Object-in-place Memory Predicted by Anterolateral
Entorhinal Cortex and Parahippocampal
Cortex Volume in Older Adults
Lok-Kin Yeung1*, Rosanna K. Olsen1,2, Bryan Hong1, Valentina Mihajlovic1,
Maria C. D’Angelo2, Arber Kacollja2, Jennifer D. Ryan1,2,
and Morgan D. Barense1,2
Abstracto
■ The lateral portion of the entorhinal cortex is one of the first
brain regions affected by tau pathology, an important biomarker
for Alzheimer disease. Improving our understanding of this
region’s cognitive role may help identify better cognitive tests
for early detection of Alzheimer disease. Based on its functional
conexión, we tested the idea that the human anterolateral en-
torhinal cortex (alERC) may play a role in integrating spatial in-
formation into object representations. We recently demonstrated
that the volume of the alERC was related to processing the spatial
relationships of the features within an object [Yeung, l. K.,
Olsen, R. K., Bild-Enkin, h. mi. PAG., D’Angelo, METRO. C., Kacollja, A.,
McQuiggan, D. A., et al. Anterolateral entorhinal cortex volume
predicted by altered intra-item configural processing. Diario de
Neurociencia, 37, 5527–5538, 2017]. en este estudio, we investi-
gated whether the human alERC might also play a role in pro-
cessing the spatial relationships between an object and its
environment using an eye-tracking task that assessed visual fixa-
tions to a critical object within a scene. Guided by rodent work,
we measured both object-in-place memory, the association of an
object with a given context [wilson, D. I., Langston, R. F.,
Schlesiger, METRO. I., Wagner, METRO., Watanabe, S., & Ainge, j. A.
Lateral entorhinal cortex is critical for novel object-context recog-
nición. Hippocampus, 23, 352–366, 2013], and object-trace
memory, the memory for the former location of objects [Tsao,
A., Moser, METRO. B., & Moser, mi. I. Traces of experience in the lateral
entorhinal cortex. Biología actual, 23, 399–405, 2013]. en un
group of older adults with varying stages of brain atrophy and
cognitive decline, we found that the volume of the alERC and
the volume of the parahippocampal cortex selectively predicted
object-in-place memory, but not object-trace memory. Estos
results provide support for the notion that the alERC may inte-
grate spatial information into object representations. ■
INTRODUCCIÓN
Lateral portions of the entorhinal cortex are among the
earliest regions to develop tau pathology, a key bio-
marker for Alzheimer disease (ANUNCIO; Khan et al., 2014;
Braak & Braak, 1991). Sucesivamente, the presence of tau pathol-
ogy here is strongly related to local gray matter loss
(Maass et al., 2017; Sepulcre et al., 2016). Consistent with
these findings, recent work from our group showed
smaller anterolateral entorhinal cortex (alERC) volumes
in ostensibly healthy older adults demonstrating early
signs of preclinical AD-related cognitive decline (Olsen
et al., 2017). Neurodegenerative changes in AD occur
years before cognitive deficits become apparent with stan-
dard neuropsychological assessments (Sperling et al.,
2011). De este modo, finding a subtle cognitive effect specifically
1universidad de toronto, 2Rotman Research Institute, Baycrest
Health Sciences, toronto
*Currently at Taub Institute, Columbia University Medical
Center, Nueva York, Nueva York.
© 2019 Instituto de Tecnología de Massachusetts
related to alERC neurodegeneration could significantly
improve early detection of AD. Sin embargo, these efforts
are limited by a lack of understanding regarding the
cognitive role of human entorhinal cortex subdivisions.
In rodents and nonhuman primates, it is well estab-
lished that distinct subregions of the entorhinal cortex
mediate two input pathways into the hippocampus. Uno
pathway originates in the ventral visual stream and
projects to the lateral entorhinal cortex (LEC) via the peri-
rhinal cortex (PRC; Naber, Caballero-Bleda, Jorritsma-
Byham, & Ingenioso, 1997; suzuki & Amaral, 1994; Ver también
Cowell, Bussey, & Saksida, 2010). The other pathway
originates in the dorsal visual stream and projects to
the medial entorhinal cortex (MEC) via the postrhinal/
parahippocampal cortex (PHC; Moser, Kropff, & Moser,
2008; Burwell, 2000). Far less is known about the function
and organization of these two pathways in humans.
Recent work suggests there exists a similar functional
parcellation of the human entorhinal cortex (Maás,
Berron, Libby, Ranganath, & Düzel, 2015; Navarro
Schröder, Haak, Zaragoza Jimenez, beckman, &
Revista de neurociencia cognitiva 31:5, páginas. 711–729
doi:10.1162/jocn_a_01385
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
oh
C
norte
_
a
_
0
1
3
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Doeller, 2015). Functional connectivity analyses reveal
that the human entorhinal cortex can be divided into
two parts: an anterior-lateral subregion (the alERC) eso
coactivates with the PRC and a posterior-medial sub-
región (the pmERC) that coactivates with the PHC. Este
suggests a functional homology between the human alERC
with the rodent LEC and between the human pmERC
with the rodent MEC.
The PMAT (posterior medial, anterior temporal) modelo
proposes that the anatomical distinction of the two path-
ways underlies a functional distinction as well (Ritchey,
Libby, & Ranganath, 2015). Bajo este modelo, the PRC–
alERC pathway is critical for representing item infor-
formación, and the PHC–pmERC pathway is critical for
representing spatial and contextual information, with both
pathways converging in the hippocampus. Funcional
neuroimaging data in humans support this model, con
greater BOLD activity in LEC when processing the identity
of a face or object and greater BOLD activity in medial
entorhinal cortex (ERC) when processing the spatial loca-
tion of that object (Berron et al., 2018; Reagh et al., 2018;
Reagh & Yassa, 2014; Schultz, verano, & Peters, 2012).
Concurrently, the representational–hierarchical model
proposes a hierarchical organization of stimulus represen-
tations of increasing complexity moving forward in each
pathway (Cowell et al., 2010). The PRC is theorized to
support object-level representations, and the hippocampus
sits even higher in the hierarchy, containing even more
complex conjunctive representations necessary to bind in-
formation across different objects, such as the spatial or
temporal relationships constituting a scene or event. Como
the alERC sits between the PRC and hippocampus, este
model suggests it supports representations more complex
than an object, but less complex than a scene.
In contrast to human fMRI studies emphasizing the
distinctiveness of the two ERC pathways, some rodent
studies have reported that the separation between them
is not absolute. Although the majority of connections
continue along their respective pathways, the rodent
LEC also has some reciprocal connections with the
MEC (van Strien, Cappaert, & Ingenioso, 2009). By analogy,
similar connections between the homologous human
alERC and pmERC might also exist. Based on these recip-
rocal connections between the two pathways, we and
others have speculated that the alERC might play a role
in integrating spatial information from the pmERC into
the object representations supported by the PRC
(Connor & Para un alfiler, 2017; Yeung et al., 2017). Two LEC
rodent studies provide support for this notion. Lesions to
the rodent LEC led to impairments in “object-in-place”
memory (es decir., memory for the association between an
object and a spatial context), but not for memory for ob-
jects or spatial contexts independently ( Wilson et al.,
2013). Además, direct recording of the rodent LEC re-
ported the presence of “object-trace cells”: Place cells
that fired specifically at the locations that had previously
contained a certain object (Tsao, Moser, & Moser, 2013).
In humans, we recently found that alERC volume was
positively related to processing the spatial relationship
of features within an object (es decir., visual fixations to the
configurally relevant region of an object; Yeung et al.,
2017). en este estudio, we sought to investigate whether
the human alERC might also play a broader role in pro-
cessing spatial information about an object, beyond the
within-object processing that we and others have ob-
served (Berron et al., 2018; Reagh et al., 2018; Yeung
et al., 2017; Reagh & Yassa, 2014; Schultz et al., 2012).
En particular, we assessed whether the integrity of the
alERC was related to associating an object with its spatial
contexto, as has been observed in rodents.
Inspired by the rodent work, we leveraged an eye-
tracking-based behavioral paradigm to test whether the
human alERC and surrounding medial-temporal lobe
(MTL) regions might also play a role in object-in-place
memory (es decir., associating an object with a particular loca-
tion in a particular context) and/or object-trace memory
(es decir., memory for the previous location of an object;
Herrero & Squire, 2008; ryan, Leung, Turk-Browne, &
Hasher, 2007; Herrero, Hopkins, & Squire, 2006; ryan &
cohen, 2004a, 2004b; ryan, Althoff, Whitlow, & cohen,
2000). As in our previous work, we used eye-tracking-
based metrics as our outcome measures, as these mea-
sures are sensitive to memory effects, which may not
reach the level of conscious awareness (Ryan et al.,
2000) and allowed us to more closely match our design
to the aforementioned rodent studies. A group of older
adult participants with varying levels of cognitive decline
incidentally viewed computer-generated scenes that were
either entirely novel, repeated identically from the pre-
vious viewing, or were manipulated such that a single
critical object was moved. This allowed us to derive
eye-tracking-based measures of object-in-place and
object-trace memory based on fixations to the location
currently or previously occupied by the object in the
manipulated scenes, respectivamente. The novel scenes were
used to assess global measures of novelty detection.
Además, we employed a recently developed manual
segmentation protocol to assess the volume of the alERC
(Olsen et al., 2017; Maass et al., 2015) and surrounding
hippocampal subfields and MTL cortices (Olsen et al.,
2013). We hypothesized that the volumes of the alERC,
the PHC, the pmERC and the hippocampal subfields,
which belong to the spatial/contextual processing path-
forma, would relate to eye-tracking-based measures of both
object-in-place memory and object-trace memory.
MÉTODOS
Participantes
Thirty-two community-dwelling older adults were re-
cruited from the community in Toronto. Data from two
participants were excluded because of eye tracker failure.
El restante 30 participants had a mean age of 72.3 años
712
Revista de neurociencia cognitiva
Volumen 31, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
oh
C
norte
_
a
_
0
1
3
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(DE = 5.2 años, range = 59–81 years; 23 mujer). Participe-
pants had previously been tested on the Montreal Cognitive
Evaluación (MoCA; Nasreddine et al., 2005) within the last
23 meses (mean = 12.1, DE = 6.8, range = 2–23 months)
and were selected to provide a distribution of MoCA
puntuaciones (mean = 25.3, DE = 3.0, range = 17–30). Dado
that MoCA is sensitive to the presence of mild cognitive
impairment, which is associated with MTL/hippocampus
volume loss (Jack et al., 1997), our intention was to select
for a participant group that had a good distribution of
cognitive abilities and MTL/hippocampal regional vol-
umes. These participants were the subset of an original
sample of 40 participants from Olsen et al. (2017) a quien
we were able to recruit for this study (es decir., eight partici-
pants were lost to follow-up). The original group of 40
participants were chosen such that 20 participants had
scored above the recommended MoCA cutoff score
(≥26) y 20 participants had scored below the MoCA
cutoff score (<25; data on these participants has previ-
ously been reported in Olsen et al. 2017; Yeung et al.,
2017). Of the 30 participants whose data we report here,
14 scored above the MoCA cutoff score, and 16 scored
below it. These two groups were matched for age (partic-
ipants in this study: t(28) = 1.29, p = .21, d = 0.237) and
years of education (participants in this study: t(28) =
0.51, p = .61, d = 0.076). Because of our efforts to match
participants above and below the MoCA cutoff score in
terms of demographic characteristics, MoCA and age were
not correlated among the 30 participants in this study
(r = −.250, p = .13). For the purposes of this study,
we were primarily interested in how MTL volume dif-
ferences related to cognitive performance, rather than
how participants who scored above/below the MoCA
threshold differed; thus, we treated all the participants as
a single group for all subsequent analyses. Participants
received a battery of neuropsychological tests to charac-
terize their cognitive status (Table 1) in an earlier session
(mean interval = 10.2 months, SD = 8.8 months). All
participants had normal or corrected-to-normal vision
(with glasses or bifocals) and were screened for color
blindness, psychological or neurological disorders, brain
damage (i.e., stroke or surgery), and metal implants,
which would have precluded MR imaging. All participants
gave informed consent. This research received ethical ap-
provals from the Research Ethics Boards of the University
of Toronto and Baycrest.
MRI Scan Parameters
High-resolution T2-weighted images were acquired in an
oblique-coronal plane, perpendicular to the long axis of
the hippocampus (echo time/repetition time = 68 msec/
3000 msec, 20–28 slices depending on head size, 512 ×
512 acquisition matrix, voxel size = 0.43 × 0.43 × 3 mm,
no skip, field of view = 220 mm), on a 3T Siemens Trio
scanner at the Rotman Research Institute at Baycrest
(Toronto, ON). The first slice was placed anterior to
the appearance of the collateral sulcus (including the
temporal pole where possible), and the last slice was
placed posterior to the hippocampal tail to ensure full
coverage of the entire hippocampus and all of the MTL
cortices included in the volumetric analyses for all
participants. To confirm slice placement, a T1-weighted
magnetization prepared rapid gradient echo whole-brain
anatomical scan (echo time/repetition time = 2.63 msec/
2000 msec, 176 slices perpendicular to the AC–PC line,
256 × 192 acquisition matrix, voxel size = 1 × 1 × 1 mm,
field of view = 256 mm) was acquired immediately before
the T2-weighted scan. The T1-weighted images were also
used to estimate total intracranial volume for head size
correction (see Volume Correction for Head Size section).
Manual Segmentation
For each participant, L. Y. manually segmented three
hippocampal subfields (CA1, dentate gyrus/CA2&3, and
subiculum) and four MTL cortices (alERC, pmERC, PRC,
and PHC) on coronal slices of the T2-wighted structural
scans (in-plane resolution: 0.43 × 0.43 mm, 3 mm between
slices) using FSLview (v3.1) (Table 2). Manual segmentation
followed the Olsen–Amaral–Palombo protocol (Olsen et al.,
2013; Palombo et al., 2013; see also the Appendix to
Yushkevich, Amaral, et al., 2015) supplemented with a
modified version of the protocol provided by Maass et al.
(2015) for the subdivisions of the entorhinal cortex (see
Figure 1 for a visualization of the segmentation protocol).
Average volumes for each manually segmented brain region
are presented in Table 3, and correlations between brain
region volumes are presented in Table 4.
We considered these particular regions for two rea-
sons. First, because these regions are directly connected
to the alERC (Burwell, 2000; Suzuki & Amaral, 1994), we
wished to explore if any observed alERC–behavior corre-
lations were mediated by its inputs and outputs. Second,
a number of these regions have been shown to be criti-
cally important in aspects of spatial memory, including
object location memory, and scene memory. PHC lesions
have been shown to impair object location memory
(Malkova & Mishkin, 2003; Bohbot et al., 1998), and the
PHC is reliably activated when viewing scenes (Epstein,
Harris, Stanley, & Kanwisher, 1999). The hippocampus
has long been known to have an important role in spa-
tial representation (e.g., O’Keefe & Dostrovsky, 1971)
and is theorized to support flexible representations of
spatial/temporal arrangements of objects (Eichenbaum
& Cohen, 2001) that underlie its role in scene memory
and perception (Lee et al., 2005). However, the ques-
tion of how structural differences in hippocampal sub-
fields might affect these cognitive roles remains to be
answered. The pmERC connects the PHC to the hippo-
campus, and direct recording work here suggests it is im-
portant for representing locations on a screen (Killian,
Jutras, & Buffalo, 2012). The PRC is involved in combining
object features into a conjunctive representation (Barense
Yeung et al.
713
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
o
c
n
_
a
_
0
1
3
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Neuropsychological Battery Results Expressed as Means (SD)
Test
MoCA (/30)
WMS-IV Logical Memory
All Participants (N = 32)
Participants Included
in Data Analysis (n = 30)
25.3 (2.9)
Slightly impaired
25.3 (2.9)
Slightly impaired
Immediate Recall Scaled Score (/20)
Delayed Recall Scaled Score (/20)
11.3 (2.8)
10.8 (2.7)
63.7%ile
59.1%ile
11.4 (2.9)
10.9 (2.8)
64.2%ile
59.3%ile
Recognition Accuracy
78.7% (17.8%)
81.8% (11.1%)
Trails A
Trails B
Digit Span Forward Score (/16)
Digit Span Backward Score (/14)
Rey–Osterrieth Complex Figure
Copy (/32)
Immediate Recall (/32)
Delayed Recall (/32)
Weschler Abbreviated Scale of Intelligence
Vocabulary (/80)
Similarities (/48)
Matrix Reasoning (/32)
Block Design (/71)
Visual Object and Spatial Perception Battery
Shape Detection (/20) (Cutoff score < 15)
Incomplete Letters (/20) (Cutoff score < 16)
Dot Counting (/10) (Cutoff score < 8)
43.9 sec
(14.1 sec)
98.90 sec
(35.3 sec)
10.1 (2.3)
6.3 (2.3)
26.8 (5.7)
11.7 (6.5)
10.4 (6.6)
58.4 (10.1)
35.8 (5.2)
20.7 (6.8)
28.8 (14.7)
19.1 (1.2)
19.2 (0.9)
9.8 (0.4)
Position Discrimination (/20) (Cutoff score < 18)
19.1 (1.7)
Number Location (/10) (Cutoff score < 7)
Cube Analysis (/10) (Cutoff score< 6)
Silhouettes (/30) (Cutoff score < 15)
Object Decision (/20) (Cutoff score < 14)
9.0 (1.7)
9.4 (1.3)
19.3 (5.3)
16.6 (2.0)
Progressive Silhouettes (/20) (Cutoff score > 15)
10.4 (3.3)
40.9%ile
52.0%ile
50.6%ile
26.9%ile
27.9%ile
37.2%ile
32.6%ile
63.9%ile
74.0%ile
72.2%ile
52.9%ile
Pass
Pass
Pass
Pass
Pass
Pass
Pass
Pass
Pass
44.1 segundo
(14.4 segundo)
95.5 segundo
(34.5 segundo)
10.2 (2.3)
6.4 (2.3)
26.6 (5.8)
12.1 (6.4)
10.7 (6.7)
58.7 (10.1)
36.0 (5.2)
20.6 (7.0)
29.3 (14.9)
19.0 (1.2)
19.2 (0.9)
9.8 (0.4)
19.1 (1.7)
9.0 (1.7)
9.3 (1.3)
19.2 (5.4)
16.7 (2.0)
10.2 (3.2)
40.6%ile
53.2%ile
51.9%ile
28.5%ile
27.6%ile
39.5%ile
34.3%ile
64.8%ile
74.9%ile
71.5%ile
53.6%ile
Pass
Pass
Pass
Pass
Pass
Pass
Pass
Pass
Pass
Subjective memory rating (Memoria
Functioning Questionnaire, /448)
290.6 (54.2)
Minimal subjective
difficulties
291.9 (55.3)
Minimal subjective
difficulties
Maximum and cutoff scores for tests are indicated in parentheses in the left column. Note that two participants did not complete the subjective
memory questionnaire. WMS-IV = Wechsler Memory Scale, 4ª edición.
et al., 2012); aquí, we are interested in how the relation-
ships between cognitive processes and alERC volumes
may be distinct from those observed with PRC volumes.
Our goal is to better understand how the alERC might
contribute to all of these spatial/object memory pro-
cesses that have long been associated with its surround-
ing MTL regions and how volumetric differences in
these regions in healthy older adults might affect those
procesos.
Intrarater and Interrater Segmentation Reliability
Intrarater reliability was established by comparing the
segmentation of five randomly selected scans by the
714
Revista de neurociencia cognitiva
Volumen 31, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
oh
C
norte
_
a
_
0
1
3
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Mesa 2. Interrater and Intrarater Reliability Measurements for Manual Segmentation
Interrater: Dice
Interrater: CPI
Intrarater: Dice
Intrarater: CPI
Subregion
CA1
Subiculum
DG/CA23
alERC
pmERC
PRC
PHC
Izquierda
.88
.85
.91
.86
.82
.87
.86
Bien
.87
.84
.90
.85
.80
.89
.84
Izquierda
.94
.89
.94
.96
.90
.98
.89
Bien
.95
.88
.99
.86
.86
.91
.95
Izquierda
.74
.67
.75
.72
.59
.74
.71
Bien
.66
.66
.73
.73
.64
.76
.77
Izquierda
.92
.81
.91
.87
.95
.98
.86
Bien
.91
.85
.96
.71
.80
.99
.96
Dice was computed for both intrarater and interrater agreement. CPI(3, k) was calculated for intrarater, and ICC(2, k) was computed for interrater
fiabilidad.
same rater (l. y.) after a delay of 1–4 months. Interrater
reliability was evaluated by comparing the segmentation
of five randomly selected scans by a second rater (R. k. o.)
to those of L. Y. Both authors were blinded to MoCA
puntaje, task performance, and the identities of participants
until after manual segmentation (including interrater and
intrarater reliability) was completed. Reliability was as-
sessed using the intraclass correlation coefficient (CPI;
which evaluates volume reliability; Shrout & Fleiss, 1979)
and the Dice metric (which also takes spatial overlap into
cuenta; Dice, 1945), computed separately for each region
in each hemisphere. CPI(3, k) was computed for intrarater
fiabilidad (consistencia), and ICC(2, k) was computed for
interrater reliability (agreement). Dice was derived using
la formula 2 * (area of intersecting region) / (area of orig-
inal segmentation + area of repeat segmentation); a Dice
overlap metric of 0 represents no overlap, whereas a met-
ric of 1 represents perfect overlap. Intrarater and interrater
reliability results are shown in Table 2. These scores are
comparable to reliability values reported in the literature
for manual segmentation of hippocampal subfields and
MTL cortices (Yushkevich, Pluta, et al., 2015; Wisse et al.,
2012) and with our previous work (Olsen et al., 2013;
Palombo et al., 2013).
Cifra 1. The modified version
of the Olsen–Amaral–Palombo
segmentation protocol used in
este estudio. Inset images depict
coronal slices of the MTL taken
at various points along the long
axis of the hippocampus (como
shown in the sagittal view in
figure at bottom left). Cifra
previously published in Olsen
et al. (2017) and reproduced
with permission.
Yeung et al.
715
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
oh
C
norte
_
a
_
0
1
3
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Mesa 3. Average Volumes (±SD, in mm3) for Each of the Manually Segmented Hippocampal Subfields and MTL Cortices (Corrected
for Head Size)
Brain Region
Hippocampus
CA1
Subiculum
DG/CA23
MTL
PRC
alERC
pmERC
PHC
All Participants (norte = 32)
Participants Included in Data Analysis (norte = 30)
1238.41 ± 149.33
1105.20 ± 192.91
1957.19 ± 351.09
4966.35 ± 1154.23
1343.32 ± 274.72
440.84 ± 112.80
3640.21 ± 636.00
1237.15 ± 154.22
1103.46 ± 194.80
1963.32 ± 357.11
4954.03 ± 1118.44
1330.96 ± 278.97
443.50 ± 113.33
3649.43 ± 655.53
Volume Correction for Head Size
Eye Tracker Setup
All manually segmented region volumes were corrected for
head size using a regression-based method to account for
differences in brain size between participants. Estimated
total intracranial volume (eTIV) was derived from the
whole-brain T1-weighted scans using FreeSurfer (v5.3;
Buckner et al., 2004). By regressing the volume of each
region with eTIV, a regression slope β was obtained for
each region (representing the effect of eTIV change on that
region’s volume). Entonces, the volume of each region was
adjusted by that participant’s eTIV using the formula
Volumeadjusted = Volumeraw + b(eTIVparticipant − eTIVmean).
The head size correction was separately computed for
each region in each hemisphere. In our previous work
with this participant group (Olsen et al., 2017), Hicimos
not observe an interaction between cognitive decline
and hemisphere. De este modo, following this previous work, volumen-
umes were summed in each region across the two hemi-
spheres, giving a single volume for each region for each
partícipe.
The experimental task was presented on a 21.2-in. lun-
itor (36 × 30 centímetro) at a resolution of 1024 × 768 píxeles
using Experiment Builder (SR Research, Mississauga,
ON). Eye-tracking measures were recorded using an
EyeLink 1000 desktop-mounted eye tracker, sampling at
una tasa de 1000 Hz, with a spatial resolution of 0.01° and an
accuracy of 0.25. Participants were positioned 55 centímetro
away from the monitor and placed their heads on a chin-
rest to limit head motion. A 9-point calibration was per-
formed before testing and was repeated until the average
gaze error was less than 1°, with no point having a gaze
error exceeding 1.5°. Before each trial, a 1-sec drift cor-
rection was performed, with a 9-point calibration being
repeated if drift error exceeded 2°.
Stimuli and ROIs
Eight categories of computer-generated household
escenas (bathrooms, bedrooms, decks, dining rooms,
Mesa 4. Pearson’s Correlations between Volumes of Manually Segmented Brain Regions
CA1
1
Subiculum
DG/CA23
.375*
1
.671**
.162
1
PRC
.366*
−.272
.468**
1
alERC
.318*
−.015
.258
.395*
1
pmERC
.347*
.590**
.219
.011
.198
1
PHC
.440**
.228
.311
.159
.201
.274
1
CA1
Subiculum
DG/CA23
PRC
alERC
pmERC
PHC
*pag < .05.
**p < .01.
716
Journal of Cognitive Neuroscience
Volume 31, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
o
c
n
_
a
_
0
1
3
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
o
c
n
_
a
_
0
1
3
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
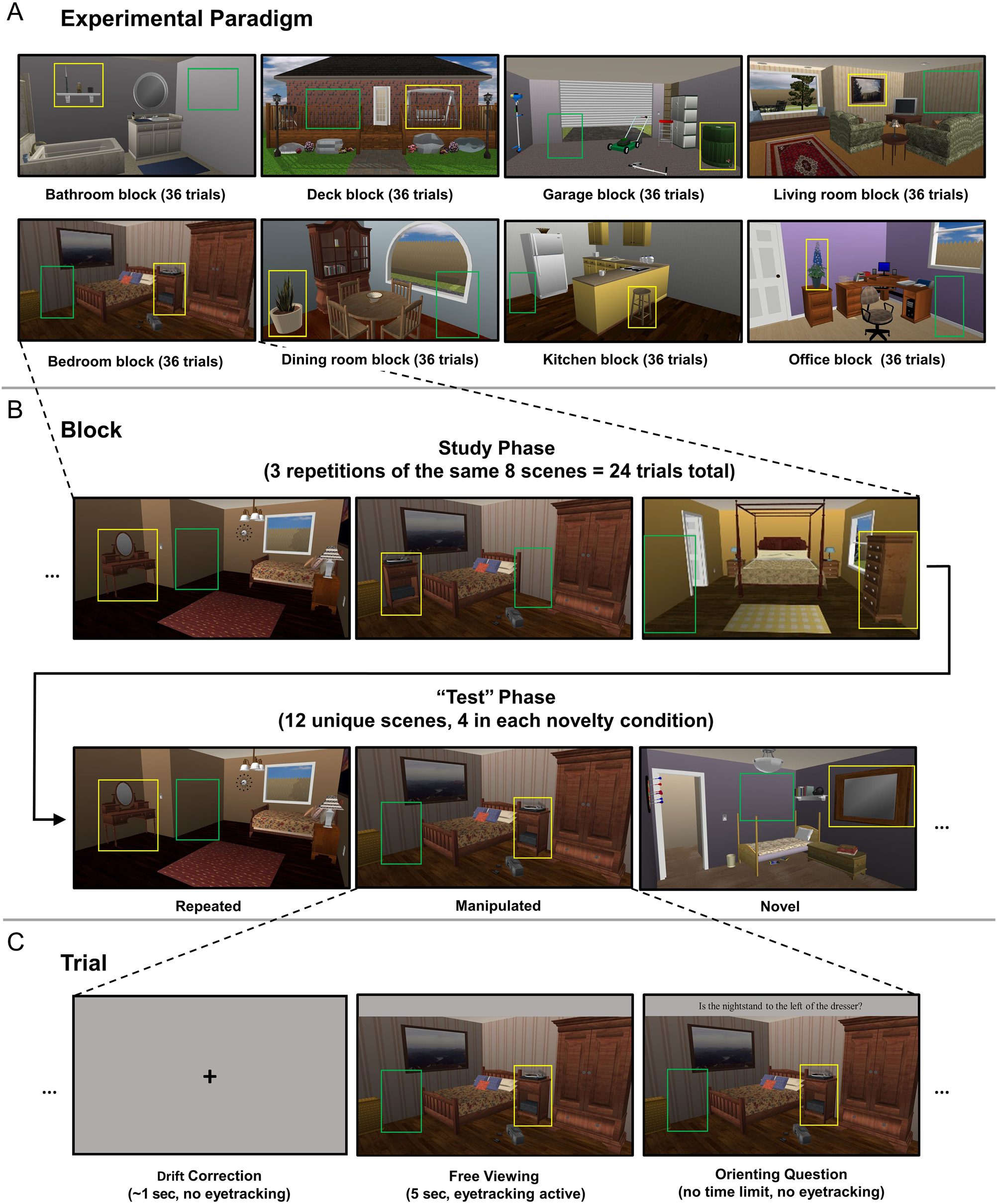
Figure 2. Schematic illustration of experimental paradigm. (A) Examples of each of the eight categories of scenes used in this study. (B) Arrangement
of trials within each block. Each block had a study phase of 24 trials (eight unique scenes, repeated three times), followed by a “test” phase
of 12 trials (four repeated scenes, four manipulated scenes, and four novel scenes). (C) Single trial timing. For all trials (in both study and “test”
phases), after drift correction, participants freely viewed a scene for 5 sec, followed by a yes/no orienting question directed to the critical
object. For illustrative purposes, in all three panels, the critical object ROI (around the critical object) is shown in yellow, and the empty ROI
(a similarly sized area covering the previous location of the critical object in manipulated scenes or an empty location in repeated/novel scenes) is
shown in green. Note that the ROIs were not visible to participants during the experiment.
Yeung et al.
717
garages, kitchens, living rooms, and offices) were used
in this study (Figure 2). All stimuli were created using
Punch! Home Design software (Encore Software). Each
scene contained thematically appropriate objects, and
all objects were unique to each individual scene. For
each scene, we created two versions—a standard and an
alternate. The alternate version of each scene was identi-
cal to the standard version in all respects, except for the
location of a critical object within the scene. The standard
version of the scene was used in all the “test” trials (i.e., it
appeared in different test conditions for different partici-
pants as a result of counterbalancing). This arrangement
allowed us to make direct comparisons between the same
standard version of the scene, regardless of which test
condition it appeared in for each individual participant.
The design followed the counterbalancing procedures
used in Ryan et al. (2000) and Smith et al. (2006). The
standard version of each scene was also used in study
trials whose scenes would be shown again in the repeated
test condition. The alternate version of a scene was used
for study trials whose corresponding test trial would be
shown in the manipulated test condition. For instance,
in Figure 2, the standard version of the scene is shown
as the “test” scene in the manipulated test condition (cen-
ter bottom), whereas the alternate version of the scene is
shown as the corresponding study trial above it. All of
the scenes measured 1028 × 518 pixels, subtending
the entire width and two thirds of the height of the
display screen. The scenes were centered on the screen
vertically.
Three rectangular ROIs were defined for each scene
(Figure 3). The “whole scene ROI” (shown in red) en-
compassed the entire scene depicted and was uniformly
1028 × 518 pixels large. The “critical object ROI” (shown
in yellow) was drawn to include only the critical object
and to minimize, as far as possible, the inclusion of parts
of any other objects in the scene. The “empty ROI”
(shown in green) covered an empty location in the scene
and was drawn to specifically minimize, as far as possible,
parts of any other objects in the scene. Importantly, the
empty ROI matched the location where the critical object
ROI had been located during the study phase for “manip-
ulated” scenes (which was simply an empty location on
the scene for the “repeated” and “novel” scenes).
Within each scene, the critical object ROI and empty
ROI were similarly, but not identically, sized. This was
necessary to ensure that the ROIs did not include, as
far as possible, any part of any other objects in the scene,
which might receive additional fixations during the
viewing period that are not directed to the critical object
or the empty location. Note that no comparisons were
made between the critical object ROI and the empty
ROI; rather, all comparisons were within the same ROI
across conditions. Across the entire stimulus set, the
mean critical object ROI had an area of 36,122 pixels
(6.80% of the scene, SD = 20,853 pixels), whereas the
mean empty ROI had an area of 37,575 pixels (7.08% of
the scene, SD = 17,778 pixels).
Eye-tracking Task
We employed an eye-tracking-based paradigm assessing
processing of objects within scenes, as assessed at vary-
ing levels of novelty, and examined how volumetric
differences in alERC (and other MTL/hippocampal re-
gions) affected object-in-place memory and object-trace
memory (Figures 2 and 3). In each trial, participants in-
cidentally viewed computer-generated scenes, depicting
household locations (e.g., bedrooms, kitchens), for 5 sec
(Figure 2C). After viewing each scene, participants were
asked to respond to a yes/no orienting question (ap-
pearing above the scene), directing attention to a critical
object in the scene (e.g., “Is the nightstand to the left of
the dresser?”). This followed the examples of Ryan et al.
(2000) and Hannula, Tranel, and Cohen (2006), who also
used orienting questions to direct viewers’ attention to a
Figure 3. An example of a single manipulated scene trial, illustrating how the proportion of fixations outcome variables were calculated. Note that
the ROIs and fixations were not visible to participants as they performed the task.
718
Journal of Cognitive Neuroscience
Volume 31, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
5
7
1
1
1
7
8
8
5
2
2
/
j
o
c
n
_
a
_
0
1
3
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
critical object within a scene. No time limit was imposed
on answering this orienting question. Visual fixations
made within the three ROIs (Figure 3) were recorded
during the 5-sec viewing period, but not during the
subsequent period when participants were asked to re-
spond to the orienting question. A brief eye tracker drift
correction (<1 sec) was performed between each trial.
The experiment organized into eight blocks of 36
trials each; all the scenes in block depicted same
type location (e.g., one consisted entirely of
bedrooms, another kitchens; Figure 2A).
Each included a study phase 24 trials (eight unique
scenes viewed three times each, were
viewed at least once before any repeated), followed
by “test” 12 (Figure 2B). It is important to
note that participants were not informed distinction
between and phases, as task instruc-
tions same across trials. In phase,
there four test conditions
(i.e., total phase), which differed
in degree novelty The con-
ditions (1) “repeated scenes,” identical
versions presented during phase;
(2) “manipulated identical to scene
presented phase, except critical
object had moved different scene; and
(3) “novel seen study
phase but type scene rest of
the kitchen kitchens).
Each repeated or manipulated shown
during corresponded specific
scene phase. For ex-
ample, 2B, (bottom left) is
identical (top
left), whereas center) is
the studied (top center), with the
exception critical object (the nightstand) was
moved from middle room left side room. Furthermore, orienting question for
each specific (including both versions the
scenes used for condition) re-
mained repetitions, did cor-
rect answer question. instance, in
Figure 2, scene
(with “Is nightstand left
of dresser?” this the
same (“yes”) alternate version scene
shown standard version
of shown manip-
ulated scene.
Across participants, condition particular
scene appeared (see Stimuli ROIs section), the
ordering blocks, correct response the
orienting counter-
balanced. we have
been using an example, half re-
ceived there a
stereo on top nightstand?” correct
answer “no” scene.
Eye-tracking Outcome Variables
We defined eye-tracking-based outcome variables
for based similar measures previ-
ously employed other studies object-scene memory
(Smith & Squire, 2008; Ryan et al., 2000, 2007; Smith
et 2006; Cohen, 2004a, 2004b; 3).
Our first primary variable “proportion
of fixations ROI.” We mea-
sure viewing assess object-in-place memory. This
measure calculated individual trial by divid-
ing number made object
ROI whole
scene ROI then averaging over a
single (i.e., repeated, manipulated, or
novel) participant. novel condition,
participants would have known the
critical (as they yet be orienting
question matching scene). However, repeated
and conditions, would
have previously (or nearly identical)
version and, thus, the
opportunity associate mentioned
in its spatial that
scene. particularly interested how pro-
portion directed ROI
differed conditions. difference
in>