Perspectivas de la computación
in Plants
Abstract Plants thrive in virtually all natural and human-adapted
environments and are becoming popular models for developing
robotics systems because of their strategies of morphological and
behavioral adaptation. Such adaptation and high plasticity offer new
approaches for designing, modelado, and controlling artificial
systems acting in unstructured scenarios. Al mismo tiempo, el
development of artifacts based on their working principles reveals
how plants promote innovative approaches for preservation
and management plans and opens new applications for
engineering-driven plant science. Environmentally mediated growth
patrones (p.ej., tropisms) are clear examples of adaptive behaviors
displayed through morphological phenotyping. Plants also create
networks with other plants through subterranean roots–fungi
symbiosis and use these networks to exchange resources or warning
signals. This article discusses the functional behaviors of plants and
shows the close similarities with a perceptron-like model that could
act as a behavior-based control model in plants. We begin by
analyzing communication rules and growth behaviors of plants; nosotros
then show how we translated plant behaviors into algorithmic
solutions for bioinspired robot controllers; and finally, discutimos
how those solutions can be extended to embrace original approaches
to networking and robotics control architectures.
Emanuela Del Dottore
Bioinspired Soft Robotics Laboratory
Istituto Italiano di Tecnologia
emanuela.deldottore@iit.it
Barbara Mazzolai
Bioinspired Soft Robotics Laboratory
Istituto Italiano di Tecnologia
barbara.mazzolai@iit.it
Palabras clave
Comportamiento, control, bioinspiration,
perceptron, network, adaptación
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
1 Introducción
Plants have inspired researchers in many fields, including robotics, for the development of ar-
tificial growing robots (Del Dottore, Sadeghi, et al., 2018) or continuum robots (Caminante, 2015)
and in informatics to develop novel heuristics for search and optimization problems (Akyol &
Alatas, 2017; l. Ma et al., 2015). Sin embargo, because of their apparent immobility and lack of brain,
there are frequently animated debates concerning possible forms of intelligence in plants (Cvrˇcková
et al., 2009; Firn, 2004; Robinson et al., 2020; Trewavas, 2003). Most definitions of intelligence
(Legg & Hutter, 2007) refer to the ability of a system to adapt to variable conditions in the surround-
ing environment. Disagreements about plant intelligence are probably due to the different means of
adaptation in nature across the animal (a widely accepted model in computation and robotics) y
plant kingdoms. In some animals, but apparently not in plants, adaptation is manifested in learning
(forming associations of ideas necessary for survival) (Estes, 2014). Plants adapt to their environ-
ments by continuously modifying their physiologies and morphologies, known as plant plasticity
or phenotypic plasticity. This is very different from the strengthening of synaptic connections,
eso es, synaptic plasticity, that takes place during learning processes in cognitive animals (Estes,
2014). In biology, adaptation refers to any process by which a system modifies itself to improve
© 2023 Instituto de Tecnología de Massachusetts.
Publicado bajo una atribución Creative Commons
4.0 Internacional (CC POR 4.0) licencia.
Artificial Life 29: 336–350 (2023) https://doi.org/10.1162/artl_a_00396
mi. Del Dottore and B. Mazzolai
Computation in Plants
performance in its environment (Holanda, 1992). This involves the transmission of information
from the environment to the organism through a selection of phenotyping or behavioral alter-
natives (Estes, 2014). On the basis of this definition, plants are organisms that act intelligently.
Through evolution, they continuously improve their adaptive strategies to survive in mutable en-
vironments (Gratani, 2014). The environment is central to plant life, desarrollo, and evolution.
Different ecological niches have generated many species variations with specialized physiological
and morphological adaptation mechanisms. Preprogrammed genetics is shaped in real time by envi-
ronmental stimuli. Plants and other organisms release traces into the environment that impact latent
plant plasticity. Intelligence is forged by the interactions between plants and the environment. Allá
is thus a clear analogy between intelligent behavior in plants and the approach to artificial intelli-
gence and robotics proposed by Rodney Brooks (1999). Brooks suggested that intelligent behavior
exists only in the observer’s eye as a phenomenon resulting from the interactions among multi-
ple perception-action modules that make up the control architecture of an embodied and situated
sistema (Arkin, 1998). He suggested the existence of behaviors, not produced by a high-level cog-
nitive process (implying representation or reasoning), but emerging from the interactions between
body and environment. The way the system behaves modifies the environment where it is situated
y, como consecuencia, its future perception-actions. His view has led to a modern approach to artificial
intelligence called new AI (Arroyos, 1999), which emphasizes the embodiment of behaviors and the
unavoidable interdependence of a physical system with its environment (Floreano & Mattiussi, 2008;
Scheier & Pfeifer, 1999). The concepts Brooks proposed then evolved over multiple paths. From a
control theory perspective, several schemas have been proposed in behavior-based control architec-
turas. They differ in how behaviors are combined or suppressed to generate purely reflexive systems
or hybrid architectures, in which reactive behaviors might be complemented with a deliberative part
(Arkin, 1998). From a physical perspective, the shaping of system behavior mediated by environ-
ment interactions has converged into embodied intelligence and morphological control concepts
(Pfeifer & Gómez, 2009; Pfeifer & Scheier, 2001). According to these theories, a physical system
embodies part of its computation by exploiting mechanical properties and material nonlinearities
to lighten the complexity of robot control. These theories express body-integrated behavior-based
controls, with behaviors combined by design. Purposeful design and choice of components and
materials are keys to programming such systems. Nature has designed embodied intelligent systems,
reaching maximal performance with plants. Plants have thus been taken as a source of inspiration
to mimic nastic movements into functional materials (Sydney Gladman et al., 2016) o, Por ejemplo,
to embody inherent adaptation to heterogenous environments in a growing robot based on addi-
tive manufacturing (Sadeghi et al., 2020). Reflexive adaptation in plants, en algunos casos, precedes a
subsequent active information processing with different levels of complexity that can be captured
in computational models. Following this idea, computation in plants is the perception processed
through multiple interacting behavioral modules or perception-action nodes, with an intelligence
emerging from these interactions. This article presents examples of different adaptive behavioral
strategies of plants (sección 2). We first provide an abstraction of the concept of plant computational
architecture by discussing selected plant behaviors, eso es, tropisms, and show how we translated
them into computational models (sección 3). We then discuss how to extend the proposed abstrac-
tion to capture computation at different levels of the plant hierarchy, up to a community of plants,
briefly discussing the case study of plant-to-plant communication mediated by mycorrhizal networks
(sección 4). We conclude with some final remarks on achievements, future directions and possible
applications of plant-inspired control strategies (sección 5). This perspective serves as a guideline
for the engineering interpretation of plant intelligent behaviors for bioinspired robotic control and
network models.
2 Behaviors in Plants
Plant behavior is the phenotypic plasticity expressed in response to external events during a plant’s
lifetime. Plants are multicellular organisms living in communities and have hierarchical structures
Artificial Life Volume 29, Número 3
337
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Cifra 1. (a) Hierarchical structure of a plant, starting from its internal arrangement and moving to its organization in
communities, where underground mycorrhizal networks interconnect with multiple organisms. (b) Apical growth in
plants. Roots and shoots have a meristematic region where cells divide. The cells produced start to elongate, pushing
the apex forward (light blue arrows). The elongation of cells is pronounced in the elongation region and stops in the
maturation region, where cell specialization for secondary growth occurs. (C) Examples of different behaviors observed
in plants. In the organs: circumnutations, stabilization, seedling, search for symbiosis. In the organism: apical dominance.
In the community: social defense mechanisms.
(Ferlian et al., 2018; Jha et al., 2021) with regular patterns, modularity, and spatial organization
(Dupuy et al., 2008; Sachs, 1991). Multiple organs (p.ej., roots and leaves) constitute a single plant.
Organs are organized into tissues, formed by cells, where molecules undergo physiological processes
instantiating internal rhythms, propagating from within the cells and crossing the higher layers of
the hierarchy (Figura 1a). Mutual interactions of the multiple layers define regulatory mechanisms
of plant functioning and behavior.
Plants have evolved several adaptive behaviors to counteract their sessile lifestyle at all levels
of their hierarchy. Because of their sessility, their strategies are very different from those used by
animals, the main one being apical growth (Figura 1b). Although plants do not locomote, they move
by growing, with cell division and elongation occurring at their apices of roots and aerial branches
for their entire lives. Based on growth, plants implement various movements to explore, navi-
puerta, and colonize their environment. Two key examples of growth-driven movements are tropisms,
which are irreversible directed growth responses to external stimuli (Hart, 1990; Muthert et al.,
2020), and circumnutations, which are reversible oscillatory growth-driven movements adopted, para
ejemplo, in the shoots of climbers to explore their surroundings and find mechanical support
(Stolarz, 2009) (Figure 1c). Growth-driven movements affect organs and portions of organs (p.ej.,
apical parts where cell division and elongation occur, named meristematic regions) (Perilli et al.,
2012). Esencialmente, directed growth is a form of foraging—hydrotropism (foraging for water),
chemotropism (foraging for nutrients), phototropism (foraging for light) (Ballaré et al., 1997;
338
Artificial Life Volume 29, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Cahill & McNickle, 2011)—and oscillatory growth is a form of exploratory behavior (p.ej.,
circumnutations in shoots of twiners) (Stolarz, 2009). Aerial branches differ from roots. por ejemplo-
amplio, phototropism is positive in the shoot and negative in roots, and within the same apparatus,
there are priorities dictated genetically with a preprogrammed architecture that are adjusted dynam-
ically with the expressed phenotype (Cahill et al., 2010; Hodge, 2004) (Figure 1c). Dominance hier-
archy behavior appears among organs of the same individual, leading to a single apical dominance
to reach faster growth and higher resource allocation (Aloni et al., 2006) (Figure 1c). En este caso, el
observed behavior is at the level of the plant, which invests energy in the most promising searching
organ. The symbiotic relationship between plant roots and fungal mycorrhizae (Ferlian et al., 2018)
is another key adaptation strategy. The plant exploits this symbiosis to procure phosphorous, entre
other mineral nutrients, and water, and the fungus uses the plant to gain carbon.
This symbiosis manifests intelligence in the form of social behaviors (Figure 1c), such as collab-
orative sharing of resources (Whiteside et al., 2019), warning of biotic attacks (Cabral et al., 2019),
competitive behaviors for the acquisition of resources (Selosse et al., 2006), and conquering an area
(Stinson et al., 2006). Plant roots release and perceive exudates in their rhizosphere (Bais et al.,
2006), similarly to volatile compounds released from the aerial part (Holopainen & Gershenzon,
2010). These molecules act as traces to communicate indirectly with other plant species, bacteria,
fungal hyphae, animals, or insects, to warn about biotic attacks or compensate for abiotic stresses
within a community of related plants. These examples of intelligent behaviors suggest cognitive
processes in plants and have sparked considerable interest in the scientific community. Para examen-
por ejemplo, plant neurobiology (Brenner et al., 2006; Segundo-Ortin & Calvo, 2022) is a new discipline to
understand how plants process information. Forms of cognitive processes, aprendiendo, and memory in
plants have been identified, highlighting the information-processing network implemented over dis-
tributed computing cells (Calvo Garzon, 2007; Mediano et al., 2021; Parise et al., 2020) and displayed
in anticipatory behaviors (Goodspeed et al., 2013; Latzel & Münzbergová, 2018). Well-accepted an-
ticipatory behaviors of plants are those connected to the circadian clock, such as the opening and
closing of some flowers and their leaves, anticipating sunshine to regulate their exposure to light
(Schwartz & Koller, 1986). Another strategy is being able to anticipate herbivore attacks. Plants can
differentiate between the sounds of chewing, wind, and insects and prime their defense responses
to vibrations induced by herbivore chewing in their proximity (Appel & Cocroft, 2014). Plants can
store and process data from the environment, integrating information over time (Meroz, 2021) a
optimize acclimation and immune defense responses (Hilker & Schmülling, 2019). Por último, todo
behaviors have a physiological basis at molecular and cellular levels (Aloni et al., 2006; Ferlian et al.,
2018; Salvi et al., 2020; Scheres & Van Der Putten, 2017). Behaviors are mediated by the perception
of the environment, with perception-action nodes distributed in plants at different levels of tissues
and cells. Perception-action nodes share similarities with sensory neurons (Scott, 1992). These are
cells connecting perception to motor cells (or motor neurons) through neural pathways composed
of other motor or internodes (Arber, 2012). It is justified to introduce sensory and motor neurons
while talking about plants if we recall that all biological cells can generate action potentials used for
signal transmission. These action potentials are generated by the migration of ions (p.ej., calcium)
across cell membranes when the concentration goes above an activation threshold (Aidley, 1998;
Volkov, 2012). Although plants do not have a nervous system, their cells trigger chemical reactions
similar to animals’ spiking neurons (k. Takahashi et al., 2021). It is not yet clear whether those
spikes also encode information used by plants, por ejemplo, at distal parts. Sin embargo, algunos de
the biosynthetic pathways underlying plant behaviors have been discovered. Tissue-dependent cal-
cium signatures (Krogman et al., 2020) may, por ejemplo, downstream gravity-dependent growth
regulación (Huang et al., 2013) or govern nutrient homeostasis (Vigani & Costa, 2019).
3 Tropisms
Plants respond to a wide range of external stimuli, and tropisms are perhaps the most evident be-
havior in plants at the level of the organs. Tropisms are directed growth responses toward attractive
Artificial Life Volume 29, Número 3
339
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
estímulos (p.ej., physical and chemical) from the environment or away from repulsive signals. For each
stimulus, there is a corresponding tropism, Por ejemplo, gravitropism (response to gravity), hy-
drotropism (agua), chemotropism (nutrients and salts), thigmotropism (mechanostimulation), y
magnetotropism (response to magnetic fields). The same type of signal can even produce different
reactions in different organs of the same plant. Por ejemplo, roots grow toward gravity (positivo
gravitropism), while shoots display negative gravitropism by growing upward. Because this results
in an organ following an attractor, tropisms are essentially foraging behaviors in plants.
Tropic responses are essentially individual behaviors acting in parallel on the plant and connect-
ing sense to actuation, eso es, the directed growth of cells. In a previous study (Sadeghi et al., 2016),
our research group exploited this behavioral concept with a motor scheme as a behavior-based
architecture (Arkin, 1998) in plant-inspired robotic roots to enable biomimetic autonomous navi-
gation for environment exploration. Motor schemes are suitable for treating continuous responses
(Arkin, 1989, 1998). A robot can have an infinite space of potential reactions to its world, a diferencia de
with subsumption architectures, where discrete behaviors (possible actions, discrete directions to
llevar) must be defined and treated as discrete states of a finite state machine. Potential field methods
can be used to implement a continuous response. Typically, potential field methods are adopted to
define the world as a vector field space of repulsive signals to avoid obstacles (Arkin, 1998; Laue &
Röfer, 2004). In behavior-based systems, the vector field is evaluated only at the time of perception
and for the close surroundings of the robot. This solution limits the computational power required
to evaluate the vector field and allows for a fast sense–actuation coupled response. Plants do not
necessarily need to anticipate and avoid obstacles. Sin embargo, a similar concept to repulsive vectors
can be postulated to define a field of attractive signals, whose preferential attractive direction can
be formulated as
y =
metro(cid:2)
yo=1
wixi − b,
(1)
where xi is the directed growth response for each of the external aggregated signals associated with
a specific behavior, i = gravitropism, hydrotropism, thigmotropism, chemotropism, etcétera; Wisconsin
is the specific weight for the aggregated signal; and b is the specific activation threshold. A sum-
mation coordinates the incoming behaviors. Given this description, y represents the global directed
growth response entering into motor nodes to command cell elongation. This equation also refers
to a single-layer perceptron model (Arkin, 1998; Reed & Marks, 1999), cual, en este caso, is used for
direct association of perception with motor control. With this control, the robot activates indepen-
dent behaviors in parallel as soon as the corresponding incoming stimulus goes above the activation
límite. Individual responses are summed together to obtain the attractive vector direction. El
respuesta, thus the direction the robot takes, depends only on the current perception, with no world
map reconstruction. In Sadeghi et al. (2016), we connected incoming environmental signals to mo-
tor control to achieve a directed differential elongation of robotic roots, seen as independent agents
of the plant. The robotic roots were equipped with humidity, temperatura, and tactile sensors, dis-
tributed around a conical tip and with an accelerometer. The roots were able to bend thanks to three
soft spring-based actuators (Figure 2a). An embedded board with a microcontroller was used to read
the sensors and connect perception to motor control (Figure 2b). In this example, all the agents are
identical, with wi fixed by the user and b set to zero. Different weights were assigned to the behav-
iors to analyze conditions to achieve different robot reactions to various settings and stimuli from
the surrounding world (Figure 2c). The robotic roots were developed to demonstrate plant roots’
exploratory and monitoring capabilities and to implement plantlike behaviors for application in soil
or air exploration. Such systems can be applied to reach a rapid localization of attractor sources,
Por ejemplo, for underground water source detection or localization of harmful signals according
to embedded receptors. We extended the same control to include tropisms shown in shoots of
climbing plants and used it to analyze the emergence of shadow avoidance, tolerance, or attraction
340
Artificial Life Volume 29, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
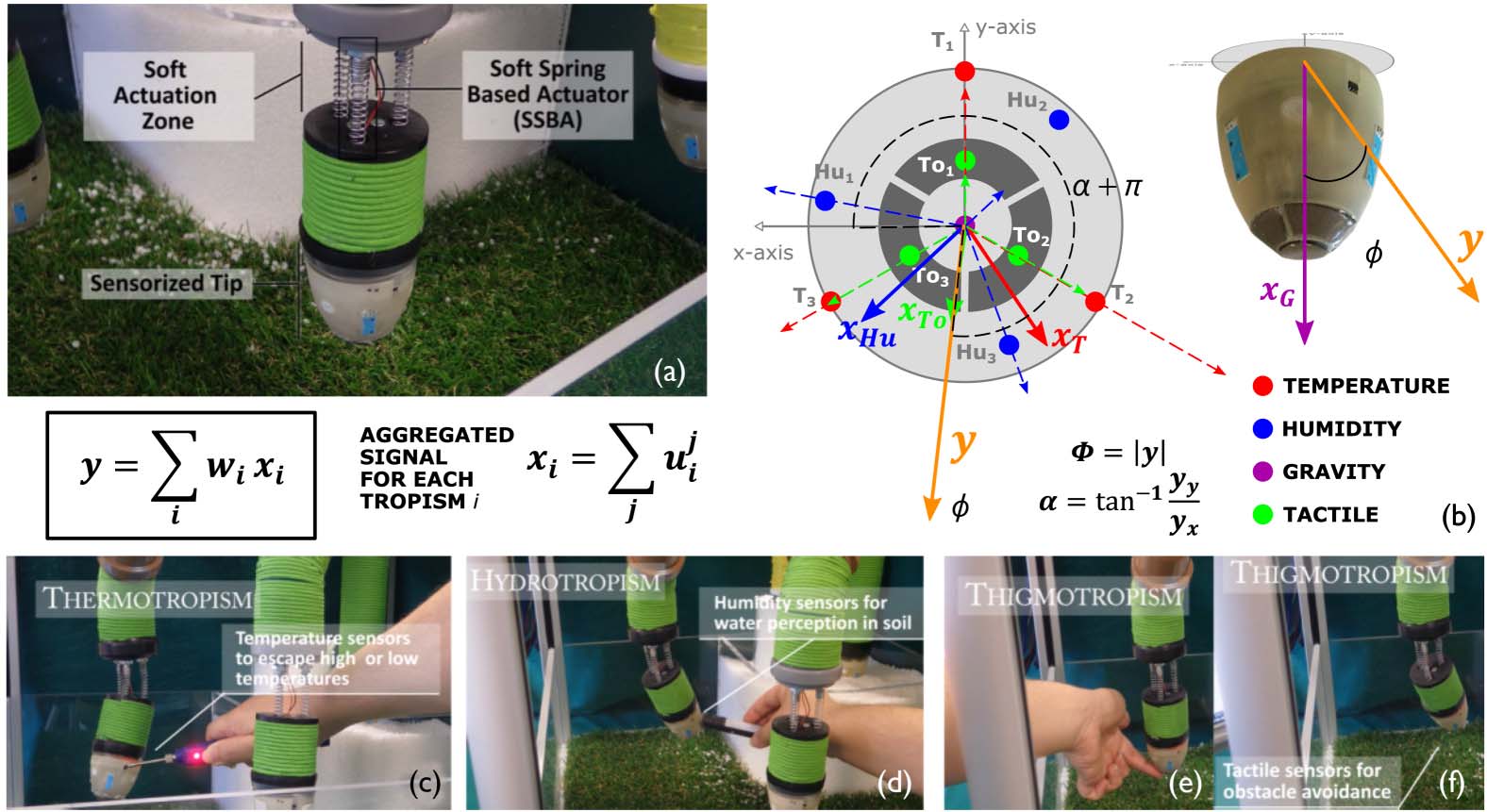
Cifra 2. (a) Example of a plant-inspired robot to imitate plant organ behaviors (Sadeghi et al., 2016). The robotic
agent in the figure has three soft springs arranged in a shaft to induce linear actuation. By differentially commanding the
motors of the springs, the robotic agent bends. (b) Robot sensor scheme and vector field generation. The tip embeds
humidity, temperatura, tactile sensors, and an accelerometer. A specific tropism (xi) results from a summation of the
vector field generated from each stimulus (u j
i , with j going over the sensors associated with tropism i). The preferential
direction to take is obtained by aggregating all the tropisms. (C) Example of the robotic agent response when stimulated
with a high-temperature source (a soldering iron). (d) Example of the robotic agent response when stimulated with a
source of humidity (wetted soil). (e–f) A sequence of the robotic agent’s response to tactile stimulation. After contact
with the finger, the robot moves away. If no other signals are present, gravitropism prevails, guiding the robotic agent
to orient itself toward the vertical.
behaviors (Del Dottore et al., 2021). Organs in plants have different specializations and thus can
offer alternative and complementary sets of behavioral responses to aid exploration strategies in
different scenarios. Además, different incoming signals undergo different internal processes in
plants (Ponce et al., 2017; Shkolnik & Fromm, 2016). This suggests that for a better understanding
and definition of plant control architecture, we might need to lower the level of abstraction and
provide signal-specific elaboration processes for each possible behavior. Gravity is perceived by
plant roots through statolitic cells located in the root cap (Cifra 3). These cells have components,
Cifra 3. An example of information processing in plants. Root apical regions perceive gravity by amyloplast sedimenta-
tion in statolitic cells. The information transduces at the molecular level, affecting protein (PIN3, PIN7) distribution and
alignment. Auxin is transported differentially on tissues through PIN distribution. Tissue-specific responses to auxin
level elongate the motor cells with an overall curvature toward gravity in plant roots.
Artificial Life Volume 29, Número 3
341
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
called amyloplasts, that move following the gravity vector. When their inclination with respect to
gravity goes over a minimum threshold (Abas et al., 2006; Swarup et al., 2005), it triggers the re-
orientation of auxin transporters at the flanks of the roots so that auxin is differentially distributed
(Furutani et al., 2020). Different auxin concentrations activate tissue-specific responses (Di Mambro
et al., 2017) to reach a macroscopic behavior called gravitropism (Ge & Chen, 2019). Differential dis-
tribution of auxin has also been observed in thigmotropic curvature responses (touch responses).
De hecho, mechanostimulation induces a polar orientation of cell auxin transporters, accelerating the
active bending response of a root encountering an obstacle (Lee et al., 2020). Mechanosensing may
also be implicated in amyloplast sedimentation perception, triggering calcium-mediated gravity re-
sponse pathways (Perbal & Driss-Ecole, 2003; k. Takahashi et al., 2021). A diferencia de, no auxin
gradient distribution has been observed in hydrotropism (Shkolnik et al., 2016). Este, sin embargo,
does not rule out the involvement of auxin in water signal transduction and transmission. De hecho,
genes in Arabidopsis plants involved in auxin signal transduction are affected during hydrotropic
respuestas (Miao et al., 2018). Amyloplast degradation is also observed in roots under water stress
(norte. Takahashi et al., 2003; Ponce et al., 2008), suggesting the need to suppress gravity signals by
hydrotropism. Amyloplast degradation is also induced in high soil salinity conditions (Sun et al.,
2008), reducing gravity perception and prioritizing escaping from dangerous conditions. Chemo-
tropism (Ferrieri et al., 2017), por otro lado, remains largely under investigated. Each nutri-
ent, metal, or salt stimulates different responses (Galvan-Ampudia & Testerink, 2011; Hodge, 2004;
j. F. Mamá, 2005; Sun et al., 2008), suggesting that chemotropism is made up of multiple subbehav-
iors. Of these subbehaviors, foraging strategies are particularly interesting (Cahill & McNickle,
2011). The Michaelis–Menten kinetics is known to drive nutrient uptake, which adapts to the
plant nutrient content (Epstein, 1972; Marschner, 2011). From this principle, in a previous article
we proposed a feedback control to adjust nutrient-specific weights and, como consecuencia, the con-
tribution of chemotropic behaviors in the overall collaborative behavior observed in a swarm of
rootlike robotic agents addressing the survival of the plant individual (Del Dottore, Mondini, et al.,
2018). Each agent acted independently from the other agents, with no overall world knowledge or
consciousness of other agents’ actions but relying only on local perception, shared resources on the
basis of each agent’s needs, and local nutrient memory. The swarm reached a balance of nutrients at
the level of the plant maximizing its wellness, without needing a central coordinator to set a global
goal or function or to act as a task allocator. Crosstalk exists between tropisms (Massa & Gilroy,
2003; h. Takahashi & Scott, 1991; norte. Takahashi et al., 2002), implying the co-contribution of dif-
ferent signals (xi with i = 1…m sensory nodes) in the achievement of a unified growth response
(φl) and justifying the use of a behavior coordination module (p.ej., cooperative summation), cual
might be guided by the same messenger (p.ej., the growth hormone auxin; Retzer et al., 2014) or by
alternative biosynthetic pathways converging into analogous responses (Muthert et al., 2020). El
single perceptron model (Ecuación 1) needs to mutate into a multilayer perceptron (Reed & Marks,
1999; Scheres & Van Der Putten, 2017) (Cifra 4), in which the first layer of hidden nodes represents
all the possible behaviors obtained with some activation function (φj, p.ej., sine law in gravitropism;
Mullen et al., 2000) for the aggregated value of weighted (wij) input signals (xi) that go above the
activation threshold (bj). To consider the possible mutual suppression of behaviors, as in the case
of hydrotropism over gravitropism, a fully connected network with all the external signals entering
the behaviors in the motor scheme architecture can be built. The wij can then be set to activate or
deactivate specific connections and define the signal summation or inhibition (negative weight). En
al mismo tiempo, the mutualistic effect of behaviors in plants that express phenotyping during their
lifetimes suggests using an adaptive module for short- and long-term memory, by which weights
of individual behaviors are adjusted to reflect adaptation over time (Cifra 4). The interrelation
among tropisms likely significantly influences circumnutation movements in plant roots (Mullen
et al., 1998; Stolarz, 2009). In agreement with the multilayer perceptron-like model proposed in
Cifra 4, we previously adopted continuous-time recurrent neural networks (a class of multilayer
perceptron) to describe tissue-dependent responses to auxin fluxes and investigate the role of cir-
cumnutations and their emergence in roots (Loshchilov et al., 2021). This study highlighted the
342
Artificial Life Volume 29, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Cifra 4. A multilayer perceptron as a model for representing plant control. The coordination among multiple stimuli-
associated behaviors produces the final growth response toward an attractor in space.
mutual role of gravitropism and thigmotropism in the occurrence of oscillatory movement patterns
and the possible role of these oscillations in exploring resources (chemotropism). Sin embargo, cómo
precisely behaviors are set, which functions model adaptation best, and what the signaling path-
ways are for each tropism and, como consecuencia, the signal processing to apply are just some of the
open issues requiring a better understanding of molecular changes during plant cell development
(Muthert et al., 2020). The multilayer perceptron architecture is a general model that can be de-
tailed for each behavior node and on the adaptive module, on the basis of new knowledge about
plant functions or according to the needs of a specific application.
4 Extended Perceptron
Plants create networks with other plants and organisms. They establish underground networks made
up of roots and fungi in the soil, called mycorrhizal networks (Simard et al., 2012). This intense inter-
connection facilitates the survival of many species by improving the colonization of large areas,
foraging, and activating defense mechanisms (Beiler et al., 2010; Selosse et al., 2006). They likely
function as an infrastructure for exchanging nutrients to recover from abiotic stress (Van’t Padje,
Bonfante, et al., 2021; Van’t Padje, Werner, et al., 2021) and propagate warning signals to prevent
biotic attacks at far distances (Alaux et al., 2020; Babikova et al., 2013). Mycorrhizae can medi-
ate competing behaviors among enemy plants and promote cooperative behaviors among relatives
(Gorzelak et al., 2015; Kiers & Heijden, 2006). This underground social network is known as the
Wood Wide Web (Beiler et al., 2010). Some attempts to improve its understanding have been made
through mechanistic, statistical network models and market or complex adaptive system theories
(Simard et al., 2012; Van’t Padje, Werner, et al., 2021). The Wood Wide Web is, En realidad, a fascinating
example of a fault-tolerant, complex adaptive system in nature (Messier et al., 2013; Simard, 2009;
Van Dorp et al., 2020). From a phytocentric point of view, plants are nodes and fungi are links in
this network. The number of nodes (the plants) in the network is dynamic. Each node is a dynamic
multilayer perceptron-like model expressing the phenotype of an individual plant. Each node has
a dynamic number of sensory inputs, varying with symbiotic connections that work as a highway
to transfer traces sent by distant nodes. Through the network, the perception of a single plant is
augmented, with the information processing, enhanced at the community level, having dynamic
connectivity patterns. The environment remains central in the dynamics of this complex system.
Computation is achieved by opening and closing symbiotic connections, source-sink transfer mech-
anisms, and possibly other active transport and regulatory mechanisms that define the information
flow and its distribution path over the network (Ezawa & saito, 2018; Jin et al., 2012). The exact
rules of message exchange, transport mechanisms, signal molecules, and regulatory pathways of
comunicación, especially concerning biotic alarm signals (Johnson & Gilbert, 2015), are still not
clear. Sin embargo, interest in understanding the plant–fungus relationship is rapidly growing due
to its crucial importance in regulating the carbon cycle and limiting the effects of climate change. Un
Artificial Life Volume 29, Número 3
343
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
engineering-driven approach can provide alternative perspectives and tools to benefit investigations.
Our computational model can be extended to describe mycorrhizal dynamics, building up process-
ing on its nodes and connections upon new knowledge about how the information is computed
and combined over the network. The extended perceptron would reflect an evolutionary state of
the forest over time, displaying intelligent behavior in the form of phenotypic adaptation to pertur-
bations in the environment. This tool would help predict alterations in an ecosystem and could be
used to define forest management strategies.
5 Conclusions
We have provided a perspective on plant behaviors and how they can become the basis of com-
putational models. These models can be exploited to develop plant-inspired control strategies and
algorithms while favoring biological investigations for a better understanding of plant behaviors.
Research on plant-inspired engineering is still underexplored and suffers from a noncomprehensive
view of plant functioning. Owing to complex interconnections among biosynthetic pathways, allá
is still no complete understanding of plant activation functions in response to environmental sig-
nal. Sin embargo, in this article we have reported some mechanisms of environmentally mediated
responses in plants, allowing us to define an abstraction for a generalizable plant like multilayer per-
ceptron that also complies with the motor scheme of behavior-based architectures. We adopted this
approach to imitate plants’ behavioral strategies for monitoring (Sadeghi et al., 2016) or foraging
(Del Dottore, Mondini, et al., 2018; Del Dottore et al., 2021) with adaptive adjustment of network
weights (Del Dottore, Mondini, et al., 2018). This control strategy could be exploited in explorative
autonomous robots, Por ejemplo, to locate resources that dissolve in gradients into the environment,
such as water, salts, or some metals in soil. Además, we have adopted continuous-time recurrent
neural networks to investigate the role of circumnutations and the reasons for their emergence in
plant roots (Loshchilov et al., 2021). Plants have already been investigated by analogy with multi-
layer perceptrons (Scheres & Van Der Putten, 2017), focusing on how such information-processing
structure can be replicated at the molecular level. Here we corroborate this idea and suggest that this
computational structure can help in the study of adaptive behavior dynamics across multiple imple-
mentation scales. Multilayer perceptron models convey computations carried out at different levels
in the hierarchical structure of plants and could be extended to dynamic computational architec-
tures to capture communication rules behind cooperative and competitive behaviors implemented
in communities of plants with mycorrhizal networks. Despite vast and growing research communi-
ties on plant biology, ecology, and mycorrhiza, hasta la fecha, such a specific biological model has rarely
been explored. Further investigations into plant and fungi physiology and molecular biology could
unravel essential functional rules that would consolidate and refine our proposed plant control ar-
chitecture by answering some key questions: How are the weights of single behaviors adjusted?
Which behaviors involve crosstalk? and What are the specific signal transduction and transmission
funciones? These questions are nested into a hierarchical organization of plants displaying behaviors
at the level of organs, organisms, and communities (Ferlian et al., 2018), appealing to the engineering
world. We believe that this approach could facilitate new solutions for autonomous environment
exploration, showing better adaptation and compliance in mutable conditions. Our approach could
also promote engineering-driven knowledge by raising more awareness for more sustainable man-
agement of resources for agricultural applications, forest management, and ecosystem restoration.
With this contribution, we hope to stimulate a cross-disciplinary discussion, which is fundamental
to boosting the development of novel technologies and stimulating overall scientific progress.
Expresiones de gratitud
This work has received funding from the European Research Council (ERC) under the European
El horizonte de la Unión 2020 research and innovation program (grant 101003304) (I-WOOD).
344
Artificial Life Volume 29, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Referencias
Abas, l., Benjamín, r., Malenica, NORTE., Paciorek, T., Wišniewska, J., Moulinier-Anzola, j. C., Sieberer, T.,
Friml, J., & Luschnig, C. (2006). Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux
facilitator PIN2 are involved in root gravitropism. Nature Cell Biology, 8(3), 249–256. https://doi.org/10
.1038/ncb1369, PubMed: 16489343
Aidley, D. j. (1998). The physiology of excitable cells. Prensa de la Universidad de Cambridge. https://doi.org/10.1017
/CBO9781139171182
Akyol, S., & Alatas, B. (2017). Plant intelligence based metaheuristic optimization algorithms. Artificial
Intelligence Review, 47(4), 417–462. https://doi.org/10.1007/s10462-016-9486-6
Alaux, P.-L., Naveau, F., Declerck, S., & Cranenbrouck, S. (2020). Common mycorrhizal network induced
JA/ET genes expression in healthy potato plants connected to potato plants infected by Phytophthora
infestans. Frontiers in Plant Science, 11, 602. https://doi.org/10.3389/fpls.2020.00602, PubMed: 32523589
Aloni, r., Aloni, MI., Langhans, METRO., & Ullrich, C. I. (2006). Role of cytokinin and auxin in shaping root
architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root
gravitropism. Annals of Botany, 97(5), 883–893. https://doi.org/10.1093/aob/mcl027, PubMed: 16473866
Appel, h. METRO., & Cocroft, R. B. (2014). Plants respond to leaf vibrations caused by insect herbivore chewing.
Oecologia, 175(4), 1257–1266. https://doi.org/10.1007/s00442-014-2995-6, PubMed: 24985883
Arber, S. (2012). Motor circuits in action: Especificación, conectividad, and function. Neurona, 74(6), 975–989.
https://doi.org/10.1016/j.neuron.2012.05.011, PubMed: 22726829
Arkin, R. C. (1989). Motor schema–based mobile robot navigation. International Journal of Robotics Research, 8(4),
92–112. https://doi.org/10.1177/027836498900800406
Arkin, R. C. (1998). Behavior-based robotics. CON prensa.
Babikova, Z., Gilbert, l., bruce, t. J., Birkett, METRO., Caulfield, j. C., Woodcock, C., Pickett, j. A., & Johnson, D.
(2013). Underground signals carried through common mycelial networks warn neighbouring plants of
aphid attack. Ecology Letters, 16(7), 835–843. https://doi.org/10.1111/ele.12115, PubMed: 23656527
Bais, h. PAG., Weir, t. l., Perry, l. GRAMO., Gilroy, S., & Vivanco, j. METRO. (2006). The role of root exudates in
rhizosphere interactions with plants and other organisms. Revisión anual de biología vegetal, 57(1), 233–266.
https://doi.org/10.1146/annurev.arplant.57.032905.105159, PubMed: 16669762
Ballaré, C. l., Scopel, A. l., & Sanchez, R. A. (1997). Foraging for light: Photosensory ecology and
agricultural implications. Plant, Cell, and Environment, 20(6), 820–825. https://doi.org/10.1046/j.1365-3040
.1997.d01-112.x
Beiler, k. J., Durall, D. METRO., Simard, S. w., Maxwell, S. A., & Kretzer, A. METRO. (2010). Architecture of the
wood-wide web: Rhizopogon spp. genets link multiple Douglas-fir cohorts. New Phytologist, 185(2), 543–553.
https://doi.org/10.1111/j.1469-8137.2009.03069.x, PubMed: 19878460
Brennero, mi. D., Stahlberg, r., Mancuso, S., Vivanco, J., Baluška, F., & Van Volkenburgh, mi. (2006). Plant
neurobiology: An integrated view of plant signaling. Trends in Plant Science, 11(8), 413–419. https://doi.org
/10.1016/j.tplants.2006.06.009, PubMed: 16843034
Arroyos, R. A. (1999). Cambrian intelligence: The early history of the new AI. CON prensa. https://doi.org/10.7551
/mitpress/1716.001.0001
Cabral, C., Wollenweber, B., António, C., & Ravnskov, S. (2019). Activity in the arbuscular mycorrhizal
hyphosphere warning neighbouring plants. Frontiers in Plant Science, 10, 511. https://doi.org/10.3389/fpls
.2019.00511, PubMed: 31057597
Cahill, j. F., Jr., & McNickle, GRAMO. GRAMO. (2011). The behavioral ecology of nutrient foraging by plants. Annual
Review of Ecology, Evolución, and Systematics, 42, 289–311. https://doi.org/10.1146/annurev-ecolsys-102710
-145006
Cahill, j. F., Jr., McNickle, GRAMO. GRAMO., Haag, j. J., Lamb, mi. GRAMO., Nyanumba, S. METRO., & Calle. Clair, C. C. (2010). Plants
integrate information about nutrients and neighbors. Ciencia, 328(5986), 1657. https://doi.org/10.1126
/science.1189736, PubMed: 20576883
Calvo Garzon, F. (2007). The quest for cognition in plant neurobiology. Plant Signaling and Behavior, 2(4),
208–211. https://doi.org/10.4161/psb.2.4.4470, PubMed: 19516990
Cvrˇcková, F., Lipavská, h., & Žárský, V. (2009). Plant intelligence: Por qué, why not or where? Plant Signaling and
Comportamiento, 4(5), 394–399. https://doi.org/10.4161/psb.4.5.8276, PubMed: 19816094
Artificial Life Volume 29, Número 3
345
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Del Dottore, MI., Mondini, A., & Mazzolai, B. (2021). Support localization strategy for growing robots aided
by light perception inspired by climbing plants. En 2021 IEEE 4th International Conference on Soft Robotics
(RoboSoft) (páginas. 105–110). IEEE. https://doi.org/10.1109/RoboSoft51838.2021.9479220
Del Dottore, MI., Mondini, A., Sadeghi, A., & Mazzolai, B. (2018). Swarming behavior emerging from the
uptake–kinetics feedback control in a plant-root-inspired robot. Applied Sciences, 8(1), 47. https://doi.org
/10.3390/app8010047
Del Dottore, MI., Sadeghi, A., Mondini, A., Mattoli, v., & Mazzolai, B. (2018). Toward growing robots:
A historical evolution from cellular to plant-inspired robotics. Frontiers in Robotics and AI, 5, 16.
https://doi.org/10.3389/frobt.2018.00016, PubMed: 33500903
Di Mambro, r., De Ruvo, METRO., Pacifici, MI., Salvi, MI., Sozzani, r., Benfey, PAG. NORTE., Busch, w., Novak, o., Ljung,
K., Di Paola, l., Marée, A. F. METRO., Costantino, PAG., Grieneisen, V. A., & Sabatini, S. (2017). Auxin minimum
triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Actas
of the National Academy of Sciences of the United States of America, 114(36), E7641–E7649.
https://doi.org/10.1073/pnas.1705833114, PubMed: 28831001
Dupuy, l., Mackenzie, J., Rudge, T., & Haseloff, j. (2008). A system for modelling cell–cell interactions
during plant morphogenesis. Annals of Botany, 101(8), 1255–1265. https://doi.org/10.1093/aob/mcm235,
PubMed: 17921524
Epstein, mi. (1972). Mineral nutrition of plants: Principles and perspectives. John Wiley.
Estes, W.. (2014). Handbook of learning and cognitive processes: volumen. 2. Conditioning and behavior theory. Psicología
Prensa. https://doi.org/10.4324/9781315770437
Ezawa, T., & saito, k. (2018). How do arbuscular mycorrhizal fungi handle phosphate? New insight into
fine-tuning of phosphate metabolism. New Phytologist, 220(4), 1116–1121. https://doi.org/10.1111/nph
.15187, PubMed: 29701874
Ferlian, o., Biere, A., Bonfante, PAG., Buscot, François., Eisenhauer, NORTE., Fernandez, I., Hause, B., Herrmann, S.,
Krajinski-Barth, F., Meier, I. C., Pozo, METRO. J., Rasmann, S., Rillig, METRO. C., Tarkka, METRO. T., van Dam, norte. METRO.,
Wagg, C., & Martinez-Medina, A. (2018). Growing research networks on mycorrhizae for mutual benefits.
Trends in Plant Science, 23(11), 975–984. https://doi.org/10.1016/j.tplants.2018.08.008, PubMed: 30241736
Ferrieri, A. PAG., Machado, R. A., Arce, C. C., Kessler, D., Baldwin, I. T., & Erb, METRO. (2017). Localized
micronutrient patches induce lateral root foraging and chemotropism in Nicotiana attenuata. Diario de
Integrative Plant Biology, 59(10), 759–771. https://doi.org/10.1111/jipb.12566, PubMed: 28650091
Firn, R. (2004). Plant intelligence: An alternative point of view. Annals of Botany, 93(4), 345–351. https://doi
.org/10.1093/aob/mch058, PubMed: 15023701
Floreano, D., & Mattiussi, C. (2008). Bio-inspired artificial intelligence: Theories, methods, and technologies. CON prensa.
Furutani, METRO., Hirano, y., Nishimura, T., Nakamura, METRO., Taniguchi, METRO., suzuki, K., Oshida, r., Kondo, C.,
Sol, S., Kato, K., Fukao, y., Hakoshima, T., & Morita, METRO. t. (2020). Polar recruitment of RLD by
LAZY1-like protein during gravity signaling in root branch angle control. Comunicaciones de la naturaleza, 11(1),
1–13. https://doi.org/10.1038/s41467-019-13729-7, PubMed: 31900388
Galvan-Ampudia, C. S., & Testerink, C. (2011). Salt stress signals shape the plant root. Current Opinion in Plant
Biología, 14(3), 296–302. https://doi.org/10.1016/j.pbi.2011.03.019, PubMed: 21511515
Ge, l., & Chen, R. (2019). Negative gravitropic response of roots directs auxin flow to control root
gravitropism. Plant, Cell, and Environment, 42(8), 2372–2383. https://doi.org/10.1111/pce.13559,
PubMed: 30968964
Goodspeed, D., Chehab, mi. w., Covington, METRO. F., & Braam, j. (2013). Circadian control of jasmonates and
salicylates: The clock role in plant defense. Plant Signaling and Behavior, 8(2), e23123. https://doi.org/10
.4161/psb.23123, PubMed: 23299428
Gorzelak, METRO. A., Asay, A. K., Pickles, B. J., & Simard, S. W.. (2015). Inter-plant communication through
mycorrhizal networks mediates complex adaptive behaviour in plant communities. AoB Plants, 7.
https://doi.org/10.1093/aobpla/plv050, PubMed: 25979966
Gratani, l. (2014). Plant phenotypic plasticity in response to environmental factors. Advances in Botany, 2014,
208747. https://doi.org/10.1155/2014/208747
Hart, j. W.. (1990). Plant tropisms: And other growth movements. Springer Science and Business Media.
Hilker, METRO., & Schmülling, t. (2019). Stress priming, memory, and signalling in plants. Plant, Cell, y
Environment, 42(3), 753–761. https://doi.org/10.1111/pce.13526, PubMed: 30779228
346
Artificial Life Volume 29, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Hodge, A. (2004). The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytologist,
162(1), 9–24. https://doi.org/10.1111/j.1469-8137.2004.01015.x
Holanda, j. h. (1992). Adaptation in natural and artificial systems: An introductory analysis with applications to biology,
control, and artificial intelligence. CON prensa. https://doi.org/10.7551/mitpress/1090.001.0001
Holopainen, j. K., & Gershenzon, j. (2010). Multiple stress factors and the emission of plant VOCs. Tendencias en
Plant Science, 15(3), 176–184. https://doi.org/10.1016/j.tplants.2010.01.006, PubMed: 20144557
Huang, S.-J., Chang, C.-L., Wang, P.-H., Tsai, M.-C., hsu, P.-H., & Chang, I.-F. (2013). A type III ACC
synthase, ACS7, is involved in root gravitropism in Arabidopsis thaliana. Journal of Experimental Botany,
64(14), 4343–4360. https://doi.org/10.1093/jxb/ert241, PubMed: 23943848
Jha, S. GRAMO., Borowsky, A. T., Col, B. J., Fahlgren, NORTE., Farmer, A., Huang, S. S. C., Karia, PAG., Libault, METRO.,
Provart, norte. J., Rice, S. l., Saura-Sanchez, METRO., agarwal, PAG., Ahkami, A. h., Anderton, C. r., Briggs, S. PAG.,
Brophy, j. A., Denolf, PAG., Di Costanzo, l. F., Exposito-Alonso, METRO., . . . Rhee, S. Y. (2021). Science forum:
Vision, challenges and opportunities for a plant cell atlas. Elife, 10, e66877. https://doi.org/10.7554/eLife
.66877, PubMed: 34491200
Jin, h., Liu, J., Liu, J., & Huang, X. (2012). Forms of nitrogen uptake, translocation, and transfer via
arbuscular mycorrhizal fungi: A review. Science China Life Sciences, 55(6), 474–482. https://doi.org/10.1007
/s11427-012-4330-y, PubMed: 22744177
Johnson, D., & Gilbert, l. (2015). Interplant signalling through hyphal networks. New Phytologist, 205(4),
1448–1453. https://doi.org/10.1111/nph.13115, PubMed: 25421970
Kiers, mi. T., & Heijden, METRO. GRAMO. V. D. (2006). Mutualistic stability in the arbuscular mycorrhizal symbiosis:
Exploring hypotheses of evolutionary cooperation. Ecology, 87(7), 1627–1636. https://doi.org/10.1890
/0012-9658(2006)87[1627:MSITAM]2.0.CO;2, PubMed: 16922314
Krogman, w., Sparks, j. A., & Blancaflor, mi. B. (2020). Cell type-specific imaging of calcium signaling in
Arabidopsis thaliana seedling roots using GCAMP3. International Journal of Molecular Sciences, 21(17), 6385.
https://doi.org/10.3390/ijms21176385, PubMed: 32887481
Latzel, v., & Münzbergová, z. (2018). Anticipatory behavior of the clonal plant Fragaria vesca. Frontiers in Plant
Ciencia, 9, 1847. https://doi.org/10.3389/fpls.2018.01847, PubMed: 30619415
Laue, T., & Röfer, t. (2004). A behavior architecture for autonomous mobile robots based on potential fields.
In RoboCup 2004: Robot Soccer World Cup VIII (páginas. 122–133). Saltador. https://doi.org/10.1007/978-3-540
-32256-6_10
Sotavento, h. J., kim, h. S., Parque, j. METRO., Dar, h. S., & Jeon, j. h. (2020). Pin-mediated polar auxin transport
facilitates root-obstacle avoidance. New Phytologist, 225(3), 1285–1296. https://doi.org/10.1111/nph
.16076, PubMed: 31336402
Legg, S., & Hutter, METRO. (2007). A collection of definitions of intelligence. Frontiers in Artificial Intelligence and
Aplicaciones, 157, 17.
Loshchilov, I., Del Dottore, MI., Mazzolai, B., & Floreano, D. (2021). Conditions for the emergence of
circumnutations in plant roots. PLoS ONE, 16(5), e0252202. https://doi.org/10.1371/journal.pone
.0252202, PubMed: 34038485
Mamá, j. F. (2005). Plant root responses to three abundant soil minerals: Silicon, aluminum and iron. Critical
Reviews in Plant Sciences, 24(4), 267–281. https://doi.org/10.1080/07352680500196017
Mamá, l., Zhu, y., Liu, y., tian, l., & Chen, h. (2015). A novel bionic algorithm inspired by plant root foraging
behaviors. Computación blanda aplicada, 37, 95–113. https://doi.org/10.1016/j.asoc.2015.08.014
Marschner, h. (2011). Marschner’s mineral nutrition of higher plants. Prensa académica.
Massa, GRAMO. D., & Gilroy, S. (2003). Touch modulates gravity sensing to regulate the growth of primary roots of
Arabidopsis thaliana. Plant Journal, 33(3), 435–445. https://doi.org/10.1046/j.1365-313X.2003.01637.x,
PubMed: 12581302
Mediano, PAG. A., Trewavas, A., & Calvo, PAG. (2021). Information and integration in plants: Towards a
quantitative search for plant sentience. Journal of Consciousness Studies, 28(1–2), 80–105.
Meroz, Y. (2021). Plant tropisms as a window on plant computational processes. New Phytologist, 229(4),
1911–1916. https://doi.org/10.1111/nph.17091, PubMed: 33219510
Messier, C., Puettmann, k. J., & Coates, k. D. (2013). Managing forests as complex adaptive systems: Building resilience
to the challenge of global change. Routledge. https://doi.org/10.4324/9780203122808
Artificial Life Volume 29, Número 3
347
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Miao, r., Wang, METRO., Yuan, w., Ren, y., li, y., zhang, NORTE., zhang, J., Kronzucker, h. J., & Xu, W.. (2018).
Comparative analysis of Arabidopsis ecotypes reveals a role for brassinosteroids in root hydrotropism. Plant
Physiology, 176(4), 2720–2736. https://doi.org/10.1104/pp.17.01563, PubMed: 29439211
Mullen, j. l., Turk, MI., Johnson, K., Wolverton, C., Ishikawa, h., Simmons, C., Söll, D., & evans, METRO. l.
(1998). Root-growth behavior of the Arabidopsis mutant RGR1: Roles of gravitropism and circumnutation
in the waving/coiling phenomenon. Plant Physiology, 118(4), 1139–1145. https://doi.org/10.1104/pp.118.4
.1139, PubMed: 9847088
Mullen, j. l., Wolverton, C., Ishikawa, h., & evans, METRO. l. (2000). Kinetics of constant gravitropic stimulus
responses in Arabidopsis roots using a feedback system. Plant Physiology, 123(2), 665–670. https://doi.org
/10.1104/pp.123.2.665, PubMed: 10859196
Muthert, l. W.. F., Izzo, l. GRAMO., Van Zanten, METRO., & Aronne, GRAMO. (2020). Root tropisms: Investigations on earth
and in space to unravel plant growth direction. Frontiers in Plant Science, 10, 1807. https://doi.org/10.3389
/fpls.2019.01807, PubMed: 32153599
Parise, A. GRAMO., Gagliano, METRO., & Souza, GRAMO. METRO. (2020). Extended cognition in plants: Is it possible? Plant Signaling
and Behavior, 15(2), 1710661. https://doi.org/10.1080/15592324.2019.1710661, PubMed: 31900033
Perbal, GRAMO., & Driss-Ecole, D. (2003). Mechanotransduction in gravisensing cells. Trends in Plant Science, 8(10),
498–504. https://doi.org/10.1016/j.tplants.2003.09.005, PubMed: 14557047
Perilli, S., Di Mambro, r., & Sabatini, S. (2012). Growth and development of the root apical meristem.
Current Opinion in Plant Biology, 15(1), 17–23. https://doi.org/10.1016/j.pbi.2011.10.006, PubMed:
22079783
Pfeifer, r., & Gómez, GRAMO. (2009). Morphological computation—Connecting brain, body, and environment. En
Creating brain-like intelligence (páginas. 66–83). Saltador. https://doi.org/10.1007/978-3-642-00616-6_5
Pfeifer, r., & Scheier, C. (2001). Understanding intelligence. CON prensa. https://doi.org/10.7551/mitpress/6979
.001.0001
Ponce, GRAMO., Corkidi, GRAMO., Eapen, D., Lledías, F., Cárdenas, l., & Cassab, GRAMO. (2017). Root hydrotropism and
thigmotropism in Arabidopsis thaliana are differentially controlled by redox status. Plant Signaling and
Comportamiento, 12(4), e1305536. https://doi.org/10.1080/15592324.2017.1305536, PubMed: 28318377
Ponce, GRAMO., Rasgado, F. A., & Cassab, GRAMO. I. (2008). Roles of amyloplasts and water deficit in root tropisms.
Plant, Cell, and Environment, 31(2), 205–217. https://doi.org/10.1111/j.1365-3040.2007.01752.x,
PubMed: 18047572
Reed, r., & Marks, R. j. (1999). Neural smithing: Supervised learning in feedforward artificial neural networks.
CON prensa. https://doi.org/10.7551/mitpress/4937.001.0001
Retzer, K., Korbei, B., & Luschnig, C. (2014). Auxin and tropisms. In Auxin and its role in plant development
(páginas. 361–387). Saltador. https://doi.org/10.1007/978-3-7091-1526-8_16
robinson, D. GRAMO., Draguhn, A., & Taiz, l. (2020). Plant “intelligence” changes nothing. Informes EMBO, 21(5),
e50395. https://doi.org/10.15252/embr.202050395, PubMed: 32301219
Sachs, t. (1991). Pattern formation in plant tissues. Prensa de la Universidad de Cambridge. https://doi.org/10.1017
/CBO9780511574535
Sadeghi, A., Del Dottore, MI., Mondini, A., & Mazzolai, B. (2020). Passive morphological adaptation for
obstacle avoidance in a self-growing robot produced by additive manufacturing. Soft Robotics, 7(1), 85–94.
https://doi.org/10.1089/soro.2019.0025, PubMed: 31592712
Sadeghi, A., Mondini, A., Del Dottore, MI., Mattoli, v., Beccai, l., Taccola, S., Lucarotti, C., Totaro, METRO., &
Mazzolai, B. (2016). A plant-inspired robot with soft differential bending capabilities. Bioinspiration and
Biomimetics, 12(1), 015001. https://doi.org/10.1088/1748-3190/12/1/015001, PubMed: 27997363
Salvi, MI., Di Mambro, r., & Sabatini, S. (2020). Dissecting mechanisms in root growth from the transition
zone perspective. Journal of Experimental Botany, 71(8), 2390–2396. https://doi.org/10.1093/jxb/eraa079,
PubMed: 32064533
Scheier, C., & Pfeifer, R. (1999). The embodied cognitive science approach. In Dynamics, synergetics, autonomous
agents: Nonlinear systems approaches to cognitive psychology and cognitive science (páginas. 159–179). Científico mundial.
https://doi.org/10.1142/9789812815354_0011
Scheres, B., & Van Der Putten, W.. h. (2017). The plant perceptron connects environment to development.
Naturaleza, 543(7645), 337–345. https://doi.org/10.1038/nature22010, PubMed: 28300110
348
Artificial Life Volume 29, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Schwartz, A., & Koller, D. (1986). Diurnal phototropism in solar tracking leaves of Lavatera cretica. Plant
Physiology, 80(3), 778–781. https://doi.org/10.1104/pp.80.3.778, PubMed: 16664701
Scott, S. A. (1992). Sensory neurons: Diversity, desarrollo, and plasticity. prensa de la Universidad de Oxford.
Segundo-Ortin, METRO., & Calvo, PAG. (2022). Consciousness and cognition in plants. Wiley Interdisciplinary Reviews:
Ciencia cognitiva, 13(2), e1578. https://doi.org/10.1002/wcs.1578, PubMed: 34558231
Selosse, M.-A., Ricardo, F., Él, X., & Simard, S. W.. (2006). Mycorrhizal networks: Des liaisons dangereuses?
Trends in Ecology and Evolution, 21(11), 621–628. https://doi.org/10.1016/j.tree.2006.07.003, PubMed:
16843567
Shkolnik, D., & Fromm, h. (2016). The Cholodny-Went theory does not explain hydrotropism. Plant Science,
252, 400–403. https://doi.org/10.1016/j.plantsci.2016.09.004, PubMed: 27717476
Shkolnik, D., Krieger, GRAMO., Nuriel, r., & Fromm, h. (2016). Hydrotropism: Root bending does not require
auxin redistribution. Molecular Plant, 9(5), 757–759. https://doi.org/10.1016/j.molp.2016.02.001,
PubMed: 26911727
Simard, S. W.. (2009). Mycorrhizal networks and complex systems: Contributions of soil ecology science to
managing climate change effects in forested ecosystems. Canadian Journal of Soil Science, 89(4), 369–382.
https://doi.org/10.4141/cjss08078
Simard, S. w., Beiler, k. J., Bingham, METRO. A., Deslippe, j. r., Philip, l. J., & Teste, F. PAG. (2012). Mycorrhizal
redes: Mechanisms, ecology and modelling. Fungal Biology Reviews, 26(1), 39–60. https://doi.org/10
.1016/j.fbr.2012.01.001
Stinson, k. A., Campbell, S. A., Powell, j. r., lobo, B. MI., Callaway, R. METRO., Thelen, GRAMO. C., Hallett, S. GRAMO.,
Prati, D., & Klironomos, j. norte. (2006). Invasive plant suppresses the growth of native tree seedlings by
disrupting belowground mutualisms. Biología PLoS, 4(5), e140. https://doi.org/10.1371/journal.pbio
.0040140, PubMed: 16623597
Stolarz, METRO. (2009). Circumnutation as a visible plant action and reaction: Physiological, cellular and molecular
basis for circumnutations. Plant Signaling and Behavior, 4(5), 380–387. https://doi.org/10.4161/psb.4.5
.8293, PubMed: 19816110
Sol, F., zhang, w., Hu, h., li, B., Wang, y., zhao, y., li, K., Liu, METRO., & li, X. (2008). Salt modulates gravity
signaling pathway to regulate growth direction of primary roots in Arabidopsis. Plant Physiology, 146(1),
178–188. https://doi.org/10.1104/pp.107.109413, PubMed: 18024552
Swarup, r., Kramer, mi. METRO., Perry, PAG., Knox, K., Leyser, h. METRO., Haseloff, J., Beemster, GRAMO. T., Bhalerao, r., &
bennett, METRO. j. (2005). Root gravitropism requires lateral root cap and epidermal cells for transport and
response to a mobile auxin signal. Nature Cell Biology, 7(11), 1057–1065. https://doi.org/10.1038/ncb1316,
PubMed: 16244669
Sydney Gladman, A., Matsumoto, mi. A., Nuzzo, R. GRAMO., Mahadevan, l., & Luis, j. A. (2016). Biomimetic 4D
printing. Nature Materials, 15(4), 413–418. https://doi.org/10.1038/nmat4544, PubMed: 26808461
Takahashi, h., & Scott, t. k. (1991). Hydrotropism and its interaction with gravitropism in maize roots.
Plant Physiology, 96(2), 558–564. https://doi.org/10.1104/pp.96.2.558, PubMed: 11538004
Takahashi, K., Takahashi, h., Furuichi, T., Toyota, METRO., Furutani-Seiki, METRO., Kobayashi, T., Watanabe-Takano,
h., Shinohara, METRO., Numaga-Tomita, T., Sakaue-Sawano, A., Miyawaki, A., & Naruse, k. (2021).
Gravity sensing in plant and animal cells. NPJ Microgravity, 7(1), 1–10. https://doi.org/10.1038/s41526
-020-00130-8, PubMed: 33558517
Takahashi, NORTE., Goto, NORTE., Okada, K., & Takahashi, h. (2002). Hydrotropism in abscisic acid, wavy, y
gravitropic mutants of Arabidopsis Thaliana. Planta, 216(2), 203–211. https://doi.org/10.1007/s00425-002
-0840-3, PubMed: 12447533
Takahashi, NORTE., Yamazaki, y., Kobayashi, A., Higashitani, A., & Takahashi, h. (2003). Hydrotropism interacts
with gravitropism by degrading amyloplasts in seedling roots of Arabidopsis and radish. Plant Physiology,
132(2), 805–810. https://doi.org/10.1104/pp.018853, PubMed: 12805610
Trewavas, A. (2003). Aspects of plant intelligence. Annals of Botany, 92(1), 1–20. https://doi.org/10.1093/aob
/mcg101, PubMed: 12740212
Van Dorp, C. h., Simard, S. w., & Durall, D. METRO. (2020). Resilience of Rhizopogon-Douglas-fir mycorrhizal
redes 25 years after selective logging. Mycorrhiza, 30(4), 467–474. https://doi.org/10.1007/s00572-020
-00968-6, PubMed: 32556666
Artificial Life Volume 29, Número 3
349
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
mi. Del Dottore and B. Mazzolai
Computation in Plants
Van’t Padje, Anouk., Bonfante, PAG., Ciampi, l. T., & Kiers, mi. t. (2021). Quantifying nutrient trade in the
arbuscular mycorrhizal symbiosis under extreme weather events using quantum-dot tagged phosphorus.
Frontiers in Ecology and Evolution, 9, 153. https://doi.org/10.3389/fevo.2021.613119
Van’t Padje, A., Werner, GRAMO. D., & Kiers, mi. t. (2021). Mycorrhizal fungi control phosphorus value in trade
symbiosis with host roots when exposed to abrupt “crashes” and “booms” of resource availability.
New Phytologist, 229(5), 2933–2944. https://doi.org/10.1111/nph.17055, PubMed: 33124078
Vigani, GRAMO., & Costa, A. (2019). Harnessing the new emerging imaging technologies to uncover the role of
signalling in plant nutrient homeostasis. Plant, Cell, and Environment, 42(10), 2885–2901. https://doi
CA2+
.org/10.1111/pce.13611, PubMed: 31286524
Volkov, A. GRAMO. (2012). Plant electrophysiology: Methods and cell electrophysiology. Springer Science and Business Media.
https://doi.org/10.1007/978-3-642-29119-7
Caminante, I. D. (2015). Biologically inspired vine-like and tendril-like robots. En 2015 Science and information
conference (SAI) (pp 714–720). IEEE. https://doi.org/10.1109/SAI.2015.7237221
Whiteside, METRO. D., Werner, GRAMO. D., Caldas, V. MI., van’t Padje, A., Dupin, S. MI., Elbers, B., Bakker, METRO., Wyatt,
GRAMO. A., Klein, METRO., Hink, METRO. A., postma, METRO., Vaitla, B., Noë, r., Shimizu, t. S., Oeste, S. A., & Kiers, mi. t.
(2019). Mycorrhizal fungi respond to resource inequality by moving phosphorus from rich to poor patches
across networks. Biología actual, 29(12), 2043–2050. https://doi.org/10.1016/j.cub.2019.04.061, PubMed:
31178314
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
9
3
3
3
6
2
1
5
4
2
6
9
a
r
t
yo
/
_
a
_
0
0
3
9
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
350
Artificial Life Volume 29, Número 3