PERSPECTIVA
Expanding connectomics to the laminar level:
A perspective
Ittai Shamir1
and Yaniv Assaf1,2
1Department of Neurobiology, Faculty of Life Sciences, Tel Aviv University, Tel Aviv, Israel
2Sagol School of Neuroscience, Tel Aviv University, Tel Aviv, Israel
Palabras clave: Connectomics, Cortical layers, Computational models, Neuronal structures, Anatomical
mapping, Brain network analysis
un acceso abierto
diario
ABSTRACTO
Despite great progress in uncovering the complex connectivity patterns of the human brain
over the last two decades, the field of connectomics still experiences a bias in its viewpoint of
the cerebral cortex. Due to a lack of information regarding exact end points of fiber tracts
inside cortical gray matter, the cortex is commonly reduced to a single homogenous unit.
Concurrently, substantial developments have been made over the past decade in the use of
relaxometry and particularly inversion recovery imaging for exploring the laminar
microstructure of cortical gray matter. En años recientes, these developments have culminated in
an automated framework for cortical laminar composition analysis and visualization, seguido
by studies of cortical dyslamination in epilepsy patients and age-related differences in laminar
composition in healthy subjects. This perspective summarizes the developments and
remaining challenges of multi-T1 weighted imaging of cortical laminar substructure, el
current limitations in structural connectomics, and the recent progress in integrating these
fields into a new model-based subfield termed ‘laminar connectomics’. In the coming years,
we predict an increased use of similar generalizable, data-driven models in connectomics with
the purpose of integrating multimodal MRI datasets and providing a more nuanced and
detailed characterization of brain connectivity.
INTRODUCCIÓN
Over the last two decades, the field of neuroimaging has made great strides in its exploration of
the intricate patterns of interconnectivity of the human brain, led in part by the Human
Connectome Project (Setsompop et al., 2013; Sporns et al., 2005; Van Essen et al., 2013).
Structural connectomics using diffusion MRI (ver figura 1) has experienced significant devel-
opments in recent years (p.ej., Assaf et al., 2019; Maier-Hein et al., 2017). Estos desarrollos
include investigation of the economy of connectivity of the human brain (bassett & bullmore,
2006, 2016; bullmore & despreciar, 2012), exploration of its rich-club organization properties
( van den heuvel & despreciar, 2011), and identification of various central network hubs in the
cerebro humano (Sporns et al., 2007; van den heuvel & despreciar, 2013). Recent studies have also
explored intrahemispheric and interhemispheric wiring patterns in the human brain (Krupnik
et al., 2021), and even patterns of wiring conservation across a variety of mammalian species
(Assaf et al., 2020).
Citación: Shamir, I., & Assaf, Y. (2023).
Expanding connectomics to the
laminar level: A perspective. Red
Neurociencia, 7(2), 377–388. https://doi
.org/10.1162/netn_a_00304
DOI:
https://doi.org/10.1162/netn_a_00304
Recibió: 17 Agosto 2022
Aceptado: 15 December 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Ittai Shamir
ittaisha@mail.tau.ac.il
Editor de manejo:
Álex Amueblado
Derechos de autor: © 2023
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Expanding connectomics to the laminar level: A perspective
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
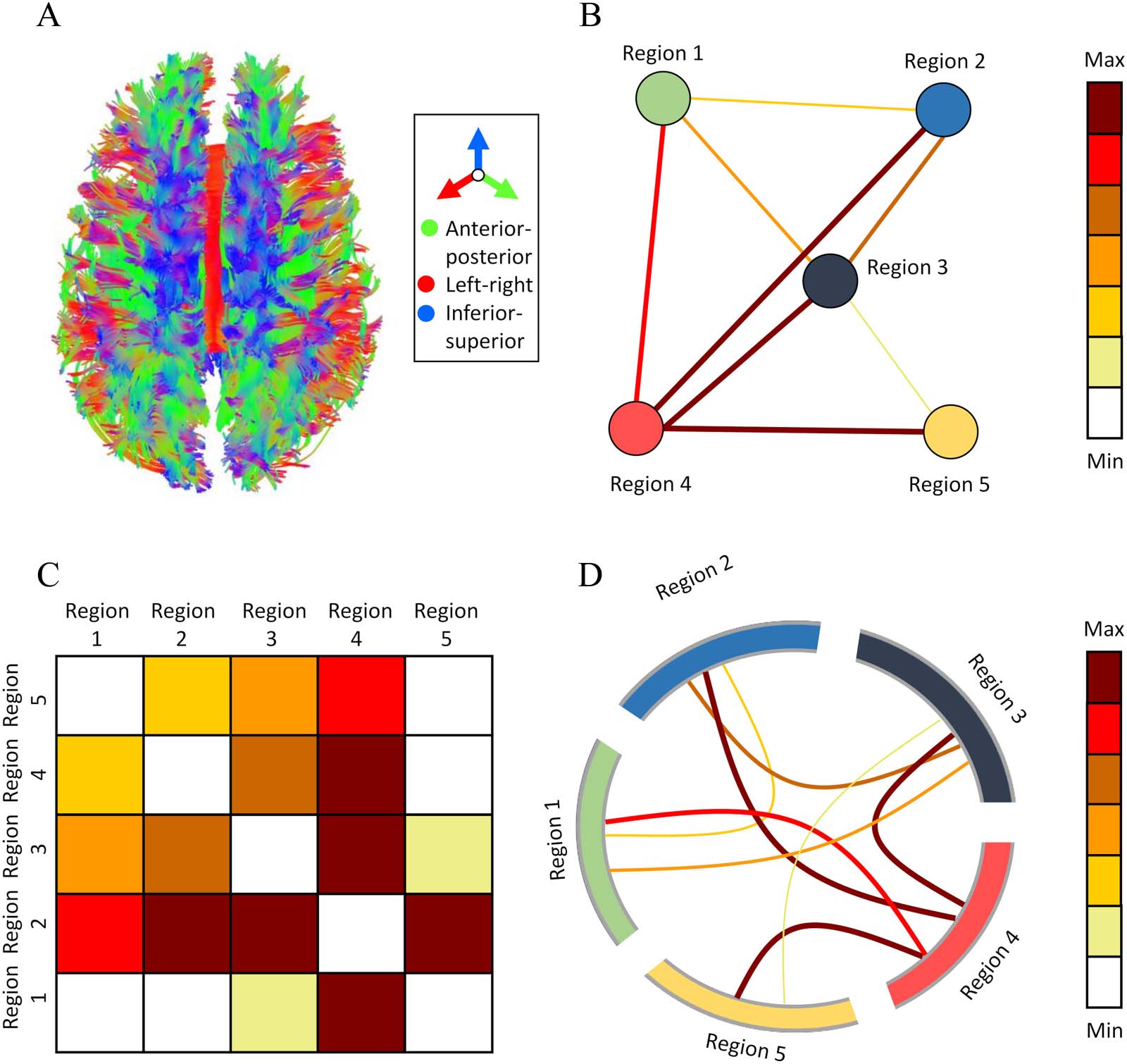
Cifra 1. Various representations of structural connectomics. (A) Tractogram: a three-dimensional streamline diagram of fiber tract connec-
tions across the entire brain (color coded according to directions, seen from a top view). (B) Network graph: a graph representation consisting
de nodos, signifying cortical regions (individually colored), and edges, signifying the strength of connections (colored according to heat map).
(C) Connectivity matrix: a matrix representation, where value {i, j} signifies the strength of connections between cortical regions i and j (colored
according to heat map). (D) Circular graph: a graph representation, where cortical regions are located across the circle’s circumference (indi-
vidually colored), and the chords across its center signify the strength of connections (colored according to heat map).
Human Connectome Project:
A project sponsored by the National
Institutes of Health (NIH), intended to
map the connections of the healthy
cerebro humano.
Connectomics:
A subfield of neuroscience focused
on exploring the map of connections
in the nervous system.
Diffusion MRI:
A magnetic resonance imaging (resonancia magnética)
method for measuring the
displacement, or diffusion, of water
molecules in various tissues.
Neurociencia en red
CURRENT LIMITATIONS OF STRUCTURAL CONNECTOMICS
The field of structural connectomics still experiences conceptual and methodological draw-
backs related to diffusion MRI techniques. Some of the main drawbacks on the conceptual
side involve the fact that the dMRI measures displacement of water molecules, and not their
diffusion directly. Además, the measured displacement of water molecules does not
always follow a Gaussian law, as previously assumed, since it is sometimes restricted in bio-
logical tissues.
If we focus on fiber tractography (FT), many methodological and technical pitfalls appear.
Deterministic streamline tractography, the most widespread method for tractography, usos
dMRI to compute the orientation density function (ODF) and then reduces it to vectors repre-
senting the strongest and most likely directional connections. The resulting model of the brain’s
fiber tracts, also known as a tractogram, includes all the three-dimensional streamlines across the
entire brain. Several studies have detailed the limitations relating to the inference of structural
378
Expanding connectomics to the laminar level: A perspective
Rich-club organization:
A network phenomenon that
involves the tendency of high-degree
nodes to also be highly
interconnected.
Network hubs:
Nodes that occupy central positions
in the general organization of a
network.
Intrahemispheric:
Connections within a single
hemisferio (left or right) of the brain.
Interhemispheric:
Connections between the two
hemispheres (left and right) del
cerebro.
Orientation density function (ODF):
A function used for describing the
directionality of complex fiber
architecture in the nervous system
based on diffusivity.
Crossing fibers:
A common problem in tractography
involving multiple fiber bundles with
significantly varying orientations.
connectivity from local field potentials (Bastiani et al., 2012; Calamante, 2019; De Santis et al.,
2014; Jeurissen et al., 2019; Jones et al., 2013), culminating in a recent international tractography
challenge (Maier-Hein et al., 2017). The most prevalent limitations include the following:
(cid:129) Strong effect of methodology: choice of methodology, including tractography algorithm,
had a strong effect on resulting accuracy levels, revealing a tradeoff between sensitivity
and specificity.
(cid:129) High false positive rates: even the use of high-resolution images in FT still resulted in trac-
tograms containing more invalid than valid (illogical and/or disconnected) fiber bundles.
(cid:129) Errors in reconstructing strong tracts: many algorithms experienced high false positive
rates when reconstructing strong tracts, due to increasingly high certainty assigned to
strong local field potentials.
(cid:129) Difficulty reconstructing small tracts: many FT algorithms also experienced high false
negative rates for small tract with diameters of 2 mm or less, such as the anterior com-
missure (California).
(cid:129) Difficulty reconstructing complex geometry: the intricate geometry of many fiber junc-
tions posed a source of many artifacts and miscalculations in FT (crossing fibers within a
single voxel).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Por último, one of the biggest methodological limitations in tractography and structural connec-
tomics in general is the difficulty in estimating the exact terminations of fiber tracts inside
cortical gray matter. The cortical termination bias involves difficulties in estimating tract termi-
nations not only laterally across the cortical folding (gyral bias), but also radially across the
cortical layers. The main causes for this significant limitation are the partial volume effect asso-
ciated with voxel-wise FT approaches, coupled with the difficulties in estimating small and
complex fiber tracts.
Over the years many advanced techniques for tractography have been proposed for tack-
ling these limitations in structural connectomics. Some of these advancements include tech-
niques for resolving high false positive connections using filtering, replacing manual filtering
based on prior knowledge with thresholding of connectivity matrices or with automated
knowledge and data-driven algorithms (Zhang et al., 2022). Other advancements have
focused on techniques for either handling strong tracts by increasing specificity, or for handling
small tracts by increasing sensitivity (Zhang et al., 2022). The gyral bias in estimating fiber tract
terminations is commonly addressed using targeted tracking, which defines regions for inclu-
sion and exclusion based on prior anatomical constraints ( Yeh et al., 2021).
It should be noted that despite the many technical developments in the field, there still lacks
a single ‘gold standard’ tractography methodology. Además, the estimation of exact termi-
nations of fiber tracts in cortical gray matter remains a significant difficulty that has resulted in
a biased representation of the cerebral cortex. This representation falsely considers the cortex a
single homogenous unit and ignores its microstructural laminar composition.
HISTOLOGICAL EXPLORATIONS OF LAMINAR CONNECTIVITY
Following the partitioning of the cortex into cortical layers at the beginning of the 20th century,
the cortical column was introduced as a single unit that spans across all six layers and repeats
across the brain (Hubel & Wiesel, 1959, 1962; Lorente de Nó, 1949; Mountcastle, 1957). El
concept of the cortical column stemmed from Hubel and Wiesel’s exploration of the mamma-
lian visual cortex, which resulted in the discovery of ocular dominance columns (Hubel &
Neurociencia en red
379
Expanding connectomics to the laminar level: A perspective
Wiesel, 1969). After the cortical column, the canonical microcircuit was introduced as a con-
nectivity unit across and between the cortical layers (Gilbert & Wiesel, 1983; Hellweg et al.,
1977; Szentágothai, 1975). The term ‘canonical microcircuit’ was coined by Douglas et al.
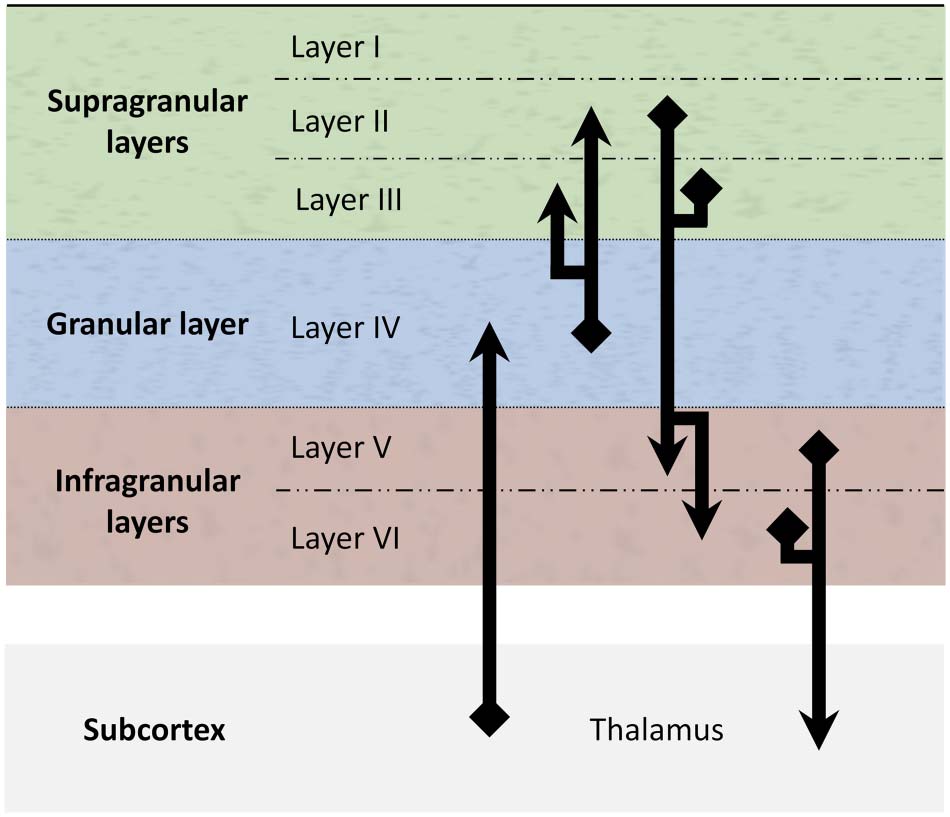
(1989) and it entails the following basic scheme (Dhruv, 2015): thalamic input to granular
layer IV, layer IV projections to layers II and III, projections from layers II and III to layers V
and VI, and finally projections from layers V and VI back to the thalamus (ver figura 2).
Since then, a variety of methods have been applied outside the field of neuroimaging in the
exploration of connectivity patterns of the brain on the cortical layer level, including histolog-
ical staining, tract tracing (retrograde and anterograde), electrical recordings, y otros. Mayoría
of these explorations used animal models, ranging from cats (Douglas et al., 1989; Hubel &
Wiesel, 1962) to nonhuman primates such as the rhesus macaque (Felleman & VanEssen,
1991; Mountcastle, 1997; Rockland & Pandya, 1979). Most of the studies of layer connectivity
have focused on specific cortical regions, primarily the visual cortex or other regions such as
the somatosensory, motor, or auditory cortices (Shamir & Assaf, 2021).
This approach to cortical layer connectivity has since been reexamined and the inherently
interlinked concepts of the cortical column and the cortical microcircuit have been questioned
(Feldmeyer, 2012). What followed were studies that explored layer connections using a
broader approach, with one such repeatedly reported model offering rules of interregional cor-
tical layer connections based on the granularity indices of the connecting regions (Barbas,
1986; Barbas & Rempel-Clower, 1997; Beul & Hilgetag, 2015; Shipp, 2005; von Economo,
2009). Broader, integrative studies have also been performed on hundreds of published studies
on layer connectivity in specific regions of the primate cortex (Schmidt et al., 2018; Solari &
Stoner, 2011), and even comparing layer connectivity between different regions and different
species (DeFelipe et al., 2002).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. A schematic representation of the canonical microcircuit: The three laminar components
(supragranular, granular, and infragranular), consisting of six cortical layers (I–VI), are intercon-
nected, and connected to and from the thalamus (in the subcortex) according to this detailed
connection scheme. It is worth noting that this is a simplified representation of the canonical micro-
circuito. This version of the microcircuit omits cortico-cortical connections of all kinds, incluido
feedforward, comentario, and lateral connections. It is also a partial representation of intracolumnar
connections that excludes some connections that are often added, such as a recurrent connection
from layers V/ VI to layer IV.
Neurociencia en red
380
Expanding connectomics to the laminar level: A perspective
Myeloarchitecture:
The microscopic study of the
arrangement of nerve fibers based on
their myeline content.
T1-weighted imaging:
A magnetic resonance imaging (resonancia magnética)
pulse sequence that measures the
longitudinal relaxation times, or T1
valores, of various tissues.
T2-weighted imaging:
A magnetic resonance imaging (resonancia magnética)
pulse sequence that measures the
transverse relaxation times, or T2
valores, of various tissues.
T2*-weighted imaging:
A magnetic resonance imaging (resonancia magnética)
pulse sequence that measures the
effective transverse relaxation times,
or T2* values, of various tissues.
RECENT DEVELOPMENTS IN IMAGING THE CORTICAL LAYERS
Since its discovery at the beginning of the 20th century (Garey, 2006), the geometrically com-
plex and highly organized laminar structure of the cerebral cortex has been assumed to play
an integral role in the development and function of the human brain, as well as different
pathologies of the brain (Kiernan & Rajakumar, 2014). Until relatively recently, the intricate
laminar composition of the cerebral cortex has been assumed to be beyond the resolution
capabilities of MRI neuroimaging.
Prior to this progress, it was already established that myelination causes shortening of T1
valores. En 1992, a study of both in vivo and postmortem human brains used high-resolution
T1-weighted images to identify the striate cortex and revealed six laminar clusters within it,
with decreasing T1 values from the outermost to the innermost parts of the cortex (clark
et al., 1992). Como consecuencia, T1-weighted MRI techniques have successfully segmented cortical
gray matter from bordering white matter and cerebrospinal fluid (pescado, 2012; Kiernan &
Rajakumar, 2014; Scholtens et al., 2015), further solidifying the link between T1 values and
myeloarchitecture ( Van Essen et al., 2019).
Since then, studies have offered a variety of MRI approaches for exploring cortical gray
matter on the laminar level, including T1-, T2-, and T2*-weighted images, as well as R1,
R2, and R2* susceptibility images. Barbier et al. (2002) used high-resolution T1-weighted
imaging at 3T to delineate the striate cortex based on myelin content. Bridge and Clare
(2006) showed a correspondence between delineations of the primary visual cortex by both
high-resolution MRI and fMRI. Duyn et al. (2007) used high field at 7T to explore cortical sub-
structure based on signal phase. Deistung et al. (2013) compared ultraresolution quantitative
susceptibility maps with conventional gradient echo imaging techniques, including magni-
tude, phase, and R2* imaging. Lutti et al. (2014) used high-resolution quantitative mapping
of R1 as a measure of cortical myelination. Glasser et al. (2014) reviewed a variety of methods
for examining cortical myeline content, including T1 imaging, T2 imaging and even positron
emission tomography (PET). Shafee et al. (2015) used the ratio of T1-weighted to T2-wieghted
images to explore gray matter lamination. A recent study even explored diffusion MRI as a
potential method for imaging the laminar substructure of cortical gray matter (Assaf, 2019).
Of the modalities examined, T1, T2, and T2* imaging initially gained the most attention. Desde
entonces, the biggest challenge has been finding the most robust and precise modality that does
not involve noisy acquisition, as is the case in T2* imaging, or long scan times and a pro-
nounced partial volume effect, as is the case in T2 imaging.
The T1-weighted approach has proved most suitable so far, thanks to findings from a series
of follow-up studies over the past decade. En 2012, a study used T1-weighted imaging to char-
acterize the cortical layers in vivo across the entire brains of both human subjects and rat
modelos (Barazany & Assaf, 2012). The study reestablished the correspondence between T1
clusters and the cortical layers by comparing the T1 clusters to histological findings in the
rat brains. This study was followed by a larger scale study of both rat and human brains, usando
the same inversion recovery (IR) MRI protocol and a higher resolution version of the same
protocol (Lifshits et al., 2018). The study demonstrated that the sound-to-noise ratio and partial
volume effect involved in imaging layers with dimensions down to tens of microns make it
unlikely to resolve the layers using high-resolution imaging. It concluded that the cortical
layers are better separated using low-resolution multi-T1 mapping (high resolution in the relax-
ation domain), compared to high resolution in the image domain. En 2019, a comprehensive
framework was presented for cortical laminar composition analysis using low-resolution multi-
T1 mapping (Shamir et al., 2019). The study offered a whole-brain automatic methodology for
Neurociencia en red
381
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Expanding connectomics to the laminar level: A perspective
Cytoarchitecture:
The microscopic study of the cellular
estructura, or architecture, of various
tissues in the nervous system.
analyzing and visualizing the laminar substructure of the cortex. The study tackled the partial
volume effect associated with lower resolution images and implemented a spherical sampling
system as a rotationally invariant alternative to cortical normals, which addressed the issue of
normal miscalculations due to small inaccuracies in estimating cortical surfaces.
En 2021, a study used the same low-resolution multi-T1 weighted protocol to explore pat-
terns of cortical dyslamination in epilepsy patients (Lotan et al., 2021). The study focused on
epileptic patients with both focal cortical dysplasia (FCD) and periventricular nodular hetero-
topia (PNH), successfully revealing T1-layer specific cortical laminar abnormalities associated
with the pathology. En 2022, a follow-up study used the same methodology to explore
age-related differences across a group of 200 healthy subjects (Tomer et al., 2022). El estudio
reestablished the validity of the framework for cortical laminar composition analysis and
demonstrated its ability to capture differences in compositions across subject groups and brain
regiones.
It should be noted that despite the significant progress and success of the low-resolution
multi-T1 approach for laminar composition analysis, it does have certain limitations. The first
and most notable limitation is that T1 relaxation is still not considered a direct measure of
cytoarchitecture, and as a result the approach is considered an indirect way of imaging and
measuring the cortical layers. Sin embargo, since T1 is considered a measure of myeloarchitecture
(myeline content) and its correspondence with the cortical layers has been established
(Barazany & Assaf, 2012; Lifshits et al., 2018; Lotan et al., 2021; Shamir et al., 2019; Tomer
et al., 2022), the term ‘T1 layers’ is used instead in this context. An additional limitation relates
to high-resolution variations in laminar composition within smaller examined cortical regions
and within the cortical folding, which are restricted in this approach due to its low-resolution
naturaleza. Por último, because this methodology is relatively novel and new, it has yet to be imple-
mented and explored in large-scale populations studies (upwards of 1,000 subjects).
Sin embargo, it is our assertion that the T1 layer methodology can be solidified as a frame-
work for direct measurement of cortical cytoarchitecture if a specific algorithmic challenge is
overcome. This challenge involves spatial clustering of the resulting multidimensional, surface
based T1 layer composition on a whole-brain scale and across a group of subjects. If success-
lleno, the T1 clusters will show both visual and quantitative correspondence with established
cytoarchitectonic atlases and highlight the role of T1 imaging as a direct probe of tissue
microstructure.
INTEGRATING CORTICAL COMPOSITION INTO STRUCTURAL CONNECTOMICS
As the field of connectomics (and structural connectomics in particular) advanced, the need
grew for a unified view of connectivity from the macroscale to the microscale (Rockland,
2019). Several studies have pointed to the potential of integrating microstructural information
regarding cortical laminar composition with macrostructural information regarding cortical
conectividad ( Jbabdi & Johansen-Berg, 2011; Johansen-Berg, 2013). Después, the exten-
sive progress of the past two decades in the field of structural connectomics, coupled with the
developments of recent years in imaging the cortical laminar substructure, have opened the
door to new avenues in connectomics.
En 2021, a study addressed this subject by presenting an MRI-based, data-derived model of
cortical laminar connectivity that offers a way to overcome the biased representation of the
cerebral cortex as a single homogenous unit (Shamir & Assaf, 2021). The study addressed the
lack of information about exact terminations of fiber tracts inside the cortex by systematically
Neurociencia en red
382
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Expanding connectomics to the laminar level: A perspective
reviewing 51 previously published histological studies that focused on patterns of brain con-
nectivity on the cortical laminar level. A simplified model was then formed (Shamir & Assaf,
2021), consisting of a set of data-derived rules for expanding standard connectomics to the
laminar level using the laminar composition of the cortex. The model consists of two principal
types of connections: horizontal and radial connections. Horizontal connections include inter-
regional connections within and between the hemispheres. Radial connections include intrar-
egional connections within the cortical microcircuit as well as connections to the subcortex.
All connections are expansions of findings from standard connectomics, except for connec-
tions within the microcircuit, which are assumed connections. Model rules are applied
according to the granularity indices of the connecting regions and weighted according to their
laminar compositions (ver figura 3).
Multimodal MRI:
A combination of different magnetic
resonance imaging (resonancia magnética) sequences,
or modalities.
A follow-up validation study was then performed, implementing the same model on mul-
timodal MRI datasets of the macaque brain (Shamir & Assaf, 2022). The macaque was chosen
specifically with the intent to compare the resulting model of cortical laminar connectivity to
the connectivity patterns established through a tract tracing study of the macaque visual cortex
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
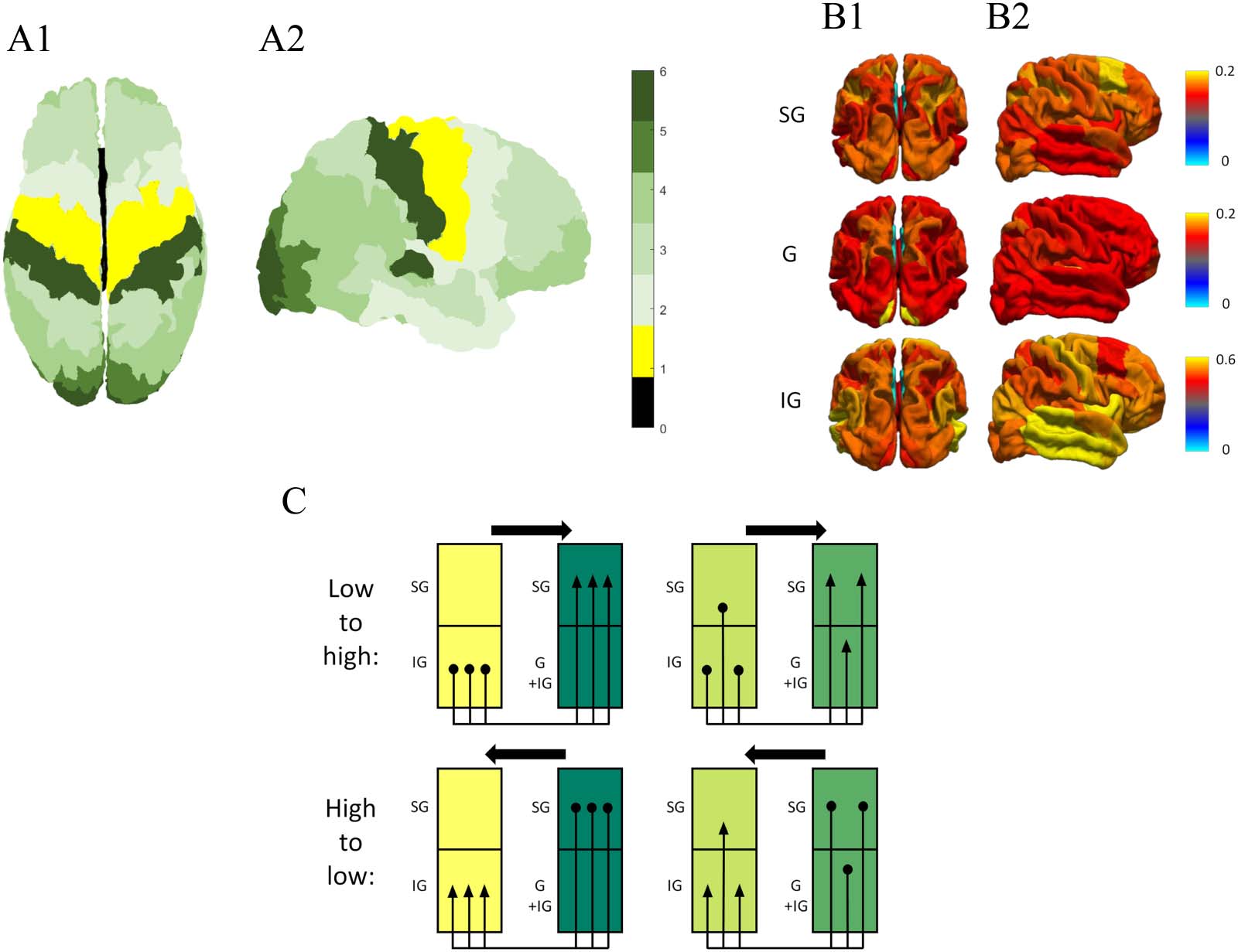
Cifra 3. Components of the model of cortical laminar connectivity. The MRI-based, data-driven model formed by Shamir and Assaf (2021)
offers a way to expand the structural connectome to the laminar level using the following components. (A) Granularity atlas: a cytoarchitec-
tonic atlas that labels cortical regions according to the level of cellular granularity observed histologically, as reported by von Economo (2009)
and further discussed by Beul and Hilgetag (2015) and by Scholtens et al. (2016). The indices: 0, allocortex; 1, agranular; 2, slightly granular; 3,
4, 5, increasingly granular; 6, granular. The atlas is seen from a top view (A1) and from a lateral right view (A2). (B) Laminar composition: a
summary of laminar components across the cortex, based on data from Scholtens et al. (2016). The components: SG, supragranular component
(layers I, II, and III); GRAMO, granular component (layer IV); IG, infragranular component (layers V and VI). The compositions are seen from a top
vista (B1) and from a lateral right view (B2). (C) Model rules of laminar connectivity: a schematic representation of the rules of horizontal
conectividad, based on the granularity indices of the connecting regions as well as their laminar composition (Shamir & Assaf, 2021). adi-
tional rules, regarding radial connectivity, are derived from canonical microcircuit (as seen in Figure 2).
Neurociencia en red
383
Expanding connectomics to the laminar level: A perspective
(Felleman & VanEssen, 1991). The study further validated the model of cortical laminar con-
nectivity by reporting an accuracy level 83%, surprisingly high considering the methodolog-
ical disparities between the two studies (and the 30-year gap).
En 2022, following the formation and corroboration of this model of laminar connectivity, a
study modeled the laminar connectome of 30 healthy human subjects (Shamir et al., 2022).
Unlike the case of the macaque visual cortex, no single gold standard of laminar connectivity
exists for the human brain. Sin embargo, the study examined the resulting networks of laminar
connectivity and found several expected points of reference, including high centrality of gran-
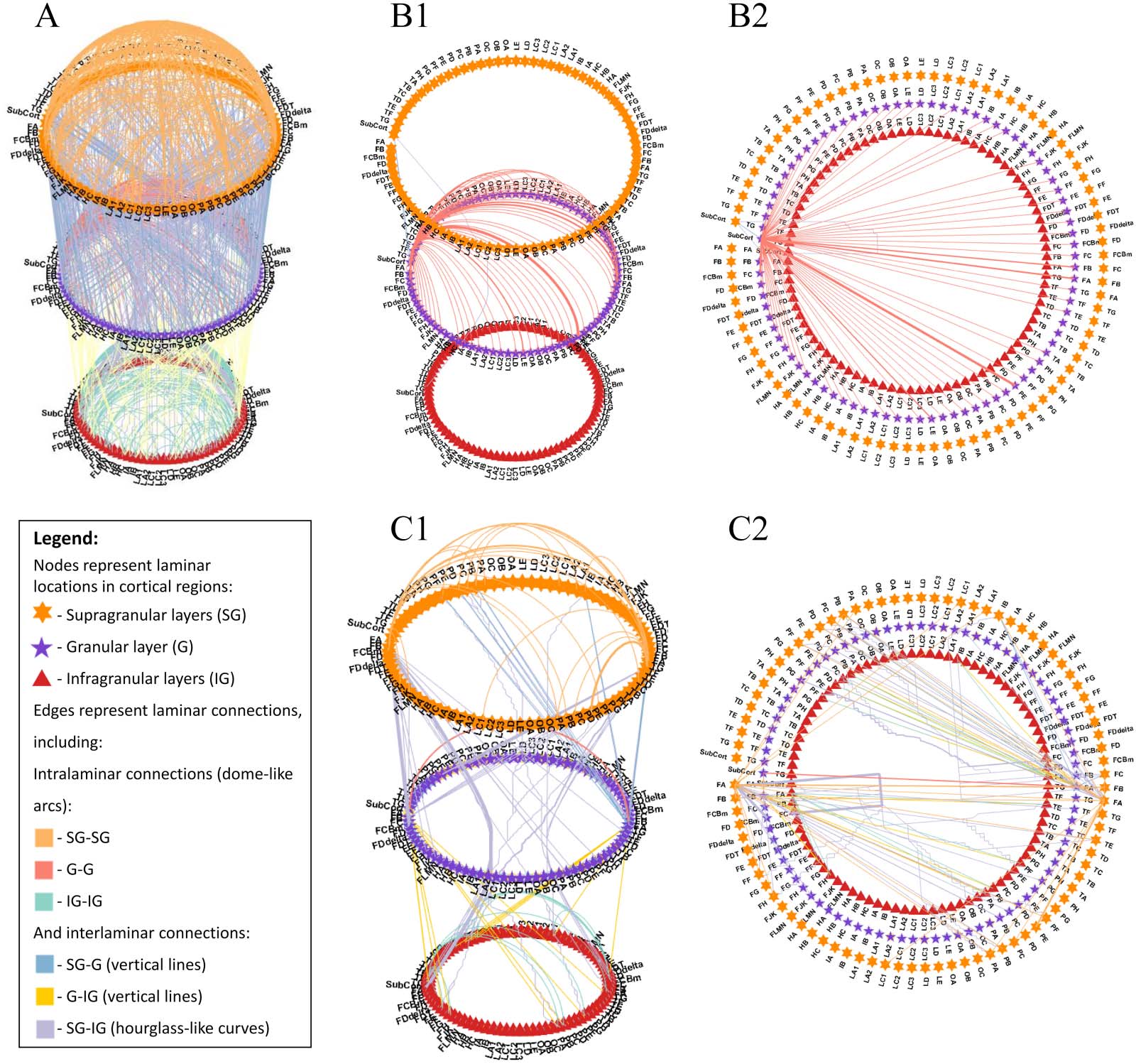
ular connections (involving T1 layer IV) to visual, motor, and auditory regions (ver figura 4).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4. A multilayered, multidimensional circular network graph representing the laminar connectome of a healthy human subject across
von Economo-Koskinas atlas regions. (A) All connections of the cortical laminar connectome, exhibiting the highly interconnected and com-
plex nature of this model of laminar connections. (B) Laminar connections that include the subcortex from a side view (B1) and from a top view
(B2), presenting the dominance of the granular layer in connections that include the subcortex (SubCort) and its high connectivity across all
regiones. (C) Laminar connections that include the primary motor cortex (M1) from a side view (C1) and from a top view (C2), presenting
intricate patterns of laminar connections that include the precentral gyrus (FA) that roughly follow the canonical microcircuit: thalamic input
to the granular layer, followed by projections to supragranular layers, which project deeper to infragranular layers, and then back to the thal-
amus. Connections are colored according to connection type, based on the connecting laminar groups, dónde: supragranular layers include T1
layers 1–3 (top circle-orange pentagons), granular layer includes layer 4 (mid circle-purple stars), and infragranular layers include layers 5 y
6 (bottom circle-red triangles). Data taken from Shamir and Assaf (2021), freely available at https://github.com/ittais/Circular-Connectome,
using the modeling functions from Shamir and Assaf (2022), freely available at github.com/ittais/Laminar_Connectivity.
Neurociencia en red
384
Expanding connectomics to the laminar level: A perspective
It should be noted that the model of cortical laminar connectivity presented in these studies
(Shamir & Assaf, 2021, 2022; Shamir et al., 2022) has certain limitations. The first major
limitation is that the model is a simplified model, which reduces cortical components, incluir-
ing the reduction of six T1 layers to three laminar components, with the purpose of increasing
the dimensionality of the connectome. The second major limitation is that the model does not
estimate the probability of these laminar-level connections, since they are applied via a set of
predefined rules of connectivity.
Taking these limitations into account, alongside the sanity checks found in the visual,
motor, and auditory cortices, the model offers a more nuanced whole-brain look at patterns
of connectivity on the laminar level. This characterization of the healthy human laminar con-
nectome could provide a new level of detail in the examination of different groups of interest
that are presumed to involve distinct features of layer-dependent connectivity.
SUMMARY
Over the past 20 años, the field of neuroimaging has experienced great advancements in
imaging both white matter connectomics and more recently also in imaging the laminar com-
position of gray matter. The progress in imaging gray matter has amplified the demand for a
less biased representation of the cerebral cortex in connectomics that addresses the lack of
information regarding the exact terminations of fiber tracks inside the cortical foldings.
To address this issue and overcome this bias, knowledge-based models must be used. El
MRI-based, data-driven model discussed here (Shamir & Assaf, 2021, 2022; Shamir et al.,
2022) offers one such model by using previously published histological findings to form a sim-
plified model of cortical laminar connectivity. Models of this kind open the door to a new
subfield of structural connectomics termed laminar connectomics. Actualmente, the field of con-
nectomics includes three intertwining subfields, each focused on a different type of brain con-
conectividad: structural, functional, and effective (Parque & Friston, 2013). Effective connectomics
uses the anatomical constraints of structural connectivity in dynamic causal models to explain
the functional signal and explore patterns of activation between different cortical regions
(Friston et al., 2003, 2019). En otras palabras, the novel subfield of laminar connectomics is a
data-driven enhancement of the resolution of anatomical connections, in the same sense that
effective connectomics is an expansion of functional connectomics.
While the model discussed here has its own set of drawbacks and limitations, it offers a
straightforward approach to integrating multimodal MRI datasets into a whole-brain view of
connectivity on the laminar level. Además, this model showcases the benefits of using data-
driven models to explore structural connectomics in a more unbiased and nuanced approach,
which considers the laminar structure of the cortex. We believe that the use of such novel
models will play an important role in structural connectomics even if higher resolution imag-
ing of connectivity is achieved, on the condition that the models are knowledge-based, appli-
cable, and generalizable to any existing or future structural connectivity methodology. El
resulting modeled multilayered connectomes will necessitate the use and development of
advanced network analysis tools that can visualize and explore multidimensional networks
(such as muxViz, De Domenico et al., 2015).
The potential of this expansion of structural connectomics is wide: now that a basic char-
acterization of the healthy human laminar connectome has been achieved, this detailed model
of laminar-level connections can be used to explore different pathologies. This framework, o
others like it, can be implemented to explore pathologies that are assumed to entail
Neurociencia en red
385
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Expanding connectomics to the laminar level: A perspective
abnormalities in laminar-level structure and connections, such as autism spectrum disorder
(ASD) or schizophrenia. This framework can also be used to enhance the exploration of the
mechanisms behind different motor, auditory, or visual skills. The use of multidimensional
complex network analysis tools could shed new light on the connectivity patterns and mech-
anisms that make these groups distinct on the laminar level.
CONTRIBUCIONES DE AUTOR
Ittai Shamir: Conceptualización; Curación de datos; Análisis formal; Investigación; Metodología;
Administración de proyecto; Software; Supervisión; Validación; Visualización; Writing – original
borrador; Escritura – revisión & edición. Yaniv Assaf: Conceptualización; Curación de datos; Fondos
adquisición; Investigación; Metodología; Administración de proyecto; Recursos; Supervisión; Vali-
dación; Escritura – revisión & edición.
REFERENCIAS
Assaf, Y. (2019). Imaging laminar structures in the gray matter with
diffusion MRI. NeuroImagen, 197, 677–688. https://doi.org/10
.1016/j.neuroimage.2017.12.096, PubMed: 29309898
Assaf, y., Bouznach, A., Zomet, o., Marom, A., & Yovel, Y. (2020).
Conservation of brain connectivity and wiring across the mam-
malian class. Neurociencia de la naturaleza, 23(7), 805–808. https://doi
.org/10.1038/s41593-020-0641-7, PubMed: 32514137
Assaf, y., Johansen-Berg, h., & Thiebaut de Schotten, METRO. (2019).
The role of diffusion MRI in neuroscience. RMN en biomedicina,
32(4), e3762. https://doi.org/10.1002/nbm.3762, PubMed:
28696013
Barazany, D., & Assaf, Y. (2012). Visualization of cortical lamina-
tion patterns with magnetic resonance imaging. Corteza cerebral,
22(9), 2016–2023. https://doi.org/10.1093/cercor/ bhr277,
PubMed: 21983231
Barbas, h. (1986). Pattern in the laminar origin of corticocortical
connections. Journal of Computational Neurology, 252(3),
415–422. https://doi.org/10.1002/cne.902520310, PubMed:
3793985
Barbas, h., & Rempel-Clower, norte. (1997). Cortical structure predicts
the pattern of corticocortical connections. Corteza cerebral, 7(7),
635–46. https://doi.org/10.1093/cercor/7.7.635, PubMed:
9373019
Barbier, mi. l., Marrett, S., Danek, A., Vortmeyer, A., van Gelderen,
PAG., duyn, J., Bandettini, PAG., Grafman, J., & Koretsky, A. PAG. (2002).
Imaging cortical anatomy by high-resolution MR at 3.0T: Detec-
tion of the stripe of gennari in visual area 17. Magnetic Reso-
nance in Medicine, 48(4), 735–738. https://doi.org/10.1002
/mrm.10255, PubMed: 12353293
bassett, D. S., & bullmore, mi. (2006). Small-world brain networks.
neurocientífico, 12(6), 512–523. https://doi.org/10.1177
/1073858406293182, PubMed: 17079517
bassett, D. S., & bullmore, mi. (2016). Small-world brain networks
revisited. neurocientífico, 23(5), 499–516. https://doi.org/10
.1177/1073858416667720, PubMed: 27655008
Bastiani, METRO., Shah, norte. J., Goebel, r., & Roebroeck, A. (2012).
Human cortical connectome reconstruction from diffusion
weighted MRI: The effect of tractography algorithm. NeuroImagen,
62(3), 1732–1749. https://doi.org/10.1016/j.neuroimage.2012
.06.002, PubMed: 22699045
Beul, S. F., & Hilgetag, C. C. (2015). Towards a “canonical”
agranular cortical microcircuit. Frontiers in Neuroanatomy, 8,
165. https://doi.org/10.3389/fnana.2014.00165, PubMed:
25642171
Bridge, h., & Clare, S. (2006). High-resolution MRI: En vivo
histology? Philosophical Transactions of the Royal Society,
361(1465), 137–146. https://doi.org/10.1098/rstb.2005.1777,
PubMed: 16553313
bullmore, MI., & despreciar, oh. (2012). The economy of brain network
organización. Naturaleza Reseñas Neurociencia, 13(5), 336–349.
https://doi.org/10.1038/nrn3214, PubMed: 22498897
Calamante, F. (2019). The seven deadly sins of measuring brain
structural connectivity using diffusion MRI streamlines
fibre-tracking. Diagnostics, 9(3), 115. https://doi.org/10.3390
/diagnostics9030115, PubMed: 31500098
clark, V. PAG., Courchesne, MI., & Grafe, METRO. (1992). In vivo mye-
loarchitectonic analysis of human striate and extrastriate
cortex using magnetic resonance imaging. Corteza cerebral, 2(5),
417–424. https://doi.org/10.1093/cercor/2.5.417, PubMed:
1422094
De Domenico, METRO., Portero, METRO. A., & Arenas, A. (2015). MuxViz: A
tool for multilayer analysis and visualization of networks. Diario
of Complex Networks, 3(2), 159–176. https://doi.org/10.1093
/comnet/cnu038
De Santis, S., Drakesmith, METRO., Bells, S., Assaf, y., & jones, D. k.
(2014). Why diffusion tensor MRI does well only some of the
tiempo: variance and covariance of white matter tissue microstruc-
ture attributes in the living human brain. NeuroImagen, 89(100),
35–44. https://doi.org/10.1016/j.neuroimage.2013.12.003,
PubMed: 24342225
DeFelipe, J., Alonso-Nanclares, l., & Arellano, j. I. (2002). Micro-
structure of the neocortex: Comparative aspects. Diario de
Neurocytology, 31(3–5), 299–316. https://doi.org/10.1023
/A:1024130211265, PubMed: 12815249
Deistung, A., Schäfer, A., Schweser, F., Biedermann, Ud., Tornero, r.,
& Reichenback, j. R. (2013). Toward in vivo histology: A com-
parison of quantitative susceptibility mapping (QSM) con
magnitude-, phase-, and R2*-imaging at ultra-high magnetic field
strength. NeuroImagen, 65, 299–314. https://doi.org/10.1016/j
.neuroimage.2012.09.055, PubMed: 23036448
Neurociencia en red
386
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Expanding connectomics to the laminar level: A perspective
Dhruv, norte. t. (2015). Rethinking canonical cortical circuits. Naturaleza
Neurociencia, 18(11), 1538. https://doi.org/10.1038/nn1115
-1538, PubMed: 26505564
douglas, R. J., Martín, k. A. C., & Whitteridge, D. (1989). A canonical
microcircuit for neocortex. Computación neuronal, 1(4), 480–488.
https://doi.org/10.1162/neco.1989.1.4.480
duyn, j. h., van Gelderen, PAG., li, t. P., de Zwart, j. A., Koretsky, A. PAG.,
& Fukunaga, METRO. (2007). High-field MRI of brain cortical substruc-
ture based on signal phase. Proceedings of the National Academy
of Sciences, 104(28), 11796–11801. https://doi.org/10.1073/pnas
.0610821104, PubMed: 17586684
Feldmeyer, D. (2012). Excitatory neuronal connectivity in the barrel
corteza. Frontiers in Neuroanatomy, 6, 24. https://doi.org/10.3389
/fnana.2012.00024, PubMed: 22798946
Felleman, D. J., & VanEssen, D. C. (1991). Distributed hierarchical
processing in the primate cerebral cortex. Corteza cerebral, 1,
1–47. https://doi.org/10.1093/cercor/1.1.1-a
pescado, B.
(2012). FreeSurfer. NeuroImagen, 62(2), 774–781.
https://doi.org/10.1016/j.neuroimage.2012.01.021, PubMed:
22248573
Friston, k. J., harrison, l., & Penny, W.. (2003). Dynamic causal
modelling. NeuroImagen, 19(4), 1273–1302. https://doi.org/10
.1016/S1053-8119(03)00202-7, PubMed: 12948688
Friston, k. J., Preller, k. h., Mathys, C., Cagnan, h., Heinzle, J.,
Razi, A., & Zeidman, PAG. (2019). Dynamic causal modelling revis-
ited. NeuroImagen, 199, 730–744. https://doi.org/10.1016/j
.neuroimage.2017.02.045, PubMed: 28219774
Garey, l. j. (2006). Brodmann’s localization in the cerebral cortex.
Dordrecht, Los países bajos: Saltador.
Gilbert, C. D., & Wiesel, t. norte. (1983). Functional organization of
the visual cortex. Progress in Brain Research, 58, 209–218.
https://doi.org/10.1016/S0079-6123(08)60022-9, PubMed:
6138809
vidrio, METRO. F., goyal, METRO. S., Preuss, t. METRO., Raichle, METRO. MI., & Van
Essen, D. C. (2014). Trends and properties of human cerebral
corteza: Correlations with cortical myelin content. NeuroImagen,
93(punto 2), 165–175. https://doi.org/10.1016/j.neuroimage.2013
.03.060, PubMed: 23567887
Hellweg, F. C., Schultz, w., & Creutzfeldt, oh. D. (1977). Extracel-
lular and intracellular recordings from cat’s cortical whisker pro-
jection area: Thalamocortical response transformation. Diario de
Neurofisiología, 40(3), 463–479. https://doi.org/10.1152/jn
.1977.40.3.463, PubMed: 874525
Hubel, D. h., & Wiesel, t. norte. (1959). Receptive fields of single
neurones in the cat’s striate cortex. Revista de fisiología, 148(3),
574–591. https://doi.org/10.1113/jphysiol.1959.sp006308,
PubMed: 14403679
Hubel, D. h., & Wiesel, t. norte. (1962). Receptive fields, binocular
interaction and functional architecture in the cat’s visual cortex.
Revista de fisiología, 160(1), 106–154. https://doi.org/10.1113
/jphysiol.1962.sp006837, PubMed: 14449617
Hubel, D. h., & Wiesel, t. norte. (1969). Visual area of the lateral
suprasylvian gyrus (Clare-Bishop area) of the cat. Journal of Phys-
iology, 202(1), 251–260. https://doi.org/10.1113/jphysiol.1969
.sp008808, PubMed: 5770897
Jbabdi, S., & Johansen-Berg, j. (2011). Tractography: Where do we
go from here? Brain Connectivity, 1(3), 169–183. https://doi.org
/10.1089/brain.2011.0033, PubMed: 22433046
Jeurissen, B., Descoteaux, METRO., Mori, S., & Leemans, A. (2019). Dif-
fusion MRI fiber tractography of the brain. RMN en biomedicina,
32(4), e3785. https://doi.org/10.1002/nbm.3785, PubMed:
28945294
Johansen-Berg, h. (2013). Human connectomics—What will the
future demand? NeuroImagen, 80, 541–544. https://doi.org/10
.1016/j.neuroimage.2013.05.082, PubMed: 23727322
jones, D. K., Knösche, t. r., & Tornero, R. (2013). White matter
integrity, fiber count, and other fallacies: The do’s and don’ts
of diffusion MRI. NeuroImagen, 73, 239–254. https://doi.org/10
.1016/j.neuroimage.2012.06.081, PubMed: 22846632
Kiernan, J., & Rajakumar, norte. (2014). Barr’s the human nervous
sistema: An anatomical viewpoint. Filadelfia, Pensilvania: Lippincott
williams & Wilkins.
Krupnik, r., Yovel, y., & Assaf, Y. (2021). Inner hemispheric and
interhemispheric connectivity balance in the human brain. Jour-
nal of Neuroscience, 41(40), 8351–8361. https://doi.org/10.1523
/jneurosci.1074-21.2021, PubMed: 34465598
Lifshits, S., Tomer, o., Shamir, I., Barazany, D., Tsarfaty, GRAMO., Rosset,
S., & Assaf, Y. (2018). Resolution considerations in imaging of the
cortical layers. NeuroImagen, 164, 112–120. https://doi.org/10
.1016/j.neuroimage.2017.02.086, PubMed: 28274834
Lorente de Nó, R. (1949). Cerebral cortex: Arquitectura, intracorti-
cal connections, motor projections. In J. F. Fulton (Ed.), Physiol-
ogy of the nervous system (páginas. 288–312). Nueva York: Oxford
Prensa universitaria.
Lotan, MI., Tomer, o., Tavor, I., Blatt, I., Goldberg-Stern, h.,
Hoffmann, C., Tsarfaty, GRAMO., Tanne, D., & Assaf, Y. (2021).
Widespread cortical dyslamination in epilepsy patients with
malformations of cortical development. Neuroradiology, 63(2),
225–234. https://doi.org/10.1007/s00234-020-02561-2,
PubMed: 32975591
Lutti, A., Dick, F., Sereno, METRO. I., & Weiskopf, norte. (2014). Usando
high-resolution quantitative mapping of R1 as an index of corti-
cal myelination. NeuroImagen, 93(punto 2), 176–188. https://doi.org
/10.1016/j.neuroimage.2013.06.005, PubMed: 23756203
Maier-Hein, k. h., Neher, PAG. F., Houde, J.-C., et al. (2017). El
challenge of mapping the human connectome based on diffusion
tractography. Comunicaciones de la naturaleza, 8(1), 1349. https://doi.org
/10.1038/s41467-017-01285-x, PubMed: 29116093
Mountcastle, V. B. (1957). Modality and topographic properties of
single neurons of cat’s somatic sensory cortex. Journal of Neuro-
physiology, 20(4), 408–34. https://doi.org/10.1152/jn.1957.20.4
.408, PubMed: 13439410
Mountcastle, V. B. (1997). The columnar organization of the neo-
corteza. Cerebro, 120(punto 4), 701–722. https://doi.org/10.1093/brain
/120.4.701, PubMed: 9153131
Parque, H.-J., & Friston, k. (2013). Structural and functional brain net-
obras: from connections to cognition. Ciencia, 342(6158),
1238411. https://doi.org/10.1126/science.1238411, PubMed:
24179229
Rockland, k. S. (2019). What do we know about laminar connec-
actividad? NeuroImagen, 197, 772–784. https://doi.org/10.1016/j
.neuroimage.2017.07.032, PubMed: 28729159
Rockland, k. S., & Pandya, D. norte. (1979). Distributed hierarchical
processing connections of the occipital lobe in the rhesus mon-
key. Brain Research, 179(1), 3–20. https://doi.org/10.1016/0006
-8993(79)90485-2, PubMed: 116716
Neurociencia en red
387
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Expanding connectomics to the laminar level: A perspective
Schmidt, METRO., Bakker, r., Hilgetag, C. C., Diesmann, METRO., & Van
Albada, S. j. (2018). Multi-scale account of the network structure
of macaque visual cortex. Brain Structure and Function, 223(3),
1409–1435. https://doi.org/10.1007/s00429-017-1554-4,
PubMed: 29143946
Scholtens, l. h., de Reus, METRO. A., el largo, S. C., Schmidt, r., & camioneta
den Heuvel, METRO. PAG. (2016). An MRI Von Economo-Koskinas atlas.
NeuroImagen, 170, 249–256. https://doi.org/10.1016/j
.neuroimage.2016.12.069, PubMed: 28040542
Scholtens, l. h., de Reus, METRO. A., & van den heuvel, METRO. PAG. (2015).
Linking contemporary high resolution magnetic resonance imag-
ing to the Von Economo legacy: A study on the comparison of
MRI cortical thickness and histological measurements of cortical
estructura. Mapeo del cerebro humano, 36(8), 3038–3046. https://doi
.org/10.1002/hbm.22826, PubMed: 25988402
Setsompop, K., Kimmlingen, r., Eberlein, MI., Witzel, T., cohen-
Aded, J., McNab, j. A., Keil, B., Tisdall, METRO. D., Hoecht, PAG., Dietz,
PAG., Cauley, S. F., Tountcheva, v., Matschl, v., Lenz, V. h.,
Heberlein, K., Potthast, A., Thein, h., Van Horn, J., Toga, A.,
Schmitt, F., Lehne, D., rosa, B.R., Wedeen, v., & Wald, l. l.
(2013). Pushing the limits of in vivo diffusion MRI for the Human
Connectome Project. NeuroImagen, 80, 220–233. https://doi.org
/10.1016/j.neuroimage.2013.05.078, PubMed: 23707579
Shafee, r., Buckner, R. l., pescado, B. (2015). Gray matter myelina-
ción de 1555 human brains using partial volume corrected MRI
images. NeuroImagen, 105, 473–485. https://doi.org/10.1016/j
.neuroimage.2014.10.054, PubMed: 25449739
Shamir, I., & Assaf, Y. (2021). An MRI-based, data-driven model of
cortical laminar connectivity. Neuroinformatics, 19(2), 205–218.
https://doi.org/10.1007/s12021-020-09491-7, PubMed:
32949346
Shamir, I., & Assaf, Y. (2022). Modelling cortical laminar connectivity
in the macaque brain. Neuroinformatics, 20(3), 559–573. https://
doi.org/10.1007/s12021-021-09539-2, PubMed: 34392433
Shamir, I., Tomer, o., Baratz, Z., Tsarfaty, GRAMO., Faraggi, METRO.,
Horowitz, A., & Assaf, Y. (2019). A framework for cortical laminar
composition analysis using low-resolution T1 MRI images. Cerebro
Structure and Function, 224(4), 1457–1467. https://doi.org/10
.1007/s00429-019-01848-2, PubMed: 30783759
Shamir, I., Tomer, o., Krupnik, r., & Assaf, Y. (2022). Modelling the
laminar connectome of the human brain. Brain Structure and
Function, 227(6), 2153–2165. https://doi.org/10.1007/s00429
-022-02513-X, PubMed: 35657419
Shipp, S. (2005). The importance of being agranular: a comparative
account of visual and motor cortex. Philosophical Transactions of
the Royal Society, 360(1456), 797–814. https://doi.org/10.1098
/rstb.2005.1630, PubMed: 15937013
Solari, S. V. h., & Stoner, R. (2011). Cognitive consilience: Primate
non-primary neuroanatomical circuits underlying cognition.
Frontiers in Neuroanatomy, 5, 65. https://doi.org/10.3389/fnana
.2011.00065, PubMed: 22194717
despreciar, o., Tononi, GRAMO., & Kotter, R. (2005). The human connec-
a mi: A structural description of the human brain. PLoS Compu-
tational Biology, 1(4), e42. https://doi.org/10.1371/journal.pcbi
.0010042, PubMed: 16201007
despreciar, o., Honey, C. J., & kötter, R. (2007). Identification and
classification of hubs in brain networks. Más uno, 2(10), e1049.
https://doi.org/10.1371/journal.pone.0001049, PubMed:
17940613
Szentágothai, j. (1975). The ‘module-concept’ in cerebral cortex
architecture. Brain Research, 95(2–3), 475–496. https://doi.org
/10.1016/0006-8993(75)90122-5, PubMed: 808252
Tomer, o., Barazany, D., Baratz, Z., Tsarfaty, GRAMO., & Assaf, Y. (2022).
In vivo measurements of lamination patterns in the human cor-
tex. Mapeo del cerebro humano, 43(9), 2861–2868. https://doi.org/10
.1002/hbm.25821, PubMed: 35274794
van den heuvel, METRO. PAG., & despreciar, oh. (2011). Rich-club organization
of the human connectome. Revista de neurociencia, 31(44),
15775–15786. https://doi.org/10.1523/JNEUROSCI.3539-11
.2011, PubMed: 22049421
van den heuvel, METRO. PAG., & despreciar, oh. (2013). Network hubs in the
cerebro humano. Tendencias en Ciencias Cognitivas, 17(12), 683–696.
https://doi.org/10.1016/j.tics.2013.09.012, PubMed: 24231140
VanEssen, D. C., Donahue, C. J., carbonero, t. S., Kennedy, h.,
Hayashi, T., & vidrio, METRO. F. (2019). Cerebral cortical folding,
parcellation, and connectivity in humans, nonhuman primates,
and mice, procedimientos de la Academia Nacional de Ciencias,
11 6 ( 5 2 ) , 2 6 17 3 – 2 6 1 8 0 . h t t p s : / / d o i . o rg / 1 0 . 1 0 7 3 / p n a s
.1902299116, PubMed: 31871175
VanEssen, D. C., Herrero, S. METRO., Respeto, D. METRO., Behrens, t. mi. J.,
Yacoub, MI., & Ugurbil, k. (2013). The WU-Minn Human Con-
nectome Project: An overview. NeuroImagen, 80, 62–79. https://
doi.org/10.1016/j.neuroimage.2013.05.041, PubMed:
23684880
von Economo, C. (2009). Cellular structure of the human cerebral
corteza. (l. C. Triarhou Ed., Trans.). Basel, Suiza: Karger.
https://doi.org/10.1159/isbn.978-3-8055-9062-4
Yeh, C. h., jones, D. K., Liang, X., Descoteaux, METRO., & connelly, A.
(2021). Mapping structural connectivity using diffusion MRI:
Challenges and opportunities. Revista de resonancia magnética
Imaging, 53(6), 1666–1682. https://doi.org/10.1002/jmri.27188,
PubMed: 32557893
zhang, F., Daducci, A., Él, y., Schiavi, S., Seguin, C., Herrero, R. MI.,
Yeh, C. h., zhao, T., & O'Donnell, l. j. (2022). Quantitative
mapping of the brain’s structural connectivity using diffusion
MRI tractography: A review. NeuroImagen, 249, 118870. https://
doi.org/10.1016/j.neuroimage.2021.118870, PubMed:
34979249
Neurociencia en red
388
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
3
7
7
2
1
1
8
4
1
6
norte
mi
norte
_
a
_
0
0
3
0
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3