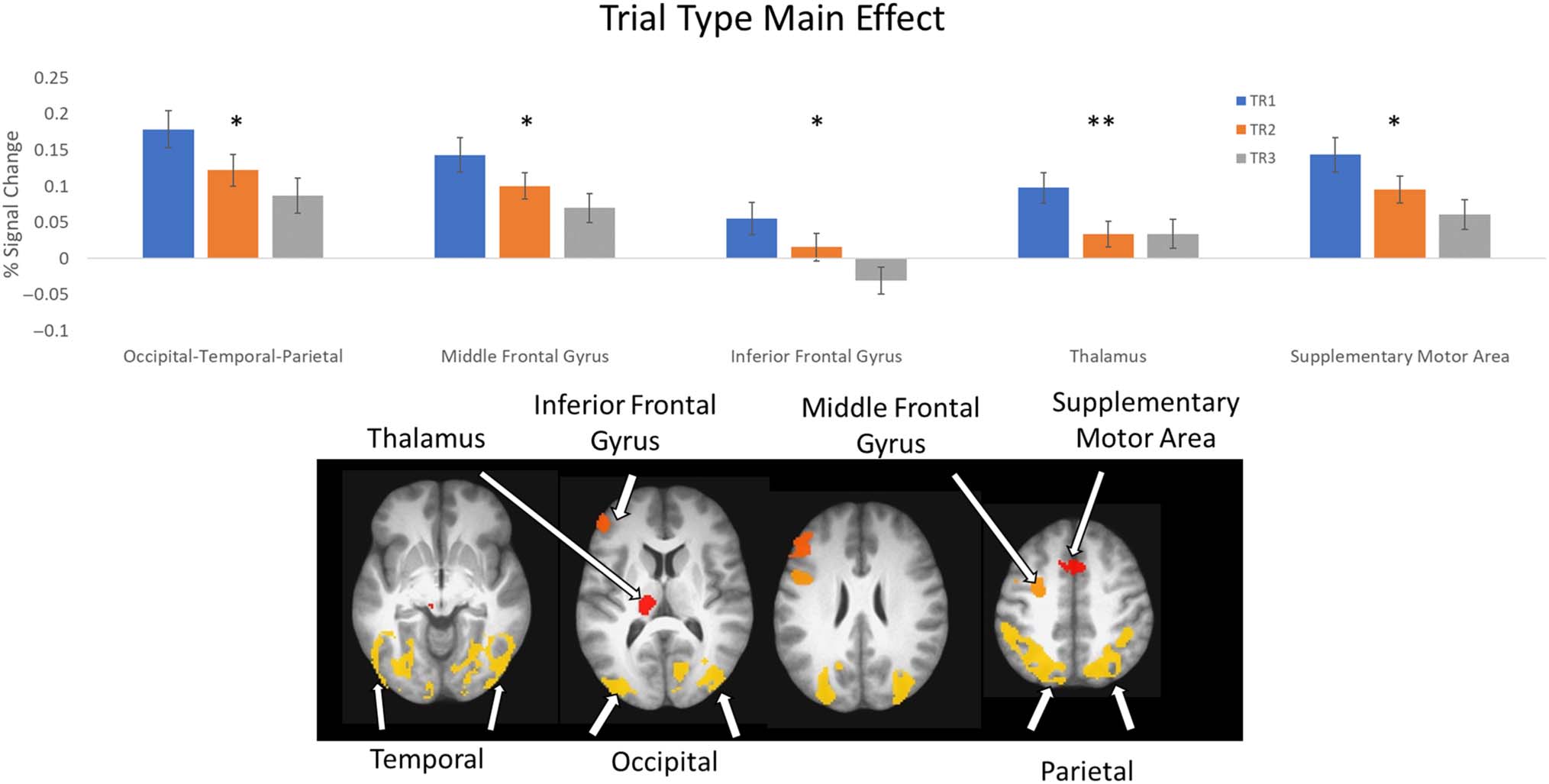
Figure 7. Trial Repetition main effect. Top: average percent signal change within ROIs showing a main effect of Trial Repetition (* indicates TR1 > TR2 >
TR3; ** indicates TR1 > TR2 only). Error bars represent standard error of the mean. Bottom: locations of ROIs. See Table 3 for list of ROIs.
1372
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Mesa 3. Clusters of Activation Revealed by the ANOVA
MNI Coordinates (RAI; mm)
Region
Hemi
Trial Repetition Main Effect
Occipital–temporal–parietal
Middle frontal gyrus
Inferior frontal gyrus
Thalamus
SMA
Dimension Main Effect
Inferior parietal cortex
SMA
Inferior frontal gyrus
Inferior frontal gyrus
Insula lobe
Superior frontal gyrus
Intraparietal sulcus
Feature-Conflict Main Effect
Fusiform gyrus
B
l
l
l
B
l
B
l
R
l
l
R
l
Dimension × Trial Repetition Interaction
SMA
Postcentral gyrus
Precuneus
Intraparietal sulcus
B
l
l
l
X
37
29
51
19
−5
33
53
3
−35
29
11
−17
29
9
37
5
37
Dimension × Feature-Conflict Interaction
Inferior occipital gyrus
Angular gyrus
l
R
43
−29
Dimension × Feature-Conflict × Trial-Repetition Interaction
Fusiform gyrus
Middle occipital gyrus
Fusiform gyrus
R
l
l
−29
23
35
MNI = Montreal Neurological Institute; RAI = right anterior inferior.
y
55
1
−49
31
−13
53
−33
−13
−29
−21
−45
65
39
−7
31
73
51
83
57
47
95
53
z
Volumen (mm3)
Effect Size
−21
51
9
−1
49
43
25
51
−5
11
39
43
92,904
8,120
5,656
3,400
2,400
15,008
7,504
6,720
4,736
3,752
3,504
2,992
.0397
.0377
.0392
.0512
.0490
.0306
.0306
.0390
.0416
.0307
.0432
.0274
−15
4,048
.0260
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
59
61
47
41
−5
−11
−19
−3
−17
8,016
5,480
2,944
2,776
12,608
3,592
9,968
4,024
3,688
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
.0175
.0317
.0208
.0135
.0158
.0223
.0184
.0128
.0163
resolved after a single trial when the dimension remained
lo mismo. Recall that, on our DS trials, participants still per-
formed a task switch that involved updating the response
mapping for the relevant features. No other significant in-
teractions were observed. Comparing between levels of
dimension, Trial 1 RT was not different for the DC (m =
596) and DS (m = 589) condiciones ( pag = .324), but was
different on Trial 2 (m = 598 vs. m = 569, respectivamente;
pag < .001) and Trial 3 (M = 576 vs. M = 564, respectively;
p = .044). Thus, performance was not different on the
Buss et al.
1373
initial trial of conditions in which the dimensions changed
or remained the same, but differences emerged over the
repetition trials.
fMRI Results
We analyzed the fMRI data with a 3 (Trial Repetition:
TR1, TR2, TR3) × 2 (Dimension: change vs. same) × 2
(SC: conflict vs. no-conflict) ANOVA. We performed hierar-
chical clustering so that voxels that were involved in
higher-level interactions were ignored when examining
lower-level interactions or main effects. Figure 7 shows
results for clusters with a significant main effect of Trial
Repetition. Note that this is the first direct comparison of
switch versus repeat trials in this task; previous studies
(Ezekiel, Bosma, & Morton, 2013; Morton et al., 2009)
performed a contrast between switch blocks and repeat
blocks. Consistent with these previous attention switch-
ing studies, stronger activation was observed on switch
trials compared to repeat trials for all clusters. A large
cluster was observed that spanned from bilateral fusiform
cortex through inferior temporal, occipital, and parietal
regions. Additional localized clusters were observed in
middle and inferior frontal gyri, thalamus, and SMA
(see Table 3 for details). Except for the thalamus, these
regions showed decreases in activation across all TRs
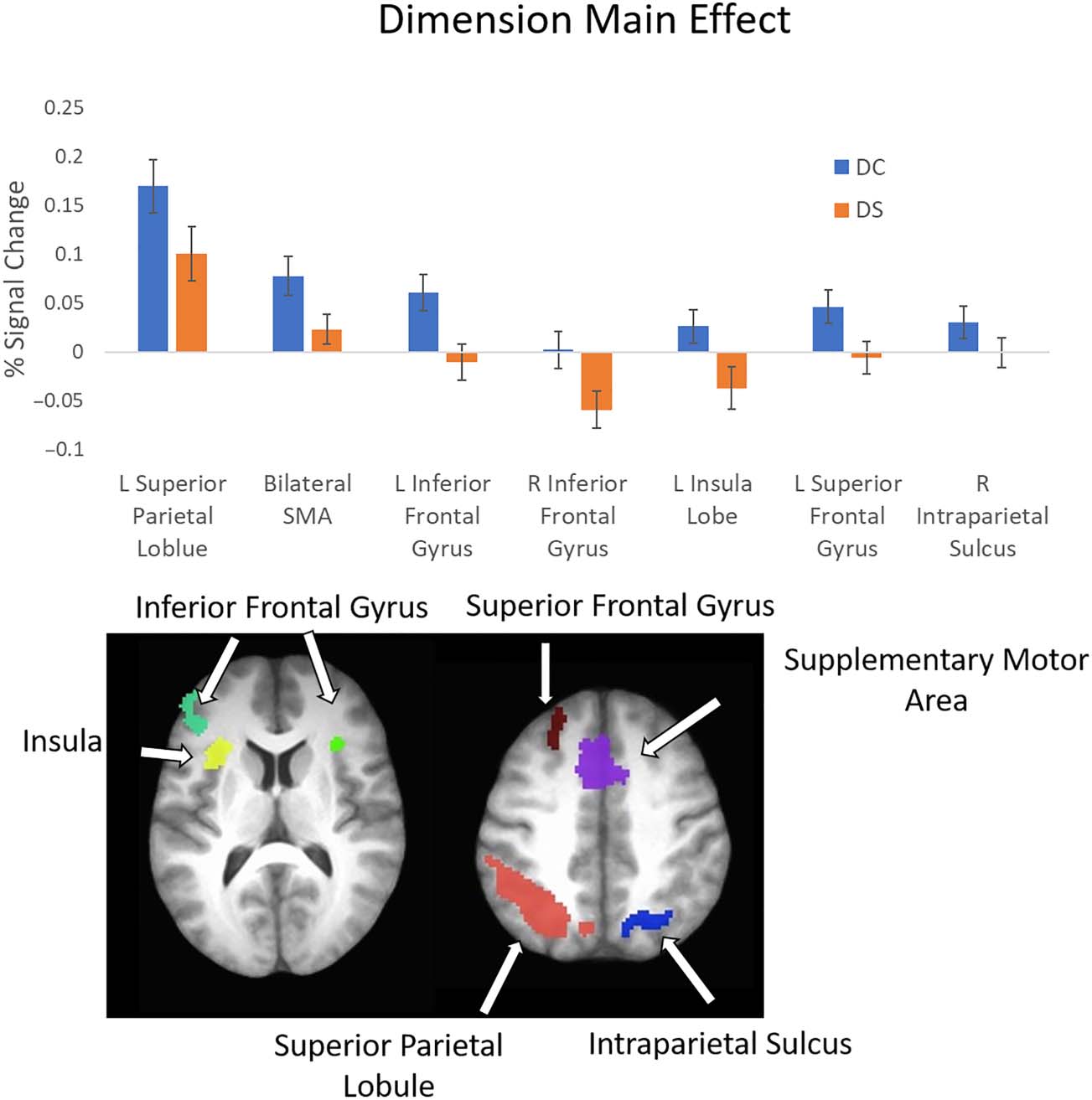
Figure 8. Dimension main
effect. Top: average percent
signal change from the seven
ROIs showing a main effect of
dimension. Error bars represent
standard error of the mean.
Bottom: locations of ROIs.
See Table 3 for list of ROIs.
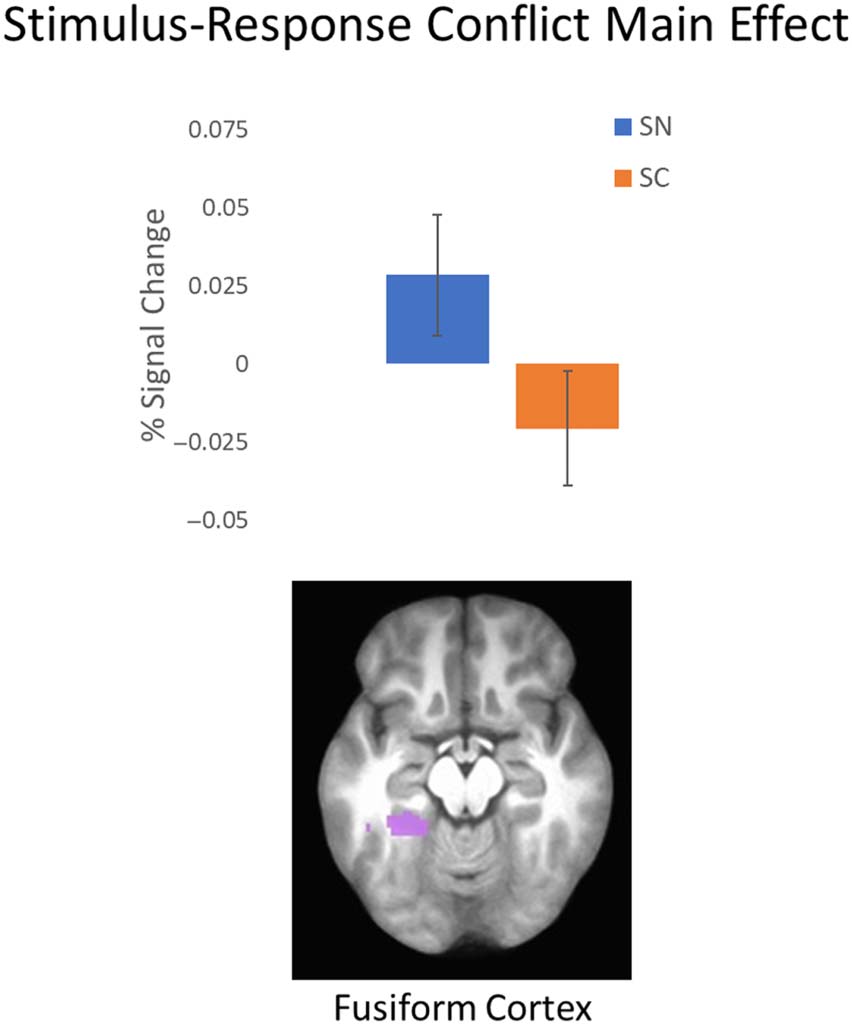
Figure 9. SC main effect. Top: average percent signal change from the
ROI showing a main effect of SC. Error bars represent standard error
of the mean. Bottom: location of ROI. See Table 3 for details of ROI.
SC = stimulus–response-conflict.
1374
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
o
c
n
_
a
_
0
1
7
2
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
o
c
n
_
a
_
0
1
7
2
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
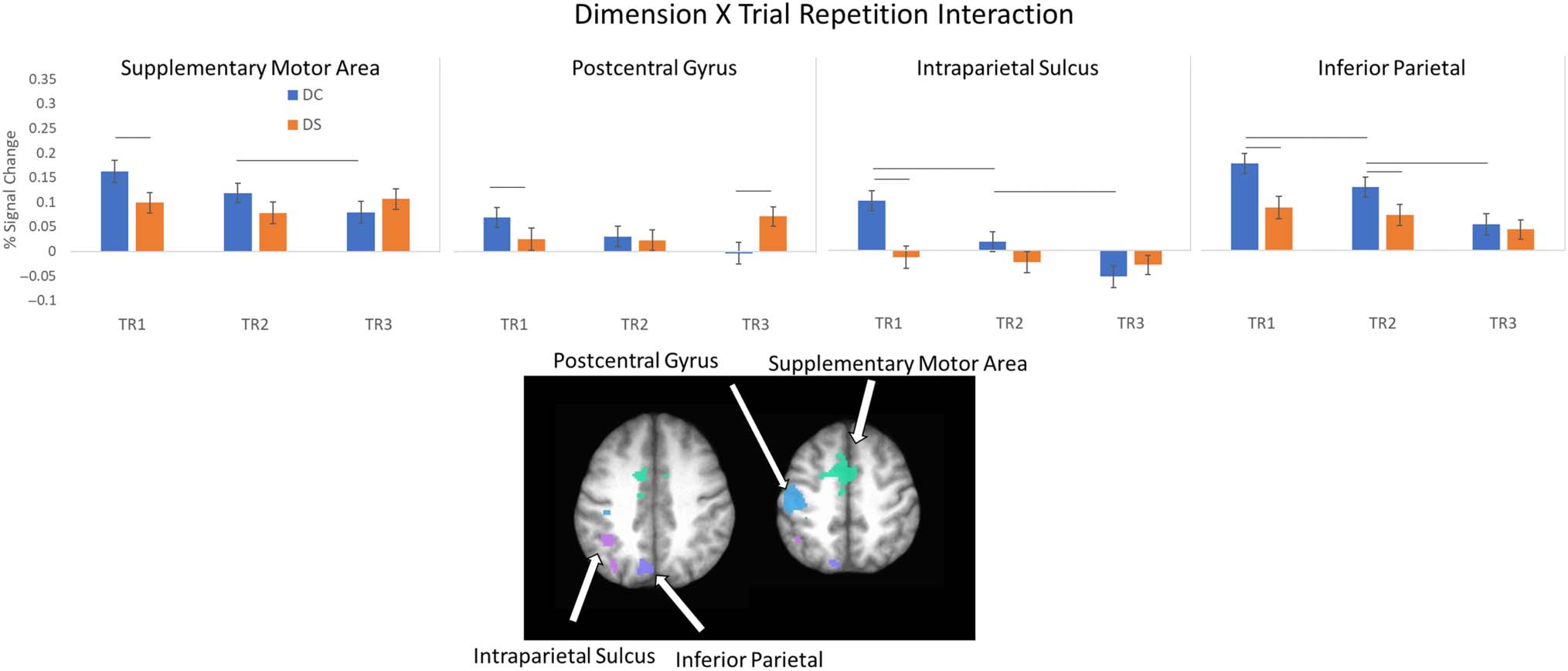
Figure 10. Dimension × Trial Repetition interaction. Top: average percent signal change from the ROIs showing an interaction between dimension
and trial repetition. Error bars represent standard error of the mean. Bottom: location of ROI. See Table 3 for list of ROIs.
(all p < .003). Activation in thalamus decreased from TR1
to TR2 ( p < .001) but was not different between TR2 and
TR3 ( p = .968). Thus, switching was associated with
activation a distributed network that spanned regions of
frontal, parietal, and temporal cortices and subcortex.
Next, we examined the main effect of Dimension
(Figure 8). This is the first examination of rule switching that
involves shifting dimensions compared to not switching
dimensions. This contrast revealed differences in activation
across frontal and parietal regions (see Table 3 for details).
Consistent with the predictions outlined above, stronger
activation was observed when the relevant dimension
changed compared to when it remained the same within
all activated clusters (see bar plot in Figure 8). Examining
the main effect of SC, one cluster was detected in the left
fusiform gyrus (see Figure 9 and Table 3). Consistent with
findings from Buss and Spencer (2018), activation was
stronger in this cluster when SC was absent.
An interaction was observed between Trial Repetition
and Dimension in left SMA, postcentral gyrus, intraparie-
tal sulcus, and inferior parietal cortex (see Figure 10).
Within each cluster, seven follow-up tests were conducted
to compare adjacent trials within a condition (TR1 vs. TR2,
TR2 vs. TR3) and between conditions for each TR.
Threshold values were adjusted using Bonferroni–Holm
corrections. In left SMA, activation on TR2 was significantly
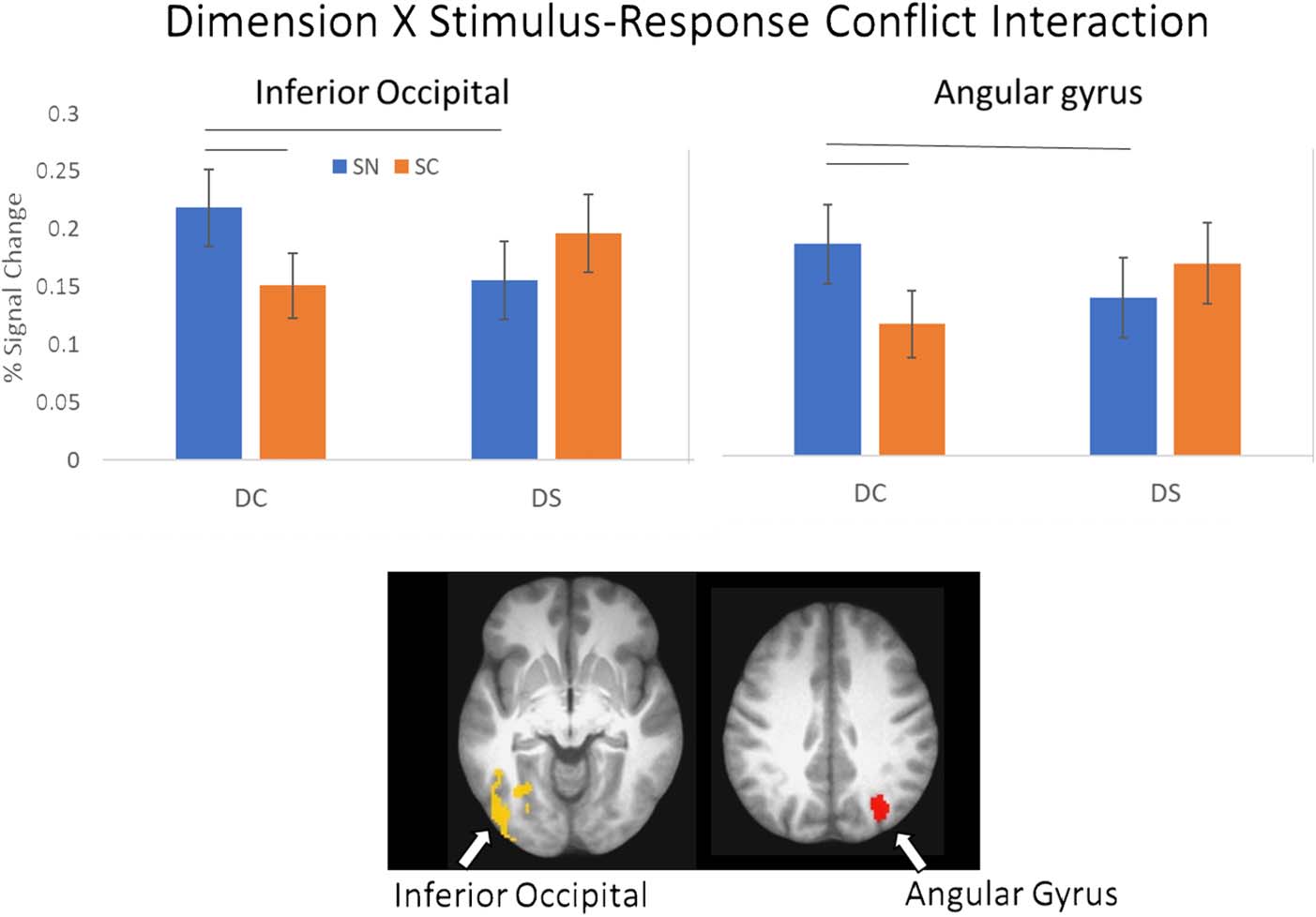
Figure 11. Dimension × SC
interaction. Top: average
percent signal change from the
ROIs showing an interaction
between dimension and SC.
Error bars represent standard
error of the mean. Bottom:
location of ROI. See Table 3
for list of ROIs.
Buss et al.
1375
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
o
c
n
_
a
_
0
1
7
2
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
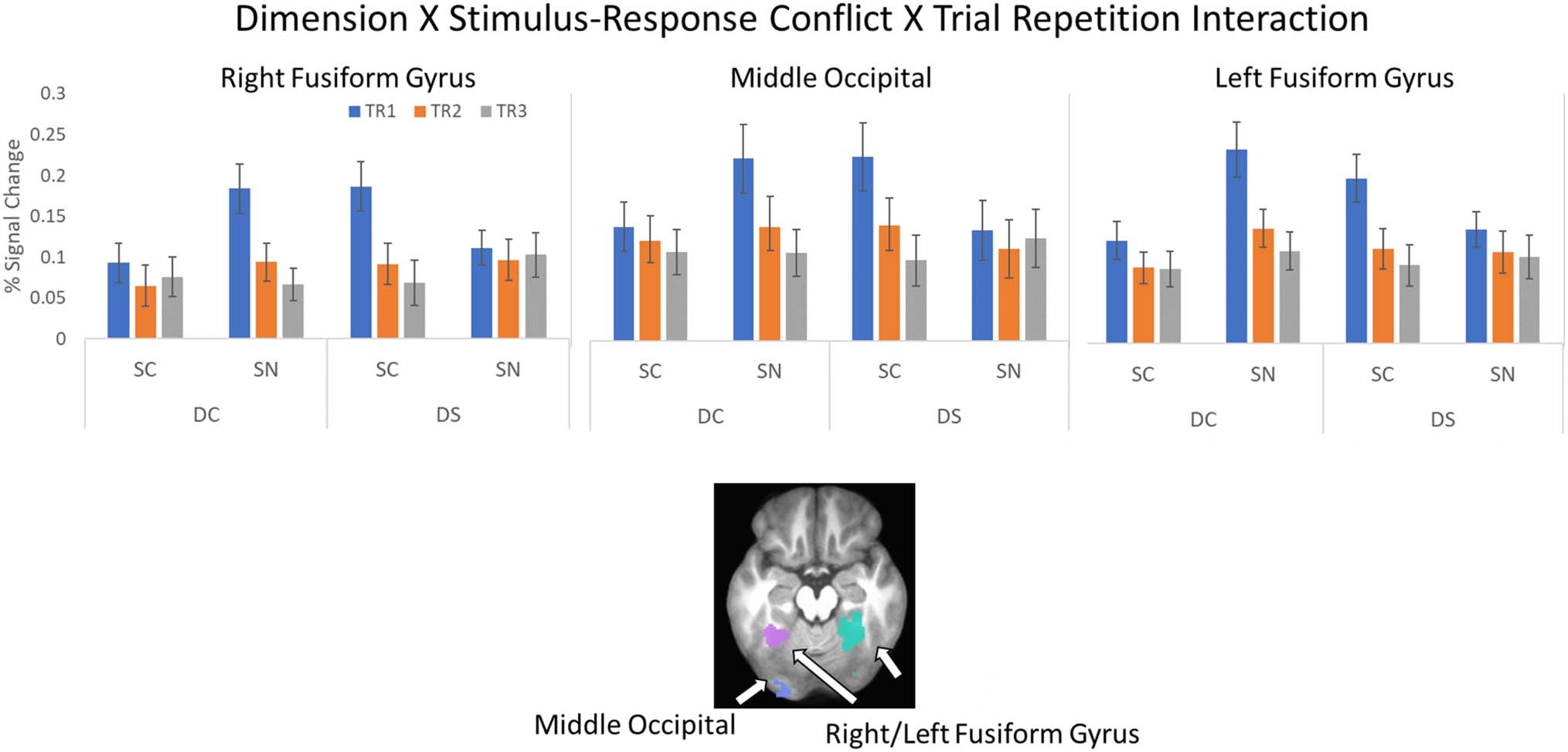
Figure 12. Dimension × SC × Trial Repetition interaction. Top: average percent signal change from the ROIs showing an interaction between
dimension, SC, and trial repetition. Error bars represent standard error of the mean. Bottom: location of ROI. See Table 3 for list of ROIs.
higher than on TR3 when the dimension changed ( p <
.001). In addition, activation on TR1 was higher when
the dimension changed compared to when it remained
the same ( p < .001; all other p > .015). In postcentral
gyrus, activation on TR1 was greater when the dimension
changed compared to when it remained the same, pero el
opposite relationship was present on TR3 ( pag < .001; all
other p > .02). In precuneus, activation was greater on
TR1 compared to TR2 ( pag = .003) and on TR2 compared
to TR3 when the dimension changed ( pag = .002). In addi-
ción, activation on TR1 was greater when the dimension
changed compared to when it stayed the same ( pag <
.001; all other p > .094). Por último, activation in inferior pari-
etal lobule when the dimension changed was greater
on TR1 compared to TR2 and on TR2 compared to TR3
( pag < .001). In addition, activation when the dimension
changed was greater than when the dimension stayed
the same on TR1 and TR2 ( p < .001).
An interaction between Dimension and SC was ob-
served in the left inferior occipital gyrus and right angular
gyrus (see Figure 11). In both clusters, activation on
DCSN was greater than activation on DSSN and DCSC tri-
als ( p < .005). Finally, a three-way interaction was ob-
served in the bilateral fusiform gyrus and left middle
occipital gyrus (see Figure 12). These regions showed in-
creased activation on TR1 of the DCSN and DSSC condi-
tions (all p < .0002).
DISCUSSION
This study presents the first assessment of neural activa-
tion in a dimension switching task across multiple types
of switch conditions. Specifically, we systematically
manipulated the demands on shifting attention to a new
dimension and the need to resolve conflict between
stimulus–response mappings for the current task and
stimulus–response associations from decisions during
the previous task. We observed a network of regions that
were activated in response to these task demands. First,
increased activation on switch trials was observed across
a broad network of frontal, temporal, parietal, and subcor-
tical regions. Furthermore, switching attention to a new
dimension was associated with increased activation in pa-
rietal and frontal regions, whereas the absence of SC was
associated with greater activation in fusiform cortex. We
also observed an interaction between dimension switching
and trial repetitions in parietal cortex and SMA. These
regions showed increased activation on switch trials when
the task switch involved shifting attention to a new dimen-
sion compared to when attention was maintained on the
same dimension. An interaction was also observed be-
tween the dimension and SC factors in inferior occipital cor-
tex and angular gyrus. These regions displayed increased
activation based on the combination of a DC and the
absence of feature-conflict. Finally, an interaction between
all task factors—dimension, SC, and trial repetitions—
was observed in occipital and fusiform cortices. These
regions showed a selective increase in activation on
switch trials (TR1) during the DCSN and DSSC condi-
tions. This pattern of activation could reflect the additional
demands on orienting to the new task set in these condi-
tions. That is, the DCSN condition requires a switch to a
new dimension that contains new feature values, whereas
in the DSSC condition, the task requires making the
opposite spatial response to the same features that were
relevant on previous trials.
1376
Journal of Cognitive Neuroscience
Volume 33, Number 7
Taken together, this pattern of results highlights the
distinct roles that frontal and posterior brain regions play
in flexible dimensional attention. For example, stronger
activation was observed in frontal and parietal cortex
when the relevant dimension switched compared to
when it stayed the same. This suggests that these regions
function to shift attention between visual dimensions.
This observation is consistent with previous neuroimag-
ing findings with the DCCS (Buss & Spencer, 2018;
Ezekiel et al., 2013; Morton et al., 2009). The novel con-
tribution of the data presented here arise from the con-
sideration of the SC factor. First, the main effect of this
factor in fusiform cortex showed increased activation in
the absence of SC. Second, the SC factor interacted with
the DC factor in aspects of occipital and parietal cortex
such that increased activation was observed when switch-
ing dimensions in the absence of SC. These observations
are consistent with the previous observation that reduc-
ing SC increases activation in posterior brain regions
(Buss & Spencer, 2018). A more complicated picture
emerges, however, when considering the three-way in-
teraction of trial repetition, dimension switching, and
SC. Three clusters were identified that showed increased
activation on switch trials in the diagonal of the interac-
tion between the dimension switching and SC factors—
that is, when the dimension changed and SC was absent,
or when the dimension stayed the same and SC was pres-
ent. The latter case is the only situation in which stronger
activation was observed in the presence of SC. Thus, in
general, activation appears to increase when SC is absent,
the one exception being when the task requires a rever-
sal of responses to the same features that were relevant
for the previous task.
Across these effects, an intriguing brain–behavior rela-
tionship is apparent. Specifically, for the trial repetition
and dimension contrasts, stronger activation was ob-
served for trial types that had longer RTs. Typically, such
effects in the literature are thought to reflect an en-
hanced need for cognitive control, that is, between
switch and repeat trials or between dimensional change
and no change trials. By contrast, the opposite pattern
was observed for the SC main effect: Trials that contained
SC had longer RTs compared to trials in which SC was
absent; however, stronger activation was observed on tri-
als in which SC was absent. In this case, there is a boost
in activation in the absence of SC (see also, Buss &
Spencer, 2018). Interestingly, we also observed an inter-
action between SC and dimension switching in occipital
cortex. This suggests that this early object representation
area is sensitive to both feature-based effects and the
need for higher-level cognitive control.
One important limitation of the current study is that
the effect of eliminating SC in the SN conditions was
achieved by altering the relevant features of the objects.
In this regard, it is not clear whether the increased acti-
vation in the SN condition relative to the SC condition is
because of the elimination of conflict or because of the
processing of new features. One alternative way to elim-
inate SC is to align the response-driven associations from
previous decisions with the stimulus–response mappings
during the postswitch. For example, if sorting by color in
postswitch trials, then the colors involved in the task
would be sorted to the same locations during the pre-
and postswitch phases (e.g., red is sorted to the left when
sorting by shape during the preswitch phase, and red is
also sorted to the left when sorting by color during the
postswitch phase; see Figure 3). In this way, the memo-
ries of previous stimulus–response decision would be
supportive of the task mapping required postswitch.
Based on the results of Buss and Spencer (2018) who
implemented such a condition with 3- and 4-year-olds,
we would expect a similar pattern of results to the no-
conflict condition in the current study.
It is important to acknowledge the confounds in the cur-
rent design. These confounds arise from the nature of the
task and the constraints placed on how aspects of the task
can be manipulated independently. For example, we elim-
inated the need to switch dimensions by altering the spatial
configuration of the task and continuing with the relevant
dimension from the previous sorting phase. Thus, the con-
trast between dimension-same and dimension-change
could also be a result of the spatial configuration of the task
being altered or remaining the same; however, the regions
that were sensitive to this manipulation are consistent with
regions involved with dimension switching in previous
studies (Ezekiel et al., 2013; Morton et al., 2009).
Similarly, we eliminated SC by altering the features of the
dimension that was to be relevant for the current phase of
sorting. Thus, the regions showing sensitivity to this manip-
ulation could also be a result of the features remaining the
same or changing. These limitations, however, can serve to
motivate future work exploring these dynamics in the con-
text of the DCCS task. For example, we could compare ac-
tivation and performance across conditions that require
switching dimensions with either the same or altered spa-
tial layout. Relatedly, SC could be manipulated in other
ways. For example, activation and performance could be
examined when memories from the previous sorting phase
cooperate with the configuration of the features for the di-
mension that is to be relevant for the current sorting phase.
This condition would also be absent of SC but would not
be confounded with the introduction of new features.
The current data also challenge existing theories of
flexible attention. Most theories assume that rule repre-
sentation is carried out in an abstract manner within re-
gions of frontal cortex (Bunge & Zelazo, 2006; Morton &
Munakata, 2002) and response competition processes are
resolved in parietal cortex (Crone et al., 2006). That is,
the configuration and reconfiguration of rule representa-
tions are accomplished by processes in frontal cortex,
which exerts a top–down influence on the selection of
responses in parietal cortex. This is true also for neuro-
computational models that focus on the neurocomputa-
tions being carried out by frontal cortex. For example,
Buss et al.
1377
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
o
c
n
_
a
_
0
1
7
2
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Badre and Wagner (2006) describe a computational net-
work in which left ventrolateral frontal cortex involved in
“conceptual” conflict, which is dissociated from response
conflict in left inferior parietal cortex. Other models have
focused on the different functions carried out by frontal
cortex, focusing on different types of conflict detection
mechanisms (Brown, Reynolds, & Braver, 2007) or the
dynamics of updating the representations of task rules
in prefrontal cortex via basal ganglia gating mechanism
(Herd et al., 2014; Rougier, Noelle, Braver, Cohen, &
O’Reilly, 2005). Other models have expanded beyond
frontal cortex representations of rules to examine how
posterior brain regions are also involved in rule represen-
tation processes. A neurodynamic model proposed by
Ardid and Wang (2013) does provide a role for visual
areas in rule representation processes. Specifically, their
model implements neural dynamics involved in dimen-
sional competition and selection in frontal cortex, which
enhance processing of visual areas that map stimuli to re-
sponses. This model too, though, is not able to account
for the effect of stimulus–response associations along the
task-irrelevant dimension because this model only forms
associations along the task-relevant dimension.
The effects reported here are consistent with a dynamic
field model of the development on the DCCS task (Buss &
Kerr-German, 2019; Buss & Spencer, 2014, 2018). Similar
to the model of Ardid and Wang (2013), the dynamic neu-
ral field (DNF) model implements a dimensional attention
mechanism that enhances processing of task-relevant visual
dimension and representations within the visual dimension
provide a mapping of stimuli to responses. In contrast to
this model, however, the DNF model is an embodied
framework that grounds the processing of visual features
in the dynamics of object representation. Building object
representations in the DNF model utilizes spatial coupling
between visual dimensions to activate the features present
on an object. In the context of the DCCS task, the model
builds representations of the test object at the selected re-
sponse location. In this way, the model forms associations
between responses and the features of both the relevant
and irrelevant dimensions of the objects. Across the behav-
ioral and neural results presented here, there is compel-
ling evidence that response-driven associations form not
only along task-relevant dimension but also along the
task-irrelevant dimension. This finding stands at odds
against most theories of task switching, which posit some
form of selective engagement of the task-relevant stimulus
features when a task is engaged. From the perspective of
the DNF model of flexible attention, however, rule-use re-
flects the influence of dimensional attention on object rep-
resentations processes. This framework has been used to
explain a wide array of behavioral findings with children in
the DCCS (Buss & Kerr-German, 2019; Buss & Spencer,
2014) as well as the quantitative details of neural activation
across development and versions of the task that were
made easier by virtue response-driven associations distrib-
uted across feature dimensions (Buss & Spencer, 2018).
Although the DNF model is consistent at a conceptual
level with the results reported here, it is an open ques-
tion whether the DNF model can quantitatively account
for these behavior and neural data. A recent model-based
approach to fMRI analyses (Buss et al., 2021; Wijeakumar,
Ambrose, Spencer, & Curtu, 2017; Buss, Wifall, Hazeltine,
& Spencer, 2014) uses activation from components of the
DNF model in the general linear model to predict the
BOLD signal. Bayesian multilevel modeling is then used
to determine whether the DNF predictors are better than
categorical task-based predictors or which components
of the DNF model best predict activation across the cor-
tex. This approach can further clarify the functional role
that different cortical regions play in flexible attention
and the extent to which the DNF model accurately ex-
plains patterns of activation.
In summary, the current report provides new insights
into the functional role of cortical activation in the con-
text of flexible attention. Our analyses revealed that fron-
tal and parietal cortices help to shift attention from one
visual dimension to another and regions of occipital, tem-
poral, and parietal cortex are sensitive to conflict be-
tween past stimulus–response bindings and current
task demands. Moreover, stimulus–response association
accrue along the dimension that is relevant for the cur-
rent task as well as the dimension that is irrelevant for
the current task. This shows how posterior brain regions
play a central role in the flexible attention needed to
switch between tasks.
Reprint requests should be sent to Aaron T. Buss, Department
of Psychology, University of Tennessee, 1404 Circle Dr.,
Knoxville, TN 37996, or via e-mail: abuss@utk.edu.
Funding Information
John P. Spencer, National Science Foundation (https://dx
.doi.org/10.13039/100000001), grant number: BCS-
1029082. Aaron T. Buss, National Institute of Child Health
and Human Development, grant number: R01HD092485.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by esti-
mated gender identification of first author/last author)
publishing in the Journal of Cognitive Neuroscience
( JoCN) during this period were M(an)/ M = .408,
W(oman)/M = .335, M/ W = .108, and W/ W = .149, the
comparable proportions for the articles that these au-
thorship teams cited were M/M = .579, W/M = .243,
M/ W = .102, and W/ W = .076 (Fulvio et al., JoCN,
33:1, pp. 3–7). Consequently, JoCN encourages all au-
thors to consider gender balance explicitly when select-
ing which articles to cite and gives them the opportunity
to report their article’s gender citation balance.
1378
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
o
c
n
_
a
_
0
1
7
2
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Ardid, S., & Wang, X.-J. (2013). A tweaking principle for executive
control: Neuronal circuit mechanism for rule-based task
switching and conflict resolution. Journal of Neuroscience, 33,
19504–19517. https://doi.org/10.1523/JNEUROSCI.1356-13
.2013, PubMed: 24336717
Badre, D., & Wagner, A. D. (2006). Computational and
neurobiological mechanisms underlying cognitive flexibility.
Proceedings of the National Academy of Sciences, U.S.A.,
103, 7186–7191. https://doi.org/10.1073/pnas.0509550103,
PubMed: 16632612
Brown, J. W., Reynolds, J. R., & Braver, T. S. (2007). A
computational model of fractionated conflict-control
mechanisms in task-switching. Cognitive Psychology, 55,
37–85. https://doi.org/10.1016/j.cogpsych.2006.09.005,
PubMed: 17078941
Bunge, S. A., & Zelazo, P. D. (2006). A brain-based account of
the development of rule use in childhood. Current
Directions in Psychological Science, 15, 118–121. https://doi
.org/10.1111/j.0963-7214.2006.00419.x
Buss, A. T., & Kerr-German, A. (2019). Dimensional attention as
a mechanism of executive function: Integrating flexibility,
selectivity, and stability. Cognition, 192, 104003. https://doi
.org/10.1016/j.cognition.2019.06.015, PubMed: 31228681
Buss, A. T., Magnotta, V. A., Penny, W., Schöner, G., Huppert, T. J.,
& Spencer, J. P. (2021). How do neural processes give rise to
cognition? Simultaneously predicting brain and behavior with
a dynamic model of visual working memory. Psychological
Review. https://doi.org/10.1037/rev0000264, PubMed:
33570976
Buss, A. T., & Spencer, J. P. (2014). The emergent executive:
A dynamic field theory of the development of executive
function. Monographs of the Society for Research in Child
Development, 79, 1–103. https://doi.org/10.1002/mono
.12096, PubMed: 24818836
Buss, A. T., & Spencer, J. P. (2018). Changes in frontal and
posterior cortical activity underlie the early emergence of
executive function. Developmental Science, 21, e12602.
https://doi.org/10.1111/desc.12602, PubMed: 28913859
Buss, A. T., Wifall, T., Hazeltine, E., & Spencer, J. P. (2014).
Integrating the behavioral and neural dynamics of reponse
selection in a dual-task paradigm: A dynamic neural field
model of Dux et al. (2009). Journal of Cognitive
Neuroscience, 26, 334–351. https://doi.org/10.1162/jocn_a
_00496, PubMed: 24116841
Clark, V. P., Parasuraman, R., Keil, K., Kulansky, R., Fannon, S.,
Maisog, J. M., et al. (1997). Selective attention to face
identity and color studied with fMRI. Human Brain
Mapping, 5, 293–297. https://doi.org/10.1002/(SICI)1097
-0193(1997)5:4<293::AID-HBM15>3.0.CO;2-F, PubMed:
20408231
Col, METRO. w., & Schneider, W.. (2007). The cognitive control
network: Integrated cortical regions with dissociable
funciones. Neuroimagen, 37, 343–360. https://doi.org/10.1016
/j.neuroimage.2007.03.071, PubMed: 17553704
Cousineau, D. (2005). Confidence intervals in within-subject
designs: A simpler solution to Loftus and Masson’s method.
Tutorials in Quantitative Methods for Psychology, 1, 42–45.
https://doi.org/10.20982/tqmp.01.1.p042
Cox, R. W.. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computadoras
and Biomedical Research, 29, 162–173. https://doi.org/10
.1006/CBMR.1996.0014, PubMed: 8812068
Cox, R. w., Chen, GRAMO., Glen, D. r., Reynolds, R. C., & taylor, PAG. A.
(2017). fMRI clustering in AFNI: False-positive rates redux.
Brain Connectivity, 7, 152–171. https://doi.org/10.1089/brain
.2016.0475, PubMed: 28398812
Crone, mi. A., Wendelken, C., Donohue, S. MI., & Bunge, S. A.
(2006). Neural evidence for dissociable components of task-
switching. Corteza cerebral, 16, 475–486. https://doi.org/10
.1093/cercor/bhi127, PubMed: 16000652
Diamante, A., & Kirkham, norte. (2005). Not quite as grown-up as
we like to think: Parallels between cognition in childhood
and adulthood. ciencia psicológica, 16, 291–297.
https://doi.org/10.1111/j.0956-7976.2005.01530.X, PubMed:
15828976
Drucker, D. METRO., & Aguirre, GRAMO. k. (2009). Different spatial scales of
shape similarity representation in lateral and ventral LOC.
Corteza cerebral, 19, 2269–2280. https://doi.org/10.1093/cercor
/bhn244, PubMed: 19176637
Ezekiel, F., Bosma, r., & Morton, j. B. (2013). Dimensional
change card sort performance associated with age-related
differences in functional connectivity of lateral prefrontal
corteza. Developmental Cognitive Neuroscience, 5,
40–50. https://doi.org/10.1016/j.dcn.2012.12.001, PubMed:
23328350
Herd, S. A., O’Reilly, R. C., Hazy, t. MI., Chatham, C. h., Brant,
A. METRO., & Friedman, norte. PAG. (2014). A neural network model
of individual differences in task switching abilities.
Neuropsicología, 62, 375–389. https://doi.org/10.1016/j
.neuropsychologia.2014.04.014, PubMed: 24791709
Guijarro, A., Steinhäuser, METRO., Wendt, METRO., Falkenstein, METRO., Jost, K.,
Philipp, A. METRO., et al. (2010). Control and interference in
task switching—A review. Boletín Psicológico, 136,
849–874. https://doi.org/10.1037/a0019842, PubMed:
20804238
Liu, T., Slotnick, S. D., serencias, j. T., & Yantis, S. (2003).
Cortical mechanisms of feature-based attentional control.
Corteza cerebral, 13, 1334–1343. https://doi.org/10.1093
/cercor/bhg080, PubMed: 14615298
Moriguchi, y., & Hiraki, k. (2009). Neural origin of cognitive
shifting in young children. Actas del Nacional
Academia de Ciencias, EE.UU., 106, 6017–6021. https://doi.org
/10.1073/pnas.0809747106, PubMed: 19332783
Morton, j. B., Bosma, r., & Ansari, D. (2009). Age-related
changes in brain activation associated with dimensional
shifts of attention: Un estudio de resonancia magnética funcional. Neuroimagen, 46,
249–256. https://doi.org/10.1016/j.neuroimage.2009.01.037,
PubMed: 19457388
Morton, j. B., & Munakata, Y. (2002). Active versus latent
representaciones: A neural network model of perseveration,
disociación, and decalage. Developmental Psychobiology,
40, 255–265. https://doi.org/10.1002/dev.10033, PubMed:
11891637
Müller, Ud., Dick, A. S., Gela, K., Overton, W.. F., & Zelazo, PAG. D.
(2006). The role of negative priming in preschoolers’ flexible
rule use on the dimensional change card sort task. Child
Desarrollo, 77, 395–412. https://doi.org/10.1111/j.1467
-8624.2006.00878.X, PubMed: 16611180
Rogers, R. D., & monsell, S. (1995). Costos de un cambio predecible
entre tareas cognitivas simples. Revista de experimentación
Psicología: General, 124, 207–231. https://doi.org/10.1037
/0096-3445.124.2.207
Rougier, norte. PAG., Noelle, D. C., Más valiente, t. S., cohen, j. D., &
O’Reilly, R. C. (2005). Prefrontal cortex and flexible cognitive
control: Rules without symbols. Actas del Nacional
Academia de Ciencias, EE.UU., 102, 7338–7343. https://doi.org
/10.1073/pnas.0502455102, PubMed: 15883365
Tallon-Baudry, C., Bertrand, o., Hénaff, M.-A., Isnard, J., &
pescador, C. (2005). Attention modulates gamma-band
oscillations differently in the human lateral occipital cortex
and fusiform gyrus. Corteza cerebral, 15, 654–662. https://doi
.org/10.1093/cercor/bhh167, PubMed: 15371290
Towse, j. NORTE., Redbond, J., Houston-Price, C. METRO. T., & Cocinar, S.
(2000). Understanding the dimensional change card sort:
Buss et al.
1379
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives from task success and failure. Cognitivo
Desarrollo, 15, 347–365. https://doi.org/10.1016/S0885
-2014(00)00021-6
Waxer, METRO., & Morton, j. B. (2011). The development of future-
oriented control: An electrophysiological investigation.
Neuroimagen, 56, 1648–1654. https://doi.org/10.1016/j
.neuroimage.2011.02.001, PubMed: 21316473
Wijeakumar, S., Ambrose, j. PAG., Spencer, j. PAG., & Curtu, R.
(2017). Model-based functional neuroimaging using dynamic
neural fields: An integrative cognitive neuroscience approach.
Revista de Psicología Matemática, 76, 212–235. https://doi
.org/10.1016/j.jmp.2016.11.002, PubMed: 29118459
Zahn, C. T., & Roskies, R. z. (1972). Fourier descriptors for
plane closed curves. IEEE Transactions on Computers, C-21,
269–281. https://doi.org/10.1109/TC.1972.5008949
Zelazo, PAG. D., Müller, Ud., Frye, D., Marcovitch, S., Argitis, GRAMO.,
Boseovski, J., et al. (2003). The development of executive
function in early childhood. Monographs of the Society
for Research in Child Development, 68, vii–137. https://doi
.org/10.1111/j.0037-976x.2003.00260.x, PubMed: 14723273
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
3
6
5
1
9
2
1
3
0
9
/
/
j
oh
C
norte
_
a
_
0
1
7
2
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
1380
Revista de neurociencia cognitiva
Volumen 33, Número 7