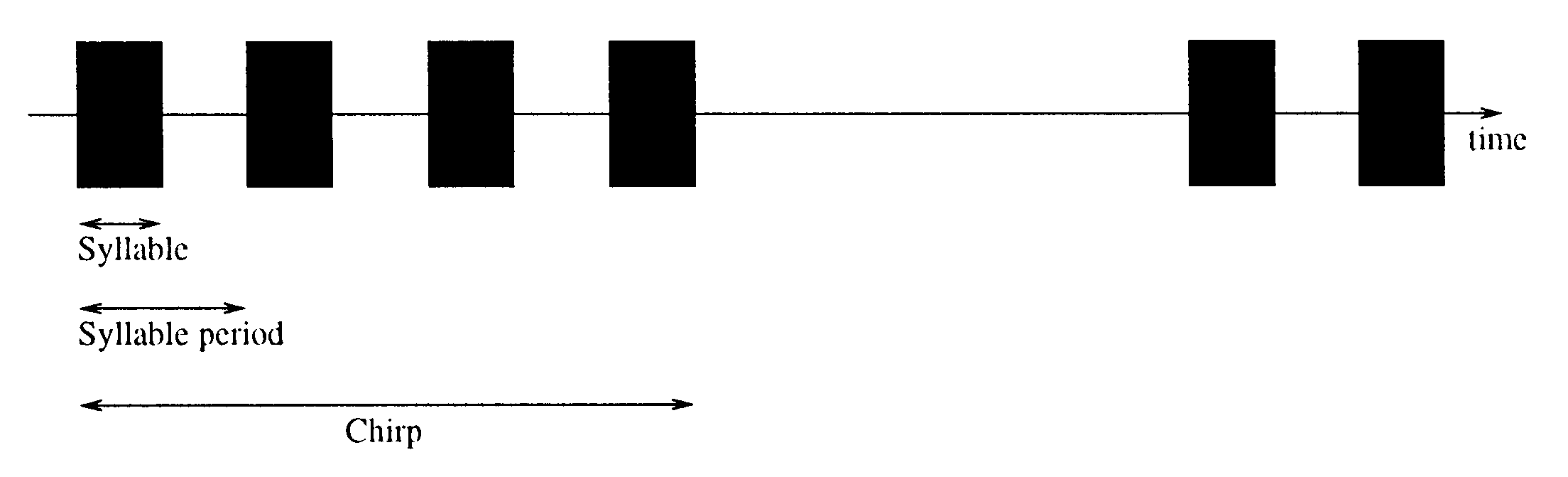Physical and Temporal Scaling
Considerations in a Robot Model
of Cricket Calling Song Preference
Abstract Behavioral experiments with crickets show that
female crickets respond to male calling songs with syllable
rates within a certain bandwidth only. We have made a robot
model in which we implement a simple neural controller that
is less complex than the controllers traditionally hypothesized
for cricket phonotaxis and syllable rate preference. El
simple controller, which had been successfully used with a
slowed and simplified signal, is here demonstrated to
función, using songs with identical parameters to those
found in real male cricket song, using an analog electronic
model of the peripheral auditory morphology of the female
cricket as the sensor. We put the robot under the same
experimental conditions as the female crickets, y eso
responds with phonotaxis to calling songs of real male
Gryllus bimaculatus. Más, the robot only responds to
songs with syllable rates within a bandwidth similar to the
bandwidth found for crickets. By making polar plots of the
heading direction of the robot, we obtain behavioral data that
can be used in statistical analyses. These analyses show that
there are statistically significant differences between the
behavioral responses to calling songs with syllable rates
within the bandwidth and calling songs with syllable rates
outside the bandwidth. This gives the verification that the
simple neural control mechanism (together with
morphological auditory matched filtering) can account for the
syllable rate preference found in female crickets. With our
robot system, we can now systematically explore the
mechanisms controlling recognition and choice behavior in
the female cricket by experimental replication.
Henrik Hautop Lund∗
The Danish National Centre
for IT-Research
University of Aarhus, Ny
Munkegade Bldg. 540
8000 Aarhus C.
Dinamarca
hhl@daimi.aau.dk
http://www.daimi.aau.dk/∼hhl
Barbara Webb
Department of Psychology
University of Nottingham
Nottingham NG7 2RD
Inglaterra, Reino Unido
Barbara.Webb@nottingham.ac.uk
John Hallam
Department of Artificial
Inteligencia
University of Edinburgh
5 Forrest Hill
Edinburgh EH1 2QL
Escocia, Reino Unido
john@dai.ed.ac.uk
Palabras clave
neuroetología, behavioral biology,
cricket phonotaxis, robotics, audi-
tory system
1 Introducción
Taxis, or approach to a sensory source, is frequently modeled in animat simulations,
as a basic behavior of animals in seeking food, compañeros, or nests. The task is commonly
described by the following abstraction: A source produces a signal, the value of which
decreases with distance (smoothly or as a step function), and an animat can sense the
value and use it to control movement to the source. Braitenburg’s [2] “vehicle 2” is an
archetypical controller for this task.
∗ The first author provided the extended neural model, made the experiments and analysis described in the article, and wrote
Secciones 3, 4, 5, 6 and parts of 2 y 7. The second author provided the original hypotheses and wrote Sections 1, parts of 2, y
7. The third author designed the auditory circuit.
C(cid:176) 1998 Instituto de Tecnología de Massachusetts
Artificial Life 4: 95–107 (1998)
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
It is often argued that such simulations model the “essence” of the behavior—as
such they should illuminate the understanding of any specific example of the behav-
ior. Sin embargo, what these abstractions generally ignore are the real physics of signal
propagation and detection. The abstract model is a reasonable representation of light,
and thus translating simulation results to robot phototaxis is quite often successful. Pero
the model is less adequate for other modalities, many of which are more salient to
real animals. Por ejemplo, in chemotaxis, the properties of the odor plume are critical
determinants of the problem [1]. Braitenberg-type control is not appropriate in a task
where the signal is highly dispersed (eliminating local gradients) and carried in specific
directions by currents in the environment (p.ej., around obstacles).
We have argued previously [10] that the common abstraction is positively misleading
when applied to the specific example of phonotaxis behavior in crickets. Here the
sensors do not respond exclusively to the relevant signal—bursts of sound of a charac-
teristic frequency and repetition rate. Sin embargo, rather than first filtering for the signal
properties and subsequently comparing strength to control turns, the directionality of
the sensors and the motor control mechanism are dependent on signal characteristics.
Por ejemplo, a tube connecting the ears transfers a phase-delayed signal from one
to the other, thus modifying the negligible amplitude difference between the sensors
of this small animal in a noisy environment. Thus we can only understand this sys-
tem through consideration of the physical properties of the task and agent, which we
believe are best investigated through building a physical model.
The same considerations suggest that the physical and temporal scales of the model
are likely to be influential in the success of certain controllers for the behavior. Nuestro
original implementation of a robot model (a LEGO robot prototype) of the cricket
suffered from a number of limitations in this regard, particularly in that the processing
of sound was relatively slow compared to the speed of movement [10]. While this
model nevertheless sufficed to demonstrate the viability of basic mechanisms, fue
difficult to make strong comparisons with data from the animal.
These limitations, and details of the construction of a new robot addressing them,
have been described in detail in [5]. We showed there that the reimplementation en-
abled us to verify experimentally assumptions that had been made, but not tested,
about how the cricket responds selectively to carrier frequency of the signal. Here we
report how the new robot led us to a revised model of neural control of phonotaxis.
Our aim was to reproduce the band-pass selectivity for repetition rate in the signal,
using temporal scales identical to those used in cricket experiments.
2 Neural Model
The original neural model was based on two properties of identified interneurons (AN1)
known to be involved in phonotaxis in the cricket [8]. Primero, the long time constant of
these neurons means that they act as low-pass filters for the temporal pattern of the
señal: Given repeated inputs they can only code distinct syllables (bursts of sound)
below a certain repetition rate (for calling song structure, ver figura 1). Segundo, el
latency to onset of firing for each syllable is dependent on the amplitude of the sound.
Thus which side fires first can be used to generate a turn toward the sound. Sin embargo,
this mechanism has two features: It will not work when syllables repeat too fast for
the low-pass filtering to track the onsets; and if the syllables repeat slowly, signals to
turn will come less often. Thus there should be a band of repetition rates for which
this mechanism works best.
We were able to demonstrate an effective “preference” for certain syllable rates using
this mechanism in the LEGO robot. Sin embargo, this depended partly on the fact that the
LEGO robot could move a significant distance between slow syllables, which because
96
Artificial Life Volume 4, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
Cifra 1. The song structure of the cricket calling song. The calling song consists of chirps, each with a number of
syllables (bursts of sound with the species-specific carrier frequency). En este caso, with equal syllable length and gap
length, the syllable duty-cycle (within a chirp) es 50%.
of the auditory processing speed lasted a second or more in the LEGO robot. Real
cricket song has repetition rates in the order of 30 Hz, and “slow syllables” to which
they no longer respond may last only 50 EM. With the new robot able to process song
at comparable rates to the cricket, slowing the song to 10 Hz or less was not sufficient
to interfere with taxis because it was still making 10 corrections a second, which was
quite adequate to get it to the sound source. Consequently we had to produce a more
complex neural control model than the one suggested in [10] and used to show the
robot’s phonotaxis to male Gryllus bimaculatus calling song in [5]. En particular, Tiene
been extended to include an extra neuron on each side that performs integration over
syllables. This simple neural system can account for much of the biological data that
show a band-pass in crickets’ phonotaxis to calling songs with different syllable rates.
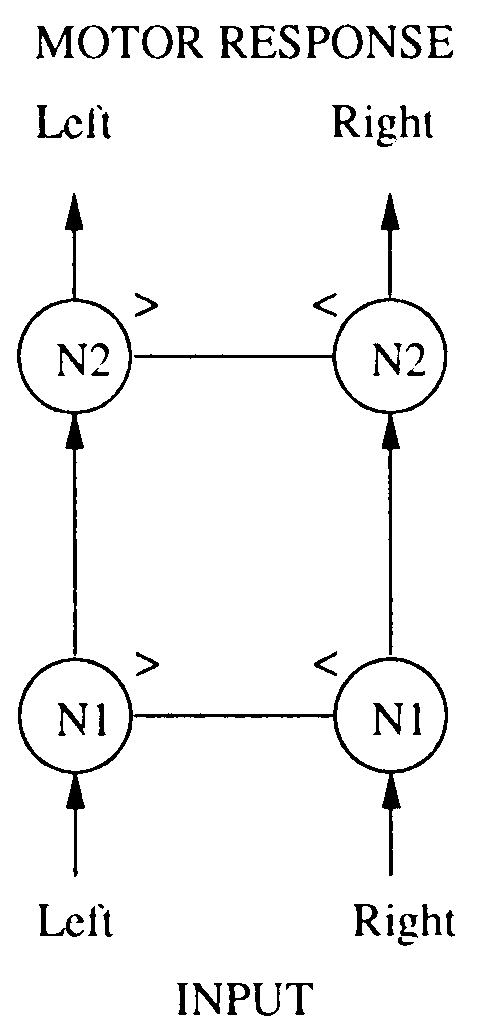
The structure of the extended neural model is as follows: Input from the auditory
sensors is fed into a neuron on each side (N1); activation can flow from N1 to another
neurona (N2), which in turn feeds activation directly to the motor on that side. El
activation within a neuron is modeled with a leaky integrator specific for that neuron,
so that the activation, A, at time t is calculated as
A(t) = α ∗ A(t − 1) + I (t),
0 < α < 1, if A(t) > Thigh, A(t) = Thigh
where I is the input to that neuron and α is the decay rate. When A(t) reaches the
upper threshold Thigh, it will “fire,” that is, pass activation on within the network. A
neuron that has produced an activation pulse cannot produce another pulse until its
activation has first decayed below a lower threshold Tlow. Thus a pulse represents an
activation onset, eso es, an increase of activation from below Tlow to above Thigh, pero
not continued activation. The decay rates and thresholds of the neurons can be varied
to produce particular kinds of temporal responsiveness.
Figures 2 y 3 show how the N1 and N2 neurons interact. Cifra 2 shows the
temporal response of neurons on the side receiving the strongest input, during three
chirps of three syllables. Each syllable activates N1 above T 1high, and each gap between
syllables allows N1 to decay below T 1low. Thus N1 will produce one pulse per syllable
as input to N2. A single pulse is not sufficient to make N2 reach T 2high, but because
the decay is slow, successive pulses at a sufficiently high rate will result in N2 reaching
límite:
in this case after three syllables or the length of the chirp. It can be seen
that N1 will act as a low-pass filter (responding only if the gaps between syllables are
large enough) and N2 as a high-pass filter (responding only if the syllables occur fast
suficiente).
The summation time to threshold in N1 will depend on the strength of the input
señal. The left N1 passes activation to the left N2 only if it reaches threshold before
the right N1, y viceversa. This means that the output of N2 indicates a louder signal,
Artificial Life Volume 4, Número 1
97
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
Cifra 2. The neural model implemented in the robot. The figure shows the neural activation of neuron N1 at the
bottom and of neuron N2 at the top over the same time scale. Three chirps each with three syllables is imagined
as input to neuron N1.
Cifra 3. The structure of the neural control mechanism. Activation from the ears is fed into N1. If N1 activation
on either left or right reaches T1high, it is compared with the activation on the contralateral side. If it is higher, el
neuron will fire activation on to N2 on the same side. N2 neurons work in a similar way.
of the right temporal pattern, and thus it can be used directly to produce a turn in the
correct direction.
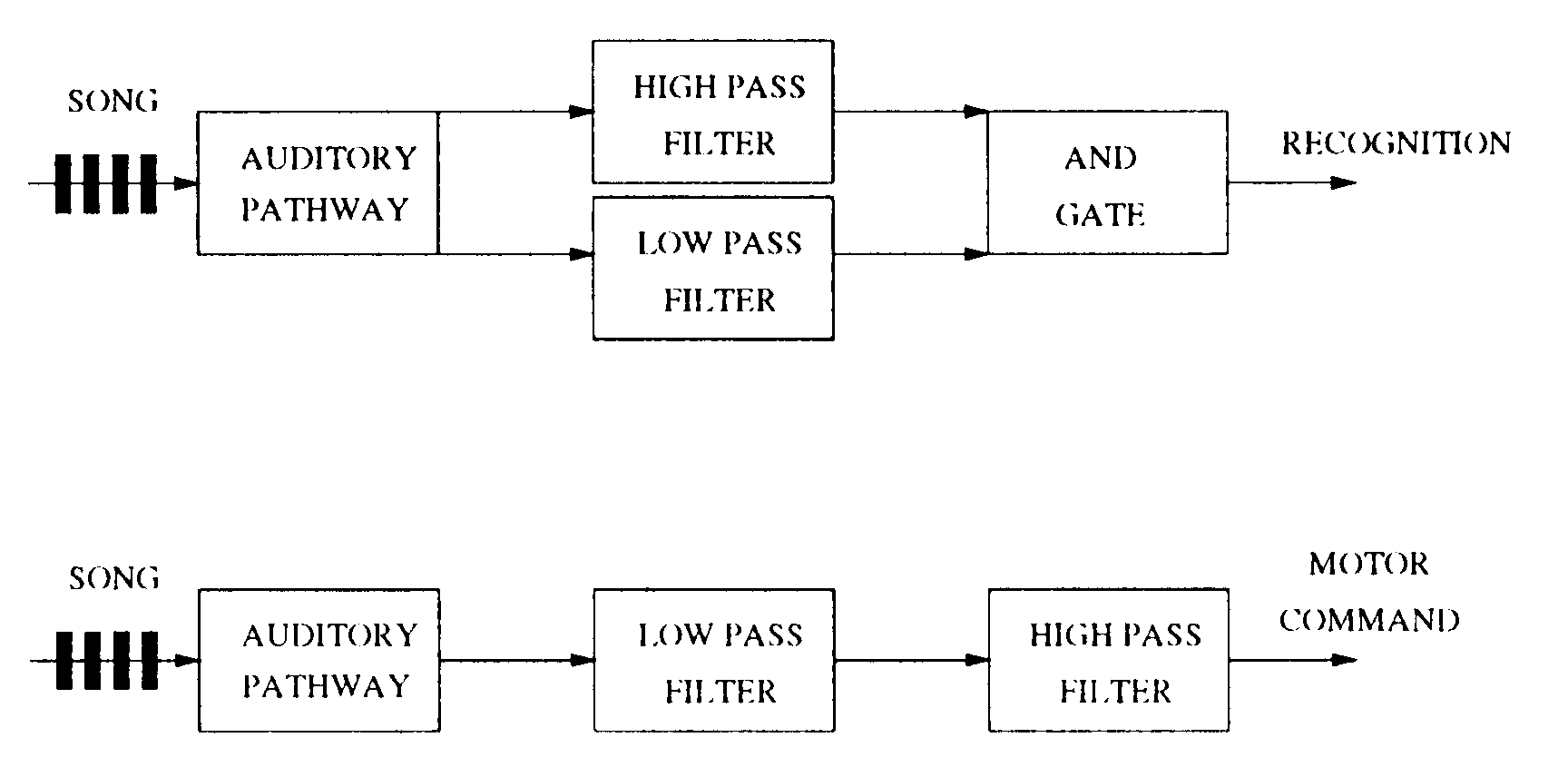
The neural model suggested here is somewhat similar to the one suggested by
Schildberger [7], who found low-pass and high-pass neurons in neurophysiological
experiments with Gryllus bimaculatus. Based on this, Schildberger suggested a neural
model with low-pass and high-pass neurons that process in parallel and then is ANDed
together to produce the band-pass filtering observed in behavioral experiments (ver
Cifra 4). Sin embargo, the model suggested here is simpler because it directly produces
the motor behavior, and not only recognition of the right syllable rate.
3 Robot Implementation
The female cricket has four auditory openings: an ear (tympanum) located on each
upper foreleg, and an auditory spiracle (or hole) on each side of the frontal section of
her body. The four are linked internally by means of tracheal tubes. Sound reaches
the tympani directly through the air and, after propagation through the internal tubes,
from the other auditory openings. The sound transduced from each tympanum by the
cricket’s auditory receptors is thus a combination of delayed and filtered signals from
the other tympanum and the spiracles arriving at the back of the tympanum with the
direct sound arriving at its outer face.
The delays and filtering performed by the auditory morphology improve the cricket’s
ability to discriminate the arrival direction of the conspecific song because the phased
combination of sounds from the different sources induces a strong directional sensitivity
98
Artificial Life Volume 4, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
Cifra 4. Top: Diagram of Schildberger’s [7] hypothesized mechanism for recognition of syllable rates. Bottom: Di-
agram of our hypothesized mechanism for both recognition and motor response for syllable rates in the right
band-width.
into the response of each tympanum. Esencialmente, sounds arriving from the same side
as the tympanum are delayed by the internal structures to arrive in antiphase with
respect to the direct path at the ipsilateral ear and in phase at the contralateral ear.
Because the sounds arriving by the two paths are subtracted (being on opposite sides
of the tympanum), the stimulus intensity at the ipsilateral ear is enhanced while at the
contralateral ear it is diminished.
In the cricket, the delays and filter characteristics of the internal auditory structures
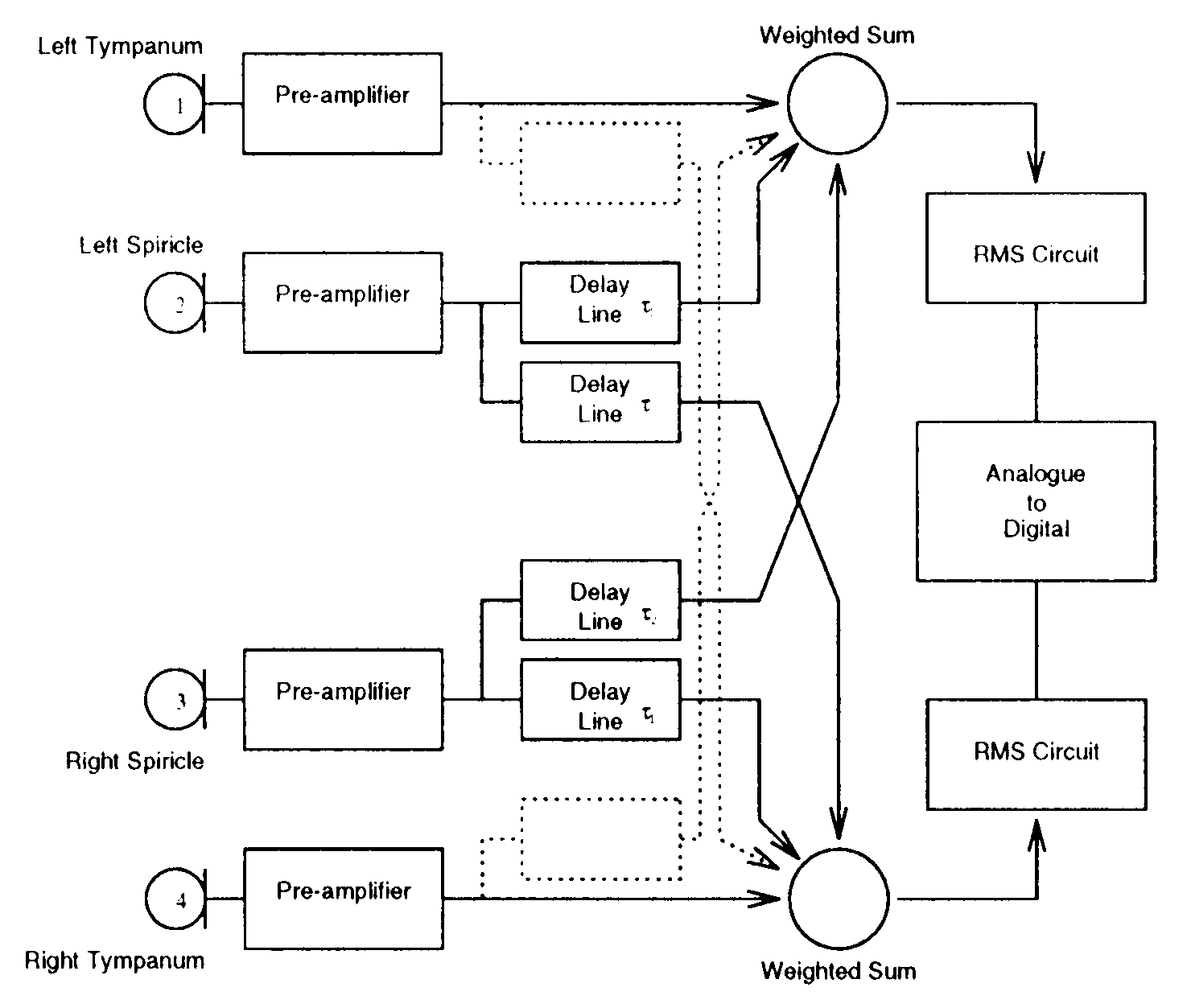
are species-specific. To model the auditory morphology of the cricket, we have built
an electronic emulation of some of these characteristics (see Figures 5 y 6). Sound
is collected by two or four microphones whose spacing is carefully controlled.
En
the present experiments, we use only two microphones to approximate the signal
contributions from the two sides of the cricket. The reason for not summing from the
opposite tympanum was to emulate biological findings that show that the opposite
tympanum makes a negligible contribution to the sum. After amplification and initial
filtering three delayed copies of the sound are generated with programmable relative
delays, which are then scaled and added together to construct a tympanal response. El
intensity of the resultant signal is transduced using an analogue-to-digital conversion
system for use by the control program. This hardware allows us to approximate the
auditory morphology of various crickets by adjusting the programmable delays and the
summing gains. It is not a perfect emulation of the insect, sin embargo: Two programmed
delays allow us to sum signals from each tympanum and both spiracles, but not from
all auditory openings; and the summation system allows us to program relative gains,
but not frequency dependent gains.
4 Phonotaxis Experiments
En [5], we used the robot with the auditory system described above to verify that the
simple neural control mechanism could account for frequency selectivity, because the
robot did phonotaxis to male cricket Gryllus bimaculatus calling song and preferred
calling songs with the right carrier frequency. Aquí, we will first replicate some of these
experiments with the extended model and then go on to the syllable rate experiments.
In the present experiments, for N1 neurons, the decay rate, α1, is set to 7/8, superior
límite, T 1high, a 900, and lower threshold, T 1low, a 600. For N2 neurons, el
decay rate, α2, is set to 63/64, upper threshold, T 2high, a 1,725. These are empirical
settings.
The robot has its auditory sensory system’s parameters set as in [5] (es decir., the two mi-
crophones are placed 18 mm apart because 18 mm corresponds to one-quarter wave-
Artificial Life Volume 4, Número 1
99
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
Cifra 5. The Khepera robot with the auditory sensors. C(cid:176) Lund, Hallam, & Webb, 1997.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 6. Simplified diagram of the ears circuit. When sound arrives at each microphone (the analogue of the
cricket’s tympanum), the received signal is preamplified. The signal is then sent with a “through delay” to the mixer
at the same side, and with a “cross delay” to the mixer at the opposite side. The mixed signal is sent through a
root-mean-square (RMS) to derive a measure of the average energy in the mixed signal, and an A/D converter to
one of the Khepera’s input channels. The same happens on the opposite side. C(cid:176) Lund, Hallam, & Webb, 1997.
length of the carrier frequency, 4.7 kHz, of male Gryllus bimaculatus calling song; el
delays are set to 53 µs (one-quarter of the period of 4.7 kHz); and the mixers subtract
the delayed signal from the contralateral side from the signal from the lateral side).
The cycle time of the controller is approximately 1.3 EM. When we play male Gryllus
bimaculatus calling song to the robot, the empirical settings of neural activation decay
and thresholds described above result in the neural effect shown in Figure 2.
100
Artificial Life Volume 4, Número 1
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
Cifra 7. The robot’s behavior when male cricket Gryllus bimaculatus calling song is emitted from the loudspeaker
at the bottom of the arena.
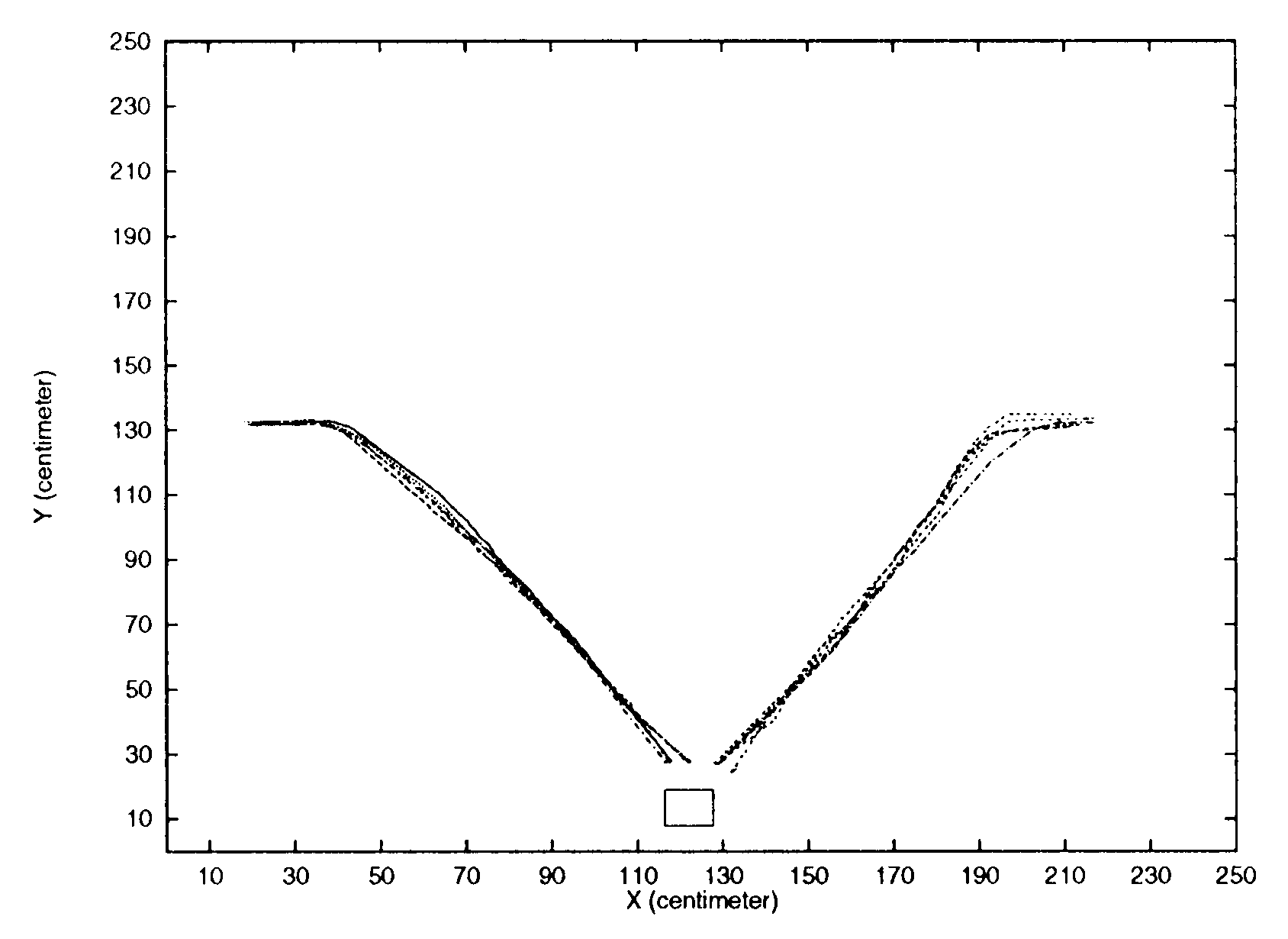
We have replicated the data from [5] with the extended neural model. The results
are shown in Figure 7.
It shows that the robot with the extended neural models
performs phonotaxis to male cricket Gryllus bimaculatus calling song1 emitted from a
loudspeaker in the arena. The figure shows 10 different runs with the robot’s starting
point alternating between 45 degrees left or right to the sound source. Both starting
points are 150 cm away from the sound source. En todos los casos, the robot moves forward
a bit, then it reacts to the calling song by turning toward the loudspeaker and moves
directly toward it.
Por eso, the robot with this neural model is attracted to male Gryllus bimaculatus
calling song. As was the case with the robot with the previous neural model, this robot
also discriminates between calling songs with the right carrier frequency (4.7 kHz)
and those with other carrier frequencies (p.ej., 6.7 kHz). (The data is not shown here
because of space limitation and emphasis on syllable rate preference. For frequency
selectivity, ver [5].
5 Syllable Rate Experiments
Syllable rate experiments with crickets have shown that the female cricket responds
only to calling songs with syllable rates within a certain band-pass. Weber and Thorson
[12] report that Gryllus campestris females to “tracking almost perfectly near 30 syllables
per second but [son] revealing reduced performance or [son] stopping entirely at rates
below 20Hz or above 40Hz” (pag. 321). These data were found using trills, eso es,
songs with no chirp structure but a continuous syllable and pause pattern (por ejemplo,
50% syllable and 50% pause). Sin embargo, Weber and Thorson [12] also report that in
performance tests for chirps with different syllable numbers, “we have uniformly found
that the females’ response increases as the number of syllables increases, as long as
syllables are delivered at the natural (30-Hz) rate” (pag. 318). Doherty [3] also found
that Gryllus bimaculatus failed to discriminate between calling songs having a 45 EM
1 The calling song was recorded at the life science department, University of Nottingham. The adult male cricket was sitting in a
sand-floored arena and was recorded using a Maplin unidirectional dynamic microphone (YU-34) on a Marantz stereo cassette
recorder (CP230) from a distance of about 20 cm. A 30 s part of these recordings is played through a host Pentium computer with
SB AWE32 sound card and was repeated twice for each experiment. The sound was fed through an amplifier to a loudspeaker
that was placed on a 240 ∗ 240 cm arena in our robot lab.
It should be noted that we did nothing to control echos from the
surrounding environment.
Artificial Life Volume 4, Número 1
101
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
syllable period and an alternative syllable period until the alternative syllable period
era 30 ms or shorter, o 55 ms or longer.
Though Popov and Shuvalov’s [6] arena experiments suggest that Gryllus campestris
are more attracted by the four-syllable calling song than those with three or five or more
syllables, other experiments [4, 12] show that chirps can be dispensed with entirely and
females can “track continuous trills of syllables delivered at rates near 30Hz. Nuestro
recent tests indicate that this ability increases with the age of the animal” ([12], pag. 318).
Regarding the change over age, Stout and McGhee [9] also conclude that adult female
A. domestica were more attracted to calling songs with a much wider range of syllable
periods than were attractive to younger females, and thus “for A. domestica females,
a central filter for SP [syllable period] would at least need a variable band-pass width
that could be influenced by other stimuli, edad, and other variables” (pag. 287).
In the following experiments, we will show how the simple neural model suggested
above together with the auditory mechanism implemented in the auditory sensor circuit
can account for this data on cricket band-pass filtering of syllable rates. Más, el
band-width is dependent only on the characteristics of the N1 and N2 neurons (a saber
the decay rate and the firing threshold) that might change over age in natural nervous
sistemas.
We made computer-generated trills of a carrier frequency of 4.7 kHz (the carrier
frequency of the Gryllus bimaculatus calling song) with different syllable rates. El
trills had syllable periods of 10, 20, 30, 40, 50, 60, 70, y 80 EM. The total sound
energy was kept constant (con un 50% duty cycle, es decir., the syllables were 50% del
syllable period).
As in the previous experiment, we replicated each experiment 10 times by placing
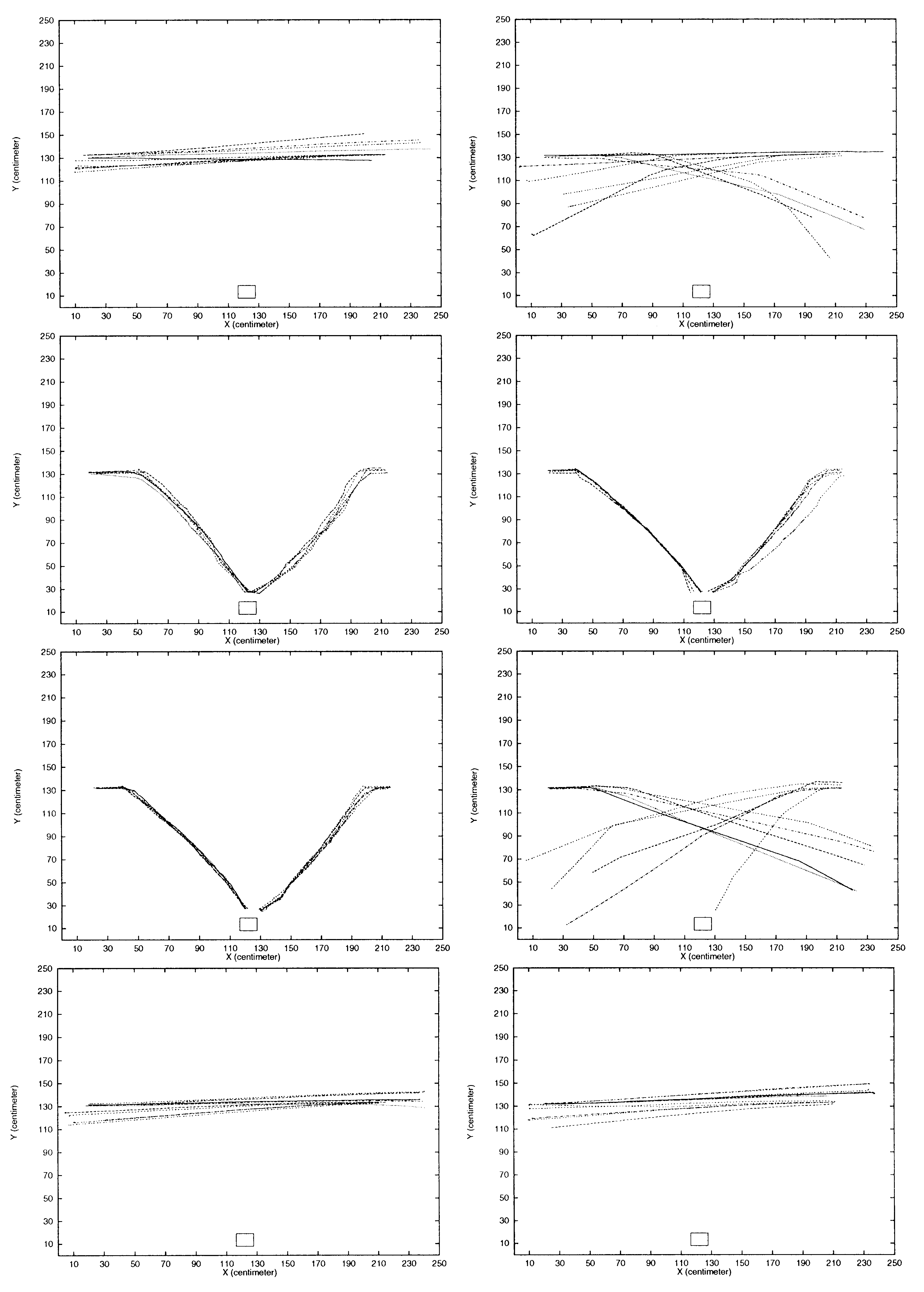
the robot at the two starting positions 5 times each (alternating between them). Cifra 8
shows the result of these experiments. When the robot was presented with the 10 EM
syllable period trill, it did not react at all but just moved with the default forward
movimiento. With the 20 ms syllable period trill, the robot would react very few times
and would not perform successful phonotaxis. Sin embargo, con 30, 40, y 50 ms syllable
period trills, the robot performed phonotaxis by reacting to the trill and moving to the
source. Con 60 ms syllable period trills, the robot reacted to the trill very few times
and managed to reach the sound source only once in the 10 carreras. With neither 70 EM
nor 80 ms syllable period trills did the robot react to the sound at all. Por eso, el
simple neural model provides an effective syllable rate band-pass filter that allowed the
En este caso, the neural
robot to perform phonotaxis only with a narrow band-pass.
band-pass filtering was such that the robot could not perform successful phonotaxis
below a syllable period of 30 ms or above 50 EM.
6 Statistical Analyses
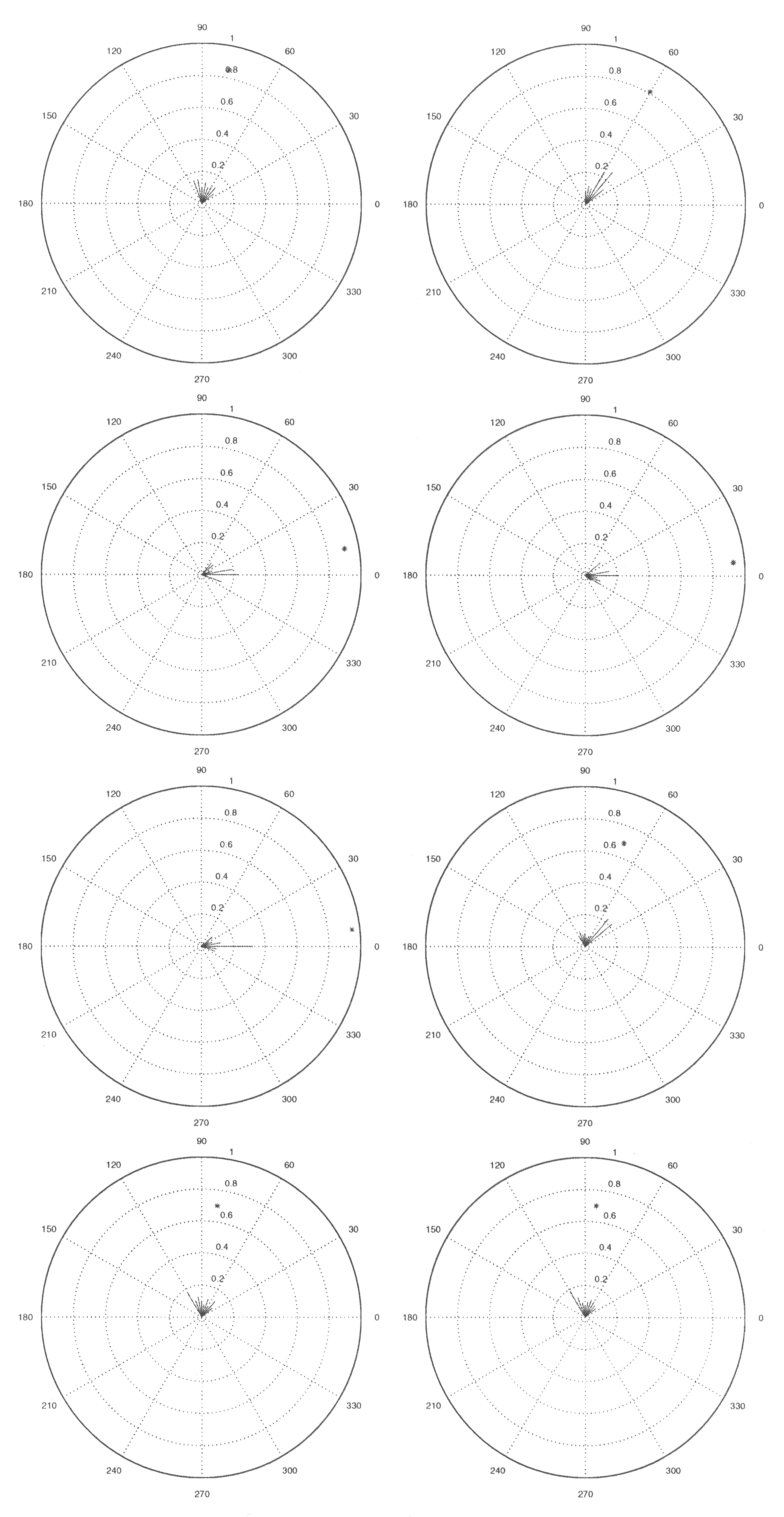
To analyze the trajectories of the robot and whether there are statistically significant
differences between performances under the different experimental setups, we plot
the robot’s heading in polar coordinates with the origin at the loudspeaker position.
Approximately each 0.3 s, we record the robot’s position and find the vector connecting
the previous position of the robot with its present position. This vector is then plotted in
the polar plot using the line from the previous position to the origin (at the loudspeaker)
as the polar reference direction. Thus if the robot heads directly toward the speaker,
its movement is plotted with zero angle. If the speaker direction is to the right of the
robot’s heading, the polar plot shows a positive angle, etcétera.
After one run, the “vector mean” of the polar plot vectors is calculated; its direction
is that of the sum of the plotted vectors, but its length is the length of the direct path
from the robot’s starting point to the loudspeaker divided by the length of the vector
102
Artificial Life Volume 4, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 8. The robot’s behavior when trills are emitted from the loudspeaker at the bottom of the arena. The duty
cycle is 50% in all experiments, while the syllable rate changes. The experiments are with syllable intervals of 10,
20, 30, 40, 50, 60, 70, y 80 EM (ordered left to right from top to bottom). C(cid:176) Lund, Hallam, & Webb, 1998).
Artificial Life Volume 4, Número 1
103
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
Mesa 1. Analysis of difference between mean heading directions in polar coordinates between experiments with
computer-generated trills with syllable periods of 10, 20, 30, 40, 50, 60, 70, y 80 EM. S indicates that there is a
statistically significant difference (significance level 0.01) between heading directions under the corresponding two
condiciones. Each experimental setup was repeated 10 times to achieve the mean heading direction. Notablemente, allá
is no statistically significant difference within 30–50 ms, and statistically significant difference between 30–50 ms and
experiments with syllable periods lying outside this interval.
syllable
período
10
20
30
40
50
60
70
80
10
20
S
S
S
S
S
S
S
30
S
S
40
S
S
50
S
S
S
S
S
S
S
S
S
S
S
70
80
S
S
S
S
S
S
S
S
60
S
S
S
S
S
S
sum. This vector mean summarizes the average heading of the robot with respect to
the loudspeaker direction and is plotted on the polar plot as a dot (*). A direct path
from starting position to the loudspeaker would therefore be plotted as (0,1)—the angle
would be 0 and the directionality of the path would be 1. Cifra 9 shows only 1 de
10 replications of each experimental setup (for space limitation reasons). En cada caso,
the robot is started on the same side of the speaker, which means that in all cases
where the robot does not react to the sound source (es decir., 10, 20, 60, 70, y 80 EM),
the vectors and mean are around 90 degrees. This is biased by the starting position,
so starting the robot to the other side of the speaker resulted in vectors and mean
around −90 degrees. For the statistical analyses, the vectors from these experiments
were subtracted from zero degrees to obtain similar testing situations.
We can then perform statistical tests on the vector means in the different groups of
experiments to achieve statistical evidence of the phonotaxis behavior (discriminación
between sounds with different syllable rates). Aquí, we use the statistical U-test and
obtain the table shown in Table 1.
For the experiments with songs with varying syllable rates, the statistical analysis
shows that there is a statistically significant difference in the robot’s performance be-
tween experiments with syllable periods in the interval 30–50 ms and all other syllable
periods. This is a verification of the biological data—but here we show it with a much
simpler control system than has been hypothesized by most biologists (p.ej., Schild-
berger, personal communication).
7 Discusión
We have hypothesized a model for explaining the band-pass filtering found in crickets.
The model is an extension of the simpler one with which we previously showed fre-
quency selectivity [5]. The experiments presented here show that the extended model
results in a behavior similar to the behavior resulted from the previous model, pero
most importantly, the syllable rate experiments show that the model results in band-
pass filtering similar to that found in biological experiments. The statistical analysis
of the heading directions shows that there is a statistical difference in heading direc-
tion between the conditions where 30–50 ms syllable rate songs are presented and the
conditions with syllable rates outside this interval.
Although we have described this as a neural model, it is more properly a “neuron-
104
Artificial Life Volume 4, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 9. The robot’s heading in experiments with songs with different syllable rates. The experiments are with
syllable intervals of 10, 20, 30, 40, 50, 60, 70, y 80 EM (ordered left to right from top to bottom). Aquí, we show
data from only 1 del 10 runs for each syllable rate. C(cid:176) Lund, Hallam, & Webb, 1998.
Artificial Life Volume 4, Número 1
105
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
like” algorithm. N1 and N2 are not intended to correspond explicitly to specific neurons
in the cricket but represent processes we believe are carried out by small numbers (3–
10) of neurons in the cricket prothoracic ganglion and brain. En [11] we have described
how these processes might be mapped onto specific neurons, and we are currently
working on implementing the spiking neuron controller described there on the new
robot base.
Sin embargo, the way in which our model controls the band-pass response to syllable
rates is a plausible “high-level” model of the cricket controller and provides a number
of useful insights for interpreting behavioral and physiological cricket research.
En
particular, it is the simple interactions of decay rates and thresholds in our model that
determine the effectiveness of different signals.
The neural band-pass filtering is dependent on the neural characteristics, a saber
the decay rate and firing thresholds of N1 and N2, and the correspondence between
the two. If T 1low is lowered, then the syllable period has to be increased for the N1
activation to drop below T 1low in between syllables, so the robot would respond only
to trills with a larger syllable period. If T 1low is set higher, then N1 would be able to
fire on trills with a lower syllable period. Sin embargo, this also depends on the decay
tasa. Por otro lado, N2 allows only trills with a syllable period up to a certain level
to pass. If the syllable period is too long, N2 activation will drop too much in between
activations from N1 and it will never be able to reach T 2high.
With the decay rates and thresholds set in these experiments, N1 works as a low-pass
filter, while N2 works as a high-pass filter of syllable rates.
This also provides a simple method by which the variation in syllable rate preference
can be explained. Small parameter changes in decay rates and thresholds can set the
preference of the cricket at different values; thus genetic predisposition to species-
specific rates can evolve. Adaptation in these parameters during the cricket’s lifetime
can explain the age-related change in bandwidth of preference reported elsewhere
[9, 12].
A more complex issue is raised by the biological data regarding the syllable duty-
ciclo: “one can alter the duty cycle. . . of the values to ca. 90%; the sound remains
attractive as long as the syllable repetition rate is near 30Hz” ([12], pag. 319). At first
glance our model suffices to explain this effect because it is only the onset of syllables
that controls behavior, and the length of syllable is irrelevant.
In fact the issue is
more complex. For a short duty-cycle, our model will produce taxis behavior provided
the amplitude of the signal is sufficient to sum to threshold (T 1high) before the short
syllable ends (this depends on details of the summation rate and the value of T 1high).
Consequently it is interesting to note that short syllables do need to be louder to be
equally attractive to female crickets. For long duty-cycles our current model is more
problematic, because the decay rate and T 1low set a minimum length of gap between
If we increase T 1low to decrease the length of gap required (p.ej., for a
syllables.
90% duty-cycle the gap would be around 5 EM), we also change the low-pass filtering
properties such that songs with syllable periods of 10 ms should be easily trackable.
We believe this points to an important experiment for cricket neuroethology: No one
tiene (a nuestro conocimiento) explored the response of auditory interneurons to long duty-
cycle songs. Unless these neurons have a more complex characteristic than low-pass
filtering, the ability to track 90% duty-cycle songs is an anomaly. De hecho, there is recent
evidence on measuring neuron response to different duty-cycles showing that a gap
de 5 ms in a 35 ms syllable interval song is not detectable in the firing pattern of AN1
neuronas (Tschuluun & Schildberger, personal communication).
Our new robot should enable us to explore a wide variety of other interesting ques-
tions in cricket behavior. One is the evolutionarily interesting issue of choice by female
crickets between differing males. Our preliminary tests in this area suggest that sen-
106
Artificial Life Volume 4, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
h. h. Lund, B. Webb, y j. Hallam
Physical and Temporal Scaling
sory bias may play a significant role. Another issue is the interaction of taxis with the
auditory escape response.
Our exploration of the phonotaxis system has re-emphsised the close relationship of
physical and temporal scales with the control systems underlying behavior. A generic
simulation approach may tell us little about real problems in approach behavior. Por
investigating a specific biological system and modeling it at a level of detail driven
by biological questions, we gain more sophisticated insights into the real problems of
sensorimotor control.
Expresiones de gratitud
This work was supported by EPSRC grant nr. GR/K 78942 and The Danish National Re-
search Councils. Facilities were provided by the Danish National Centre for IT Research,
and the Universities of Edinburgh and Nottingham.
Referencias
1. Campana, W.. J., Kipp, l. r., & collins, R. D. (1995). The role of chemo-orientation in search
behaviour. En R. t. Carde & W.. j. Campana (Editores.), Chemical ecology of insects 2 Nueva York:
Chapman and Hall.
2. Braitenberg, V. (1984). Vehicles: Experiments in synthetic psychology. Cambridge, MAMÁ: CON
Prensa.
3. Doherty, j. A. (1985). Phonotaxis in the cricket, Gryllus bimaculatus degeer: Comparison
of choice and no-choice paradigms. Journal of Comparative Physiology A, 157, 279–289.
4. Doherty, j. A. (1985). Temperature coupling and trade-off phenomena in the acoustic
communication system of the cricket, Gryllus bimaculatus degeer (gryllidae). Diario de
Experimental Biology, 114, 17–35.
5. Lund, h. h., Webb, B., & Hallam, j. (1997). A robot attracted to the cricket species Gryllus
bimaculatus. In P. Husbands & I. harvey (Editores.), Proceedings of the Fourth European
Conference on Artificial Life (páginas. 246–255). Cambridge, MAMÁ: MIT Press/Bradford Books.
6. Popov, A. v., & Shuvalov, V. F. (1977). Phonotactic behaviour of crickets. Diario de
Comparative Physiology A, 119, 111–128.
7. Schildberger, k. (1984). Temporal selectivity of identified auditory neurons in cricket brain.
Journal of Comparative Physiology A, 155, 171–185.
8. Schildberger, K., & Horner, METRO. (1988). The function of auditory neurons in cricket
phonotaxis I. Influence of hyperpolarization of identified neurons on sound localization.
Journal of Comparative Physiology A, 163, 621–631.
9. Stout, j. F., & McGhee, R. W.. (1988). Attractiveness of the male Acheta domesticus calling
song to females II. The relative importance of syllable period, intensidad, and chirp rate.
Journal of Comparative Physiology A, 164, 277–287.
10. Webb, B. (1995). Using robots to model animals: A cricket test. Robotics and Autonomous
Sistemas, 16, 117–134.
11. Webb, B, & Scutt, t. (1997). A simple latency-dependent spiking-neuron model of cricket
phonotaxis. Manuscript submitted for publication.
12. Weber, T., & Thorson, j. (1989). Phonotactic behavior of walking crickets. In F. Huber,
t. mi. moore, & W.. Loher (Editores.), Cricket behaviour and neurobiology (páginas. 310–339). Ítaca,
Nueva York: Prensa de la Universidad de Cornell.
Artificial Life Volume 4, Número 1
107
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
/
4
1
9
5
1
6
6
1
5
9
6
1
0
6
4
5
4
6
9
8
5
6
8
4
6
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3