Toward Open-Ended Fraternal
Transitions in Individuality
Abstract The emergence of new replicating entities from the union
of simpler entities characterizes some of the most profound events
in natural evolutionary history. Such transitions in individuality are
essential to the evolution of the most complex forms of life. Così,
understanding these transitions is critical to building artificial systems
capable of open-ended evolution. Alas, these transitions are
challenging to induce or detect, even with computational organisms.
Here, we introduce the DISHTINY (Distributed Hierarchical
Transitions in Individuality) platform, which provides simple cell-like
organisms with the ability and incentive to unite into new individuals
in a manner that can continue to scale to subsequent transitions.
The system is designed to encourage these transitions so that they can
be studied: Organisms that coordinate spatiotemporally can maximize
the rate of resource harvest, which is closely linked to their
reproductive ability. We demonstrate the hierarchical emergence of
multiple levels of individuality among simple cell-like organisms that
evolve parameters for manually designed strategies. During evolution,
we observe reproductive division of labor and close cooperation
among cells, including resource-sharing, aggregation of resource
endowments for propagules, and emergence of an apoptosis response
to somatic mutation. Many replicate populations evolved to direct
their resources toward low-level groups (behaving like multicellular
individuals), and many others evolved to direct their resources toward
high-level groups (acting as larger-scale multicellular individuals).
Matthew Andres Moreno*
Michigan State University
mmore500@msu.edu
Charles Ofria
Michigan State University
ofria@msu.edu
Keywords
Open-ended evolution, major transitions
in evolution, fraternal transitions,
multicellularity, cooperation, evolutionary
dynamics
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
1 introduzione
Artificial life researchers design systems that exhibit properties of biological life in order to better
understand their dynamics and, often, to apply these principles toward engineering applications such
as artificial intelligence [4]. Studies of evolution have been of particular interest to the community,
especially in regard to how organisms are produced with increasing sophistication and complexity
[11]. This particular subject is often described as open-ended evolution. Although precise definitions
and measures of open-ended evolution are still being established, this term is generally understood
to refer to evolving systems that exhibit the continued production of novelty [19]. Evolutionary
transitions in individuality, which are key to the complexification and diversification of biological
* Corresponding author.
Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(S) and do not necessarily
reflect the views of the National Science Foundation.
© 2019 Massachusetts Institute of Technology Artificial Life 25: 117–133 (2019) doi:10.1162/artl_a_00284
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
life [14], have been highlighted as key research targets with respect to the question of open-ended
evolution [3, 17]. In an evolutionary transition of individuality, a new, more complex replicating
entity is derived from the combination of cooperating replicating entities that have irrevocably en-
twined their long-term fates [20]. In particular, we focus on fraternal transition in individuality,
events where closely related kin come together or stay together to form a higher-level organism
[15]. Eusocial insect colonies and multicellular organisms exemplify this phenomenon [14]. Like
the definition of open-ended evolution, the notion of what constitutes an evolving individual is
not concretely established. Commonly indicated features include: close coordination and coopera-
zione, reproductive division of labor, reproductive bottlenecks, and loss of ability to replicate inde-
pendently [5, 7].
Our appreciation of fraternal transitions in individuality benefits from experimental work probing
the origins of multicellularity. In the biological domain, Ratcliff et al. have demonstrated evolution of
multicellularity in yeast, deriving fraternal clusters of cells that cling together in order to maximize
their settling rate [16]. The contributions of Goldsby and collaborators are particularly notable
among computational artificial life work on the origins of multicellularity. Their evolutionary exper-
iments track a population composed of demes, distinct spatial domains inhabited by clonal colonies of
cells. Two distinct types of reproduction occur: (1) cells reproducing within demes and (2) deme
reproduction, where a target deme is sterilized and then reinoculated with genetic material from the
parent deme. With such methods, Goldsby et al. have studied division of labor [8, 9], the origin of
soma [10], and the evolution of morphological development [11]. We aspire to complement deme-
based approaches with a framework where higher-level individuality unfolds via cellular reproduc-
tions within a single unified space. In particular, we are interested in the potential for such a system
to undergo nested hierarchical transitions.
Major challenges in studying evolutionary transitions in individuality include (1) determining the
environmental conditions that will promote such a transition and then (2) recognizing that a tran-
sition has occurred. In order to begin exploring transitions in individuality, we must devise a system
in which we expect such transitions to occur repeatably and in a detectable manner. Once we can
consistently induce and observe evolutionary transitions in individuality, we may subsequently pro-
ceed to relax aspects of such a system to explore in greater detail what conditions are necessary to
induce transitions and how transitions can be detected. For now, we will focus on these initial goals
in the context of fraternal transitions in individuality.
A tal fine, we introduce the DISHTINY (Distributed Hierarchical Transitions in Individuality)
platform, which seeks to achieve the evolution of transitions in individuality by explicitly registering
organisms in cooperating groups that coordinate spatiotemporally to maximize the harvest of a re-
source. Detection of such a transition in DISHTINY is accomplished by identifying resource-
sharing and reproductive division of labor among organisms registered to the same cooperating
group. We designed this system such that hierarchical transitions across an arbitrary number of levels
of individuality can be selected for and meaningfully detected. We have focused this system on a rigid
form of major transition using simple organisms, but the underlying principles can be applied to a
wide range of artificial life systems. Inoltre, DISHTINY is decentralized and amenable to mas-
sive parallelization via distributed computing. We believe that such scalability—with respect to both
concept and implementation—is an essential consideration in the pursuit of artificial systems capable
of generating complexity and novelty rivaling that of biological life via open-ended evolution [1, 2].
2 Methods
In order to demonstrate that the DISHTINY platform selects for detectable hierarchical transitions
in individuality, we performed experiments where cell-like organisms evolved parameters to control
manually designed strategies such as resource-sharing, reproductive decision making, and apoptosis.
We will first cover the design of the DISHTINY platform and then describe the simple cell-like
organisms we used to evaluate the platform.
118
Artificial Life Volume 25, Numero 2
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
Figura 1. Activation signaling and net resource collection for three different-size same-channel networks during a
resource wave event. At the top, a resource wave is depicted propagating over three updates and then ceasing for four
updates (da sinistra a destra). In row (UN), a small two-cell channel-signaling group (far left, in green) is activated; tracking the
resource wave (top) yields a small net resource harvest (far right). In row (B), an intermediate-size 13-cell channel-
signaling group yields a high net resource harvest. Finalmente, in row (C), a large 29-cell channel-signaling group incurs a
net negative resource harvest. In rows (UN), (B), E (C), dark purple indicates the active state, light purple indicates
the quiescent state, and white indicates the ready state.
2.1 DISHTINY
DISHTINY allows cell-like organisms to replicate across a toroidal grid. Over discrete time steps
(updates), the cells can collect a continuous-valued resource. Once sufficient resource has accrued,
cells may pay 8.0 units of resource to place a daughter cell on an adjoining tile of the toroidal grid
(cioè., reproduce), replacing any existing cell already there. As cells reproduce, they can choose to
include offspring in the parentʼs cooperating signaling channel group or force offspring to create a
new cooperating signaling channel group.
As shown at the top of Figure 1, resources appear at a single point and then spread outwards,
update by update, in a diamond-shaped wave, disappearing when the expanding wave reaches a
predefined limit. Cells must be in a costly activated state to collect resource as it passes. The cell
at the starting position of a resource wave is automatically activated, and will send the activate signal
to neighboring cells on the same signaling channel. The newly activated cells, in turn, activate their
own neighbors registered to the same signaling channel. Neighbors registered to other signaling
channels do not activate. Each cell, after sending the activation signal, enters a temporary quiescent
state so as not to reactivate from the signal. In this manner, cells sharing a signaling channel activate
in concert with the expanding resource wave. As shown Figure 1(UN), (B), the rate of resource collection
for a cell is determined by the size and shape of its same-channel signaling network; small or
fragmented same-channel signaling networks will frequently miss resource as it passes by.
Each cell pays a resource cost when it activates. This cost is outweighed by the resource collected,
so that cells that activate in concert with a resource wave derive a net benefit. Recall, Anche se, Quello
resource waves have a limited extent. Cells that activate outside the extent of a resource wave or
activate out of sync with the resource wave (due to an indirect path from the cell that originated
the signal) pay the activation cost but collect no resource. Cells that frequently activate erroneously
use up their resource and die. In our implementation, organisms that accrue a resource debt of −11 or
greater are killed. This erroneous activation scenario is depicted in Figure 1(C).
Artificial Life Volume 25, Numero 2
119
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
In this manner, Goldilocks—not too small and not too big—signaling networks are selected for.
Based on a randomly chosen starting location, resource wave starting points (seeds) are tiled over
the toroidal grid in such a way that the extents of the resource waves touch, but do not overlap. Tutto
waves start and proceed synchronously; when they complete, the next resource waves are seeded.
This process ensures that selection for Goldilocks same-channel signaling networks is uniformly
distributed over the toroidal grid.
Cells control the size and shape of their same-channel signaling group through strategic reproduction.
Three choices are afforded: whether to reproduce at all, where among the four adjoining tiles of
the toroidal grid to place their offspring, and whether the offspring should be registered to the
parentʼs signaling channel or be given a random channel ID (in the range 1 A 222). No guarantees
are made about the uniqueness of a newly generated channel ID, but chance collisions are rare.
Hierarchical levels are introduced into the system through multiple separate, but overlaid, instan-
tiations of this resource-wave–channel-signaling scheme. We refer to each independent resource-
wave–channel-signaling system as a level. In our experiments, we allowed two such levels, identified
here as level one and level two. On level one, resource waves extended a radius of three toroidal
tiles. On level two they extended a radius of 24 toroidal tiles. On both levels, activated cells netted
+1.0 resource from a resource wave, but suffered an activation penalty of −5.0 if no resource was
available. Due to the different radii of resource waves on different levels, level one selects for small
same-channel signaling networks and level two selects for large same-channel signaling networks.
Cells were marked with two separate channel IDs, one for level one and another for level two.
We enforced hierarchical nesting of same-channel signaling networks during reproduction: Daughter
cells may inherit neither channel ID, just the level-two channel ID, or both channel IDs. Daughter
cells may not inherit only the level-one channel ID while having a different level-two channel ID.
The distribution of IDs across the level-two and level-one channels can be envisioned by analogy to
political countries and territories. Each country (cioè., level-two channel network) may have one or
many territories (cioè., level-one channel networks). Tuttavia, no territory spans more than one
country. Figura 2 depicts hierarchically nested channel states at the end of three evolutionary runs.
Channel IDs enable straightforward detection of an evolutionary transition in individuality.
Because common channel IDs may only arise systematically through inheritance, common channel
IDs indicate a close hereditary relationship in addition to a close cooperative relationship. Because
new channel IDs arise first in a single cell, same-channel signaling networks are reproductively
bottlenecked, ensuring meaningful reproductive lineages at the level of the same-channel signaling
rete. To recognize an evolutionary transition in individuality, we therefore evaluate:
1. Do cells with the same channel ID choose to share resources (per esempio., cooperate)?
2.
Is there division of reproductive labor between members of the same channel (per esempio., do
cells at the interior of a network cede reproduction to those at the periphery)?
If these conditions are met among cells sharing the same level-one channel, a first-level transition in in-
dividuality may have occurred. Likewise, if these conditions are met among cells sharing the same level-
two channel, a second-level transition in individuality may have occurred. In either case, observation of
altruistic behavior, such as an apoptosis response to mutation, would further evidence a transition.
2.2 Organisms
We performed our experiments using cell-like organisms composed of 15 floating-point parame-
ters, each controlling a specific manually designed strategy component pertinent to transitions in
individuality (viz., reproductive division of labor, resource pooling, apoptosis, propagule genera-
zione, and propagule endowment). These particular cell-like organisms are in no way inherent in the
DISHTINY platform, but were merely developed to study transitions using as simple a model
system as feasible. On reproduction, we applied mutation to each parameter independently with
probability 0.00005.
120
Artificial Life Volume 25, Numero 2
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. End state of same-channel signaling networks in replicates where resource was exclusively allocated to first-
level channel pools (UN), was split evenly between first- and second-level channel pools (B), and was primarily allocated to
second-level channel pools (C). Level-one channels are coded by color saturation, and level-two channels are coded by
color hue. A single cell-like organism occupies each grid tile except for black tiles, which are empty. Level-one same-
channel groups appear as uniformly colored clumps, bounded by a white border. Level-two same-channel groups appear
as same-hue amalgamations of level-one groups, bounded by a black border.
The aversion parameters (A1 and A2) allow cells to avoid reproducing over neighbors sharing the
same signaling channel. Specifically, they control the probability that a cell declines to supplant a
neighbor sharing the same level-one (A1) or level-two (A2) channel ID. If a cell declines to place
its offspring in all four adjoining tiles, it does not reproduce. Mutation is performed by a redraw
from the uniform distribution U (−0.5, 1.5) clamped to the range [0, 1].
The resource allocation parameters control the proportion of resources that go to the cellʼs stockpile
(Pc), its level-one channelʼs resource pool (P1), or its level-two channelʼs resource pool (P2). These
parameters are initialized by a draw from U (−1.0, 2.0) clamped to the range [0, 1] and mutated by
addition of a normal value drawn from N (0.0, 0.2), with the result clamped to the range [0, 1]. IL
set Pc , P1, P2 is always normalized to sum to 1.
Artificial Life Volume 25, Numero 2
121
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
Channel resource pools are identical to an organismʼs individual stockpile, except that any deficit
is distributed evenly among the individual organismʼs stockpile. On every update, cells can spend
from their individual stockpile to reproduce, or from a channel pool, with priority given to cells
nearest to the centroid of that poolʼs members. Thereby, pool-funded reproduction fills in a
same-channel signaling network from the inside out and helps produce diamond-shaped same-
channel signaling networks. (Distance is measured using the taxicab metric.)
Channel cap parameters C1 and C2 regulate the size of same-channel signaling networks. When an
organism reproduces, it checks the size of its level-one signaling network against C1, and the size of
its level-two signaling group against C2. If neither cap is met or exceeded, then the organism will
produce an offspring sharing both of its channel IDs. If only the C1 cap is exceeded, then the or-
ganism will produce an offspring with new level-one channel ID but identical level-two channel ID.
Finalmente, if the C2 cap is exceeded, then the organism will produce an offspring with new IDs for both
channels. For level-one caps, these parameters are initialized by a draw from U (0.0, 16.0). For level-
two caps, they are initialized by a draw from U (0.0, 128.0). Both are mutated by addition of a value
drawn from N (0.0, 24.0) with the result clamped to be non-negative.
The endowment parameters Ec , E1, and E2 determine the amount of resource provided to offspring.
This endowment is paid as an additional cost by the cell stockpile (or same-channel resource pool)
funding a reproduction. The full amount of the received endowment is divided between the daugh-
ter cellʼs stockpile, level-one same-channel resource pool, and level-two same-channel resource pool
according to the offspringʼs resource allocation parameters. Ec is the endowment amount paid to an
offspring that shares both channel IDs of the parent; E1 is the endowment paid to an offspring that
shares just the level-two channel ID of the parent; and E2 is the endowment paid to an offspring
that shares neither the level-one nor the level-two channel ID of the parent. Endowed resources
help new-channel propagules to rapidly grow their signaling network in order to begin collecting
resource at a rate competitive with other well-established same-channel signaling networks. In order
that adequate resource remain to ensure parental stability, endowment was paid out only after twice
the endowment amount had been accrued (leaving an amount of resource equal to the endowment
remaining with the parent). Cell-level endowments are initialized by a draw from U (0.0, 5.0). Level-
one endowments are initialized by a draw from U (0.0, 80.0). Level-two endowments are initialized
by a draw from U (0.0, 405.0). All endowments are mutated by addition of a value drawn from N
(0.0, 10.0) with the result clamped to be non-negative.
Parameters Mc , M1, and M2 control the apoptosis response to mutation. Each time that a mutation occurs
during reproduction, the mutated offspring attempts suicide with probability Mc if it shares both channel
IDs of its parent, probability M1 if it shares just the level-two channel ID of its parent, and probability
M2 if it shares neither channel ID of the parent. The Mx value applied is from the offspringʼs geno-
type after mutation. Attempted suicide succeeds 90% of the time. This capacity enables first- O
second-level individuals to combat somatic mutation. Initialization and mutation of each of these
parameters is performed by a redraw from the distribution U (−0.5, 1.5) clamped to the range [0, 1].
Finalmente, parameters S1 and S2 fine-tune site choice for offspring placement. If an organism is placing an
offspring with identical channel IDs, with probability S1 the four possible sites for offspring place-
ment are considered in order of increasing distance from the centroid of the parentʼs level-one
signaling network. If an organism is placing an offspring with identical level-two channel ID but
different level-one channel ID, with probability S2 the four possible sites for offspring placement
are considered in order of increasing distance from the centroid of the parentʼs level-two same-
channel signaling network. Otherwise, the four possible sites for offspring placement are considered
in a random order. Initialization and mutation are performed by a draw from the distribution
U (−0.5, 1.5) clamped to the range [0, 1].
2.3 Treatments
Our standard treatment was designed to assess the evolutionary trajectories of populations in
DISHTINY. We seeded each tile on the 120 × 120 toroidal grid with a randomized organism
122
Artificial Life Volume 25, Numero 2
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
and ran the simulation for 20 million updates. In order to facilitate turnover, we culled the
population intermittently. Starting at update 500,000, and every 50,000 updates thereafter, we ran-
domly selected second-level channel IDs and killed all cells with that channel ID, continuing until
almeno 5% of grid tiles were empty. We performed 50 replicates within this treatment. On average,
each cellular generation took just over 500 updates. Across all successive 10,000 update segments
of all replicates, the mean number of cellular generations elapsed per 10,000 updates was 19.2 con
a standard deviation of 2.7 cellular generations per 10,000 updates.
In order to untangle the influence of same-channel signaling networks with respect to kin recog-
nition versus cooperation to increase the resource collection rate, we performed control evolutionary
trials where same-channel signaling networks did not affect the cellular resource collection rate.
Under control conditions, same-channel signaling networks just helped cells recognize other related
cells. In our implementation, this treatment corresponded to a constant per-update inflow of 0.02
resource units into all cells. All cells were activated (in order to take up the resource) at all updates,
and no cost for activation was assessed. We chose this resource inflow rate in order to approximately
match the cellular generation rate of the control treatment to that of the standard treatment. In cont-
trol runs, each cellular generation took around 450 updates. Across all successive 10,000 update seg-
ments of all replicates, the mean number of cellular generations elapsed per 10,000 updates was 22.0
with a standard deviation of 2.0 cellular generations per 10,000 updates. Due to checkpoint-restart
failures on our compute cluster, control experiments were curtailed at 3 million updates. All other
aspects of control runs, including culling and the functionality of all lifestyle parameters, were other-
wise identical to standard conditions. We performed 50 replications of the control treatment.
In standard evolutionary runs, we observed a spectrum of evolved resource-caching strategies.
To assess the relative fitness of these evolved organisms, we ran competitions between the most-
common genotype from three standard evolutionary runs. The first genotype allocated resource ex-
= 1.0), the second split resource evenly
clusively to its first-level same-channel resource pool (cioè., P1
= 0.5), and the third allocated
between its first-level and second-level resource pools (cioè., P1
resource primarily to the second-level resource pool (cioè., P2 > P1). (No most-common genotypes
allocated resource exclusively to the second-level resource pool.) We seeded each competition with
three copies of each genotype, uniformly spaced over the 120 × 120 toroidal grid with random
arrangement. Each competition lasted 2 million updates. We performed 50 runs in this experiment.
= P2
2.4 Implementation
We performed our computational experiments at the Michigan State University High Performance
Computing Center. Each replicate of standard evolutionary experiments required approximately six
days of compute time to reach 20 million updates. Each replicate of control evolutionary experi-
ments expended approximately two days of compute time to reach 3 million updates. Control runs
were somewhat slower than standard runs, perhaps due to increased computational overhead asso-
ciated with bookkeeping for the larger same-channel groups that evolved under the control condi-
zioni. Each replicate of competition experiments consumed approximately ten hours of compute
time. For standard evolutionary experiments, data processing required approximately four hours
of compute time per run. Other data processing was computationally negligible.
We implemented our experimental system using the Empirical library for scientific software de-
velopment in C++, available at https://github.com/devosoft/Empirical. The code used to perform
and analyze our experiments, our figures, data from our experiments, and a live in-browser demo of
our system is available via the Open Science Framework at https://osf.io/ewvg8/.
3 Results and Discussion
3.1 Standard Evolutionary Experiments
A spectrum of resource allocation strategies, ranging from (io) allocation purely to level-one same-
channel resource pools to (ii) primarily to level-two same-channel resource pools, were observed at
Artificial Life Volume 25, Numero 2
123
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
the conclusion of different runs of our evolutionary simulation (mean cellular generation 37,168 con
standard deviation 4,684). We interpret these outcomes as ranging between individuality at the level
of first-level same-channel groups to individuality at the level of second-level same-channel groups.
Figura 2 shows the level-one and level-two signaling networks at the end of runs where first-, split-,
and second-level resource allocation evolved, rispettivamente. First-level allocators form somewhat ir-
regular level-two amalgamations of diverse level-one networks. Second-level allocators form highly
regular diamond-shaped level-two signaling networks. Split-allocation individuals exhibit a level-two
phenotype of intermediate regularity. Figura 3 shows a time series of signaling network snapshots in
an evolutionary run where second-level individuality evolved.
Tavolo 1 summarizes the most-common genotypes observed at the end of our evolutionary sim-
ulazioni. In the standard treatment, all evolved genotypes had A2 fixed at 1.0. So, reproduction over
cells sharing the same level-two channel was universally avoided; genotypes evolved so that cells
declined to reproduce when they were located in the interior of level-two same-channel signaling
networks.
Tuttavia, a variety of resource-caching strategies evolved. The most-abundant genotypes at the
end of nine evolutionary runs exclusively cached resource in organismsʼ level-one signaling net-
= 1.0). We observed strategies where resource was primarily, but not entirely,
workʼs pool (cioè., P1
cached in an organismʼs level-one signaling network pool (cioè., 1.0 > P1 > P2) as the most-abundant
genotype at the end of seven evolutionary runs. In one run, the most-abundant final genotype split
=
resources evenly between an organismʼs level-one and level-two signaling network pools (P1
0.5). Finalmente, we observed strategies where resource was primarily, but not entirely, cached in an
= P2
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 3. Progression of same-channel level-one and level-two signaling networks states in an evolutionary run where
level-two resource sharing evolved. Level-one channels are coded by color saturation, and level-two channels are coded
by color hue. A single cell-like organism occupies each grid tile except for black tiles, which are empty. Level-one same-
channel groups appear as uniformly colored clumps, bounded by a white border. Level-two same-channel groups appear
as same-hue amalgamations of level-one groups, bounded by a black border.
124
Artificial Life Volume 25, Numero 2
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
D
N
e
e
H
T
T
UN
e
P
sì
T
o
N
e
G
T
N
UN
D
N
tu
B
UN
–
T
S
o
M
e
H
T
F
o
S
e
tu
l
UN
v
N
UN
e
M
e
H
T
D
N
UN
)
"
S
R
o
T
io
T
e
P
M
o
C
“
(
S
T
N
e
M
io
R
e
P
X
e
N
o
io
T
io
T
e
P
M
o
C
R
o
F
S
D
e
e
S
S
UN
D
e
S
tu
S
e
P
sì
T
o
N
e
G
e
T
UN
R
e
M
tu
N
e
S
T
N
e
M
G
e
S
l
e
B
UN
T
o
w
T
T
S
o
M
T
F
e
l
e
H
T
.
l
1
e
B
UN
T
D
R
UN
D
N
UN
T
S
R
o
F
S
e
tu
l
UN
v
e
P
sì
T
o
N
e
G
N
UN
e
M
–
N
o
io
T
UN
l
tu
P
o
P
e
H
T
e
T
UN
R
e
M
tu
N
e
S
T
N
e
M
G
e
S
l
e
B
UN
T
T
S
o
M
T
H
G
io
R
e
H
T
.
sì
G
e
T
UN
R
T
S
io
G
N
H
C
UN
C
–
e
C
R
tu
o
S
e
R
sì
B
D
e
N
o
io
T
io
T
R
UN
P
H
T
o
B
,
)
io
T
N
UN
N
M
o
D
.
S
N
o
io
T
UN
R
e
N
e
G
R
UN
l
tu
l
l
e
C
D
e
S
P
UN
l
e
l
io
)
sì
e
T
UN
M
X
o
R
P
P
UN
(
D
N
UN
T
N
tu
o
C
e
T
UN
D
P
tu
l
e
T
tu
o
S
B
UN
H
T
o
B
T
UN
D
e
H
C
T
UN
M
,
)
"
N
UN
e
M
.
P
o
P
l
o
R
T
N
o
C
“
(
S
T
N
e
M
T
UN
e
R
T
l
o
R
T
N
o
C
D
N
UN
)
"
"
N
UN
e
M
.
P
o
P
“
N
UN
e
M
“
(
S
N
tu
R
sì
R
UN
N
o
io
T
tu
o
v
e
l
F
o
(
S
l
UN
io
R
T
sì
R
UN
N
o
io
T
tu
o
v
e
l
)
.
D
S
.
±
(
N
UN
e
M
.
P
o
P
l
o
R
T
N
o
C
)
.
D
S
.
±
(
N
UN
e
M
.
P
o
P
)
.
D
S
.
±
(
io
T
N
UN
N
M
o
D
N
UN
e
M
S
R
o
T
io
T
e
P
M
o
C
l
l
UN
l
l
UN
l
l
UN
1
P
≥
2
P
2
P
>
1
P
>
0
1
.
0
1
.
=
1
P
7
7
3
±
6
2
6
6
2
7
6
±
9
6
0
6
9
2
7
±
0
7
6
6
6
4
3
3
±
5
1
3
9
3
4
4
4
3
±
6
4
3
5
3
3
8
1
3
±
1
4
8
0
3
1
P
>
2
P
7
0
5
7
4
1
P
=
2
P
2
5
8
3
3
0
1
.
=
1
P
0
2
9
9
2
.
N
e
G
l
l
e
C
M
3
0
5
M
3
0
5
M
3
3
.
0
5
M
0
2
4
3
M
0
2
7
M
0
2
9
4
3
0
.
±
6
5
0
.
5
3
0
.
±
3
5
0
.
7
3
0
.
±
3
5
0
.
6
4
0
.
±
7
5
0
.
7
4
0
.
±
0
5
0
.
5
3
0
.
±
3
2
0
.
1
0
0
.
±
9
9
0
.
0
0
0
.
±
0
0
1
.
0
0
0
.
±
0
0
1
.
0
0
0
.
±
0
0
1
.
0
0
0
.
±
0
0
1
.
0
0
0
.
±
0
0
1
.
0
0
0
.
±
0
0
0
.
2
0
0
.
±
2
0
0
.
3
0
0
.
±
2
0
0
.
5
0
0
.
±
3
0
0
.
0
0
0
.
±
0
0
0
.
0
0
0
.
±
0
0
0
.
7
3
0
.
±
6
5
0
.
4
2
0
.
±
2
4
0
.
5
2
0
.
±
2
4
0
.
6
1
0
.
±
8
2
0
.
7
0
0
.
±
0
6
0
.
0
0
0
.
±
0
0
1
.
7
3
0
.
±
4
4
0
.
4
2
0
.
±
6
5
0
.
4
2
0
.
±
6
5
0
.
4
1
0
.
±
9
6
0
.
7
0
0
.
±
0
4
0
.
0
0
0
.
±
0
0
0
.
.
7
1
2
±
.
6
8
2
0
6
0
.
±
1
2
3
.
3
6
0
.
±
1
2
3
.
9
6
0
.
±
3
0
3
.
3
3
0
.
±
8
3
3
.
0
6
0
.
±
0
9
3
.
M
0
2
1
9
8
0
.
0
0
1
.
0
0
0
.
0
0
0
.
0
0
1
.
4
0
2
.
M
0
2
1
0
0
0
.
0
0
1
.
0
0
0
.
0
5
0
.
0
5
0
.
5
4
3
.
M
0
2
1
0
0
0
.
0
0
.
l
0
0
0
.
0
0
1
.
0
0
0
.
3
1
3
.
.
5
3
2
1
±
.
0
4
8
4
.
3
5
5
±
.
8
5
9
1
.
1
8
5
±
.
5
1
0
2
.
6
3
7
±
.
6
1
7
2
.
3
5
4
±
.
7
2
9
1
.
1
1
7
±
.
6
0
3
2
.
2
0
9
2
.
6
8
3
2
.
2
3
3
2
.
D
P
U
N
1
UN
2
UN
C
P
1
P
2
P
1
C
2
C
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Artificial Life Volume 25, Numero 2
125
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
)
.
.
D
S
±
(
N
UN
e
M
.
P
o
P
l
o
R
T
N
o
C
)
.
.
D
S
±
(
N
UN
e
M
.
P
o
P
)
.
.
D
S
±
(
io
T
N
UN
N
M
o
D
N
UN
e
M
S
R
o
T
io
T
e
P
M
o
C
)
D
e
tu
N
io
T
N
o
C
(
.
1
l
e
B
UN
T
l
l
UN
l
l
UN
l
l
UN
1
P
≥
2
P
2
P
>
1
P
>
0
1
.
0
1
.
=
1
P
7
7
3
±
6
2
6
6
2
7
6
±
9
6
0
6
9
2
7
±
0
7
6
6
6
4
3
3
±
5
1
3
9
3
4
4
4
3
±
6
4
3
5
3
3
8
1
3
±
1
4
8
0
3
M
3
0
5
M
3
0
5
M
3
3
.
0
5
M
0
2
4
3
M
0
2
7
M
0
2
9
8
0
1
.
±
0
5
1
.
5
0
1
.
±
1
2
1
.
7
0
1
.
±
4
1
1
.
5
7
0
.
±
1
2
0
.
9
5
0
.
±
4
4
0
.
7
3
0
.
±
9
2
0
.
.
2
2
2
±
.
9
8
2
.
9
5
1
±
.
2
9
1
.
2
6
1
±
.
1
8
1
5
0
7
.
±
2
6
4
.
.
0
2
1
±
.
3
1
2
.
7
1
2
±
.
2
7
4
1
P
>
2
P
7
0
5
7
4
M
0
2
1
0
2
4
.
0
8
4
.
1
P
=
2
P
2
5
8
3
3
M
0
2
1
4
1
0
.
.
7
1
1
0
1
.
=
1
P
0
2
9
9
2
M
0
2
1
7
8
0
.
.
4
3
3
.
3
6
6
±
.
3
7
1
3
.
2
5
6
±
.
7
2
0
3
.
5
6
6
±
.
0
3
0
3
.
9
8
6
±
.
4
5
2
3
.
0
7
5
±
.
1
3
8
2
.
3
4
9
±
.
2
1
3
2
.
1
1
2
3
.
4
7
9
3
.
4
1
4
3
3
2
0
.
±
8
1
0
.
1
3
0
.
±
9
3
0
.
2
3
0
.
±
9
3
0
.
5
3
0
.
±
7
6
0
.
1
3
0
.
±
4
7
0
.
1
4
0
.
±
3
3
0
.
3
3
0
.
±
8
4
0
.
5
3
0
.
±
1
5
0
.
7
3
0
.
±
2
5
0
.
8
3
0
.
±
8
6
0
.
6
4
0
.
±
5
6
0
.
1
4
0
.
±
2
5
0
.
6
3
0
.
±
3
5
0
.
2
3
0
.
±
7
4
0
.
3
3
0
.
±
7
4
0
.
2
4
0
.
±
0
5
0
.
7
3
0
.
±
2
5
0
.
9
3
0
.
±
5
4
0
.
4
3
0
.
±
7
4
0
.
4
3
0
.
±
0
4
0
.
6
3
0
.
±
9
3
0
.
2
4
0
.
±
7
4
0
.
0
4
0
.
±
5
5
0
.
8
3
0
.
±
5
6
0
.
4
3
0
.
±
5
5
0
.
4
3
0
.
±
6
4
0
.
4
3
0
.
±
7
4
0
.
9
3
0
.
±
5
4
0
.
9
3
0
.
±
5
3
0
.
3
4
0
.
±
1
5
0
.
6
6
0
.
0
4
0
.
0
0
1
.
0
0
1
.
6
4
0
.
0
0
1
.
0
0
1
.
4
4
0
.
0
0
1
.
1
0
0
.
1
1
0
.
0
0
0
.
0
0
0
.
0
0
0
.
0
0
0
.
.
N
e
G
l
l
e
C
.
D
P
U
N
C
E
1
E
2
E
C
M
1
M
2
M
1
S
2
S
126
Artificial Life Volume 25, Numero 2
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
organismʼs level-two signaling network pool (cioè., 1.0 > P2 > P1) as the most-abundant genotype
at the end of 33 evolutionary runs.
We suspect that a tradeoff between growth rate and long-term stability prompted the universal
allocation of at least some resource to level-one pools and/or cell stockpiles. Cell and level-one
resource caching might function something like saving for a rainy day. Because reproduction over
level-two channel mates was universally avoided, cells and level-one same-channel networks situated
in the interior of a larger level-two same-channel network do not expend their resource pools unless
that larger level-two same-channel network is damaged, exposing them to directly adjacent cells of a
different level-two channel. Così, resource accumulates in cell stockpiles and level-one pools until
the level-two same-channel network comes under stress. Split allocation might also represent
hedging against defection of a second-level channel mate via somatic mutation.
Infatti, we did observe selection for apoptosis in the 41 replicates where the dominant genotype
employed second-level resource caching. In these replicates, the average population-mean value
of Mc was 0.68 with standard deviation 0.33, significantly greater than the value Mc = 0.5 we would
expect in the absence of a selective pressure on apoptosis response to mutation ( P < 0.001,
bootstrap test).
To assess whether heavy second-level resource allocators, which we characterize as higher-level
individuals, were more likely to employ apoptosis to mitigate somatic mutation, we examined the
relationship between first- and second-level resource pooling and cellular apoptosis at the conclusion
of our 50 replicate evolutionary trials. We observed a significant negative correlation between
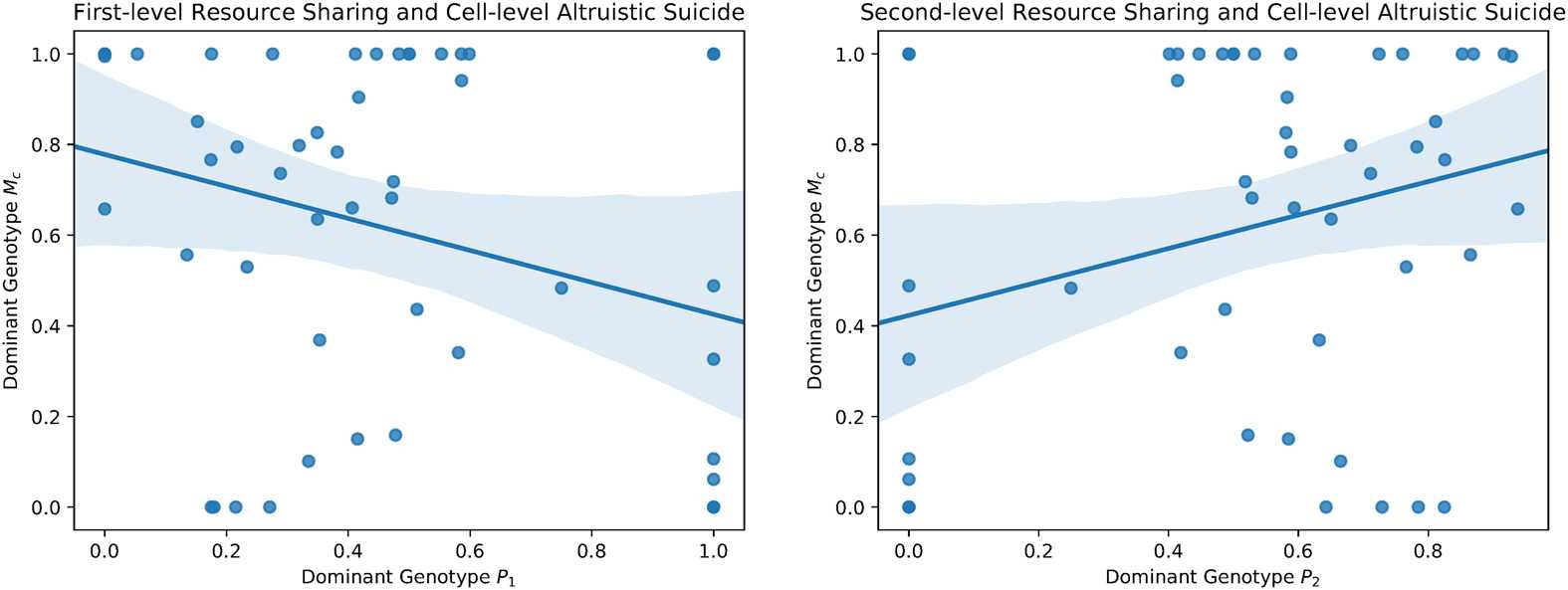
dominant genotype P1 and Mc ( p < 0.05; bootstrap test; Figure 4(a)) and a significant positive cor-
relation between dominant genotype P2 and Mc ( p < 0.05; bootstrap test; Figure 4(b)). This result
suggests that second-level individuals, in particular, relied on apoptosis to mitigate somatic mutation.
We also assessed whether higher-level individuals provided larger resource endowments to their
second-level propagules (offspring sharing neither the level-one nor the level-two channel ID with
the parent). We examined the relationship between first- and second-level resource pooling and
dominant genotype second-level propagule endowment at the conclusion of our 50 replicate evo-
lutionary trials. We observed a significant negative correlation between dominant genotype P1 and
E2 (p < 0.05; bootstrap test) and a significant positive correlation between dominant genotype P2
and E2 (p < 0.05; bootstrap test). Second-level individuals might provide larger endowments to
propagules simply because of a greater capacity to collect resource or perhaps because of stronger
selection for well-endowed offspring when competing against other second-level individuals.
Figure 4. Plots of dominant resource caching strategies and dominant apoptosis strategies. A bootstrapped 95% confi-
dence interval for the fit is shaded. Both correlations are statistically significant (p < 0.05; bootstrap test).
Artificial Life Volume 25, Number 2
127
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
a
r
t
l
/
_
a
_
0
0
2
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
M. A. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
This result prompts the reverse question: Do lower-level individuals provide larger resource
endowments to first-level propagules (offspring that do not share level-one channel ID with the
parent but may or may not share level-two channel ID with the parent)? Indeed, we observed a
significant positive correlation between first-level resource sharing and first-level endowment
( p < 0.0001; bootstrap test) and a significant negative correlation between second-level resource
sharing and first-level endowment ( p < 0.0001; bootstrap test). Cells that pool resource with their
smaller level-one same-channel group tend to invest more heavily in the direct offshoots of their
level-one same-channel group than cells that pool resource with their larger level-two same-channel
group. This observation suggests that, although cells do not directly displace their level-one channel
mates, competitive dynamics between them may be at play.
3.2 Competition Experiments
Next, we wanted to compare first-, second-, and split-level allocators to determine which genotype
was the most fit. We ran competition experiments between dominant genotypes from evolutionary
runs representative of each of these strategies. To prevent further evolution, we disabled mutation
for these experiments. To represent first-level allocators, we selected randomly from the nine pure
first-level allocator dominant genotypes we observed. To represent the split-level allocators, we
selected the single dominant genotype where resource was partitioned exactly evenly between first-
and second-level channel pools. To represent second-level allocators, we selected the dominant
genotype with the largest second-level allocation proportion. Table 1 enumerates the three represen-
tative genotypes used. Figure 5 shows a time series of signaling network snapshots in a competition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
a
r
t
l
/
_
a
_
0
0
2
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Progression of same-channel level-one and level-two signaling network states in a competition run. We
seeded the grid with three copies of each of three champion genotypes from evolutionary trials. Then, with mutation
disabled to prevent further evolution, the genotypes competed. Level-one channels are coded by color saturation, and
level-two channels are coded by color hue. A single cell-like organism occupies each grid tile except for black tiles, which
are empty. Level-one same-channel groups appear as uniformly colored clumps, bounded by a white border. Level-two
same-channel groups appear as same-hue amalgamations of level-one groups, bounded by a black border.
128
Artificial Life Volume 25, Number 2
M. A. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
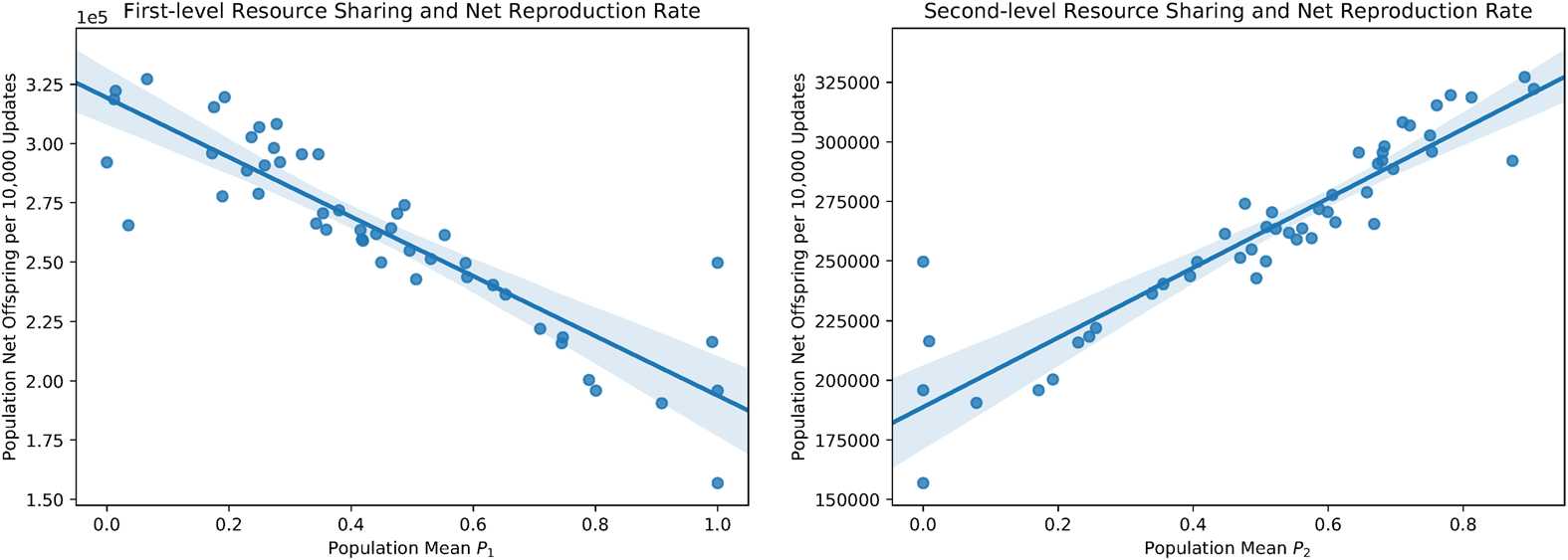
Figure 6. Mean resource caching strategies and net reproduction rate across populations. A bootstrapped 95% confi-
dence interval for the fit is shaded. Both correlations are statistically significant (p < 0.0001; bootstrap test).
experiment run. Colonies of each genotype can be seen to grow from each seed and then clash,
ultimately yielding a population dominated by second-level allocators.
Indeed, the second-level resource-caching strategy became most-abundant in all 50 trials. Across
the 50 replicates, at update 1.5 million (cellular generation 3489 with standard deviation 40) the
second-level resource-caching strategy constituted 90.2%, with standard deviation 3.8%, of the com-
peting population of cells. In the absence of mutation, second-level allocators tend to exhibit greater
fitness than split- and first-level allocators ( p < 0.0001; two-tailed exact test).
In competition experiments, however, higher-level individuals likely benefited from elimination
of somatic mutation. To assess the relative fitness of first- and second-level individuals without
mutation disabled, we examined the relationship between first- and second-level resource pooling
and the rate of cellular reproduction at the end of each of the 50 replicate evolutionary trials
performed. We observed a significant negative correlation between mean P1 and cellular reproduc-
tion rate ( p < 0.0001; bootstrap test; Figure 6a) and a significant positive correlation between mean
P2 and cellular reproduction rate ( p < 0.0001; bootstrap test; Figure 6b). This result suggests
that second-level allocators tend to collect resource more effectively than split- and first-level
allocators.
3.3 Control Evolutionary Experiments
Under control conditions where resource was distributed evenly to all cells regardless of same-
channel group configuration, split-level resource caching also evolved. Split-level allocation was
the most common strategy at update 3 million in all replicates. Strategies where resource was pri-
marily, but not entirely, cached in an organismʼs level-one signaling network pool (i.e., 1.0 > P1 > P2)
were most-abundant at the end of 33 evolutionary runs, and strategies where resource was primarily,
but not entirely, cached in an organismʼs level-two signaling network pool (cioè., 1.0 > P2 > P1) were
most-abundant at the end of 17 evolutionary runs. As shown in Table 1, the average population
mean of P1 is greater in the control treatment than in the standard treatment at time points matched
by absolute elapsed update count and approximate elapsed cellular generations, but this difference is
not statistically significant.
Consistent with the standard treatment, we observed strong selection against direct reproductive
competition between channel mates at update 3 million in the control treatment. Nearly all most-
= 1.0),
common genotypes completely avoided reproducing over level-two channel mates (cioè., A2
except for a single most-common genotype where a very slim probability of reproducing over
level-two channel mates was allowed (A2
= 0.996).
Artificial Life Volume 25, Numero 2
129
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
UN
R
T
l
/
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
UN
R
T
l
/
_
UN
_
0
0
2
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
M. UN. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
The emergence of resource-sharing and competition avoidance under control conditions suggests
kin recognition alone can prompt some aspects of higher-level individuality. Tuttavia, we observed
selection against the apoptosis response to mutation, Mc, under control conditions. Across 50 replicates
of the control treatment, the average population mean of Mc was 0.18 with standard deviation 0.23—
significantly less than the value Mc = 0.5 expected without selective pressure against apoptosis
response to mutation ( P < 0.0001, two-tailed t-test). Indeed, the population-mean Mc for control
runs was also significantly reduced compared to the standard treatment at time points matched by
absolute elapsed update count ( p < 0.001; two-tailed t-test) and by approximate elapsed cellular
generations ( p < 0.001; two-tailed t-test). Perhaps, under control conditions, the apoptosis response
to mutation is disfavored because kin groups stand to lose less from mutant members (i.e., the
resource penalty for excessive same-channel network expansion is absent). It appears that, at least
in our system, kin recognition alone does not suffice to prompt full-fledged fraternal transitions in
individuality.
In the absence of resource penalties for erroneous activation under control conditions, we also
observed the evolution of larger same-channel groups. At update 3 million, the most-common geno-
types encoded a level-two same-channel cap C2 of 484.0 cells with standard deviation of 123.5.
Compared to the standard treatment, control runs exhibited larger mean level-two same-channel
caps C2 at time points matched by absolute elapsed update count ( p < 0.0001; two-tailed t-test)
and approximate elapsed cellular generations ( p < 0.0001; two-tailed t-test). Even at 20 million
updates, when evolution had elapsed around six times as many cellular generations in the standard
treatment as in the control treatment at update 3 million, the mean level-two same-channel cap C2
reached only 262.9 with standard deviation 72.2 under the standard treatment. This is significantly
smaller than mean C2 under the control treatment at update 3 million ( p < 0.0001; two-
tailed t-test). Figure 7 depicts the comparatively large same-channel level-two groups present at the
end of a control run. Table 1 summarizes the most-common genotypes observed under the control
treatment.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
a
r
t
l
/
_
a
_
0
0
2
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. End state (update 3000000, cell gen. 6916) of same-channel signaling networks evolved under the control
treatment. Level-one channels are coded by color saturation and level-two channels are coded by color hue. A single
cell-like organism occupies each grid tile except for black tiles, which are empty. Level-one same-channel groups appear
as uniformly-colored clumps, bounded by a white border. Level-two same-channel groups appear as same-hue amalgam-
ations of level-one groups, bounded by a black border.
130
Artificial Life Volume 25, Number 2
M. A. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
4 Conclusion
Using simple organisms that evolve parameters for a set of manually designed strategies, we have
demonstrated that DISHTINY selects for genotypes that exhibit high-level individuality. We
observed a spectrum of first- and second-level individuality among evolutionary outcomes. Specif-
ically, we observed
1.
2.
3.
reproductive division of labor among members of the same channel (i.e., individuals
enveloped in a same-channel signaling network ceded reproduction to those at the
periphery),
cooperation between members of the same channel (i.e., pooling of resource on same-
channel signaling networks),
reproductive bottlenecking (i.e., groups of cells sharing a channel ID descend from a single
originator of that channel ID), and
4.
suppression of somatic mutation via apoptosis coincident with second-level individuality.
Competition experiments revealed that second-level individuals usually outcompete lower-level
individuals. The magnitude of resource endowment for propagules was also correlated with second-
level individuality.
Although shifts in individuality to level-one and level-two signaling networks were both observed,
the question of whether these transitions were truly hierarchical in nature is debatable. That is, it is
not clear whether level-one individuality was to some extent preserved in or necessary for the emer-
gence of level-two individuality. Given the nature of the manually designed strategies for resource
pooling and reproductive division of labor, level-two resource pooling and division of labor could
readily leapfrog over level-one resource pooling and division of labor and, in many ways, seemed to
completely supersede those level-one efforts.
We believe that this is a shortcoming of the manual design of behaviors for which simple cell-like
organisms evolved parameters, not of the DISHTINY platform itself. We have nevertheless
demonstrated that DISHTINY ultimately selects for high-level individuality. We are eager to work
with more sophisticated cell-like organisms capable of arbitrary computation via genetic program-
ming in order to pursue more open-ended evolutionary experiments. We will also test the implica-
tions of relaxing current arbitrary restrictions that artificially promote transitions, such as the
hierarchical nesting of same-channel signaling networks and the explicitly defined signaling networks
themselves, leaving these details to evolution to figure out. Further work will provide valuable
insight into scientific questions relating to major evolutionary transitions such as the role of
preexisting phenotypic plasticity [6, 12], preexisting environmental interactions, preexisting repro-
ductive division of labor, and how transitions relate to increases in organizational [8], structural,
and functional [10] complexity.
We believe that such an approach also provides a unique opportunity to fundamentally advance
artificial life with respect to open-ended evolution. Fundamental to this goal is scale. The
DISHTINY platform potentially scales to select for an arbitrary number of hierarchical levels of
individuality, not just the two hierarchical levels explored in these experiments. For example, a third
separate, but overlaid, resource-wave–channel-signaling layer could incentivize level-two individuals
to unite into level-three individuals and so on and so forth. Importantly, the platform is implemented
in a decentralized manner and can comfortably scale as additional computing resources are provided.
Parallel computing is widely exploited in evolutionary computing, where subpopulations are farmed
out for periods of isolated evolution or single genotypes are farmed out for fitness evaluation
[13, 18]. DISHTINY presents a more fundamental parallelization potential: principled parallelization
of the evolving individual phenotype at arbitrary scale (i.e., a high-level individual as a large collection
of individual cells on the toroidal grid). Such parallelization will be key to realizing evolving com-
putational systems with scale—and, perhaps, complexity—approaching biological systems.
Artificial Life Volume 25, Number 2
131
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
a
r
t
l
/
_
a
_
0
0
2
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
M. A. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
Acknowledgments
Thanks to members of the DEVOLAB, in particular Michael J. Wiser, for feedback on statistical
methods, and to Heather Goldsby for concept and editing feedback. This research was supported in
part by NSF grants DEB-1655715 and DBI-0939454, and by Michigan State University through the
computational resources provided by the Institute for Cyber-Enabled Research. This material is
based upon work supported by the National Science Foundation Graduate Research Fellowship
under Grant No. DGE-1424871.
References
1. Ackley, D. H. (2016). Indefinite scalability for living computation. In D. Schuurmans & M. Wellman
(Eds.), Proceedings of the Thirtieth Association for the Advancement of Artificial Intelligence Conference on Artificial
Intelligence (pp. 4142–4146). Palo Alto, CA: AAAI Press.
2. Ackley, D. H., & Cannon, D. C. (2011). Pursue robust indefinite scalability. In M. Welsh (Ed.), Proceedings of
the 13th USENIX Conference on Hot Topics in Operating Systems (pp. 8–13). Berkeley, CA: USENIX
Association.
3. Banzhaf, W., Baumgaertner, B., Beslon, G., Doursat, R., Foster, J. A., McMullin, B., De Melo, V. V.,
Miconi, T., Spector, L., Stepney, S., & White, R. (2016). Defining and simulating open-ended novelty:
Requirements, guidelines, and challenges. Theory in Biosciences, 135(3), 131–161.
4. Bedau, M. A. (2003). Artificial life: Organization, adaptation and complexity from the bottom up. Trends in
Cognitive Sciences, 7(11), 505–512.
5. Bouchard, F. (2013). What is a symbiotic superindividual and how do you measure its fitness. In
F. Bouchard & P. Huneman (Eds.), From groups to individuals: Evolution and emerging individuality
(pp. 243–264). Cambridge, MA: MIT Press.
6. Clune, J., Ofria, C., & Pennock, R. T. (2007). Investigating the emergence of phenotypic plasticity in
evolving digital organisms. In F. A. e Costa, L. M. Rocha, E. Costa, I. Harvey, & A. Coutinho (Eds.),
Proceedings of the 9th European Conference on Advances in Artificial Life (pp. 74–83). Berlin, Heidelberg:
Springer-Verlag.
7. Ereshefsky, M., & Pedroso, M. (2015). Rethinking evolutionary individuality. Proceedings of the National
Academy of Sciences of the U.S.A., 112(33), 10126–10132.
8. Goldsby, H. J., Dornhaus, A., Kerr, B., & Ofria, C. (2012). Task-switching costs promote the evolution
of division of labor and shifts in individuality. Proceedings of the National Academy of Sciences of the U.S.A.,
109(34), 13686–13691.
9. Goldsby, H. J., Knoester, D. B., & Ofria, C. (2010). Evolution of division of labor in genetically
homogenous groups. In M. Pelikan & J. Branke (Eds.), Proceedings of the 12th Annual Conference on Genetic and
Evolutionary Computation (pp. 135–142). New York: ACM.
10. Goldsby, H. J., Knoester, D. B., Ofria, C., & Kerr, B. (2014). The evolutionary origin of somatic cells under
the dirty work hypothesis. PLOS Biology, 12(5), e1001858.
11. Goldsby, H. J., Young, R. L., Hofmann, H. A., & Hintze, A. (2017). Increasing the complexity of solutions
produced by an evolutionary developmental system. In M. Pelikan & J. Branke (Eds.), Proceedings of the
Genetic and Evolutionary Computation Conference Companion (pp. 57–58). New York: ACM.
12. Lalejini, A., & Ofria, C. (2016). The evolutionary origins of phenotypic plasticity. In C. Gershenson,
T. Froese, J. M. Siqueiros, W. Aguilar, E. Izquierdo, & H. Sayama (Eds.), Proceedings of the Artificial Life
Conference 2016 (pp. 372–379). Cambridge, MA: MIT Press.
13. Lin, S. C., Punch, W. F., & Goodman, E. D. (1994). Coarse-grain parallel genetic algorithms:
Categorization and new approach. In Proceedings of the Sixth Institute of Electrical and Electronics Engineers
Symposium on Parallel and Distributed Processing (pp. 28–37). New York: IEEE.
14. Maynard Smith, J., & Szathmáry, E. (1997). The major transitions in evolution. New York: Oxford University
Press.
15. Queller, D. C. (1997). Cooperators since life began. The Quarterly Review of Biology, 72(2), 184–188.
16. Ratcliff, W. C., Denison, R. F., Borrello, M., & Travisano, M. (2012). Experimental evolution of
multicellularity. Proceedings of the National Academy of Sciences of the U.S.A., 109(5), 1595–1600.
132
Artificial Life Volume 25, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
a
r
t
l
/
_
a
_
0
0
2
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
M. A. Moreno and C. Ofria
Toward Open-Ended Fraternal Transitions in Individuality
17. Ray, T. S., & Thearling, K. (1996). Evolving parallel computation. Complex Systems, 10(3), 229–237.
18. Real, E., Moore, S., Selle, A., Saxena, S., Suematsu, Y. L., Tan, J., Le, Q. V., & Kurakin, A. (2017). Large-
scale evolution of image classifiers. In D. Precup & Y. W. Teh (Eds.), Proceedings of the 34th International
Conference on Machine Learning (pp. 2902–2911). Sydney, Australia: PMLR.
19. Taylor, T., Bedau, M., Channon, A., Ackley, D., Banzhaf, W., Beslon, G., Dolson, E., Froese, T.,
Hickinbotham, S., Ikegami, T., et al. (2016). Open-ended evolution: Perspectives from the OEE workshop
in York. Artificial Life, 22(3), 408–423.
20. West, S. A., Fisher, R. M., Gardner, A., & Kiers, E. T. (2015). Major evolutionary transitions in
individuality. Proceedings of the National Academy of Sciences of the U.S.A, 112(33), 10112–10119.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
5
2
1
1
7
1
8
9
6
7
0
0
a
r
t
l
/
_
a
_
0
0
2
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Artificial Life Volume 25, Number 2
133