Métodos
Bridging global and local topology in whole-brain
networks using the network statistic jackknife
Teague R. Henry
1, Kelly A. Duffy1, Marc D. Rudolph1, Mary Beth Nebel2,3,
Stewart H. Mostofsky2,3,4, and Jessica R. Cohen1
1Department of Psychology and Neuroscience, University of North Carolina at Chapel Hill, Chapel Hill, CAROLINA DEL NORTE, EE.UU
2Center for Neurodevelopmental and Imaging Research, Kennedy Krieger Institute, baltimore, Maryland, EE.UU
3Department of Neurology, Johns Hopkins School of Medicine, baltimore, Maryland, EE.UU
4Department of Psychiatry and Behavioral Science, Johns Hopkins School of Medicine, baltimore, Maryland, EE.UU
Palabras clave: Red, Estadísticas, resonancia magnética funcional, Conectividad funcional, Graph theory, Whole-brain
análisis, Jackknife
un acceso abierto
diario
ABSTRACTO
Whole-brain network analysis is commonly used to investigate the topology of the brain
using a variety of neuroimaging modalities. This approach is notable for its applicability to a
large number of domains, such as understanding how brain network organization relates to
cognition and behavior and examining disrupted brain network organization in disease. A
benefit to this approach is the ability to summarize overall brain network organization with a
single metric (p.ej., global efficiency). Sin embargo, important local differences in network
structure might exist without any corresponding observable differences in global topology,
making a whole-brain analysis strategy unlikely to detect relevant local findings. En cambio,
using local network metrics can identify local differences, but are not directly informative of
differences in global topology. Aquí, we propose the network statistic (NS) jackknife
estructura, a simulated lesioning method that combines the utility of global network analysis
strategies with the ability to detect relevant local differences in network structure. Nosotros
evaluate the NS jackknife framework with a simulation study and an empirical example
comparing global efficiency in children with attention-deficit/hyperactivity disorder (TDAH)
and typically developing (TD) niños. The NS jackknife framework has been implemented
in a public, open-source R package, netjack, available at
https://cran.r-project.org/package=netjack.
INTRODUCCIÓN
Describing the brain as a network, an approach termed network neuroscience (bassett &
despreciar, 2017), is a powerful method that allows regions distributed across the entire brain
to be incorporated into a single model describing overall brain topology. This method has pro-
vided insights into whole-brain disruption in neurological and psychiatric diseases (Proporcionó,
bullmore, & Brilla, 2017; Proporcionó, Brilla, & romper la lanza, 2015), the dynamic processes
underlying cognition (cohen & D'Esposito, 2016; Verano, Tambini, Sadaghiani, Keilholz, &
cohen, 2018), and changes across development (Baum et al., 2017; Grayson & Fair, 2017).
Researchers using a network neuroscience framework can draw upon the rich methodological
and statistical literature in graph theory and network analysis, grounding the approach in a
well-established mathematical framework.
Sin embargo, whole-brain analysis methods, as commonly applied in network neuroscience,
often fail at what Hallquist and Hillary (Hallquist & Hillary, 2018) refer to as “matching theory
Citación: Henry, t. r., Duffy, k. A.,
Rudolph, METRO. D., Nebel, METRO. B.,
Mostofsky, S. h., & cohen, j. R. (2020).
Bridging global and local topology in
whole-brain networks using the
network statistic jackknife. Red
Neurociencia, 4(1), 70–88. https://
doi.org/10.1162/netn_a_00109
DOI:
https://doi.org/10.1162/netn_a_00109
Supporting Information:
https://doi.org/10.1162/netn_a_00109
https://CRAN.R-project.org/
package=netjack
Recibió: 4 Junio 2019
Aceptado: 12 Septiembre 2019
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Teague Rhine Henry
trhenry@email.unc.edu
Editor de manejo:
Richard Betzel
Derechos de autor: © 2019
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Jackknife:
A nonparametric statistical technique
that involves repeatedly reestimating
a parameter value after the removal
of single observations.
Whole-brain (global)
network statistic:
A statistic derived from a whole-brain
network that describes an aspect of
global topology.
Global topology:
The overall shape and structure of a
network.
to scale.” This refers to the fact that whole-brain network analysis methods operate at a global
resolution, while relevant differences might be very localized. En otras palabras, there may be
differences at a local scale (es decir., within a single subnetwork) that would be missed when cal-
culating a single, whole-brain summary measure that describes global topology. En cambio,
observed global differences in topology could be driven by a specific local difference (es decir., un
observed overall difference in network integration could be driven by a difference limited to
a single subnetwork). This mismatch in resolution, por lo tanto, could obscure meaningful differ-
ences in network structure. While local network statistics, such as participation coefficient of
a single node, can be used to identify local differences of interest, these local statistics do not
directly assess a localized difference in global topology, and as such are not ideal for matching
a global theory to a local scale. In the present manuscript, we develop the network statistic
(NS) jackknife as a general method to localize global network statistics across a variety of
network metrics and brain features of interest, thus allowing researchers to better address this
resolution issue.
The NS jackknife method (available for use at https://cran.r-project.org/package=netjack)
proposed here allows researchers to identify specific brain regions, connections, or subnet-
obras (es decir., local features) that drive global network analysis results or, conversely, that are not
detected by global network analysis. By doing so, our method can detect local differences in
network structure using the same summary metrics that a global analysis would use, si arriba-
posed to relying on local metrics that do not explicitly link to the global metrics in question.
This approach bridges the gap between a global and a local scope of analysis by allowing
researchers to apply global analysis strategies in a way that reveals differences at local scopes,
even when those differences would not be detected with the global analysis method. We eval-
uate the performance of the NS jackknife with a simulation study and demonstrate the utility
of this method with an empirical example comparing resting-state brain network organization
in children with attention-deficit/hyperactivity disorder (TDAH) and age-matched typically
developing (TD) niños.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Global Versus Local Topology
There are many network statistics that can be used to analyze the global topology of a network;
for simplicity we focus on two widely used network statistics as examples. Primero, we use mod-
ularity, or a measure of a network’s segregation into multiple communities (or subnetworks;
Hombre nuevo, 2006). Segundo, we use global efficiency, a statistic that assesses the overall inte-
gration of a network (latora & marchiori, 2001). These two statistics have led to great insight
regarding brain organization, in particular for supporting the hypothesis that the balance be-
tween network segregation (dense, within-subnetwork connectivity) and network integration
(communication across distinct subnetworks) is critical for cognition (for reviews and canon-
ical articles, see the following: cohen & D'Esposito, 2016; Brillar & Poldrack, 2017; despreciar,
2013; despreciar, Chialvo, Kaiser, & Hilgetag, 2004).
Sin embargo, a key issue for whole-brain analysis is its poor detection of local differences in
topología. Por ejemplo, if a whole-brain analysis produces differences in overall network topol-
ogy between groups or conditions, it does not provide insight into where in the brain these
differences might arise. Más, if a whole-brain analysis produces no differences in overall
network topology, that does not preclude the existence of important local differences. Este
property of whole-brain network analysis is less than ideal as it ignores local features, cual
can include specific brain regions, specific connections between brain regions, or specific
subnetworks. A local topological difference arising from one region/connection/subnetwork
Neurociencia en red
71
The network statistic jackknife
Specificity:
When differences in global topology
are being driven by specific
differences in local topology, como
opposed to globally distributed
diferencias.
Equifinality:
When different network
configurations can lead to the same
global network statistic.
versus another could have vastly different implications for empirical inferences and resultant
clinical applications.
There are several existing graph theory metrics that operate on a local scale. Para examen-
por ejemplo, within-module degree indicates which nodes are more central to their communities, y
participation coefficient indicates which nodes connect different communities (Guimerà &
Nunes Amaral, 2005). Además, there are a variety of statistical models for identifying
nodes and other network features of importance. One of the most widely used is the network-
based statistic method (NBS; Brilla, Proporcionó, & bullmore, 2010), which identifies differences
between groups by examining maximally different sets of edges. The degree-based statistic
method of Yoo and colleagues (Yoo et al., 2017) is more focused than that of the NBS method,
and identifies highly central nodes that are related to a hypothesis of interest. Another recent
acercarse, that of screen filtering (Meskaldji et al., 2015), allows researchers to analyze group
differences in nodal and edge-wise metrics; sin embargo, this method as currently implemented
is restricted to nodal, edge-wise or subnetwork statistics as it relies on correcting a collection
of statistical tests. All of these methods allow researchers to assess local differences between
groups and thus are powerful tools for understanding different features of network organi-
zación, but none of these methods assess local drivers of global topological properties of a
network. Por ejemplo, a common method for examining group differences in local modular
structure is examining differences in participation coefficient for individual nodes or subsets
de nodos. While nodes that exhibit a participation coefficient difference might correspond to
nodes that are relevant to differences in global modularity, this is by no means assured. Local
metrics and tests can be related to global metrics (es decir., participation coefficient can be related
to whole-brain modularity), but these relations are not one-to-one, and a difference in local
properties does not necessarily imply a difference in global properties. Similarmente, previous sta-
tistical models that identify network elements of importance do not identify these elements
based on their relative contribution to global topology, but rather based on their impacts on
local topology. These properties of local metrics and existing local statistical models suggest
a need for a framework that can assess differences in global topology and directly link these
global differences to local structures.
The disconnect that arises between the global and the local scope can be due to specificity,
in which whole-brain network results are driven by specific connections, regiones, or subnet-
obras; or to equifinality, in which strikingly different network configurations can lead to the
same whole-brain summary metric values. These two concepts can be related, as equifinality
can be caused by specific differences in local topology that do not lead to differences in sum-
mary metrics describing global topology. The NS jackknife, proposed here, provides a solution
to these issues. This statistical approach allows researchers to simultaneously analyze differ-
ences in global topology and localize these differences to specific brain regions, connections,
or subnetworks.
Network Specificity and Equifinality
Specificity is the network principle whereby differences in observed whole-brain or whole-
network topology can be driven by specific differences in local topology, as opposed to glob-
ally distributed differences. A whole-brain network difference can be specific if, Por ejemplo,
one group of interest has greater connectivity within one specific functional subnetwork than
a different group, and this increase in connectivity is substantial enough that it alone drives a
significant difference in global modularity. En cambio, a whole-brain network difference can
be nonspecific if the difference in topology is due to a diffuse pattern of local differences.
Neurociencia en red
72
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Por ejemplo, if one group has higher connectivity between all pairs of subnetworks, this can
result in a difference in global efficiency that cannot be localized to any given subset of net-
obras. One key issue with whole-brain network analysis is that it is not capable of determining
whether an observed topological difference is specific or nonspecific. This property has been
termed the theory-to-scale issue (Hallquist & Hillary, 2018).
This lack of specificity is complicated by the fact that, when calculating various summary
statistics for brain networks, typical approaches in network neuroscience can produce the
same network statistics for different network configurations. Equifinality, a term borrowed from
system theory (Bertalanffy, 1950), is the network principle whereby the same result can be
achieved by many different means. Aquí, we use it to emphasize the fact that the same in-
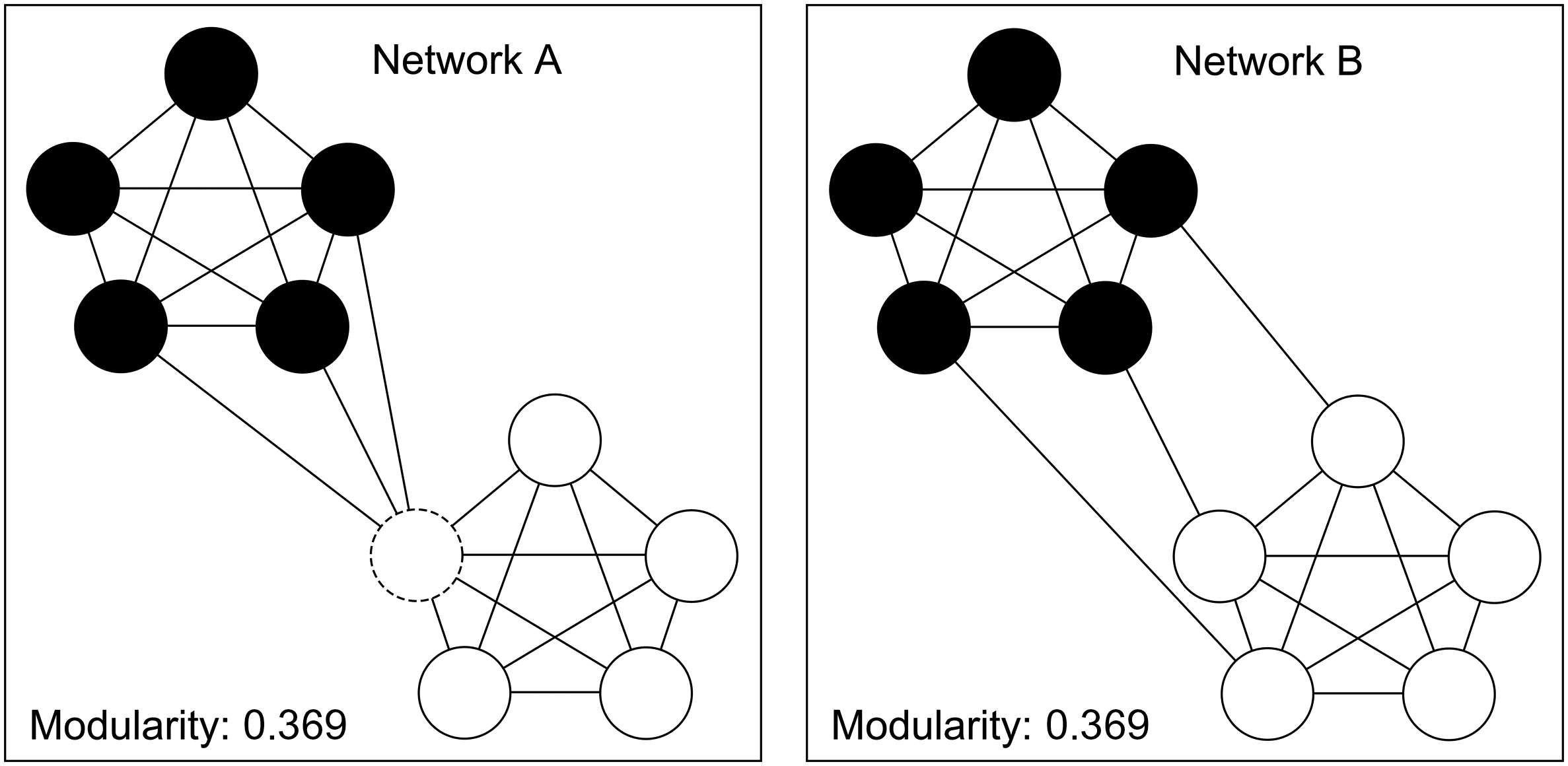
ference can result from different global network configurations. Por ejemplo, Cifra 1 muestra
two networks, each consisting of two subnetworks. The difference between these networks is
in how these subnetworks are connected. In Network A, the communities are connected via a
hub node (dotted circle), while for Network B the subnetworks are connected more diffusely.
Whole-network modularity of the two networks in Figure 1 is identical, yet this result masks
the fact that the local structure of the inter-subnetwork connections in the two networks is quite
diferente. This demonstrates the possible disconnect between whole-brain findings (es decir., global
alcance) and specific local differences in topology (es decir., local scope).
Specific versus nonspecific whole-brain findings occur with regularity in neuroimaging re-
buscar. Por ejemplo, individuals suffering from Alzheimer’s disease exhibit reductions in de-
fault mode network connectivity as well as global loss of a small-world network structure
(Pievani, de Haan, Wu, Seeley, & Frisoni, 2011). This analysis of whole-brain small-world
structure differentiates patients with Alzheimer’s disease from healthy control participants but
provides no information as to the role default mode network connectivity plays in that global
finding. The NS jackknife method has the ability to quantify whether reductions of whole-brain
small-world structure in Alzheimer’s disease are driven by the default mode network alone, son
widely distributed across all subnetworks in the brain, or are driven by a small number of sub-
redes.
Cifra 1. Two networks with the same value of modularity yet very different inter-subnetwork
patterns of connectivity. Note that Network A contains a single connector hub, or a node that links
two communities (dashed circle), while Network B does not.
Neurociencia en red
73
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Virtual/simulated lesioning:
A technique in neuroimaging that
removes components of a network
during analysis to examine the
impact of those components on
global topology.
Network robustness:
How much a network’s global
topology changes when nodes/edges
are removed. This removal is
typically done in a random or
semirandom fashion to assess the
network’s robustness to attack or
damage.
These specific local differences with prominent global differences found in Alzheimer’s
disease can be contrasted with the finding that individuals with autism spectrum disorder
(ASD) tend to exhibit increased whole-brain integration, as assessed with global efficiency,
as compared with healthy control participants (Luis, Theilmann, Townsend, & evans, 2013;
Rudie & Dapretto, 2013). While other research has shown specific differences between indi-
viduals with ASD and healthy control participants, for example within the salience network
(Di Martino et al., 2009), the finding of increased functional integration appears be a result of
more diffuse and weaker functional connections distributed across the entire brain, and thus a
whole-brain phenomenon (Roine et al., 2015; Rudie & Dapretto, 2013). While a whole-brain
network analysis would accurately detect this group difference, it would give no indication
whether the difference was due to a widely distributed whole-brain difference or driven by
specific subnetworks or connections, a conclusion that could be made with use of the NS jack-
knife. These examples emphasize that whole-brain network analysis strategies are designed to
examine a global scope but lack the ability to determine the specificity of any given finding
(Hallquist & Hillary, 2018).
Localization of Whole-Brain Analysis Using the NS Jackknife
The NS jackknife localizes whole-brain network analyses by iteratively removing elements of
the network, such as edges, nodos, or subnetworks, and recalculating the target network statis-
tic without those elements. By sweeping through the whole network, as well as assessing the
impact of the removal across subjects, the NS jackknife creates an empirical distribution of
network element effects, allowing a researcher to localize specific features of brain network
organization that contribute most strongly to a given whole-brain network statistic. The NS
jackknife was inspired by the traditional jackknife estimator used extensively in nonparametric
statistics and can be considered a virtual lesioning technique. We are using the term “jackknife”
not only because the NS jackknife relies on the leave-one-out approach of the traditional jack-
knife, but also because like the traditional jackknife, the NS jackknife is a general statistical
technique that can be applied to any question that entails localizing a global network statistic.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
A jackknife in statistics is a resampling technique used to adjust for outliers using a leave-
one-out approach (Efron, 1982; Tukey, 1958). This procedure removes a single observation,
such as a single subject’s data, from a sample, recalculates the effect estimate, then adds the
removed observation back into the sample and removes another; it iterates through the sample
in this fashion. In standard sampling situations, in which each observation is the behavioral data
of a single individual, this method is useful for detecting potential outliers. Jackknife techniques
have been previously used in network science to assess the overall robustness of a network
statistic to node removal (Snijders & Borgatti, 1999). Sin embargo, previous uses of this technique
have focused on single instances of a network (es decir., one social network) and on obtaining
estimates of a network statistic, rather than on assessing the importance of particular network
structures in a set of multiple networks (es decir., multiple brain networks).
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
While a standard jackknife procedure removes individual subjects in turn from an analysis,
we are proposing here to remove elements of a graph, such as nodes, bordes, or subnetworks.
Here for the sake of example, we focus on the removal of entire functional subnetworks, o
tightly interconnected sets of regions of interest (ROI) thought to underlie various aspects of
cognition, such as the default mode network (Greicius, Krasnow, Reiss, & menón, 2003) or the
salience network (Seeley et al., 2007). Once a subnetwork is removed, the network statistic is
recalculated, and the procedure continues with the removal of another subnetwork. Specific
versions of this procedure have been used in previous neuroimaging research (p.ej., Achard,
Neurociencia en red
74
The network statistic jackknife
Salvador, Whitcher, Suckling, & bullmore, 2006; Henry, Dichter, & Gates, 2018); we develop
a general framework here. críticamente, while our NS jackknife estimator is based on previous
work in simulated lesioning, and at its core is a simulated lesioning method (Achard et al.,
2006; Alstott, romper la lanza, Hagmann, Cammoun, & despreciar, 2009; Crucitti, latora, marchiori,
& Rapisarda, 2004), it is focused on localization of network effects rather than the overall
robustness of a network. More specifically, while robustness focuses on understanding the
overall resiliency of a network to attack, localization attempts to understand the impact of the
removal of specific elements on the network. Robustness and localization are highly related,
and the NS jackknife differs from previous simulated lesioning methods mainly in interpretation
and in its specific focus on understanding group differences. Además, while the focus here
is on testing group differences without subject- or network-level covariates, the NS jackknife
framework can be extended to more complex models that can account for covariates and
other design factors. En este sentido, the NS jackknife is an extension of the simulated lesioning
technique into a general testing framework for localizing global network statistics. Finalmente, nosotros
provide an easily accessible implementation of the NS jackknife in the netjack package.
The NS Jackknife
To describe the NS jackknife in more precise terms, dejar {G1, . . . Gn} be the observed networks
of n subjects. sea f (·) be the functional form of some network statistic, such as modularity,
and let k be the indicator of a feature of interest (such as a subnetwork), taken from some set
K of size |k|. Gi[−k] is the network of individual i with feature k removed. More concretely, si
we were analyzing functional connectivity networks, then Gi would be the whole-brain func-
tional connectivity network of individual i, and Gi[−DMN] would be the functional connectivity
network of individual i with all the nodes of the default mode network removed.
The set of original network statistics is f (·) applied to all G1, . . . Gn, and this n-length vector
is denoted as ˆf . The jackknife estimate with regard to feature k is then f (·) applied to all
G1[−k], . . . , Gn[−k]; this length n vector is denoted as ˆf−k. Finalmente, the difference between the
original network statistic and the jackknife estimator with regard to structure k is denoted as
ˆd−k.
Applying this procedure to the |k| features of interest (p.ej., all functional subnetworks) leads
a |k| jackknife estimate vectors and |k| difference vectors for each subject. If subjects are
(B)
divided between Groups A and B, we denote the estimate and difference vectors as ˆf
−k
and ˆd
(A)
−k
, ˆf
(A)
−k , ˆd
(B)
−k .
There are several methodological considerations when using the NS jackknife. The first is
that network statistics are a priori dependent on network size and density (del barrio, estampar,
& Daffertshofer, 2010); comparing network statistics across different subsets of networks of
different sizes can lead to spurious findings. In the NS jackknife framework, this issue is handled
by comparing networks of the same size (as in the group test) or changes in network statistics
(as in the differential impact test).
Además, the NS jackknife produces a series of dependent multiple comparisons. In tra-
ditional jackknife settings, observations removed at each iteration are independent of all other
observaciones. Sin embargo, in the NS jackknife the removal of individual elements from every sub-
ject’s network and the subsequent reestimation of network statistics results in a series of depen-
dent statistical tests. Because of the dependent nature of these tests, and the risk for inflation
because of multiple comparisons, the Benjamani-Hochberg (BH) procedure is an appropriate
false discovery rate (FDR) correction for multiple comparisons (Benjamini & Hochberg, 1995).
Neurociencia en red
75
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Other multiple comparisons corrections that allow for dependent tests, such as the Benjamani-
Yekutieli (BY; Benjamini & Yekutieli, 2001) correction, are also appropriate, while the Bonfer-
roni correction would not be, as it does not account for the presence of dependent tests. If the
network components of interest are nodes or edges, and the subnetwork membership of these
components is known, the screen filtering approach of Meskaldji et al. (2015) provides a way
of correcting for multiple comparisons that results in more power to detect effects than the BH
or BY corrections, and should be preferentially used.
All analyses and graphical outputs in this manuscript use the netjack open source R pack-
edad (Henry, 2018), a package tailor-made for performing the NS jackknife in a simple to use,
customizable fashion.
Localizing Group Differences and Differential Impact
The primary use of the NS jackknife is to localize global network statistic differences be-
tween groups to specific subnetworks, nodos, or edges. Consider the example of comparing
the whole-brain modularity for a given set of subnetworks between two clinical groups. In ad-
dition to quantifying differences across the groups in global modularity using standard network
Estadísticas, the NS jackknife allows for two additional questions of substantive interest:
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
1. Group differences: Given the removal of a subnetwork, are the two groups’ mean global
values significantly different from each other? This corresponds to a hypothesis of the
following:
(A)
ˆf
−k i 6= E h
2. Differential impact: Is there a differential impact of the removal of a subnetwork between
(B)
ˆf
−k i .
E h
the two groups? This corresponds to a hypothesis of the following:
E h
(A)
ˆd
−k i 6= E h
(B)
ˆd
−k i .
We illustrate the use of these two tests in the following simulated example.
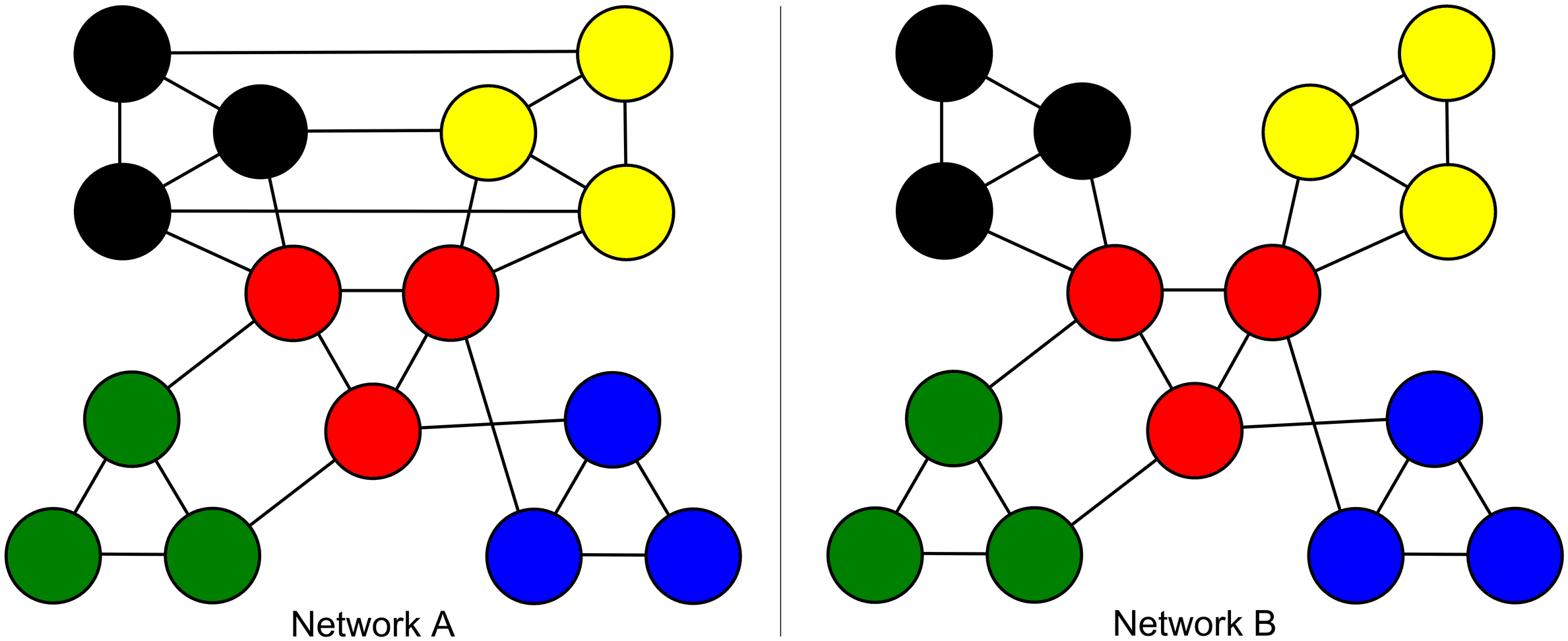
Data generation. For Group A, the sample of networks was generated using a stochastic
block model for binary undirected networks (Nowicki & Snijders, 2001), with probabilities set
to create the observed community structure (0.9 dentro, 0.5 entre). The sample contained
10 “subjects.” For Group B, data generation used the same procedure and the same base prob-
abilities (0.9 dentro, 0.5 entre), with additional connections between the yellow and black
groups set to 0.5 probabilidad (ver figura 2).Group B, like Group A, contained 10 “subjects.”
Análisis. The NS jackknife was used to quantify the effect of the removal of each of the five
subnetworks on each subject’s overall network modularity. For the group differences analysis,
a two-sample t test was conducted to determine whether the updated modularity due to the
removal of any given subnetwork was significantly different across groups. For the differential
impact analysis, a two-sample t test was conducted on the difference in modularity due to the
removal of any given subnetwork to determine whether there was a differential impact across
the groups in removing each subnetwork. For the differential impact analysis, p values were
corrected for five tests using the BH correction for dependent multiple comparisons. Group
difference test p values are uncorrected.
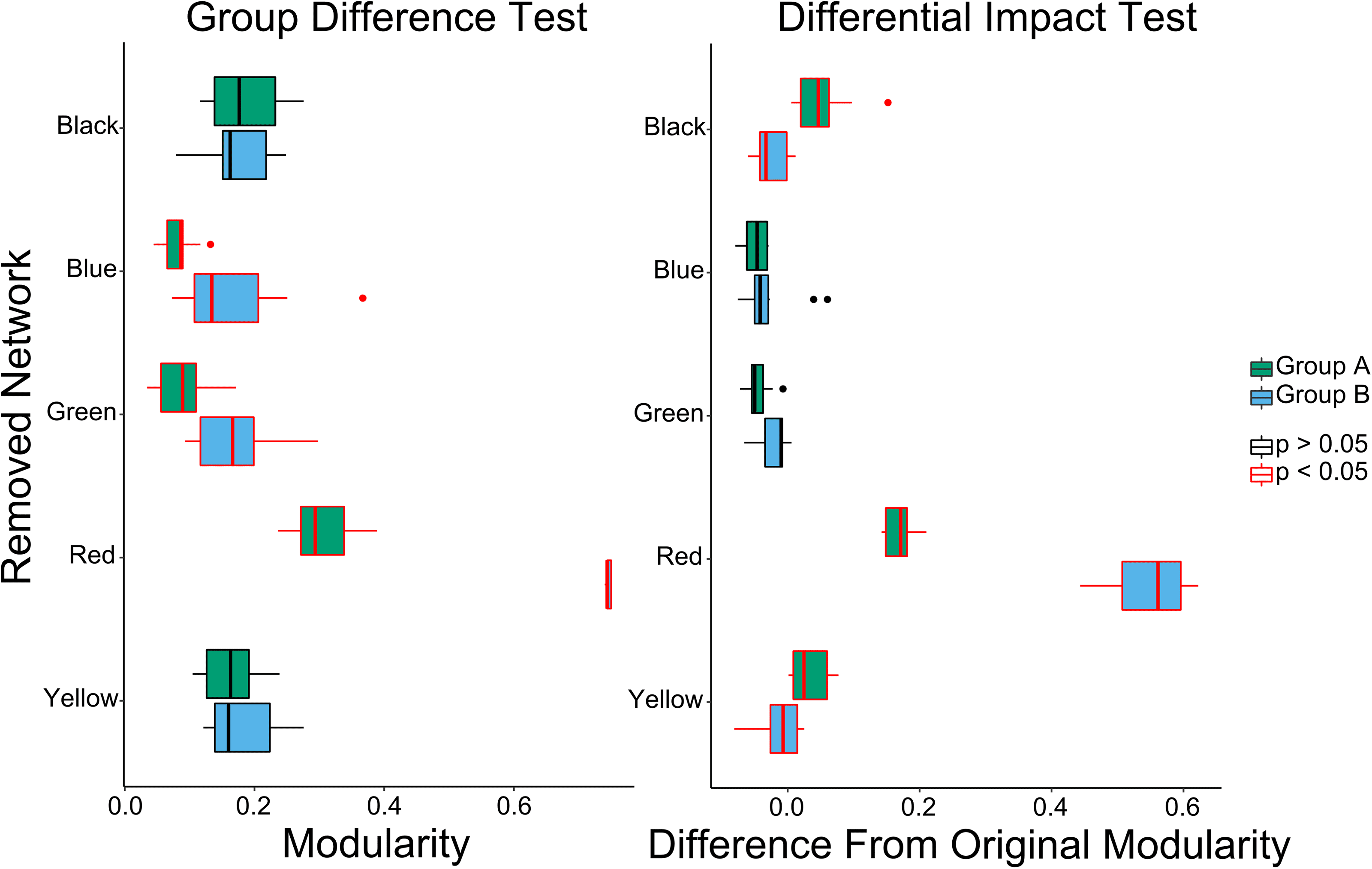
Resultados. Whole-brain modularity was significantly higher in Group B than in Group A
(t(13.41) = 2.9288, pag < 0.05; average modularity 0.1974 and 0.1332, respectively). For the
group differences analysis, the whole-brain group difference (B > A) remained after removal of
Neurociencia en red
76
The network statistic jackknife
Cifra 2. Two modular networks. The red module acts as a central hub for both networks, mientras
in Network A the black and yellow modules are directly interconnected.
the blue, verde, or red subnetworks, while removal of the black or yellow subnetworks resulted
in no group difference in modularity (Cifra 3, left panel). For the differential impact analysis,
removal of the black and yellow subnetworks resulted in an increased change in modularity for
Group A as compared with Group B. Removal of the red subnetwork resulted in an increased
change in modularity for Group B as compared with Group A (although modularity increased
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3. Group differences (left figure) and differential impact (right figure) of subnetwork removal
on modularity of Network A (in blue) and Network B (in green) from Figure 2. Central line is the
median, box is the interquartile range (IQR), and whiskers represent 1.5 ∗ IQR. Points represent
values greater than 2 SD from the mean. Both group difference and differential impact tests suggest
that the red network is significantly more centrally integrated in Group B than in Group A. El
differential impact tests additionally show that removal of the black or yellow network causes more
of an increase in modularity for Group A than Group B, suggesting that these subnetworks are more
integrated into the whole network in Group A.
Neurociencia en red
77
The network statistic jackknife
for both groups), and removal of the blue and green subnetworks resulted in a comparable
(and near-zero) change in modularity across the groups (Cifra 3, right panel).
Interpretation. The red subnetwork was highly integrated in both Group A and Group B;
sin embargo, its removal led to less of an impact on modularity in Group A than Group B. El
black and yellow subnetworks were more integrated into the rest of the network in Group A
than Group B.
Comparing group differences with differential impact analysis permits one to directly assess
the impact that local differences can have on global findings. To summarize, the removal of
the red subnetwork corresponded to a significant group difference as well as a significant
differential impact, suggesting that its removal led to the networks becoming more different
from each other. A diferencia de, the removal of the yellow or black subnetworks led to significant
differential impact but no significant group differences, indicating that the removal of either of
these subnetworks led to the networks becoming more similar to one another.
Simulation Study: Specificity and Sensitivity of the NS Jackknife
A simulation study assessing the specificity and sensitivity of the NS jackknife to the magni-
tude of group differences, sample size, and network size was conducted. Results show that NS
jackknife has good sensitivity to detect small group differences in the generating model at mod-
erate sample sizes (50 subjects per group) and larger networks (250 nodos) and shows excel-
lent sensitivity for larger group differences across multiple conditions. In a condition in which
groups had different whole-network statistics that were driven by specific local differences,
NS jackknife detected differences in differential impact across all subnetworks (Condition 4,
Figura complementaria 5), and an examination of effect sizes correctly identified which subnet-
works had the most impact on the network statistic (Figura complementaria 6). In all conditions
probado, network size was an important predictor of power, with larger networks demonstrating
greater power to detect small effects.
One key conclusion from this simulation study regards the interpretation of NS jackknife
results when the tested groups are significantly different in their whole-brain statistic. En esto
caso, the NS jackknife tends to detect differences upon the removal of all the network elements
being jackknifed on. En este caso, two routes of interpretation are viable. The first is to examine
the effect sizes of differences to determine which component of the network contributes most
to the differences between the groups (es decir., which has the largest effect size). The second is to
determine whether any subnetwork was not found to have a significant differential impact, en
which case it can be concluded that those subnetworks were not significant contributors to
the observed group difference.
For full details of this simulation study, see the Supporting Information.
Empirical Example: Reduced Integration of Subnetworks in ADHD
This empirical example is a proof of concept demonstrating how the use of the NS jackknife
provides important information about differences in functional brain network organization in
children with ADHD and TD children. ADHD is a neurodevelopmental disorder characterized
by difficulty with sustaining attention, as well as excessive impulsive or hyperactive behavior
(American Psychiatric Association, 2013). Existing research indicates that whole-brain func-
tional and structural topology are disrupted in individuals with ADHD, as are specific local
differences in connectivity (for reviews, see the following: Cao, Shu, Cao, Wang, & Él, 2014;
Neurociencia en red
78
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Henry & cohen, 2019). Individuals with ADHD exhibit higher whole-brain functional segrega-
tion than healthy control participants (Lin et al., 2014; Wang y cols., 2009). In terms of specific
local differences, there appears to be disruptions in networks such as the default mode, fron-
toparietal, atención, visual, and motor subnetworks (Castellanos & Proal, 2012). Given these
findings at the global and local level, a comparison of individuals with ADHD and healthy
control participants is a useful test case for the NS jackknife. In the following analyses, nosotros
examine global efficiency (latora & marchiori, 2001) of resting-state functional MRI scans in
children with ADHD as compared with TD children. Global efficiency is a measure of func-
tional integration. Previous studies have mixed findings with regard to differences in global
efficiency, either showing that individuals with ADHD have slightly lower global efficiency
than healthy control participants (Lin et al., 2014) or no differences in global efficiency (Wang
et al., 2009). In this example, we attempt to localize potential differences in global efficiency
to specific functional subnetworks in an effort to learn more about these inconsistencies in the
prior literature.
MÉTODOS
Sample Characteristics and Data Acquisition
Participants included 80 children with ADHD between 8 y 12 years of age (male N = 55
and female N = 25) y 208 age-matched TD children (male N = 150 and female N = 58).
For details on study inclusion criteria, see Barber and colleagues (2015).
All data were collected at the Kennedy Krieger Institute (baltimore, Maryland) using a Philips
3T Achieva MRI scanner. High-resolution T1-weighted 3D MPRAGE images were acquired
with the following parameters: repetition time (TR) = 8.05 EM, tiempo de eco (EL) = 3.76 EM, flip
angle = 8◦, matrix 256 × 256, field of view (FOV) = 200 mm, and slice thickness 1 mm. Para
resting-state data, 21 participants had 128 T2*-weighed echoplanar images (EPIs), mientras 167
participants had 156, both collected with the following parameters: TR = 2.5 s, TE = 30 EM,
flip angle = 70◦, matrix 96 × 96, and FOV = 256 mm.
Preprocesamiento
The resting-state images were preprocessed using FMRIPREP version 1.0.7 (Esteban et al.,
2018), a Nipype-based tool (Gorgolewski et al., 2011). Each T1-weighted volume was cor-
rected for intensity nonuniformity using N4BiasFieldCorrection v2.1.0 (Tustison et al., 2010)
and skull-stripped using antsBrainExtraction.sh v2.1.0 (using the OASIS template). Brain sur-
faces were reconstructed using recon-all from FreeSurfer v6.0.1 (Valle, pescado, & Sereno, 1999),
and the brain mask estimated previously was refined with a custom variation of the method
to reconcile ANTs-derived and FreeSurfer-derived segmentations of the cortical graymatter of
Mindboggle (Klein et al., 2017). Spatial normalization to the ICBM 152 Nonlinear Asymmet-
ric template version 2009c (fonov, evans, McKinstry, almíli, & collins, 2009) was performed
through nonlinear registration with the antsRegistration tool of ANTs v2.1.0 (Avants, Epstein,
Grossman, & Gee, 2008), using brain-extracted versions of both the T1-weighted volume
and template. Brain tissue segmentation of cerebrospinal fluid (CSF), whitematter (W.M.), y
graymatter (GM) was performed on the brain-extracted T1-weighted image using FAST
(zhang, Brady, & Herrero, 2001).
Functional data were slice time corrected using 3dTshift from AFNI v16.2.07 (Cox, 1996)
and motion corrected using mcflirt (FSL v5.0.9; Jenkinson, Bannister, Brady, & Herrero, 2002).
Neurociencia en red
79
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
This was followed by coregistration to the corresponding T1-weighted image using boundary-
based registration (Greve & pescado, 2009) con 9 degrees of freedom, using bbregister (FreeSurfer
v6.0.1). Motion-correcting transformations, BOLD-to-T1-weighted transformation, and T1-
weighted-to-template (MNI) warp were concatenated and applied in a single step using
antsApplyTransforms (ANTs v2.1.0) using Lanczos interpolation. Frame-wise displacement
(FD) (Power et al., 2013) was calculated for each functional run using the implementation
of Nipype.
Many internal operations of FMRIPREP use Nilearn (Abraham et al., 2014), principally
within the BOLD-processing workflow. For more details of the pipeline see https://fmriprep.
readthedocs.io/en/latest/workflows.html. This description of the preprocessing pipeline was
generated at http://fmriprep.readthedocs.io/en/latest/citing.html.
Motion Processing and Spectral Filtering
Following preprocessing with FMRIPREP, the resting-state scans were corrected for nuisance
variables, including motion, WM signal, and CSF signal. This correction proceeded in the
following steps. Timepoints that exceeded 0.3mm FD (Fuerza, Barnes, Snyder, Schlaggar, &
Petersen, 2012) were flagged, as were the timepoints immediately preceding and following the
flagged timepoint. Interpolation across these flagged timepoints was conducted using spectral
interpolación (Ciric et al., 2017). Following the spectral interpolation, a high-pass filter at 0.008
MHz was applied to the voxel time series containing the interpolated timepoints. The flagged
timepoints were then set to missing for all voxels. A nuisance regression set consisting of the 6
degrees of motion and mean WM and CSF signal, as well as the temporal derivatives, quadratic
expansions, and the quadratic expansions of the temporal derivatives of these 8 regressors, era
high-pass filtered at 0.008 MHz, as per the suggestion of Ciric, Satterthwaite, and colleagues
(Ciric et al., 2017; Satterthwaite et al., 2013). The filtered nuisance regressors were used to
regress out nuisance signal from the scrubbed and filtered voxel time series. Finalmente, subjects
that had more than 30% of their data lost because of scrubbing were removed from the analysis,
as were subjects who exceeded 0.2 mm mean FD overall. This resulted in a final sample of
34 children with ADHD and 121 TD children, with mean percentage timepoints lost 8% y
10% respectivamente. This difference in percentage timepoints lost was nonsignificant (t(48.423) =
−1.47, pag = 0.14). Además, the mean FD in the final sample was not significantly different
between children with ADHD and TD children (t(49.902) = 1.27, pag = 0.21, ADHD mean =
0.131, TD mean = 0.121).
ROI Extraction and Network Construction
From the preprocessed and motion-corrected resting-state images, ROI time series were
extracted using a commonly used functional brain atlas (Power et al., 2011). This atlas
consists of 264 ROI, 234 of which are classified into 13 functional subnetworks. Estos
functional subnetworks and abbreviations are as follows: auditory (AUD), cerebellar (CER),
cingulo-opercular (CO), default mode (DMN), atención dorsal (DAN), frontoparietal (FP),
memory (MEM), prominencia (SAL), somatomotor hand (SM Hand), somatomotor mouth (SM
Mouth), subcortical (SUB), atención ventral (VAN), and visual (VIS). From these ROI time series,
234 × 234 correlation matrices were computed. These correlation matrices were then con-
verted to absolute values and thresholded at a correlation magnitude of 0.35 to create binary
connectivity networks. The pattern of findings below was consistent for a band of thresholds
de 0.25 a 0.4.
Neurociencia en red
80
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Network Statistics and Jackknife
In this application, we use global efficiency as our network statistic. Global efficiency is de-
fined as the average of the inverse shortest path length between all nodes in the network (latora
& marchiori, 2001), and can be thought of as a measure of global integration. We applied the
jackknife over the 13 functional subnetworks in the functional brain atlas. The two tests previ-
ously described were then applied to the jackknifed data. This allowed us to assess (a) si
the removal of a functional subnetwork led to a significant difference in global efficiency be-
tween children with ADHD and TD children; y (b) whether the impact of the removal of
a functional subnetwork on functional integration was different for children with ADHD as
compared with TD children.
Jackknife Estimate and Group Comparisons
Independent two-sample t tests were conducted to compare global efficiency values across the
entire sample, and to compare global efficiency values and differential impacts across groups.
Differential impact results were corrected for multiple dependent comparisons using the BH
correction, as described in the above simulations.
RESULTADOS
Group differences in whole-brain global efficiency: There was no group difference in global
efficiency for children with ADHD and TD children (t(5037) = 0.99, pag = 0.32; TDAH
mean = 0.29, TD mean = 0.28).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4. Group differences in global efficiency between children with ADHD and TD children
after the removal of a given subnetwork. Networks are presented here in alphabetical order. Central
line is the median, box is the interquartile range (IQR), and whiskers represent 1.5 ∗ IQR. Points
represent values more than 2 SD away from the mean. Removal of the SUB network resulted in
children with ADHD having significantly higher global efficiency than TD children.
Neurociencia en red
81
The network statistic jackknife
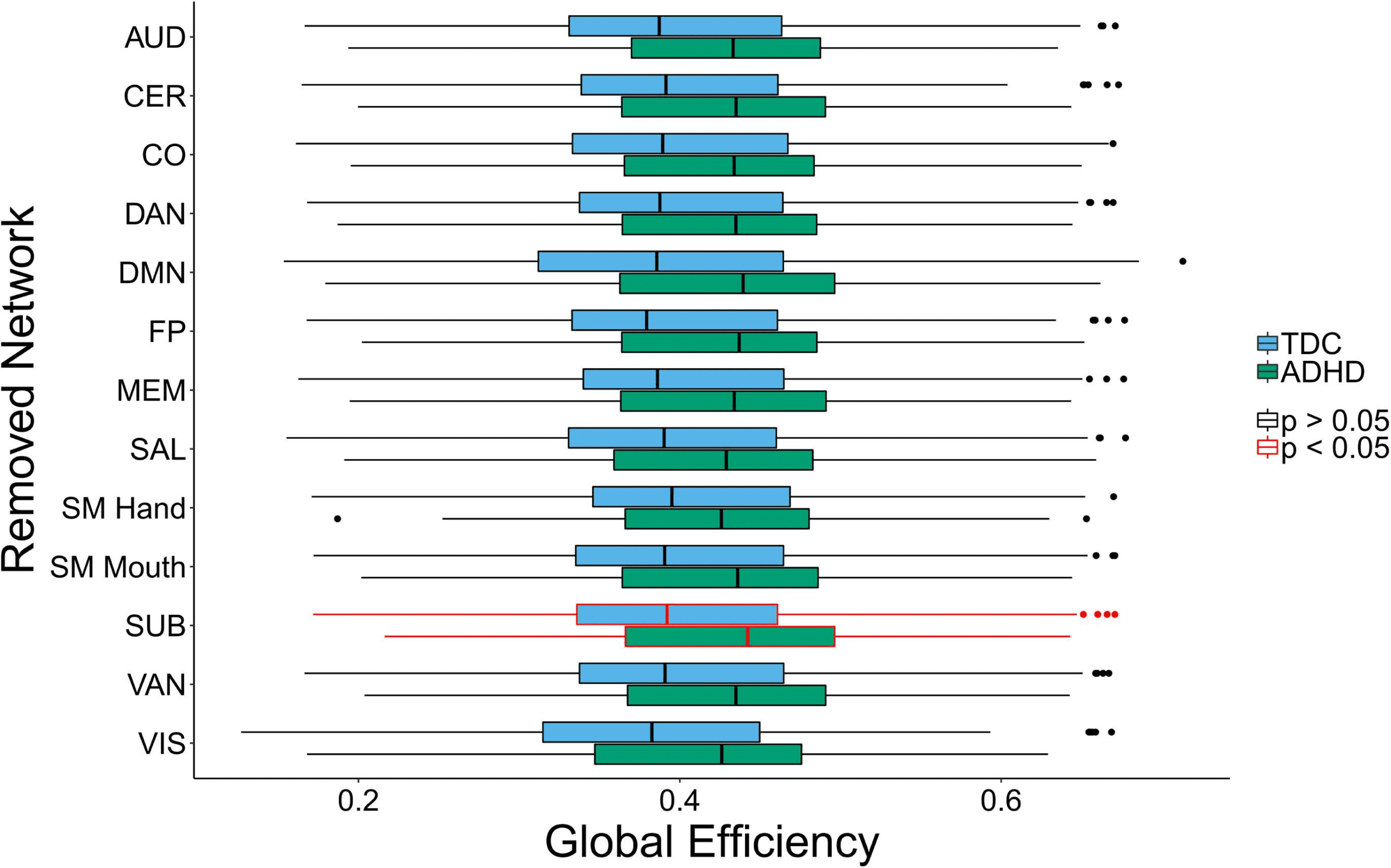
Group differences by subnetwork removal: Próximo, we examined group differences in global
efficiency between children with ADHD and TD children after removing each functional
subnetwork.
Removal of the SUB subnetwork led to a significant difference between children with ADHD
and TD children with respect to global efficiency (Cifra 4). Específicamente, children with ADHD
had greater global efficiency than TD children after SUB subnetwork removal. Mesa 1 en el
Supporting Information contains the numerical differences and p values. It should be noted that
these results are best interpreted when combined with the differential impact results below.
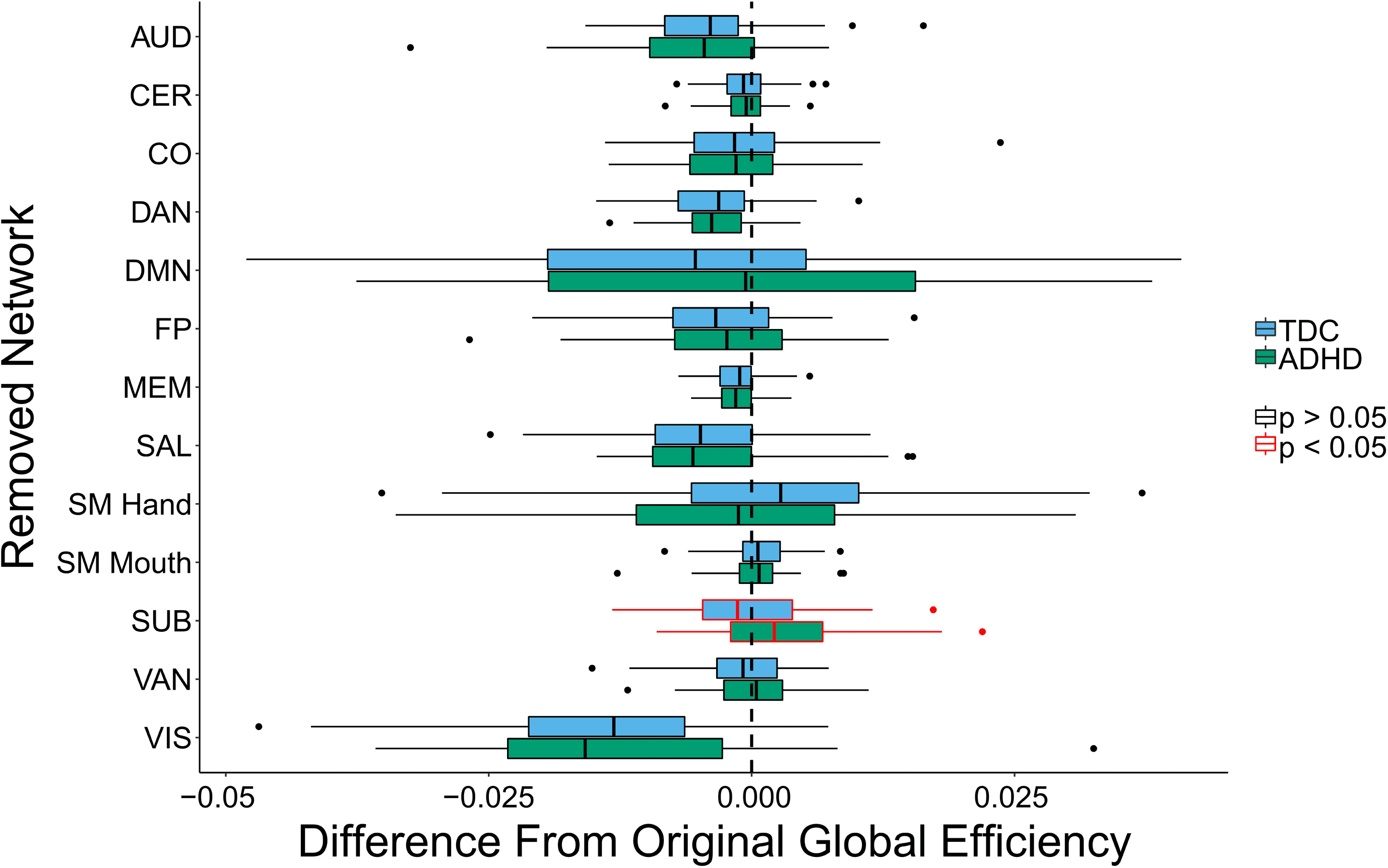
Differential impact of subnetwork removal: Finalmente, we examined whether the removal of
any of the functional subnetworks led to a differential change in global efficiency for children
with ADHD as compared with TD children.
The only subnetwork that exhibited significant differential impact after adjusting for mul-
tiple comparisons was the SUB subnetwork. Children with ADHD exhibited an increase in
global efficiency after SUB subnetwork removal, while TD children exhibited a slight decrease
(Cifra 5). These findings suggest that the SUB subnetwork has lower integration with the global
network in children with ADHD. The lower integration of the SUB subnetwork in children with
ADHD is consistent with prior findings of disruptions in cortico-striatal-thalamic-cortical loops
in ADHD (para una revisión, see Posner, Parque, & Wang, 2014). En tono rimbombante, this finding is an ex-
ample of equifinality, as the difference in the local topology of the subcortical network did
not lead to an observed difference in whole-brain global efficiency. Mesa 3 in the Supporting
Information contains the numerical differences and p values for this analysis.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 5. Differential impact on global efficiency when removing subnetworks between children
with ADHD and TD children. Central line is the median, box is the interquartile range (IQR), y
whiskers represent 1.5 ∗ IQR. Points represent values more than 2 SD away from the mean. Removal
of the subcortical subnetwork (SUB) led to a greater increase in global efficiency in children with
ADHD as compared with TD children, suggesting that the SUB subnetwork is less integrated into the
whole-brain network in children with ADHD. Significant differences were based on BH corrected
p values.
Neurociencia en red
82
The network statistic jackknife
DISCUSIÓN
Whole-brain network analysis is a powerful tool for assessing brain topology, yet it lacks the
ability to localize findings related to global topology to a specific location. This issue, com-
bined with the fact that robust local differences do not necessarily result in observable global
differences in topology, suggests that whole-brain network analysis has a substantial blind
spot. Local graph theory analysis metrics can alleviate this but lack the ability to directly relate
local findings to global topology. In this article, we introduced the NS jackknife, a flexible tool
for localizing global differences in network topology. Our framework links the global scope to
the local scope (Hallquist & Hillary, 2018) and is flexible enough to operate on any global
network statistic. We implemented this framework in an open source R package, netjack
to make it accessible to re-
(Henry, 2018; https://CRAN.R-project.org/package=netjack)
searchers. Our simulation study evaluating the performance of the NS jackknife showed that
the method has excellent sensitivity to local differences in network topology across a variety of
condiciones, while keeping the false discovery rate at a nominal level (0.05; see the Supporting
Información).
The NS jackknife is an extension of the simulated lesioning approach that offers a variety of
advantages to neuroimaging researchers interested in examining group differences in network
topología. The NS jackknife extends the simulated lesioning approach beyond its most common
application, examining the robustness of a network to attack, thus enabling researchers to
examine how individual elements of a given network contribute to a global network statistic.
It can be applied to any global network statistic under study, which allows researchers to extend
existing analysis pipelines rather than requiring implementation of a new set of analyses. El
specificity of a global analysis can be tested at any level of resolution (subnetwork, nodo,
borde), and resulting significance tests are appropriately corrected for multiple comparisons,
making this an easy tool to implement and interpret. Finalmente, while the focus here was on binary
networks derived from functional MRI data, the NS jackknife can easily be applied to weighted
functional networks or to structural networks derived from diffusion-weighted imaging.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
t
.
There are several further methodological developments for the NS jackknife. While we fo-
cused on comparisons between two groups of subjects, future versions of the NS jackknife
toolbox will allow for within-subject across-time comparisons, testing covariates and control-
ling for potential confounds, and analyzing factorial grouping structures. The NS jackknife
framework can be extended to generalized linear regression approaches as well as to allow
for dependencies, such as those that arise from multiple collection sites. This is an active area
of development, with implementations being introduced in the near future. Although the NS
jackknife as currently presented examines network statistics as the outcome, it is likewise nat-
ural to consider network statistics as predictors of some distal outcome, such as behavior or
symptomology. Our framework can be extended to handle cases where some distal outcome
is of interest and would allow an assessment of the importance of local differences in network
topology with regard to that distal outcome.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Finalmente, our empirical example produced results that were consistent with prior results
suggesting little difference in global metrics of functional integration (es decir., global efficiency)
between children with ADHD and TD children. En tono rimbombante, our use of the NS jackknife un-
covered an occurrence of equifinality, in which a local group difference did not lead to a
global group difference. Específicamente, an analysis of group differences and differential impact of
subnetwork removal showed that the subcortical subnetwork was less integrated into the whole-
brain network in children with ADHD, consistent with prior literature (Hong et al., 2014).
Neurociencia en red
83
The network statistic jackknife
Limitations and Considerations
A key consideration for the use of the NS jackknife is in the use of weighted networks or
networks formed from white matter tractography. The NS jackknife is agnostic to the type of
network (weighted vs. binario, undirected vs. directed), but special care needs to be taken
when using weighted networks to ensure that the appropriate network statistics are used. Para
ejemplo, when global efficiency is computed on correlation networks, it often exhibits scaling
issues due to taking the inverse of correlations that are close to zero. If those correlations are
removed during the NS jackknife, the resulting difference in global efficiency could simply be
due to that scaling, and not due to any true difference in network structure. This is an issue for
any weighted graph analysis and is not limited to the NS jackknife approach.
NS jackknife as a general framework has a limitation that users need to consider. As most
tests of interest involve multiple comparisons, a multiple comparison correction is necessary to
reduce the false discovery rate. Because of this, as the number of network features jackknifed
aumenta, power to detect any given difference due to the removal of a single network feature
decreases. This therefore limits the scope of an NS jackknife application. In smaller samples,
NS jackknife is likely best applied to questions regarding functional subnetworks, en el cual
the number of subnetworks is relatively small and potential impacts are large. For larger
estudios, particularly studies utilizing public datasets such as the Human Connectome Project
(Van Essen et al., 2013) or the Autism Brain Imaging Data Exchange (ABIDE) conjuntos de datos
(Di Martino et al., 2017; Di Martino et al., 2014), it becomes more feasible to examine dif-
ferences at the level of nodes or even edges. Another strategy that researchers with smaller
samples can employ is examining more localized networks with the NS jackknife. por ejemplo-
amplio, conducting an analysis limiting the network under consideration to the default mode
network and removing nodes and edges of that smaller network rather than of the whole-brain
network. This particular limitation, sin embargo, is inherent to neuroimaging analysis given the
high-dimensional nature of the data. This speaks to a general need for the development of
analytic methods of high-dimensional data dealing with issues of global to local scoping as
discutido anteriormente. One potential solution to this problem is to use the screen filtering method
of Meskaldji and colleagues (2015) to adjust the p values for tests of difference at the node or
edge level with respect to subnetwork groupings.
For pedagogical purposes the manuscript focused on detecting local differences in single
canonical functional subnetworks; sin embargo, the NS jackknife can be applied to combinations
of canonical subnetworks as well combinations of nodes or edges. One key consideration
here is that of combinatoric complexity. When using the NS jackknife to test removal of com-
binations of multiple subnetworks or nodes, the number of possible combinations increases
factorially. To avoid this, the NS jackknife can be used in a confirmatory way, with researchers
specifying a set of combinations of network structures informed by previous literature or a pre-
vious exploratory application of NS jackknife, then testing for local differences with that set
of combinations. Finalmente, the NS jackknife does not require the use of canonical subnetworks
as demonstrated here. Sample-specific parcellations derived from some other method can be
used instead. The only requirement here is that a given network is defined on the same nodes
for all subjects.
There are two additional considerations for researchers interested in using the NS jackknife
estructura. The first is that of network construction. The NS jackknife assumes a fixed network
estructura, and that the only changes that are occurring are due to the removal of specific
network features. This means that network estimation techniques, such as regularized partial
correlations, are to be used before the NS jackknife is applied, and not applied to the jackknifed
Neurociencia en red
84
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
networks themselves. This ensures that any localized effect can be interpreted as due to the
removal of the specific structure only, and not an analytical artifact caused by a reestimation
of the network.
The second consideration is that of the density/strength dependency. As mentioned earlier,
network statistics (such as average degree) are dependent on the size and density of the network
(van Wijk et al., 2010). Tal como, it is expected that the network statistics would change when
structures are removed. This potentially causes issues with interpretation, where an observed
group difference might simply be due to differences in density of the removed component
rather than any topological difference. One potential solution to this interpretational concern
is to add the network element’s density or strength as a covariate to the jackknife models.
Research into adjustments for density/strength dependency is ongoing.
CONCLUSIÓN
In closing, the NS jackknife framework bridges the gap between global network analysis and
local differences in topology, making it an ideal tool with which to investigate the specificity
of observed differences in whole-brain topology. Además, it allows researchers to explore
local network features in the absence of a global result while still using the tools, terminology,
and interpretation of network neuroscience at the global level. Finalmente, future developments will
extend the NS jackknife to more complex modeling frameworks and settings, further extending
its utility within the neuroimaging community.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00109.
The NS jackknife method is available at https://cran.r-project.org/package=netjack (Henry,
2018).
CONTRIBUCIONES DE AUTOR
Teague Henry: Análisis formal; Investigación; Metodología; Software; Visualización; Writing –
Original Draft; Writing – Review & Editing. Kelly A. Duffy: Curación de datos; Recursos; Writing –
Revisar & Editing. Marc D. Rudolph: Writing – Original Draft; Writing – Review & Editing. Mary
Beth Nebel: Curación de datos; Writing – Review & Editing. Stewart H. Mostofsky: Curación de datos;
Adquisición de financiación; Writing – Review & Editing. Jessica R. cohen: Conceptualización; Fondos
adquisición; Recursos; Supervisión; Writing – Original Draft; Writing – Review & Editing.
INFORMACIÓN DE FINANCIACIÓN
Jessica R. cohen, National Institute of Mental Health (US), Award ID: R00MH102349. Stewart
h. Mostofsky, National Institute of Mental Health (US), Award ID: R01MH085328. Stewart
h. Mostofsky, National Institute of Mental Health (US), Award ID: R01MH078160. Mary Beth
Nebel, National Institute of Mental Health (US), Award ID: K01MH109766.
REFERENCIAS
Abrahán, A., Pedregosa, F., Eickenberg, METRO., Gervais, PAG., Mueller,
A., Kossaifi, J., . . . Varoquaux, GRAMO. (2014). Machine learning for
neuroimaging with scikit-learn. Frontiers in Neuroinformatics, 8.
https://doi.org/10.3389/fninf.2014.00014
Achard, S., Salvador, r., Whitcher, B., Suckling, J., & bullmore,
mi. (2006). A resilient, low-frequency, small-world human brain
functional network with highly connected association cortical
hubs. Revista de neurociencia, 26(1), 63–72. https://doi.org/10.
1523/JNEUROSCI.3874-05.2006
Alstott, J., romper la lanza, METRO., Hagmann, PAG., Cammoun, l., & despreciar,
oh. (2009). Modeling the impact of lesions in the human brain.
Biología Computacional PLoS, 5(6), e1000408. https://doi.org/10.
1371/journal.pcbi.1000408
Neurociencia en red
85
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
American Psychiatric Association. (2013). Diagnostic and statistical
manual of mental disorders (5th ed.). Arlington, Virginia: Americano
Psychiatric Publishing.
Avants, B. B., Epstein, C. l., Grossman, METRO., & Gee, j. C. (2008). Sym-
metric diffeomorphic image registration with cross-correlation:
Evaluating automated labeling of elderly and neurodegenerative
cerebro. Análisis de imágenes médicas, 12(1), 26–41. https://doi.org/10.
1016/j.media.2007.06.004
Barber, A. D., jacobson, l. A., Wexler, j. l., Nebel, METRO. B., Caffo,
B. S., Pekar, j. J., & Mostofsky, S. h. (2015). Connectivity support-
ing attention in children with attention deficit hyperactivity dis-
orden. NeuroImagen: Clinical, 7, 68–81. https://doi.org/10.1016/j.
nicl.2014.11.011
bassett, D. S., & despreciar, oh. (2017). Network neuroscience. Naturaleza
Neurociencia, 20(3), 353–364. https://doi.org/10.1038/nn.4502
Baum, GRAMO. l., círico, r., roalf, D. r., Betzel, R. F., moore, t. METRO.,
(2017). Modular
Shinohara, R. T.,
segregation of structural brain networks supports the develop-
ment of executive function in youth. Biología actual, 27(11),
1561–1572.e8. https://doi.org/10.1016/j.cub.2017.04.051
. Satterthwaite, t. D.
.
.
Benjamini, y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society. Serie B (Methodological),
57(1), 289–300. https://doi.org/10.2307/2346101
Benjamini, y., & Yekutieli, D. (2001). The control of the false discov-
ery rate in multiple testing under dependency. Annals of Statistics,
29(4), 1165–1188. https://doi.org/10.1214/aos/1013699998
Bertalanffy, l. Von. (1950). An outline of general system theory.
British Journal for the Philosophy of Science, 1(2), 134–165. https://
doi.org/10.1093/bjps/I.2.134
Cao, METRO., Shu, NORTE., Cao, P., Wang, y., & Él, Y. (2014). Imag-
ing functional and structural brain connectomics in attention-
deficit/hyperactivity disorder. Molecular Neurobiology, 50(3),
1111–1123. https://doi.org/10.1007/s12035-014-8685-x
Castellanos, F. X., & Proal, mi. (2012). Large-scale brain systems in
TDAH: Beyond the prefrontal-striatal model. Trends in Cogni-
tive Sciences, 16(1), 17–26. https://doi.org/10.1016/j.tics.2011.
11.007
círico, r., Lobo, D. h., Fuerza, j. D., roalf, D. r., Baum, GRAMO. l.,
Ruparel, K., . . . Satterthwaite, t. D. (2017). Benchmarking of
participant-level confound regression strategies for the control
of motion artifact in studies of functional connectivity. Neuro-
Image,154, 174–187. https://doi.org/10.1016/j.neuroimage.2017.
03.020
cohen, j. r., & D'Esposito, METRO. (2016). The segregation and integra-
tion of distinct brain networks and their relationship to cognition.
Revista de neurociencia, 36(48), 12083–12094. https://doi.org/
10.1523/JNEUROSCI.2965-15.2016
Cox, R. W.. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29(3), 162–173. https://doi.org/10.1006/
cbmr.1996.0014
Crucitti, PAG., latora, v., marchiori, METRO., & Rapisarda, A. (2004). Error
and attack tolerance of complex networks. Physica A: Statistical
Mechanics and Its Applications, 340(1–3), 388–394. https://doi.
org/10.1016/j.physa.2004.04.031
Valle, A. METRO., pescado, B., & Sereno, METRO. I. (1999). Cortical surface-
based analysis:
I. Segmentation and surface reconstruction.
NeuroImagen, 9(2), 179–194. https://doi.org/10.1006/nimg.1998.
0395
Di Martino, A., O’Connor, D., Chen, B., Alaerts, K., anderson, j. S.,
Assaf, METRO., . . . Milham, METRO. PAG. (2017). Enhancing studies of the
connectome in autism using the autism brain imaging data ex-
change II. Scientific Data, 4(170010). https://doi.org/10.1038/
sdata.2017.10
Di Martino, A., Shehzad, Z., Kelly, C., roy, A. K., Gee, D. GRAMO., Uddin,
l. P., . . . Milham, METRO. PAG. (2009). Relationship between cingulo-
insular functional connectivity and autistic traits in neurotypical
adultos. Revista americana de psiquiatría, 166(8), 891–899. https://
doi.org/10.1176/appi.ajp.2009.08121894
Di Martino, A., yan, C. G.-G., li, P., Denio, MI., Castellanos,
F. X., Alaerts, K., . . . Milham, METRO. PAG. (2014). The autism brain
imaging data exchange: Towards a large-scale evaluation of the
intrinsic brain architecture in autism. Molecular Psychiatry, 19(6),
659–667. https://doi.org/10.1038/mp.2013.78
Efron, B. (1982). The jackknife, the bootstrap and other resampling
planes. Filadelfia, Pensilvania: Society for Industrial and Applied Math-
ematics. https://doi.org/10.1137/1.9781611970319
Esteban, o., Markiewicz, C., Blair, R. w., Moody, C., Isik, A. I.,
Aliaga, A. MI., . . . Gorgolewski, k. j. (2018). FMRIPrep: A ro-
bust preprocessing pipeline for functional MRI. BioRxiv, 306951.
https://doi.org/10.1101/306951
fonov, v., evans, A., McKinstry, r., almíli, C., & collins, D. (2009).
Unbiased nonlinear average age-appropriate brain templates
from birth to adulthood. NeuroImagen, 47(1), S102. https://doi.
org/10.1016/S1053-8119(09)70884-5
Proporcionó, A., bullmore, mi. T., & Brilla, A. (2017). Opportunities and
challenges for psychiatry in the connectomic era. Biological Psy-
chiatry: Cognitive Neuroscience and Neuroimaging, 2(1), 9–19.
https://doi.org/10.1016/j.bpsc.2016.08.003
Proporcionó, A., Brilla, A., & romper la lanza, METRO. (2015). The connectomics of
brain disorders. Naturaleza Reseñas Neurociencia, 16(3), 159–172.
https://doi.org/10.1038/nrn3901
Gorgolewski, K., Burns, C. D., Madison, C., clark, D., Halchenko,
Y. o., Waskom, METRO. l., & Ghosh, S. S. (2011). Nipype: A flexible,
lightweight and extensible neuroimaging data processing frame-
work in Python. Frontiers in Neuroinformatics, 5(13). https://doi.
org/10.3389/fninf.2011.00013
Grayson, D. S., & Fair, D. A. (2017). Development of large-scale
functional networks from birth to adulthood: A guide to the neu-
roimaging literature. NeuroImagen, 160, 15–31. https://doi.org/
10.1016/j.neuroimage.2017.01.079
Greicius, METRO. D., Krasnow, B., Reiss, A. l., & menón, V. (2003).
Functional connectivity in the resting brain: A network analy-
sis of the default mode hypothesis. Actas del Nacional
Academia de Ciencias, 100(1), 253–258. https://doi.org/10.1073/
pnas.0135058100
Greve, D. NORTE., & pescado, B. (2009). Accurate and robust brain image
alignment using boundary-based registration. NeuroImagen, 48(1),
63–72. https://doi.org/10.1016/j.neuroimage.2009.06.060
Guimerà, r., & Nunes Amaral, l. A. (2005). Functional cartography
of complex metabolic networks. Naturaleza, 433(7028), 895–900.
https://doi.org/10.1038/nature03288
Neurociencia en red
86
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Hallquist, METRO. NORTE., & Hillary, F. GRAMO. (2018). Graph theory approaches
to functional network organization in brain disorders: A critique
for a brave new small-world. Neurociencia en red, 1(Agosto),
243741. https://doi.org/10.1101/243741
Henry, t. R.
(2018). Netjack: Tools for working with samples
of networks. Retrieved from https://cran.r-project.org/package=
netjack
Henry, t. r., & cohen, j. R. (2019). Dysfunctional brain network
organization in neurodevelopmental disorders. In P. j. Laurienti,
B. Munsell, & GRAMO. Wu (Editores.), Connectomics: Métodos, mathemat-
ical models and applications. Elsevier.
Henry, t. r., Dichter, GRAMO. S., & Gates, k. (2018). Age and gender
effects on intrinsic connectivity in autism using functional in-
tegration and segregation. Biological Psychiatry: Cognitive Neu-
roscience and Neuroimaging, 3(5), 414–422. https://doi.org/10.
1016/j.bpsc.2017.10.006
hong, S. B., Brilla, A., Proporcionó, A., Parque, S., Cual, Y. h., Parque,
METRO. h., . . . kim, j. W.. (2014). Connectomic disturbances in
attention-deficit/hyperactivity disorder: A whole-brain tractogra-
phy analysis. Biological Psychiatry, 76(8), 656–663. https://doi.
org/10.1016/j.biopsych.2013.12.013
Jenkinson, METRO., Bannister, PAG., Brady, METRO., & Herrero, S. (2002). Improved
optimization for the robust and accurate linear registration and
motion correction of brain images. NeuroImagen, 17(2), 825–841.
https://doi.org/10.1016/S1053-8119(02)91132-8
Klein, A., Ghosh, S. S., Bao, F. S., Giard, J., Häme, y., Stavsky,
MI., . . . Keshavan, A. (2017). Mindboggling morphometry of hu-
man brains. Biología Computacional PLoS, 13(2). https://doi.org/
10.1371/journal.pcbi.1005350
Verano, A., Tambini, A., Sadaghiani, S., Keilholz, S., & cohen, j. R.
(2018). Spontaneous cognitive processes and the behavioral vali-
dation of time-varying brain connectivity. Neurociencia en red,
1–57. https://doi.org/10.1162/NETN_a_00037
latora, v., & marchiori,METRO. (2001). Efficient behavior of small-worldnet-
obras. Physical Review Letters, 87(19), 198701-1-198701–198704.
https://doi.org/10.1103/PhysRevLett.87.198701
Luis, j. D., Theilmann, R. J., Townsend, J., & evans, A. C. (2013).
Network efficiency in autism spectrum disorder and its relation to
brain overgrowth. Frontiers in Human Neuroscience, 7. https://
doi.org/10.3389/fnhum.2013.00845
lin, PAG., Sol, J., Yu, GRAMO., Wu, y., Cual, y., Liang, METRO., & Liu, X. (2014).
Global and local brain network reorganization in attention-
deficit/hyperactivity disorder. Brain Imaging and Behavior, 8(4),
558–569. https://doi.org/10.1007/s11682-013-9279-3
Meskaldji, D. MI., Vasung, l., Romascano, D., Thiran, j. PAG., Hagmann,
PAG., Morgenthaler, S., & Van De Ville, D. (2015). Improved stat-
istical evaluation of group differences in connectomes by
screening-filtering strategy with application to study matura-
tion of brain connections between childhood and adolescence.
NeuroImagen,108, 251–264. https://doi.org/10.1016/j.neuroimage.
2014.11.059
Hombre nuevo, METRO. mi. j. (2006). Modularity and community structure in
redes. procedimientos de la Academia Nacional de Ciencias, 103(23),
8577–8582. https://doi.org/10.1073/pnas.0601602103
Nowicki, K., & Snijders, t. A. B. (2001). Estimation and predic-
tion for stochastic blockstructures. Journal of the American Statis-
tical Association, 96(455), 1077–1087. https://doi.org/10.1198/
016214501753208735
Pievani, METRO., de Haan, w., Wu, T., Seeley, W.. w., & Frisoni, GRAMO. B.
(2011). Functional network disruption in the degenerative de-
mentias. The Lancet Neurology, 10(9), 829–843,. https://doi.org/
10.1016/S1474-4422(11)70158-2
posner, J., Parque, C., & Wang, z. (2014). Connecting the dots: A
review of resting connectivity MRI studies in attention-deficit/
hyperactivity disorder. Neuropsychology Review, 24(1), 3-15.
https://doi.org/10.1007/s11065-014-9251-z
Fuerza, j. D., Barnes, k. A., Snyder, A. Z., Schlaggar, B. l., &
Petersen, S. mi. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject mo-
ción. NeuroImagen, 59(3), 2142–2154. https://doi.org/10.1016/j.
neuroimage.2011.10.018
Fuerza, j. D., cohen, A. l., nelson, S. METRO., Peluca, GRAMO. S., Barnes, k. A.,
Church, j. A., . . . Petersen, S. mi. (2011). Functional network or-
ganization of the human brain. Neurona, 72(4), 665–678. https://
doi.org/10.1016/j.neuron.2011.09.006
Fuerza, j. D., Mitra, A., Laumann, t. o., Snyder, A. Z., Schlaggar,
B. l., & Petersen, S. mi. (2013). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImagen, 84,
320–341. https://doi.org/10.1016/j.neuroimage.2013.08.048
Roine, Ud., Roine, T., Salmi, J., Nieminen-von Wendt, T., Tani,
PAG., Leppämäki, S., . . . Sams, METRO. (2015). Abnormal wiring of
the connectome in adults with high-functioning autism spectrum
disorder. Molecular Autism, 6(1), 65. https://doi.org/10.1186/
s13229-015-0058-4
Rudie,
j. D., & Dapretto, METRO.
(2013). Convergent evidence of
brain overconnectivity in children with autism? Informes celulares, 5(3),
565–566. https://doi.org/10.1016/j.celrep.2013.10.043
J., Calkins, METRO. MI.,
Satterthwaite, t. D., eliot, METRO. A., Gerraty, R. T., Ruparel, K.,
Loughead,
(2013). Un
.
improved framework for confound regression and filtering for
control of motion artifact in the preprocessing of resting-state
functional connectivity data. NeuroImagen, 64(1), 240–256. https://
doi.org/10.1016/j.neuroimage.2012.08.052
. Worf, D. h.
.
Seeley, W.. w., menón, v., Schatzberg, A. F., Keller, J., guantero, GRAMO. h.,
Kenna, h., . . . Greicius, METRO. D. (2007). Dissociable intrinsic con-
nectivity networks for salience processing and executive control.
Revista de neurociencia, 27(9), 2349–2356. https://doi.org/10.
1523/JNEUROSCI.5587-06.2007
Brillar, j. METRO., & Poldrack, R. A. (2017). Principles of dynamic
network reconfiguration across diverse brain states. NeuroIm-
edad, 180(punto. B), 396–405. https://doi.org/10.1016/j.neuroimage.
2017.08.010
Snijders, t. A. B., & Borgatti, S. PAG. (1999). Non-parametric standard
errors and tests for network statistics. Connections, 22(2), 1–10.
despreciar, oh. (2013). Network attributes for segregation and integra-
tion in the human brain. Opinión actual en neurobiología, 23(2),
162–171. https://doi.org/10.1016/j.conb.2012.11.015
despreciar, o., Chialvo, D. r., Kaiser, METRO., & Hilgetag, C. C. (2004).
Organization, development and function of complex brain net-
obras. Tendencias en Ciencias Cognitivas, 8(9), 418–425. https://doi.
org/10.1016/j.tics.2004.07.008
Tukey, j. W.. (1958). Bias and confidence in not-quite large sample.
Annals of Mathematical Statistics, 29, 614.
Neurociencia en red
87
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The network statistic jackknife
Tustison, norte.
J., Avants, B. B., Cocinar, PAG. A., Zheng, y., Egan,
A., Yushkevich, PAG. A., & Gee, j. C. (2010). N4ITK: Improved
N3 bias correction. IEEE Transactions on Medical Imaging, 29(6),
1310–1320. https://doi.org/10.1109/TMI.2010.2046908
VanEssen, D. C., Herrero, S. METRO., Respeto, D. METRO., Behrens, t. mi. J.,
Yacoub, MI., & Ugurbil, k. (2013). The WU-minn human connec-
tome project: An overview. NeuroImagen, 80, 62–79. https://doi.
org/10.1016/j.neuroimage.2013.05.041
del barrio, B. C. METRO., estampar, C. J., & Daffertshofer, A. (2010). Com-
paring brain networks of different size and connectivity density
using graph theory. PLoS ONE, 5(10), e13701. https://doi.org/10.
1371/diario.pone.0013701
Wang, l., Zhu, C., Él, y., Zang, y., Cao, P., zhang, h., . . . Wang,
Y. (2009). Altered small-world brain functional networks in chil-
dren with attention-deficit/hyperactivity disorder. Human Brain
Cartografía, 30(2), 638–649. https://doi.org/10.1002/hbm.20530
Yoo, K., Sotavento, PAG., Chung, METRO. K., Sohn, W.. S., Chung, S. J., Na, D. l.,
. . . jeong, Y. (2017). Degree-based statistic and center persistency
for brain connectivity analysis. Mapeo del cerebro humano, 38(1),
165–181. https://doi.org/10.1002/hbm.23352
Brilla, A., Proporcionó, A., & bullmore, mi. t. (2010). Network-based
statistic: Identifying differences in brain networks. NeuroImagen,
53(4), 1197–1207. https://doi.org/10.1016/j.neuroimage.2010.
06.041
zhang, Y. J., Brady, METRO., & Herrero, S. mi. (2001). Segmentation of brain
MR images through a hidden Markov random field model and the
expectation-maximization algorithm. IEEE Transactions on Med-
ical Imaging, 20, 45–57.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
7
0
1
9
2
0
5
3
8
norte
mi
norte
_
a
_
0
0
1
0
9
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurociencia en red
88