Medial Prefrontal Cortex: Adding Value
to Imagined Scenarios
Wen-Jing Lin, Aidan J. Horner, James A. Bisby, and Neil Burgess
Abstracto
■ The medial prefrontal cortex (mPFC) is consistently impli-
cated in the network supporting autobiographical memory.
Whereas more posterior regions in this network have been re-
lated to specific processes, such as the generation of visuospa-
tial imagery or the association of items and contexts, el
functional contribution of the mPFC remains unclear. Cómo-
alguna vez, the involvement of mPFC in estimation of value during
decision-making suggests that it might play a similar role in
memory. We investigated whether mPFC activity reflects the
subjective value of elements in imagined scenarios. Participantes
in an MRI scanner imagined scenarios comprising a spatial con-
texto, a physiological state of need (p.ej., thirst), and two items that
could be congruent (p.ej., drink) or incongruent (p.ej., alimento) con
the state of need. Memory for the scenarios was tested outside
the scanner. Our manipulation of subjective value by imagined
need was verified by increased subjective ratings of value for
congruent items and improved subsequent memory for them.
Consistent with our hypothesis, fMRI signal in mPFC reflected
the modulation of an item’s subjective value by the imagined
physiological state, suggesting the mPFC selectively tracked
subjective value within our imagination paradigm. Further anal-
yses showed uncorrected effects in non-mPFC regions, incluir-
ing increased activity in the insula when imagining states of
need, the caudate nucleus when imagining congruent items,
and the anterior hippocampus/amygdala when imagining subse-
quently remembered items. We therefore provide evidence that
the mPFC plays a role in constructing the subjective value of the
components of imagined scenarios and thus potentially in
reconstructing the value of components of autobiographical
recollection. ■
INTRODUCCIÓN
Autobiographical memories (AMs) define who we are
and depend on a network of brain regions including
the hippocampus, parahippocampal gyrus, retrosplenial
corteza, posterior parietal cortices, and medial prefron-
tal cortex (mPFC; p.ej., Addis, Moscovitch, Crawley, &
McAndrews, 2004; Piolino et al., 2004; Maguire, 2001; Nadel
& Moscovitch, 1997). Research into the neural mecha-
nisms underlying AM has focused on closely related con-
cepts of imagery for spatial context (p.ej., Burgess, Maguire,
& O’Keefe, 2002), “scene construction” (p.ej., Hassabis,
Kumaran, & Maguire, 2007), “episodic future thinking”
(p.ej., Addis, Wong, & Schacter, 2007), “self-projection”
(Buckner & Carroll, 2007), and item-to-context binding
(Eichenbaum, Yonelinas, & Ranganath, 2007). Además
to the long-recognized hippocampal role in AM (Howard &
Eichenbaum, 2013; Squire & Zola-Morgan, 1991; O’Keefe &
Nadel, 1978; Scoville & Milner, 1957), this research has pro-
posed specific functional roles for posterior brain regions.
The parahippocampus, retrosplenial cortex, and the rest of
Papez’s circuit have been ascribed roles in the genera-
tion of visuospatial imagery (Byrne, Becker, & Burgess,
2007), whereas medial-temporal regions have been impli-
cated in storing items and context beyond the spatial
University College London
domain (Eichenbaum et al., 2007). Además, lateral pa-
rietal and prefrontal areas have been ascribed roles in atten-
tional and working memory components of AM tasks
( Johnson, suzuki, & Rugg, 2013; Rugg & Vilberg, 2013;
Cabeza, Ciaramelli, Olson, & Moscovitch, 2008; simons
et al., 2008; Wagner, shannon, Kahn, & Buckner, 2005).
Sin embargo, less is known regarding the functional role
of mPFC in AM. In decision-making, mPFC responses are
believed to represent the subjective value of chosen items
relative to potential alternatives (Rushworth, Noonan,
Boorman, Walton, & Behrens, 2011). Activity in mPFC is
correlated with the value of the chosen item, irrespective
of whether the items are food (Gross et al., 2014; Hare,
Camerer, & Rangel, 2009), agua (Bouret & Richmond,
2010), monetary reward (Nicolle et al., 2012; Boorman,
Behrens, lana rica, & Rushworth, 2009), physical action,
engaging activities (Gross et al., 2014), or abstract figures
(Glascher, Hampton, & O’Doherty, 2009). The mPFC is
also associated with self-referential thought, incluido
memory (Levin, 2004; Macrae, Moran, Heatherton,
Banfield, & kelly, 2004; Vogeley et al., 2004; Johnson
et al., 2002; Gusnard, Akbudak, Shulman, & Raichle, 2001),
leading to the recent suggestion that ventromedial pFC
(vmPFC) helps to establish the personal value, affective
quality, or significance of self-related information (Benoit,
Szpunar, & Schacter, 2014; D’Argembeau, 2013; Lebreton
et al., 2013).
© 2015 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 no portado (CC POR 3.0) licencia
Revista de neurociencia cognitiva 27:10, páginas. 1957–1967
doi:10.1162/jocn_a_00836
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
oh
3
C
3
norte
0
_
6
a
/
_
j
0
oh
0
C
8
norte
3
6
_
a
pag
_
d
0
0
b
8
y
3
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
.
F
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Given the association between memory and imagery, él
is interesting that imagery can interact with subjective
value and can influence our motivation for satisfying
basic needs, such as food consumption (larson, Redden,
& Elder, 2014; Morewedge, Huh, & Vosgerau, 2010).
Además, imagining future scenarios can influence
decision-making by changing the subjective value of
choices (Lebreton et al., 2013; Benoit, Gilbert, &
Burgess, 2011; Peters & Büchel, 2010). De este modo, imagining
oneself in a hungry state may raise the subjective value
of food items. En cambio, human memory can be influ-
enced by the value or motivational salience of the to-be-
remembered stimuli (Erwin & Ferguson, 1979). Para
instancia, fasting people have enhanced memory for food
pictures (morris & Dolan, 2001). De este modo, we infer that
memory for items could also be modulated by their value
in imagined scenarios.
Building on these previous studies, we hypothesized
that mPFC plays a role in AM and self-related imagery by
providing the subjective value of elements of a scene—a
function not ascribed to more posterior parts of the AM
network. To test this hypothesis, we designed a paradigm
in which the subjective value of items within imagined
scenarios could be manipulated experimentally during
resonancia magnética funcional. We required participants to imagine being in a cur-
rent context and state (as opposed to imaging a future
scenario, see Benoit et al., 2014) and subsequently imag-
ine seeing, but importantly not consuming (cf. Bruto
et al., 2014), objects that were congruent or incongruent
with the imagined state of need. We reasoned that the
imagined current state of need would modulate the sub-
jective value of the unconsumed objects and that mPFC
activity would correlate with this state-modulated subjec-
tive value.
MÉTODOS
Participantes
Twenty right-handed participants were recruited from
the University College London student population. Uno
did not finish the task, so the data reported here concern
the remaining 19 Participantes (12 women). The mean age
of the remaining participants was 21.7 años (DE = 2.68,
range = 19–27). All participants gave written informed
consent to participate, in accordance with the local ethics
committee (1825/003). One participant did not complete
the postscan memory task, so the results from the mem-
ory analyses are based on 18 Participantes.
Stimuli and Design
Four different physiological states of need were used:
thirst, coldness, hunger, and tiredness. A neutral state
was used as a baseline condition (instruction for neutral
estado: Imagine you are just fine. You are not in any state
of need but just in an ordinary condition.). Twelve spatial
contexts were used: beach, cocina, desert, campos, class-
habitación, airplane, forest, oficina, library, playground, church,
and ship. These were included to make the imagined sce-
narios more realistic and because, without instruction,
participants would be likely to imagine uncontrolled
backgrounds to facilitate imagery. Había 60 state–
context combinations, with each appearing only once
durante el 60 trials of the imagery task.
Pictures from four categories were used as items; cada
category contained items that were usually used to satisfy
one of the four physiological states of need. The first cat-
egory contained water, juice, beer, and other beverages
used to quench thirst. The second category contained
items that were able to be used to help people resist cold
clima, such as fireplace, hot drink, and winter clothes.
Another category contained food, and the final category
contained items used for taking a rest or relieving tired-
ness included a bed, couch, bathtub, etcétera. Allá
eran 180 item pictures in total, consisting of 45 pictures
per category. Among these pictures, 120 appeared in the
imagery task and another 60 served as new items during
an old–new recognition test. The assignment of pictures
to old items and new items was counterbalanced across
Participantes. All pictures were obtained from FreeDigital-
Photos.net (www.freedigitalphotos.net/).
In the imagery task, each trial contained one state–context
combination presented as cue words and also two item
pictures (see Figure 1A for an example of trial presenta-
tion order). The relationship between the participant’s
current imagined state and each item picture during a
single trial could either be congruent or incongruent.
For a congruent item, the type of item presented would
meet the participant’s current need created by the imag-
ined state. Por ejemplo, a food picture would be classified
as congruent if the state was hunger, but incongruent if
the state was tired, cold, or thirsty. Note that “incongru-
ent” items were irrelevant rather than opposite to the
current state of need. Ambiguous items were never used
as “incongruent items” (p.ej., a hot drink was not used
in thirst trials). From the two item pictures, sequentially
presented during each trial, either item could be congru-
ent or incongruent with the current state. This provided
four possible combinations: congruent–congruent,
incongruent–incongruent, congruent–incongruent, y
incongruent–congruent. En tono rimbombante, all four combina-
tions of items occurred in pseudorandom order across
ensayos, allowing us to identify the effects of an individual
items’ subjective value, as modulated by its congruency
with the imagined state. Among the 120 item pictures
presented during the imagery task, 24 served as neutral
pictures as they occurred in a neutral state. An alternative
would be to use items unrelated to any of the physiolog-
ical states, but such items would be intrinsically different
to the congruent items in the study. El restante 96
pictures were equally assigned as congruent or incongru-
ent items.
1958
Revista de neurociencia cognitiva
Volumen 27, Número 10
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
oh
3
C
3
norte
0
_
6
a
/
_
j
0
oh
0
C
8
norte
3
6
_
a
pag
_
d
0
0
b
8
y
3
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
F
/
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
oh
3
C
3
norte
0
_
6
a
/
_
j
0
oh
0
C
8
norte
3
6
_
a
pag
_
d
0
0
b
8
y
3
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
F
t
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
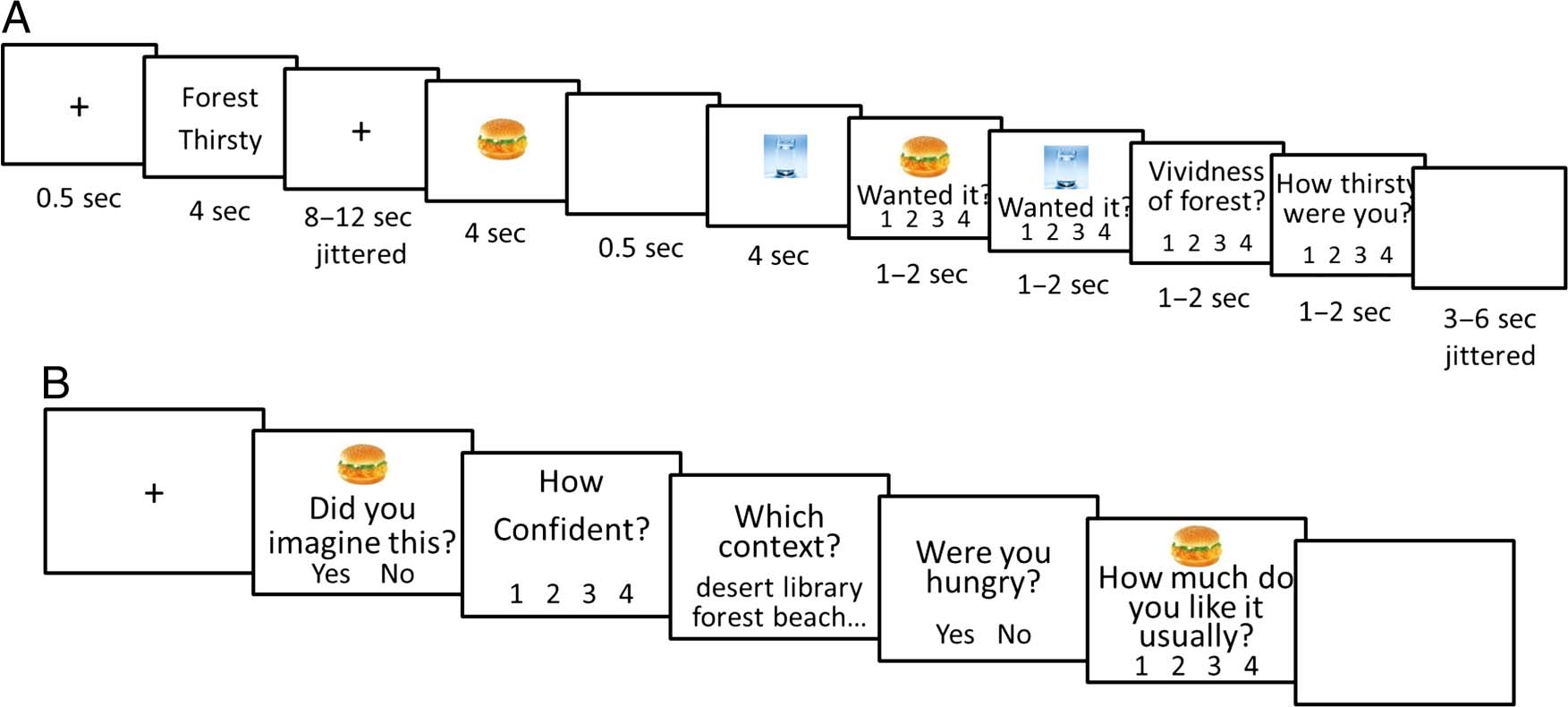
Cifra 1. Procedure of the imagery task (A) and the memory task (B).
Procedimiento
Imagery Task
Participants were provided with task instructions before
scanning and completed a number of practice trials out-
side the scanner. The entire imagery task, consisting of
60 imagery trials, was equally divided into two sessions,
and scanning lasted for 1 hr in total, including acquisition
of a structural scan. See Figure 1A for an illustration of
stimulus presentation for the imagery task. Each trial be-
gan with a fixation cross at the center of the screen,
which was replaced by a pair of state–context cue words
después 0.5 segundo. Participants were instructed to vividly imag-
ine the context and state according to the cue words pro-
vided. The state–context cue words were presented for
4 segundo, and then a fixation cross appeared again (for 8–
12 segundo, jittered), during which the participants were in-
structed to continue imagining. Próximo, two pictures were
presented sequentially, each for 4 sec separated by a
0.5-sec blank screen. Participants were required to incor-
porate each presented item into their imagined scenario
during the trial. Participants were explicitly instructed
to not imagine consuming these items to satisfy their
imagined state and its associated need. Por ejemplo, ellos
were required to imagine seeing (but not consuming) a
chicken burger in a forest while they were thirsty (as in
Figura 1A). After a further blank screen (1–4 sec, jit-
tered), participants made four simple ratings, one at a
tiempo. The first two ratings asked participants to rate
how much they had wanted each item when they initially
saw it during the trial. The last two separately rated how
vividly they had imagined the current state and context.
All ratings used a 4-point scale (1 = not at all, 4 = very
mucho). Each trial ended with a final blank screen (3–
6 segundo, jittered). Visual stimuli were presented by MATLAB
(The MathWorks, Natick, MAMÁ) and COGENT 2000 toolbox
(www.vislab.ucl.ac.uk/cogent.php).
Memory Task
The memory task took place outside the scanner after
the imagery task was completed. Each trial consisted of
a 500-msec fixation cross followed by a picture of an item,
and participants were required to judge whether the pic-
ture had been presented in the imagery task or not (es decir.,
old/new item recognition judgment) and how confident
they were of their answer (Figure 1B shows an illustration
of the memory task). If participants answered “new,” par-
ticipants were then asked how much they like that item
in their daily lives. If an item was judged “old,” two fur-
ther source memory questions were presented to the
participant to test memory for the associated state and
contexto. To test state, one of the state words (hunger,
thirst, tired, cold, or neutral) was presented, and partici-
pants judged whether that state was the one they had
been asked to imagine when the recognized item picture
had appeared in the imagery task. The correct answer
was yes for 50% of trials, and within these trials, 40% de
the state words were congruent with the tested item,
40% were incongruent, y 20% were neutral. Para el
context source memory test, todo 12 of the contexts were
listed to allow participants to select the one which had
accompanied the recognized item picture. The trial ended
with the daily subjective rating. Había 180 memory
trials in total (120 with “old” items and 60 with “new”
elementos). Twelve alternative forced choice is an efficient
way to test memory for the spatial context of an item’s
presentation but could not be used to test memory for
the physiological state, because a simple strategy of
guessing the congruent state would artificially inflate per-
rendimiento (p.ej., choosing “thirst” when presented with a
drink). In this situation, choosing a congruent state
would be correct in 40% of trials, a neutral state would
be correct in 20% of trials, and the three incongruent
states would be correct in 13% of trials. To avoid this,
Lin et al.
1959
we tested participants with yes/no cued recognition of a
single state that was chosen to be correct 50% del
tiempo, irrespective of its congruence with the item.
fMRI Data Acquisition and Preprocessing
Functional imaging was performed on a 3T scanner
(Siemens TIM Trio, Siemens, Berlina, Alemania) durante
the imagery task. The functional data were acquired with
a gradient-echo EPI sequence (repetition time = 3.36 segundo,
echo time = 30 mseg, flip angle = 90°, resolution = 3 × 3 ×
3 mm, 64 × 74, 48 slices per volume). El numero total de
volumes in each run varied across participants because of
the variation of RT for each rating (the mean number of vol-
umes was 332 per session). A high-resolution T1-weighted
3-D structural image (1 mm3) was acquired after two ses-
sions of functional scans. A double-echo FLASH fieldmap
sequence was also recorded.
Functional images were processed and analyzed with
SPM8 (Wellcome Trust Centre for Neuroimaging, Londres,
Reino Unido, www.fil.ion.ucl.ac.uk/spm/software/spm8/). The first
five volumes of each scan were discarded for T1 equilibra-
ción. Preprocessing procedures included bias correction,
realignment, unwarping, coregistration, slice timing cor-
rection, and normalization to the MNI template using the
Dartel toolbox. EPI images were smoothed with an iso-
tropic 8 mm FWHM Gaussian kernel. One of the partici-
pant’s fieldmap scan was not collected, so the unwarping
procedure was skipped in their data.
Análisis de los datos
The preprocessed functional images were analyzed with
general linear models (GLMs). We estimated five GLMs
for different purposes. All GLMs included six movement
regressors for each session, estimated during realign-
mento, as well as two further regressors modeling each
session. On the basis of our strong a priori hypothesis
about the mPFC and vmPFC, we performed small-volume
correction (SVC) within a combined anatomical mask
of these regions: bilateral mPFC and vmPFC (volumen
∼ 53,493 mm3). This mask was derived from the AAL atlas
(Tzourio-Mazoyer et al., 2002), as implemented in
the WFU PickAtlas Tool (Maldjian, Laurienti, kraft, &
Burdette, 2003). This mask contained superior frontal gy-
rus, medial frontal gyrus, anterior cingulate, and cingulate
gyrus. Within this small volume, we report effects that sur-
vive p < .05 FWE correction. For completeness, we also
report effects at p < .001 uncorrected across the whole
brain; however, caution is needed in interpretation of
these effects.
The first model (GLM1) was a parametric modulation
analysis, searching for regions that correlated with the
subjective value of an item during imagined states of
need. The first-level model contained seven regressors
per session: (1) imagining a state of need, (2) imagining
a neutral state, (3) imagining an item in a state of need,
(4) a parametric modulator of the item regressor based on
the participant’s subjective value of each item, (5) imagin-
ing an item in a neutral state, (6) intertrial interval (ITI)
periods, and (7) key presses. Trial periods were modeled
with a boxcar function for the entire length of each period
(e.g., the 4 sec of imaging an item), convolved with the
canonical hemodynamic response function. The second-
level analysis was a one-sample t test on the parameter
estimates from the parametric modulator (Regressor 4)
averaged across the two sessions. For the parametric
modulation, we used the subjective rating of each item
when imagined in the state of need of the current trial mi-
nus the subjective rating of the item in the participant’s
daily life, given after the scanning session. This calculation
allowed us to control for variations in the participants’
baseline preference for the various items. The range of
these normalized subjective ratings was from −3 to 3.
The second model (GLM2) was used for comparing
imagination of congruent items versus incongruent items
(given that the first GLM collapsed across these condi-
tions to maximize power in our parametric modulation
analysis) and also for comparing imagining states of
needs versus neutral states. This model included seven
regressors per session: (1) imagining a state of need,
(2) imagining a neutral state, (3) imagining a congruent
item in a state of need, (4) imagining an incongruent
item in a state of need, (5) imagining an item in a neutral
state, (6) ITI periods, and (7) key presses. Parameter es-
timates for regressors (1) to (4) were averaged across the
two sessions and entered into a second-level model. A
separate regressor was also included for each individual
subject that consisted of a “1” for each condition for that
specific participant (i.e., subject effects). A third model
(GLM3) aimed to test the subsequent memory effect
for imagined items. The model was similar to GLM1 but
replaced the subjective value parametric modulator with
a modulator based on subsequent memory. The model
included six regressors per session: (1) imagining a state
of need, (2) imagining a neutral state, (3) imagining an
item (in either a state of need or neutral state), (4) a para-
metric modulator of the previous regressor based on
subsequent memory for the item, (5) ITI periods, and
(6) key presses. Note that the parametric modulator for
subjective value was applied to item imagination during a
state of need, not during neural states, as we were specif-
ically interested in how states of need modulated subjec-
tive value. The parametric modulator for subsequent
memory was applied to all item imagination trials (includ-
ing neutral states) to maximize power. Subsequent mem-
ory was parameterized as a transformed confidence rating
to maximize sensitivity. Participants’ 1–4 confidence rat-
ings for old and new items at test were transformed into
a measure of successful memory performance by com-
bining ratings for item “hits” with negative ratings for
item “misses” (e.g., a “miss” given a confidence rating
of 4 would become −4 in the parametric modulator).
The second-level analysis was a one-sample t test on
1960
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the parameter estimates from the parametric modulator
(Regressor 4) averaged across the two sessions.
The final two models (GLM4 and GLM5) aimed to test
the subsequent memory effect for the state of need
(GLM4) and the context (GLM5) in which items were
imagined (i.e., two types of source memory). GLM4 con-
tained seven regressors per session: (1) imagining a state
of need, (2) imagining a neutral state, (3) item imagina-
tion trials for which the item and state of need are sub-
sequently remembered, (4) item imagination trials for
which the item but not the state is remembered, (5) item
imagination trials for which the item is not remembered,
(6) ITI periods, and (7) key presses. GLM5 was similar
to GLM4 but split item imagination trials (Regressors 3–
5) by whether the context (rather than the state) was re-
membered. Second-level models for each GLM were
paired t tests comparing either state or context hits ver-
sus misses (Regressors 3 and 4) averaged across the two
sessions.
Note that we built separate GLMs for each analysis of
interest. This was due to the overlapping nature of cer-
tain regressors. In particular, the categorical congruent
versus incongruent contrast correlated with the related,
but more sensitive, item-by-item parametric modulation
of value by state. Furthermore, the parametric modula-
tors relating to subsequent memory and subjective value
were also correlated. Despite the overlapping nature of
these regressors of interest, our separate GLMs revealed
distinct patterns of activity.
RESULTS
Behavioral Results
The Subjective Value of Items in Imagery
To demonstrate that our manipulation of imagined state
worked, a three-way repeated-measure ANOVA with
Situation (two levels: everyday rating and rating during
imagery), Rating (1–4), and category (congruent,
incongruent, and neutral) was performed. The three-
way interaction was significant (F(6, 102) = 18.70, p <
.001), so we performed further analyses that revealed that
the distributions of ratings differed between categories for
ratings during imagery (Rating × Category, F(6, 102) =
27.40, p < .001), but not for everyday ratings (Rating ×
Category, F(6, 102) = .66, p = .68). Thus, it was only when
participants imagined being in a specific state of need that
the subjective value of the objects differed between our
“congruent” and “incongruent” conditions. Table 1 shows
that a greater proportion of congruent items had positive
subjective value (controlling for baseline value, i.e., rating
of imagined value—everyday rating; 39.67%) whereas
most incongruent items had negative subjective values
(60.25%). This suggests that our participants indeed
followed the instruction to imagine the assigned state of
need and that those imagined states influenced the subjec-
tive value of the item on that trial.
We also carried out a two-way repeated-measure
ANOVA with Congruency between the state question word
and item (congruent and incongruent) and Rating (1–4) as
within-subject variables to test whether the preceding
state question might bias ratings (e.g., “hungry” increasing
ratings for food items). There was no significant in-
teraction between Congruency and Rating (F(3, 51) =
.40; p = .75), suggesting that the everyday value ratings
were not influenced by the preceding source memory
questions.
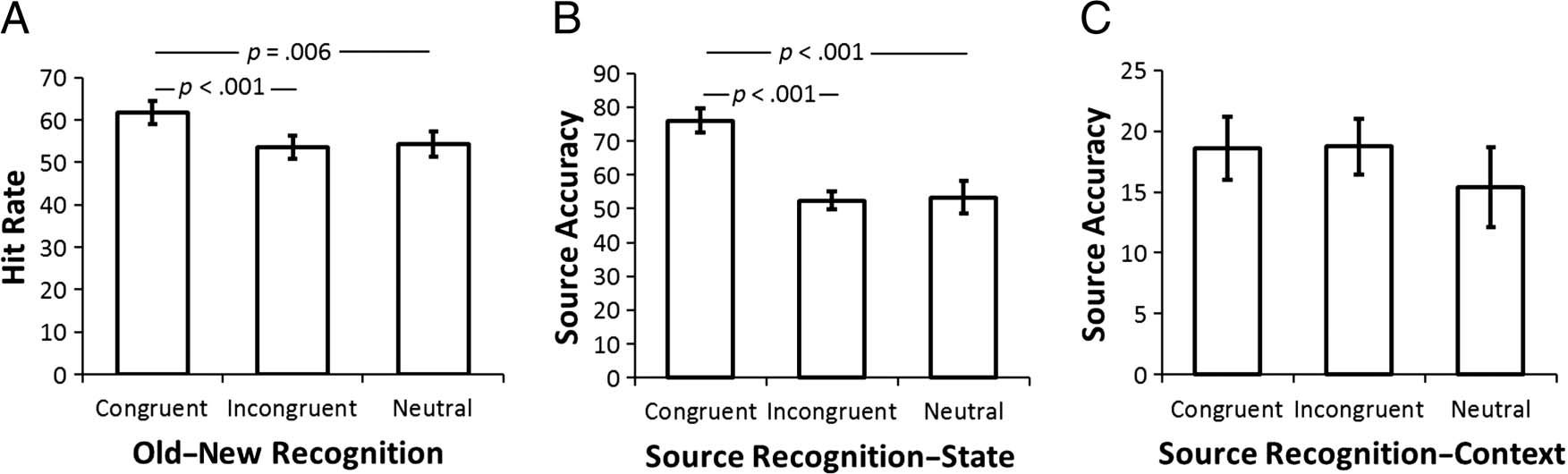
Old–New Recognition
A one-way repeated-measure ANOVA across Congruency
(congruent, incongruent, and neutral) was carried out to
test for differences in hit rate among different categories
of items. The results revealed a significant main effect of
congruency (F(2, 34) = 9.01, p < .001; see Figure 2A for
memory performance). Pairwise comparisons showed
that hit rate was higher for congruent items than for in-
congruent (t(17) = 5.16, p < .001) and neutral (t(17) =
3.14, p = .006) items. However, there was no signif-
icant difference between incongruent and neutral items
(t(17) = .35, p = .73). This result suggests that partici-
pants had better memory for items that were able to ful-
fill their needs in the imagined state. Participants showed
a high correct rejection rate for new items (87%). Table 2
shows confidence ratings across all responses.
For completeness, we checked whether our results
varied with the order in which items were presented
within a trial. We ran a two-way repeated-measure ANOVA
with Order of presentation (two levels: first or second) and
Category (three levels: congruent, incongruent, and neu-
tral) as within-subject factors on the subjective ratings
and subsequent memory scores. The results show that
the order of presentation during encoding did not affect
item memory (Order, F(1, 17) = .20, p = .66; Category,
Table 1. Percentage of Subjective Values of Items during Imagery (−3 to +3), according to Whether They Were Imagined in a
Congruent, Incongruent, or Neutral State of Need, Controlling for Baseline Value ( Value during Imagery—Everyday Value)
Congruent
Incongruent
Neutral
−3
2.36%
6.63%
6.35%
−2
5.20%
18.64%
18.03%
−1
17.66%
34.98%
30.90%
0
35.10%
27.31%
28.32%
+1
23.27%
8.93%
11.43%
+2
11.95%
2.94%
4.97%
+3
4.45%
0.58%
0.00%
Lin et al.
1961
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Behavioral results for the memory task. (A) Mean values of hit rate in the item recognition memory task. (B and C) Mean performance in
the source recognition task for the state of need (B) and the spatial context (C). Error bars represent ±1 SEM.
F(2, 34) = 9.22, p = .001; Order × Category, F(2, 34) =
1.89, p = .17), and there was a nonsignificant trend to-
ward lower ratings for the first item versus the second
item (Order × Category, F(2, 34) = 1.05, p = .36;
order, F(1, 17) = 3.92, p = .06).
Source Memory
Source memory performance for correctly associating the
imagined state with the recognized item was analyzed
using a one-way ANOVA across levels of Congruency.
We found a significant main effect of Item congruency
(F(2, 34) = 17.30, p < .001). Pairwise comparisons
showed that the conditional state source performance
hit rate (% correct source memory for the state associ-
ated with items correctly recognized as “old”) for congru-
ent items was significantly higher than for incongruent
items (t(17) = 6.16, p < .001) and neutral items (t(17) =
5.44, p < .001), whereas there was no significant differ-
ence between the latter two categories (t(17) = .17,
p = .864; see Figure 2B). Although participants showed
a response bias toward accepting the state (answering
“yes”) when it was congruent (55.6% responses were
yes) or neutral (54.2% yes) relative to the item and
“no” when it was incongruent (41% responses for incon-
gruent items were no), this response bias could not ac-
count for our results (the correct proportion of “yes”
responses being 50% in both cases).
Analysis of source memory performance for the imag-
ined spatial context (e.g., “beach”) within the recognized
Table 2. Percentage of Hits, Misses, False Alarms, and Correct
Rejections across Confidence Ratings (1–4) in the Old–New
Item Recognition Task
Hit
3.9
14.1
20.1
61.9
Miss
16.7
19.8
26.8
36.7
1
2
3
4
False Alarm
Correct Rejection
17.0
29.1
29.1
24.8
10.4
15.3
23.5
50.7
item showed no significant main effect of Item congru-
ency (F(2, 34) = .889, p = .42; see Figure 2C for context
source memory performance). It is possible that this re-
flects the irrelevance of spatial context to the subjective
ratings that the participants are required to give on each
trial or that any small effects of congruency on context–
source memory were obscured by low levels of per-
formance (chance = 8%) although performance was
above chance in each category (congruent: t(17) =
3.96, p = .001; incongruent: t(17) = 4.48, p < .001; neu-
tral: t(17) = 2.14, p = .047).
In general, behavioral results supported our predic-
tion. Subjective values of items support the validity of
our imagined need paradigm. We also saw greater recog-
nition performance for congruent than incongruent
items and better memory for the imagined state of con-
gruent than incongruent items. Thus, we observed better
memory performance for items when their value was
congruent with the imagined state.
fMRI Results
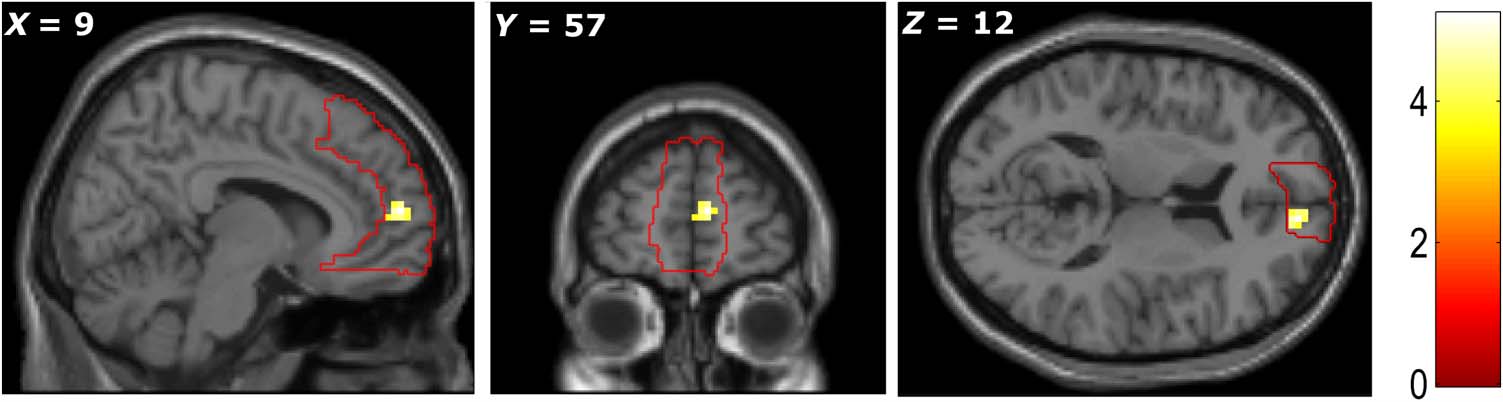
Subjective Value of Items in Imagery (GLM1)
First, we focused on the main prediction of our study:
that the subjective value of items in imagined scenarios
would correlate with the BOLD response in the mPFC.
To isolate imagined value from differences in the intrinsic
values of the items used, we calculated the participant’s
subjective value for the item when imagining it in the cur-
rent state of need minus their subjective value for the
same item in their daily life. This parametric modulator
revealed an effect in the mPFC (+9, +57, +12, Z =
3.98; p < .05 FWE SVC). We therefore provide evidence
that mPFC represents the values of elements in imagined
scenarios, controlling for variations in their intrinsic value
in other situations (Figure 3).
Given the complexity of our imagination task, it is im-
portant to rule out other explanations for our main mPFC
result. This is particularly important given the overlapping
nature of certain experimental factors (see Methods). In
short, none of our subsequent analyses showed an effect
1962
Journal of Cognitive Neuroscience
Volume 27, Number 10
Figure 3. The activation of
mPFC during imagination of an
object during a state of need
varied according to the extent
to which the subjective value of
item was modulated by the
imagined state of need. All
peaks significant at p < .001,
uncorrected (color bar indicates
t statistic). (The red line depicts
the area of mPFC mask used in
SVC analysis.)
in mPFC, even at a lenient p < .001 uncorrected thresh-
old. However, these analyses did reveal effects in other re-
gions at this threshold. We report these results for
completeness but note that they should be treated with
caution given that they do not survive correction for mul-
tiple comparisons.
and second item, finding a nonsignificant trend toward a
greater effect of state congruency for the first versus sec-
ond item in the vmPFC (−3, 33, −12; p = .083 FWE
SVC). However, these could not influence the findings
themselves, as our manipulation of state congruency
was counterbalanced across items.
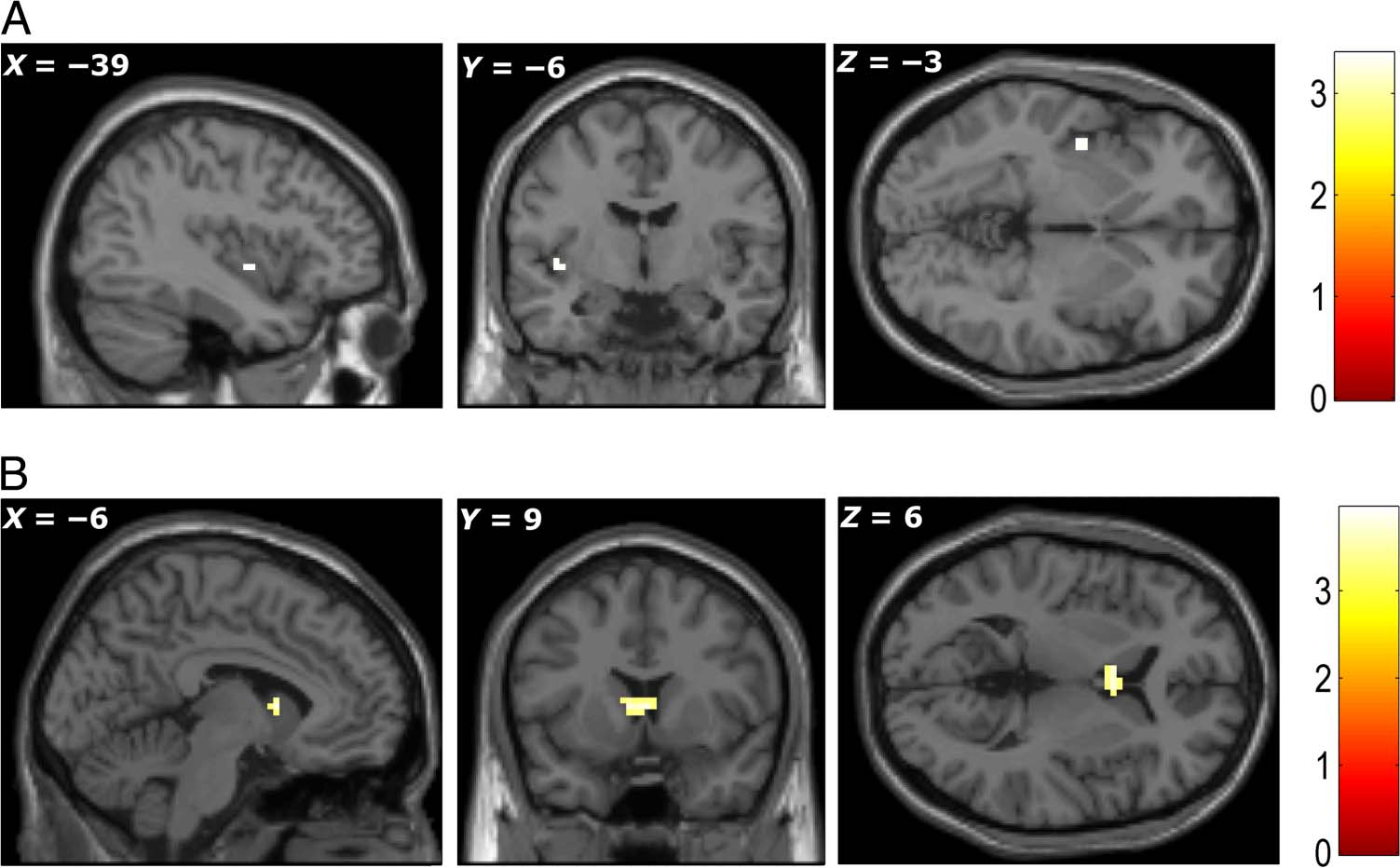
Imagining States of Need and Item Congruency with
Need (GLM2)
Compared with imagination of a neutral state, imagina-
tion of states of physiological need showed greater activa-
tion in bilateral insula (MNI coordinates of peak
activations: −39, −6, −3, Z = 3.27; +45, +15, +3, Z =
3.15; p < .001, uncorrected; Figure 4A). By contrasting
imagery for congruent versus incongruent items, we iden-
tified a region in the basal ganglia—the caudate nucleus
(+3, +9, +6, Z = 3.60; −6, +9, +6, Z = 3.56, p < .001,
uncorrected; Figure 4B). Because congruent items had
higher subjective value than incongruent ones, we also car-
ried out an SVC analysis for the congruent–incongruent
contrast in the mPFC ROI but found no significant effect.
We also investigated whether the fMRI correlates of an
item’s value or state congruency varied between the first
Subsequent Memory Effects (GLM3)
This parametric modulation analysis showed that BOLD
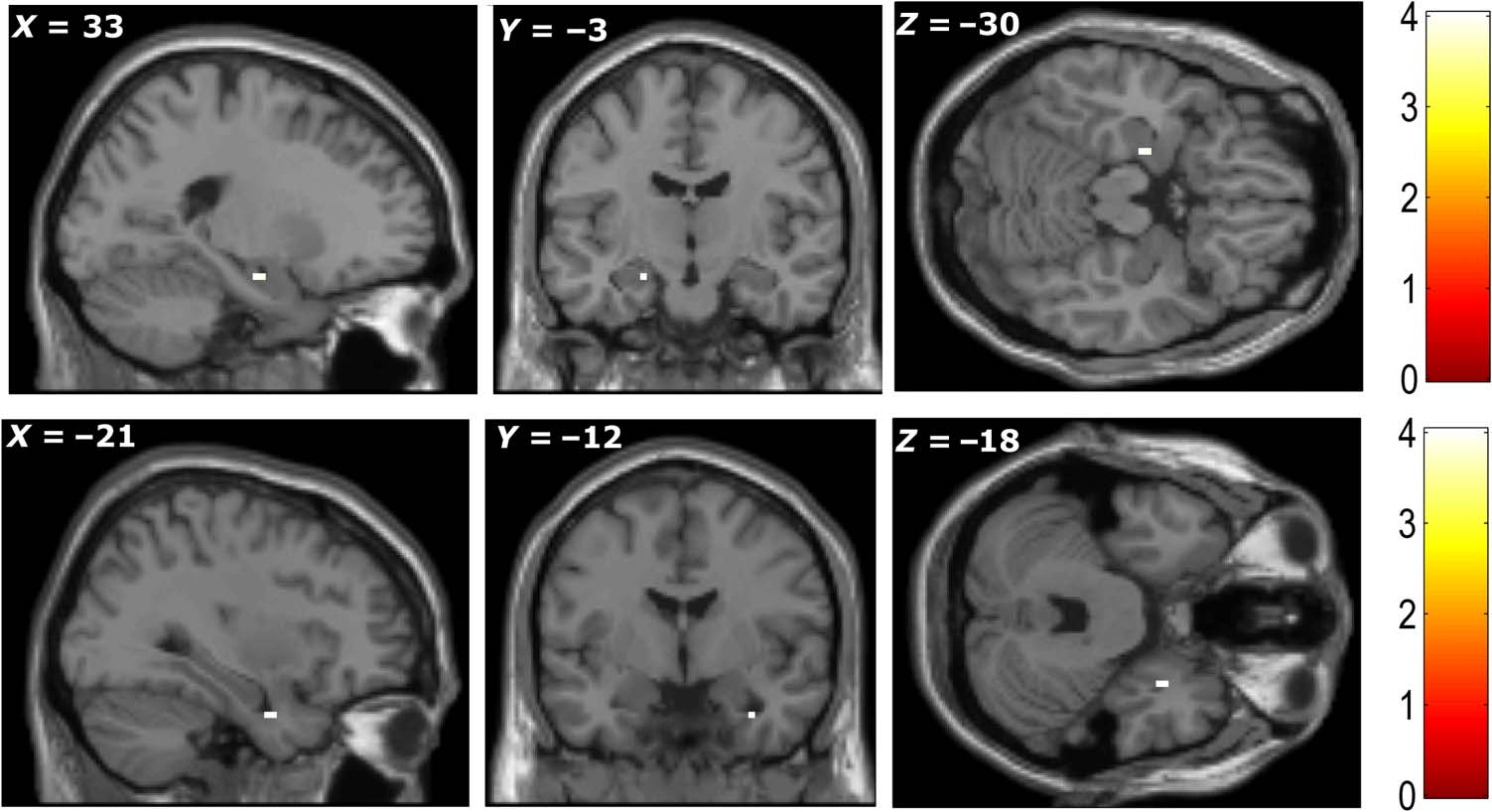
signal in the right amygdala (+33, −3, −30; Z = 3.27)
and left anterior hippocampus (−21, −12, −18; Z =
3.33), when participants were imagining items, were sig-
nificantly correlated with participants’ subsequent mem-
ory ( p < .001, uncorrected; Figure 5). Note that our
subsequent memory modulator combined categorical
subsequent memory status (i.e., hits and misses) with
subjective confidence, revealing linear increases in BOLD
response from −4 (high confidence misses) to +4 (high
confidence hits). No other significant activity was re-
vealed in this analysis.
No significant activations were found corresponding to
subsequent source memory effects for state (GLM4) or
for context (GLM5), that is, the comparisons of imagery
Figure 4. (A) Bilateral insula
showed higher activation when
participants were imagining
states of need compared to
imagining the neutral state.
(B) The caudate showed greater
activation for imagining a
state-congruent item than a
state-incongruent item. All
peaks significant at p < .001,
uncorrected (color bars indicate
t statistic).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Lin et al.
1963
Figure 5. Greater activity
was seen during encoding in
left hippocampus (top row)
and right amygdala (bottom
row) for items that were
subsequently correctly
recognized with high
confidence compared to
subsequently nonrecognized
items. All peaks significant at
p < .001, uncorrected (color
bars indicate t statistic).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
for items that became source hits versus source misses.
This may reflect a lack of power, given the relatively low
trial numbers in specific conditions (i.e., source misses
for state), and the absence of a parametric measure like
the confidence ratings used for item memory.
DISCUSSION
We were interested in the potential role of mPFC in con-
tributing subjective value to the contents of imagery. Our
paradigm provides a way to measure this by manipulating
subjective value of imagined items with respect to imag-
ined physiological need. The behavioral results suggest
that the manipulation was valid, and the imaging results
support the hypothesis that mPFC activity reflects the
subjective value of elements in imagined scenarios.
The manipulation of imagined need succeeded in al-
tering the subjective value of elements within imagined
scenarios in that participants indicated higher ratings
for items congruent with (i.e., likely to satisfy) the state
of need. Subsequent recognition memory for items also
supports the success of our manipulation. Items that
were able to fulfill people’s imaginary needs showed
greater subsequent memory, both in being better recog-
nized and being better associated to the state of need in
which they were presented. This could be because to
imagine a congruent item in the imagined scenario is
more consistent with our daily life experiences and this
enabled participants to have a richer imagination. Simi-
larly, congruent items might fit more readily into a preex-
isting “schema” allowing for a more rapid integration of the
item and imagined state (Tse et al., 2007; Bransford &
Johnson, 1972; Bartlett, 1932). Equally, congruent items
might have been better remembered because more valu-
able scenarios tend to be more strongly represented in
memory-related areas (Lebreton, Jorge, Michel, Thirion,
& Pessiglione, 2009; Wittmann et al., 2005).
The instruction to imagine states of physiological need
was accompanied by increased activity in the insula com-
pared to neutral states, albeit at an uncorrected threshold.
This would be consistent with studies showing insular
activation corresponding to interoception of actual physi-
ological states (Craig, 2003), including thermo sensa-
tion (Craig, Chen, Bandy, & Reiman, 2000) and hunger
(Tataranni et al., 1999). One might wonder whether peo-
ple are able to imagine themselves in different physiolog-
ical states, because physiological states are not usually
Table 3. Whole-brain fMRI Analysis Results
Region
Cluster Size
x
y
z
Peak Z Score
The Subjective Value of Items in Imagery
Right mPFC
Anterior cingulate
Left ventral mPFC
18
6
8
9
0
57
12
27 −9
−12
45
3
3.98*
3.1
3.63
Imagine States of Need >Imagine Neutral State
Left insula
Right insula
6
1
−39 −6 −3
45
15
3
3.27
3.15
Congruent >Incongruent
Caudate
21
3
9
6
3.6
Subsequent Memory Effect
Left hippocampus
Right amygdala
3
2
−21 −12 −18
33 −3 −30
3.33
3.27
All peaks reached an uncorrected significant level of p = .001.
*p value was <.05 at the cluster level with SVC.
1964
Journal of Cognitive Neuroscience
Volume 27, Number 10
thought to be under cognitive control. However, involun-
tary physiological signs can be influenced by imagination,
for example, pupil dilation can be affected by imagining
dark or light environments (Laeng & Sulutvedt, 2014).
We were interested in the process by which subjective
value is afforded to an item within an imagined scenario.
To investigate this, we looked for an fMRI signal match-
ing the modulation of an item’s subjective value by the
imagined state of need, that is, a regressor formed from
the subjective rating of the item when imagined as part of
a specific scenario minus the subjective rating of that
item in daily life. We found activity following this pattern
in mPFC, both in a more superior region and the ventral
region of mPFC (albeit at an uncorrected threshold for
the latter region; see Table 3). This is consistent with
our hypothesis for the role of mPFC in imagery. Thus,
beyond the representation of the subjective value of
choices in decision-making, the mPFC may also play a
role in representing the value of items in imagined sce-
narios more generally. This more general role might be-
gin to explain its involvement in AM retrieval or episodic
future thinking, as well as tasks with an implied compo-
nent of choice such as planning. Indeed, mPFC activation
has been seen together with hippocampal activation
during the imagination of rewarding future situations in
a decision task (Lebreton et al., 2013).
In general, congruent items were rated as more valu-
able than incongruent ones. Congruent items might be
valuable because of their utility in a specific context
(i.e., a congruent state; Hare, Malmaud, & Rangel,
2011) or because congruent items are more self-relevant
in a congruent state (D’Argembeau, 2013). Could the re-
sults we observed in mPFC be caused by semantic con-
gruency effect? To examine the effect of semantic
congruency itself, we simply compared the imagination
of explicitly congruent or incongruent items, finding ac-
tivity in the caudate nucleus (but not in mPFC, where the
difference in activity was some way below threshold, at
p = .06 uncorrected). Thus, there is little support for a se-
mantic interpretation of the mPFC activity we observed.
The representation of the combined scenario may involve
the striatum, via increased consolidation of the congruent
state–item association, consistent with some rodent stud-
ies of consolidation (Pennartz et al., 2004). Alternatively,
the striatal activation may reflect the involvement of these
areas in reward-related processing (e.g., Knutson, Rick,
Wimmer, Prelec, & Loewenstein, 2007), in the sense that
the imagined interaction with the congruent item seems
more rewarding in nature (although we forbade imagined
consummation of items).
The behavioral results demonstrate a higher recogni-
tion rate for congruent items. This memory effect could
relate to schema theory: perhaps the encoding of new
information (i.e., a congruent item) benefits from being
congruent rather than incongruent with the existing sce-
nario. The mPFC has been implicated in incorporating
new information into existing knowledge structures (van
Kesteren et al., 2013; van Kesteren, Ruiter, Fernández, &
Henson, 2012; Tse et al., 2011; Benchenane et al., 2010;
van Kesteren, Fernández, Norris, & Hermans, 2010). How-
ever, mPFC did not show a significant subsequent memory
effect. Subsequent memory for items was related to activ-
ity in the anterior medial-temporal lobe during encoding,
consistent with several previous studies implicating the
hippocampus (e.g., Wagner et al., 1998). Our subsequent
memory effects also extended into the amygdala. This may
be consistent with a role for the amygdala in item memory
(Farovik, Place, Miller, & Eichenbaum, 2011; Kensinger,
Addis, & Atapattu, 2011; Ranganath, 2010; Kensinger &
Schacter, 2006) or with amygdala involvement in enhanc-
ing memory for items with affective salience (Hamann, Ely,
Grafton, & Kilts, 1999) or intrinsic value as a reinforcer
(Rolls, 2005). Unfortunately, we did not have enough sta-
tistical power to analyze subsequent memory effects sep-
arately in congruent, neutral, and incongruent items to
address these possibilities.
The recollection of autobiographical information has
been associated with a network of brain regions. Al-
though many posterior regions have a hypothesized func-
tional role within this network (e.g., Schacter et al., 2012;
Hassabis & Maguire, 2009; Byrne et al., 2007; Cabeza &
St Jacques, 2007), the mPFC has received somewhat less
attention. AMs tend to be highly personal and value-laden.
For example, we are more likely to remember the experi-
ence of having a cup of hot tea after walking outdoors for
hours on a cold winter day than having a cup of tea on an
ordinary afternoon. Given its association with value in
decision-making and with the value afforded by imagined
scenarios in this study and related studies (Benoit et al.,
2014; Gross et al., 2014; Winecoff et al., 2013; Nieuwenhuis
& Takashima, 2011), mPFC activity may reflect the value of
recollected information (see also D’Argembeau, 2013).
This is perhaps one reason why mPFC is typically not seen
in more traditional episodic memory tasks, such as word
recognition, where memory for such items may be high,
but little value is associated with the retrieved items.
Indeed, the subjective value associated with items may
be one critical difference between typical autobiographical
and episodic memory tasks.
To conclude, we have developed a new paradigm for
looking at the interaction of imagery and value. We have
validated it behaviorally via subjective value ratings and
subsequent memory effects. Supporting our hypothesis,
we found activity in the mPFC corresponding to the sub-
jective value that an item is afforded by the imagined sce-
nario. This suggests an extension of the well-known role
of mPFC in representing value during decision-making
and offers a potential explanation of its involvement in
imagery and AM retrieval.
Acknowledgments
We thank the Medical Research Council U.K., the Wellcome
Trust, and the Taiwan Government scholarship for studying
Lin et al.
1965
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
abroad for funding, and the Wellcome Trust Centre for Neuro-
imaging at University College London for providing facilities.
Reprint requests should be sent to Neil Burgess, Institute of
Cognitive Neuroscience, University College London, 17 Queen
Square, London WC1N 3AR, United Kingdom, or via e-mail:
n.burgess@ucl.ac.uk.
REFERENCES
Addis, D. R., Moscovitch, M., Crawley, A. P., & McAndrews,
M. P. (2004). Recollective qualities modulate hippocampal
activation during autobiographical memory retrieval.
Hippocampus, 14, 752–762.
Addis, D. R., Wong, A. T., & Schacter, D. L. (2007).
Remembering the past and imagining the future: Common
and distinct neural substrates during event construction
and elaboration. Neuropsychologia, 45, 1363–1377.
Bartlett, F. C. (1932). Remembering. Cambridge: Cambridge
University Press.
Benchenane, K., Peyrache, A., Khamassi, M., Tierney, P. L.,
Gioanni, Y., Battaglia, F. P., et al. (2010). Coherent theta
oscillations and reorganization of spike timing in the
hippocampal-prefrontal network upon learning. Neuron,
66, 921–936.
Benoit, R. G., Gilbert, S. J., & Burgess, P. W. (2011). A neural
mechanism mediating the impact of episodic prospection on
farsighted decisions. Journal of Neuroscience,
31, 6771–6779.
Benoit, R. G., Szpunar, K. K., & Schacter, D. L. (2014).
Ventromedial prefrontal cortex supports affective future
simulation by integrating distributed knowledge.
Proceedings of the National Academy of Sciences, U.S.A.,
111, 16550–16555.
Boorman, E. D., Behrens, T. E. J., Woolrich, M. W., &
Rushworth, M. F. S. (2009). How green is the grass on the
other side? Frontopolar cortex and the evidence in favor of
alternative courses of action. Neuron, 62, 733–743.
Bouret, S., & Richmond, B. J. (2010). Ventromedial and orbital
prefrontal neurons differentially encode internally and
externally driven motivational values in monkeys. Journal of
Neuroscience, 30, 8591–8601.
Bransford, J. D., & Johnson, M. K. (1972). Contextual
prerequisites for understanding: Some investigations of
comprehension and recall. Journal of Verbal Learning and
Verbal Behavior, 11, 717–726.
Buckner, R. L., & Carroll, D. C. (2007). Self-projection and the
brain. Trends in Cognitive Sciences, 11, 49–57.
Burgess, N., Maguire, E. A., & O’Keefe, J. (2002). The human
hippocampus and spatial and episodic memory. Neuron,
35, 625–641.
Byrne, P., Becker, S., & Burgess, N. (2007). Remembering
the past and imagining the future: A neural model of
spatial memory and imagery. Psychological Review, 114,
340–375.
Cabeza, R., Ciaramelli, E., Olson, I. R., & Moscovitch, M. (2008).
The parietal cortex and episodic memory: An attentional
account. Nature Reviews Neuroscience, 9, 613–625.
Cabeza, R., & St Jacques, P. (2007). Functional neuroimaging
of autobiographical memory. Trends in Cognitive Sciences,
11, 219–227.
Craig, A. D. (2003). Interoception: The sense of the
physiological condition of the body. Current Opinion in
Neurobiology, 13, 500–505.
Craig, A. D., Chen, K., Bandy, D., & Reiman, E. M. (2000).
Thermosensory activation of insular cortex. Nature
Neuroscience, 3, 184–190.
D’Argembeau, A. (2013). On the role of the ventromedial
prefrontal cortex in self-processing: The valuation
hypothesis. Frontiers in Human Neuroscience, 7, 372.
Eichenbaum, H., Yonelinas, A. P., & Ranganath, C. (2007). The
medial temporal lobe and recognition memory. Annual
Review of Neuroscience, 30, 123–152.
Erwin, R. J., & Ferguson, E. D. (1979). The effect of food and
water deprivation and satiation on recognition. The
American Journal of Psychology, 92, 611–626.
Farovik, A., Place, R. J., Miller, D. R., & Eichenbaum, H. (2011).
Amygdala lesions selectively impair familiarity in recognition
memory. Nature Neuroscience, 14, 1416–1417.
Glascher, J., Hampton, A. N., & O’Doherty, J. P. (2009).
Determining a role for ventromedial prefrontal cortex in
encoding action-based value signals during reward-related
decision making. Cerebral Cortex, 19, 483–495.
Gross, J., Woelbert, E., Zimmermann, J., Okamoto-Barth, S.,
Riedl, A., & Goebel, R. (2014). Value signals in the
prefrontal cortex predict individual preferences across
reward categories. The Journal of Neuroscience, 34,
7580–7586.
Gusnard, D. A., Akbudak, E., Shulman, G. L., & Raichle, M. E.
(2001). Medial prefrontal cortex and self-referential mental
activity: Relation to a default mode of brain function.
Proceedings of the National Academy of Sciences, U.S.A.,
98, 4259–4264.
Hamann, S. B., Ely, T. D., Grafton, S. T., & Kilts, C. D.
(1999). Amygdala activity related to enhanced memory
for pleasant and aversive stimuli. Nature Neuroscience,
2, 289–293.
Hare, T. A., Camerer, C. F., & Rangel, A. (2009). Self-control in
decision-making involves modulation of the vmPFC valuation
system. Science, 324, 646–648.
Hare, T. A., Malmaud, J., & Rangel, A. (2011). Focusing attention
on the health aspects of foods changes value signals in
vmPFC and improves dietary choice. Journal of
Neuroscience, 31, 11077–11087.
Hassabis, D., Kumaran, D., & Maguire, E. A. (2007). Using
imagination to understand the neural basis of episodic
memory. Journal of Neuroscience, 27, 14365–14374.
Hassabis, D., & Maguire, E. A. (2009). The construction system
of the brain. Philosophical Transactions of the Royal Society,
Series B, Biological Sciences, 364, 1263–1271.
Howard, M. W., & Eichenbaum, H. (2013). The hippocampus,
time, and memory across scales. Journal of Experimental
Psychology: General, 142, 1211–1230.
Johnson, J. D., Suzuki, M., & Rugg, M. D. (2013). Recollection,
familiarity, and content-sensitivity in lateral parietal cortex:
A high-resolution fMRI study. Frontiers in Human
Neuroscience, 7, 219.
Johnson, S. C., Baxter, L. C., Wilder, L. S., Pipe, J. G., Heiserman,
J. E., & Prigatano, G. P. (2002). Neural correlates of self-
reflection. Brain, 125, 1808–1814.
Kensinger, E. A., Addis, D. R., & Atapattu, R. K. (2011).
Amygdala activity at encoding corresponds with memory
vividness and with memory for select episodic details.
Neuropsychologia, 49, 663–673.
Kensinger, E. A., & Schacter, D. L. (2006). Amygdala activity is
associated with the successful encoding of item, but not
source, information for positive and negative stimuli. The
Journal of Neuroscience, 26, 2564–2570.
Knutson, B., Rick, S., Wimmer, G. E., Prelec, D., & Loewenstein,
G. (2007). Neural predictors of purchases. Neuron, 53,
147–156.
Laeng, B., & Sulutvedt, U. (2014). The eye pupil adjusts to
imaginary light. Psychological Science, 25, 188–197.
Larson, J. S., Redden, J. P., & Elder, R. S. (2014). Satiation from
sensory simulation: Evaluating foods decreases enjoyment
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1966
Journal of Cognitive Neuroscience
Volume 27, Number 10
of similar foods. Journal of Consumer Psychology, 24,
188–194.
Lebreton, M., Bertoux, M., Boutet, C., Lehericy, S., Dubois, B.,
Fossati, P., et al. (2013). A critical role for the hippocampus in
the valuation of imagined outcomes. PLoS Biology, 11,
e1001684.
Lebreton, M., Jorge, S., Michel, V., Thirion, B., & Pessiglione, M.
(2009). An automatic valuation system in the human brain:
Evidence from functional neuroimaging. Neuron, 64,
431–439.
Levine, B. (2004). Autobiographical memory and the self in
time: Brain lesion effects, functional neuroanatomy, and
lifespan development. Brain and Cognition, 55, 54–68.
Macrae, C. N., Moran, J. M., Heatherton, T. F., Banfield, J. F., &
Kelley, W. M. (2004). Medial prefrontal activity predicts
memory for self. Cerebral Cortex, 14, 647–654.
Maguire, E. A. (2001). Neuroimaging studies of
autobiographical event memory. Philosophical Transactions
of the Royal Society, Series B, Biological Sciences, 356,
1441–1451.
Maldjian, J. A., Laurienti, P. J., Kraft, R. A., & Burdette, J. H.
(2003). An automated method for neuroanatomic and
cytoarchitectonic atlas-based interrogation of fMRI data sets.
Neuroimage, 19, 1233–1239.
Morewedge, C. K., Huh, Y. E., & Vosgerau, J. (2010). Thought
for food: Imagined consumption reduces actual
consumption. Science, 330, 1530–1533.
Morris, J. S., & Dolan, R. J. (2001). Involvement of human
amygdala and orbitofrontal cortex in hunger-enhanced
memory for food stimuli. Journal of Neuroscience, 21,
5304–5310.
Nadel, L., & Moscovitch, M. (1997). Memory consolidation,
retrograde amnesia and the hippocampal complex. Current
Opinion in Neurobiology, 7, 217–227.
Nicolle, A., Klein-Flügge, M. C., Hunt, L. T., Vlaev, I., Dolan, R. J.,
& Behrens, T. E. J. (2012). An agent independent axis for
executed and modeled choice in medial prefrontal cortex.
Neuron, 75, 1114–1121.
Nieuwenhuis, I. L. C., & Takashima, A. (2011). The role of the
ventromedial prefrontal cortex in memory consolidation.
Behavioural Brain Research, 218, 325–334.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a
cognitive map. Oxford: Clarendon Press.
Pennartz, C. M. A., Lee, E., Verheul, J., Lipa, P., Barnes, C. A., &
McNaughton, B. L. (2004). The ventral striatum in off-line
processing: Ensemble reactivation during sleep and
modulation by hippocampal ripples. Journal of
Neuroscience, 24, 6446–6456.
Peters, J., & Büchel, C. (2010). Episodic future thinking
reduces reward delay discounting through an enhancement
of prefrontal-mediotemporal interactions. Neuron,
66, 138–148.
Piolino, P., Giffard-Quillon, G., Desgranges, B., Chételat, G.,
Baron, J. C., & Eustache, F. (2004). Re-experiencing old
memories via hippocampus: A PET study of autobiographical
memory. Neuroimage, 22, 1371–1383.
Ranganath, C. (2010). A unified framework for the functional
organization of the medial temporal lobes and the
phenomenology of episodic memory. Hippocampus, 20,
1263–1290.
Rolls, E. T. (2005). Emotion explained. Oxford: Oxford
University Press.
Rugg, M. D., & Vilberg, K. L. (2013). Brain networks underlying
episodic memory retrieval. Current Opinion in
Neurobiology, 23, 255–260.
Rushworth, M., Noonan, M., Boorman, E., Walton, M., &
Behrens, T. (2011). Frontal cortex and reward-guided learning
and decision-making. Neuron, 70, 1054–1069.
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng,
R. N., & Szpunar, K. K. (2012). The future of memory:
Remembering, imagining, and the brain. Neuron, 76,
677–694.
Scoville, W. B., & Milner, B. (1957). Loss of recent memory after
bilateral hippocampal lesions. Journal of Neurology,
Neurosurgery, and Psychiatry, 20, 11–21.
Simons, J. S., Peers, P. V., Hwang, D. Y., Ally, B. A., Fletcher,
P. C., & Budson, A. E. (2008). Is the parietal lobe necessary for
recollection in humans? Neuropsychologia, 46, 1185–1191.
Squire, L. R., & Zola-Morgan, S. (1991). The medial temporal
lobe memory system. Science, 253, 1380–1386.
Tataranni, P. A., Gautier, J. F., Chen, K., Uecker, A., Bandy, D.,
Salbe, A. D., et al. (1999). Neuroanatomical correlates of
hunger and satiation in humans using positron emission
tomography. Proceedings of the National Academy of
Sciences, U.S.A., 96, 4569–4574.
Tse, D., Langston, R. F., Kakeyama, M., Bethus, I., Spooner,
P. A., Wood, E. R., et al. (2007). Schemas and memory
consolidation. Science, 316, 76–82.
Tse, D., Takeuchi, T., Kakeyama, M., Kajii, Y., Okuno, H.,
Tohyama, C., et al. (2011). Schema-dependent gene
activation and memory encoding in neocortex. Science,
333, 891–895.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neuroimage, 15, 273–289.
van Kesteren, M. T. R., Beul, S. F., Takashima, A., Henson, R. N.,
Ruiter, D. J., & Fernández, G. (2013). Differential roles for
medial prefrontal and medial temporal cortices in schema-
dependent encoding: From congruent to incongruent.
Neuropsychologia, 51, 2352–2359.
van Kesteren, M. T. R., Fernández, G., Norris, D. G., & Hermans,
E. J. (2010). Persistent schema-dependent hippocampal-
neocortical connectivity during memory encoding and
postencoding rest in humans. Proceedings of the National
Academy of Sciences, U.S.A., 107, 7550–7555.
van Kesteren, M. T. R., Ruiter, D. J., Fernández, G., & Henson,
R. N. (2012). How schema and novelty augment memory
formation. Trends in Neurosciences, 35, 211–219.
Vogeley, K., May, M., Ritzl, A., Falkai, P., Zilles, K., & Fink, G. R.
(2004). Neural correlates of first-person perspective as one
constituent of human self-consciousness. Journal of
Cognitive Neuroscience, 16, 817–827.
Wagner, A. D., Schacter, D. L., Rotte, M., Koutstaal, W., Maril, A.,
Dale, A. M., et al. (1998). Building memories: Remembering
and forgetting of verbal experiences as predicted by brain
activity. Science, 281, 1188–1191.
Wagner, A. D., Shannon, B. J., Kahn, I., & Buckner, R. L. (2005).
Parietal lobe contributions to episodic memory retrieval.
Trends in Cognitive Sciences, 9, 445–453.
Winecoff, A., Clithero, J. A., Carter, R. M., Bergman, S. R., Wang,
L., & Huettel, S. A. (2013). Ventromedial prefrontal cortex
encodes emotional value. Journal of Neuroscience,
33, 11032–11039.
Wittmann, B. C., Schott, B. H., Guderian, S., Frey, J. U., Heinze,
H. J., & Düzel, E. (2005). Reward-related fMRI activation
of dopaminergic midbrain is associated with enhanced
hippocampus-dependent long-term memory formation.
Neuron, 45, 459–467.
Lin et al.
1967
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
9
0
5
/
7
1
1
9
9
5
4
7
9
/
5
1
3
8
7
8
o
3
c
3
n
0
_
6
a
/
_
j
0
o
0
c
8
n
3
6
_
a
p
_
d
0
0
b
8
y
3
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1