Neural Coding of Visual Objects Rapidly Reconfigures to
Reflect Subtrial Shifts in Attentional Focus
Lydia Barnes1
, Erin Goddard2
, and Alexandra Woolgar1,3
Abstracto
■ Every day, we respond to the dynamic world around us by
choosing actions to meet our goals. Flexible neural populations
are thought to support this process by adapting to prioritize
task-relevant information, driving coding in specialized brain
regions toward stimuli and actions that are currently most
important. Respectivamente, human fMRI shows that activity pat-
terns in frontoparietal cortex contain more information about
visual features when they are task-relevant. Sin embargo, if this
preferential coding drives momentary focus, Por ejemplo, a
solve each part of a task in turn, it must reconfigure more
quickly than we can observe with fMRI. Aquí, we used multivar-
iate pattern analysis of magnetoencephalography data to test
for rapid reconfiguration of stimulus information when a new
feature becomes relevant within a trial. Participants saw two dis-
plays on each trial. They attended to the shape of a first target
then the color of a second, or vice versa, and reported the
attended features at a choice display. We found evidence of
preferential coding for the relevant features in both trial phases,
even as participants shifted attention mid-trial, commensurate
with fast subtrial reconfiguration. Sin embargo, we only found this
pattern of results when the stimulus displays contained multiple
objects and not in a simpler task with the same structure. El
data suggest that adaptive coding in humans can operate on a
fast, subtrial timescale, suitable for supporting periods of
momentary focus when complex tasks are broken down into
simpler ones, but may not always do so. ■
INTRODUCCIÓN
Human cognition is remarkably flexible. We can fluidly
direct our focus toward what we need for our current goal,
seamlessly adapt to changes in our environment, and gen-
eralize from what we know to solve new problems. Several
lines of research suggest that this flexibility emerges from
activity in frontoparietal cortex. Cognitively challenging
tasks elicit robust activity in the “multiple demand”
(Maryland) system—a distributed network of frontal and
parietal cortex recruited by a wide range of tasks (Assem,
vidrio, VanEssen, & Duncan, 2020; Fedorenko, Duncan,
& Kanwisher, 2013; Duncan, 2010). Damage to this system
linearly predicts fluid intelligence scores ( Woolgar,
Duncan, Manes, & Fedorenko, 2018; Woolgar et al.,
2010), which in turn powerfully predict how well we are
able to acquire new skills.
The characteristic adaptability of frontoparietal regions
means that they are ideally suited to supporting flexible
cognition. Por ejemplo, patterns of activity in the MD sys-
tema, measured with fMRI, adapt to code information that
is relevant for the current task. MD patterns can encode
many different aspects of a task (p.ej., visual: Jackson,
Rich, williams, & Woolgar, 2016; vibrotactile: Woolgar
& Zopf, 2017; for a review see Woolgar, Jackson, & Duncan,
1University of Cambridge, Reino Unido, 2University of New
South Wales, Sídney, Australia, 3Macquarie University, Sídney,
Australia
2016), commensurate with a high degree of mixed selec-
tivity in these regions (Fusi, Molinero, & Rigotti, 2016; Rigotti
et al., 2013). Además, MD coding for task-relevant stimuli
is enhanced when stimuli are more difficult to discriminate
( Woolgar, williams, & Rich, 2015; Woolgar, Hampshire,
Thompson, & Duncan, 2011) and changes to prioritize
information that is at the focus of attention ( Jackson &
Woolgar, 2018; Woolgar, williams, et al., 2015). Activity
in at least one MD region appears to be causal for facili-
tating task-relevant information processing elsewhere in
the MD system ( Jackson, Feredoes, Rich, Lindner, &
Woolgar, 2021). This may provide a source of bias to
more specialized brain regions, Por ejemplo, a través de
task-dependent connectivity (Cole y col., 2013; ver, p.ej.,
the work of Baldauf & Desimone, 2014). Como consecuencia,
adaptive coding has been proposed as a central compo-
nent of goal-directed attention, biasing sensory and
motor brain regions to perceive and respond to informa-
tion that is relevant to our current task.
A key outstanding question concerns the temporal scale
of this process. Aquí, we explore the “attentional epi-
sodes” account of flexible behavior (Duncan, 2013), cual
predicts a fast temporal scale. This account draws on stud-
ies of human and artificial intelligence to propose that flex-
ible behavior rests on our ability to break a complex task
down into a series of simpler parts, and to focus, moment-
to-moment, on the information needed for each part
(Duncan, Chilinski, mitchell, & Bhandari, 2017; Duncan,
© 2022 Instituto de Tecnología de Massachusetts. Published under
a Creative Commons Attribution 4.0 Internacional (CC POR 4.0) licencia.
Revista de neurociencia cognitiva 34:5, páginas. 806–822
https://doi.org/10.1162/jocn_a_01832
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
oh
C
norte
_
a
_
0
1
8
3
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
2013; Duncan, Schramm, Thompson, & Dumontheil,
2012). En efecto, there is some evidence that this ability
may underpin performance on novel problem-solving
tareas. Por ejemplo, explicitly breaking a complex task into
simple parts removes the performance gap between peo-
ple with high and low fluid intelligence scores (Duncan
et al., 2017; see also the work of O’Brien, mitchell, Duncan,
& holmes, 2020). In this matrix reasoning study, participar-
pants viewed a 2 × 2 grid with three of the four squares
filled with an image. They had to abstract relationships
between the images to fill in the remaining square. Images
consisted of multiple features. In the second half of the
experimento, each feature was presented separately. Estos
segmented problems were trivial for participants to solve,
regardless of whether they struggled or performed well on
the difficult, unsegmented problems. This led the authors
to propose that participants who were able to solve the
unsegmented problems were better able to mentally
break them down into their relevant parts. Adaptive cod-
ing could be a key component of this segmentation by
driving momentary focus toward subsets of the available
information in turn.
From these studies, it seems intuitive that flexible cog-
nition involves identifying simple problems that we can
solve and addressing them in an ordered sequence. Cómo-
alguna vez, we do not have clear insight into whether codes
reconfigure quickly enough to prioritize relevant informa-
tion throughout a task. The bulk of research on adaptive
coding in humans uses fMRI. Although these studies show
trial-to-trial shifts in what information can be discriminated
from activity patterns (p.ej., Woolgar, williams, et al., 2015;
Woolgar, Hampshire, et al., 2011), the coarse temporal
resolution of fMRI does not support precise, subsecond
measurement of changes in task information.
Time-resolved methods, such as electrophysiology,
EEG, and magnetoencephalography (MEG), offer promis-
ing evidence for rapid changes in task representation.
Nonhuman primate studies show that the same frontal
neurons can encode object identity and then location
within a single trial, as monkeys attended to what and then
where an object was (Rao, Rainer, & Molinero, 1997). Estos
data demonstrate that the neural population can systema-
tically change its activity pattern in synchrony with the
tarea. Sin embargo, they are taken from highly trained mon-
keys and could rely on a learned response rather than
instantaneous shifts in a flexible brain system. More recent
work by Spaak, Watanabe, Funahashi, and Stokes (2017)
demonstrates that, even when the same information is
encoded across phases of a task, neurons in primate lateral
pFC dynamically update what they encode. This dynamic
reallocation of selectivity within a trial makes plausible
rapid shifts in the information that these adaptive brain
regions represent. In humans, stronger coding for visual
features when they are task-relevant compared to task-
irrelevant emerges in MEG data as early as 100 msec from
stimulus onset (Goddard, Carlson, & Woolgar, 2022;
Moerel, Rich, & Woolgar, 2021; Battistoni, Kaiser, Hickey,
& Peelen, 2020; Wen, Duncan, & mitchell, 2019), with sus-
tained coding of the relevant feature emerging around
200–400 msec in the MEG/EEG signal (Goddard et al.,
2022; Grootswagers, robinson, Shatek, & Carlson, 2021;
Moerel et al., 2021; Yip, Cheung, Ngan, Wong, & Wong,
2021). This provides preliminary evidence that population
codes for task-relevant features develop rapidly, pero esto
previous time-resolved human neuroimaging work did
not require participants to shift their attention within tri-
como, so we do not know how rapidly information codes
update to redirect attention in each part of a task.
Rapid reorganization of information coding within a task
has been proposed as key component of how we solve
complex tasks, but the neural correlates of this have not
yet been studied in the human brain. Aquí, we test the
dynamic adaptation of task representations when what is
relevant changes within single trials. We used MEG to track
shifts in adaptive coding with subsecond precision across
fragments of two rapidly changing tasks. Considerando el
strong association between task difficulty and the brain
regions implicated in adaptive coding (Crittenden &
Duncan, 2014; Fedorenko et al., 2013), we tested this
at two levels of attentional demand. In Experiment 1,
we used simple stimuli to track preferential coding of
relevant information under low attentional demands.
In Experiment 2, we used a complex stimulus space,
abstracted decisions, and the presence of distractors to
track preferential coding of relevant information under
high attentional demands. Across both experiments, nosotros
asked whether neural codes for relevant stimulus informa-
tion rapidly reconfigure when what is relevant changes
mid-trial.
MÉTODOS
Participantes
Participants were selected to (a) have normal or corrected-
to-normal visual acuity and normal color vision, (b) ser
right-handed, (C) have no exposure to fMRI in the previous
week, (d) have no nonremovable metal objects, y (mi)
have no history of neurological damage or current psycho-
active medication. Prospective participants were informed
of the study’s selection criteria, aims, and procedure,
through a research participation site.
For Experiment 1, 20 Participantes (17 women, 3 hombres,
mean age 25 ± 6 años) were recruited from the paid par-
ticipant pool at Macquarie University (Sídney). They gave
written informed consent before participating and were
paid AUD$30 for their time. Ethics approval was obtained
from the Human Research Ethics Committee at Macquarie
Universidad (5201300602).
For Experiment 2, 20 Participantes (16 women, 4 hombres,
mean age 31 ± 12 años) were recruited from the volun-
teer panel at the MRC Cognition and Brain Sciences Unit
(Cambridge). They gave written informed consent before
each testing session and were paid GBP£40 for their time.
Barnes, Goddard, and Woolgar
807
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
oh
C
norte
_
a
_
0
1
8
3
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Participants were additionally asked to only volunteer if
they had existing structural MRI scans on the panel data-
base. Two participants took part before completing a
structural scan; one obtained a scan through another study
conducted at the MRC Cognition and Brain Sciences Unit,
and the other returned for a separate MRI session as part
of this study. This participant gave written informed con-
sent before completing the structural scan and was paid
an additional GBP£20 for this component of their time.
Ethics approval was obtained from the Psychology
Research Ethics Committee at the University of Cambridge
(PRE.2018.101).
Estímulos
Stimuli were created in MATLAB (The MathWorks, v2012b)
and presented with Psychtoolbox (Kleiner et al., 2007;
Brainard, 1997). In Experiment 1, they were displayed with
an InFocus IN5108 LCD back projection monitor at a
viewing distance of 113 cm. In Experiment 2, they were
displayed with a Panasonic PT-D7700 projector at a viewing
distance of 150 cm.
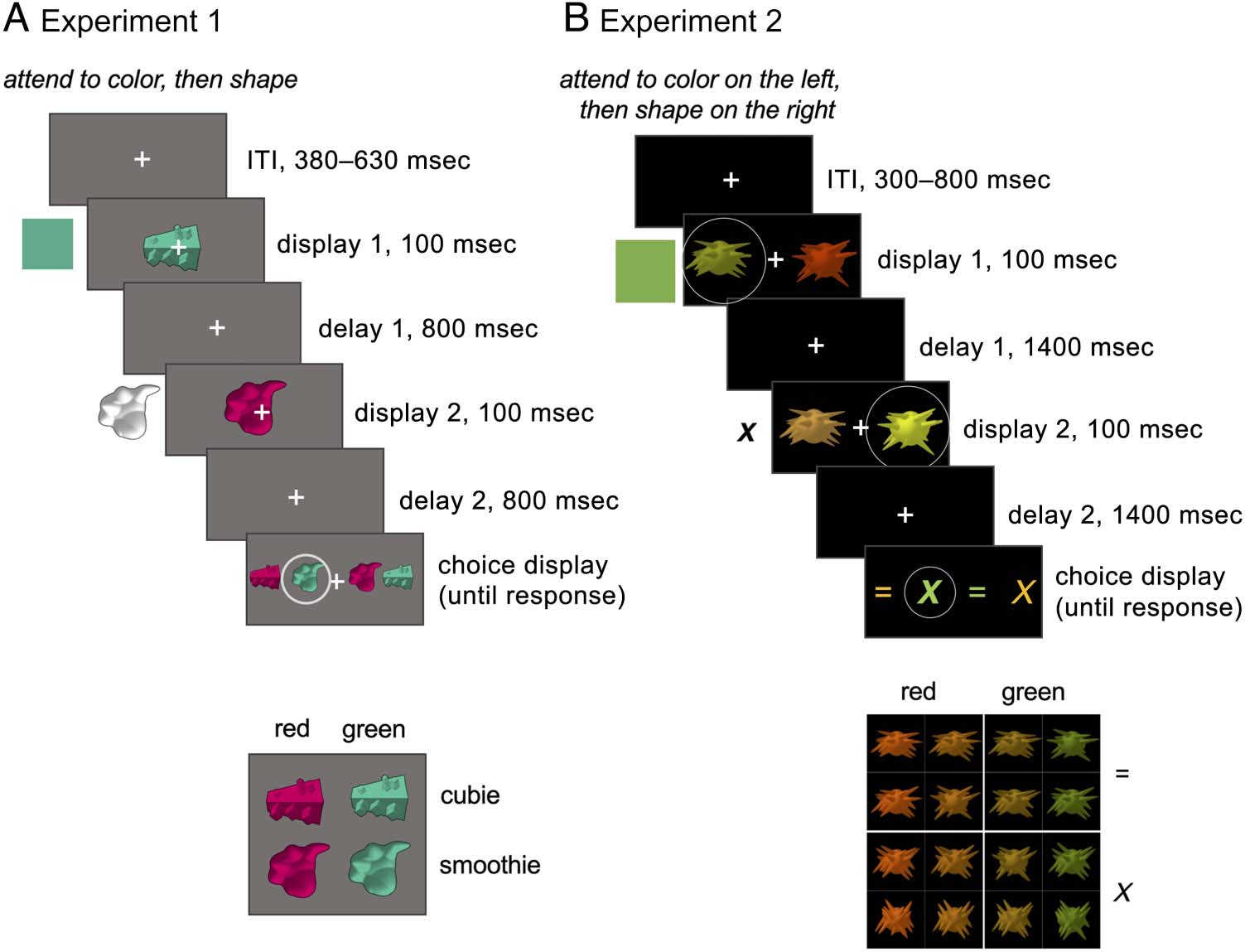
Experimento 1 stimuli consisted of four novel objects
(Op de Beeck, Panadero, DiCarlo, & Kanwisher, 2006; ver
Cifra 1) that were either “cubie” or “smoothie” shaped,
and green or red (rojo, verde, blue 0–194-155 and 224–0-
98). Colors were chosen for high chromatic variation and
strong contrast against the dark gray background (rojo,
verde, blue 30–30-30).
Experimento 2 stimuli consisted of 16 novel “spiky”
objects, adapted from the Op de Beeck et al. (2006)
“spiky” stimuli, selected at four points on a spectrum of
red to green, and upright to flat (Goddard et al., 2022).
Color values were numerically equally spaced in u0v0 color
space between [0.35, 0.53] y [0.16, 0.56]. Shapes were
also equally spaced to create four steps in orientation from
upright to flat. Each step included 100 shape exemplars,
with different spikes indicating the orientation, to discour-
age participants from judging orientation based on a
single spike.
Tarea
Experimento 1 used simple displays and stimuli, optimized
for strong visual signals. Each block began with a written
cue instructing participants to attend to the color of the
first object and the shape of the second object, or vice
versa. On each trial, participants viewed two brief displays
(100 mseg), each followed by a delay (800 mseg; ver
Cifra 1). Finalmente, they were prompted to select an object
from a choice display that comprised the combination of
the remembered features. All four objects appeared on the
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
oh
C
norte
_
a
_
0
1
8
3
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Stimuli and example trials for Experiments 1 y 2. Relevant information for each epoch is shown beside the display. A shows an example
trial for Experiment 1, with a single object on each display. In this trial, the relevant features are “green” (Target 1) and “smoothie” (Target 2),
resulting in a “green smoothie” response on the choice display. Stimuli could be red, verde, “cubie,” or “smoothie.” B shows an example trial for
Experimento 2, in which the participant was cued to attend to color on the left and then shape on the right. The relevant features were thus green and
“X,” leading to a response of “green X” on the choice display. Stimuli varied in four steps from red to green, and from X to =, but were assigned to
binary red / verde, X / = categories. Circles represent the focus of attention and correct choice and were not shown to participants.
808
Revista de neurociencia cognitiva
Volumen 34, Número 5
choice display, and participants selected the object that
matched the color and shape they had extracted from
the preceding displays. Por ejemplo, under the rule
“attend shape, then color,” if the first object was a “cubie”
and the second object was “red,” the target on the choice
display was a red cubie. Participants indicated their choice
by pressing one of four buttons on a bimanual fiber optic
response pad operated with the four fingers of the right
mano. The mapping from object location to response but-
ton was intuitive (far left button for far left object, etc.) y
consistent across trials; sin embargo, the arrangement of the
four objects on the choice display varied to prevent partic-
ipants preparing a motor response until the display screen
was shown. Stimulus arrangements were presented in
pseudorandom order and balanced within each rule such
that all stimuli on the second display were equally pre-
ceded by each stimulus on the first display, and the correct
choice pertained equally to all motor responses. Objects
were sampled with replacement, meaning that the same
object could appear in both stimulus displays, but partici-
pants could not use the trial sequence to anticipate when
this would occur. If a participant made three consecutive
incorrect or slow responses (> 3 segundo), the task was paused
and the cue was presented again until the participant ver-
bally confirmed that they understood the rule for that
block. Average accuracy and response times were dis-
played at the end of each block.
Experimento 2 followed the structure of Experiment 1,
but used simultaneously presented objects and subtler
stimulus discriminations, optimized for high attentional
load. For this experiment, each display contained two
objects. Participants were cued to both a location and fea-
tura, Por ejemplo, “attend to shape on the right, then color
on the left.” Relevant location and feature always changed
from Display 1 to Display 2, creating four possible rules.
Delay periods were increased to 1500 msec to allow accu-
rate responses, following piloting of the task. Participantes
judged the color and shape category of the target
objects’ features. The choice display contained the sym-
bols X and =, presented in the average of the two “red”
colors and the average of the two “green” colors, to rep-
resent the four possible answers. These symbols were
chosen to encourage participants to make category-level
decisions about the objects. As in Experiment 1, the spa-
tial arrangement of the items on the choice display was
updated on each trial.
Procedimiento
Experimento 1
Each participant first completed four blocks of 10 práctica
trials outside the shielded room. These were identical to
test trials except that (a) participants received feedback
of “correct,” “incorrect,” or a red screen signifying a slow
respuesta (> 3 segundo), on every trial, (b) display durations
in the first two practice blocks were slowed from 100 a
500 msec to ease participants into the task, y (C)
response key codes were marked on the choice display
to train participants in the location-response mapping.
Once in the MEG scanner, participants completed eight
blocks of 96 trials each, with feedback at the end of each
block. Each block lasted approximately 7 mín.. Blocks
alternated between the two rules, “attend shape, entonces
color” and “attend color, then shape,” with the order
counterbalanced across participants.
Experimento 2
Participants learned the stimulus categories (red vs. verde,
upright vs. flat) and the task in a separate training session.
Training could be on the day of or the day before the scan-
ner session. Training consisted of two blocks of 50 cate-
gory learning trials, in which they saw a single object for
100 msec and pressed a button to indicate its shape or
color category. They then began training on the core
tarea. Within-trial delay periods began at 4 sec and
reduced to 1.5 sec in three steps (3 segundo, 2 segundo, y
1.5 segundo). Participants completed a minimum of 10 ensayos
at each of the four speeds for each of the four rules
(es decir., al menos 40 trials per rule). Después 10 trials were com-
pleted, the speed increased when the participant got eight
trials correct in any 10 consecutive trials. Feedback was
given on each trial by a brighter fixation cross for correct
responses and a blue fixation cross for incorrect
respuestas, shown for the first 100 msec of the posttrial
interval. This procedure trained each participant to the
same criterion without penalizing them for errors early
in the block.
Once in the MEG, participants completed four blocks,
each corresponding to a single rule and comprising 258
ensayos, lasting approximately 20 mín.. Rule order was bal-
anced across participants.
MEG Data Acquisition
Experimento 1
We acquired MEG data in the Macquarie University KIT-
MEG laboratory using a whole-head horizontal dewar with
160 coaxial-type first-order gradiometers with a 50-mm
base (Model PQ1160R-N2; KIT; Uehara et al., 2003;
Kado et al., 1999) in a magnetically shielded room
(Fujihara Co. Limitado.). Primero, the tester fit the participant
with a cap containing five head position indicator coils.
The location of the nasion, left and right pre-auricular,
and each of the head position indicators were digitized
with a Polhemus Fastrak digitiser. This information was
copied to the data acquisition computer to track head
position during data collection. Participants lay supine
during the scan and were positioned with the top of the
head just touching the top of the MEG helmet. Any change
in head position relative to the start of the session was
checked and recorded after four blocks. MEG data were
recorded at 1000 Hz.
Barnes, Goddard, and Woolgar
809
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
oh
C
norte
_
a
_
0
1
8
3
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Experimento 2
We acquired MEG data with the MRC Cognition and Brain
Sciences’ Elekta-Neuromag 306-sensor Vectorview system
with active shielding. Ground and reference EEG elec-
trodes were placed on the cheek and nose. Bipolar elec-
trodes for eye movements were placed at the outer canthi,
above and below the left eye. Heartbeat electrodes were
on the left abdomen and right shoulder. Scalp EEG were
also applied for a separate project. Head position indica-
tors were placed on top of the EEG cap. Both head shape
and the location of the head position indicators were dig-
itized with a Polhemus Fastrak digitiser. Head position was
recorded continuously through the scan and viewed after
each block to ensure that the top of the participant’s head
stayed within 6 cm of the top of the helmet in the dewar
(mean movement across task 3.94 mm, range 0.5–15 mm).
Because targets in this experiment could appear to either
side of fixation, we also recorded eye movements with an
EyeLink 1000 eye tracker, which we calibrated before each
block. If we observed more information about the stimu-
lus at the relevant location, eye-tracking data would allow
us to measure the contribution of gaze. Sin embargo, nuestro
primary analysis compared features at the same location,
so we did not include the eye-tracking data here.
Analyses
MEG Processing
Because of active shielding and artifacts from continuous
head position indicators, data from Experiment 2 eran
first processed with Neuromag’s proprietary filtering soft-
mercancía (Maxfilter, 2010). We applied temporal signal space
separation to remove environmental artifacts, used contin-
uous head position information to correct for head move-
ment within each block, and reoriented each block to the
subjects’ initial head position.
All other processing was the same across experiments.
We used a minimal preprocessing pipeline to minimize
the chance of removing meaningful data. This was espe-
cially appropriate in our case, as our planned multivariate
analyses are typically robust to noise (Grootswagers,
Wardle, & Carlson, 2016). MEG data were imported into
MATLAB v2018b using Fieldtrip (Oostenveld, Fries,
Maris, & Schoffelen, 2011) and bandpass filtered (0.01–
200 Hz). Trials were epoched from a 100-msec prestimu-
lus baseline to the maximum possible trial duration (Exp 1:
4800 mseg, Exp 2: 5000 mseg). Principle component anal-
ysis was applied the data, retaining the first components
that together captured 99% of the variance. All sensors
were included in the analysis.
At the request of a reviewer, we also repeated the anal-
yses for Experiment 2 with additional independent com-
ponent analysis to remove heart- and eye-related artifacts.
We then used systematic averaging before decoding (p.ej.,
averaging across red and green trials when decoding
forma) to ensure a balanced test and training set. Estos
additional analyses (data not shown) produced compa-
rable results to what we report here with minimal
preprocessing.
MEG Decoding
We used multivariate pattern analysis to trace the informa-
tion about rule, color, and shape in each task phase. Nosotros
then compared the information about color when it was
relevant and irrelevant, repeating the comparison for
forma. Following previous studies, we expected that rule
información, which was known before each trial, would be
present throughout the trial and increase briefly after
visual displays (Goddard et al., 2022; Hebart, Bankson,
Harel, Panadero, & Cichy, 2018). We predicted that preferen-
tial coding would be reflected in improved decoding of
visual features when they were relevant, compared to irrel-
evant (Goddard et al., 2022; Grootswagers et al., 2021;
Moerel et al., 2021; Yip et al., 2021; Battistoni et al.,
2020; Wen et al., 2019; Hebart et al., 2018). Increased color
information when color was relevant would indicate that
information was flexibly coded according to task demands.
Our critical comparison, entonces, was how this happened for
the two task phases. If information about the relevant fea-
ture was prioritize d in both task epochs, this would indi-
cate that preferential coding can reconfigure in line with
subsecond shifts in what is relevant to the task.
We first trained a linear classifier (linear discriminant
análisis; see the work of Grootswagers et al., 2016) en
labeled data from two feature rules—“attend color, entonces
shape” and “attend shape, then color”—using all but one
trial from each category. We then tested whether the
weights that the classifier had learned to discriminate
the training data generalized to the remaining unobserved
ensayos. We repeated the process, leaving out a different
pair of trials each time, until all trials had acted as the test
datos. We then averaged the classification accuracy across
all test sets.
For color and shape classification, we trained a linear
classifier on labeled data from two categories—for exam-
por ejemplo, “red” and “green”—using all but one trial from each
categoría, for each feature rule separately. For Experi-
mento 2, we decoded pairs of shape or color, at a fixed
ubicación, for each feature and location rule. Por ejemplo,
we took trials under the rule “attend color on the left, entonces
shape on the right.” For items on the left on the first dis-
play, we decoded strong red versus yellow red, yellow red
versus yellow green, and so on for all six pairs of color. Nosotros
then averaged classifier accuracy across the six pairs into a
single measure of color information coding in the left
hemifield under this rule. We repeated this for each rule
to obtain four traces of left hemifield color information
codificación, representing color information when that location
and feature were relevant or irrelevant. We conducted the
same pairwise decoding and averaging for color in the
right hemifield. Conducting the analyses for each hemi-
field separately minimized the requirement for the
810
Revista de neurociencia cognitiva
Volumen 34, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
oh
C
norte
_
a
_
0
1
8
3
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
classifier to generalize patterns over space. Finalmente, nosotros
averaged the four traces of left hemifield color information
coding with the corresponding right hemifield traces to
produce a single trace for each attention condition:
“attended location, attended feature” (the task-relevant
trace), “attended location, unattended feature,” “unat-
tended location, attended feature,” and “unattended loca-
ción, unattended feature.” The two traces for color (o
forma) information at the attended location parallel the
two traces for each target in Experiment 1 and form the
central part of our analysis.
Statistical Tests
We tested whether decoding accuracy scores were above
chance using a null distribution generated from the data.
To generate this, we permuted the predicted class labels
so that they were randomly assigned over trials (Bae &
Luck, 2019). We calculated decoding accuracy as above
and repeated the process 10,000 times to produce a
decoding distribution for each participant and each
comparación. We then sampled 10,000 times across par-
ticipants’ null distributions to form a group-level null
distribución. At each time point, we calculated t-scores
for classification accuracy relative to the null distribution
(Stelzer, Chen, & Tornero, 2013). We used a threshold-
free cluster statistic (threshold step 0.1; Herrero &
Nichols, 2009) to flexibly set a cluster-forming threshold
to identify peaks in the t-score time course that were
more strong and/or sustained than expected from the
null distribution ( pag < .05). This maximizes sensitivity
to peaks that are most likely to reflect meaningful
change while down-weighting peaks that are small or
transient (Noble, Scheinost, & Constable, 2020; Vastano,
Ambrosini, Ulloa, & Brass, 2020; Pernet, Latinus,
Nichols, & Rousselet, 2015; Mensen & Khatami, 2013;
Smith & Nichols, 2009). We then used this threshold
to correct for multiple comparisons at the cluster level
across the whole trial. Decoding onset was the onset of
the first cluster for which decoding accuracy was reliably
above chance.
For between-conditions comparisons, we contrasted
the decoding trace for the target when it was the relevant
or irrelevant feature using a two-sided t test, implemented
in CoSMoMVPA (Oosterhof, Connolly, & Haxby, 2016)
with threshold-free cluster enhancement and a threshold
step of 0.1 ( p < .05; Smith & Nichols, 2009; Figures 4
and 5).
For Experiment 2, we also conducted secondary analy-
ses to assess the combined effects of spatial- and feature-
selective, as reported in the work of Goddard et al. (2022).
We conducted 2 × 2 ANOVAs to test, for each time bin,
whether stimulus color and shape information coding
was boosted (1) at the relevant compared to irrelevant
location, (2) when that stimulus feature was relevant for
the task compared to when it was irrelevant, and (3) when
both feature and location were relevant compared to all
other attention conditions. We quantified these as main
effects of Spatial and Feature-Selective Attention, and as
a planned comparison between the coding of the reported
feature at the attended location and the coding of that fea-
ture at that location in the other three attention condi-
tions (following our prediction from Goddard et al.,
2022). For example, we contrasted decoding for color
on the left when people were attending to color on the
left, with decoding for color on the left when attending
to shape on the left, color on the right, and shape on
the right. We present the results of these secondary anal-
yses in Figure 6.
Lastly, in Experiment 2, we asked whether attentional
effects had similar temporal profiles in Epoch 1 and Epoch
2 of the trial. We epoched the stimulus decoding traces for
the target, separately around the first and second stimulus
displays (0–1500 msec), using the same pretrial baseline
(−100 to 0 msec) for all traces. This created four overlaid
traces, a relevant and an irrelevant feature trace for
Epoch 1 and Epoch 2. We conducted a 2 × 2 ANOVA
with main effects of Relevance and Epoch. An interaction
term tested our hypothesis that preferential coding of
relevant information emerges earlier, or is more substan-
tial, in one epoch compared to the other.
RESULTS
Behavioral Performance
In Experiment 1, median accuracy was 93.3% (SD 7.5%),
with median RT of 829.2 msec (SD 210.7 msec). In Exper-
iment 2, median accuracy was 75.9% (SD 10.9%), with
median RT of 665.2 msec (SD 92.1 msec). In both tasks,
chance accuracy was 25%.
Rule Information Coding
We trained a classifier to discriminate between feature
attention rules (“attend shape, then color” from “attend
color, then shape”) from MEG data to extract a time course
of rule information coding (Figure 2). Because the rule was
cued at the start of the block, we expected that partici-
pants might prepare their task set in advance of the stim-
ulus display. We anticipated that rule information would
be more decodable after each display, when the rule could
be applied to extract relevant information (as in the work
of Goddard et al., 2022). Indeed, rule information coding
emerged early in both experiments, increasing after each
stimulus onset, and remaining above chance throughout
the trial. Rule information coding gradually ramped up
after each display in Experiment 1, whereas in Experi-
ment 2, rule information coding was elevated through-
out the trial and peaked steeply after each display. For
Experiment 2, we collapsed the feature rule analysis
over locations to mirror Experiment 1 (Figure 2). We
also decoded the location rule (i.e., “attend left, then
Barnes, Goddard, and Woolgar
811
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Feature rule decoding (“attend color then shape” vs. “attend shape then color”) for Experiment 1 (A) and Experiment 2 (B). Vertical gray
patches mark the stimulus displays and the maximum possible duration of the choice display. Vertical dotted lines mark the median response time
with one quartile on either side. Horizontal gray lines show chance (50%) bounded by the 95% confidence interval for the null mean, which we
estimated from permutation-based null data. Time points at which decoding was reliably different to the null based on threshold-free cluster
correction are marked below the trace in brown.
Figure 3. Location rule decoding (“attend left then right” vs. “attend right then left”) for Experiment 2. Vertical gray patches mark the stimulus
displays and the maximum possible duration of the choice display. Vertical dotted lines mark the median response time with one quartile on either
side. Horizontal gray lines show chance (50%) bounded by the 95% confidence interval for the null mean, which we estimated from permutation-
based null data. Time points at which decoding was reliably different to the null based on threshold-free cluster correction are marked below the
trace in brown.
812
Journal of Cognitive Neuroscience
Volume 34, Number 5
Figure 4. Color (A) and shape
(B) decoding for Experiment 1.
(A) and (B) show decoding
traces for the first and second
targets in the upper and lower
panels. Decoding accuracies are
shown for each feature when it
was relevant (blue) or irrelevant
(orange) for the task. Gray bars
mark the stimulus and response
display durations. Vertical lines
show the median response
time, ± one quartile. Times at
which decoding was greater
than chance, p < .05 using a
cluster-based correction for
multiple comparisons, are
marked below each trace in the
corresponding color. Relevant
information coding did not
reliably exceed coding for the
irrelevant feature at any time
point.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
right” and “attend right, then left”), which we show in
Figure 3 for completeness.
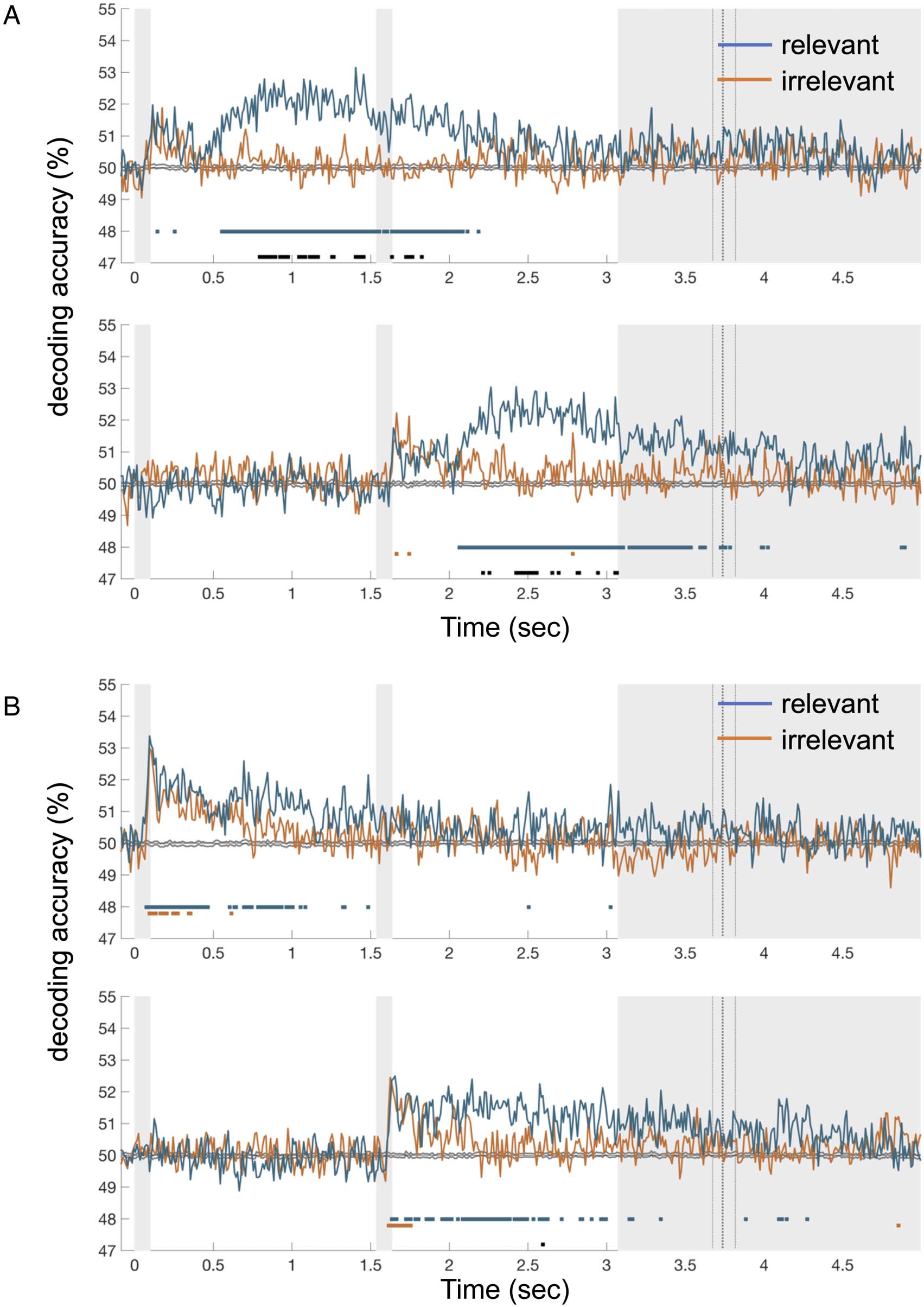
Preferential Coding of Visual Features
Next, we examined the time course with which we could
decode stimulus color and shape from the pattern of
MEG activity. We quantified this separately when a feature
was relevant or irrelevant for the participant’s task so that
we could examine the effect of attention on coding of this
information. We predicted that both relevant and irrelevant
stimulus features would be decodable from the sensor data,
but that each feature would be more readily decoded when
it was relevant compared to when it was irrelevant, partic-
ularly at later time points (Goddard et al., 2022; Moerel
et al., 2021; Hebart et al., 2018). In Experiment 1, robust
decoding of stimulus information emerged rapidly after
the onset of each display, remaining through the initial part
of the delay phase for each epoch (Figure 4). Contrary to
our prediction, however, in Experiment 1, there was no reli-
able evidence of preferential coding of the currently rele-
vant information, in either task epoch, for color or shape
information (Figure 4). We subsequently applied a Bayesian
analysis of preferential coding, comparing evidence for
preferential coding to a point nil, and using a one-sided,
medium width (r = .707) Cauchy prior over the interval
[0 Inf], following Teichmann, Moerel, Baker, and
Grootswagers (2021). This interval favors detection of small
effects, as the bulk of the prior distribution is close to the
null value of 0. This analysis showed strong evidence for the
null at most time points (Bayes factor < .1), for all features.
Few or no time points showed strong evidence (Bayes
factor > 10) in favor of the hypothesis that decoding
accuracy was higher when the feature was task-relevant.
Barnes, Goddard, and Woolgar
813
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
oh
C
norte
_
a
_
0
1
8
3
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 5. Color (A) and shape (B) decoding for Experiment 2. (A) y (B) show decoding traces for the first and second targets in the upper and
lower panels. Decoding accuracies are shown for each feature when it was relevant (azul) or irrelevant (naranja) for the task. Gray bars mark the
stimulus and response display durations. Vertical lines show the median response time, ± one quartile. Times at which decoding was greater than
chance, pag < .05, using a cluster-based correction for multiple comparisons, are marked below each trace in the corresponding color. Times at
which relevant information coding was reliably above coding for the irrelevant target feature (threshold-free cluster correction, p < .05) are
marked in black.
814
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
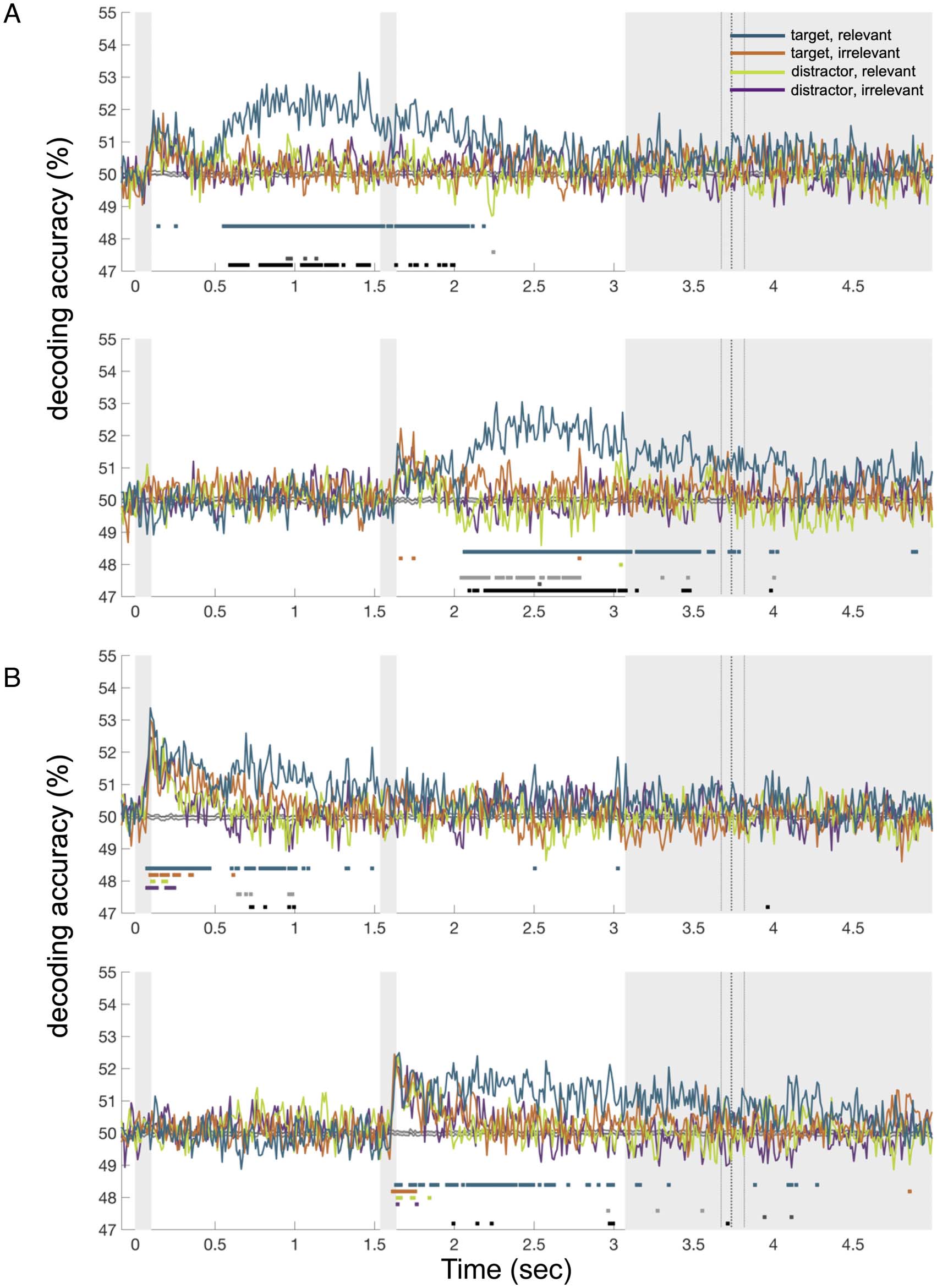
Figure 6. Experiment 2 color (A) and shape (B) decoding for the target and distractor objects on each display. Traces represent decoding accuracy
for colors or shapes at the attended location (blue = relevant feature, orange = irrelevant feature), data repeated from Figure 5, as well as at the
unattended location (green = attended feature, purple = unattended feature). Times at which each trace was reliably different to chance, at p < .05
with a threshold-free cluster correction for multiple comparisons, are marked in the corresponding color. grayscale markers indicate times with a
statistically reliable effect of spatial attention (target vs. distractor, light gray), feature attention (attended vs. unattended feature, dark gray), or
interaction between spatial and feature attention (relevant feature of target vs. all other features, black).
Barnes, Goddard, and Woolgar
815
information in both epochs, although this was statistically
reliable only in the second epoch. A follow-up analysis
revealed no reliable difference between the preferential
coding of color and shape.
As a secondary analysis, we additionally considered cod-
ing of the features of the distractor object. All four traces
(relevant and irrelevant feature of target and distractor)
are shown in Figure 6. Color and shape information was
briefly decodable in all four attention conditions, after
which there was a sustained preferential coding of the rel-
evant target feature compared to the average of all other
features (Figure 6, black lines). Where there were main
effects of spatial or feature-selective attention, they tended
to be accompanied or quickly followed by an interaction of
the two attention types. Moreover, when, in an exploratory
analysis, we directly compared coding of the irrelevant fea-
ture of the target with those of the distractor, or the rele-
vant with irrelevant feature of the distractor, there are no
time points where the difference was significant. This
implies no advantage for the irrelevant information at
the relevant location, or for the relevant information
at the relevant location. This replicates similar findings in
the work of Goddard et al. (2022), in which main effects of
spatial and feature attention emerged briefly before an
interaction showed preferential coding specifically for
the information that participants needed to retain.
Rapid Coding of Features across Epochs
To compare the dynamics of attentional prioritization
across the two epochs, we took the decoding traces for
the target in each epoch of Experiment 2 and aligned them
in time. We anticipated that the effect of attention
(enhancement of relevant information) might develop
later in Epoch 2, which reflected a subtrial shift of attention
when participants had less time to prepare what they
would attend to. However, preferential coding for rele-
vant information in Epoch 2 was comparable to Epoch 1
(Figure 7). We did not observe a main effect of epoch, or
an interaction between epoch and relevance. This does
not rule out the possibility that shifting attention mid-trial
incurs some delay in preferential coding in other circum-
stances, for example, with more difficult tasks or a shorter
within-trial interstimulus interval. However, it demon-
strates that humans can rapidly reconfigure their neural
codes to prioritize coding of a new stimulus dimension
mid-trial, even while holding the previously attended
stimulus information in mind. Commensurate with non-
human primate work, this highlights our capacity to
dynamically code task-relevant information.
DISCUSSION
Understanding how task-sensitive neural codes reconfig-
ure is a key step in tracing how the brain supports adaptive
behavior. Here, we conducted two experiments to ask
whether the brain can rapidly reconfigure neural codes
Figure 7. Color (A) and shape (B) decoding for both epochs
superimposed. Blue and orange color indicate relevant and irrelevant
features, and solid and dotted lines indicate Epochs 1 and 2,
respectively. For each trace, time points that reliably differ from chance
are marked with colored squares (Epoch 1) or diamonds (Epoch 2).
There was no reliable difference between epochs, or interaction
between epoch and relevance.
Experiment 2 stimulus decoding was similarly rapid
(Figure 5). Although less pronounced (potentially
because of the busier displays and more subtle color
and shape differences), initial stimulus decoding peaks
followed a similar time course to Experiment 1. For cod-
ing of color, there was an initial stimulus-driven
response peaking at 100 msec, which was similar when
that information was relevant or irrelevant, and which
occurred for both epochs, although these peaks did
not reach statistical significance. For shape, the pattern
was broadly similar and statistically significant, with an
initial stimulus-driven response at 100 msec from each
display onset. Critically, in contrast to Experiment 1, in
Experiment 2, we now saw evidence of additional, sus-
tained, preferential coding of relevant information.
Whereas decoding for the target’s color remained close
to chance when that feature was irrelevant, coding for
the same information when it was relevant was higher
and sustained (Figure 5). Coding of relevant color informa-
tion was reliably different to chance and to the irrelevant
feature trace from approximately 500 msec after stimulus
presentation and was sustained into the subsequent trial
epoch. We observed the same pattern for shape decod-
ing, with a sustained response only for the relevant
816
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
for relevant stimulus features when what is relevant
changes. In both experiments, participants judged the
shape, then color, or vice versa of two targets presented
in sequence. When shape and color judgments were easy
(Experiment 1), we observed strong coding of all object
information. We found no reliable evidence for preferen-
tial coding of task-relevant features. By contrast, when the
shape and color judgments were difficult and additional
distractors were present (Experiment 2), we did see pref-
erential coding for the relevant feature. Crucially, stronger
coding for the relevant feature occurred in both phases of
the trial, although participants were shifting attention
between features mid-trial.
Tracing this process with MEG allowed us to see the
temporal evolution of preferential coding in the human
brain, showing with millisecond resolution how attention
emerges and redirects. Even with this precise temporal
detail, Experiment 2 demonstrated a remarkably similar
time course for selection of relevant information for the
first and second stimuli. We might expect that preferential
encoding of the relevant feature in the second epoch
would be slower and/or less selective than in the first.
For example, a lag or reduction in selectivity could reflect
residual attention to the feature that was relevant for the
first epoch, or time taken to transition to selective encod-
ing of the second feature. Instead, we did not find any evi-
dence of slower or reduced selectivity in the second
epoch, suggesting that, in this paradigm, reconfiguration
was fast enough for the relevant feature of the second
stimulus to be selected as efficiently as for the first. These
findings indicate that, when adaptive coding is engaged,
task-relevant information is preferentially coded with
remarkable speed even as task demands change within
single trials. This provides possible infrastructure for the
fast, subtrial switching of attentional sets necessary for a
goal-directed behavior (Duncan, 2013).
Although participants successfully performed both
tasks, Experiment 1 did not elicit reliable preferential cod-
ing of relevant over irrelevant stimulus features. Curiously,
both tasks showed strong and sustained representation of
the rule (“attend color, then shape”), although only one
task showed an effect of rule on stimulus coding. Current
explanations of top–down control emphasize both main-
taining task information and enhancing relevant stimulus
information. For example, both rule and relevant stimulus
information can typically be decoded from MD regions
in human fMRI ( Woolgar & Zopf, 2017; Jackson et al.,
2016; Woolgar, Afshar, Williams, & Rich, 2015; Woolgar,
Thompson, Bor, & Duncan, 2011) and from frontal cortex
in nonhuman primate single-unit recordings (Stokes et al.,
2013; Everling, Tinsley, Gaffan, & Duncan, 2006). Disrupt-
ing prefrontal function causes reduction in task-relevant
information coding ( Jackson et al., 2021), and incorrect
rule or stimulus information coding predicts incorrect
behavioral responses ( Woolgar, Dermody, Afshar,
Williams, & Rich, 2019). Moreover, the structure of frontal
stimulus information predicts subsequent occipital
stimulus information as attentional selection of relevant
features emerges (Goddard et al., 2022). In view of these
findings, it is plausible that selection occurs through rule
information that is maintained by domain-general regions,
which in turn selectively enhance relevant stimulus infor-
mation in both domain-general and task-specific regions.
In contrast, in Experiment 1, we observed a dissociation:
clear rule coding, but no evidence of enhanced coding of
the relevant stimulus features, although the rule defined
which stimulus features participants should attend to.
Rule decoding increased after the stimulus displays in
both tasks, particularly in Experiment 2. These increases
could reflect neural responses diverging as participants
applied the feature rule to the stimuli, in a way that did
not enhance coding of the relevant stimulus features to
an extent that our methods could reliably detect. Con-
versely, increases in rule decoding could be related to a
more general shift, such as the widespread reduction in
cortical response variance at the onset of a stimulus
(Churchland et al., 2010). This highlights the utility of
tracing both attentional rule information and rule-related
changes in stimulus information, to characterize the
impact of the rule on attentional selection. As Experiment
1 shows, the presence of decodable attentional rules does
not necessarily translate to preferential coding of relevant
stimulus information.
There were several differences between the two exper-
iments that may have contributed to the different results.
Experiment 2 was more difficult: Participants responded
well above chance level in both tasks, but overall perfor-
mance was lower in Experiment 2 even after intensive
training on the task. In Experiment 1, stimuli were drawn
from a set of four objects, with strongly differentiated
colors and shapes, and a single object was shown on each
display. Because of this small stimulus set, on 25% of trials,
the objects on Display 1 and Display 2 were identical, mak-
ing the task trivial. On the remaining trials, participants
had to select differential information from each display
to respond accurately. However, there was significantly
less information on each display, and less confusability
among colors and shapes, than in Experiment 2. Thus,
responding to the relevant information could well engage
different attentional mechanisms across the two tasks.
Increased selection with increased stimulus complexity
is a common theme in many theories of attention. For
example, behavioral data demonstrate that although par-
ticipants can find and respond to targets more quickly in
simple displays compared to complex displays, they are
also more easily influenced by salient distractors (Lavie,
1995; Lavie & Tsal, 1994). Neuroimaging evidence also
suggests that distractors are not processed as deeply when
a task becomes more difficult: BOLD activity associated
with a distractor stimulus category no longer differentiates
repeating and unrepeating distractors when target visibil-
ity drops (Yi, Woodman, Widders, Marois, & Chun, 2004).
Load theory (Lavie, Beck, & Konstantinou, 2014; Lavie,
1995), takes these findings to argue that selection is
Barnes, Goddard, and Woolgar
817
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
qualitatively different for simple and complex stimuli. In
simple environments, perceptual capacity not spent on
relevant information spills over to other stimuli. As com-
plexity increases, through the number, similarity, or visibil-
ity of the stimuli, we voluntarily direct our fixed capacity
toward relevant features and ignore salient distractors.
Load theory does not strictly specify that all features that
fall within perceptual capacity limits are equally repre-
sented. Based on behavioral responses to distractors
under low load, we might predict that relevant and irrele-
vant features in simple displays are equally encoded, so
that preferential coding only occurs when we exceed our
perceptual capacity. Our differential findings in Experi-
ments 1 and 2 could be consistent with this view, if Exper-
iment 1 displays fell within most participants’ perceptual
capacity while Experiment 2 displays exceeded it. How-
ever, neuroimaging data so far do not support the idea that
we require complex displays to engage preferential cod-
ing. Indeed, multivariate analyses of fMRI data show that
relevant feature coding in visual cortex ( primary visual
area and the lateral occipital complex) can be enhanced
in simple displays, with this enhancement extending to
frontoparietal cortex when stimulus discrimination is diffi-
cult ( Jackson et al., 2016; Woolgar, Williams, et al., 2015).
Recent sensor-space MEG data also show enhanced cod-
ing of the relevant stimulus category (objects or letters)
although the displays contained only two easily distin-
guishable objects (Grootswagers et al., 2021). Based on
these previous results, we would predict that feature-
selective attention produces a relative enhancement of rel-
evant perceptual information in simple displays, although
both relevant and irrelevant information can be perceived
and recalled. This raises an interesting question: If both
simple and complex displays can elicit preferential coding
(that we can detect with both fMRI and MEG), why is stim-
ulus coding in our Experiment 1 unaffected by relevance?
Theories focusing on the object-based nature of atten-
tion (Baldauf & Desimone, 2014; Chen, 2012) may offer a
better explanation for why coding two features of a single
object, as in our Experiment 1, and coding two objects, as
in the work of Grootswagers et al. (2021), would follow dif-
ferent rules. Behavioral studies demonstrate that we can
often report irrelevant features of a target object without
any apparent performance cost, suggesting that all fea-
tures of the object are processed in parallel before we
chose specific elements to respond to (Chen, 2012;
Duncan, 1984). Under this object-based account of atten-
tion, it is unsurprising that we did not observe different
responses to the same visual feature when it was the rele-
vant or irrelevant dimension of a target object. Rather, we
should expect to see preferential coding of the target
object over the distractor. We can see this in the work of
Goddard et al. (2022), in which a spatial attention effect
emerges before coding of the relevant target feature out-
strips all other traces. This same pattern is suggested by
our secondary analyses, where brief main effects of spatial
attention emerge before preferential coding of the relevant
target feature (Figure 6, Epoch 2 color and Epoch 1 shape).
However, object-based accounts struggle to account for
the preferential coding of single dimensions of stimuli
(e.g., Jackson & Woolgar, 2018; Jackson et al., 2016),
which we observed at later time points in Experiment 2.
Biased competition (Reynolds, Chelazzi, & Desimone,
1999; Kastner, Weerd, Desimone, & Ungerleider, 1998;
Desimone & Duncan, 1995) provides a possible unifying
framework for the load-driven and object-based character-
istics of attention. Similar to load theory, this account pro-
poses that complex stimuli trigger attentional selection.
Rather than appealing to a threshold for perceptual capac-
ity, biased competition suggests that, as distinct represen-
tations of stimulus features in early visual cortex feed
forward to shared neural populations in higher visual cor-
tex, competition emerges for what feature will be repre-
sented at the higher level, forcing selection to occur (Scalf,
Torralbo, Tapia, & Beck, 2013; Reynolds, O’Reilly, Cohen,
& Braver, 2012; Desimone & Duncan, 1995). Because inte-
gration co-occurs with broadening receptive fields, even
spatially segregated shapes can project to the same neu-
rons and compete for in-depth processing. In our study,
the two-object displays of difficult-to-discriminate stimuli
in Experiment 2 might elicit more competition than the
single-object displays in Experiment 1, creating the oppor-
tunity for selection, even within the target objects.
Importantly, Duncan (2006) integrates space-, object-,
and feature-based attention under the biased competition
framework, highlighting that competition drives selection
across disparate forms of attention, which can operate
independently or in concert. This broader perspective of
attention as a family of processes implemented through
biased competition has since been embraced by Kravitz
and Behrmann (2011), who demonstrate that space-,
object-, and feature-based attention can combine to
enhance object processing. Combined effects of spatial
and feature-based attention have also been observed in
nonhuman primates’ lateral intraparietal area (Ibos &
Freedman, 2016). Goddard et al. (2022) similarly show
multiplicative effects of spatial and feature-selective atten-
tion give rise to selective coding of only the relevant fea-
ture at the relevant location. Using the same stimuli, we
replicated this finding, showing that coding of the relevant
feature at the relevant location is enhanced relative to the
irrelevant feature at that location (Figure 5) and the rele-
vant and irrelevant features of the distractor (secondary
analyses, Figure 6), whereas there was no advantage for
the irrelevant feature at the relevant location, or relevant
feature at the irrelevant location.
From a broader perspective, each of these theories
incorporates the suggestion that selection processes do
not always alter stimulus representations. In Experiment 1,
we saw that people were able to perform a task that
required selection without visible impact on the repre-
sentation of stimulus features. This was consistent with
the idea that there were enough resources to process
both aspects of those stimuli to a sufficient level before
818
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
choosing what would impact behavior. According to the
theories above, this capacity to process multiple stimulus
features to a high level could depend on the number of
features, on object binding, or on a lack of competition,
each of which could have facilitated neural coding of stim-
uli in Experiment 1. Neural network simulations addition-
ally offer some insight into the cost of selection, showing
that strong coding of currently relevant task features
induces slow reconfiguration to code subsequently rele-
vant information (Musslick, Jang, Shvartsman, Shenhav, &
Cohen, 2018). Therefore, there may be a computational
benefit to avoiding brain-wide reconfiguration of attentional
sets (e.g., within trials) where possible. An adaptive system
may be characterized not only by the ability to flexibly
prioritize processing of currently relevant information, but
the flexibility to only do so when processing demands
require it.
We should highlight that the two experiments in this
study differed in aspects other than the number and com-
plexity of the stimuli. The experiments were coded,
recruited, and run at different testing sites, meaning that
the participants, screens, and scanners were unique to
each. We were careful to control the stimulus parameters
and match the data preprocessing. However, we cannot
rule out the possibility that some property of the partici-
pant group or scanning equipment impacted the results.
In addition, we extended the poststimulus delay periods
in Experiment 2 relative to Experiment 1, to account for
a large increase in task difficulty. This makes it difficult
to directly compare the two tasks. A within-subject study
with matched timings will be important in the future to sta-
tistically compare preferential coding in simple and com-
plex tasks, and narrow down the circumstances in which
patterns do or do not rapidly reconfigure within single
trials.
An interesting question is whether we would see the
same rapid reconfiguration of what information is prefer-
entially encoded in a less stable context. Here, participants
applied the same rule (e.g., “attend color, then shape”)
throughout a block of more than 200 trials, before switch-
ing to a new rule. This has the advantage of allowing
participants to prepare for each trial, enabling us to use
the rapid preferential coding of relevant information in
Epoch 1 as a baseline against which to compare Epoch
2. However, the repeating rule could have more extensive
consequences. An interleaved rule design (e.g., cued
trial-by-trial) could potentially uncover limits to rapid
reconfiguration, for example, if people struggle to quickly
prioritize new information without warning, or are unable
to fully prepare one or both parts of the task in advance. In
addition, it is well established that frontoparietal BOLD
activation is sensitive to difficulty, typically with a U-shaped
function, where activation peaks when tasks are difficult
but not overwhelming ( Van Snellenberg et al., 2015; Jaeggi
et al., 2007; Callicott et al., 1999). Thus, to the extent that
the current results reflect the engagement of this net-
work, it seems likely that the additional challenge of
reconfiguring task sets on each trial would further impact
the results depending where on this function the task
sits. Further empirical work is needed to establish the
extent to which our results generalize to other designs.
Here, we have shown that human adaptive population
codes can reconfigure within a single trial. This supports
the current theory, which emphasizes the potential of
focusing on each step in a task to produce complex and
creative behavior. Surprisingly, where attention effects
were seen, the dynamics were comparable for between-
trials and within-trial shifts of attentional focus. This pro-
vides a potential neural substrate for the rapid creation
of attentional episodes in multipart tasks. However, signif-
icant effects of attention were only obtained in a demand-
ing version of the task. Although many factors differed
between the experiments, the difference could reflect
the inherent cost of reconfiguring attention, meaning that
it is not always an optimal strategy to engage. Future work
will be important to identify what conditions push us
toward preferentially coding the relevant information.
Spatio-temporally resolved methods, such as source-
reconstructed MEG or MEG-fMRI fusion (Moerel et al.,
2021; Mohsenzadeh, Mullin, Lahner, Cichy, & Oliva,
2019; Cichy, Pantazis, & Oliva, 2016), paired with system-
atic manipulation of task difficulty, could further elucidate
how domain-general and task-specific brain regions inter-
act to select relevant information under varying task
demands. Rapid stimulus streams or self-directed atten-
tion shifting could further probe how rapidly the brain
can reconfigure neural codes for preferential processing.
Furthermore, relating the speed of reconfiguration to
measures of fluid ability could clarify the functional impor-
tance of adaptive coding timescales. Together with our
findings, this will offer rich insight into the biological bases
of a mind that adapts to connect our goals with the world
around us.
Reprint requests should be sent to Lydia Barnes, MRC Cog-
nition and Brain Sciences Unit, University of Cambridge, 15
Chaucer Road, Cambridge, CB2 7EF UK, or via e-mail:
lydiabarnes01@gmail.com.
Funding Information
Medical Research Council (https://dx.doi.org/10.13039
/501100000265), grant number: SUAG/052/G101400.
Macquarie University (https://dx.doi.org/10.13039
/501100001230), grant number: Research Training Path-
way Scholarship. Australian Research Council (https://dx
.doi.org/10.13039/501100000923), grant numbers:
DE200100139, FT170100105.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
Barnes, Goddard, and Woolgar
819
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .549, W/M = .137,
M/ W = .059, and W/ W = .255.
REFERENCES
Assem, M., Glasser, M. F., Van Essen, D. C., & Duncan, J. (2020).
A domain-general cognitive core defined in multimodally
parcellated human cortex. Cerebral Cortex, 30, 4361–4380.
https://doi.org/10.1093/cercor/bhaa023, PubMed: 32244253
Bae, G.-Y., & Luck, S. J. (2019). Decoding motion direction
using the topography of sustained ERPs and alpha
oscillations. Neuroimage, 184, 242–255. https://doi.org/10
.1016/j.neuroimage.2018.09.029, PubMed: 30223063
Baldauf, D., & Desimone, R. (2014). Neural mechanisms of
object-based attention. Science, 344, 424–427. https://doi.org
/10.1126/science.1247003, PubMed: 24763592
Battistoni, E., Kaiser, D., Hickey, C., & Peelen, M. V. (2020). The
time course of spatial attention during naturalistic visual
search. Cortex, 122, 225–234. https://doi.org/10.1016/j.cortex
.2018.11.018, PubMed: 30563703
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163
/156856897X00357, PubMed: 9176952
Callicott, J. H., Mattay, V. S., Bertolino, A., Finn, K., Coppola, R.,
Frank, J. A., et al. (1999). Physiological characteristics of
capacity constraints in working memory as revealed by
functional MRI. Cerebral Cortex, 9, 20–26. https://doi.org/10
.1093/cercor/9.1.20, PubMed: 10022492
Chen, Z. (2012). Object-based attention: A tutorial
review. Attention, Perception, & Psychophysics,
74, 784–802. https://doi.org/10.3758/s13414-012-0322-z,
PubMed: 22673856
Churchland, M. M., Yu, B. M., Cunningham, J. P., Sugrue, L. P.,
Cohen, M. R., Corrado, G. S., et al. (2010). Stimulus onset
quenches neural variability: A widespread cortical
phenomenon. Nature Neuroscience, 13, 369–378. https://doi
.org/10.1038/nn.2501, PubMed: 20173745
Cichy, R. M., Pantazis, D., & Oliva, A. (2016). Similarity-based
fusion of MEG and fMRI reveals spatio-temporal dynamics in
human cortex during visual object recognition. Cerebral
Cortex, 26, 3563–3579. https://doi.org/10.1093/cercor
/bhw135, PubMed: 27235099
Cole, M. W., Reynolds, J. R., Power, J. D., Repovs, G., Anticevic,
A., & Braver, T. S. (2013). Multi-task connectivity reveals
flexible hubs for adaptive task control. Nature Neuroscience,
16, 1348–1355. https://doi.org/10.1038/nn.3470, PubMed:
23892552
Crittenden, B. M., & Duncan, J. (2014). Task difficulty
manipulation reveals multiple demand activity but no frontal
lobe hierarchy. Cerebral Cortex, 24, 532–540. https://doi.org
/10.1093/cercor/bhs333, PubMed: 23131804
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222. https://doi.org/10.1146/annurev.ne.18.030195
.001205, PubMed: 7605061
Duncan, J. (1984). Selective attention and the organization of
visual information. Journal of Experimental Psychology:
General, 113, 501–517. https://doi.org/10.1037/0096-3445
.113.4.501, PubMed: 6240521
Duncan, J. (2006). EPS mid-career award 2004: Brain
mechanisms of attention. Quarterly Journal of Experimental
Psychology, 59, 2–27. https://doi.org/10.1080
/17470210500260674, PubMed: 16556554
Duncan, J. (2010). The multiple-demand (MD) system of the
primate brain: Mental programs for intelligent behaviour.
Trends in Cognitive Sciences, 14, 172–179. https://doi.org/10
.1016/j.tics.2010.01.004, PubMed: 20171926
Duncan, J. (2013). The structure of cognition: Attentional
episodes in mind and brain. Neuron, 80, 35–50. https://doi
.org/10.1016/j.neuron.2013.09.015, PubMed: 24094101
Duncan, J., Chylinski, D., Mitchell, D. J., & Bhandari, A. (2017).
Complexity and compositionality in fluid intelligence.
Proceedings of the National Academy of Sciences, U.S.A.,
114, 5295–5299. https://doi.org/10.1073/pnas.1621147114,
PubMed: 28461462
Duncan, J., Schramm, M., Thompson, R., & Dumontheil, I.
(2012). Task rules, working memory, and fluid intelligence.
Psychonomic Bulletin & Review, 19, 864–870. https://doi.org
/10.3758/s13423-012-0225-y, PubMed: 22806448
Everling, S., Tinsley, C. J., Gaffan, D., & Duncan, J. (2006).
Selective representation of task-relevant objects and locations
in the monkey prefrontal cortex. European Journal of
Neuroscience, 23, 2197–2214. https://doi.org/10.1111/j.1460
-9568.2006.04736.x, PubMed: 16630066
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad
domain generality in focal regions of frontal and parietal
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 110, 16616–16621. https://doi.org/10.1073/pnas
.1315235110, PubMed: 24062451
Fusi, S., Miller, E. K., & Rigotti, M. (2016). Why neurons mix:
High dimensionality for higher cognition. Current Opinion
in Neurobiology, 37, 66–74. https://doi.org/10.1016/j.conb
.2016.01.010, PubMed: 26851755
Goddard, E., Carlson, T. A., & Woolgar, A. (2022). Spatial and
feature-selective attention have distinct, interacting effects on
population-level tuning. Journal of Cognitive Neuroscience,
34, 290–312. https://doi.org/10.1162/jocn_a_01796, PubMed:
34813647
Grootswagers, T., Robinson, A. K., Shatek, S. M., & Carlson,
T. A. (2021). The neural dynamics underlying prioritisation of
task-relevant information. Neurons, Behavior, Data Analysis,
and Theory, 5, 1–17. https://doi.org/10.51628/001c.21174
Grootswagers, T., Wardle, S. G., & Carlson, T. A. (2016).
Decoding dynamic brain patterns from evoked responses: A
tutorial on multivariate pattern analysis applied to time series
neuroimaging data. Journal of Cognitive Neuroscience, 29,
677–697. https://doi.org/10.1162/jocn_a_01068, PubMed:
27779910
Hebart, M. N., Bankson, B. B., Harel, A., Baker, C. I., & Cichy,
R. M. (2018). The representational dynamics of task and
object processing in humans. eLife, 7, e32816. https://doi.org
/10.7554/eLife.32816, PubMed: 29384473
Ibos, G., & Freedman, D. J. (2016). Interaction between spatial
and feature attention in posterior parietal cortex. Neuron, 91,
931–943. https://doi.org/10.1016/j.neuron.2016.07.025,
PubMed: 27499082
Jackson, J., Feredoes, E., Rich, A. N., Lindner, M., & Woolgar, A.
(2021). Concurrent neuroimaging and neurostimulation
reveals a causal role for dlPFC in coding of task-relevant
820
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
information. Communications Biology, 4, 1–16. https://doi
.org/10.1038/s42003-021-02109-x, PubMed: 34002006
Jackson, J., Rich, A. N., Williams, M. A., & Woolgar, A. (2016).
Feature-selective attention in frontoparietal cortex:
Multivoxel codes adjust to prioritize task-relevant
information. Journal of Cognitive Neuroscience, 29,
310–321. https://doi.org/10.1162/jocn_a_01039, PubMed:
27626230
Jackson, J., & Woolgar, A. (2018). Adaptive coding in
the human brain: Distinct object features are encoded by
overlapping voxels in frontoparietal cortex. Cortex, 108,
25–34. https://doi.org/10.1016/j.cortex.2018.07.006, PubMed:
30121000
Jaeggi, S. M., Buschkuehl, M., Etienne, A., Ozdoba, C., Perrig,
W. J., & Nirkko, A. C. (2007). On how high performers keep
cool brains in situations of cognitive overload. Cognitive,
Affective, & Behavioral Neuroscience, 7, 75–89. https://doi
.org/10.3758/CABN.7.2.75, PubMed: 17672380
Kado, H., Higuchi, M., Shimogawara, M., Haruta, Y., Adachi, Y.,
Kawai, J., et al. (1999). Magnetoencephalogram systems
developed at KIT. IEEE Transactions on Applied
Superconductivity, 9, 4057–4062. https://doi.org/10.1109/77
.783918
Kastner, S., Weerd, P. D., Desimone, R., & Ungerleider, L. G.
(1998). Mechanisms of directed attention in the human
extrastriate cortex as revealed by functional MRI. Science,
282, 108–111. https://doi.org/10.1126/science.282.5386.108,
PubMed: 9756472
Kleiner, M., Brainard, D. H., Pelli, D. G., Ingling, A., Murray, R.,
& Broussard, C. (2007). What’s new in psychtoolbox-3.
Perception, 36, 1–16.
Kravitz, D. J., & Behrmann, M. (2011). Space-, object-, and
feature-based attention interact to organize visual scenes.
Attention, Perception, & Psychophysics, 73, 2434–2447.
https://doi.org/10.3758/s13414-011-0201-z, PubMed:
22006523
Lavie, N. (1995). Perceptual load as a necessary condition for
selective attention. Journal of Experimental Psychology, 21,
451–468. https://doi.org/10.1037/0096-1523.21.3.451,
PubMed: 7790827
Lavie, N., Beck, D. M., & Konstantinou, N. (2014). Blinded by
the load: Attention, awareness and the role of perceptual
load. Philosophical Transactions of the Royal Society of
London, Series B: Biological Sciences, 369, 20130205. https://
doi.org/10.1098/rstb.2013.0205, PubMed: 24639578
Lavie, N., & Tsal, Y. (1994). Perceptual load as a major
determinant of the locus of selection in visual attention.
Perception & Psychophysics, 56, 183–197. https://doi.org/10
.3758/BF03213897, PubMed: 7971119
Maxfilter (2.2). (2010). [Computer software]. Elekta Neuromag.
Mensen, A., & Khatami, R. (2013). Advanced EEG analysis using
threshold-free cluster-enhancement and non-parametric
statistics. Neuroimage, 67, 111–118. https://doi.org/10.1016/j
.neuroimage.2012.10.027, PubMed: 23123297
Moerel, D., Rich, A. N., & Woolgar, A. (2021). Selective
attention and decision-making have separable neural bases in
space and time. BioRxiv. https://doi.org/10.1101/2021.02.28
.433294
Mohsenzadeh, Y., Mullin, C., Lahner, B., Cichy, R. M., & Oliva, A.
(2019). Reliability and generalizability of similarity-based
fusion of MEG and fMRI data in human ventral and dorsal
visual streams. Vision, 3, 8. https://doi.org/10.3390
/vision3010008, PubMed: 31735809
Musslick, S., Jang, J. S., Shvartsman, M., Shenhav, A., & Cohen,
J. D. (2018). Constraints associated with cognitive control
and the stability-flexibility dilemma. In Proceedings of the
40th Annual Meeting of the Cognitive Science Society
(pp. 806–811). Madison, WI.
Noble, S., Scheinost, D., & Constable, R. T. (2020). Cluster
failure or power failure? Evaluating sensitivity in
cluster-level inference. Neuroimage, 209, 116468.
https://doi.org/10.1016/j.neuroimage.2019.116468, PubMed:
31852625
O’Brien, S., Mitchell, D. J., Duncan, J., & Holmes, J. (2020).
Cognitive segmentation and fluid reasoning in childhood.
PsyArXiv. https://doi.org/10.31234/osf.io/dt84m
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis of
MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
Oosterhof, N. N., Connolly, A. C., & Haxby, J. V. (2016).
CoSMoMVPA: Multi-modal multivariate pattern analysis of
neuroimaging data in MATLAB/GNU octave. Frontiers in
Neuroinformatics, 10, 27. https://doi.org/10.3389/fninf.2016
.00027, PubMed: 27499741
Op de Beeck, H. P., Baker, C. I., DiCarlo, J. J., & Kanwisher,
N. G. (2006). Discrimination training alters object
representations in human extrastriate cortex. Journal of
Neuroscience, 26, 13025–13036. https://doi.org/10.1523
/JNEUROSCI.2481-06.2006, PubMed: 17167092
Pernet, C. R., Latinus, M., Nichols, T. E., & Rousselet, G. A.
(2015). Cluster-based computational methods for mass
univariate analyses of event-related brain potentials/fields: A
simulation study. Journal of Neuroscience Methods, 250,
85–93. https://doi.org/10.1016/j.jneumeth.2014.08.003,
PubMed: 25128255
Rao, S. C., Rainer, G., & Miller, E. K. (1997). Integration of what
and where in the primate prefrontal cortex. Science, 276,
821–824. https://doi.org/10.1126/science.276.5313.821,
PubMed: 9115211
Reynolds, J. H., Chelazzi, L., & Desimone, R. (1999).
Competitive mechanisms subserve attention in macaque
areas V2 and V4. Journal of Neuroscience, 19, 1736–1753.
https://doi.org/10.1523/JNEUROSCI.19-05-01736.1999,
PubMed: 10024360
Reynolds, J. R., O’Reilly, R. C., Cohen, J. D., & Braver, T. S.
(2012). The function and organization of lateral prefrontal
cortex: A test of competing hypotheses. PLoS One, 7, e30284.
https://doi.org/10.1371/journal.pone.0030284, PubMed:
22355309
Rigotti, M., Barak, O., Warden, M. R., Wang, X.-J., Daw, N. D.,
Miller, E. K., et al. (2013). The importance of mixed selectivity
in complex cognitive tasks. Nature, 497, 585–590. https://doi
.org/10.1038/nature12160, PubMed: 23685452
Scalf, P. E., Torralbo, A., Tapia, E., & Beck, D. M. (2013).
Competition explains limited attention and perceptual
resources: Implications for perceptual load and dilution
theories. Frontiers in Psychology, 4, 243. https://doi.org/10
.3389/fpsyg.2013.00243, PubMed: 23717289
Smith, S. M., & Nichols, T. E. (2009). Threshold-free cluster
enhancement: Addressing problems of smoothing, threshold
dependence and localisation in cluster inference.
Neuroimage, 44, 83–98. https://doi.org/10.1016/j.neuroimage
.2008.03.061, PubMed: 18501637
Spaak, E., Watanabe, K., Funahashi, S., & Stokes, M. G. (2017).
Stable and dynamic coding for working memory in primate
prefrontal cortex. Journal of Neuroscience, 37, 6503–6516.
https://doi.org/10.1523/JNEUROSCI.3364-16.2017, PubMed:
28559375
Stelzer, J., Chen, Y., & Turner, R. (2013). Statistical inference
and multiple testing correction in classification-based
multi-voxel pattern analysis (MVPA): Random permutations
and cluster size control. Neuroimage, 65, 69–82. https://doi
.org/10.1016/j.neuroimage.2012.09.063, PubMed: 23041526
Barnes, Goddard, and Woolgar
821
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Stokes, M. G., Kusunoki, M., Sigala, N., Nili, H., Gaffan, D., &
Duncan, J. (2013). Dynamic coding for cognitive control in
prefrontal cortex. Neuron, 78, 364–375. https://doi.org/10
.1016/j.neuron.2013.01.039, PubMed: 23562541
Teichmann, L., Moerel, D., Baker, C., & Grootswagers, T.
(2021). An empirically-driven guide on using Bayes factors for
M/EEG decoding. BioRxiv. https://doi.org/10.1101/2021.06.23
.449663
Uehara, G., Adachi, Y., Kawai, J., Shimogawara, M., Higuchi, M.,
Haruta, Y., et al. (2003). Multi-channel SQUID systems for
biomagnetic measurement. IEICE Transactions on
Electronics, E86-C, 43–54.
Van Snellenberg, J., Slifstein, M., Read, C., Weber, J., Thompson,
J., Wager, T., et al. (2015). Dynamic shifts in brain network
activation during supracapacity working memory task
performance. Human Brain Mapping, 36, 1245–1264.
https://doi.org/10.1002/hbm.22699, PubMed: 25422039
Vastano, R., Ambrosini, E., Ulloa, J. L., & Brass, M. (2020). Action
selection conflict and intentional binding: An ERP study.
Cortex, 126, 182–199. https://doi.org/10.1016/j.cortex.2020.01
.013, PubMed: 32088407
Wen, T., Duncan, J., & Mitchell, D. J. (2019). The time-course of
component processes of selective attention. Neuroimage,
199, 396–407. https://doi.org/10.1016/j.neuroimage.2019.05
.067, PubMed: 31150787
Woolgar, A., Afshar, S., Williams, M. A., & Rich, A. N. (2015).
Flexible coding of task rules in frontoparietal cortex: An
adaptive system for flexible cognitive control. Journal of
Cognitive Neuroscience, 27, 1895–1911. https://doi.org/10
.1162/jocn_a_00827, PubMed: 26058604
Woolgar, A., Dermody, N., Afshar, S., Williams, M. A., & Rich,
A. N. (2019). Meaningful patterns of information in the brain
revealed through analysis of errors. BioRxiv. https://doi.org
/10.1101/673681
Woolgar, A., Duncan, J., Manes, F., & Fedorenko, E. (2018).
Fluid intelligence is supported by the multiple-demand
system not the language system. Nature Human Behaviour,
2, 200–204. https://doi.org/10.1038/s41562-017-0282-3,
PubMed: 31620646
Woolgar, A., Hampshire, A., Thompson, R., & Duncan, J. (2011).
Adaptive coding of task-relevant information in human
frontoparietal cortex. Journal of Neuroscience, 31,
14592–14599. https://doi.org/10.1523/JNEUROSCI.2616-11
.2011, PubMed: 21994375
Woolgar, A., Jackson, J., & Duncan, J. (2016). Coding of visual,
auditory, rule, and response information in the brain: 10
Years of multivoxel pattern analysis. Journal of Cognitive
Neuroscience, 28, 1433–1454. https://doi.org/10.1162/jocn_a
_00981, PubMed: 27315269
Woolgar, A., Parr, A., Cusack, R., Thompson, R., Nimmo-Smith,
I., Torralva, T., et al. (2010). Fluid intelligence loss linked to
restricted regions of damage within frontal and parietal
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 107, 14899–14902. https://doi.org/10.1073/pnas
.1007928107, PubMed: 20679241
Woolgar, A., Thompson, R., Bor, D., & Duncan, J. (2011).
Multi-voxel coding of stimuli, rules, and responses in human
frontoparietal cortex. Neuroimage, 56, 744–752. https://doi
.org/10.1016/j.neuroimage.2010.04.035, PubMed: 20406690
Woolgar, A., Williams, M. A., & Rich, A. N. (2015). Attention
enhances multi-voxel representation of novel objects in
frontal, parietal and visual cortices. Neuroimage, 109,
429–437. https://doi.org/10.1016/j.neuroimage.2014.12.083,
PubMed: 25583612
Woolgar, A., & Zopf, R. (2017). Multisensory coding in the
multiple-demand regions: Vibrotactile task information is
coded in frontoparietal cortex. Journal of Neurophysiology,
118, 703–716. https://doi.org/10.1152/jn.00559.2016,
PubMed: 28404826
Yi, D.-J., Woodman, G. F., Widders, D., Marois, R., & Chun,
M. M. (2004). Neural fate of ignored stimuli: Dissociable
effects of perceptual and working memory load. Nature
Neuroscience, 7, 992–996. https://doi.org/10.1038/nn1294,
PubMed: 15286791
Yip, H. M. K., Cheung, L. Y. T., Ngan, V. S. H., Wong, Y. K., &
Wong, A. C.-N. (2021). The effect of task on object processing
revealed by EEG decoding. BioRxiv, https://doi.org/10.1101
/2020.08.18.255018
822
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
0
6
2
0
0
4
6
4
9
/
j
o
c
n
_
a
_
0
1
8
3
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3