PERSPECTIVA
What Does Dorsal Cortex Contribute
to Perception?
Erez Freud 1, Marlene Behrmann 2, and Jacqueline C. Snow 3
1Department of Psychology and the Centre for Vision Research, York University
2Department of Psychology and the Neuroscience Institute, Carnegie Mellon University
3Department of Psychology, The University of Nevada Reno
Palabras clave:
affordance
two visual pathways, object
recognition, vision-for-perception, vision-for-action,
ABSTRACTO
According to the influential “Two Visual Pathways” hypothesis, the cortical visual system is
segregated into two pathways, with the ventral, occipitotemporal pathway subserving object
percepción, and the dorsal, occipitoparietal pathway subserving the visuomotor control of
acción. Sin embargo, growing evidence suggests that the dorsal pathway also plays a functional
role in object perception. In the current article, we present evidence that the dorsal pathway
contributes uniquely to the perception of a range of visuospatial attributes that are not
redundant with representations in ventral cortex. We describe how dorsal cortex is recruited
automatically during perception, even when no explicit visuomotor response is required.
En tono rimbombante, we propose that dorsal cortex may selectively process visual attributes that can
inform the perception of potential actions on objects and environments, and we consider
plausible developmental and cognitive mechanisms that might give rise to these
representaciones. Tal como, we consider whether naturalistic stimuli, such as real-world solid
objects, might engage dorsal cortex more so than simplified or artificial stimuli such as
images that do not afford action, and how the use of suboptimal stimuli might limit our
understanding of the functional contribution of dorsal cortex to visual perception.
TWO VISUAL PATHWAYS
The cortical visual system of human and nonhuman primates is anatomically segregated into
two pathways, each of which is hierarchical in nature. These two pathways originate in early
visual cortex and then diverge, with one projecting to the ventral cortex and extending to
the lateral and inferior surfaces of the temporal lobes, and the other projecting to the dorsal
cortex and extending to the posterior and superior aspects of the parietal lobes (Mishkin &
Ungerleider, 1982).
A través de los años, influential cognitive neuroscience theories have postulated that the two
pathways are not only structurally dissociated from each other but are also functionally disso-
ciated. Initially, it was suggested that the functional dissociation was based on the differential
sensitivity of the pathways to specific visual attributes of the input. According to this divi-
sion of labor, the “what” ventral pathway processes information about object properties (es decir.,
un acceso abierto
diario
Citación: Freud, MI., behrman, METRO., &
Snow, j. C. (2020). What Does Dorsal
Cortex Contribute to Perception?
Mente abierta: Discoveries in Cognitive
Ciencia, 4, 40–56. https://doi.org/10.
1162/opmi_a_00033
DOI:
https://doi.org/10.1162/opmi_a_00033
Recibió: 6 Enero 2020
Aceptado: 20 Puede 2020
Conflicto de intereses: Los autores
declaran no tener ningún conflicto de
interés.
Autor correspondiente:
Erez Freud
efreud@yorku.ca
Derechos de autor: © 2020
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
shape/geometry, and surface properties such as texture and color) and promotes “object vi-
sión,” while the “where” dorsal pathway encodes information about the location of the object
in space and promotes “spatial vision” (Mishkin & Ungerleider, 1982; Mishkin, Ungerleider, &
Macko, 1983). Roughly a decade later, this framework underwent revision with the recasting
of the role of the pathways not based on the type of input information to which the path-
way is responsive but, en cambio, by the contribution of each pathway to different functions or
output requirements. On this reconceptualized view, the ventral pathway is responsible for
vision-for-perception (es decir., the “what” pathway), while the dorsal pathway is responsible for
vision-for-action (es decir., the “how” pathway) (Goodale & Milner, 1992). Just as was true for the
what/where division of labor, the what/how distinction has been reinforced by decades of re-
search with studies employing diverse methods, including neuroimaging, neuropsychology,
and psychophysics (for a recent review, see Goodale & Milner, 2018).
En años recientes, sin embargo, there is growing evidence that neither of these frameworks
adequately captures the functional capabilities of each pathway. críticamente, the evidence sug-
gests that the dorsal pathway may not only be engaged in vision-for-action computations but
might also play a role in object perception (Erlikhman, Caplovitz, Gurariy, Medina, & Snow,
2018; Freud, Plaut, & behrman, 2016), specifically in diverse perceptual functions such as
3D perception, localization of objects, and spatiotemporal integration.
In the current perspective, we review the evidence for the involvement of the dorsal
pathway in the computation of four different visuospatial attributes, a saber, profundidad, orientación,
structure-from-motion, and shape, even under conditions that are independent of goal-directed
comportamiento. We also consider evidence consistent with the view that dorsal representations con-
tribute uniquely to visual perception and are not simply redundant with information computed
in ventral cortex. Además, we discuss some aspects of visual development that might give
rise to the emergence of these dorsal representations. We then argue that naturalistic visual in-
puts, such as real-world graspable solids, may engage the dorsal pathway more strongly than
simplified or artificial stimuli such as pictures or computerized images, which are often used in
vision science, and that the use of these suboptimal stimuli may have contributed to the under-
emphasis on the dorsal pathway in visual perception. Finalmente, we propose a new perspective
on dorsal cortex function that centers on the premise that the potential for action offered by
visual stimuli in naturalistic visual environments might shape the computations carried out by
the dorsal pathway. These computations are then utilized to support action but can be used
more extensively to support perception, también.
THE DORSAL PATHWAY PROCESSES VISUOSPATIAL ATTRIBUTES
In contrast with earlier characterizations of dorsal cortex that emphasize how visual inputs
are used primarily to guide online control of action, research over the past two decades has
revealed that the dorsal pathway is involved in processing visuospatial attributes of the vi-
sual input (for a discussion on the functional parcellation of the dorsal pathway, see Box 1).
En tono rimbombante, this perceptual contribution of dorsal cortex is apparent even under conditions
where no explicit visuomotor response is needed (for review, see Erlikhman et al., 2018;
Freud et al., 2016). Below we describe some of the key findings from this literature (ver figura 1
for a summary).
Three-Dimensional (3D) Depth Perception
The processing of depth information is imperative for the successful recognition of 3D
objects, cual, under naturalistic conditions, need to be recognized from different distances
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
41
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
.
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
Box 1. Anatomical and functional parcellations of the dorsal pathway.
In this review, we often refer to “dorsal pathway representations.” However, it is important
to note that the dorsal pathway territory includes significant portions of the occipital and
the parietal lobes and is, por lo tanto, composed of heterogeneous anatomical and functional
subregions. These regions differ in their relative contribution to perceptual and visuomo-
tor functions. The principal parcellation of the parietal lobe includes the postcentral gyrus,
the superior parietal lobule, the inferior parietal lobule, the parietal operculum, and the in-
traparietal sulcus (IPS). Además, each of these regions can be further divided to smaller
subregions (p.ej., the IPS can be functionally subdivided to five or six regions based on
retinotopic maps; Sereno et al., 1995; Swisher, Halko, Merabet, McMains, & Somers, 2007).
Respectivamente, previous studies have recognized the neural heterogeneity of the dorsal path-
way and characterized the changes in neural representations of objects along this pathway.
Kravitz and colleagues (2011) have provided one of the most comprehensive accounts
on the functional subdivision of the dorsal pathway. This account, which relies mostly on
animal studies, suggests that the dorsal pathway is composed of three sub-pathways: El
parieto–prefrontal pathway that supports spatial working memory, the parieto–premotor path-
way that supports visually guided action, and the parieto–medial temporal pathway that
primarily promotes spatial navigation. En tono rimbombante, the functional architecture of the three
sub-pathways corresponds to the structural connections of each sub-pathway with other cor-
tical structures.
The notion that the structural connectivity of regions along the dorsal pathway can
dictate their relative contribution to perception and action is also mirrored in the idea that
object representations in the dorsal pathway follow a posterior-anterior gradient of the dorsal
pathway (Freud, Culham, Plaut, & behrman, 2017; Freud et al., 2016). En particular, posición-
terior regions of the IPS are structurally and functionally coupled with the ventral pathway
(Freud, Rosenthal, Ganel, & Avaro, 2015; Stepniewska, Cerkevich, & Aquél, 2015). Por eso,
those regions likely contribute more to perceptual functions, while more anterior regions,
which are coupled with premotor, motor, and somatosensory cortices (Rushworth, Behrens,
& Johansen-Berg, 2006), are more important for visuomotor transformations.
Por último, different regions of parietal cortex are postulated to be involved in computations
related to other cognitive functions that are not purely visual. This includes functions such
as attention (behrman, Geng, & Shomstein, 2004; Buschman & Kastner, 2015; Greenberg
et al., 2012), multisensory integration (Ghazanfar & Schroeder, 2006), working memory
(D'Esposito & Postle, 2015), and numerical computation (harvey, Klein, Petridou, &
Dumoulin, 2013). One possibility is that the involvement of parietal cortex in these functions
might reflect the extent to which spatial attributes are relevant for the specific processes,
Por ejemplo, spatial representations that underlie the conceptualization of a number line.
The mapping between the nature of the information represented in dorsal cortex and the
emergent function remains to be fully explicated.
and viewpoints (Cox, 2014). Sensitivity of the dorsal pathway to this type of varying infor-
formación, previously thought to be the sole purview of ventral cortex, has been documented
in many studies with both human and nonhuman primates (for reviews, see Orban, 2011;
Orban, Janssen,& Vogels, 2006). These studies have revealed responses to depth information
induced from both monocular (Georgieva, Todd, Peeters, & Orban, 2008; Nelissen et al., 2009;
Vanduffel et al., 2002) and binocular (es decir., disparity) señales (Georgieva, Peeters, Kolster, Todd, &
Orban, 2009; Janssen, Vogels, & Orban, 2000; Yoshioka, Doi, Abdolrahmani, & Fujita, 2020),
and have also uncovered responses to the global 3D structure of objects (Freud, Ganel, et al.,
2017; Freud, Rosenthal, Ganel, & Avaro, 2015). Finalmente, activation in the dorsal pathway, como
revealed using fMRI in humans, is observed in mental rotation (Gauthier et al., 2002), un proceso
that is closely associated with computations related to 3D structural information. Similarmente,
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
42
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
.
/
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
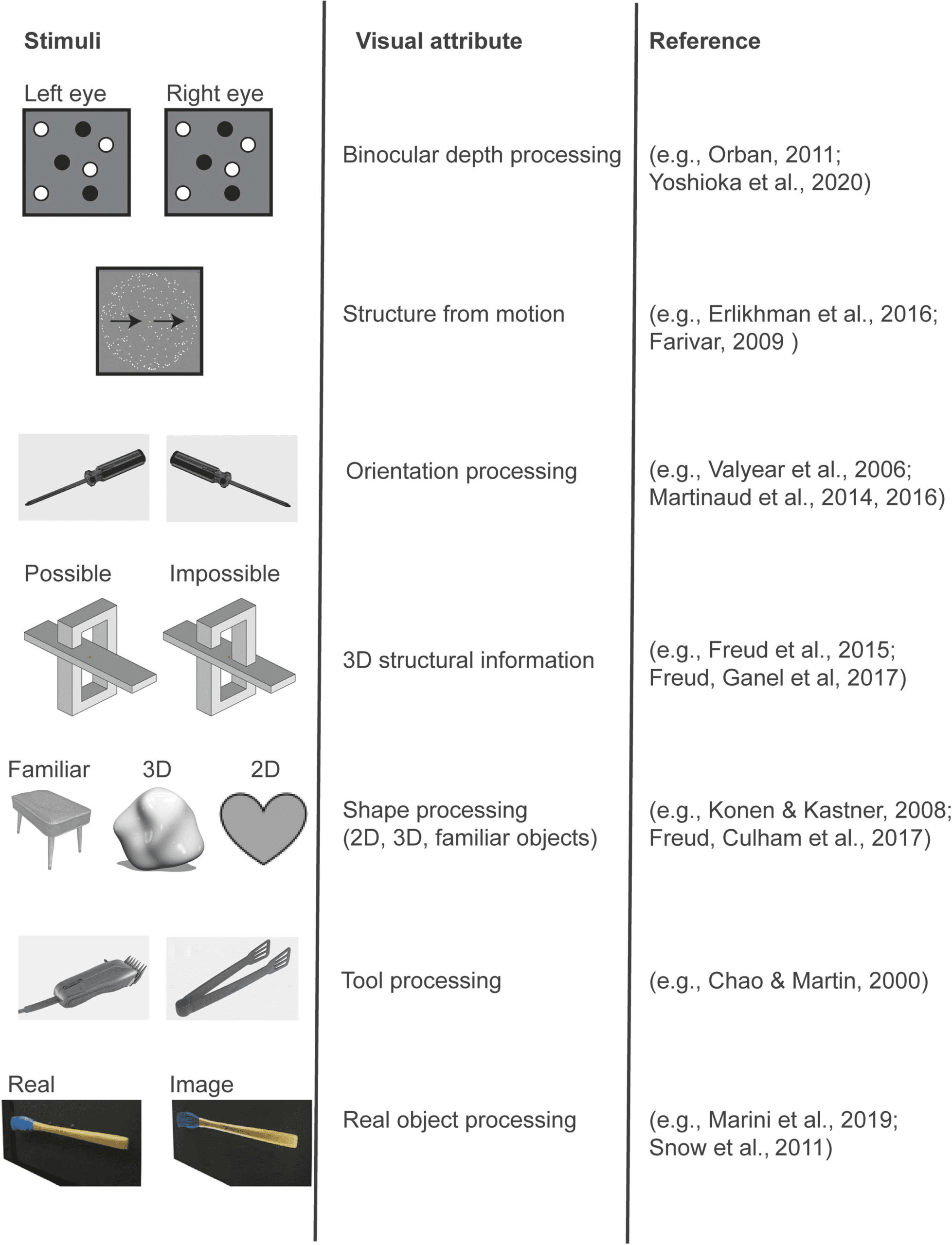
Cifra 1. The dorsal pathway is sensitive to different visual attributes. Regions along the dorsal
pathway process different visual attributes. The left column provides examples of the experimental
stimuli that have been used. The middle column summarizes the visual attribute that is assumed to
be processed by the dorsal pathway. The right column provides representative references.
the processing of configurally manipulated faces, which is related to volume derivation, elic-
its greater activation in the dorsal cortex than featured-based processing (Zachariou, Nikas,
Safiullah, Gotts, & Ungerleider, 2016).
Sin embargo, it might be argued that sensitivity to depth information in the dorsal pathway
is solely in the service of action and visuomotor transformations. Sin embargo, recent behavioral
evidence from healthy observers (Freud, robinson, & behrman, 2018), studies of brain-
damaged patients with visual agnosia (Freud, Ganel, et al., 2017), and nonhuman primates
(Van Dromme, Premereur, Verhoef, Vanduffel, & Janssen, 2016), undermine this alternative
hypothesis by showing that the dorsal pathway contributes to, and might even be necessary
para, successful visuospatial perception (Berryhill, Fendrich, & Olson, 2009; Medina, Jax, &
(2017) showed that patients with visual ag-
Coslett, 2020). En particular, Freud, Ganel, et al.
nosia following damage to ventral cortex (with extensive damage in some cases) were still able
to derive the 3D structural information of objects and evinced preserved sensitivity of dorsal re-
sponses to this information. Consistently, reversible inactivation of the caudal IPS (es decir., a region
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
43
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
in the dorsal pathway) in monkeys led to impaired 3D perception (Van Dromme et al., 2016).
Visuospatial impairments have also been found in humans with lesions to the dorsal pathway
(Berryhill et al., 2009; Medina et al., 2020), further supporting a causal relationship between
dorsal cortex function and object perception.
Orientation
Another visual attribute to which dorsal cortex shows sensitivity is the orientation of a stimulus
y, por supuesto, representing the orientation is necessary for grasping behavior. Sin embargo, ori-
entation is critical for perception too, Por ejemplo, to distinguish between the enantiomorph
letters “p” and “q,” which share geometric forms that are mirror images of each other. Damage
to parietal cortex, Por ejemplo, following a right occipitoparietal hematoma (Martinaud et al.,
2014), or after a lesion affecting the posterior temporal gyrus and the inferior parietal lobule
(Martinaud et al., 2016), results in persistent impairments in orientation and mirror-orientation
Procesando. Asimismo, neuroimaging findings from healthy individuals have demonstrated that
the dorsal transverse occipital sulcus, but not the ventral cortex, is sensitive to the orientation
of objects (Valyear, Culham, Sharif, Westwood, & Goodale, 2006) and to mirror transforma-
tions of scenes (Dilks, Julian, Kubilius, Spelke, & Kanwisher, 2011). These data reveal that the
dorsal pathway is also involved in computing orientation for the purpose of perception, pro-
viding further support for the role of dorsal cortex in perceptual processing, independent of
goal-directed actions.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
.
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Structure-From-Motion
Motion is one of the most powerful cues for the perception of objects in dynamic, 3D, natural
entornos. Motion information can be used to compute the distance of an object, to inte-
grate spatial information over time and to retrieve the 3D structure of objects, and regions of
the dorsal pathway that are activated by 3D information are also sensitive to motion cues (para
a detailed review, see Erlikhman et al., 2018).
The computations of Structure-from-Motion (SfM) are of particular interest, because they
are thought to be the responsibility of the dorsal pathway and, por lo tanto, independent from
computations carried out by the ventral pathway (Farivar, 2009). SfM is achieved by exploit-
ing the varying velocities of different points on the surface of the object. Movement-selective
regiones (middle temporal area, MONTE; and medial superior temporal area, MST) (Sugihara,
Murakami, shenoy, andersen, & Komatsu, 2002), together with regions in posterior parietal
cortex in humans (Erlikhman, Gurariy, Mruczek, & Caplovitz, 2016; Vanduffel et al., 2002),
are involved in extracting SfM. Neuropsychological investigations offer converging evidence
for the role of the dorsal pathway in the perception of SfM. Por ejemplo, patients with oc-
cipitoparietal lesions, but not those with occipitotemporal lesions, are selectively impaired in
perceiving 3D SfM and in detecting global motion patterns (Vaina, 1989) (see also preserved
biological motion perception in individuals with ventral lesions, Gilaie-Dotan, Saygin, Lorenzi,
rees, & behrman, 2015). En tono rimbombante, this impairment in SfM perception did not reflect a gen-
eral deficit in motion processing, as the patients with occipitoparietal lesions were still able to
detect local motion.
Shape
Shape processing, standardly attributed to the ventral pathway, is perhaps one of the most in-
triguing test cases for the role of dorsal cortex in perception. Shape processing provides the
foundation for a diverse range of perceptual and visuomotor behaviors and involves higher-level
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
44
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
integration of multiple visual cues. Whereas early neuroimaging research identified shape-
selectivity in the lateral occipital and posterior fusiform cortices within the ventral occipi-
totemporal (vOT) pathway (Grill-Spector et al., 1998; Malach et al., 1995), later studies also
uncovered shape selectivity in parietal cortex. These initial observations of shape selectivity in
dorsal cortex were attributed to the involvement of attention (Kourtzi & Kanwisher, 2000), o
to the association of certain types of visual stimuli such as tools (versus non-tool objects) con
visuomotor actions (chao & Martín, 2000; Mruczek, von Loga, & Kastner, 2013). Sin embargo,
more recent evidence has revealed shape selectivity in multiple regions of the dorsal pathway
for non-tool objects (Bracci & Op de Beeck, 2016; Freud, Culham, Plaut, & behrman, 2017),
for novel objects with no semantic associations (Freud et al., 2015; Konen & Kastner, 2008),
and even for 2D line-drawings of objects or basic shapes (Konen & Kastner, 2008; Sereno &
Maunsell, 1998).
Many studies have been devoted to elucidating the nature of shape representations de-
rived by the dorsal-response profile and a number of important conclusions have been reached.
Primero, dorsal shape selectivity is not uniform across parietal cortex (Kravitz et al., 2011), pero
bastante, follows a representational gradient, with greater shape sensitivity in more posterior
regions compared with more anterior regions that are tuned to visuomotor aspects (Freud,
Culham, Plaut, & behrman, 2017; ver figura 2 and Box 1). Segundo, although shape repre-
sentations in the dorsal pathway can be modulated by input from (Mahon, Kumar, & Almeida,
2013) or by damage confined to (Freud & behrman, 2020) the ventral pathway, these dorsal
representations are not just epiphenomenal or a byproduct resulting from cascaded signals
from the ventral pathway. Studies using high–temporal resolution methods, like ERP, MEG,
and single-cell recording, have revealed that dorsal shape-selectivity signals precede those of
ventral shape signals (collins, Freud, Kainerstorfer, Cao, & behrman, 2019; Lehky & Sereno,
2007; Liu, Wang, zhou, Ding, & luo, 2017), making it unlikely that dorsal activation is merely
projected from ventral shape computations. También, recent developmental neuroimaging find-
ings in normally developing children ranging in age from 6 a 21 años, reveals that dorsal
cortex matures earlier than ventral cortex, suggesting that representations in dorsal cortex are
unlikely to be constrained a priori by ventral signals (Ciesielski et al., 2019). Finalmente, based
on human fMRI data, the representational similarity between shape-selective responses in the
two visual pathways can be greater than the similarity within each of the pathways, further
emphasizing the importance of both pathways in shape processing (Freud, Culham, Plaut,
& behrman, 2017). Sin embargo, unlike ventral representations, dorsal representations are
more adaptive than ventral representations, and they are subject to task demands. Por ejemplo,
fMRI classification accuracies of object category from dorsal region decreased when partici-
pants completed a task that was unrelated to the objects, while classification accuracies from
ventral regions were more invariant of the task (Vaziri-Pashkam & Xu, 2017, 2018; Xu, 2018).
What Gives Rise to Dorsal Pathway Representations?
Some stimulus attributes processed in dorsal cortex (reviewed above), such as SfM, appear to
be unique and under the sole purview of the dorsal visual pathway, whereas others, como
orientation and shape, are represented by both dorsal and ventral pathways. One might then
ask what the functional contribution of dorsal representations is to visual perception and to
what extent is this contribution distinct from ventral cortex? Does dorsal cortex contribute to
visual perception by processing the elements considered above or are there additional contri-
butions? Even though there is overlap in the visual attributes processed by the two pathways,
dorsal and ventral cortex may still contribute in different ways (Konen & Kastner, 2008). Para
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
45
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
.
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
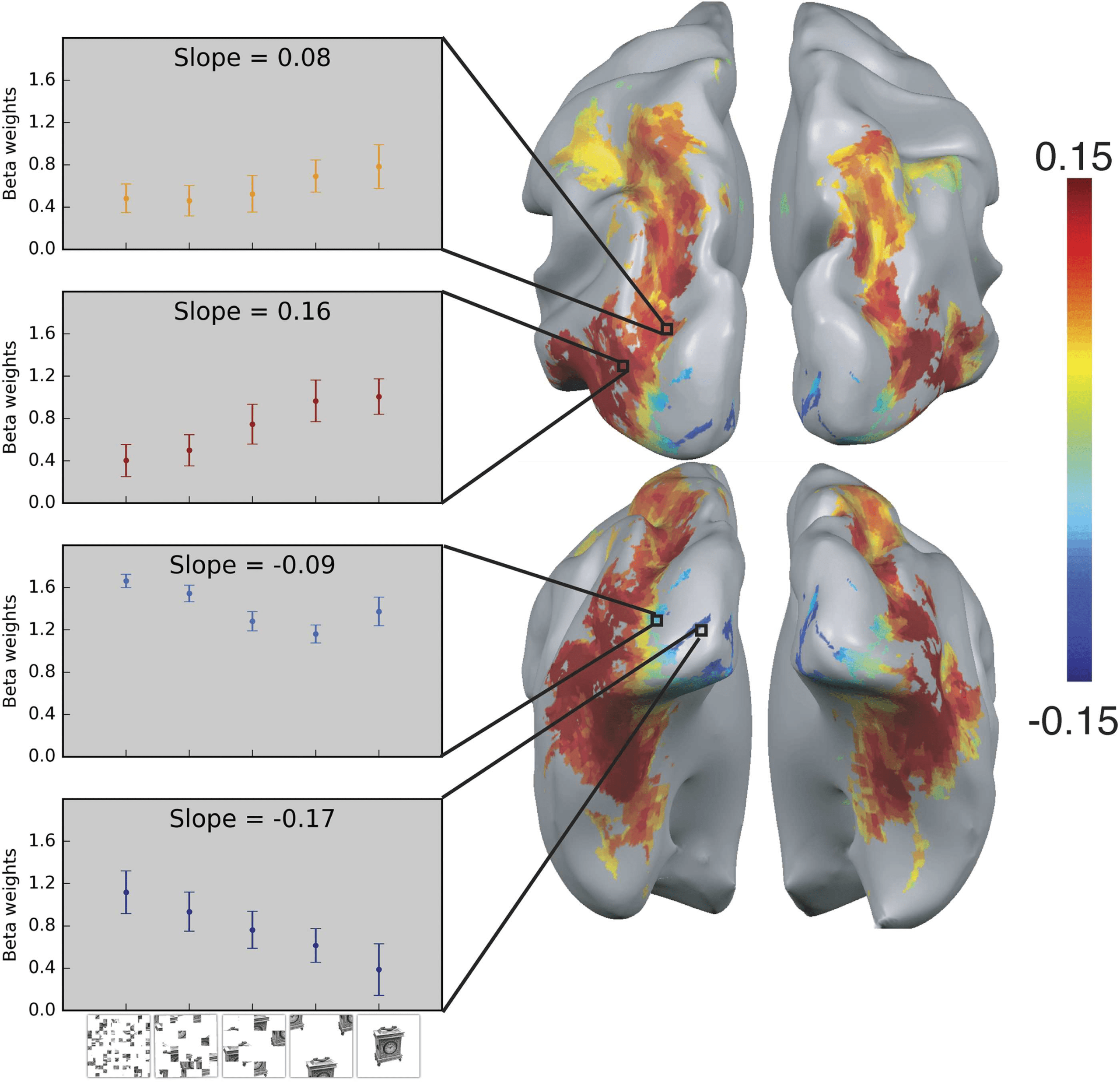
Cifra 2. Shape processing along the dorsal pathway. Shape sensitivity along both pathways was
explored using fMRI by utilizing a parametric scrambling manipulation in which shape information
is degraded gradually (left panel, fila inferior). The activation profile of four representative clusters
(10 voxels each) is plotted and the color of the bars reflects the slope value of each cluster (left panel).
Shape sensitivity is projected on an inflated brain from a superior view (upper right panel) and from
a posterior-inferior view (lower right panel). Warm colors signify voxels that are shape sensitive,
with activation increasing as a function of object coherence. En cambio, cold colors reflect low
shape sensitivity (negative slopes) or greater sensitivity for scrambled than intact images. Notablemente,
shape sensitivity was observed not only along the ventral pathway but also in the dorsal pathway
(Freud, Culham, et al., 2017).
ejemplo, dorsal cortex might use these inputs to guide perception in a qualitatively differ-
ent fashion than ventral cortex. En otras palabras, even though largely similar visual inputs are
utilized in both visual streams, they may still serve different perceptual purposes.
One possibility is that dorsal cortex processes certain visual attributes because the at-
tributes offer useful physical constraints for potential actions with objects and environments.
Específicamente, the visual attributes reviewed above, including 3D depth, SfM, and geometric
forma, may be selectively extracted for further processing by dorsal cortex because they spec-
ify regularities in the natural world that can be exploited to inform and constrain the perception
of environmental opportunities for action. Real-world environments consist of a complex 3D
array of tangible solids of different shapes, sizes, and weights, and of course, observers move
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
46
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
around the environment, thereby changing the relative egocentric distances between the body
and other solids. Notablemente, the idea that the potential for action can promote dorsal cortex
processing is also consistent with neuropsychological and behavioral studies that documented
enhanced perceptual abilities for objects that are placed near the hands and therefore are more
relevant for actions (Brockmole, Davoli, Abrams, & Witt, 2013; Cosman & Vecera, 2010; Gozli,
Oeste, & Pratt, 2012).
En tono rimbombante, this perspective of dorsal cortex function bears some resemblance to the
“vision-for-action” and “vision-for-perception” framework (Goodale & Milner, 1992) in that it
emphasizes the different output requirements of the two pathways. But the key difference lies
in the distinction that the dorsal pathway is recruited not only during “in-the-moment” actions
but also during visual perception for detecting potential opportunities for action with objects
and environments, whether or not an action is planned or executed. This view dovetails with
“ecological” frameworks of perception that emphasize that visual perception serves to facilitate
action in real-world contexts (Gibson, 1979), and evolutionary arguments that the human brain
has presumably evolved to allow us to perceive and interact with real objects and environments
(Un cuerpo & Kalaska, 2010; Heft, 2013).
Such a “potential action” mechanism for dorsal contributions to visual perception, en
which representations are modulated by attributes such as 3D shape, viewpoint, egocen-
tric distance, and physical size, also contrasts with theoretical accounts of the ventral pro-
cessing stream in which responses to objects remain predominantly stable or “invariant,"
despite changes in viewing conditions that alter object appearance but not identity (Bar, 2004;
Grill-Spector, Kourtzi, & Kanwisher, 2001; Grill-Spector & Malach, 2004; Kravitz, Vinson, &
Panadero, 2008; Logothetis & Sheinberg, 1996). Because subtle changes in visual parameters can
have a dramatic influence on behavior, it seems advantageous to have perceptual responses
or representations in dorsal cortex that are largely stable (es decir., not invariant) when a stimulus
conveys visual cues that signify concrete physical attributes (Holler, Fabbri, & Snow, 2020).
Por ejemplo, knowing the absolute egocentric distance of an object from the body is critical
for determining whether the object lies within reach (Jeannerod, 1981) and for pre-shaping
the hand for grasping (Castiello, 2005; Jeannerod, 1986; Smeets & Brennero, 1999). Sin embargo,
these conjectures contrast with evidence for “content rich,” complex representations of be-
haviorally relevant objects in dorsal cortex that appear to be represented in a highly abstract
manner (jeong & Xu, 2016; Xu, 2018). Por ejemplo, images of task-relevant faces and cars
are represented invariantly in human parietal cortex despite changes in visual parameters such
as background scene, viewpoint or size (jeong & Xu, 2016). Sin embargo, as we outline later (ver
Dorsal Cortex May Play a Unique Role in Perceiving Action-Relevant Stimuli and Environ-
mentos), it remains to be seen whether dorsal representations respond more or less invariantly
depending upon the format in which the stimulus is displayed (Holler et al., 2020; Snow et al.,
2011), and whether or not the stimulus classes investigated are items that would typically be
grasped and manipulated with the hands. It also remains an empirical question as to whether
shape images (es decir., a picture of a baseball or a mug) would stimulate dorsal cortex more strongly
if they were displayed so that the retinal size matched the real-world size, or indicated that
the object was an appropriate size for manipulation. Although recent neuropsychological ev-
idence suggests that manipulations of image size alone may not be sufficient to engage dorsal
corteza (Holler, behrman, & Snow, 2019), it remains to be seen whether additional cues to
tamaño, such as stereoscopic depth or background scene context, could engage dorsal cortex more
effectively.
Powerful links between perception and action emerge over time, and over the course
of development, in parallel with repeated experience with objects in the natural environment
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
47
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
.
/
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
(Bertenthal, 1996; Fan et al., 2020; Lockman & Kahrs, 2017). These encounters may be the
source of such bidirectional perception–action interactions. One compelling demonstration of
how goal-directed actions shape perception comes from disruptions to visual perception in
human observers who actively or passively explored the natural environment while wearing
vision-adjusting prism lenses (Held & Bossom, 1961; Held & Mikaelian, 1964). La necesidad de
coupling perception and action over development is even more clearly demonstrated in the
early “kitten-carousel” study by Held and Hein (1963) in which two kittens reared in darkness
from birth were harnessed to opposite ends of a horizontal arm in a carousel. Both animals
received the same visual stimulation but one kitten was harnessed with its feet touching the
ground, allowing it to walk actively around the carousel, while the other was harnessed with
its feet on a platform and was moved passively by the “self-moving” kitten partner. Más tarde, cuando
the kittens were removed from the carousel, the self-moving kitten demonstrated normal vi-
sual perception, while the passively moving kitten showed abnormalities in visually guided
paw placements, as well as in depth and distance perception. An analogous finding from hu-
mans comes from the study of individuals whose sight was restored surgically (Held et al.,
2011). The key question was whether previously blind individuals would be able to recognize
visually an object that had only been perceived tactilely. The individuals failed to recognize
the object visually immediately after their sight was restored, but crossmodal mappings devel-
oped rapidly, and after only 5 days of natural real-world visual experience their crossmodal
matching ability was close to ceiling (Held et al., 2011). This suggests that real-world action
can tune or recalibrate this intersensory matching process rather rapidly. (See Box 2 for an
elaborated discussion on the development of dorsal object representations.)
One particularly persuasive piece of evidence for the role of dorsal cortex in object
percepción, entirely independent of action, would be findings that show that dorsal cortex is
engaged in object perception in patients who are paralyzed or who never had use of their
manos. Such data would support the view that object perception is not solely contingent on
the presence of action or on action-relevant affordances (Gibson, 1979). Although we have
not been able to find such data, one finding worth noting is that patients born without hands
(dysplasia) and who use their feet instead of hands, appear to have typical motor selectivity
(albeit in inferior parietal lobule, rather than in motor cortex) leading to the suggestion that
high-level representations are effector-invariant (and perhaps affordance based) (Striem-Amit,
Vannuscorps, & Caramazza, 2018; Vannuscorps & Caramazza, 2016). Relacionado, in these in-
dividuals, it is also the case that motor cortex or other regions (like the inferior parietal lobule)
show motor selectivity of the compensatory effector (Striem-Amit et al., 2018; Vannuscorps,
Wurm, Striem-Amit, & Caramazza, 2019). A second set of related studies with dysplasic partic-
ipants has reported the finding of normal hand- and tool-selectivity and their overlap in ventral
corteza, reflecting functional organization in ventral cortex even in the absence of sensorimotor
experiencia (Striem-Amit, Vannuscorps, & Caramazza, 2017). As evident, although the above
findings are of interest, the most compelling data of dorsal-based object perception in the ab-
sence of any action constraint have not been reported as yet. Such a finding would further
cement the claim of independence of dorsal object representations.
We have suggested above that dorsal cortex selectively processes certain visual attributes
because of the constraints they impose on perception in the service of action, and that this
perception–action relationship appears to follow a global posterior-to-anterior visual-to-motor
gradient, with more posterior regions such as caudal intraparietal area (CIP) devoted to pro-
cessing features such as 3D curvature, and more anterior regions such as anterior intrapari-
etal sulcus (aIPS) involved in computing the metrics required for motor actions with objects
(Culham & Valyear, 2006; Fabbri, Stubbs, Cusack, & Culham, 2016; Shmuelof & Zohary, 2005;
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
48
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
.
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
Box 2. Developmental emergence of dorsal object representations.
One source of data that might inform our understanding of the relative contribution of dorsal
object representations and its coupling to action comes from studies of the developmental
emergence of dorsal cortex and motor development. The critical hypothesis is that if dor-
sal object representations are configured or constrained by action, then one might expect
that the dorsal pathway would not represent object perception until after motor systems are
mature enough to permit interaction with objects (see section What Gives Rise to Dorsal
Pathway Representations? for related questions addressed in adults with movement impair-
mentos). The early emergence of reaching behaviors is consistent with the notion that action
such as reaching not only influences action perception (Cannon, Woodward, Gredebäck,
von Hofsten, & Turek, 2012; Sommerville, Woodward, & Needham, 2005) but also influ-
ences visual perception in important ways, especially during early development (Needham
& Libertus, 2011).
Although there has not been a study conducted across ages (either longitudinal or
cross-sectional) that is specifically and systematically focused on the chronological emer-
gence of perception vis-à-vis action, there do exist data that can shed light on this matter.
Por ejemplo, several studies have argued that prehension in infants can significantly guide
the infants’ learning about objects and their properties. In one such study, infants were di-
vided into two groups, one of which was able to pick up objects using active “sticky” mittens
and the other of which simply watched their parents interact with the objects (Libertus &
Needham, 2010). Only the infants from the former condition showed changes in their visual
exploration of agents and objects in a live setting, leading to the conclusion that early action
can influence reaching as well as visual behavior. A similar finding was obtained in a later
study in which infants played with toys. One group of infants again was fitted with sticky mit-
tens while a second group had passive “nonsticky” mittens. In advance of this, infants were
given a teether to explore. Only infants who were in the active sticky mittens condition, pero
not those in the passive condition, demonstrated an increase in looking at and exploring the
teether more after than before training (Needham, Wiesen, Hejazi, Libertus, & Christopher,
2017). Y, relatedly, young infants either manipulated objects actively themselves or re-
ceived objects passively presented to them (Needham & Libertus, 2011). Spontaneous ori-
enting toward faces and objects was only evident in the former and not in the latter group,
revealing a potential link between manual engagement and the development of orienting
toward faces.
These findings argue for the influence of early actions on perception. There are also
studies offering evidence for slightly later development of visual perception of 3D objects,
which would allow for earlier emergence of action skills. Por ejemplo, infants only show sen-
sitivity to 3D object completion at 6 months of age (with girls at only 9 months of age) (Soska
& Johnson 2008, 2013). Por último, there are some data showing that even if action does not pre-
cede perception, these skills emerge concurrently. Por ejemplo, object completion abilities
emerge in conjunction with developing motor skills, most notably, independent sitting and
visual-manual exploration, as these two skills predict looking behavior in an incomplete
3D object task (Soska, Adolph, & Johnson, 2010). Juntos, these studies argue in favor of
action-based representations helping configure perceptual representations.
Sin embargo, differentiating the contribution of action entirely independent from that
of perception is not trivial. Por ejemplo, in one study, infants aged 5 a 15 months viewed
different sets of balls; one set was rigid in structure and required a full-hand power grasp, y
the other set was nonrigid and could be acquired with full precision grasp just with fingertips
(Barrett, Traupman, & Needham, 2008). Reaching movement was evaluated prior to contact
with the ball and the results showed very different forms of prehension for the two sets of balls.
This led to the conclusion that visuomotor constraints that are present early in development
led to the appropriate differentiation of the grasp. En otras palabras, the reaching (or action
solo) was not sufficient and visual information about the balls contributed to alterations
in reaching. En suma, reaching definitive conclusions about the emerging sequence of action
versus perception and their influence on each other is not always possible, and while action
certainly appears to modulate perception, the converse may likely be true as well.
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
49
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
Rígido & Zohary, 2008; also see Freud et al., 2016). Through this lens, the notion of perception
in the service of action could perhaps be characterized as a Bayesian prior that is scaled by
object size in the context of body space. Behavioral and neurophysiological findings point
to the existence of a network of areas along occipitoparietal cortex, including cortical areas
in humans such as V3A, V7/IPS0, and IPS, that selectively process object attributes that are
important for action (Erlikhman et al., 2018). Notablemente, many of these regions of parietal cortex
represent actions within the co-ordinate reference frame of specific effectors, such as the eyes,
manos, or arms (andersen & Buneo, 2002; andersen, Snyder, Bradley, & Xing, 1997; Colby &
Goldberg, 1999; Gallivan & Culham, 2015).
There also appears to be an emphasis within dorsal cortex on the representation of ego-
centric distance and peripersonal space—the area around the body that is within reach of
the arms (Duhamel, Bremmer, Hamed, & Graf, 1997; Galati et al., 2000; Galletti, Battaglini, &
Fattori, 1993; Rizzolatti, Fadiga, Gallese, & Fogassi, 1996; Vallar et al., 1999). En tono rimbombante, uno
región, the superior occipitotemporal cortex (SPOC), selectively responds to graspable objects
that are within reach of the observer, but not when they lie outside of reach, even when no
physical grasping action is required toward the object (Cavina-Pratesi et al., 2018; Gallivan,
Cavina-Pratesi, & Culham, 2009). Similarmente, some dorsal regions tuned to stereo-depth show
greater sensitivity to manipulations of egocentric versus allocentric distance (Neggers, Van der
Lubbe, Ramsey, & postma, 2006; Neri, Bridge, & Heeger, 2004). Convergent evidence for
the involvement of dorsal cortex in egocentric distance perception comes from neuropsycho-
logical patients with lesions of parietal cortex (Cerebro, 1941). Some patients show selective
impairments in estimating the distance from the body to nearby objects, or placing objects
at specific distances from the body (Berryhill et al., 2009), although such deficits could also
reflect more fundamental impairments in spatial or orientation processing that are limited to
the egocentric reference frame.
Dorsal Cortex May Play a Unique Role in Perceiving Action-Relevant Stimuli and Environments
Despite the fact that, in everyday life, humans predominantly interact with solid objects in
real-world 3D environments, visual perception has classically been studied in experimental
psychology and neuroscience using impoverished stimuli in the form of static two-dimensional
(2D) images of objects. One problem with this approach, particularly for understanding the
functional role of dorsal cortex and its potential role in coding potential actions with objects
and environments, is that 2D images convey little, if any, information about the types of visual
attributes that dorsal cortex is uniquely sensitive to, such as 3D shape and depth, SfM, egocen-
tric distance, and real-world size. Por ejemplo, from the perspective of an observer looking
at a planar image of an object on a computer screen, the distance to the projection surface is
conocido, but not the distance to the depicted object. The real-world size of the stimuli used in
studies of visual perception is further obscured when objects are presented as pictorial cutouts
abstracted from their corresponding background, and when the retinal extent of items that are
typically large in the world (es decir., a horse) is matched with items that are orders of magnitude
smaller (es decir., a butterfly) (Bracci & Op de Beeck, 2016; Cichy, Kriegeskorte, Jozwik, van den
Bosch, & Charest, 2019; Konkle & Oliva, 2012; Kriegeskorte, Mur, & Bandettini, 2008).
Curiosamente, convergent evidence from studies using a range of empirical approaches,
including behavioral psychophysics, resonancia magnética funcional, EEG, and neuropsychology, has begun to highlight
fundamental differences in the way real objects and computerized images are processed dur-
ing perception (Gerhard, Culham, & Schwarzer, 2016; Holler et al., 2019, 2020; escuderos,
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
50
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
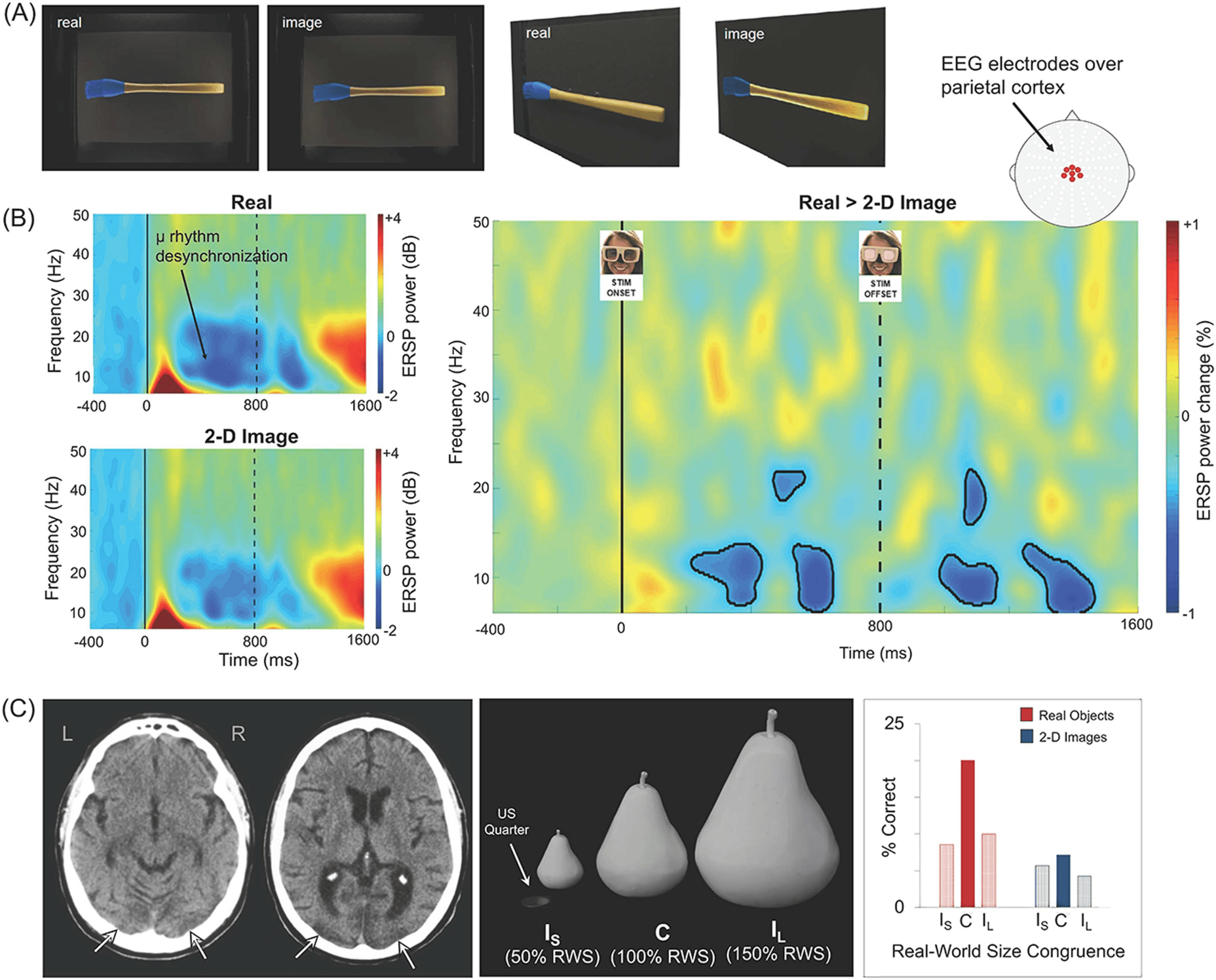
Cifra 3. The processing of real objects (A). Stimuli used in an EEG study by Marini et al.
(2019) to investigate whether real-world objects
elicit distinct visuomotor brain dynamics compared to matched 2D images of the same items. In the study, EEG recordings were collected
while observers viewed real tools and 2D images of tools. When viewed from the front, the stimuli were matched closely for apparent size,
distancia, fondo, illumination, and color (left panel). For illustrative purposes, the same stimuli are shown (right panel) from an oblique
perspectiva. (B). Real objects increased the strength and duration of activation in brain networks involved in automatic action planning relative
to images of objects. In the study described, Marini et al. (2019) decomposed EEG signals to reveal desynchronization of the µ (“mu”) ritmo
(8–13 Hz)—a frequency-specific change associated with the automatic transformation of visual object information into action representations.
Left panels illustrate event-related spectral perturbation (ERSP) power recorded from central electrodes positioned over parietal cortex, mostrado
separately for real objects (upper panel) and 2D images (lower panel). Although both stimulus formats elicited µ rhythm desynchronization, este
effect was stronger and more sustained for real objects in comparison to 2D images (right panel, black outlines demarcate areas of statistical
significance). The stronger µ desynchronization for the real objects (versus images) was apparent during stimulus presentation and persisted
after stimulus offset (demarcated by dashed vertical line) (Marini et al., 2019). (C). Real-world size coding of solid objects in a patient with
visual agnosia. JW, a patient with bilateral lesions to occipitotemporal cortex (left panel) was presented with solid objects and 2D computerized
images of the same items that were scaled to be incongruent smaller (Es), congruente (C), or incongruent larger (IL) than typical real-world size
(middle panel). While recognition of images was extremely poor, real object recognition was surprisingly preserved, but only when physical
size matched real-world size (right panel) (Holler et al., 2019).
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
51
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
macdonald, Culham, & Snow, 2016), memory (Snow, Skiba, Coleman, & Berryhill, 2014), en-
tention (Gomez et al., 2017), and decision-making (Romero, Compton, Cual, & Snow, 2018).
En tono rimbombante, these effects may be driven by the selective recruitment of dorsal cortex during
perceptual processing. Por ejemplo, passively viewing real objects triggers stronger and more
prolonged automatic motor preparation signals than does viewing matched images of the same
objects, as measured by high-density EEG over parietal cortex, particularly in the hemisphere
contralateral to the dominant hand (Marini et al., 2019) (see Figure 3A–Figure 3B).
Compellingly, evidence from neuropsychology further suggests that dorsal cortex is criti-
cally involved in perceiving object shape and size, specifically for real objects (but not images).
Holler et al. (2019) examined object recognition in patients with severe visual agnosia result-
ing from bilateral lesions of shape processing areas in ventral cortex (see Figure 3C). A pesar de
the patients were severely impaired in their ability to recognize 2D images of objects, ellos
showed a striking preservation in their ability to recognize real-world exemplars of the same
estímulos. críticamente, sin embargo, the recognition advantage shown by the patients for the real ob-
jects was only apparent when the physical size of the (real) objects was consistent with the
typical real-world size. Recognition of objects whose physical size deviated from real-world
size was severely impaired and similar to 2D images. Analogous manipulations of the visual
size of 2D computerized images did not modulate recognition performance. These findings are
all consistent with the claim that visual representations in dorsal cortex may be constrained by
real-world properties of objects and it is these representations that then primarily serve action
as well as perception.
CONCLUSIONS
En resumen, we have reviewed evidence that dorsal cortex is particularly sensitive to a range
of visuospatial attributes including SfM, 3D geometric shape, orientación, and size. Tenemos
also advanced the idea that dorsal cortex selectively processes these visual attributes because
of the constraints they impose for perception in the service of action, y eso, while these
constraints are predominantly relevant in the context of real-world objects and environments,
they can also mediate vision for perception. This characterization of the role of dorsal cortex
for perception contrasts with traditional frameworks that ascribed a more monolithic role that
centered upon action alone, as well as with conceptualizations of the role of ventral cortex,
in which object processing operates predominantly to achieve invariant responses to objects
to stabilize conscious perception despite changing visual conditions.
One of the reasons why it has been difficult to characterize fully the role of dorsal cortex
is that working with more naturalistic conditions and real objects in the laboratory presents a
number of practical and empirical challenges that are not encountered when using image dis-
plays (Romero & Snow, 2019). Whether or not dorsal cortex is particularly tuned to processing
actionable solid objects or is also tuned for perception independent of action, and whether
dorsal cortex responds only or specifically to visual properties that are relevant to action, son
questions of outstanding importance that require careful investigation. To further advance our
understanding of the two cortical pathways, future experiments should include a wider range
of required behaviors and stimuli.
INFORMACIÓN DE FINANCIACIÓN
JCS, National Eye Institute, Award ID: R01EY026701, the National Science Foundation (NSF),
Award ID: 1632849; and the Clinical Translational Research Infrastructure Network, Award ID:
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
52
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
17-746Q-UNR-PG53-00. MB, National Eye Institute, Award ID: RO1EY027018. EF, Natural
Sciences and Engineering Research Council of Canada (501100000038), Award ID: NA, y
Vision Science to Applications (VISTA) program funded by the Canada First Research Excel-
lence Fund (CFREF, 2016–2023) (EF).
CONTRIBUCIONES DE AUTOR
EF: Conceptualización: Equal; Visualización: Equal; Writing – Original Draft: Equal; Writing –
Revisar & Editing: Equal. MB: Conceptualización: Equal; Writing – Revisar & Editing: Equal.
JCS: Conceptualización: Equal; Visualización: Equal; Writing – Original Draft: Equal; Writing –
Revisar & Editing: Equal.
REFERENCIAS
andersen, R. A., & Buneo, C. A. (2002). Intentional maps in posterior
parietal cortex. Revisión anual de neurociencia, 25, 189–220.
andersen, R. A., Snyder, l. h., Bradley, D. C., & Xing, j. (1997). Mul-
timodal representation of space in the posterior parietal cortex
and its use in planning movements. Annual Review of Neuro-
ciencia, 20, 303–330.
Bar, METRO. (2004). Visual objects in context. Nature Reviews Neuro-
ciencia, 5, 617–629.
Barrett, t. METRO., Traupman, MI., & Needham, A. (2008). Infants’ visual
anticipation of object structure in grasp planning. Infant Behavior
and Development, 31(1), 1–9.
behrman, METRO., Geng, j. J., & Shomstein, S. (2004). Parietal cortex
and attention. Opinión actual en neurobiología, 14, 212–217.
https://doi.org/1016/j.conb.2004.03.012
Berryhill, METRO. MI., Fendrich, r., & Olson, I. R. (2009). Impaired dis-
tance perception and size constancy following bilateral occipito-
parietal damage. Experimental Brain Research, 194, 381–393.
https://doi.org/10.1007/s00221-009-1707-7
Bertenthal, B. I. (1996). Origins and early development of percep-
ción, acción, and representation. Annual Review of Psychology,
47, 431–459.
Bracci, S., & Op de Beeck, h. (2016). Dissociations and associations
between shape and category representations in the two visual
pathways. The Journal of Neuroscience, 36, 432–444. https://
doi.org/10.1523/JNEUROSCI.2314-15.2016
Cerebro, W.. R.
(1941). Visual disorientation with special reference
to lesions of the right cerebral hemisphere. Cerebro, 64, 244–272.
https://doi.org/10.1093/brain/64.4.244
Brockmole, j. r., Davoli, C. C., Abrams, R. A., & Witt, j. k. (2013).
The world within reach: Effects of hand posture and tool use on
visual cognition. Current Directions in Psychological Science, 22,
38–44. https://doi.org/10.1177/0963721412465065
Buschman, t. J., & Kastner, S. (2015). From behavior to neural dy-
namics: An integrated theory of attention. Neurona, 88, 127–144.
Cannon, mi. NORTE., Woodward, A. l., Gredebäck, GRAMO., von Hofsten, C., &
Turek, C. (2012). Action production influences 12-month-old in-
fants’ attention to others’ actions. Developmental Science, 15(1),
35–42.
Castiello, Ud.. (2005). The neuroscience of grasping. Reseñas de naturaleza
Neurociencia, 6, 726–736.
Cavina-Pratesi, C., Connolly, j. D., Monaco, S., Figley, t. D., Milner,
A. D., Schenk, T., & Culham, j. C. (2018). Human neuroimaging
reveals the subcomponents of grasping, reaching and pointing
comportamiento. Corteza, 98, 128–148.
chao, l. l., & Martín, A. (2000). Representation of manipulable man-
made objects in the dorsal stream. NeuroImagen, 12, 478–484.
https://doi.org/10.1006/nimg.2000.0635
Cichy, R. METRO., Kriegeskorte, NORTE., Jozwik, k. METRO., van den Bosch, j. j. F.,
& Charest, I. (2019). The spatiotemporal neural dynamics under-
lying perceived similarity for real-world objects. NeuroImagen, 194,
12–24. https://doi.org/10.1016/j.neuroimage.2019.03.031
Ciesielski, k. t. r., Stern, METRO. MI., Diamante, A., Kan, S., Busa, mi. A.,
Goldsmith, t. MI., van der Kouwe, A., pescado, B., & rosa, B. R.
(2019). Maturational changes in human dorsal and ventral visual
redes. Corteza cerebral, 29, 5131–5149. https://doi.org/10.
1093/cercor/bhz053
Un cuerpo, PAG., & Kalaska, j. F. (2010). Neural mechanisms for interacting
with a world full of action choices. Annual Review of Neuro-
ciencia, 33, 269–298.
Colby, C. l., & Goldberg, METRO. mi. (1999). Space and attention in pari-
etal cortex. Revisión anual de neurociencia, 22, 319–349.
collins, MI., Freud, MI., Kainerstorfer, j. METRO., Cao, J., & behrman, METRO.
(2019). Temporal dynamics of shape processing differentiate
Diario de
contributions of dorsal and ventral visual pathways.
Neurociencia Cognitiva, 31, 821–836.
Cox, D. D.
Cosman, j. D., & Vecera, S. PAG. (2010). Attention affects visual percep-
tual processing near the hand. ciencia psicológica, 21, 1254–1258.
(2014). Do we understand high-level vision? Actual
Opinion in Neurobiology, 25, 187–193. https://doi.org/10.1016/
j.conb.2014.01.016
Culham, j. C., & Valyear, k. F. (2006). Human parietal cortex in ac-
ción. Opinión actual en neurobiología, 16, 205–212.
D'Esposito, METRO., & Postle, B. R. (2015). The cognitive neuroscience
de memoria de trabajo. Annual Review of Psychology, 66, 115–142.
Dilks, D. D., Julian, j. B., Kubilius, J., Spelke, mi. S., & Kanwisher,
norte.
(2011). Mirror-image sensitivity and invariance in object
and scene processing pathways. The Journal of Neuroscience,
31, 11305–11312. https://doi.org/10.1523/JNEUROSCI.1935-
11.2011
Duhamel, J.-R., Bremmer, F., Hamed, S. B., & Graf, W.. (1997). Spa-
tial invariance of visual receptive fields in parietal cortex neurons.
Naturaleza, 389, 845–848.
Erlikhman, GRAMO., Caplovitz, GRAMO. PAG., Gurariy, GRAMO., Medina, J., & Snow,
j. C. (2018). Towards a unified perspective of object shape and
motion processing in human dorsal cortex. Consciousness and
Cognición, 64, 106–120.
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
53
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
Erlikhman, GRAMO., Gurariy, GRAMO., Mruczek, R. MI., & Caplovitz, GRAMO. PAG. (2016).
The neural representation of objects formed through the spa-
tiotemporal integration of visual transients. NeuroImagen, 142,
67–78.
Fabbri, S., Stubbs, k. METRO., Cusack, r., & Culham, j. C. (2016). Dis-
entangling representations of object and grasp properties in the
cerebro humano. The Journal of Neuroscience, 36, 7648–7662.
Admirador, j. MI., Wammes, j. D., Gunn, j. B., Yamins, D. l. K., Norman,
k. A., & Turk-Browne, norte. B. (2020). Relating visual production
and recognition of objects in human visual cortex. The Jour-
nal of Neuroscience, 40, 1710–1721. https://doi.org/10.1523/
JNEUROSCI.1843-19.2019
Farivar, R. (2009). Dorsal–ventral integration in object recognition.
Brain Research Reviews, 61(2), 144–153.
Freud, MI., & behrman, METRO.
(2020). Altered large-scale organiza-
tion of shape processing in visual agnosia. Corteza, 129, 423–435.
https://doi.org/10.1016/j.cortex.2020.05.009
Freud, MI., Culham, j. C., Plaut, D. C., & behrman, METRO. (2017). El
large-scale organization of shape processing in the ventral and
dorsal pathways. ELife, 6, e27576. https://doi.org/10.7554/eLife.
27576
Freud, MI., Ganel, T., Shelef, I., Hammer, METRO. D., Avaro, GRAMO., &
behrman, METRO. (2017). Three-dimensional representations of ob-
jects in dorsal cortex are dissociable from those in ventral cortex.
Corteza cerebral, 27, 422–434. https://doi.org/10.1093/cercor/
bhv229
Freud, MI., Plaut, D. C., & behrman, METRO. (2016). “What” is happen-
ing in the dorsal visual pathway. Tendencias en Ciencias Cognitivas,
20, 773–784. https://doi.org/10.1016/j.tics.2016.08.003
Freud, MI., robinson, A. K., & behrman, METRO. (2018). More than ac-
ción: The dorsal pathway contributes to the perception of 3-D
estructura. Revista de neurociencia cognitiva, 30, 1047–1058.
Freud, MI., Rosenthal, GRAMO., Ganel, T., & Avaro, GRAMO. (2015). Sensitivity
to object impossibility in the human visual cortex: Evidencia de
conectividad funcional. Revista de neurociencia cognitiva, 27,
1029–1043. https://doi.org/10.1162/jocn_a_00753
Galati, GRAMO., Lobel, MI., Vallar, GRAMO., Berthoz, A., Pizzamiglio, l., & Le
Bihan, D. (2000). The neural basis of egocentric and allocentric
coding of space in humans: A functional magnetic resonance
estudiar. Experimental Brain Research, 133(2), 156–164. https://doi.
org/10.1007/s002210000375
Galletti, C., Battaglini, PAG. PAG., & Fattori, PAG. (1993). Parietal neurons en-
coding spatial locations in craniotopic coordinates. Experimental
Brain Research, 96, 221–229.
Gallivan, j. PAG., Cavina-Pratesi, C., & Culham, j. C.
Is that
within reach? FMRI reveals that the human superior parieto-
occipital cortex encodes objects reachable by the hand. The Jour-
nal of Neuroscience, 29, 4381–4391.
(2009).
Gallivan, j. PAG., & Culham, j. C. (2015). Neural coding within human
brain areas involved in actions. Current Opinion in Neurobiol-
ogia, 33, 141–149. https://doi.org/10.1016/j.conb.2015.03.012
J., anderson, A. w.,
Skudlarski, PAG., & Sangre, j. C. (2002). BOLD activity during mental
rotation and viewpoint-dependent object recognition. Neurona,
34, 161–171.
I., Hayward, W.. GRAMO., Tarr, METRO.
Gauthier,
Georgieva, S. S., Peeters, r., Kolster, h., Todd, j. T., & Orban, GRAMO. A.
(2009). The processing of three-dimensional shape from disparity
en el cerebro humano. The Journal of Neuroscience, 29, 727–742.
https://doi.org/10.1523/JNEUROSCI.4753-08.2009
Georgieva, S. S., Todd, j. T., Peeters, r., & Orban, GRAMO. A. (2008). El
extraction of 3D shape from texture and shading in the human
cerebro. Corteza cerebral, 18, 2416–2438. https://doi.org/10.1093/
cercor/bhn002
Gerhard, t. METRO., Culham, j. C., & Schwarzer, GRAMO.
(2016). Distinto
visual processing of real objects and pictures of those objects in
7-to 9-month-old infants. Fronteras en psicología, 7, 827.
Ghazanfar, A., & Schroeder, C. (2006). Is neocortex essentially mul-
tisensory? Tendencias en Ciencias Cognitivas, 10, 278–285. https://doi.
org/10.1016/j.tics.2006.04.008
Gibson, j. j. (1979). The ecological approach to visual perception.
Bostón, MAMÁ: Houghton Mifflin.
Gilaie-Dotan, S., Saygin, A. PAG., Lorenzi, l. J., rees, GRAMO., & behrman,
METRO. (2015). Ventral aspect of the visual form pathway is not crit-
ical for the perception of biological motion. Actas de la
Academia Nacional de Ciencias, 112, E361–E370.
Gómez, J., barnett, METRO. A., Natu, v., Mezer, A., Palomero-Gallagher,
NORTE., Weiner, k. S., Amunts, K., Zilles, K., & Grill-Spector, k.
(2017). Microstructural proliferation in human cortex is coupled
with the development of face processing. Ciencia, 355, 68–71.
Goodale, METRO. A., & Milner, A. D. (1992). Separate visual pathways
for perception and action. Trends in Neurosciences, 15, 20–25.
https://doi.org/0166-2236(92)90344-8
Goodale, METRO. A., & Milner, A. D.
(2018). Two visual pathways—
Where have they taken us and where will they lead in future?
Corteza, 98, 283–292.
Gozli, D. GRAMO., Oeste, GRAMO. l., & Pratt, j.
(2012). Hand position alters
vision by biasing processing through different visual pathways.
Cognición, 124, 244–250. https://doi.org/10.1016/j.cognition.
2012.04.008
Greenberg, A. S., Petrificar, T., Chiu, Y.-C., Yantis, S., Schneider,
w., & behrman, METRO.
(2012). Visuotopic cortical connectiv-
ity underlying attention revealed with white-matter tractography.
The Journal of Neuroscience, 32, 2773–2782. https://doi.org/10.
1523/JNEUROSCI.5419-11.2012
Grill-Spector, K., Kourtzi, Z., & Kanwisher, norte.
(2001). The lateral
occipital complex and its role in object recognition. Vision Re-
buscar, 41, 1409–1422. https://doi.org/S0042-6989(01)00073-6
Grill-Spector, K., Kushnir, T., Hendler, T., Edelman, S., Itzchak,
y., & Malach, R.
(1998). A sequence of object-processing
stages revealed by fMRI in the human occipital lobe. Humano
Mapeo cerebral, 6, 316–328. https://doi.org/10.1002/(CIENCIA)1097-
0193(1998)6:4<316::AID-HBM9>3.0.CO;2-6
Grill-Spector, K., & Malach, R. (2004). The human visual cortex. Un-
nual Review of Neuroscience, 27, 649–677. https://doi.org/10.
1146/annurev.neuro.27.070203.144220
harvey, B. METRO., Klein, B. PAG., Petridou, NORTE., & Dumoulin, S. oh. (2013).
Topographic representation of numerosity in the human parietal
corteza. Ciencia, 341, 1123–1126.
Heft, h. (2013). Environment, cognition, and culture: Reconsider-
ing the cognitive map. Journal of Environmental Psychology, 33,
14–25.
Held, r., & Bossom, j. (1961). Neonatal deprivation and adult re-
arrangement: Complementary techniques for analyzing plastic
sensory-motor coordinations. Journal of Comparative and Physi-
ological Psychology, 54, 33–37.
Held, r., & Hein, A. (1963). Movement-produced stimulation in the
development of visually guided behavior. Journal of Comparative
and Physiological Psychology, 56, 872–876.
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
54
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
/
.
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
Held, r., & Mikaelian, h.
(1964). Motor-sensory feedback versus
need in adaptation to rearrangement. Perceptual and Motor Skills,
18, 685–688.
Held, r., Ostrovsky, y., de Gelder, B., Gandhi, T., Ganesh, S.,
Mathur, Ud., & Sinha, PAG. (2011). The newly sighted fail to match
seen with felt. Neurociencia de la naturaleza, 14, 551–553. https://doi.
org/10.1038/nn.2795
Holler, D. MI., behrman, METRO., & Snow, j. C. (2019). Real-world size
coding of solid objects, but not 2-D or 3-D images, in visual ag-
nosia patients with bilateral ventral lesions. Corteza, 119, 555–568.
https://doi.org/10.1016/j.cortex.2019.02.030
Holler, D. MI., Fabbri, S., & Snow, j. C. (2020). Object responses are
highly malleable, rather than invariant, with changes in object
apariencia. Scientific Reports, 10(1), 1–14.
Janssen, PAG., Vogels, r., & Orban, GRAMO. A.
(2000). Selectivity for 3D
shape that reveals distinct areas within macaque inferior tempo-
ral cortex. Ciencia, 288, 2054–2056. https://doi.org/10.1126/
science.288.5473.2054
Jeannerod, METRO. (1981). Intersegmental coordination during reaching
at natural visual objects. Attention and Performance IX, 9, 153–168.
Jeannerod, METRO. (1986). The formation of finger grip during prehen-
sión. A cortically mediated visuomotor pattern. Behavioural Brain
Investigación, 19, 99–116.
jeong, S. K., & Xu, Y. (2016). Behaviorally relevant abstract object
identity representation in the human parietal cortex. The Jour-
nal of Neuroscience, 36, 1607–1619. https://doi.org/10.1523/
JNEUROSCI.1016-15.2016
Konen, C. S., & Kastner, S. (2008). Two hierarchically organized neu-
ral systems for object information in human visual cortex. Naturaleza
Neurociencia, 11, 224–231. https://doi.org/10.1038/nn2036
Konkle, T., & Oliva, A. (2012). A real-world size organization of object
responses in occipitotemporal cortex. Neurona, 74, 1114–1124.
Kourtzi, Z., & Kanwisher, norte. (2000). Cortical regions involved in perceiv-
ing object shape. The Journal of Neuroscience, 20, 3310–3318.
Kravitz, D. J., Saleem, k. S., Panadero, C. I., & Mishkin, METRO. (2011). A new
neural framework for visuospatial processing. Reseñas de naturaleza
Neurociencia, 12, 217–230. https://doi.org/10.1038/nrn3008
Kravitz, D. J., Vinson, l. D., & Panadero, C. I.
(2008). How position
dependent is visual object recognition?. Trends in Cognitive Sci-
ences, 12(3), 114–122.
Kriegeskorte, NORTE., Mur, METRO., & Bandettini, PAG. A. (2008). Representa-
tional similarity analysis-connecting the branches of systems neuro-
ciencia. Frontiers in Systems Neuroscience, 2, 4. https://doi.org/
10.3389/neuro.06.004.2008
Lehky, S. r., & Sereno, A. B.
(2007). Comparison of shape en-
coding in primate dorsal and ventral visual pathways. Diario de
Neurofisiología, 97, 307–319.
Libertus, K., & Needham, A. (2010). Teach to reach: The effects of
active vs. passive reaching experiences on action and perception.
Vision Research, 50, 2750–2757.
Liu, l., Wang, F., zhou, K., Ding, NORTE., & luo, h. (2017). Perceptivo
integration rapidly activates dorsal visual pathway to guide local
processing in early visual areas. Biología PLoS, 15, e2003646.
Lockman, j. J., & Kahrs, B. A. (2017). New insights into the devel-
opment of human tool use. Current Directions in Psychological
Ciencia, 26, 330–334.
Logothetis, norte. K., & Sheinberg, D. l. (1996). Visual object recogni-
ción. Revisión anual de neurociencia, 19, 577–621.
Mahon, B. Z., Kumar, NORTE., & Almeida, j. (2013). Spatial frequency
tuning reveals interactions between the dorsal and ventral vi-
Revista de neurociencia cognitiva, 25, 862–871.
sual systems.
https://doi.org/10.1162/jocn_a_00370
Malach, r., Reppas, j. B., Benson, R. r., Kwong, k. K., Jiang, h.,
Kennedy, W.. A., Ledden, PAG. J., Brady, t. J., rosa, B. r., & Tootell,
R. B. (1995). Object-related activity revealed by functional mag-
netic resonance imaging in human occipital cortex. Actas
of the National Academy of Sciences of the United States of
America, 92, 8135–8139.
Marini, F., Breeding, k. A., & Snow, j. C. (2019). Distinct visuo-motor
brain dynamics for real-world objects versus planar images. Neuro-
Image, 195, 232–242.
https://doi.org/10.1016/j.neuroimage.
2019.02.026
Martinaud, o., Mirlink, NORTE., Bioux, S., Bliaux, MI., Champmartin, C.,
Pouliquen, D., Cruypeninck, y., Hannequin, D., & Gérardin, mi.
(2016). Mirrored and rotated stimuli are not the same: A neu-
ropsychological and lesion mapping study. Corteza, 78, 100–114.
https://doi.org/10.1016/j.cortex.2016.03.002
Martinaud, o., Mirlink, NORTE., Bioux, S., Bliaux, MI., Lebas, A., Gerardin,
MI., & Hannequin, D. (2014). Agnosia for mirror stimuli: A new
case report with a small parietal lesion. Archives of Clinical
Neuropsychology, 29, 724–728. https://doi.org/10.1093/arclin/
acu032
Medina, J., Jax, S. A., & Coslett, h. B. (2020). Impairments in ac-
tion and perception after right intraparietal damage. Corteza, 122,
288–299.
Mishkin, METRO., & Ungerleider, l. GRAMO.
(1982). Contribution of striate
inputs to the visuospatial functions of parieto-preoccipital cortex
in monkeys. Behavioural Brain Research, 6, 57–77.
Mishkin, METRO., Ungerleider, l. GRAMO., & Macko, k. A.
(1983). Object
vision and spatial vision: Two cortical pathways. Trends in Neu-
rosciences, 6, 414–417.
Mruczek, R. MI., von Loga, I. S., & Kastner, S. (2013). The represen-
tation of tool and non-tool object information in the human in-
traparietal sulcus. Revista de neurofisiología, 109, 2883–2896.
https://doi.org/10.1152/jn.00658.2012
Needham, A., & Libertus, k. (2011). Embodiment in early development.
Wiley Interdisciplinary Reviews: Ciencia cognitiva, 2(1), 117–123.
Needham, A. w., Wiesen, S. MI., Hejazi, j. NORTE., Libertus, K., &
(2017). Characteristics of brief sticky mittens
Diario de
Christopher, C.
training that lead to increases in object exploration.
Experimental Child Psychology, 164, 209–224.
Neggers, S. F., Van der Lubbe, R. h., Ramsey, norte. F., & postma, A.
(2006). Interactions between ego-and allocentric neuronal rep-
resentations of space. NeuroImagen, 31, 320–331.
Nelissen, K., Joly, o., Durand, J.-B., Todd, j. T., Vanduffel, w.,
& Orban, GRAMO. A.
(2009). The extraction of depth structure from
shading and texture in the macaque brain. Más uno, 4, e8306.
Neri, PAG., Bridge, h., & Heeger, D. j. (2004). Stereoscopic processing
of absolute and relative disparity in human visual cortex. Diario
of Neurophysiology, 92, 1880–1891.
Orban, GRAMO. A. (2011). The extraction of 3D shape in the visual system
of human and nonhuman primates. Annual Review of Neuro-
ciencia, 34, 361–388. https://doi.org/10.1146/annurev-neuro-
061010-113819
Orban, GRAMO. A., Janssen, PAG., & Vogels, R. (2006). Extracting 3D struc-
ture from disparity. Trends in Neurosciences, 29, 466–473.
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
55
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
.
/
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
/
.
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Dorsal Pathway Contributes to Perception Freud, behrman, Snow
Rizzolatti, GRAMO., Fadiga, l., Gallese, v., & Fogassi, l. (1996). Premo-
tor cortex and the recognition of motor actions. Cognitive Brain
Investigación, 3, 131–141.
Romero, C. A., Compton, METRO. T., Cual, y., & Snow, j. C. (2018). El
real deal: Willingness-to-pay and satiety expectations are greater
for real foods versus their images. Corteza, 107, 78–91. https://
doi.org/10.1016/j.cortex.2017.11.010
Romero, C. A., & Snow, j. C. (2019). Methods for presenting real-
Diario
world objects under controlled laboratory conditions.
of Visualized Experiments, 148, 59762. https://doi.org/10.3791/
59762
Rushworth, METRO. F. S., Behrens, t. mi. J., & Johansen-Berg, h. (2006).
Connection patterns distinguish 3 regions of human parietal cor-
tex. Corteza cerebral, 16, 1418–1430. doi:10.1093/cercor/bhj079
Sereno, A. B., & Maunsell, j. h. R. (1998). Shape selectivity in pri-
mate lateral intraparietal cortex. Naturaleza, 395, 500–503. https://
doi.org/10.1038/26752
Sereno, METRO. I., Valle, A. METRO., Reppas, j. B., Kwong, k. K., Belliveau,
j. w., Brady, t. J., rosa, B. r., & Tootell, R. B. (1995). Borders of
multiple visual areas in humans revealed by functional magnetic
resonance imaging. Ciencia, 268, 889–893.
Shmuelof, l., & Zohary, mi.
(2005). Dissociation between ventral
and dorsal fMRI activation during object and action recognition.
Neurona, 47, 457–470.
Smeets, j. B., & Brennero, mi. (1999). A new view on grasping. Motor
Control, 3, 237–271.
Snow, j. C., Pettypiece, C. MI., McAdam, t. D., McLean, A. D.,
Stroman, PAG. w., Goodale, METRO. A., & Culham, j. C. (2011). Bring-
ing the real world into the fMRI scanner: Repetition effects for
pictures versus real objects. Scientific Reports, 1, 130.
Snow, j. C., Skiba, R. METRO., Coleman, t. l., & Berryhill, METRO. mi. (2014).
Real-world objects are more memorable than photographs of ob-
jects. Frontiers in Human Neuroscience, 8, 837. http://www.
ncbi.nlm.nih.gov/pmc/articles/PMC4202719/
Sommerville, j. A., Woodward, A. l., & Needham, A. (2005). C.A-
tion experience alters 3-month-old infants’ perception of others’
comportamiento. Cognición, 96(1), B1–B11.
Soska, k. C., Adolph, k. MI., & Johnson, S. PAG. (2010). Systems in de-
velopment: Motor skill acquisition facilitates three-dimensional
object completion. Developmental Psychology, 46, 129–138.
https://doi.org/10.1037/a0014618
Soska, k. C., & Johnson, S. PAG.
(2008). Development of three-
dimensional object completion in infancy. Child Development,
79, 1230–1236. https://doi.org/10.1111/j.1467-8624.
2008.01185.X
Soska, k. C., & Johnson, S. PAG. (2013). Development of three-dimen-
sional completion of complex objects. Infancy, 18, 325–344.
escuderos, S. D., macdonald, S. NORTE., Culham, j. C., & Snow, j. C. (2016).
Priming tool actions: Are real objects more effective primes than
pictures? Experimental Brain Research, 234, 963–976.
Rígido, A., & Zohary, mi.
(2008). Parietal mapping of visuomotor
transformations during human tool grasping. Corteza cerebral,
18, 2358–2368.
Stepniewska, I., Cerkevich, C. METRO., & Aquél, j. h.
(2015). Cortical
connections of the caudal portion of posterior parietal cortex in
prosimian Galagos. Corteza cerebral, 26, 2753–2777.
Striem-Amit, MI., Vannuscorps, GRAMO., & Caramazza, A.
(2017).
Sensorimotor-independent development of hands and tools se-
Actas del Nacional
lectivity in the visual cortex.
Academia de Ciencias, 114, 4787–4792.
Striem-Amit, MI., Vannuscorps, GRAMO., & Caramazza, A. (2018). Plastic-
ity based on compensatory effector use in the association but not
primary sensorimotor cortex of people born without hands. Pro-
ceedings of the National Academy of Sciences, 115, 7801–7806.
Sugihara, h., Murakami, I., shenoy, k. v., andersen, R. A., &
(2002). Response of MSTd neurons to simulated
Revista de neurofisiología,
Komatsu, h.
3D orientation of rotating planes.
87, 273–285.
Swisher, j. D., Halko, METRO. A., Merabet, l. B., McMains, S. A., &
Somers, D. C. (2007). Visual topography of human intraparietal
sulcus. The Journal of Neuroscience, 27(20), 5326–5337. https://
doi.org/10.1523/JNEUROSCI.0991-07.2007
Vaina, l. METRO.
(1989). Selective impairment of visual motion inter-
pretation following lesions of the right occipito-parietal area in
humanos. Cibernética biológica, 61, 347–359.
Vallar, GRAMO., Lobel, MI., Galati, GRAMO., Berthoz, A., Pizzamiglio, l., &
(1999). A fronto-parietal system for computing
Le Bihan, D.
the egocentric spatial frame of reference in humans. Experi-
mental Brain Research, 124, 281–286. https://doi.org/10.1007/
s002210050624
Valyear, k. F., Culham, j. C., Sharif, NORTE., Westwood, D., & Goodale,
METRO. A. (2006). A double dissociationbetween sensitivity to changes
in object identity and object orientation in the ventral and dor-
sal visual streams: A human fMRI study. Neuropsicología, 44,
218–228.
Van Dromme, I. C., Premereur, MI., Verhoef, B.-E., Vanduffel, w., &
Janssen, PAG. (2016). Posterior parietal cortex drives inferotemporal
activations during three-dimensional object vision. Biología PLoS,
14, e1002445. https://doi.org/10.1371/journal.pbio.1002445
Vanduffel, w., Fize, D., Peuskens, h., Denys, K., Sunaert, S., Todd,
j. T., & Orban, GRAMO. A. (2002). Extracting 3d from motion: Diferencias
in human and monkey intraparietal cortex. Ciencia, 298, 413–415.
https://doi.org/10.1126/science.1073574
Vannuscorps, GRAMO., & Caramazza, A. (2016). Typical action percep-
tion and interpretation without motor simulation. Actas de
the National Academy of Sciences, 113, 86–91.
Vannuscorps, GRAMO., Wurm, METRO., Striem-Amit, MI., & Caramazza, A.
(2019). Large-scale organization of the hand action observation
network in individuals born without hands. Corteza cerebral, 29,
3434–3444.
Vaziri-Pashkam, METRO., & Xu, Y. (2017). Goal-directed visual process-
ing differentially impacts human ventral and dorsal visual repre-
sentaciones. The Journal of Neuroscience, 36, 8767–8782.
Vaziri-Pashkam, METRO., & Xu, Y. (2018). An information-driven 2-pathway
characterization of occipitotemporal and posterior parietal visual
object representations. Corteza cerebral, 29, 2034–2050.
Xu, Y. (2018). A tale of two visual systems: Invariant and adaptive
visual information representations in the primate brain. Annual
Review of Vision Science, 4, 311–336.
Yoshioka, t. w., Doi, T., Abdolrahmani, METRO., & Fujita, I. (2020). Spe-
cialization of mid-tier stages of dorsal and ventral pathways in
stereoscopic processing. BioRxiv.
Zachariou, v., Nikas, C., Safiullah, z. NORTE., Gotts, S. J., & Ungerleider,
l. GRAMO. (2016). Spatial mechanisms within the dorsal visual path-
way contribute to the configural processing of faces. Cerebral
Corteza, 27, 4124–4138. https://doi.org/10.1093/cercor/bhw224
MENTE ABIERTA: Descubrimientos en ciencia cognitiva
56
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
oh
pag
metro
i
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
d
oh
i
/
i
/
.
/
1
0
1
1
6
2
oh
pag
metro
_
a
_
0
0
0
3
3
1
8
6
8
4
1
6
oh
pag
metro
_
a
_
0
0
0
3
3
pag
d
.
/
i
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3