PET Evidence for a Role for Striatal Dopamine in the
Attentional Blink: Functional Implications
Heleen A. Slagter1, Rachel Tomer2, Bradley T. Christian3,
Andrew S. Fox3, Lorenza S. Colzato4, Carlye R. King3,
Dhanabalan Murali3, and Richard J. Davidson3
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
Abstracto
■ Our outside world changes continuously, Por ejemplo, cuando
driving through traffic. An important question is how our brain
deals with this constant barrage of rapidly changing sensory
input and flexibly selects only newly goal-relevant information
for further capacity-limited processing in working memory.
The challenge our brain faces is experimentally captured by
the attentional blink (AB): an impairment in detecting the sec-
ond of two target stimuli presented in close temporal proximity
among distracters. Many theories have been proposed to ex-
plain this deficit in processing goal-relevant information, con
some attributing the AB to capacity limitations related to en-
coding of the first target and others assigning a critical role to
on-line selection mechanisms that control access to working
memory. The current study examined the role of striatal dopa-
mine in the AB, given its known role in regulating the con-
tents of working memory. Específicamente, participants performed
an AB task and their basal level of dopamine D2-like receptor
binding was measured using PET and [F-18]fallypride. As pre-
dicted, individual differences analyses showed that greater
D2-like receptor binding in the striatum was associated with
a larger AB, implicating striatal dopamine and mechanisms
that control access to working memory in the AB. Específicamente,
we propose that striatal dopamine may determine the AB
by regulating the threshold for working memory updating, pro-
viding a testable physiological basis for this deficit in gating
rapidly changing visual information. A challenge for current
models of the AB lies in connecting more directly to these neuro-
biological data. ■
INTRODUCCIÓN
In an ever-changing world, our senses are continuously
bombarded with more information than our brain can
process up to the level of awareness. Por eso, the ability
to rapidly select goal-relevant information when it occurs
and inhibit irrelevant or distracting information is central
to goal-directed behavior. The challenge our brain faces
when presented with an overwhelming amount of infor-
mation to analyze is captured by one of the most studied
attentional phenomena in the literature: the so-called
attentional blink (AB) deficit (raimon, Shapiro, & Arnell,
1992). This deficit occurs when people have to detect two
target stimuli (T1 and T2) presented in close temporal
succession and among distracter events. Específicamente, cuando
T2 follows T1 within 100–500 msec, it often goes un-
noticed. Different models have been proposed to explain
this deficit in selective information processing, with many
early models attributing the AB to limitations in processing
capacity (Shapiro, raimon, & Arnell, 1997). The general
idea underlying these models (p.ej., Chun & Potter, 1995)
is that, to become available for conscious report, a stimulus
1University of Amsterdam, 2University of Haifa, 3Universidad de
Wisconsin, 4Universidad de Leiden
has to be encoded and consolidated in working memory,
a process that is assumed to consume limited processing
recourses. If these resources are allocated to consolida-
tion of T1, fewer resources are available for T2, representación
the sensory representation of T2 more susceptible to
decay and interruption by distracters, so that T2 is less
likely to be maintained and reported later.
Todavía, limited-capacity models have difficulty explaining
recent behavioral findings showing that individuals can
easily process up to four targets presented in close tem-
poral proximity, as long as there is no intervening dis-
tracter stimulus (Di Lollo, Kawahara, Shahab Ghorashi,
& Enns, 2005). Esta observación, as well as others (p.ej.,
Olivers, van der Stigchel, & Hulleman, 2007; Kawahara,
Kumada, & Di Lollo, 2006), have stimulated the develop-
ment of novel theories that posit that the AB results from
on-line selection mechanisms that act in response to dis-
tracting input rather than being the result of T1-induced
cognitive resource depletion (for recent reviews, ver
Martens & Wyble, 2010; Dux & Marois, 2009). Por ejemplo,
it has been postulated that individuals apply an atten-
tional set (or filter) for targets and against distracters and
that the AB results from disruption of the input control fil-
ter by T1 processing (Di Lollo et al., 2005) or an inhibitory
response meant to suppress D1, but which accidentally
© 2012 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 24:9, páginas. 1932–1940
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
oh
8
C
3
norte
4
_
4
a
/
_
j
0
oh
0
C
2
norte
5
5
_
a
pag
_
d
0
0
b
2
y
5
gramo
5
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
t
.
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
suppresses the subsequently presented T2 (Olivers &
Meeter, 2008).
Hasta ahora, the majority of AB accounts make inferences
about the exact processes that contribute to the AB solely
based on the indirect measure of performance. Estudios
examining the neurobiological mechanisms underlying
the AB may provide additional information critical for
understanding this deficit in gating rapidly changing vi-
sual information. The current study examined the role
of striatal dopamine in the AB given its known role in
controlling information flow into working memory (p.ej.,
Cools & DʼEsposito, 2011; Franco, Loughry, & OʼReilly,
2001; Más valiente & cohen, 1999). Many AB theories empha-
size the importance of working memory for consolidat-
ing target information (Martens & Wyble, 2010; Dux
& Marois, 2009). En efecto, behavioral work has shown
that the size of the AB is related to operating resources
within working memory, including oneʼs ability to keep
irrelevant information out of working memory (Arnell,
alimenta, MacLean, & Gicante, 2010; Arnell & Stubitz, 2010;
Martens & Valchev, 2009; Dux & Marois, 2008; Colzato,
Spapé, Pannebakker, & Hommel, 2007). Notablemente, un gran
body of research has linked working memory operations
to interactions between the striatum and frontal cortex
and has revealed a critical role for the neurotransmitter
dopamine in regulating these interactions. En particular,
whereas frontal dopamine appears critical for maintain-
ing working memory representations and thus stability of
comportamiento, striatal dopamine is thought to be critical for
the updating of frontal working memory representations
and hence, flexibility of behavior (p.ej., Cools & DʼEsposito,
2011; Frank et al., 2001; Más valiente & cohen, 1999). En efecto,
striatal dopamine has been shown to promote cognitive
flexibility by allowing updating of newly relevant repre-
sentations in working memory and preventing gating of
nonrelevant representations (p.ej., Wang, Vijayraghavan, &
Goldman-Rakic, 2004; Frank et al., 2001). On the basis of this
trabajar, we previously hypothesized that striatal dopamine
may play an important role in the AB by regulating which
stimuli enter working memory (Colzato, Slagter, de Rover,
& Hommel, 2011; Colzato, Slagter, Spapé, & Hommel,
2008). Consistent with this hypothesis, we found that indi-
vidual AB size is predicted by spontaneous eye blink rate, a
marker of striatal dopaminergic functioning (Karson, 1983),
and by the C957T polymorphism at the DRD2 gene (associ-
ated with striatal D2 receptors; Colzato et al., 2008, 2011).
These observations provide preliminary, albeit indirect,
evidence for a role of striatal dopamine in the AB.
The current study tested the hypothesis that striatal
dopamine and in particular striatal D2 receptors play a role
in the AB more directly. Para tal fin, participants took
part in three sessions: (1) an AB task session, (2) a PET ses-
sion in which baseline D2-like (D2/D3) receptor availabil-
ity was measured using the ligand [F-18]fallypride at rest,
y (3) an MRI session for anatomical localization purposes.
Our main prediction was that individual variation in striatal
D2-like receptor binding would predict individual AB size.
Notablemente, D2 receptors are expressed primarily in the striatal
indirect pathway (Gerfen, 1992), which is thought to prevent
gating of irrelevant representations into working memory
(Hazy, Franco, & OʼReilly, 2007a; OʼReilly & Franco, 2006).
An association between striatal D2-like receptor binding
and AB size would thus indicate that striatal dopaminergic
functioning may represent an important neurobiological
mechanism underlying distracter suppression efficiency
and hence the ability to rapidly and flexibly gate only newly
relevant information into working memory for further pro-
cessing in the AB task.
MÉTODOS
Participantes
Fourteen right-handed adults (nine women, 19–29 years,
edad media = 20.2 años) participated in this study. Solo
healthy participants without history of developmental
disorders, head trauma, psychiatric or neurological dis-
ease, and drug abuse were included. All had normal or
corrected-to-normal visual acuity. Participants were asked
to abstain from eating, drinking (other than water), y
smoking for minimally 4 hr before the PET session. Ellos
were compensated for participation. The study was ap-
proved by the local ethics committee.
Experimental Design and Procedure
Data were collected in three sessions, separated by 1–
4 semanas: a behavioral session, a PET imaging session, y
an MRI session.
Behavioral Session
Participants performed an AB task adopted from Slagter
et al. (Slagter, Lutz, Greischar, Nieuwenhuis, & Davidson,
2009; Slagter et al., 2007). They had to identify two digits
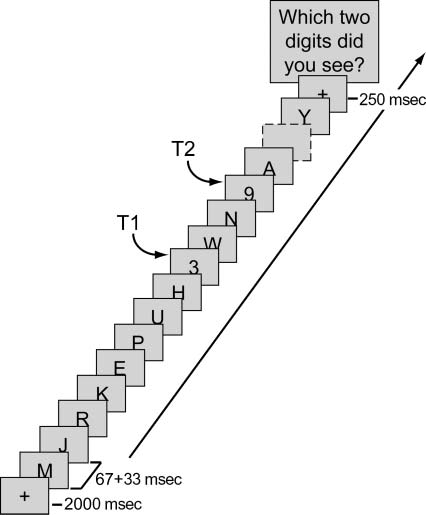
presented in a rapid stream of letters (Cifra 1). Cada prueba
started with a fixation-plus sign (+; 2000 mseg). Después
a blank interval (250 mseg), the stimulus stream began,
consisting of 20 elementos, each presented in black on a gray
fondo (rojo, verde, azul: 70, 70, 70) at the center
of the screen (16 point Times New Roman) para 67 mseg,
followed by a 33-msec blank. Each letter was randomly
drawn (without replacement) from the alphabet (excepto
I, oh, S, and Z). Digits were drawn randomly from the set
1–9. T1 position was varied randomly between 7 y 9.
T2 was presented directly thereafter (Lag 1) or after two,
four, or seven distracters (at Lags 3, 5, o 8, respectivamente).
Previous work using a highly similar task has shown a large
AB at Lag 3 compared with Lag 8 posición (Colzato et al.,
2008, 2011). Both targets were to be reported 250 mseg
after the stream by pressing the corresponding digit keys.
Participants first practiced the task for 24 ensayos. Entonces, ellos
performed two blocks of 48 trials each, Resultando en 24 ensayos
per lag condition, all intermixed within blocks.
Slagter et al.
1933
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
oh
8
C
3
norte
4
_
4
a
/
_
j
0
oh
0
C
2
norte
5
5
_
a
pag
_
d
0
0
b
2
y
5
gramo
5
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
F
.
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
MRI Data Acquisition Session
Anatomical brain images were acquired on a 3T-GE Signa
scanner, equipped with high-speed gradients and a whole-
head transmit–receive quadrature birdcage headcoil (GE
Medical Systems). Anatomical scans consisted of a high-
resolution 3-D T1-weighted, inversion–recovery fast-
gradient-echo image (inversion time = 600 mseg, in-plane
resolution = 256 × 256, campo de visión = 240 mm, 124 ×
1.1 mm axial slices), and a T2-weighted, fast-spin echo
imagen (in-plane resolution = 256 × 256, campo de visión =
240 mm, 81 × 2 mm sagittal slices).
Análisis de los datos
Behavioral Data Analysis
T1 and T2 accuracy data were submitted to separate
repeated-measures ANOVAs with Lags (1, 3, 5, y 8)
as a within-subject factor. T2 accuracy was based only
on those trials in which T1 was correctly reported.
Brain Imaging Data Analysis
PET data were reconstructed using a filtered back-projection
algorithm with sinogram trimming; axial and in-plane
smoothing (4 mm Gaussian filter) to a voxel size of
1.84 mm × 1.84 mm × 2.43 mm; and corrected for ran-
dom events, annihilation radiation attenuation, deadtime,
scanner normalization, and scatter radiation. The recon-
structed PET time series was then inspected and corrected
for head motion using the SPM2 coregistration (www.fil.
ion.ucl.ac.uk/spm) algorithm based on frame-to-frame
coregistration to an early integrated reference image. El
cerebellar time–activity curve was extracted from the PET
data based on an ROI drawn on the cerebellar lobes of the
early PET data. Parametric images of distribution volume
ratios (DVRs) were generated using this time course to
represent the behavior of the radiotracer in brain regions
with negligible binding (Mukherjee et al., 2002). El
DVR parameter represents an index that is proportional
to the concentration of available D2-like binding sites
(Bmax), given by the relationship:
DVR ¼ ðBmax=KDÞfND þ 1;
where KD is the apparent (in vivo) equilibrium disso-
ciation constant and fND is the free fraction of radiotracer
in the brain tissue (Innis et al., 2007). A multilinear ap-
proach was used to generate the DVR estimates using the
data starting at 39 mín. (t*) until the end of the acquisition
(Ichise, Toyama, Innis, & carson, 2002; Logan et al., 1996).
The DVR parametric images were spatially coregistered
to the same participantʼs T1-weighted MRI images using
the FMRIB Software Library (FSL) linear registration tool
(Jenkinson & Herrero, 2001).
T1-weighted MRI images were manually masked to
exclude nonbrain tissues. These “skull-stripped” images
Cifra 1. The attentional blink task. Participants had to detect two
objetivos (digits) in a rapid stream of distracters (letters). T2 followed
T1 at Lags 1, 3, 5, o 8.
PET Data Acquisition Session
PET data were acquired using a Siemens EXACT HR+
PET scanner in 3-D mode (septa retracted) and the high-
affinity D2/D3 radioligand [F-18]fallypride. Individual dif-
ferences in D2-like receptor availability as measured by
[F-18]fallypride are stable across time (Mukherjee et al.,
2002) and thus provide an index of stable individual differ-
ences in striatal dopaminergic neurotransmission. [F-18]
fallypride expresses the highest affinity for D2 receptors,
but also binds to D3 receptors and (albeit weakly) to D4
receptors. The subjects were positioned head first, supine
with the cantho-meatal line parallel to the in-plane field
of view. The head rests in the scanner head holder extend-
ing from the patient bed and held in place by surgical tape
placed firmly across the subjectʼs forehead. A 5-min trans-
mission scan was then acquired to correct for the attenua-
tion of the gamma rays within the tissue. The acquisition of
the dynamic [F-18]fallypride PET scan was initiated with
the injection of radioligand (237 ± 43 MBq). The synthesis
de [F-18]fallypride was carried out using previously re-
ported methods (Mukherjee, Cual, El, & Marrón, 1995).
The final sterile 0.9% saline solution of [F-18]fallypride
was produced with radiochemical purity greater than
95% and specific activity of 227 ± 140 GBq/micromol.
A 150-min dynamic acquisition was acquired, initiating
with the 30-sec bolus infusion of radiotracer. The time
series were binned into six 1-min frames and 48 3-mín.
marcos.
1934
Revista de neurociencia cognitiva
Volumen 24, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
oh
8
C
3
norte
4
_
4
a
/
_
j
0
oh
0
C
2
norte
5
5
_
a
pag
_
d
0
0
b
2
y
5
gramo
5
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
F
/
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
for each participant were coregistered to the MNI-152 tem-
plate packaged with FSL. The resulting standard-space,
skull-stripped T1-weigted MRI images were averaged
to create single-participant templates in standard space.
Small differences in brain morphology were accounted
for by performing a nonlinear warp using FSLʼs nonlinear
registration tool (www.fmrib.ox.ac.uk/analysis/techrep/
tr07ja2/tr07ja2.pdf) to align each participantʼs standard-space
skull-stripped T1-weighted MRI images to their single-subject
template. These transformations were then combined
with the DVR to T1-weighted MRI transforms and ap-
plied to the DVR images to create binding images. Ser-
cause differences observed in across-subject analyses
might result from individual differences in brain anatomy
( Woodward et al., 2009; Oakes et al., 2007) en vez de
true differences in binding, we assessed the probability of
gray matter (GMP) at each voxel in each participantʼs brain
based on the skull-stripped standard-space T1-weighted
MRIs using FSLʼs automated segmentation tool (zhang,
Brady, & Herrero, 2001). Before statistical analyses, all im-
ages were blurred using an 8-mm FWHM Gaussian fil-
ter to account for potential across participant differences
in anatomy.
Our main prediction was that striatal D2-like recep-
tor binding would be predictive of individual differences
in AB size, defined as the difference in T2 accuracy be-
tween Lag 8 and Lag 3 ensayos. To examine this prediction,
we correlated individual differences in receptor binding
with individual differences in AB size (while controlling
for GMP at each voxel) using Spearmanʼs rank correlation).
Only voxels with an average dopamine binding value of 1.2
or greater (es decir., greater than 1.2 times the cerebellar bind-
ing rate) were included in the correlation analysis to ex-
clude most white matter regions. Significance testing was
carried out via a two-stage permutation testing (Nichols
& holmes, 2002). At the first stage (voxel level), as a first
step, GMP was regressed out of D2-like receptor binding.
Entonces, participant identity was randomly shuffled, y el
Spearmanʼs correlation between AB size and the residua-
lized binding values was computed again at each voxel.
This was repeated 1,000 veces, generating a distribution
of correlation coefficients at each voxel under the null hy-
pothesis of no relationship between AB size and D2-like re-
ceptor binding (while controlling for GMP). Statistical Z
values were taken as the normalized distance of the real
correlation coefficient compared with the null distribution.
Voxels with a Z value greater than 2.6 ( pag < .005) were re-
tained as being significant at the voxel level. In the second
stage (cluster level), Z values were computed based on one
of the 1,000 random permutation iterations, and the statis-
tical map was thresholded again. This time, the number of
voxels in the largest suprathreshold cluster was stored. This
was repeated 500 times, generating a distribution of max-
imum cluster sizes under the null hypothesis. The cluster
threshold was defined as the standardized distance from
the mean of the maximum cluster distribution correspond-
ing to p < .05 (20 contiguous voxels).
RESULTS
Behavioral Results
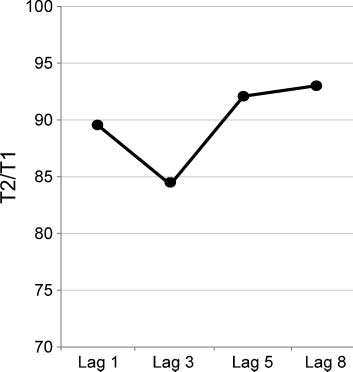
As expected and shown in Figure 2, participants exhibited
a marked AB, with lower performance at Lag 3 than at
Lags 1, 5, and 8 (main effect of Lag: F(3, 39) = 3.6, p =
.022). This pattern was confirmed by post hoc polynomial
contrasts revealing a significant cubic trend, F(1, 13) = 4.8,
p = .046. Furthermore, in line with the literature (e.g.,
Martens, Munneke, Smid, & Johnson, 2006), there was
large variability among participants in the size of the AB,
as indexed by the difference in T2 accuracy between Lags
8 and 3. AB size varied across individuals between −7.8%
and 40.7%. T1 accuracy did not differ significantly be-
tween lags, F(3, 39) < 1, p = .69. T1 accuracy was overall
high: 99%, 99%, 100%, and 99% at Lags 1, 3, 5, and 8,
respectively.
D2-like Receptor Binding and AB Size
Our main prediction was that individual variation in
AB size would be associated with individual variation in
striatal D2-like receptor binding, as measured in a separate
session using PET and the radioligand [F-18]fallypride
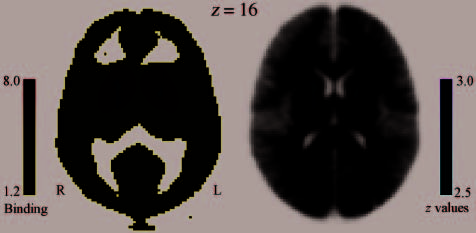
during resting conditions. Confirming this prediction, voxel-
wise correlation analysis revealed a significant correlation—
after correction for GMP—between AB size and D2-like
receptor binding in the dorsal striatum, bilaterally and
in the vicinity of the caudate nucleus and globus pallidus
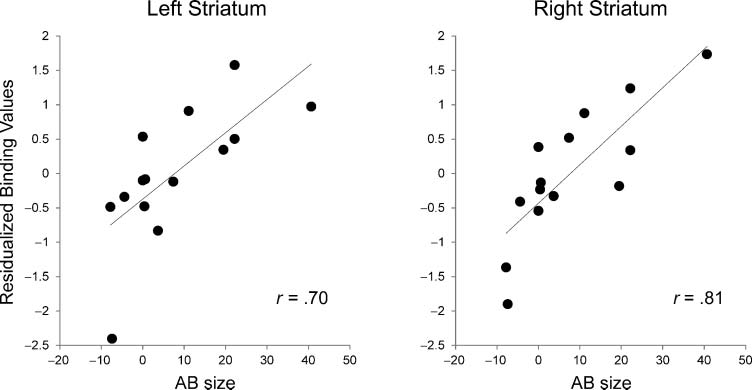
(Figure 3). More specifically, in both the left and the right
striatum, higher binding was associated with a larger AB to
T2, as can be seen in Figure 4. This scatter plot displays
the correlation between AB size and binding for the voxel
showing the highest correlation for each striatal cluster
separately. The xyz MNI coordinates of the voxel showing
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
o
8
c
3
n
4
_
4
a
/
_
j
0
o
0
c
2
n
5
5
_
a
p
_
d
0
0
b
2
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. AB task performance. Average detection rates of T2 (contingent
on T1 detection) are displayed as a function of lag. This figure shows a
marked decrease in T2 detection rate at Lag 3, that is, an AB.
Slagter et al.
1935
binding, and the AB, a deficit in processing two meaningful
events presented in close temporal proximity and among
distracter events. As predicted, increased D2-like receptor
availability in the striatum was associated with a larger AB
to T2. This novel finding supports the idea that striatal
dopaminergic neurotransmission; hence, mechanisms in-
volved in dynamically regulating the contents of working
memory play an important role in the AB. It corroborates
previous, more indirect evidence implicating the striatum,
and in particular the striatal D2 pathway, in the AB (Colzato
et al., 2008, 2011; Slagter, Johnstone, Beets, & Davidson,
2010).
Increased D2-like receptor binding in the striatum in
individuals with a relatively large AB may correspond to
higher receptor densities or lower levels of (competing)
endogenous dopamine or both. It is notable in this respect
that genetic overexpression of striatal D2 receptors in
rodents causes impairments in set shifting (Kellendonk
et al., 2006). Furthermore, a lower basal level of endoge-
nous striatal dopamine in individuals with a large AB would
be in line with our previous observation that these indi-
viduals show a relatively low spontaneous eye blink rate
(sEBR), an index of striatal dopaminergic function (Colzato
et al., 2008; Karson, 1983).
An important question for future research is how pre-
cisely striatal dopamine may determine the AB. As men-
tioned in the Introduction, previous work has shown that
high levels of striatal dopamine promote cognitive flexi-
bility by permitting updating of frontal working memory
representations, whereas low striatal dopamine levels are
associated with cognitive stability and little updating
in response to novel relevant information (for recent re-
views, see, e.g., Cools & DʼEsposito, 2011; Maia & Frank,
2011). Our observation that more D2-like receptor bind-
ing (hence, likely a lower level of [competing] endogenous
dopamine) was generally associated with a larger AB is in
line with the notion that low striatal dopamine levels pro-
mote stabilization of frontal task-relevant representations
Figure 3. Regions of the striatum showing a significant relationship
between D2-like receptor binding and individual AB size. Displayed
is the axial section (z = 16) of the statistical map of the correlation
between individual [F-18]fallypride binding and AB size overlaid
upon (on the left) the group-averaged binding map and (on the
right) the average MRI of all participants in MNI space.
the highest correlation (r(12) = .70, p = .005) within the
left striatal cluster were: −16, −2, 18. The right striatal clus-
ter contained two peaks. The xyz MNI coordinates of
the first peak (r(12) = .81, p < .001) were 18, −8, 18, and
the xyz MNI coordinates of the second peak (r(12) = .71,
p = .005) were 20, 12, 12.
In contrast to several other D2 radioligands (e.g.,
[C-11]raclopride), [F-18]fallypride provides sufficient
signal-to-noise ratio to quantify D2-like binding in extra-
striatal areas, such as cortex, where the concentration of
D2 receptors is much lower than that in the striatum.
Yet, no other brain region exhibited a relationship (nega-
tive or positive) between AB size and D2-like receptor
availability, even at a more lenient voxel-level threshold
of p < .01.
DISCUSSION
In this PET study, we examined the relationship between
striatal dopamine, as indexed by baseline D2-like receptor
Figure 4. Striatal D2-like
receptor binding predicted
AB size across participants.
Correlation plots showing
the significant cross-subject
relationship between D2-like
receptor binding and AB size,
separately for the left (left
panel) and the right (right
panel) striatum. Binding
values are based on the
peak voxel within each
striatal cluster. Non-ranked
values are shown for
illustrative purposes.
1936
Journal of Cognitive Neuroscience
Volume 24, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
o
8
c
3
n
4
_
4
a
/
_
j
0
o
0
c
2
n
5
5
_
a
p
_
d
0
0
b
2
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(Cools & DʼEsposito, 2011; Maia & Frank, 2011). How-
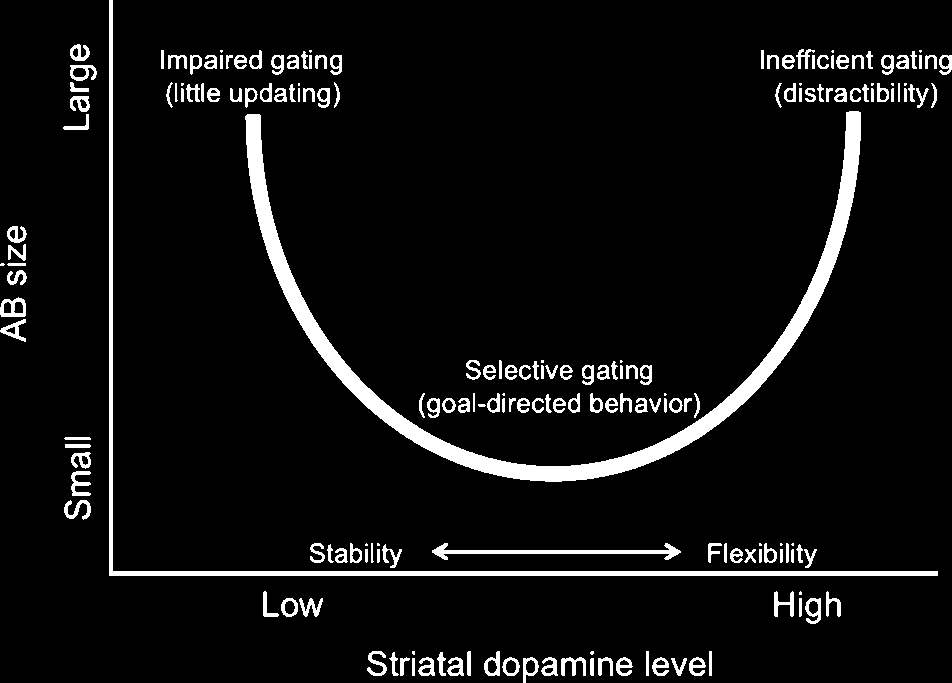
ever, previous research also indicates that excessivly high
striatal dopamine levels may impair performance as well,
and it has been suggested that the relationship between
cognitive performance and striatal dopamine level follows
a U-shaped function (summarized in Cools & DʼEsposito,
2011), where both too little and too much striatal dopa-
mine can hurt performance due to, respectively, impaired
gating and inefficient gating (and contaminant distractibil-
ity). Our data, although indicative of a linear relationship,
are not inconsisent with a U relationship between D2-like
receptor binding and AB size, as it is conceivable that
healthy subjects display a relatively restricted range in
baseline dopamine level during resting conditions. On
the basis of the larger literature on working memory and
dopamine, we therefore hypothesize that striatal dopa-
mine more likely exhibits a U relationship with AB size
(see Figure 5), such that intermediate tonic dopamine lev-
els promote optimal gating (or selective target updating)
and hence a small AB,whereas both excessively low and ex-
cessively high tonic dopamine levels produce a large AB,
albeit for different reasons. Whereas high dopamine levels
will lead to more overall working memory updating and
concomitant distractibility and a large AB (i.e., they also
induce distracter updating, causing interference in work-
ing memory), very low levels will also be associated with
a large AB due to a general poor updating ability (not
enough dopamine to update even target information).
Note that in the latter case in particular (general poor
Figure 5. Hypothesized U relationship between tonic dopamine
level and AB size. Dopamine level is hypothesized to determine
the AB by regulating the threshold for working memory updating.
Specifically, we postulate that AB size exhibits a U relationship with
striatal dopamine level, such that (i) excessively high dopamine
levels will result in inefficient gating (they also induce distracter
updating, causing interference in working memory) and hence a
large AB, (ii) intermediate dopamine levels will promote optimal
gating (selective target updating) and hence a small AB, and (iii)
very low levels will be associated with poor updating ability (not
enough dopamine to update even target information) and both
poor T1 accuracy and a large AB to T2.
updating ability), one would also expect poor T1 accuracy.
In the current study, with healthy individuals and T1 accu-
racy at ceiling level (99–100%), no relationship between
D2-like binding and T1 accuracy was observed. Yet, it is
of note that in the study by Colzato et al. (2008), the group
of subjects with relatively low sEBR scores not only dis-
played a larger AB to T2 but also lower T1 accuracy com-
pared with the group of subjects with relatively high sEBR
scores. This observation is in line with the idea that low
striatal dopamine levels lead to an overall impairment in
updating ability. Yet, future research in which dopamine
level is experimentally manipulated across a wide range
is necessary to determine whether or not the relationship
between AB size and dopamine level follows the proposed
U-shaped function.
In summary, we hypothesize that striatal dopamine
determines the AB by regulating the threshold for work-
ing memory updating. In this framework, the AB can be
due both to a general poor updating ability, when the
threshold for working updating is set too strongly (and
striatal dopamine levels are very low), and to distracter
interference in working memory, when the threshold
for updating is set too lenient (and striatal dopamine lev-
els are very high). This account, which highlights a neuro-
biological mechanism through which rapid input control
might be accomplished, is in line with the notion that
the AB results from online selection mechanisms that
control information flow into working memory rather
than being the result of T1-induced cognitive resource
depletion (see also below). Yet, future studies, includ-
ing pharmacological, patient, and neurocomputational
studies, are necessary to determine the precise relation-
ship (i.e., linear or U-shaped) between basal striatal dopa-
mine activity and the ability to gate only newly task-relevant
information into working memory under rapidly changing
input conditions, such as the AB. This work should also
address the relationship between tonic dopamine levels
and phasic dopamine responses. Phasic dopamine in the
striatum has been related to selective reinforcement of
gating for task-relevant stimulation (e.g., Maia & Frank,
2011), and large phasic responses may hence promote
selective target updating in the AB task. Future work
should also look into the possible role of striatal dopa-
mine receptor features in the AB. As pointed out above,
D2-like receptor binding is proportional to both receptor
density and endogenous dopamine, and on the basis of
the current data alone, it is unclear to what extent the
AB may be related to endogenous dopamine level, D2
receptor density, and/or other receptor features, such as
affinity. Thus, more research is necessary for fine tuning
the “dopamine hypothesis” of the AB and may ultimately
lead to a mechanistic and neurobiologically grounded
model of the AB.
It is noteworthy that the regions in which D2-like re-
ceptor binding predicted AB size fall within the associa-
tive striatum. This part of the striatum forms loops with
associative regions of the cortex, such as dorsolateral pFC
Slagter et al.
1937
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
o
8
c
3
n
4
_
4
a
/
_
j
0
o
0
c
2
n
5
5
_
a
p
_
d
0
0
b
2
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(Draganski et al., 2008; Joel & Weiner, 2000). Notably,
PET and fMRI studies have linked the associative striatum
to flexible cognitive control, including operations such as
working memory updating and inhibition of irrelevant in-
formation (e.g., Kühn et al., in press; Bäckman et al.,
2011; Dahlin, Neely, Larsson, Bäckman, & Nyberg, 2008;
McNab & Klingberg, 2008). Although striatal dopamine
has been linked to many functions other than WM gating
and our measure, D2-like receptor binding, hence does not
provide a pure measure of WM gating, the location of our
effects in the associative striatum is in line with our hypoth-
esis that striatal dopamine determines the AB by regulating
the threshold for WM updating.
It is of further note that the D2 receptor is expressed
primarily in the striatal indirect pathway, which traverses
the pallidum and subthalamic nucleus before targeting
BG output structures (Gerfen, 1992). It has been posited
that the striatum facilitates or suppresses cortical actions
(e.g., working memory updating) via separate direct and
indirect pathways, respectively, that project to frontal cor-
tex and that the likelihood that a cognitive representation
is selected for updating of working memory in the frontal
cortex is proportional to the difference between activity
for that representation in the direct and indirect pathways
(Hazy, Frank, & OʼReilly, 2007b). Indeed, the indirect path-
way has previously been shown to play a critical role in fil-
tering out distracting information from being updated into
working memory (McNab & Klingberg, 2008; Moustafa,
Sherman, & Frank, 2008; Frank & OʼReilly, 2006; OʼReilly
& Frank, 2006). Given the critical role of the striatal D2/
indirect pathway in inhibiting distracting information, it is
notable that recent behavioral studies have shown that
individual differences in the ability to inhibit distracting
information determine AB size (Arnell & Stubitz, 2010;
Martens & Valchev, 2009; Dux & Marois, 2008). In addition,
the here-observed relationship between D2-like receptor
binding and AB size may thus provide a neurobiological
mechanism through which selection mechanisms control
responses to distracting input and provide support for
filter-based accounts, where the AB reflects the workings
of a mechanism designed to filter out nonrelevant infor-
mation (e.g., Olivers & Meeter, 2008; Di Lollo et al.,
2005; Raymond et al., 1992). One of these models (Olivers
& Meeter, 2008) actually postulated a possible role for
the BG in the AB based on previous findings showing
that a prefrontal/BG network regulates access to working
memory (McNab & Klingberg, 2008). Yet, our finding
could be consistent with models that attribute the AB
to the shielding of T1 processing as well (e.g., Taatgen,
Juvina, Schipper, Borst, & Martens, 2009; Wyble, Bowman,
& Nieuwenstein, 2009; Hommel et al., 2006; Jolicoeur
& DellʼAcqua, 1998). By preventing gating of irrelevant
representations, striatal D2 receptors may also promote
undisturbed consolidation of T1 in working memory.
In either case, the observed relationship between D2-
like receptor binding and the AB suggests that mecha-
nisms involved in regulating the contents of working
memory and in particular mechanisms related to sup-
pressing irrelevant information contribute significantly to
this phenomenon. A challenge for current computational
and theoretical accounts of the AB lies in connecting more
directly with these neurobiological data. Our results also
synthesize the AB literature with the largely separate litera-
ture associated with working memory and may thus in ad-
dition inform models of working memory that have not
yet had to confront rapid presentation of stimuli (e.g., Hazy
et al., 2007a; Frank et al., 2001) by providing information
regarding the timing of mechanisms involved in gating of
information.
A role for striatal dopamine in the AB also receives
support, albeit more indirectly, from several previous
studies. First, as mentioned in the introduction, Colzato
and colleagues found that sEBR, a marker of striatal do-
pamine function (Karson, 1983), predicted individual
AB size (Colzato et al., 2008) and, furthermore, that the
C957T polymorphism at the DRD2 gene (associated with
striatal D2 receptors) was associated with AB size whereas
polymorphisms associated with frontal dopamine and
norepinephrine were not (Colzato et al., 2011). In addi-
tion, people high in WM operation span—a measure asso-
ciated with striatal dopaminergic activity (Cools, Gibbs,
Miyakawa, Jagust, & DʼEsposito, 2008)—display a smaller
AB (Colzato et al., 2007). Finally, Olivers and Nieuwenhuis
(2006) reported a reduced AB when, before trial onset,
participants viewed pictures of positive affective content,
which are thought to stimulate the dopaminergic system
(Ashby, Isen, & Turken, 1999). Using PET imaging, the
current data extend these findings by providing more
direct evidence for a role of striatal dopamine and, spe-
cifically, the indirect striatal pathway in the AB. Future re-
search in which striatal dopamine levels are manipulated
pharmacologically is necessary to determine more pre-
cisely how striatal dopamine may control gating of rapidly
changing visual input.
Yet, it should be noted that it is unlikely that any single
neuromodulatory mechanism can explain the many ex-
perimental factors that are known to modulate the AB.
Other neurotransmitters and in particular norepinephrine,
given its role in temporal attention (Aston-Jones & Cohen,
2005), likely also contribute to this phenomenon. For ex-
ample, whereas striatal dopamine may modulate the thresh-
old for working memory updating, norephinephrine may
enhance cortical target processing (Nieuwenhuis, Gilzenrat,
Holmes, & Cohen, 2005). Nevertheless, together with pre-
vious findings (Colzato et al., 2008, 2011), the current data
suggest an important role for striatal dopamine and, spe-
cifically, the striatal indirect pathway in the AB and, more
generally, in flexibly prioritizing relevant over irrelevant
information.
Acknowledgments
This work was supported by the Netherlands Organization for
Scientific Research (NWO VIDI grant to H. A. S., NWO VENI
1938
Journal of Cognitive Neuroscience
Volume 24, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
o
8
c
3
n
4
_
4
a
/
_
j
0
o
0
c
2
n
5
5
_
a
p
_
d
0
0
b
2
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
grant to L. S. C.) and a grant from the National Institute of Mental
Health (MH43454 to R. J. D.). We thank two anonymous re-
viewers for their helpful comments.
Reprint requests should be sent to Heleen A. Slagter, Department
of Psychology, University of Amsterdam, Weesperplein 4, 1018 XA
Amsterdam, the Netherlands, or via e-mail: h.a.slagter@uva.nl.
REFERENCES
Arnell, K. M., Stokes, K. A., MacLean, M. H., & Gicante, C.
(2010). Executive control processes of working memory
predict attentional blink magnitude over and above
storage capacity. Psychological Research, 74, 1–11.
Arnell, K. M., & Stubitz, S. M. (2010). Attentional blink
magnitude is predicted by the ability to keep irrelevant
material out of working memory. Psychological Research,
74, 457–467.
Ashby, F. G., Isen, A. M., & Turken, A. U. (1999). A
neuropsychological theory of positive affect and its
influence on cognition. Psychological Review, 106,
529–550.
Aston-Jones, G., & Cohen, J. D. (2005). An integrative theory
of locus coeruleus-norepinephrine function: Adaptive gain
and optimal performance. Annual Review of Neuroscience,
28, 403–450.
Bäckman, L., Nyberg, L., Soveri, A., Johansson, J., Andersson,
M., Dahlin, E., et al. (2011). Effects of working-memory
training on striatal dopamine release. Science, 333, 718.
Braver, T. S., & Cohen, J. D. (1999). Dopamine, cognitive
control, and schizophrenia: The gating model. Progress
in Brain Research, 121, 327–349.
Chun, M. M., & Potter, M. C. (1995). A two-stage model for
multiple target detection in rapid serial visual presentation.
Journal of Experimental Psychology: Human Perception
and Performance, 21, 109–127.
Colzato, L. S., Slagter, H. A., de Rover, M., & Hommel, B. (2011).
Dopamine and the management of attentional resources:
Genetic markers of striatal D2 dopamine predict individual
differences in the attentional blink. Journal of Cognitive
Neuroscience, 23, 3576–3585.
Colzato, L. S., Slagter, H. A., Spapé, M. M. A., & Hommel, B.
(2008). Blinks of the eye predict blinks of the mind.
Neuropsychologia, 46, 3179–3183.
Colzato, L. S., Spapé, M., Pannebakker, M. M., & Hommel, B.
(2007). Working memory and the attentional blink: Blink
size is predicted by individual differences in operation
span. Psychonomic Bulletin & Review, 14, 1051–1057.
Cools, R., & DʼEsposito, M. (2011). Inverted-U-shaped
dopamine actions on human working memory and
cognitive control. Biological Psychiatry, 69, e113–e125.
Cools, R., Gibbs, S. E., Miyakawa, A., Jagust, W., & DʼEsposito, M.
(2008). Working memory capacity predicts dopamine
synthesis capacity in the human striatum. The Journal
of Neuroscience: The Official Journal of the Society for
Neuroscience, 28, 1208–1212.
Dahlin, E., Neely, A. S., Larsson, A., Bäckman, L., & Nyberg, L.
(2008). Transfer of learning after updating training mediated
by the striatum. Science, 320, 1510–1512.
Di Lollo, V., Kawahara, J., Shahab Ghorashi, S. M., & Enns, J. T.
(2005). The attentional blink: Resource depletion or temporary
loss of control? Psychological Research, 69, 191–200.
Draganski, B., Kherif, F., Klöppel, S., Cook, P. A., Alexander,
D. C., Parker, G. J. M., et al. (2008). Evidence for segregated
and integrative connectivity patterns in the human basal
ganglia. The Journal of Neuroscience: The Official Journal
of the Society for Neuroscience, 28, 7143–7152.
Dux, P. E., & Marois, R. (2008). Distractor inhibition predicts
individual differences in the attentional blink. PloS One, 3,
e3330.
Dux, P. E., & Marois, R. (2009). The attentional blink:
A review of data and theory. Attention, Perception &
Psychophysics, 71, 1683–1700.
Frank, M. J., Loughry, B., & OʼReilly, R. C. (2001). Interactions
between frontal cortex and basal ganglia in working memory:
A computational model. Cognitive, Affective & Behavioral
Neuroscience, 1, 137–160.
Frank, M. J., & OʼReilly, R. C. (2006). A mechanistic account
of striatal dopamine function in human cognition:
Psychopharmacological studies with cabergoline and
haloperidol. Behavioral Neuroscience, 120, 497–517.
Gerfen, C. R. (1992). The neostriatal mosaic: Multiple levels
of compartmental organization. Journal of Neural
Transmission. Supplementum, 36, 43–59.
Hazy, T. E., Frank, M. J., & OʼReilly, R. C. (2007a). Towards
an executive without a homunculus: Computational models
of the prefrontal cortex/basal ganglia system. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 362, 1601–1613.
Hazy, T. E., Frank, M. J., & OʼReilly, R. C. (2007b). Towards
an executive without a homunculus: Computational
models of the prefrontal cortex/basal ganglia system.
Philosophical Transactions of the Royal Society of
London, Series B, Biological Sciences, 362, 1601–1613.
Hommel, B., Kessler, K., Schmitz, F., Gross, J., Akyürek, E.,
Shapiro, K., et al. (2006). How the brain blinks: Towards
a neurocognitive model of the attentional blink.
Psychological Research, 70, 425–435.
Ichise, M., Toyama, H., Innis, R. B., & Carson, R. E. (2002).
Strategies to improve neuroreceptor parameter estimation
by linear regression analysis. Journal of Cerebral Blood
Flow and Metabolism: Official Journal of the International
Society of Cerebral Blood Flow and Metabolism, 22,
1271–1281.
Innis, R. B., Cunningham, V. J., Delforge, J., Fujita, M., Gjedde, A.,
Gunn, R. N., et al. (2007). Consensus nomenclature for
in vivo imaging of reversibly binding radioligands. Journal
of Cerebral Blood Flow and Metabolism: Official Journal
of the International Society of Cerebral Blood Flow and
Metabolism, 27, 1533–1539.
Jenkinson, M., & Smith, S. (2001). A global optimisation
method for robust affine registration of brain images.
Medical Image Analysis, 5, 143–156.
Joel, D., & Weiner, I. (2000). The connections of the dopaminergic
system with the striatum in rats and primates: An analysis with
respect to the functional and compartmental organization of
the striatum. Neuroscience, 96, 451–474.
Jolicoeur, P., & DellʼAcqua, R. (1998). The demonstration
of short-term consolidation. Cognitive Psychology, 36,
138–202.
Karson, C. N. (1983). Spontaneous eye-blink rates and
dopaminergic systems. Brain: A Journal of Neurology,
106, 643–653.
Kawahara, J.-I., Kumada, T., & Di Lollo, V. (2006). The
attentional blink is governed by a temporary loss of
control. Psychonomic Bulletin & Review, 13, 886–890.
Kellendonk, C., Simpson, E. H., Polan, H. J., Malleret, G.,
Vronskaya, S., Winiger, V., et al. (2006). Transient and
selective overexpression of dopamine D2 receptors in
the striatum causes persistent abnormalities in prefrontal
cortex functioning. Neuron, 49, 603–615.
Kühn, S., Schmiedek, F., Noack, H., Wenger, E., Bodammer,
N. C., Lindenberger, U., et al. (in press). The dynamics
of change in striatal activity following updating training.
Human Brain Mapping. doi:10.1002/hbm.22007
Slagter et al.
1939
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
o
8
c
3
n
4
_
4
a
/
_
j
0
o
0
c
2
n
5
5
_
a
p
_
d
0
0
b
2
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Logan, J., Fowler, J. S., Volkow, N. D., Wang, G. J., Ding, Y. S.,
& Alexoff, D. L. (1996). Distribution volume ratios without
blood sampling from graphical analysis of PET data. Journal
of Cerebral Blood Flow and Metabolism: Official Journal
of the International Society of Cerebral Blood Flow and
Metabolism, 16, 834–840.
Maia, T. V., & Frank, M. J. (2011). From reinforcement learning
models to psychiatric and neurological disorders. Nature
Neuroscience, 14, 154–162.
Martens, S., Munneke, J., Smid, H., & Johnson, A. (2006).
Quick minds donʼt blink: Electrophysiological correlates
of individual differences in attentional selection. Journal
of Cognitive Neuroscience, 18, 1423–1438.
Martens, S., & Valchev, N. (2009). Individual differences
in the attentional blink. The important role of irrelevant
information. Experimental Psychology, 56, 18–26.
Martens, S., & Wyble, B. (2010). The attentional blink:
Past, present, and future of a blind spot in perceptual
awareness. Neuroscience and Biobehavioral Reviews,
34, 947–957.
McNab, F., & Klingberg, T. (2008). Prefrontal cortex and
basal ganglia control access to working memory. Nature
Neuroscience, 11, 103–107.
Moustafa, A. A., Sherman, S. J., & Frank, M. J. (2008). A
dopaminergic basis for working memory, learning and
attentional shifting in Parkinsonism. Neuropsychologia,
46, 3144–3156.
Mukherjee, J., Christian, B. T., Dunigan, K. A., Shi, B.,
Narayanan, T. K., Satter, M., et al. (2002). Brain imaging
of 18F-fallypride in normal volunteers: Blood analysis,
distribution, test-retest studies, and preliminary assessment
of sensitivity to aging effects on dopamine D-2/D-3 receptors.
Synapse, 46, 170–188.
Mukherjee, J., Yang, Z. Y., Das, M. K., & Brown, T. (1995).
Fluorinated benzamide neuroleptics—III. Development of
(S)-N-[(1-allyl-2-pyrrolidinyl)methyl]-5-(3-[18F]fluoropropyl)-2,
3-dimethoxybenzamide as an improved dopamine D-2
receptor tracer. Nuclear Medicine and Biology, 22, 283–296.
Nichols, T. E., & Holmes, A. P. (2002). Nonparametric
permutation tests for functional neuroimaging: A primer
with examples. Human Brain Mapping, 15, 1–25.
Nieuwenhuis, S., Gilzenrat, M. S., Holmes, B. D., & Cohen,
J. D. (2005). The role of the locus coeruleus in mediating
the attentional blink: A neurocomputational theory. Journal
of Experimental Psychology: General, 134, 291–307.
Oakes, T. R., Fox, A. S., Johnstone, T., Chung, M. K., Kalin, N.,
& Davidson, R. J. (2007). Integrating VBM into the general
linear model with voxelwise anatomical covariates.
Neuroimage, 34, 500–508.
Olivers, C. N. L., & Meeter, M. (2008). A boost and bounce
theory of temporal attention. Psychological Review, 115,
836–863.
Olivers, C. N. L., & Nieuwenhuis, S. (2006). The beneficial
effects of additional task load, positive affect, and instruction
on the attentional blink. Journal of Experimental Psychology:
Human Perception and Performance, 32, 364–379.
Olivers, C. N. L., van der Stigchel, S., & Hulleman, J. (2007).
Spreading the sparing: Against a limited-capacity account
of the attentional blink. Psychological Research, 71,
126–139.
OʼReilly, R. C., & Frank, M. J. (2006). Making working
memory work: A computational model of learning
in the prefrontal cortex and basal ganglia. Neural
Computation, 18, 283–328.
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992).
Temporary suppression of visual processing in an RSVP
task: An attentional blink? Journal of Experimental
Psychology: Human Perception and Performance,
18, 849–860.
Shapiro, K. L., Raymond, J. E., & Arnell, K. M. (1997).
The attentional blink. Trends in Cognitive Sciences,
1, 291–296.
Slagter, H. A., Johnstone, T., Beets, I. A. M., & Davidson, R. J.
(2010). Neural competition for conscious representation
across time: An fMRI study. PloS One, 5, e10556.
Slagter, H. A., Lutz, A., Greischar, L. L., Francis, A. D.,
Nieuwenhuis, S., Davis, J. M., et al. (2007). Mental training
affects distribution of limited brain resources. PLoS Biology,
5, e138.
Slagter, H. A., Lutz, A., Greischar, L. L., Nieuwenhuis, S.,
& Davidson, R. J. (2009). Theta phase synchrony
and conscious target perception: Impact of intensive
mental training. Journal of Cognitive Neuroscience, 21,
1536–1549.
Taatgen, N. A., Juvina, I., Schipper, M., Borst, J. P., & Martens, S.
(2009). Too much control can hurt: A threaded cognition
model of the attentional blink. Cognitive Psychology, 59,
1–29.
Wang, M., Vijayraghavan, S., & Goldman-Rakic, P. S. (2004).
Selective D2 receptor actions on the functional circuitry
of working memory. Science, 303, 853–856.
Woodward, N. D., Zald, D. H., Ding, Z., Riccardi, P.,
Ansari, M. S., Baldwin, R. M., et al. (2009). Cerebral
morphology and dopamine D2/D3 receptor distribution
in humans: A combined [18F]fallypride and voxel-based
morphometry study. Neuroimage, 46, 31–38.
Wyble, B., Bowman, H., & Nieuwenstein, M. (2009). The
attentional blink provides episodic distinctiveness: Sparing
at a cost. Journal of Experimental Psychology: Human
Perception and Performance, 35, 787–807.
Zhang, Y., Brady, M., & Smith, S. (2001). Segmentation of brain
MR images through a hidden Markov random field model and
the expectation-maximization algorithm. IEEE Transactions
on Medical Imaging, 20, 45–57.
1940
Journal of Cognitive Neuroscience
Volume 24, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
9
2
4
1
/
9
9
3
/
2
1
1
9
9
3
4
2
4
/
3
1
2
7
2
7
o
8
c
3
n
4
_
4
a
/
_
j
0
o
0
c
2
n
5
5
_
a
p
_
d
0
0
b
2
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1