Visual Distraction Disrupts Category-tuned Attentional
Filters in Ventral Visual Cortex
Blaire Dube, Lasyapriya Pidaparthi, and Julie D. Golomb
Astratto
■ Our behavioral goals shape how we process information via
attentional filters that prioritize goal-relevant information, dictat-
ing both where we attend and what we attend to. When some-
thing unexpected or salient appears in the environment, Esso
captures our spatial attention. Extensive research has focused
on the spatiotemporal aspects of attentional capture, but what
happens to concurrent nonspatial filters during visual distraction?
Here, we demonstrate a novel, broader consequence of distrac-
zione: widespread disruption to filters that regulate category-
specific object processing. We recorded fMRI while participants
viewed arrays of face/house hybrid images. On distractor-absent
trials, we found robust evidence for the standard signature of
category-tuned attentional filtering: greater BOLD activation in
fusiform face area during attend-faces blocks and in parahippo-
campal place area during attend-houses blocks. Tuttavia, on trials
where a salient distractor (white rectangle) flashed abruptly around
a nontarget location, not only was spatial attention captured, Ma
the concurrent category-tuned attentional filter was disrupted,
revealing a boost in activation for the to-be-ignored category. Questo
disruption was robust, resulting in errant processing—and early
SU, prioritization—of goal-inconsistent information. These find-
ings provide a direct test of the filter disruption theory: that in
addition to disrupting spatial attention, distraction also disrupts
nonspatial attentional filters tuned to goal-relevant information.
Inoltre, these results reveal that, under certain circumstances,
the filter disruption may be so profound as to induce a full rever-
sal of the attentional control settings, which carries novel impli-
cations for both theory and real-world perception. ■
INTRODUCTION
Our visual environments are too complex to process in their
entirety. To compensate, we filter incoming visual informa-
tion based on current behavioral goals, using attention to
selectively process only the most relevant information
(per esempio., Desimone & Duncan, 1995; Bundesen, 1990). One
way that we filter information is by prioritizing objects based
on high-level attributes such as category: When searching
for a friend on a busy street, for instance, we prioritize the
faces in the scene at the expense of less relevant categories
such as houses. Category-based attention ensures that
important information is prioritized for attentional selec-
tion early (Nako, Wu, & Eimer, 2014; Zhang & Luck,
2009) and simultaneously throughout space (Liu & Hou,
2011; Liu & Mance, 2011; White & Carrasco, 2011; Sàenz,
Buraĉas, & Boynton, 2003; Saenz, Buracas, & Boynton,
2002), increasing search efficiency (per esempio., speed).
Such category-tuned filters are partially subserved
by category-specific neural regions in ventral visual
cortex, such as the fusiform face area (FFA; Kanwisher,
McDermott, & Chun, 1997; Mccarthy, Puce, Gore, & Truett,
1997) and the parahippocampal place area (PPA; Epstein &
Kanwisher, 1998). These regions respond preferentially to
viewing images of faces and scene stimuli, rispettivamente, E
are sensitive to attentional manipulations. Neural evidence
The Ohio State University
© 2022 Istituto di Tecnologia del Massachussetts
of object-based attentional filtering has been demonstrated
using categorically defined stimuli that overlap in space
(cioè., “hybrid” stimuli of semitransparent superimposed
images) as well as during binocular rivalry and mental
imagery, with BOLD responses enhanced in FFA relative
to PPA when observers attend to faces and in PPA relative
to FFA when observers attend to houses (Baldauf &
Desimone, 2014; Hsieh, Colas, & Kanwisher, 2012;
O’Craven & Kanwisher, 2000; Serences, Schwarzbach,
Courtney, Golay, & Yantis, 2004; Tong, Nakayama,
Vaughan, & Kanwisher, 1998). This category-based atten-
tional modulation has been linked to frontoparietal gamma
synchrony (Baldauf & Desimone, 2014), attentional shift
signals (Serences et al., 2004), and functional connectivity
(Turk-Browne, Norman-Haignere, & Mccarthy, 2010).
Allo stesso tempo, efficient goal-directed behavior also
relies heavily on spatial attention (Nako et al., 2014; Liu
& Mance, 2011; White & Carrasco, 2011; Zhang & Luck,
2009; Saenz et al., 2002, 2003; Hoffman & Nelson, 1981).
Goal-directed spatial attention and feature- or category-
based filters can coexist (Stein & Peelen, 2017), allowing
us to tune attention to task-relevant objects in attended
locations. Tuttavia, many of these studies have over-
looked a critical problem: Spatial attention is not static;
in the real world, dynamic and distracting information also
competes for selection. What are the consequences for
category-based attention when distracting information
appears in the search environment?
Journal of Cognitive Neuroscience 34:8, pag. 1521–1533
https://doi.org/10.1162/jocn_a_01870
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
C
N
_
UN
_
0
1
8
7
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Most prior work on visual distraction has focused on dis-
ruptions to spatial attention: The presence of distractors
increases search RTs (Theeuwes & Burger, 1998; Folk,
Remington, & Johnston, 1992; Remington, Johnston, &
Yantis, 1992; Yantis & Jonides, 1984; Jonides & Irwin,
1981), attracts eye movements (Liesefeld, Liesefeld,
Töllner, & Müller, 2017; Henderson, 2003; Theeuwes,
Kramer, Hahn, Irwin, & Zelinsky, 1999; Theeuwes, Kramer,
Hahn, & Irwin, 1998), and elicits electrophysiological
markers of covert spatial attention (Bacigalupo & Luck,
2019; Wang, van Driel, Ort, & Theeuwes, 2019; Luck
& Hillyard, 1994) during visual search. More recent
work has started to evaluate broader consequences of
visual distraction, Per esempio, demonstrating that atten-
tional capture can also distort feature perception (Chen,
Leber, & Golomb, 2019). Tuttavia, what about other types
of concurrent nonspatial attentional control? Recentemente,
Dube and Golomb (2021) proposed a filter disruption the-
ory, suggesting that in addition to disrupting spatial atten-
tion filters, visual distraction also disrupts currently active
nonspatial filters. Dube and Golomb (2021) used a behav-
ioral memory-driven capture paradigm (Olivers, Meijer, &
Theeuwes, 2006) to demonstrate that distraction disrupts
the filter that prioritizes relevant information for visual
working memory ( VWM) encoding, resulting in irrelevant
distractor features intruding into VWM. Tuttavia, IL
most direct prediction of this theory remains untested:
that distraction could even disrupt a prolonged, robust
attentional control setting like a stable category-based fil-
ter, causing a temporary increase in neural processing of
task-irrelevant information across the visual field.
In the current study, we use face–house hybrid stimuli
and fMRI to directly evaluate this key prediction of the filter
disruption theory: in addition to capturing spatial attention,
visual distraction also momentarily disrupts a category-
tuned attentional filter, causing the selection and processing
of stimuli from a task-irrelevant category that would other-
wise be ignored. We predicted that, on distractor-absent
trials, we would observe the standard signature of atten-
tional modulation: greater FFA activity on attend-faces
blocks relative to attend-houses blocks and greater PPA
activity on attend-houses blocks relative to attend-faces
blocks. Critically, on trials where a salient distractor appears,
we consider two competing hypotheses: (1) Spatial atten-
tion is captured, but the category filter remains intact, result-
ing in a behavioral capture effect (per esempio., slower RTs), Ma
preserved neural selectivity (per esempio., greater FFA during
attend-faces blocks), versus (2) spatial attention is captured
and the category-tuned attentional filter is also broken,
resulting in the errant processing of the irrelevant object
categoria (cioè., houses during attend-faces blocks).
METHODS
Overview
the target via a white border. Participants were instructed
to attend to one stimulus category (cioè., attend-faces
blocks and attend-houses blocks) and determine whether
the identity of the target stimulus on the current trial
matched that of the previous trial. On some trials, a salient
distractor (a white dotted line) appeared briefly around
one of the other three hybrid images. To examine
category-tuned attentional filtering, we measured BOLD
activation in the FFA and the PPA. Critically, here, we are
not testing whether a distracting image is processed dur-
ing attentional capture. Piuttosto, we are evaluating whether
attentional capture breaks a long-term (across-block) cat-
egory filter, causing the incidental processing of the irrel-
evant object category within the hybrid images.
Participants
Fifteen participants participated in the study (eight
women, seven men; mean age = 23.5 years, range =
19–31 years) for monetary compensation ($10/hr for the behavioral prescreen, $15/hr for the scanning session).
One participant was an author but was scanned before
their role in or knowledge of the study. All participants
reported normal or corrected-to-normal vision, provided
informed consent, and were prescreened for MRI eligi-
bility. The study protocol was approved by the Ohio
State University Biomedical Sciences institutional review
board.
The sample size was selected with the goal of extensive
within-participant sampling and was chosen to be suffi-
cient to detect the characteristically large effect sizes for
univariate comparisons of FFA/PPA activity, comparable
to prior studies (Peters, Roelfsema, & Goebel, 2012; Yi,
Kelley, Marois, & Chun, 2006; Wojciulik, Kanwisher, &
Driver, 1998). The approach of collecting relatively smaller
sample sizes, but with extensive numbers of trials for each
participant, is an alternative way of increasing statistical
power (Naselaris, Allen, & Kay, 2021) and carries certain
benefici, including the ability to assess the robustness of
effects at the individual participant level. In our design,
each of our participants completed 576 experimental tri-
COME, and we show both group- and individual-level results
in our data figures.
Stimuli and Task
Stimuli were generated and presented using MATLAB
(Mathworks, Inc.) and the Psychophysics Toolbox
(Brainard, 1997) and were presented on a gray back-
ground for both the experimental and localizer tasks. Par-
ticipants viewed all stimuli from a distance of 74 cm via a
mirror 45° above the head coil.
Experimental Task
On each trial, participants viewed an array of four
face/house hybrid images, one of which was defined as
See Figure 1 for a trial schematic. Experimental stimuli
were grayscale face/ house hybrid images, made by
1522
Journal of Cognitive Neuroscience
Volume 34, Numero 8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
C
N
_
UN
_
0
1
8
7
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
C
N
_
UN
_
0
1
8
7
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
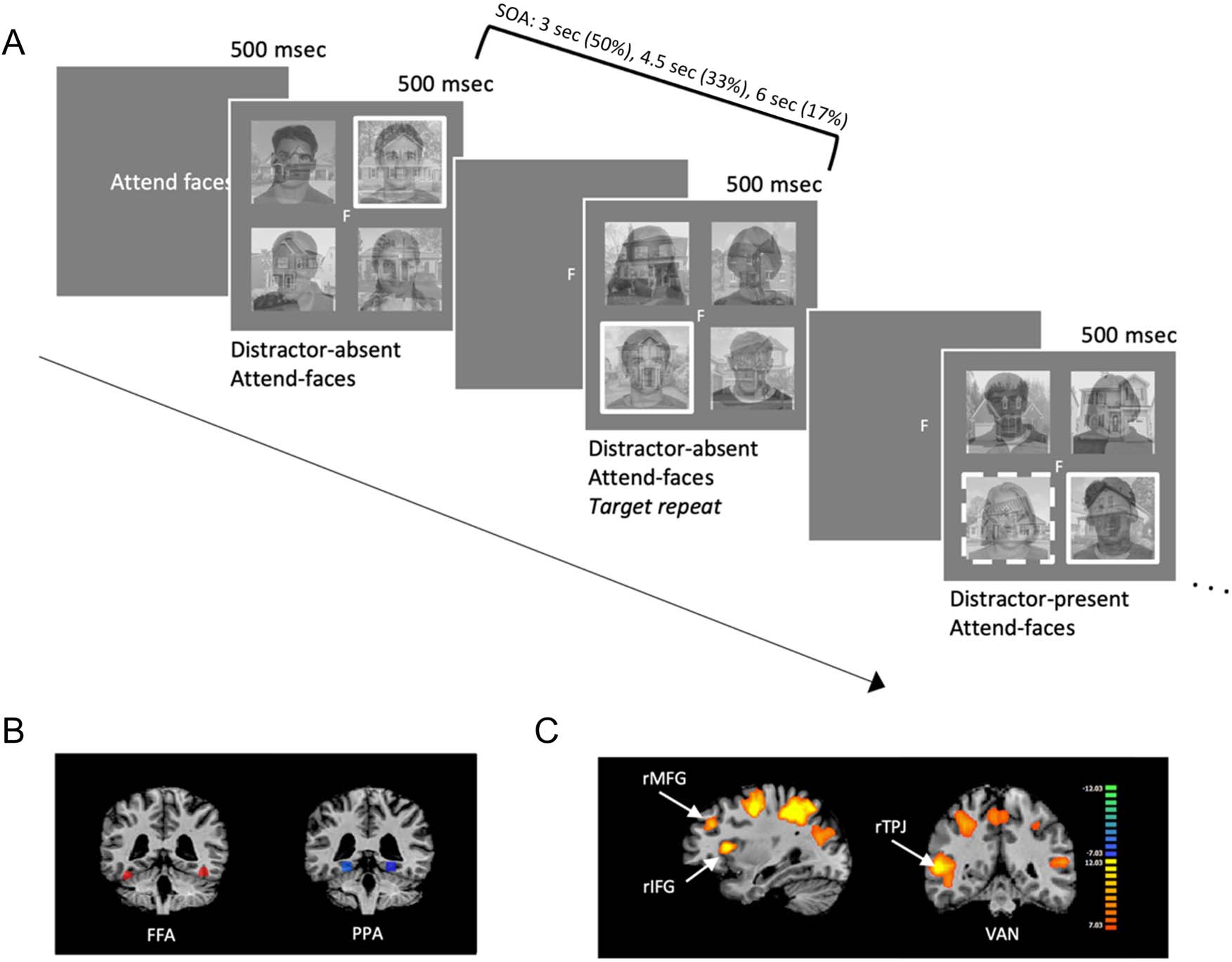
Figura 1. (UN) The experimental trial sequence. In the scanner, participants performed a 1-back task based on the relevant category (here attend-
faces), for the images appearing in the target box (white border). Participants responded with a “1” if the relevant image did not repeat and “2” if it
did. On 25% of trials, a salient abrupt-onset distractor (dotted white border) appeared around one of the three nontarget hybrid images. The salient
distractor appeared 50 msec after the onset of the stimulus display for that trial and remained on-screen for 100 msec. Participants were instructed to
ignore this salient distractor when it appeared. (B) The a priori FFA and PPA ROIs for a sample participant. (C) A whole-brain contrast showing
activation on salient-distractor-present contrasted with salient-distractor-absent trials, highlighting the VAN, with the rMFG, rIFG, and rTPJ ROIs
labeled.
overlaying a 50% transparent face image over a 50% trans-
parent house image. Hybrids were created from a bank of
160 total images (80 faces, as used in Golomb and
Kanwisher [2012] E 80 houses retrieved from a Google
image search and converted to grayscale), and face/house
image pairs were generated randomly and separately for
each participant (2304 unique hybrid images per partici-
pant: four per trial, 96 experimental trials per run, six
runs). Each experimental run consisted of two blocks of
48 trials: an attend-faces block and an attend-houses block
(order counterbalanced across runs; run order generated
randomly across participants). Each block began with the
500-msec presentation of an instruction screen (“Attend
faces” or “Attend houses”). The central fixation point on
each subsequent trial was either a letter “F” or “H” (size =
1°) to indicate the current target category, and this
remained on-screen for the duration of the block.
On each trial, four face/house hybrid images (size =
5° × 5°, eccentricity = 2.5°) were presented simulta-
neously in a 2 × 2 grid for 500 msec. One of the images
was framed with a white border, identifying it as the target
for that trial. Participants were instructed to perform a
1-back task on the image inside this target box, based on
the relevant category (the face or house); questo è, is the cur-
rent target face (or house) the same face (or house) as the
previous trial’s target? Participants responded on the
button box with a “2” if the target repeated and “1” if it
did not. The target (relevant-category image inside the
target box) repetition frequency was 20% per block. IL
nonrelevant category image inside the target box repeated
independently with the same frequency, as did both the
relevant and nonrelevant category images in the three
nontarget locations. Participants performed 48 trials in a
row for each block, with trials separated by an ISI of either
3 sec (50% of trials), 4.5 sec (33% of trials), O 6 sec
(17% of trials). The central fixation point (F or H) remained
on-screen for the duration of the block.
Our critical manipulation was that, SU 25% of trials (12
trials per block, randomly intermixed but never back-to-
back), a salient abrupt-onset distractor also appeared in
Dube, Pidaparthi, and Golomb
1523
the display. The salient distractor was a thick dotted white
border that flashed briefly around one of the three nontar-
get image locations (onset time = 50 msec after array
onset, duration = 100 msec). Participants were instructed
to ignore this dotted border when it appeared.
Participants completed six experimental runs (6.75 min
each). This resulted in 216 attend-faces distractor-absent
trials, 216 attend-houses distractor-absent trials, 72
attend-faces distractor-present trials, E 72 attend-
houses distractor-present trials. The location of the target
Immagine, whether the salient distractor appeared, and the
location of the salient distractor were all randomized
across trials. We note that the task was intentionally diffi-
cult (the hybrid images substantially increase perceptual
load, and the 1-back task requires working memory
resources and a strong attentional control setting) A
further incentivize attentional filtering.
Functional Localizer Task
Participants also completed two experimental runs of a
standard functional localizer task to localize FFA and PPA
ROIs. Participants viewed blocks of faces or houses (single
images presented sequentially at screen center, sized 10°)
and performed a 1-back task. Images were presented for
300 msec with a 500-msec ISI, and 1-back (repetition) fre-
quency was 10%. There were 20 trials per block, and par-
ticipants performed 13 blocks per run (five house blocks,
five face blocks, and three fixation blocks during which
only a fixation point appeared).
fMRI Acquisition
The study was carried out at the Ohio State University
Center for Cognitive and Behavioral Brain Imaging with
a Siemens Prisma 3-T MRI scanner using a 32-channel
phase array receiver head coil. Functional data were
acquired using a T2-weighted gradient-echo sequence
(repetition time = 1500 msec, echo time = 28 msec, flip
angle = 70°). We used multiband whole-brain coverage
aligned to the anterior commissure–posterior commis-
sure (48 slices, 3 × 3 × 3 mm voxel, 10% gap, multiband
factor = 3). We also collected a high-resolution MPRAGE
anatomic scan at 1-mm3 resolution for each participant.
Each participant was scanned in one 2-hr session.
fMRI Preprocessing
The fMRI data were preprocessed with Brain Voyager QX
(Brain Innovation). All functional data were corrected for
slice acquisition time and head motion, temporally filtered
(general linear model [GLM] Fourier; two cycles), and spa-
tially smoothed with a Gaussian kernel of 4-mm FWHM.
Runs with abrupt motion (>1 mm) were discarded from
analyses, resulting in the removal of a single run from
three separate participants and all six experimental runs
for a single participant and leaving a final n = 14. Data
for each participant were normalized into Talairach space
(Talairach & Tournoux, 1998).
ROIs
Our main analyses focused on two a priori functionally
defined ROIs: the FFA and the PPA. We localized the FFA
and PPA using standard procedures: Using data acquired
from the independent localizer scan, for each participant,
we defined a bilateral face-selective FFA in the mid-
fusiform gyrus that responded more strongly to faces than
houses and a bilateral house-selective PPA in the parahip-
pocampal gyrus that responded more strongly to houses
than faces. The resultant ROIs were contiguous voxels
(cluster threshold = 10) that demonstrated categorical
preference at a significance of p < 10−3 (uncorrected).
For two participants, we were unable to localize left FFA
at this threshold, so we only included right FFA ROIs.
For all other participants, we averaged across left and right
ROIs for FFA and PPA analyses. The reported pattern of
results is the same for left and right ROIs.
We performed additional exploratory analyses to evalu-
ate the effects of the salient distractor in the ventral atten-
tion network ( VAN). The VAN was identified based on a
contrast of distractor-present trials relative to distractor-
absent trials regardless of attended category. We localized
ROIs in right TPJ (rTPJ), right middle frontal gyrus (rMFG),
and right inferior frontal gyrus (rIFG), well-known regions
of the VAN that detect salient events outside the current
focus of attention (Shulman et al., 2009; Indovina &
Macaluso, 2007; Serences et al., 2005; Downar, Crawley,
Mikulis, & Davis, 2001; Corbetta, Kincade, Ollinger,
Mcavoy, & Gordon, 2000). We defined these ROIs for each
participant using a leave-one-subject-out procedure to
avoid circularity concerns (Esterman, Tamber-Rosenau,
Chiu, & Yantis, 2010). We iteratively left a single partici-
pant out of the group GLM and performed a whole-brain
analysis on all trials on the group-level (N − 1) data to
localize the relevant ROIs for the left-out participant,
contrasting distractor-present trials relative to distractor-
absent trials. From this contrast, we defined a set of
contiguous voxels (cluster threshold = 30) in the rTPJ,
rMFG, and rIFG for each participant that demonstrated
substantially greater activation on distractor-present
trials relative to distractor-absent trials at a significance of
p < 10−12 (uncorrected). These secondary ROIs were only
analyzed for the final exploratory tests contrasting the
first versus second half of trials: We report results from
all three ROIs but focus our discussion on the rTPJ as a
representative ROI from the VAN given its robust role in
detecting unexpected events of potential behavioral
relevance (i.e., a distractor that is visually similar/related
to the target [Natale, Marzi, & Macaluso, 2010; Hu, Bu,
Song, Zhen, & Liu, 2009; Hampshire, Duncan, & Owen,
2007; Serences et al., 2005]) and as a “reset” mechanism
for reorienting (Corbetta, Patel, & Shulman, 2008).
Figure 1B and C shows all ROIs.
1524
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analyses
Behavior
Average RTs for the 1-back task were calculated per condi-
0—a
tion using only correct trials. We also calculated d
sensitivity metric based on signal detection theory—as a
0 measures
measure of accuracy for each condition. Here, d
sensitivity to a target (i.e., a repeat trial) based on the num-
ber of hits (correct recognition of a repeat trial) relative to
false alarms (erroneously identifying a nonrepeat trial as a
repeat; Stanislaw & Todorov, 1999). We also report the
results of an exploratory analysis on behavioral data in
which we compare RT results from the first half of a block
to the second half of a block to evaluate for evidence of
habituation to the salient onset distractor (i.e., Won &
Geng, 2020) within a given control setting.
fMRI
Univariate fMRI analyses were carried out using Brain
Voyager QX (Brain Innovation). Using the functionally
defined FFA and PPA ROIs for each participant, we applied
a GLM to evaluate the mean BOLD response ( beta
weights) for each experimental condition, ROI, and partic-
ipant. Incorporated in the GLM were regressors for each of
the four experimental conditions (attend-faces distractor-
absent, attend-faces distractor-present, attend-houses
distractor-absent, and attend-houses distractor-present)
plus an instruction condition (“attend face” or “attend
house” text before each block). “Distractor-present” and
“distractor-absent” here and in the remainder of the text
refer to the presence/absence, respectively, of the salient
abrupt-onset distractor. The beta weights for the four
experimental conditions were submitted to repeated-
measures ANOVAs and paired t tests. We conducted
analyses within each ROI separately and also combined
across ROIs by averaging the BOLD responses for each
ROI’s preferred and nonpreferred conditions.
We first performed planned paired-samples t tests to
compare activation on attend-faces trials versus attend-
houses trials in the distractor-absent conditions to estab-
lish baseline filtering in each ROI and then performed
the same contrasts for the distractor-present conditions.
We then conducted 2 (Target Category: attend-faces vs.
attend-houses) × 2 (Distractor Condition: absent vs.
present) repeated-measures ANOVAs in FFA and PPA
ROIs. Of particular importance in this analysis is the Target
Category × Distractor Condition interaction, which pro-
vides insight into whether attentional filtering patterns
are substantially different on distractor-present relative
to distractor-absent trials.
Next, we combined across FFA and PPA ROIs to evaluate
activation reflecting the processing of the preferred
stimulus category (FFA activity on attend-faces trials, PPA
activity on attend-houses trials) relative to the nonpre-
ferred stimulus category (PPA activity on attend-faces tri-
als, FFA activity on attend-houses trials) across distractor
conditions. We performed an additional 2 (Target Category:
attend-preferred vs. attend-nonpreferred) × 2 (Distractor
Condition: absent vs. present) x 2 (ROI: FFA vs. PPA)
repeated-measures ANOVA.
As a measure of filter strength, we also calculated an
attentional filtering index for each ROI, participant, and
distractor condition using the formula below, where
higher positive values indicate stronger, more effective
attentional filtering.
ð
ð
Attentional Filtering Index ¼ Attpreferred − Attnonpreferred
Attpreferred þ Attnonpreferred
We performed a 2 (ROI: FFA vs. PPA) × 2 (Distractor Con-
dition: absent vs. present) ANOVA on attentional filtering
indices. We followed up with post hoc one sample t tests to
evaluate whether attentional filtering was significantly
nonzero in each distractor condition across ROIs.
Þ
Þ
Finally, we performed exploratory analyses to evaluate
changes in the first half versus the second half of experi-
mental blocks (block phase: first half vs. second half ),
for the FFA and PPA analyses described above, as well
as the rTPJ ROI. For this, we applied a GLM with
eight regressors (attend-faces distractor-absent block
Phase 1, attend-faces distractor-absent block Phase 2;
etc.), plus an instruction condition, to evaluate mean
BOLD response for each of the original four experimental
conditions by block phase (first half vs. second half ), per
participant.
Effect sizes (mean and 90% CI) are provided for all
analyses.
RESULTS
Behavioral and Neural Confirmation of
Attentional Capture
To confirm that the salient distractor captured spatial
attention as intended, we compared mean RTs (Table 1)
across conditions in the 1-back task. We subjected RTs
to a 2 (Distractor Condition: absent vs. present) × 2 (Tar-
get Category: attend-faces vs. attend-houses) repeated-
measures ANOVA. We observed a significant main effect
of Distractor Condition, F(1, 13) = 41.75, p < .001, ηp
2 =
.76 (90% CI [0.97, 2.33]), such that RTs were slower on
distractor-present trials relative to distractor-absent trials,
demonstrating reliable attentional capture. There was also
a significant main effect of Target Category, F(1, 13) =
47.92, p < .001, ηp
2 = .79 (90% CI [0.53, 0.86]), such that
RTs were generally slower in the attend-houses conditions
relative to the attend-faces conditions, although the lack of
a significant interaction, F(1, 13) = 0.01, p = .93, ηp
2 = .001
(90% CI [0, 0.03]), suggests that the distractor costs were
comparable across categories.
The response accuracy (Table 1) data trended in the
same patterns as RT, suggesting no speed–accuracy trade-
offs: here, a significant main effect of Distractor Condition,
F(1, 13) = 6.73, p = .02, ηp
2 = .34 (90% CI [0.03, 0.56]),
with a nonsignificant main effect of Target Category, F(1,
Dube, Pidaparthi, and Golomb
1525
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Summary of Mean RT (sec), Hit Rate (HR), False Alarm (FA) Rate, and Accuracy (d
0)
Attend-faces,
Distractor-absent
Attend-faces,
Distractor-present
Attend-houses,
Distractor-absent
Attend-houses,
Distractor-present
RT (SD)
HR (SD)
FA (SD)
d0 (SD)
0.99 (0.18)
0.50 (0.17)
0.10 (0.11)
1.40 (0.63)
1.12 (0.22)
0.39 (0.17)
0.11 (0.13)
0.94 (0.85)
1.11 (0.20)
0.47 (0.15)
0.13 (0.10)
1.05 (0.51)
1.24 (0.26)
0.31 (0.24)
0.11 (0.08)
0.75 (0.87)
Data for all four experimental conditions. d0 is calculated per participant from hit rate and false alarm rate using d0 = ZHIT − ZFA where z represents
the respective z transformations (Macmillan & Creelman, 1990).
13) = 1.78, p = .21, ηp
2 = .12 (90% CI [0, 0.38]). Again,
there was no significant interaction, F(1, 13) = 0.22, p =
.65, ηp
2 = .02 (90% CI [0, 0.22]).
A whole-brain contrast of distractor-present versus
distractor-absent trials also confirmed previously reported
neural patterns associated with attentional capture,
namely, increased activation in the VAN, including the
rTPJ, rMFG, and rIFG (Figure 1C).
Neural Processing Via Category-tuned Attentional
Filters in FFA and PPA
The key question this study was designed to address was
whether the attentional capture effects were accompanied
by changes to category-tuned filters. In both FFA and PPA
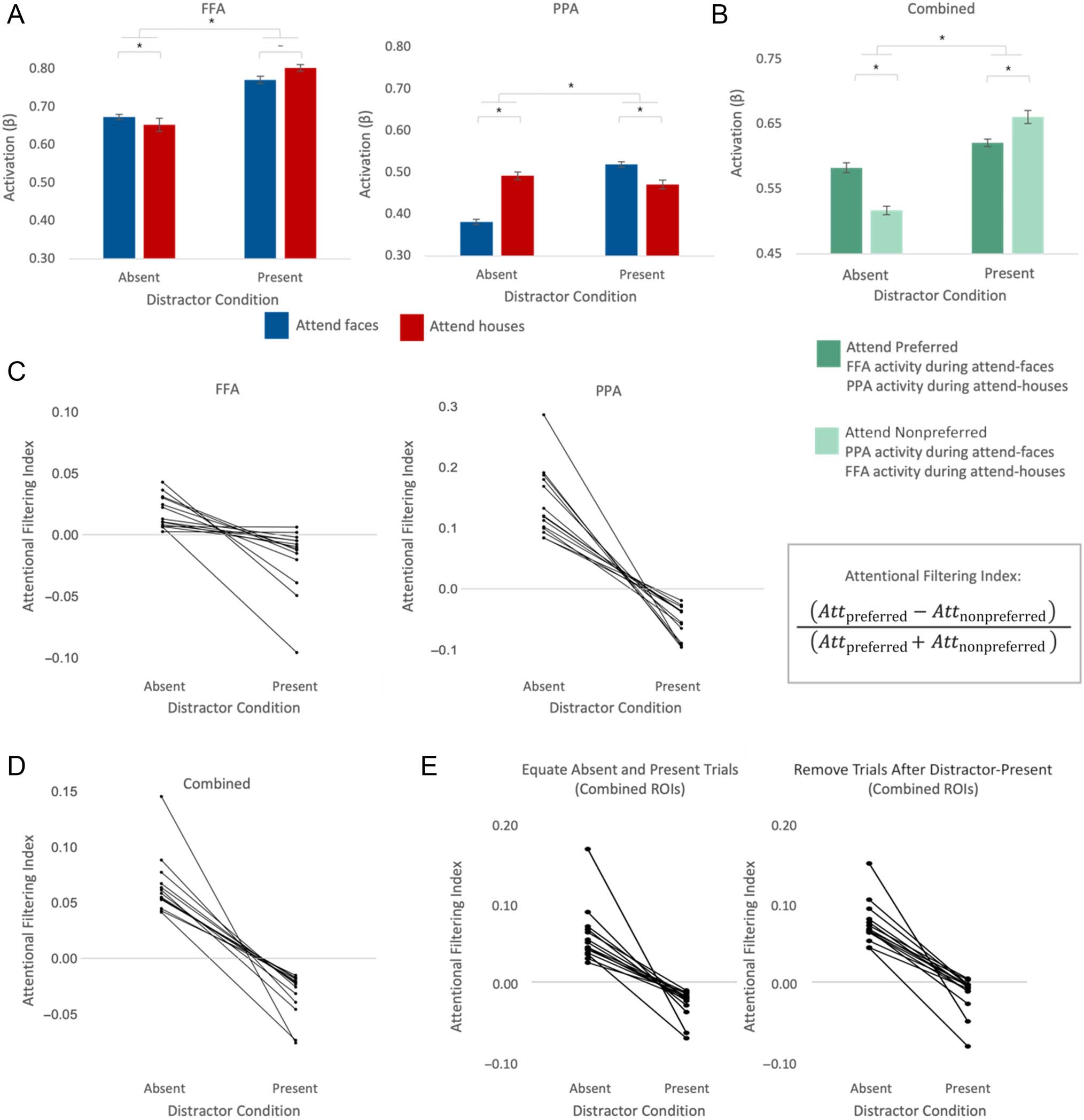
(Figure 2A), we observed the expected pattern of standard
category-based attentional modulation on distractor-
absent trials: significantly greater FFA activation on
attend-faces trials relative to attend-house trials, t(13) =
5.70, p < .001, d = 1.52 (90% CI [0.73, 2.29]) and signifi-
cantly greater PPA activation on attend-houses trials rela-
tive to attend-faces trials, t(13) = 13.6, p < .001, d =
3.64 (90% CI [2.15, 5.10]). This baseline comparison
confirms that, in the absence of a salient visual distractor,
participants were effectively attending the target category
and filtering out the irrelevant nontarget category in the
hybrid images.
Strikingly, on distractor-present trials, this pattern was
dramatically altered in both FFA and PPA. In fact, if any-
thing, the filtering pattern was reversed: FFA activation
to the hybrid images was numerically greater on attend-
houses blocks relative to attend-faces blocks, t(13) =
−1.90, p = .08, d = −0.51 (90% CI [−1.06, 0.06]), and
PPA activation was significantly greater on attend-faces
blocks relative to attend-houses blocks, t(13) = 10.86,
p < .001, d = 2.90 (90% CI [1.67, 4.11]). This suggests that,
under conditions of distraction, the filter that prioritized
the to-be-attended category was not only disrupted but
also reversed, causing the incidental prioritization of the
faces in the hybrid face/house images on attend-houses
trials and the incidental prioritization of the houses in
the hybrid images on attend-faces trials. The Target
Category × Distractor Condition interactions were signif-
icant in both ROIs, FFA: F(1, 13) = 9.21, p = .01, ηp
2 = .42
(90% CI [0.07, 0.61]); PPA: F(1, 13) = 178.34, p < .001,
ηp
2 = .93 (90% CI [0.83, 0.95]).
Figure 2B also shows the analysis combining across
ROIs, with data sorted into preferred versus nonpreferred
stimulus category (ANOVA statistics in Table 2). In addi-
tion to corroborating the significant effects above, there
was a significant main effect of ROI, with ROI also modu-
lating the other effects, such that overall BOLD activation
was generally greater in FFA, but PPA showed greater
differences between conditions.
To quantify attentional filter strength more directly, we
calculated an attentional filtering index for each ROI and
distractor condition for each participant (Figure 2C; com-
bined ROI in Figure 2D). Consistent with the above
results, we observed a significant main effect of distractor
condition, F(1, 13) = 112.71, p < .001, ηp
2 = .90 (90% CI
[0.75, 0.93]), with the filter disruption significant in both
ROIs: FFA distractor absent versus present: t(13) = 4.21,
p = .001, d = 1.13 (90% CI [0.54, 1.68]); PPA distractor
absent versus present: t(13) = 8.88, p < .001, d = 2.37
(90% CI [1.47, 3.23]). Critically, we observed robust filter
disruption in all 14 participants in both ROIs. Moreover,
the attentional filtering index was significant (greater
than zero) in both FFA and PPA on distractor-absent trials,
t(13) = 5.14, p < .001, d = 1.37 (90% CI [0.62, 2.10]), and
t(13) = 9.22, p < .001, d = 2.46 (90% CI [1.38, 3.52]), and
significantly less than zero on distractor-present trials,
t(13) = −2.65, p = .02, d = −0.71 (90% CI [−1.29,
−0.11]), and t (13) = −7.44, p < .001, d = −1.99 (90%
CI [−2.90, −1.06]).
To confirm the stability of the filter disruption in FFA
and PPA, we ran two control analyses (Figure 2E). First,
we equated for number of trials across conditions by ran-
domly selecting an equal number of distractor-absent trials
as distractor-present trials. The results were the same:
Attentional filtering for the combined ROIs was signifi-
cantly disrupted (i.e., weaker on distractor-absent relative
to distractor-present trials), t(13) = 6.52, p < .001, d =
1.74 (90% CI [1.01, 2.43]), with the disruption present in
14 of 14 participants. Furthermore, attentional filtering
was greater than zero on distractor-absent trials, t(13) =
6.00, p < .001, d = 1.60 (90% CI [1.05, 2.10]), and signif-
icantly less than zero on distractor-present trials, t(13) =
−5.40, p < .001, d = −1.44 (90% CI [−1.91, −0.92]).
1526
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Summary of fMRI BOLD results. (A) Activation levels in the FFA and PPA across all experimental conditions and (B) activation in combined
ROI across all experimental conditions. (C) Attentional filtering index (the proportion of total activation reflecting the processing of the preferred
stimulus category) for the FFA and PPA and combined in (D) where each data point represents a single participant. This is a measure of filtering
effectiveness—see box in the figure for calculation. A negative slope indicates filter disruption. Values below zero indicate filter reversal. (E) Results
from two control analyses, plotting attentional filtering index for the combined ROI. Error bars are within-participant standard error (Morey, 2008).
Second, we removed trials immediately after distractor-
present trials. Again, attentional filtering for the combined
ROIs was significantly disrupted on distractor-present tri-
als: t (13) = 8.76, p < .001, d = 2.34 (90% CI [1.45, 3.18]),
with the disruption present in 14 of 14 participants, and
attentional filtering was significantly greater than zero on
distractor-absent trials, t(13) = 10.00, p < .001, d = 2.68
(90% CI [1.88, 3.38]), and significantly less than zero on
distractor-present trials, t(13) = −2.37, p = .034, d =
−0.63 (90% CI [−1.00, −0.25]).
Exploratory Analysis: Distractor Habituation across
a Block
As a final, exploratory analysis, we asked whether the
observed filter disruption effect might decrease over time.
Dube, Pidaparthi, and Golomb
1527
Table 2. Statistics for the 2 (Target Category: Attend-preferred vs. Attend-nonpreferred) × 2 (Distractor Condition: Absent vs.
Present) × 2 (ROI: FFA vs. PPA) Repeated-Measures ANOVA Carried Out on BOLD Activation in FFA and PPA
ROI
Target category (preferred vs. nonpreferred)
Distractor condition
ROI × Target Category
ROI × Distractor Condition
Target Category × Distractor Condition
ROI × Target Category × Distractor Condition
df
1, 13
1, 13
1, 13
1, 13
1, 13
1, 13
1, 13
F
18.93
8.68
63.79
16.39
13.49
105.05
24.61
p
< .001
.011
< .001
.001
.003
< .001
< .001
ηp
2 [90% CI]
.59 [.23, .73]
.4 [.06, .61]
.83 [.61, .89]
.56 [.19, .71]
.51 [.15, .68]
.89 [.74, .93]
.65 [.31, .77]
There is prior evidence that the interference produced by
a salient distractor—both with respect to RT costs ( Won
& Geng, 2020) and oculomotor responses (Bonetti &
Turatto, 2019; Chelazzi, Marini, Pascucci, & Turatto,
2019; Turatto, Bonetti, & Pascucci, 2018)—can decrease
after repeated or prolonged exposure. Such habituation
is typically studied over the course of an experiment when
a salient onset distractor remains consistent. In the current
task, whereas the salient onset distractor stimulus (dotted
white border) remained consistent over the entire exper-
iment, the attentional filter/control setting had to be
updated at the start of each new block, such that the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
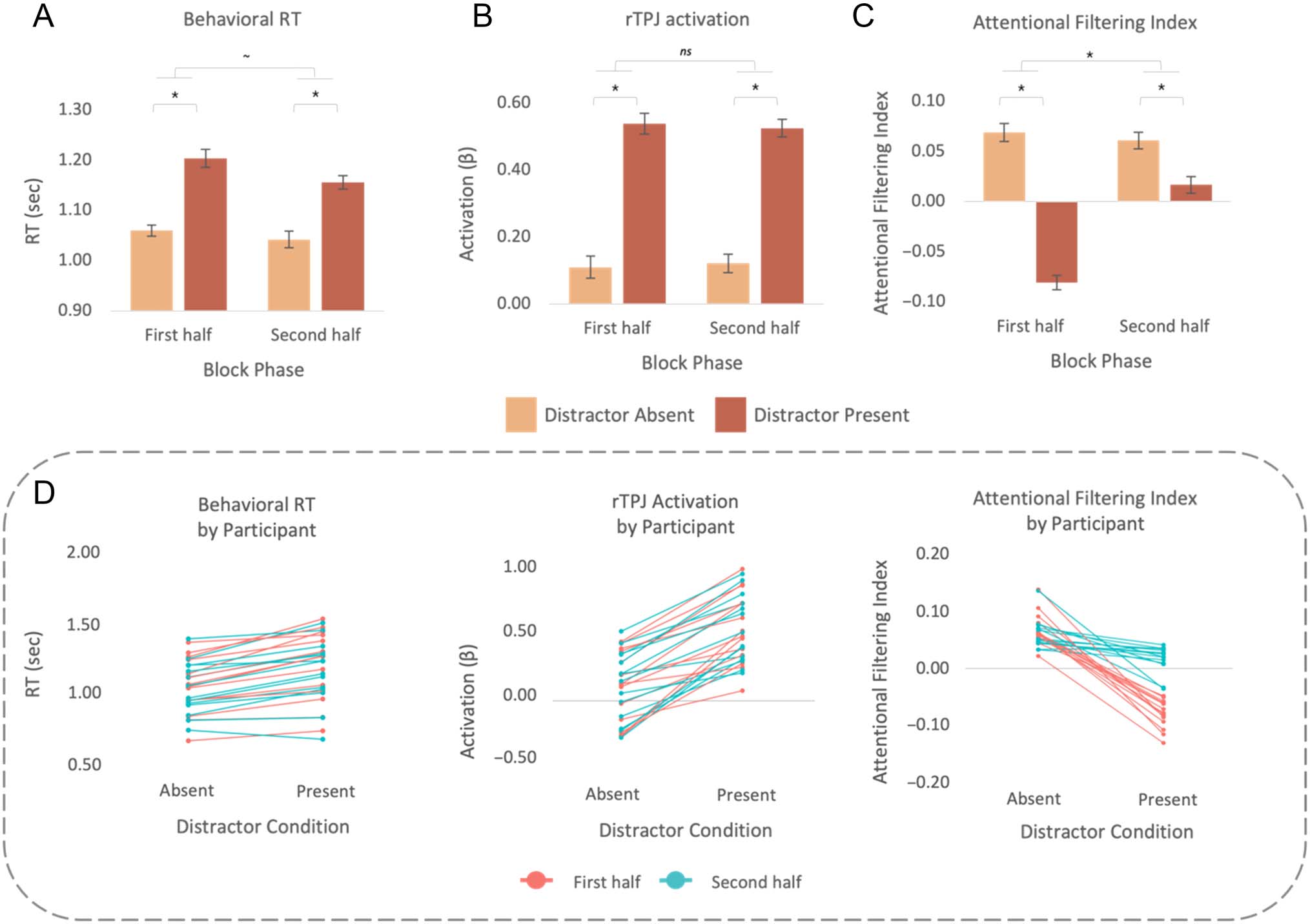
Figure 3. Summary of exploratory analyses of the first versus second halves of blocks. (A) Behavioral RT by distractor condition and block phase.
(B) rTPJ activation by distractor condition and block phase. (C) Attentional filter strength (via the attentional filtering index) across distractor
conditions and block phase, collapsed across ROI condition. (D) Participant-level RT, rTPJ activation, and the filtering index by block phase (where
a negative slope indicates filter disruption and values below zero indicate filter reversal). Error bars are within-participant standard error (Morey,
2008).
1528
Journal of Cognitive Neuroscience
Volume 34, Number 8
to-be-filtered (nontarget) category reset accordingly.
Thus, to explore if the consequences of attentional cap-
ture on category-tuned filtering (i.e., incidentally pro-
cessing the nontarget category) might be mitigated over
time, we separated our data into trials occurring in the
first half versus last half of each block. We compared (1)
the category-tuned attentional filtering index (combined
across FFA and PPA), (2) a behavioral measure of attentional
capture (RT for distractor-present vs. distractor-absent
trials), and (3) a generic neural measure of attentional cap-
ture (rTPJ activation for distractor-present vs. distractor-
absent trials). All three measures are shown across block
phase at both the group and participant levels in Figure 3.
In brief, the filter disruption cost in FFA/ PPA was
reduced for the second half of the block relative to the
first [Block Phase × Distractor Condition interaction,
F(1, 13) = 74.95, p < .001, ηp
2 = .85 (90% CI [0.57,
0.88])]. The behavioral capture cost (RT) was also reduced
marginally (Phase × Distractor interaction, F(1, 13) =
3.48, p = .08, ηp
2 = .21 (90% CI [0, 0.46]), but interestingly,
the generic neural capture measure did not vary (Phase ×
Distractor interaction, F < 1.43, p > .25).
In terms of the attentional filtering index (Figure 3C), SU
distractor-absent trials, attentional filtering was strong (sig-
nificantly greater than zero) both early and later in the
block, T(13) = 9.09, P < .001, d = 2.43 (90% CI [1.51,
3.3]), and t(13) = 8.61, p < .001, d = 2.3 (90% CI [1.42,
3.13], respectively). On distractor-present trials, however,
there was a notable difference over time: The attentional
filter was disrupted and reversed (attentional filtering was
significantly less than zero) early in the block, t(13) =
−12.15, p < .001, d = −3.25 (90% CI [−4.34, −2.09]),
but only more mildly disrupted (with no reversal) later
in the block. In the second half of the block, there was still
significant filter disruption, with the filtering index
reduced on distractor-present compared to distractor-
absent trials [paired-samples t test: t(13) = 3.62, p =
.003, d = 0.97 (90% CI [0.42, 1.49])], but the filtering index
remained positive [significantly greater than zero, t(13) =
2.59, p = .02, d = 0.69 (90% CI [0.19, 1.17])]. These results
reveal that the distractor produced a significant cost to
attentional filtering both early and later in a block, but this
cost was substantially mitigated later in the block, where
distraction disrupted but no longer reversed the filter.
Intriguingly, the same analysis carried out on rTPJ BOLD
activation yielded a different result (Figure 3B): Activation
in rTPJ was reliably greater on distractor-present trials rel-
ative to distractor-absent trials, F(1, 13) = 74.82, p < .001,
ηp
2 = .85 (90% CI [0.66, 0.90]), but it was unaffected by
block phase [in terms of both the lack of interaction and
no significant main effect of block phase: F(1, 13) = 0.01,
p = .92, ηp
2 = .001 (90% CI [0, 0.06])], suggesting that the
neural response to the salient distractor itself remained
equivalently strong over time (relative to the block onset),
yet the impact of this distractor on attentional filtering
and processing of the hybrid images was apparently
reduced. The rTPJ pattern was consistent across the
other localized ROIs of the VAN (rMFG and rIFG Block
Phase × Distractor interaction, ps > .69).
DISCUSSION
We have long known that visual distraction disrupts spatial
Attenzione (Jonides & Irwin, 1981), but recently, the conse-
quences are being understood to be broader (Dube &
Golomb, 2021; Chen et al., 2019). The current findings
reveal even more fundamental consequences for visual
distraction. We show that distraction also disrupts a con-
currently maintained category-tuned filter, interrupting
the prioritization of goal-relevant information in the visual
scene. This is particularly notable because the filter here
reflected a prolonged, robust attentional control setting,
and the disruption resulted in a temporary preference
for task-irrelevant information. Inoltre, the filter dis-
ruption was reliable enough to be seen at the individual
participant level for all participants tested.
This study was designed to directly test the filter disrup-
tion theory proposed in Dube and Golomb (2021). Dube
and Golomb (2021) demonstrated that visual distraction
(by a similar abrupt onset distractor as the current study)
causes the incidental encoding of distractor features, pre-
sumably by disrupting the filter that controls VWM encod-
ing. When performing two sequential visual search tasks in
which color was irrelevant, memory-driven capture (exac-
erbated attentional capture when visual information
matches the contents of VWM; Olivers et al., 2006) era
elicited in the second search when the color of a singleton
matched the distractor color from the first search, suggest-
ing that the irrelevant Search 1 distractor color intruded
into memory and subsequently biased attention. How-
ever, although participants were told to ignore color, there
was not an explicit instruction to filter irrelevant features
from VWM encoding. Accordingly, we designed the cur-
rent study to investigate a more robust attentional filter,
such that the task required a long-term (cioè., sustained
for a several minute block), explicit category-tuned filter.
The results of this study extend the theoretical implica-
tions of Dube and Golomb (2021) in important ways,
revealing that attentional capture not only disrupts spatial
attention but also disrupts control over nonspatial filters
that regulate behavior, such that activity in ventral visual
cortex no longer reflects prioritization of goal-relevant
informazione.
The results thus provide direct support for the filter
disruption theory: During distraction, spatial attention is
captured and the category-tuned attentional filter is also
broken, resulting in the errant processing of the irrelevant
object category. Is this boost in irrelevant category pro-
cessing occurring primarily within the hybrid image at
the distractor location, or is the category-tuned filter dis-
rupted globally across the display? Although the current
experiment was not designed to assess spatial selectivity
(cioè., we did not control eye movements, and our localizer
task used large, central stimuli), we conducted some
Dube, Pidaparthi, and Golomb
1529
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
C
N
_
UN
_
0
1
8
7
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
exploratory analyses capitalizing on the known contralat-
eral organization of visual cortex—that in both FFA and
PPA, neural activation is greater for stimuli presented on
the contralateral side of the visual display (Hemond,
Kanwisher, & Op de Beeck, 2007). We separately analyzed
FFA/PPA activation by hemisphere for the subset of trials
where the target and distractor were on opposite sides of the
display and found the same pattern of category tuning and
disruption in both hemispheres: UN 2 (distractor-present
vs. distractor-absent) × 2 (preferred vs. nonpreferred
target category) × 2 (contra-target vs. contra-distractor
hemisphere) ANOVA carried out on attentional filtering
indices yielded a significant Distractor Condition × Target
Category interaction, F(1, 13) = 80.4, P < .001, ηp
2 =
.86 (90% CI [0.67, 0.91]), but no three-way interaction,
F(1, 13) = 0.94, p = .35, ηp
2 = .07 (90% CI [0, 0.31]),
suggesting that the filter disruption effect did not interact
with hemisphere. Given the already coarse contralateral
organization of the FFA and PPA and limitations of the
current experimental design, these hemisphere-based
analyses do not allow us the resolution to inspect activa-
tion specific to the exact distractor or target locations and
should be taken as exploratory, but these data suggest
that the filter disruption is not limited solely to the
distractor location. Such an effect would be consistent
with the idea of a nonspatial category-tuned filter that
operates globally across the visual field. It has been well
established that feature-based attention operates quickly
and globally in parallel across a visual scene, indepen-
dently of spatial attention, allowing for a preliminary
“scan” of the visual scene to help tune a spatial saliency
map based on stimulus relevance (Liu & Mance, 2011;
Serences & Boynton, 2007; Saenz et al., 2002; Treue &
Martínez Trujillo, 1999). It is possible that nonspatial
categorical filters may work in a similar way. If so, the
current results suggest that, when the spatial atten-
tional filter is broken, the category-tuned attentional
filter may be similarly disrupted at both the location of the
distractor and elsewhere, a conjecture that could be
directly tested in future studies with more spatially
sensitive methods.
In terms of neural mechanisms or signatures of distrac-
tion, many studies have focused on the role of the fronto-
parietal dorsal attention network and VAN, with the dorsal
attention network (including the intraparietal sulcus and
FEFs) active during voluntary, goal-based orienting of
attention and the VAN (including the rTPJ and right ventral
frontal cortex) acting as a circuit breaker to the dorsal
network when attention is reoriented to salient events
outside the current focus, as during attentional capture
(Shulman et al., 2009; Indovina & Macaluso, 2007;
Serences et al., 2005; Downar et al., 2001; Corbetta et al.,
2000). Our whole-brain contrasts of distractor-present ver-
sus distractor-absent trials confirmed the involvement of
the VAN in the current task. However, whereas prior
studies have primarily focused analyses on these networks
to assess questions related to spatial capture and recovery,
here we demonstrate that category-selective visual object
processing areas are also influenced by this disruption to
control. How, exactly, the VAN communicates with
category-specific regions in ventral visual cortex to disrupt
filtering is an open question: Regions such as the FFA and
PPA may receive direct modulatory signals via feedback
from parietal VAN regions, or the “circuit breaker” signal
may instead be sent to earlier visual regions responsible
for basic visuospatial processing and then fed forward to
higher-level FFA and PPA.
Strikingly, we found that observers do not simply disen-
gage from the current attentional filter setting during
attentional capture; they appear to incidentally adopt an
errant filter setting. Specifically, in both FFA and PPA, acti-
vation reflecting the processing of the current nontarget
category (i.e., houses on attend-faces blocks) was tempo-
rarily greater than activation reflecting the current target
category. In conceptualizing the current experiment and
in the filter disruption theory framework more broadly,
we predicted that the category-tuned filter might be dis-
rupted under conditions of distraction. We did not predict
the disruption would be so extreme as to cause filter rever-
sal. The discovery that activity in ventral visual cortex
instead prioritizes goal-inconsistent information immedi-
ately after distraction raises new and interesting theoreti-
cal consequences of distraction that would have been
difficult to disentangle behaviorally. For example, behav-
ioral accuracy in this task declined substantially on
distractor-present compared to distractor-absent trials,
which could be consistent with any of the following
causes: spatial capture of attention away from the target
(attentional capture), a disruption of attentional focus to
the target category (filter disruption), and/or incidentally
prioritizing the nontarget stimulus category over the
target category (filter reversal). A unique appeal of this
neuroimaging approach is that we are able to assess the
processing of truly task-irrelevant information to reveal
new potential consequences of distraction, both predicted
and unpredicted, laying strong theoretical groundwork
for future studies.
Although the filter reversal was an unexpected finding
and not the main emphasis of this article, it was present
in all of our participants, and such a robust finding begs
speculation about why/how such a reversal could occur.
One possibility is that the reversal effect may be a result
of conditioning participants to switch between opposing
control settings. Over the course of the experiment,
participants alternated between only two goal states:
attend-faces and attend-houses. Under these conditions,
we speculate that when an observer loses control over
the goal-consistent filter (i.e., attend-faces), they may
automatically revert to the alternative goal (i.e., attend-
houses). An intriguing question for future study would
be to evaluate the nature of filter disruption in a task
requiring more than a binary attend Category A/ignore
Category B setting, for example, if more than two catego-
ries and/or possible attentional settings were involved.
1530
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Although the current data do not allow for more than
speculation about what is causing the reversal, they do
allow us to rule out at least one alternative explanation.
Specifically, the reversal does not seem to be an incidental
consequence of adaptation because of attending the same
category over a several minute block of trials. Our explor-
atory block phase analysis argues strongly against this: If
the filter reversal were because of adaptation, the reversal
should be more evident in the second half of a block than
the first half. This is the opposite of what we found: The
filter was disrupted more severely (inducing the reversal)
in the first half of a block.
Interestingly, this finding from the block phase analysis
suggests that filter disruption is weaker in the second half
of a block relative to the first half and that, over the course
of a block, participants regain some control over the filter
on distractor-present trials. Intriguingly, although the filter
disruption in PPA and FFA was less severe in the second
half of a block, our rTPJ analysis did not show the same
reduction: rTPJ activation was significantly stronger on
distractor-present relative to distractor-absent trials (as
expected from prior studies; Downar et al., 2001), and
the magnitude of this effect remained constant through-
out the block. As such, it appears that spatial attentional
capture was strong for the duration of the block, but its
consequences—to both attentional filtering and, to a
lesser extent, RTs—were mitigated over time. This is
notable given recent interest in learned distractor
suppression—more specifically, the finding that the
effects of distractors can be attenuated with increased
exposure ( Won & Geng, 2020) and the debate over proac-
tive versus reactive suppression (see Chelazzi et al., 2019,
for a review). Given the ability to simultaneously measure
rTPJ/ VAN activation, a neural filtering index in object-
processing areas, and behavioral RT, the paradigm intro-
duced here may be a useful future tool in examining topics
related to distractor habituation and other open questions
about attentional capture, contributing to our growing
knowledge of the broader consequences of attentional
capture and distraction.
Open Practices Statement
The experiment reported here was not formally preregis-
tered, but the design and analysis plan were proposed
before data collection as part of a Natural Sciences and
Engineering Research Council of Canada postdoctoral
fellowship grant by B. D. Deidentified data are available
on OSF via the link here.
Acknowledgments
This work was supported by grants from the National Institutes
of Health (R01-EY025648) and the National Science Foundation
(NSF 1848939) to J. G. and Natural Sciences and Engineering
Research Council of Canada postdoctoral fellowship to B. D.
Reprint requests should be sent to Blaire Dube, Department of
Psychology, The Ohio State University, Columbus, OH 43210,
or via e-mail: dube.25@osu.edu.
Author Contributions
Blaire Dube: Conceptualization; Data curation; Formal
analysis; Funding acquisition; Investigation; Methodology;
Project administration; Writing—Original draft;
Writing—Review & editing. Lasyapriya Pidaparthi: Data
curation; Formal analysis; Investigation; Project adminis-
tration; Writing—Original draft; Writing—Review & edit-
ing. Julie D. Golomb: Conceptualization; Formal analysis;
Funding acquisition; Investigation; Methodology; Project
administration; Resources; Software; Supervision;
Writing—Original draft; Writing—Review & editing.
Funding Information
Blaire Dube, Natural Sciences and Engineering Research
Council of Canada (htt ps://dx.doi.org /10.1 303 9
/501100000038), grant number: postdoctoral fellowship.
Julie D. Golomb, National Science Foundation (https://
dx.doi.org/10.13039/100000001), grant number: NSF
1848939; National Institutes of Health (https://dx.doi.org
/10.13039/100000002), grant number: R01-EY025648.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender
identification of first author/last author) publishing in
the Journal of Cognitive Neuroscience ( JoCN) during this
period were M(an)/M = .407, W(oman)/M = .32, M/ W =
.115, and W/ W = .159, the comparable proportions for the
articles that these authorship teams cited were M/M =
.549, W/M = .257, M/ W = .109, and W/ W = .085 (Postle
and Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN
encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance.
REFERENCES
Bacigalupo, F., & Luck, S. J. (2019). Lateralized suppression of
alpha-band EEG activity as a mechanism of target processing.
Journal of Neuroscience, 39, 900–917. https://doi.org/10
.1523/JNEUROSCI.0183-18.2018, PubMed: 30523067
Baldauf, D., & Desimone, R. (2014). Neural mechanisms of
object-based attention. Science, 344, 424–427. https://doi.org
/10.1126/science.1247003, PubMed: 24763592
Bonetti, F., & Turatto, M. (2019). Habituation of oculomotor
capture by sudden onsets: Stimulus specificity, spontaneous
recovery and dishabituation. Journal of Experimental
Psychology: Human Perception and Performance, 45,
Dube, Pidaparthi, and Golomb
1531
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
264–284. https://doi.org/10.1037/xhp0000605, PubMed:
30570321
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163
/156856897X00357, PubMed: 9176952
Bundesen, C. (1990). A theory of visual attention. Psychological
Review, 97, 523–547. https://doi.org/10.1037/0033-295X.97.4
.523, PubMed: 2247540
Chelazzi, L., Marini, F., Pascucci, D., & Turatto, M. (2019).
Getting rid of visual distractors: The why, when, how, and
where. Current Opinion in Psychology, 29, 135–147. https://
doi.org/10.1016/j.copsyc.2019.02.004, PubMed: 30856512
Chen, J., Leber, A. B., & Golomb, J. D. (2019). Attentional
capture alters feature perception. Journal of Experimental
Psychology: Human Perception and Performance, 45,
1443–1454. https://doi.org/10.1037/xhp0000681, PubMed:
31464467
Corbetta, M., Kincade, J. M., Ollinger, J. M., Mcavoy, M. P., &
Gordon, L. (2000). Voluntary orienting is dissociated from
target detection in human posterior parietal cortex. Nature
Neuroscience, 3, 521. https://doi.org/10.1038/73009
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting
system of the human brain: From environment to theory of
mind. Neuron, 58, 306–324. https://doi.org/10.1016/j.neuron
.2008.04.017, PubMed: 18466742
Desimone, R., & Duncan, J. S. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222. https://doi.org/10.1146/annurev.ne.18.030195
.001205, PubMed: 7605061
Downar, J., Crawley, A. P., Mikulis, D. J., & Davis, K. D. (2001).
The effect of task relevance on the cortical response to
changes in visual and auditory stimuli: An event-related fMRI
study. Neuroimage, 14, 1256–1267. https://doi.org/10.1006
/nimg.2001.0946, PubMed: 11707082
Dube, B., & Golomb, J. D. (2021). Perceptual distraction causes
visual memory encoding intrusions. Psychonomic Bulletin
and Review, 28, 1592–1600. https://doi.org/10.3758/s13423
-021-01937-6, PubMed: 34027621
Epstein, R., & Kanwisher, N. (1998). A cortical representation
the local visual environment. Nature, 392, 598–601. https://
doi.org/10.1038/33402, PubMed: 9560155
Esterman, M., Tamber-Rosenau, B. J., Chiu, Y. C., & Yantis, S.
(2010). Avoiding non-independence in fMRI data analysis:
Leave one subject out. Neuroimage, 50, 572–576. https://doi
.org/10.1016/j.neuroimage.2009.10.092, PubMed: 20006712
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992).
Involuntary covert orienting is contingent on attentional
control settings. Journal of Experimental Psychology:
Human Perception and Performance, 18, 1030–1044.
https://doi.org/10.1037/0096-1523.18.4.1030, PubMed:
1431742
Golomb, J. D., & Kanwisher, N. (2012). Higher level visual
cortex represents retinotopic, not spatiotopic, object
location. Cerebral Cortex, 22, 2794–2810. https://doi.org/10
.1093/cercor/bhr357, PubMed: 22190434
Hampshire, A., Duncan, J., & Owen, A. M. (2007). Selective
tuning of the blood oxygenation level-dependent response
during simple target detection dissociates human
frontoparietal subregions. Journal of Neuroscience, 27,
6219–6223. https://doi.org/10.1523/JNEUROSCI.0851-07
.2007, PubMed: 17553994
Hemond, C. C., Kanwisher, N. G., & Op de Beeck, H. P. (2007).
A preference for contralateral stimuli in human object- and
face-selective cortex. PLoS One, 2, 3–7. https://doi.org/10
.1371/journal.pone.0000574, PubMed: 17593973
Henderson, J. M. (2003). Human gaze control during real-world
scene perception. Trends in Cognitive Sciences, 7, 498–504.
https://doi.org/10.1016/j.tics.2003.09.006, PubMed: 14585447
Hoffman, J. E., & Nelson, B. (1981). Spatial selectivity in visual
search. Perception & Psychophysics, 30, 283–290. https://doi
.org/10.3758/BF03214284, PubMed: 7322804
Hsieh, P. J., Colas, J. T., & Kanwisher, N. G. (2012). Pre-stimulus
pattern of activity in the fusiform face area predicts face
percepts during binocular rivalry. Neuropsychologia, 50,
522–529. https://doi.org/10.1016/j.neuropsychologia.2011.09
.019, PubMed: 21952195
Hu, S., Bu, Y., Song, Y., Zhen, Z., & Liu, J. (2009). Dissociation
of attention and intention in human posterior parietal cortex:
An fMRI study. European Journal of Neuroscience, 29,
2083–2091. https://doi.org/10.1111/j.1460-9568.2009.06757.x,
PubMed: 19453626
Indovina, I., & Macaluso, E. (2007). Dissociation of stimulus
relevance and saliency factors during shifts of visuospatial
attention. Cerebral Cortex, 17, 1701–1711. https://doi.org/10
.1093/cercor/bhl081, PubMed: 17003078
Jonides, J., & Irwin, D. E. (1981). Capturing attention.
Cognition, 10, 145–150. https://doi.org/10.1037/13239-002
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The
fusiform face area: A module in human extrastriate cortex
specialized for face perception. Journal of Neuroscience, 17,
4302–4311. https://doi.org/10.1523/JNEUROSCI.17-11-04302
.1997, PubMed: 9151747
Liesefeld, H. R., Liesefeld, A. M., Töllner, T., & Müller, H. J.
(2017). Attentional capture in visual search: Capture and
post-capture dynamics revealed by EEG. Neuroimage, 156,
166–173. https://doi.org/10.1016/j.neuroimage.2017.05.016,
PubMed: 28502842
Liu, T., & Hou, Y. (2011). Global feature-based attention to
orientation. Journal of Vision, 11, 1–8. https://doi.org/10
.1167/11.10.1, PubMed: 21900371
Liu, T., & Mance, I. (2011). Constant spread of feature-based
attention across the visual field. Vision Research, 51, 26–33.
https://doi.org/10.1016/j.visres.2010.09.023, PubMed:
20887745
Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during visual
search: Evidence from human electrophysiology. Journal
of Experimental Psychology: Human Perception and
Performance, 20, 1000–1014. https://doi.org/10.1037/0096
-1523.20.5.1000, PubMed: 7964526
Macmillan, N. A., & Creelman, C. D. (1990). Response bias:
Characteristics of detection theory, threshold theory, and
“nonparametric” indexes. Psychological Bulletin, 107,
401–413. https://doi.org/10.1037/0033-2909.107.3.401
Mccarthy, G., Puce, A., Gore, J. C., & Truett, A. (1997). Face-
specific processing in the human fusiform gyms. Journal of
Cognitive Neuroscience, 9, 605–610. https://doi.org/10.1162
/jocn.1997.9.5.605, PubMed: 23965119
Morey, R. D. (2008). Confidence intervals from normalized data:
A correction to Cousineau (2005). Tutorials in Quantitative
Methods for Psychology, 4, 61–64. https://doi.org/10.20982
/tqmp.04.2.p061
Nako, R., Wu, R., & Eimer, M. (2014). Rapid guidance of visual
search by object categories. Journal of Experimental
Psychology: Human Perception and Performance, 40,
50–60. https://doi.org/10.1037/a0033228, PubMed: 23796065
Naselaris, T., Allen, E., & Kay, K. (2021). Extensive sampling for
complete models of individual brains. Current Opinion in
Behavioral Sciences, 40, 45–51. https://doi.org/10.1016/j
.cobeha.2020.12.008
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Natale, E., Marzi, C. A., & Macaluso, E. (2010). Right
temporal–parietal junction engagement during spatial
reorienting does not depend on strategic attention control.
Neuropsychologia, 48, 1160–1164. https://doi.org/10.1016/j
.neuropsychologia.2009.11.012, PubMed: 19932706
O’Craven, K. M., & Kanwisher, N. (2000). Mental imagery of
faces and places activates corresponding stimulus-specific
1532
Journal of Cognitive Neuroscience
Volume 34, Number 8
brain regions. Journal of Cognitive Neuroscience, 12,
1013–1023. https://doi.org/10.1162/08989290051137549,
PubMed: 11177421
Olivers, C. N. L., Meijer, F., & Theeuwes, J. (2006). Feature-
based memory-driven attentional capture: Visual working
memory content affects visual attention. Journal of
Experimental Psychology: Human Perception and
Performance, 32, 1243–1265. https://doi.org/10.1037/0096
-1523.32.5.1243, PubMed: 17002535
Peters, J. C., Roelfsema, P. R., & Goebel, R. (2012). Task-
relevant and accessory items in working memory have
opposite effects on activity in extrastriate cortex. Journal of
Neuroscience, 32, 17003–17011. https://doi.org/10.1523
/JNEUROSCI.0591-12.2012, PubMed: 23175851
Remington, R. W., Johnston, J. C., & Yantis, S. (1992).
Involuntary attentional capture by abrupt onsets. Perception
& Psychophysics, 51, 279–290. https://doi.org/10.3758
/BF03212254, PubMed: 1561053
Saenz, M., Buracas, G. T., & Boynton, G. M. (2002). Global
effects of feature-based attention in human visual cortex.
Nature Neuroscience, 5, 631–632. https://doi.org/10.1038
/nn876, PubMed: 12068304
Sàenz, M., Buraĉas, G. T., & Boynton, G. M. (2003). Global
feature-based attention for motion and color. Vision
Research, 43, 629–637. https://doi.org/10.1016/S0042-6989
(02)00595-3, PubMed: 12604099
Serences, J. T., & Boynton, G. M. (2007). Feature-based
attentional modulations in the absence of direct visual
stimulation. Neuron, 55, 301–312. https://doi.org/10.1016/j
.neuron.2007.06.015, PubMed: 17640530
Serences, J. T., Schwarzbach, J., Courtney, S. M., Golay, X., &
Yantis, S. (2004). Control of object-based attention in human
cortex. Cerebral Cortex, 14, 1346–1357. https://doi.org/10
.1093/cercor/bhh095, PubMed: 15166105
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth,
H. E., & Yantis, S. (2005). Coordination of voluntary and
stimulus-driven attentional control in human cortex.
Psychological Science, 16, 114–122. https://doi.org/10.1111/j
.0956-7976.2005.00791.x, PubMed: 15686577
Shulman, G. L., Astafiev, S. V., Franke, D., Pope, D. L. W.,
Snyder, A. Z., McAvoy, M. P., et al. (2009). Interaction of
stimulus-driven reorienting and expectation in ventral and
dorsal frontoparietal and basal ganglia-cortical networks.
Journal of Neuroscience, 29, 4392–4407. https://doi.org/10
.1523/JNEUROSCI.5609-08.2009, PubMed: 19357267
Stanislaw, H., & Todorov, N. (1999). Calculation of signal
detection theory measures. Behavior Research Methods,
Instruments, & Computers, 3, 37–149. https://doi.org/10.3758
/BF03207704, PubMed: 10495845
Stein, T., & Peelen, M. V. (2017). Object detection in natural
scenes: Independent effects of spatial and category-based
attention. Attention, Perception, & Psychophysics, 79,
738–752. https://doi.org/10.3758/s13414-017-1279-8, PubMed:
28138945
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic atlas
of the human brain. 3-Dimensional proportional system: An
approach to cerebral imaging. New York: Thieme.
Theeuwes, J., & Burger, R. (1998). Attentional control during
visual search: The effect of irrelevant singletons. Journal
of Experimental Psychology: Human Perception and
Performance, 24, 1342–1353. https://doi.org/10.1037/0096
-1523.24.5.1342, PubMed: 9778827
Theeuwes, J., Kramer, A. F., Hahn, S., & Irwin, D. E. (1998). Our
eyes do not always go where we want them to go: Capture of
the eyes by new objects. Psychological Science, 9, 379–385.
https://doi.org/10.1111/1467-9280.00071
Theeuwes, J., Kramer, A. F., Hahn, S., Irwin, D. E., & Zelinsky,
G. J. (1999). Influence of attentional capture on oculomotor
control. Journal of Experimental Psychology: Human
Perception and Performance, 25, 1595–1608. https://doi.org
/10.1037/0096-1523.25.6.1595, PubMed: 10641312
Tong, F., Nakayama, K., Vaughan, J. T., & Kanwisher, N. (1998).
Binocular rivalry and visual awareness in human extrastriate
cortex. Neuron, 21, 753–759. https://doi.org/10.1016/S0896
-6273(00)80592-9, PubMed: 9808462
Treue, S., & Martínez Trujillo, J. C. (1999). Feature-based
attention influences motion processing gain in macaque
visual cortex. Nature, 399, 575–579. https://doi.org/10.1038
/21176, PubMed: 10376597
Turatto, M., Bonetti, F., & Pascucci, D. (2018). Filtering visual
onsets via habituation: A context-specific long-term memory
of irrelevant stimuli. Psychonomic Bulletin & Review, 25,
1028–1034. https://doi.org/10.3758/s13423-017-1320-x,
PubMed: 28547537
Turk-Browne, N. B., Norman-Haignere, S. V., & Mccarthy, G.
(2010). Face-specific resting functional connectivity between
the fusiform gyrus and posterior superior temporal sulcus.
Frontiers in Human Neuroscience, 4, 1–15. https://doi.org
/10.3389/fnhum.2010.00176, PubMed: 21151362
Wang, B., van Driel, J., Ort, E., & Theeuwes, J. (2019).
Anticipatory distractor suppression elicited by statistical
regularities in visual search. Journal of Cognitive
Neuroscience, 31, 1535–1548. https://doi.org/10.1162/jocn_a
_01433, PubMed: 31180265
White, A. L., & Carrasco, M. (2011). Feature-based attention
involuntarily and simultaneously improves visual
performance across locations. Journal of Vision, 11, 1–10.
https://doi.org/10.1167/11.6.15, PubMed: 21602553
Wojciulik, E., Kanwisher, N., & Driver, J. (1998). Covert visual
attention modulates face-specific activity in the human
fusiform gyrus: fMRI study. Journal of Neurophysiology,
79, 1574–1578. https://doi.org/10.1152/jn.1998.79.3.1574,
PubMed: 9497433
Won, B. Y., & Geng, J. J. (2020). Passive exposure attenuates
distraction during visual search. Journal of Experimental
Psychology: General, 149, 1987–1995. https://doi.org/10.1037
/xge0000760, PubMed: 32250138
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and
selective attention: Evidence from visual search. Journal
of Experimental Psychology: Human Perception and
Performance, 10, 601–621. https://doi.org/10.1037/0096-1523
.10.5.601, PubMed: 6238122
Yi, D. J., Kelley, T. A., Marois, R., & Chun, M. M. (2006).
Attentional modulation of repetition attenuation is
anatomically dissociable for scenes and faces. Brain
Research, 1080, 53–62. https://doi.org/10.1016/j.brainres
.2006.01.090, PubMed: 16507300
Zhang, W., & Luck, S. J. (2009). Feature-based attention modulates
feedforward visual processing. Nature Neuroscience, 12,
24–25. https://doi.org/10.1038/nn.2223, PubMed: 19029890
Dube, Pidaparthi, and Golomb
1533
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
2
1
2
0
3
3
1
6
5
/
/
j
o
c
n
_
a
_
0
1
8
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3