Theta-band Oscillations in the Middle Temporal Gyrus
Reflect Novel Word Consolidation
Iske Bakker-Marshall1, Atsuko Takashima1,2, Jan-Mathijs Schoffelen1, Janet G. van Hell3,
Gabriele Janzen1, and James M. McQueen1,2
Astratto
■ Like many other types of memory formation, novel word
learning benefits from an offline consolidation period after the
initial encoding phase. A previous EEG study has shown that re-
trieval of novel words elicited more word-like-induced electro-
physiological brain activity in the theta band after consolidation
[Bakker, I., Takashima, A., van Hell, J. G., Janzen, G., & McQueen,
J. M. Changes in theta and beta oscillations as signatures of novel
word consolidation. Journal of Cognitive Neuroscience, 27,
1286–1297, 2015]. This suggests that theta-band oscillations play
a role in lexicalization, but it has not been demonstrated that this
effect is directly caused by the formation of lexical representa-
zioni. This study used magnetoencephalography to localize the
theta consolidation effect to the left posterior middle temporal
gyrus (pMTG), a region known to be involved in lexical storage.
Both untrained novel words and words learned immediately be-
fore test elicited lower theta power during retrieval than existing
words in this region. After a 24-hr consolidation period, the differ-
ence between novel and existing words decreased significantly,
most strongly in the left pMTG. The magnitude of the decrease
after consolidation correlated with an increase in behavioral
competition effects between novel words and existing words with
similar spelling, reflecting functional integration into the mental
lexicon. These results thus provide new evidence that consolida-
tion aids the development of lexical representations mediated by
the left pMTG. Theta synchronization may enable lexical access by
facilitating the simultaneous activation of distributed semantic,
phonological, and orthographic representations that are bound
together in the pMTG. ■
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
Novel word learning, like other types of memory encod-
ing, benefits from an offline consolidation period after
exposure. For instance, behavioral work has shown that
only after a delay of at least several hours do novel words
acquire the ability to enter into lexical competition with
phonologically and orthographically similar existing words
during speech processing (Bakker, Takashima, van Hell,
Janzen, & McQueen, 2014; Gaskell & Dumay, 2003; Dumay
& Gaskell, 2007, 2012) and visual processing (Bakker et al.,
2014). Allo stesso modo, novel words start priming visually pre-
sented, semantically related existing words after a delay
(van der Ven, Takashima, Segers, & Verhoeven, 2015;
Tamminen & Gaskell, 2013).
In line with neurocognitive complementary learning
systems (CLS) models of memory consolidation (Frankland
& Bontempi, 2005; McClelland, McNaughton, & O’Reilly,
1995; Squire & Alvarez, 1995; Marr, 1970), it has been
argued that novel words are initially encoded as episodic
memories by a fast-learning hippocampal mechanism and
only gradually become integrated into the neocortical
lexicon (Davis & Gaskell, 2009). This offline consolidation
1Radboud University Nijmegen, 2Max Planck Institute for Psy-
cholinguistics, Nijmegen, 3Pennsylvania State University
© 2018 Istituto di Tecnologia del Massachussetts
process transforms isolated, episodic memories into stable,
lexical representations that interact with other words dur-
ing language use. These representations are no longer
modality specific, as indicated by the finding that visually
acquired novel words interact with existing words in audi-
tory tasks, and vice versa (Bakker et al., 2014). The CLS
account of word learning thus predicts a qualitative change
in the neural representation of novel words, with consoli-
dation leading to increasingly word-like retrieval processes.
In previous work (Bakker, Takashima, van Hell, Janzen,
& McQueen, 2015), we tested this hypothesis in the
time–frequency domain using EEG and demonstrated that
consolidated novel words indeed elicited more word-
like oscillatory brain responses than recently learned
words in the theta band (4–8 Hz) over left-hemisphere
sensors. Given that scalp-recorded EEG is not optimized
for accurate estimation of distributed neural sources, IL
question that arises is what the neural substrate of this
theta effect is. If consolidation facilitates the formation
of lexical representations, enhanced retrieval activity for
consolidated novel words should be observed in a rela-
tively focal network of left-lateralized perisylvian regions
known to be involved in lexical processing, as will be dis-
cussed below. Tuttavia, the data leave open the possibility
that theta synchronization reflects orthogonal processes,
Per esempio, those related to episodic retrieval. The current
Journal of Cognitive Neuroscience 30:5, pag. 621–633
doi:10.1162/jocn_a_01240
study sought to address this issue by using magneto-
encephalography (MEG) source localization techniques to
identify the sources of the oscillatory signatures of lexical
consolidation.
In our previous EEG study (Bakker et al., 2015), IL
pattern of theta power modulations induced by the visual
presentation of a word was taken as a measure of lexical
activation. A larger theta power increase versus a pre-
stimulus baseline is typically observed in response to words
compared with pseudowords, both auditorily (Krause et al.,
2006) and visually (Marinkovic, Rosen, Cox, & Kovacevic,
2012). Semantically rich words also elicit larger theta power
increases compared with function words (Bastiaansen,
van der Linden, ter Keurs, Dijkstra, & Hagoort, 2005). IL
topography of this power increase is further sensitive to
word meaning, possibly reflecting the somatotopic organi-
zation of semantic information (Bastiaansen, Oostenveld,
Jensen, & Hagoort, 2008). These findings suggest that
theta synchronization plays a role in lexical retrieval,
which in turn implies that consolidated novel words
should exhibit more word-like theta responses than re-
cently learned words.
To test this prediction, participants in the Bakker et al.
(2015) study were trained on two sets of novel and exist-
ing words paired with definitions, one set on each of
2 consecutive days. After the second training session,
EEG responses were recorded as participants made se-
mantic decisions to words from the two trained sets or
a set of untrained novel and existing words. It was there-
fore possible to contrast responses to completely novel
parole (the “untrained” condition), novel words learned
only before test (the “recent” condition), and novel
words that had an opportunity for offline consolidation
as they had been learned 1 day before testing (the “re-
mote” condition) against existing words with the same
level of training and exposure. In line with previous find-
ing, a lexicality effect was observed, as reflected by a
larger theta power increase over left-hemisphere tempo-
ral sensors in response to untrained existing words than
to untrained novel words (cioè., pseudowords). This lexi-
cality effect was smaller in the recent condition but was
no longer present for words trained a day earlier (the re-
mote condition). This suggests that the retrieval process
became more word-like with consolidation. We speculated
that this pattern may reflect the gradual formation of a
lexical representation.
The question that arises is where the neural substrate of
the theta-related lexicalization process might be located.
A likely candidate is the left posterior middle temporal
gyrus (pMTG), which many current models of word pro-
cessing view as a lexical “hub” that mediates the mapping
of word forms onto distributed semantic information
(Gow, 2012; Lau, Phillips, & Poeppel, 2008; Hickok &
Poeppel, 2004, 2007). The role of the left pMTG in lexical
processing is supported by fMRI evidence showing an
enhanced BOLD response to words relative to pseudo-
parole (Prabhakaran, Blumstein, Myers, Hutchison, & Britton,
2006) as well as by semantic priming effects in both fMRI
and EEG (see Lau et al., 2008, for a review). Damage to
the left pMTG typically results in word finding problems
combined with spared perceptual and conceptual abili-
ties, indicating specific involvement in lexical access (Vedere
Gow, 2012, for a review). Inoltre, Marinkovic et al.
(2012) estimated the source of the theta power difference
between visually presented words and pseudowords in
their MEG data to be in the left temporal cortex (although
this source extended into the left inferior frontal lobe).
Previous fMRI work on word learning has accordingly
demonstrated an increased BOLD signal in left pMTG
involvement after consolidation (Takashima, Bakker,
van Hell, Janzen, & McQueen, 2014). In this study, par-
ticipants learned a set of novel spoken words and per-
formed a recognition task immediately after learning as
well as 24 hr later. After the consolidation period, left
pMTG activation in response to correctly recognized novel
words increased relative to the response observed in the
immediate scanning session. Postconsolidation func-
tional connectivity between the auditory cortex and this
MTG region was further enhanced for those participants
who exhibited stronger behavioral evidence of lexical
integration. These data, as well as those of Takashima,
Bakker, van Hell, Janzen, and McQueen (2017), who exam-
ined consolidation of newly learned words after a 1-week
delay, are in line with the hypothesis that lexical storage
in the left pMTG develops during offline consolidation.
The current study was designed to test the hypothesis
that the consolidation effect we previously observed in
the theta band is driven by an increase in left pMTG in-
volvement, which would support the CLS claim that con-
solidation of novel words facilitates the lexicalization of
their representations. As in Bakker et al. (2015), partici-
pants were trained on written novel and existing words
SU 2 consecutive days, after which MEG responses to
the learned words plus an untrained set were recorded
during a semantic decision task. Behavioral measures of
lexical competition and semantic priming were obtained
to assess the functional integration of novel words into the
existing lexicon. The difference in theta power elicited
by novel versus existing words was computed at each
level of training. We then used a beamformer approach
(Gross et al., 2001) to identify the most likely neural
generators of these lexicality effects and asked whether
the change between recent and remote words could be
localized to the left pMTG. Because we previously ob-
served additional, less robust consolidation-dependent
changes in beta (16–21 Hz) power (Bakker et al., 2015), lex-
icality effects in this frequency band were also investigated.
METHODS
Participants
Twenty-nine right-handed (as assessed by an abridged
version of the Edinburgh Handedness Inventory;
622
Journal of Cognitive Neuroscience
Volume 30, Numero 5
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Oldfield, 1971) native speakers of Dutch (eight men),
aged 18–35 years (mean = 23 years), participated in
the experiment in return for course credit or monetary
compensation. Participants had no history of neuro-
logical or language-related disorders and reported hav-
ing normal or corrected-to-normal vision and hearing.
One male participant and one female participant were
removed from the MEG analyses because of excessive
movement, and one female participant was removed
because of large eye-movement-related artifacts. One
female participant was removed from all analyses be-
cause of experimenter error.
Design
Participants were trained on two different sets of novel
and existing words, one set on each of 2 consecutive days
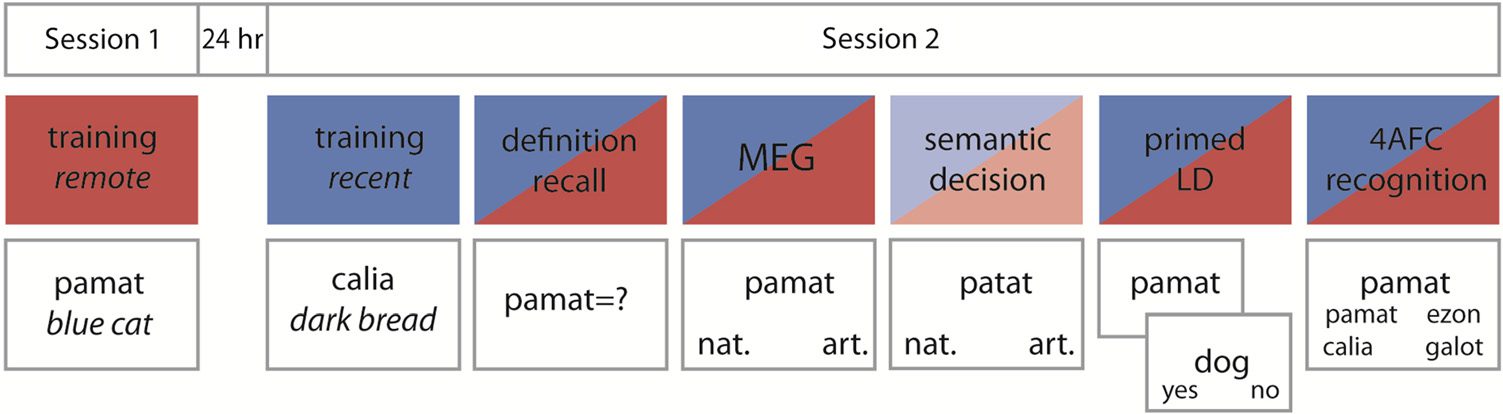
(Guarda la figura 1 for an overview of tasks). Immediately after
the second training session, participants performed a
semantic decision task in the MEG scanner. This task
contained the trained novel and existing words from
the first session (remote condition), the trained novel
and existing words from the second session (recent con-
dizione), and a set of novel and existing words that were
not part of the trained set (untrained condition). Questo
design allowed us to compare the effect of consolidation
on the difference between novel and existing words
within a single recording session, using the existing words
as a baseline for each novel condition to control for pro-
cesses related to episodic rather than lexical retrieval.
Finalmente, two behavioral tasks measured lexical competition
between novel and existing words (semantic decision on
the base words) and semantic priming from novel to exist-
ing words (primed lexical decision). All materials were
presented in the visual modality only.
Materials
The materials largely overlapped with those used by
Bakker et al. (2015), but for the purpose of the semantic
decision task on the base words (see Procedure section),
some items were adapted such that the base words of
half of the novel words in each list referred to natural
objects and half referred to artifacts. Four lists of 20 novel
parole (Vedi la tabella 1) of four to seven letters (mean = 5.2
letters) were derived from Dutch words by substituting
one letter, Per esempio, “pamat” from “patat” (chips). Base
words had no or few orthographic neighbors (mean = 2.7)
and had a frequency of 1–112 per million (mean = 12.7 per
million) in the CELEX database (Baayen, Piepenbrock, &
Gulikers, 1995). The substituted letter was in the first posi-
tion in 17 parole, between the second and penultimate
positions in 45 parole, and in the last position in 18 parole.
The four lists were matched on number of neighbors,
word length, and frequency of the base words.
Two lists of 20 definitions were created to provide the
novel words’ meanings, in part based on Tamminen and
Gaskell (2013) and largely identical to those used by
Bakker et al. (2015). Each definition consisted of an
existing object category paired with two distinguishing
caratteristiche, Per esempio, “A cat that has stripes and is bluish
gray,” and thus described a novel subcategory of an exist-
ing concept.
For each participant, two of the four lists of novel
words and both lists of definitions served as the to-be-
learned material, one in each of two learning sessions
(recent and remote). The pairing of novel words and def-
initions was randomized for each participant. The third
list of novel words was used as the untrained condition
in the MEG task, and the base words of the fourth list
served as the untrained condition in the semantic deci-
sion task. The pairing of lists and tasks/conditions was
rotated across participants.
Three lists of 20 existing Dutch words (Vedi la tabella 2) Di
four to eight letters (mean = 5.8 letters) with a frequency
of 1–195 per million (mean = 32.7 per million) were
created and matched on frequency and length. Each
existing word was presented with a realistic definition
(per esempio., “lemon: a yellow, sour-tasting fruit”). Each partici-
pant saw two of the three lists of existing words and defi-
nitions, one in each of the two learning sessions. The third
list served as the untrained existing condition in the MEG
task.
For the purpose of the behavioral primed lexical decision
task, three semantically related existing Dutch target words
were selected for each novel word meaning’s category label
Figura 1. Schematic overview
of tasks. The bottom row
gives an abbreviated example
of a trial. Participants learned a
set of novel and existing words
in Session 1 (remote) and a
second set of novel and existing
parole 24 hr later in Session 2
(recente). Tests in Session 2
included both the recent
(indicated by blue) and remote
(indicated by red) sets as well
as a set of untrained (novel and existing) words in the MEG task. The semantic decision task was performed on the base words from which the
novel words were derived (indicated by lighter colors). LD = lexical decision; nat. = natural; art. = artifact.
Bakker-Marshall et al.
623
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 1. Novel Word Stimuli and Their Existing Dutch
Base Words
Novel
Base
Translation Novel
Base
Translation
List 1
celmo
cello
ablas
atlas
cello
atlas
List 2
eglo
iglo
radak
radar
igloo
radar
hile
file
traffic jam waraf
karaf
carafe
alcum album album
abulet
amulet
amulet
dokane douane customs
indet
index
index
alara
alarm alarm
asaalt
asfalt
asfalt
mobot
robot
robot
saltris
salaris
salary
trefe
niald
trede
step
naald
needle
catera
camera camera
meibel meubel
item of
mobilia
tadi
taxi
taxi
fleb
fles
bottle
nobra
cobra
cobra
lepia
lepra
lepra
frec
uzer
fret
uier
ferret
udder
prema
poema
puma
sprug
spuug
spit
porin
porie
pore
relma
reuma
rheumatism
lagine
lawine
avalanche
astia
astma
asthma
halik
havik
hawk
puzil
pupil
pupil
vidus
virus
virus
stelet
skelet
skeleton
ziloer
zilver
silver
ramijn
ravijn
ravine
alimaat klimaat climate
okrel
oksel
armpit
ratuur
natuur nature
gole
golf
wave
List 3
List 4
tosto
tosti
toast
bumier bumper bumper
pamat
patat
chips
kiosa
kiosk
kiosk
kohma
komma comma
merro metro metro
assel
asiel
asylum/
mossee moskee mosque
shelter
keno
fiehe
jenu
kano
fiche
canoo
chip
palaar
pilaar
pillar
pialm
psalm psalm
menu menu
oriel
orgel
organ
ofera
opera
opera
pontein fontein fountain
hemb
hemd
vest
perzon perron
platform
teno
tent
tent
kantoog kantoor office
elane
eland moose
ezon
calia
cavia
guinea pig
fnoe
ozon
gnoe
ozone
gnu
ananak ananas pineapple
inoor
ivoor
ivory
elster
ekster magpie
lamboe bamboe bamboo
Tavolo 1. (continued )
Novel
Base
Translation Novel
Base
Translation
gjord
arwt
fjord
erwt
fiord
pea
galot
galop
gallop
perdik
perzik
peach
taroe
tarwe
wheat
nund
rund
cow
vitroen citroen lemon
moerak moeras
swamp
maverie materie matter
ozel
ezel
donkey
winc
vento
vento
riviet
rivier
river
(per esempio., DOG for “pamat,” if the given definition was a type
of cat). As much as possible, targets were taken from a
Dutch database of word associations (De Deyne &
Storms, 2008) O, in case the prime word was unavailable
in that database, from the Florida Free Association Norms
(Nelson, McEvoy, & Schreiber, 1998). Target words were
composed of 3–10 letters (mean = 5.2 letters), with a fre-
quency of 1–1084 per million (mean = 71.7 per million).
The two lists of novel word meanings were matched for
target length and frequency. No target words occurred
in any of the definitions or as a base word of one of the
novel words. Three unrelated prime–target pairs were
created for each meaning by shuffling the list of target
parole.
Procedure
Training and Memory Tests
The training and memory test procedures were identical
to Bakker et al. (2015). Briefly, training consisted of a
round of exposure to each word–definition pair, followed
by two rounds of a series of four training blocks: (1) two-
alternative forced-choice word–definition matching where
definitions were the cues and words were the choices,
three trials for each item; (2) two-alternative forced-choice
word–definition matching where words were the cues
and definitions were the choices, three trials for each
item; (3) recall of words cued by definitions; E (4) recall
of definitions cued by words. All responses were typed on
the computer. Feedback was provided on each trial. In
total, participants received 17 exposures of each word–
definition pair. Presentation of novel and existing items
was mixed, and item order was randomized for each
block.
After the training phase in the second session, partici-
pants performed a definition recall block without feed-
back, in which the learned novel and existing words
from both training sessions served as cues to recall their
meanings. This block was included to reactivate the re-
mote condition and to minimize perceptual effects of
recency of exposure as well as to measure memory for
the remote set without intervening exposure. Given that
this task is highly demanding, we also administered a
624
Journal of Cognitive Neuroscience
Volume 30, Numero 5
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 2. Existing Word Stimuli
Stimulus
dweil
vampier
garage
sigaar
dokter
herfst
hengst
pleister
strand
kreeft
suiker
tijger
duivel
fiets
slang
vlieg
dochter
molen
keuken
baard
List 1
List 2
List 3
Translation
Stimulus
Translation
Stimulus
Translation
mop
vampire
garage
cigar
doctor
autumn
stallion
plaster
beach
lobster
sugar
tiger
devil
bike
snake
fly
daughter
mill
kitchen
beard
applaus
oorlog
vulkaan
monnik
dwerg
walvis
bijbel
planeet
bezem
hotel
insect
zwaan
tante
ridder
kelner
konijn
vinger
winter
slee
pauw
applause
war
vulcano
monk
midget
whale
bible
planet
broom
hotel
insect
swan
aunt
knight
waiter
rabbit
finger
winter
sled
peacock
reptiel
borstel
tomaat
gorilla
bliksem
tapijt
koffie
oester
piano
ladder
appel
druif
sneeuw
tulp
varken
ballet
banaan
feest
winkel
zolder
reptile
brush
tomato
gorilla
lightning
carpet
coffee
oyster
piano
ladder
apple
grape
snow
tulip
pig
ballet
banana
party
shop
attic
four-alternative forced-choice (4AFC) word–definition
matching task at the end of the testing session to confirm
that both sets of words could at least still be recognized.
The 4AFC task only included the novel words.
MEG Task
The MEG task required participants to make a natural/
artifact decision on the 20 novel and 20 existing words
from the three conditions: remote, recente, and untrained.
Each item was presented five times, for a total of 100 trials
per condition. Words were presented at the center of the
screen, in black on a gray background. A trial consisted of
a fixation screen for 1200 msec, presentation of the word
for a randomly jittered period of 1400–1800 msec, and a
response prompt for 2000 msec or until the participant
responded. Participants pressed one of two buttons,
using their left hand, to indicate whether the word
referred to a natural or manmade object. For untrained
novel objects, which had no meaning, they were instructed
to guess. Each trial was followed by a 1200-msec period for
blinking.
Semantic Decision on Base Words
The semantic decision task was designed to measure lex-
ical competition from the learned novel words with their
existing orthographic neighbors—the base words. Partic-
ipants made a speeded natural/artifact decision to the
base words of the 40 learned novel words as well as to
a control set of 20 base words from the fourth list of
novel words (cioè., not the list that was on the “untrained”
condition in the MEG task). A slower response to base
words with novel competitors (per esempio., “patat” [chips] Quando
“pamat” has been learned) as compared with base words
without any novel competitors is taken to indicate lexical
competition and thus integration of the novel word into
the lexicon. A set of 80 filler items was included to dis-
tract participants from the relation between the base
words and the learned novel words. Half of the items
in each condition required a “natural” response; and half,
an “artifact” response. Trials consisted of a 500-msec fixa-
tion cross and a 500-msec blank screen, followed by pre-
sentation of the stimulus word for 2000 msec or until
button press. Responses were measured until 1500 msec
after target onset. Participants used their left and right
Bakker-Marshall et al.
625
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
index fingers to indicate their responses, with the allo-
cation of response buttons being counterbalanced across
participants.
Primed Lexical Decision Task
The primed lexical decision task measured the ability of
novel words to speed up responses to semantically related
existing words, indicating their semantic integration into
the lexicon. Each trained novel word was presented as a
prime once with each of three semantically related targets
and once with each of three unrelated targets. Inoltre,
each trained word was presented six times as a prime with
a different nonword target, derived from unrelated exist-
ing target words by substitution of one letter. Così, there
were 240 nonword and 240 word response trials, for a total
Di 480. A trial consisted of a 500-msec fixation screen and
presentation of the prime in lower case letters for 250 msec,
followed by an interval of a blank screen for 250 msec and
presentation of the target in capital letters for 1500 msec
or until button press. Participants were instructed to
indicate as fast as possible whether the target word was
a real Dutch word or not by pressing a button under
their left index finger for “no” or their right index finger
for “yes.”
MEG Acquisition and Analysis
MEG data were acquired from 275 axial gradiometers
(CTF VSG MedTech) digitized at a sampling frequency
Di 1200 Hz, after analog low-pass filtering at 300 Hz. Head
position was tracked in real time using two coils attached
to the ear plugs and one placed at the nasion. Position of
the head was readjusted relative to the start of the re-
cording during breaks. Eye movements were recorded
using the SR Research Eyelink 1000 on the left eye.
Data analysis was performed using FieldTrip (Oostenveld,
Fries, Maris, & Schoffelen, 2011). The raw data were seg-
mented into epochs containing the baseline period and
the word presentation period (1400 msec prestimulus to
1200 msec poststimulus). Trials containing muscle artifacts,
a head position that was more than 6 mm displaced from
the starting position, and SQUID jumps were removed
(4%). Heartbeat and eye movement components were
subsequently identified using independent component
analysis and removed from the data. Consistently noisy
channels were also removed (the same three channels for
all participants and a fourth channel for one participant).
For sensor level analysis, synthetic planar gradients were
computed to facilitate topographical interpretation when
comparing across participants (Bastiaansen & Knösche,
2000). Time–frequency representations of power were
estimated for frequencies between 4 E 30 Hz using a
sliding window of 500 msec in steps of 50 msec, multiplied
with a Hanning taper. Normalized differences of power
between novel and existing words [(novel − existing) /
(novel + existing)] were computed for each level of train-
ing (untrained, recente, and remote). Normalizing mini-
mizes biases based on potential overall power differences
between conditions, for instance, due to a difference in
recency of exposure, number of trials, or presence of arti-
facts. As in Bakker et al. (2015), a window of 500–600 msec
was selected for statistical analysis, which due to the length
of the sliding window, is influenced by data between 250
E 850 msec. Power was averaged across left temporal
channels (see Figure 4A), based on the topography of
the theta effect in Bakker et al.
A Dynamic Imaging of Coherent Sources scalar beam-
forming approach was used to estimate the sources of
theta oscillations (Gross et al., 2001). T1-weighted ana-
tomical MRIs with 1-mm isotropic voxels were obtained
on a 1.5-T Siemens (Erlangen, Germany) Avanto scanner
for all participants and used to construct realistic volume
conduction models as well as individual volumetric source
models. The latter were based on a regular 3-D grid with a
1-cm resolution, created from the Montreal Neurological
Institute template brain, giving 2982 grid points within
the brain. The participants’ scans were normalized to the
Montreal Neurological Institute template brain, and the
inverse transformation was applied to the template grid,
such that a given grid point location in a participant cor-
responded to the same grid point location in volumetrically
normalized space.
For source localization of theta and beta effects, dati
from a 300- to 800-msec time window were multiplied
with a Hanning taper (leading to frequency smoothing
of ∼2 Hz) and transformed to the frequency domain
using a fast Fourier transform. This time window was
chosen to be centered around the peaks of the channel
level effects observed in Bakker et al. (2015) and to con-
tain at least two cycles of the frequency of interest. For
the theta band, a center frequency of 6 Hz was used,
and for the beta band, a center frequency of 18 Hz was
used. We used a common spatial filter across conditions
to obtain a power estimate for each grid point and con-
dition that is unbiased by potential differences (ad esempio
noise level or number of trials) between conditions. Questo
was achieved by averaging the data across all conditions
and creating a condition-averaged cross-spectral density
(CSD) matrix. From the CSD and the leadfield (the for-
ward model of sensor distribution of cortical sources),
we constructed a common spatial filter. This filter was
then applied to the CSD matrix of each condition sepa-
rately, resulting in a power estimate for each grid point
and condition. The normalized difference between novel
and existing words was then computed for each level of
training.
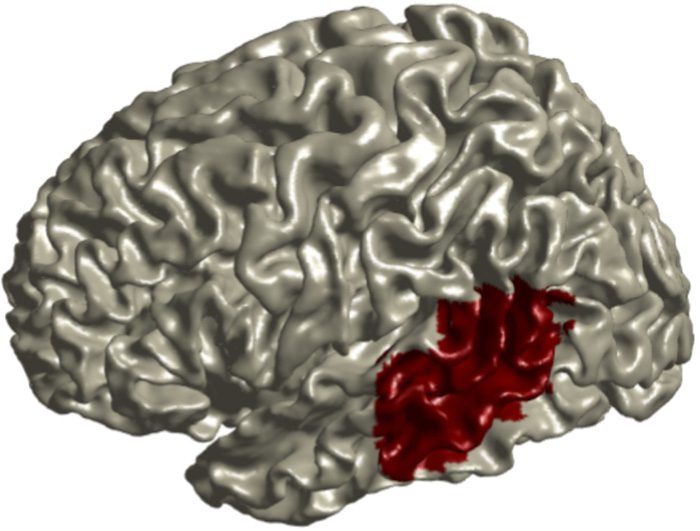
The ROI analysis of the left pTMG used a mask created
by cutting a straight line across the left MTG region of the
automated anatomical labeling (AAL) atlas (Tzourio-
Mazoyer et al., 2002) at the middle y value along the
anterior–posterior axis (Guarda la figura 2). The AAL atlas was
interpolated to the template grid, and each participant’s
theta power estimates for the grid points corresponding
626
Journal of Cognitive Neuroscience
Volume 30, Numero 5
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Recognition of novel words when cued by their defini-
tions in the 4AFC task at the end of the second session
was successful for 75% (±17.6%) of the recent words and
70.4% (±16.2%) of the remote words. This level of per-
formance suggests that, although some forgetting had
occurred, the meanings of most words of both sets were
still retrievable at the time of testing.
MEG Task
The novel untrained condition was excluded from behav-
ioral analysis of the semantic decision task performed
during the MEG recording, as these words were mean-
ingless and participants were instructed to give a random
response to them. Overall accuracy across the other con-
ditions was 88.4% (recent novel: 90.6%, remote novel:
84.8%; recent existing: 88%, remote existing: 88.4%).1 Dopo
the procedure taken in our previous EEG study (Bakker
et al., 2015), errors and RTs above or below 1.5 SDs from
the mean were removed for RT analysis (19.1%). A Day ×
Lexicality ANOVA did not reveal an interaction or main
effects (all ps > .4).
Semantic Priming
Lexical decision accuracy in the priming task was 93.6%.
RTs were analyzed using an ANOVA with factors Related-
ness (related/unrelated prime) and Day (recent/remote).
Errors and RTs below or above 1.5 SDs from the mean
were removed (14.4%). The RT analysis did not reveal a
main effect of Relatedness (cioè., an overall priming effect;
F(1, 27) = 0.25, p = .621) or an effect of Day (F(1, 27) =
0.148, p = .704). The interaction, Tuttavia, was signifi-
cant, reflecting a change in the direction of the priming
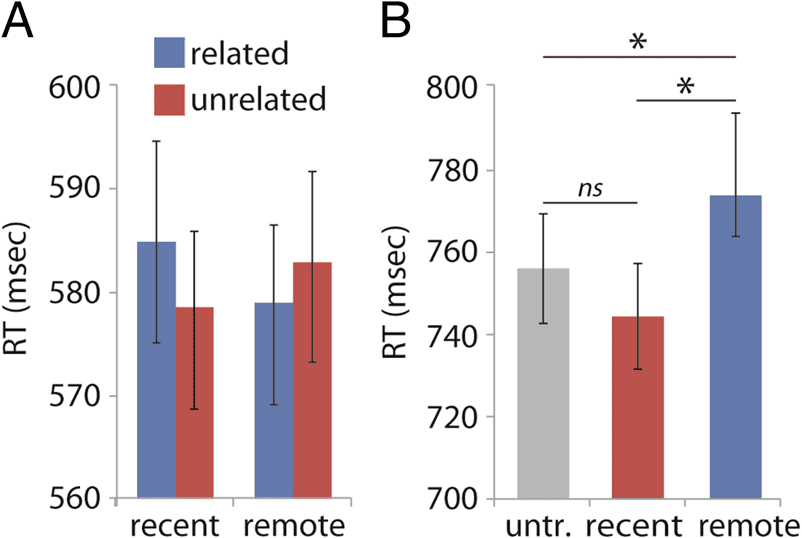
effect (F(1, 27) = 4.297, p = .048). In the recent condi-
zione, a numerical slowing down (8 msec) was observed
for related prime–target pairs (T(27) = 1.7, p = .101, eval-
uated at α = .025 to correct for multiple comparisons
following the sequential Holm–Bonferroni [H-B] method;
Holm, 1979). In the remote condition, the expected facil-
itation occurred (5 msec) but did not reach significance
(T(27) = 1.214, p = .235, H-B α = .05). This pattern sug-
gests that the novel words’ ability to prime existing words
did increase with consolidation but did not (yet) reach
the point at which they significantly facilitated recognition
(see Figure 3A).
Lexical Competition
Lexical competition between the novel words and the
base words from which they had been derived (per esempio.,
“patat,” chips, the base word of “pamat”) was tested with
a one-way ANOVA on the three conditions: remote (novel
neighbor learned on Day 1), recente (novel neighbor
learned on Day 2), and untrained (no novel neighbor).
Accuracy was 87.3%, and errors and RTs below or above
1.5 SDs from the mean were removed from the RT analysis
Bakker-Marshall et al.
627
Figura 2. Left posterior MTG mask used for ROI analysis, created
by cutting a straight line across the left MTG region of the AAL atlas
(Tzourio-Mazoyer et al., 2002) at the middle y value along the
anterior–posterior axis.
to the left pMTG mask were extracted. The averages were
entered into a repeated-measures ANOVA.
To establish whether any theta effects were driven
mainly by induced or evoked activity, we repeated our
main statistical analysis on the phase-locked portion of
the response. To compute the theta power of the evoked
risposta, we averaged the single-trial Fourier coefficients
and took the magnitude squared of the result. Given that
Fourier transformation is a linear operation, this proce-
dure is mathematically equivalent to single trial averaging
in the time domain, followed by Fourier transformation.
To compare the evoked theta power across the three
conditions, we calculated the theta-band spectral power
of the condition-specific evoked response in the 300- A
800-msec time window. We then performed a repeated-
measures ANOVA on the mean theta power values, COME
before.
RESULTS
Behavioral Results
Training and Memory Performance
In the last block of training, participants could produce
the correct category name (per esempio., “cat”) when cued by a
novel word in 97.1% (±5%) of items on Day 1 E
97.7% (±4%) on Day 2. The number of definition fea-
tures given for novel words was 87.8% (±10.3%) SU
Day 1 E 89.7% (±10.4%) on Day 2. Così, participants
successfully learned the novel word meanings and per-
formed similarly on both sets.
In the definition recall task without feedback, presented
immediately after the training procedure on Day 2, 94.1%
(±7.6%) of the features were recalled in the novel recent
condition; E 71% (±23.3%), in the novel remote condi-
zione. The correct category (per esempio., “cat”) was given in 95.7%
(±6.8%) of the novel recent words and 72.5% (±20.8%) Di
the novel remote words. As expected, performance for
existing words was at ceiling.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
C
N
_
UN
_
0
1
2
4
0
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
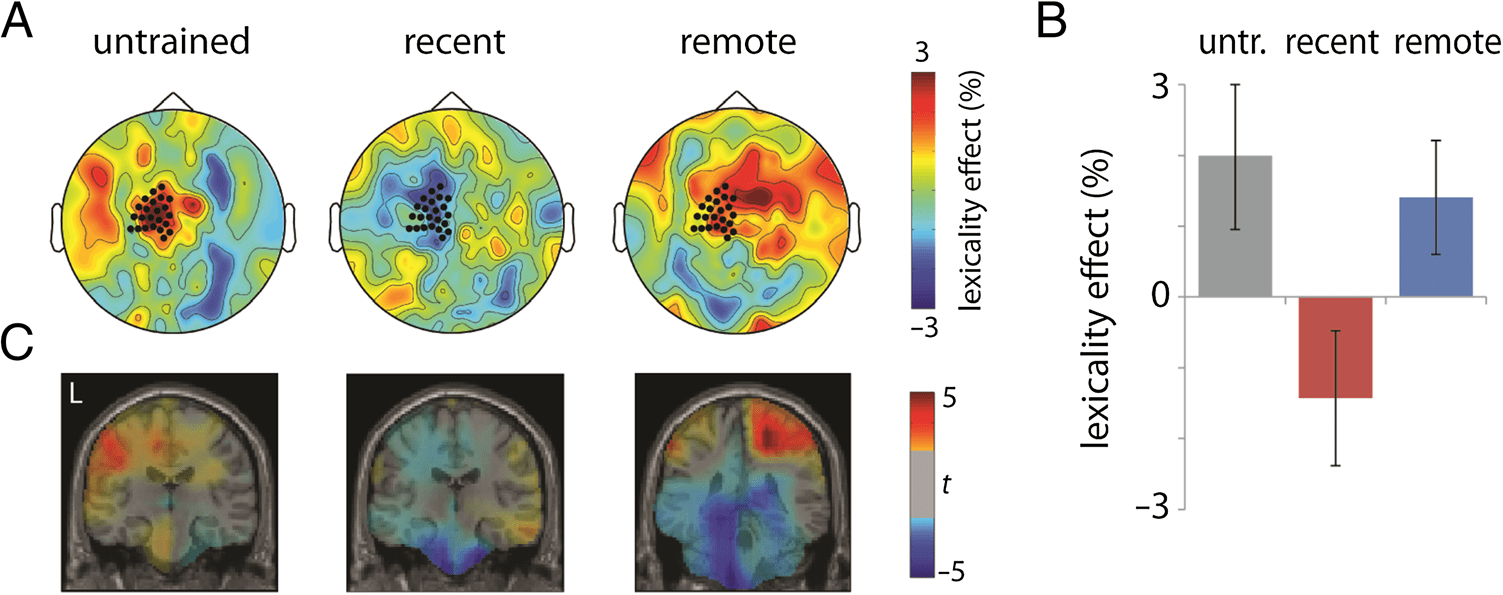
Condition was nonsignificant (F(2, 48) = 1.265, p =
.292). One-sample t tests for each of the three conditions
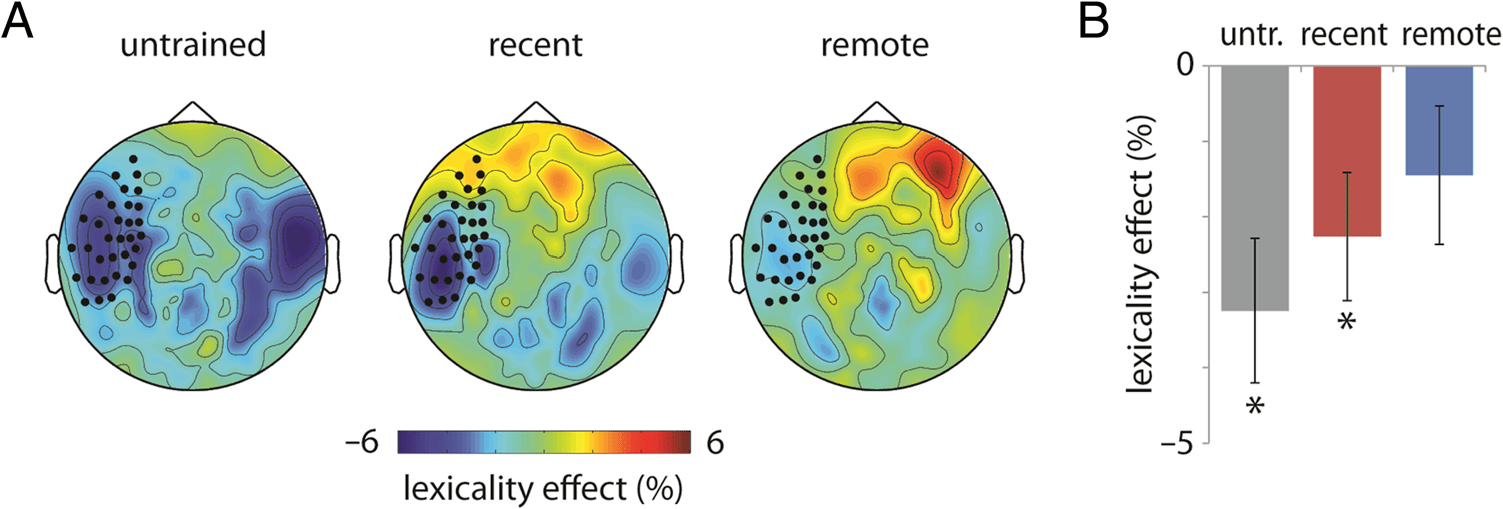
confirmed that untrained novel words elicited lower theta
power than existing words (T(24) = 3.423, p = .002, H-B
α = .017). This lexicality effect remained significant for
recent words (T(24) = 2.660, p = .014, H-B α = .025)
but was absent in the remote condition (T(24) = 1.555,
p = .133, H-B α = .05; Guarda la figura 4).
Our main hypothesis was that theta lexicality effects in
the left pMTG decrease with consolidation. We therefore
extracted the power estimates for the grid points cor-
responding to the anatomical left pMTG2 for each condi-
tion and performed a one-way repeated-measures ANOVA
on the regional averages (see Figure 5A). This analysis
revealed a significant change in the magnitude of the lex-
icality effect across conditions (F(2, 48) = 3.832, p =
.029). One-sample t tests on the normalized differences
per condition showed that both untrained and recent
novel words elicited lower theta responses in the pMTG
than existing words (untrained: T(24) = 4.35, P < .001,
H-B α = .017; recent: t(24) = 3.457, p = .002, H-B α =
.025). In the remote condition, this lexicality effect was
no longer significant (t(24) = 0.286, p = .777, H-B α =
.05). To quantify the change in magnitude of lexicality
effects, we then compared the novel-existing difference
for untrained words with the novel-existing difference for
recent words. This difference was not reliable (t(24) =
0.424, p = .676, H-B α = .05), suggesting that training
alone did not significantly reduce the lexicality effect.
Comparing the differences for recent and remote words
revealed a marginally significant decrease in lexicality ef-
fects (t(24) = 2.258, p = .033, H-B α = .025), indicating
that the interaction effect was largely driven by a difference
between the recent and remote conditions.
To control for the possibility that the pMTG effects re-
sulted from bleeding of activation from adjacent areas,
the main one-way ANOVA on the normalized differences
was repeated in the left superior temporal gyrus (STG)
and inferior temporal gyrus (ITG), angular gyrus, supra-
marginal gyrus, and inferior parietal lobule (as defined in
the AAL atlas; Tzourio-Mazoyer et al., 2002). None of these
Figure 3. Behavioral integration results. (A) RTs to related and
unrelated prime–target pairs for the remote and recent conditions in
the primed lexical decision task. Errors bars denote standard errors.
(B) RTs to the existing base words of untrained (untr.), recent,
and remote novel words in the semantic decision task. Error bars
denote standard errors.
(21.7%). ANOVA revealed a main effect of Condition (F(2,
54) = 4.478, p = .016). Responses in the remote condition
were 18 msec slower than those in the untrained condi-
tion, indicating that novel words (e.g., “pamat”) entered
into lexical competition with their existing neighbors
(e.g., “patat”; t(27) = 2.332, p = .027, marginally significant
at H-B α = .025). In contrast, there was no sign of com-
petition in the recent condition (t(27) = 1.078, p = .291,
H-B α = .05). The difference between the two effects
(remote–untrained vs. recent–untrained) was significant
(t(27) = 2.693, p = .012; see Figure 3B).
MEG Results
The change over time in channel-level theta power (4–
8 Hz) in the 500- to 600-msec time window over left
fronto-temporal channels was analyzed with a one-way
ANOVA on the normalized differences between novel
and existing words in each of the three conditions, as
in Bakker et al. (2015). This analysis replicated the pat-
tern observed in that study, although the main effect of
Figure 4. Lexicality effects
(novel − existing / novel +
existing) in the theta band
(4–8 Hz) for untrained (untr.),
recent, and remote words,
averaged across participants.
(A) Sensor topography of
the average lexicality effect,
averaged across 500–600 msec.
Blue indicates a negative
difference in power (less
synchronization for novel
words than existing words).
(B) Lexicality effects averaged
across the left fronto-temporal
channels highlighted in A,
based on Bakker et al. (2015).
628
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
c
n
_
a
_
0
1
2
4
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
c
n
_
a
_
0
1
2
4
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
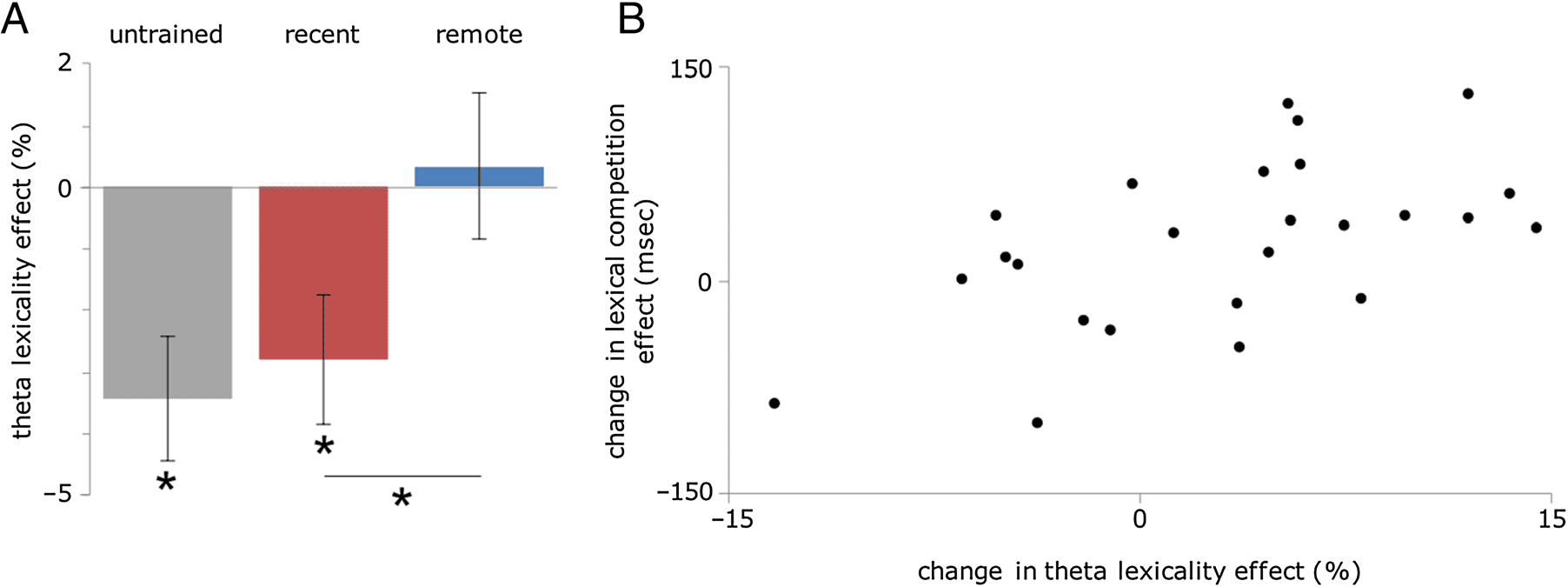
Figure 5. (A) Lexicality effects (novel − existing / novel + existing) in the theta band (4–8 Hz) averaged across the left pMTG ROI at source
level. Error bars denote standard error. (B) Correlation between the decrease in lexicality effects in the left pMTG and the increase in competition
effects (difference between remote and recent effects) after consolidation.
regions exhibited a change in lexicality effects across con-
ditions (all ps > .069), confirming that the effect is specific
to the MTG. Secondo, we investigated whether the observed
theta effect could be explained by time-locked (per esempio., N400-
related) activity. We did not observe a change in lexicality
effects within the evoked part of the activity (F(2, 48) =
1.869, p = .165), suggesting that the effect was driven at
least predominantly by induced activity.
To investigate the relation between theta power and
behavioral integration effects, we computed the correla-
tion between the magnitude of the theta lexicality effect
in both learned conditions and the magnitude of the be-
havioral priming and competition effects. No correlations
were observed between the theta lexicality effect and the
priming effect for remote or recent words or between the
theta lexicality effect and the competition effect for remote
or recent words (all ps > .5). Tuttavia, the degree to which
the theta lexicality effect decreased after consolidation (Rif-
mote lexicality effect vs. recent lexicality effect) was corre-
lated with the increase in competition effects after
consolidation (remote competition effect vs. recent compe-
tition effect; R(23) = .56, P < .001). This suggests that a
more word-like theta response in the remote condition is
related to the emergence of competition effects from those
words (see Figure 5B). No correlation was observed be-
tween the change in theta lexicality effects and the change
in priming effects, possibly because the priming effect re-
mained weak even in the remote condition (see Figure 3).
Given our strong prior hypothesis regarding the left
pMTG, we restricted ROI analysis and correlations with
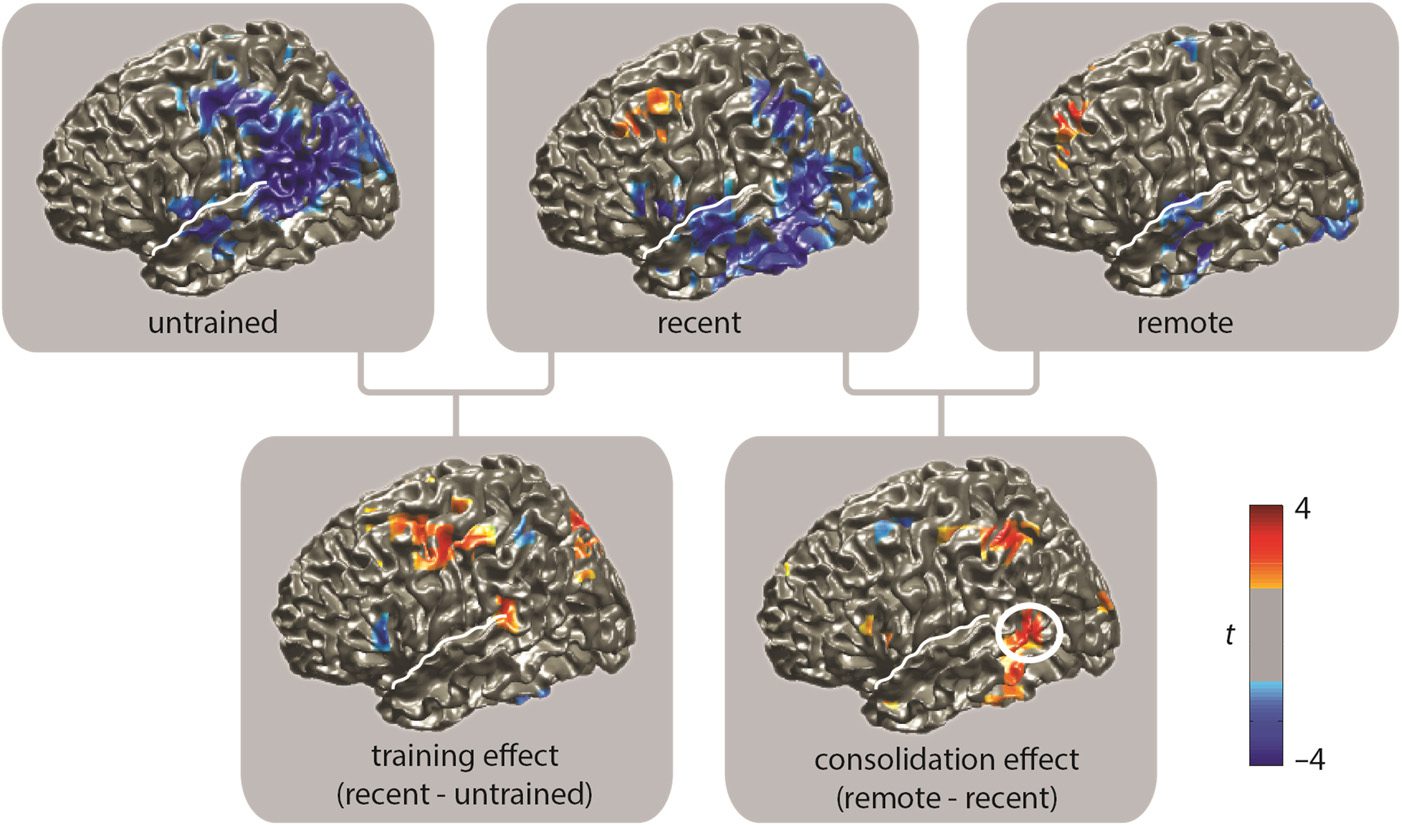
Figure 6. Estimated source
activity for the lexicality
effects, computed for
4–8 Hz and 300–800 msec.
Uncorrected parametric
t values (thresholded at
p = .05) are plotted to illustrate
the most consistent source
estimates. (Note that the use
of t values here is merely a
way to visually illustrate the
relative consistency of the
effects across different areas,
and no conclusions about the
significance of these effects
can be drawn from this figure.)
The white line indicates the
location of the Sylvian fissure
for orientation purposes. The
top row shows the lexicality
effect per condition as a
T statistic (novel-existing),
with blue indicating lower theta power for novel words as compared with existing words and red indicating higher theta power for novel than
existing words. The bottom left shows the change in magnitude of the lexicality effect between the untrained and recent conditions. Red colors
signify a decrease of the lexicality effect in the recent condition, that is, an effect of training. The bottom right shows the change in magnitude of
the lexicality effect between the recent and remote conditions. Red here indicates a decrease of the lexicality effect in the remote condition, that is,
a consolidation effect. The white circle indicates the location of the largest effect, which is estimated to be in the posterior MTG.
Bakker-Marshall et al.
629
Figure 7. Lexicality effects
(novel − existing/novel +
existing) in the beta band
(16–20 Hz) for untrained
(untr.), recent, and remote
words. Red indicates a
positive difference in power
(more synchronization for
novel than existing words);
blue, the opposite. (A)
Topography of the lexicality
effect, averaged across
300–500 msec. (B) Lexicality
effects averaged across the
left fronto-temporal channels
highlighted in A, based
on Bakker et al. (2015).
(C) Estimated sources of beta lexicality effects. Colors follow the same conceptual interpretation as in A, but t values (thresholded at p = .05)
are plotted to illustrate the most reliable source estimates.
behavioral effects to this region. However, to illustrate
the overall spatial pattern of lexicality effects in the theta
band, Figure 6 shows a surface rendering of the estimated
source activity of the difference between novel and exist-
ing words in each condition, expressed as a T statistic.
Note that these values are uncorrected and serve only
to illustrate the topography of the effect, not as a test of
significance. Furthermore, the spatial resolution of MEG
is limited, and the anatomical location of sources is there-
fore only an approximation. Untrained words exhibited
the strongest lexicality effect in the region of the left STG
and MTG, extending medially into the medial-temporal
lobe and dorsally into the angular gyrus, supramarginal
gyrus, and precentral and postcentral gyrus as well as in
the right medial-temporal lobe. In the recent condition,
the largest difference was estimated to be around the
ITG and fusiform gyrus, MTG, and anterior STG, extending
into the inferior frontal gyrus. The difference in theta
power also seems to be reflected in the region of the infe-
rior parietal lobule and postcentral gyrus. The peak voxel
for the smaller lexicality effect in the remote condition
was estimated to be in the left fusiform gyrus, and this
source extended into the medial-temporal lobe and ante-
rior part of the ITG. The right fusiform gyrus also exhibited
a lexicality effect. As illustrated at the bottom left of Fig-
ure 6, training did not reduce the lexicality effect in the
left pMTG. In contrast, comparison of the effects in the
remote versus recent conditions showed that lexicality
effects after consolidation were most reduced (the highest
t values were observed) in the posterior part of the left
MTG, consistent with the MTG ROI analysis (bottom right
of Figure 6). A second peak was observed in the left infe-
rior parietal lobule.
Unlike in our previous EEG data (Bakker et al., 2015),
we did not observe a clear consolidation effect in the
lower beta band (Figure 7). On the basis of the EEG find-
ings, we analyzed 16- to 20-Hz activity in a 300- to 500-msec
window over left central channels. In line with the earlier
findings, this revealed numerically weaker beta desyn-
chronization (i.e., higher beta power) for untrained novel
words compared with existing words (t(24) = 2.072, p =
.049, H-B α = .017). The source of this effect (at ∼18 Hz)
was localized most strongly to the left postcentral gyrus.
However, there was a numerical effect in the opposite di-
rection for the recent condition (t(24) = 1.591, p = .125,
H-B α = .025) and a numerical difference in the same direc-
tion for the remote condition (t(24) = 1.695, p = .103, H-B
α = .05). The estimated source of the effect for remote
words was in the right postcentral gyrus.
DISCUSSION
The current study tested the hypothesis that the develop-
ment of lexical representations of novel words causes a
more word-like pattern of theta synchronization in the
left pMTG after consolidation. As expected, the sensor
level data show that untrained novel words (i.e., pseudo-
words) elicited lower theta synchronization than existing
words in the left fronto-temporal sensors. The estimated
sources of this effect comprised a mostly left-lateralized
network of temporal and parietal regions. ROI analysis
of the pMTG revealed that training of novel words only
before test (recent condition) did not reduce the differ-
ence between theta responses to novel and existing
words. In contrast, the lexicality effect in the pMTG was
significantly reduced for responses to words learned 24 hr
previously (remote condition), which no longer differed
from responses to existing words. Comparing the magni-
tude of the lexicality effect for recent versus remote words
further revealed that the largest decrease occurred in the
pMTG (Figure 6), confirming the hypothesis that this re-
gion supports the formation and integration of lexical
representations for novel words. Although the limited
spatial resolution of MEG prohibits detailed conclusions
about the anatomical location of sources, and several
language-related areas may contribute to this effect (such
as the STG, ITG, and inferior parietal lobule), the whole-
brain analysis suggests that the largest decrease occurred
in the posterior part of the left MTG.
630
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
c
n
_
a
_
0
1
2
4
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
These results replicate our earlier observation of de-
creased theta lexicality effects after consolidation (Bakker
et al., 2015) and link this theta effect to fMRI data impli-
cating the left MTG (and, in particular, its posterior half )
in novel word consolidation (Takashima et al., 2014,
2017). The posterior MTG has been proposed to function
as a lexical “association area,” which maps between
word-form representations and semantic information dis-
tributed throughout the cortex (Gow, 2012; Lau et al.,
2008; Hickok & Poeppel, 2004, 2007). Theta synchroni-
zation may be one of the mechanisms by which these
widely distributed representations are simultaneously
activated, enabling them to be accessed as a single lexical
item. On this view, the activation of such a coherent
word-specific network bound together by the pMTG
would lead to local theta synchronization, producing
the scalp-level power increase over left temporal and
frontal channels that was observed here and in other
studies (Bakker et al., 2015; Marinkovic et al., 2012;
Bastiaansen et al., 2005, 2008; Krause et al., 2006). The
degree to which form- and meaning-related information
for a given word is available and accessible via the pMTG
thus predicts the level of theta power during recognition
of that word. Training a novel word with its meaning ini-
tiates changes in the pMTG, but only after a consolida-
tion period are the new connections strong enough to
elicit word-like oscillatory responses. It remains to be de-
termined if, for each novel word, a new representation is
formed in the pMTG that is linked to word-form and
semantic (and syntactic) representations of that word
(e.g., a lemma representation; see, e.g., Levelt, Roelofs,
& Meyer, 1999). Alternatively, the pMTG may serve to
connect the different types of lexical representations
located elsewhere without the formation of a new repre-
sentation such as a lemma. On either view, however,
what is consolidated is the way in which the novel word
is integrated, not only with knowledge about that word
(e.g., about its form or meaning) but also with knowl-
edge about other words.
The consolidation of lexical representations has been
claimed to underlie the emergence of behavioral evi-
dence of interaction between novel and existing words,
such as lexical competition (Bakker et al., 2014; Dumay &
Gaskell, 2007, 2012; Gaskell & Dumay, 2003) and semantic
priming (van der Ven et al., 2015; Tamminen & Gaskell,
2013). In line with these studies, the present data revealed
a competition effect between novel words and their exist-
ing orthographic neighbors in a semantic decision task.
This competition effect was found for novel words learned
the previous day, but not for words learned immediately
before test. Such delayed competition effects have gen-
erally been interpreted as evidence for the transformation
of initially episodic memory traces toward neocortically
integrated lexical representations (Davis & Gaskell, 2009).
On the basis of the behavioral data alone, however, it is
difficult to exclude the possibility that consolidation of
the episodic trace itself may increase its accessibility and
enable competition effects. Here, we show that the in-
crease in competition after consolidation correlated with
the decrease in pMTG theta lexicality effects, suggesting
that novel words that elicited more word-like theta re-
sponses were better integrated in the existing lexicon. This
novel finding provides further empirical support for the
assumption that novel words’ ability to interact with exist-
ing words relies on the formation of lexical representations
and/or the links between them, rather than the strengthen-
ing of an episodic memory trace.
Given that competition is generally assumed to occur
at the modality-specific lexeme level (see Bakker et al.,
2015, for a discussion), a question that arises from this
finding is how the pMTG contributes to the competition
process itself. One potential explanation is that the
pMTG lemma representation itself does not play a direct
role in the competition process, and the correlation arises
from the fact that the lexicality effect on pMTG theta
power and the behavioral competition effect both result
from the same general consolidation process that trans-
forms episodic memory traces into distributed neo-
cortical representations. Alternatively, the availability of
a lemma representation may strengthen competition
indirectly through top–down activation of lexemes and/
or sublexical representations. Distinguishing between
these explanations is beyond the scope of the current
design but remains an interesting question for future
investigations.
The current data are consistent with the CLS claim that
neocortical lexical links are established slowly during off-
line consolidation (Davis & Gaskell, 2009; McClelland
et al., 1995). In this framework, the MTG can be seen as
gradually taking over the binding function of the hippo-
campus as novel words are consolidated. However, recent
behavioral data suggest that novel words are able to inter-
act with existing words immediately after training when
training encourages integration (Coutanche & Thompson-
Schill, 2014; Lindsay & Gaskell, 2013; Szmalec, Page, &
Duyck, 2012) or when competition is measured with a
test that is more sensitive to word-specific competition
(Kapnoula, Packard, Gupta, & McMurray, 2015). This may
indicate that lexicalization in the pMTG occurs imme-
diately during learning but requires further offline strength-
ening to produce behavioral effects large enough to be
observed with RT methods. This more quantitative neo-
cortical view of lexicalization would be in line with findings
that information that is easily related to prior knowledge
relies to a much smaller extent on the hippocampus than
completely unrelated information, both during encoding
and later retrieval (van Kesteren, Ruiter, Fernández, &
Henson, 2012; Tse et al., 2007). Future work should
attempt to characterize the precise contribution of the
hippocampal and neocortical systems in word learning.
The current data suggest that theta connectivity between
the hippocampus and neocortical structures before and
after consolidation may provide a useful measure for
this question.
Bakker-Marshall et al.
631
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
c
n
_
a
_
0
1
2
4
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
In contrast to Bakker et al. (2015), we did not observe
a clear pattern of lexicalization in the lower beta band.
Untrained novel words elicited weaker beta desynchro-
nization than existing words, in line with proposals that
beta desynchronization reflects retrieval of semantic infor-
mation (for a review, see Hanslmayr, Staudigl, & Fellner,
2012). Whereas previous semantic beta effects have been
localized to the left inferior frontal gyrus (Hanslmayr
et al., 2011; Meeuwissen, Takashima, Fernández, & Jensen,
2011), the present effect appeared to be generated by
the left postcentral gyrus. This is commensurate with an
explanation in terms of motor preparation and response
certainty (Alegre et al., 2004). However, the observed pat-
tern is unlikely to reflect only a difference in motor re-
sponse preparation as participants responded with their
left hand, which should produce a right-lateralized effect.
It is possible therefore that the beta effect for untrained
novel words does reflect a memory-related process. We
observed a decreased lexicality effect in the recent con-
dition, but the effect for remote words surprisingly re-
turned to the level of untrained words. The source of
this latter effect appeared to be more right lateralized.
This pattern is difficult to interpret, but in any case, it does
not support a role for beta desynchronization in lexicali-
zation. Future work may be able to shed light on the role
of beta desynchronization through the use of a task that
does not require any motor response.
In conclusion, the work reported here demonstrates
that the left pMTG is associated with a consolidation-
dependent development toward more word-like theta
responses to novel words. The decrease in the difference
in theta power between novel and existing words was
found to be correlated with a postconsolidation increase
in competition effects, reflecting functional integration of
novel words with their existing neighbors. This suggests
that theta synchronization enables distributed informa-
tion to be integrated into lexical representations bound
by the pMTG and that the incorporation of novel words
into this system benefits from offline consolidation. The
current work thus brings together previous observations
of behavioral consolidation effects (e.g., Bakker et al.,
2014; Tamminen & Gaskell, 2013; Dumay & Gaskell, 2007,
2012; Gaskell & Dumay, 2003), increased pMTG activa-
tion in fMRI (Takashima et al., 2014), and more word-
like theta responses in EEG after consolidation of novel
words (Bakker et al., 2015).
Acknowledgments
This research was funded by The Netherlands Organization
for Scientific Research (NWO) Brain and Cognition Grant
433-09-239.
Reprint requests should be sent to Iske Bakker-Marshall,
Wellcome Centre for Integrative Neuroimaging, University of
Oxford, FMRIB, John Radcliffe Hospital, Oxford OX3 9DU,
United Kingdom, or via e-mail: iske.marshall@psy.ox.ac.uk.
Notes
1. Note that performance on existing words was not perfect
because, even with familiar words, it is not always trivial to
make a natural/artifact decision, especially under time pressure.
Participants differed, for example, in their responses to words
such as “monk,” with some reasoning that monks are artifacts
because the word refers to a cultural concept, whereas others
considered monks natural as they are humans.
2. Using the whole left MTG mask from the AAL atlas did not
change the pattern or significance level of any of these effects.
REFERENCES
Alegre, M., Gurtubay, I. G., Labarga, A., Iriarte, J., Valencia, M.,
& Artieda, J. (2004). Frontal and central oscillatory changes
related to different aspects of the motor process: A study in go/
no-go paradigms. Experimental Brain Research, 159, 14–22.
Baayen, R. H., Piepenbrock, R., & Gulikers, L. (1995). The
CELEX lexical database [webcelex]. Philadelphia, PA:
University of Pennsylvania Linguistic Data Consortium.
Bakker, I., Takashima, A., van Hell, J. G., Janzen, G., & McQueen,
J. M. (2014). Competition from unseen or unheard novel
words: Lexical consolidation across modalities. Journal of
Memory and Language, 73, 116–130.
Bakker, I., Takashima, A., van Hell, J. G., Janzen, G., & McQueen,
J. M. (2015). Changes in theta and beta oscillations as
signatures of novel word consolidation. Journal of Cognitive
Neuroscience, 27, 1286–1297.
Bastiaansen, M. C. M., & Knösche, T. R. (2000). Tangential
derivative mapping of axial MEG applied to event-related
desynchronization research. Clinical Neurophysiology, 111,
1300–1305.
Bastiaansen, M. C. M., Oostenveld, R., Jensen, O., & Hagoort, P.
(2008). I see what you mean: Theta power increases are
involved in the retrieval of lexical semantic information. Brain
and Language, 106, 15–28.
Bastiaansen, M. C. M., van der Linden, M., ter Keurs, M., Dijkstra,
T., & Hagoort, P. (2005). Theta responses are involved in
lexical–semantic retrieval during language processing. Journal
of Cognitive Neuroscience, 17, 530–541.
Coutanche, M. N., & Thompson-Schill, S. L. (2014). Fast
mapping rapidly integrates information into existing memory
networks. Journal of Experimental Psychology: General,
143, 2296–2303.
Davis, M. H., & Gaskell, M. G. (2009). A complementary systems
account of word learning: Neural and behavioural evidence.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 364, 3773–3800.
De Deyne, S., & Storms, G. (2008). Word associations: Norms
for 1,424 Dutch words in a continuous task. Behavior
Research Methods, 40, 198–205.
Dumay, N., & Gaskell, M. G. (2007). Sleep-associated changes in
the mental representation of spoken words. Psychological
Science, 18, 35–39.
Dumay, N., & Gaskell, M. G. (2012). Overnight lexical
consolidation revealed by speech segmentation. Cognition,
123, 119–132.
Frankland, P. W., & Bontempi, B. (2005). The organization of
recent and remote memories. Nature Reviews Neuroscience,
6, 119–130.
Gaskell, M. G., & Dumay, N. (2003). Lexical competition and
the acquisition of novel words. Cognition, 89, 105–132.
Gow, D. W. (2012). The cortical organization of lexical
knowledge: A dual lexicon model of spoken language
processing. Brain and Language, 121, 273–288.
Gross, J., Kujala, J., Hämäläinen, M., Timmermann, L.,
Schnitzler, A., & Salmelin, R. (2001). Dynamic imaging of
632
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
c
n
_
a
_
0
1
2
4
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
coherent sources: Studying neural interactions in the human
brain. Proceedings of the National Academy of Sciences,
U.S.A., 98, 694–699.
Hanslmayr, S., Staudigl, T., & Fellner, M.-C. (2012). Oscillatory
power decreases and long-term memory: The information
via desynchronization hypothesis. Frontiers in Human
Neuroscience, 6, 74.
Hanslmayr, S., Volberg, G., Wimber, M., Raabe, M., Greenlee,
M. W., & Bäuml, K.-H. T. (2011). The relationship between
brain oscillations and BOLD signal during memory formation:
A combined EEG–fMRI study. Journal of Neuroscience, 31,
15674–15680.
Hickok, G., & Poeppel, D. (2004). Dorsal and ventral streams:
A framework for understanding aspects of the functional
anatomy of language. Cognition, 92, 67–99.
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8, 393–402.
Holm, S. (1979). A simple sequential rejective multiple test
procedure. Scandinavian Journal of Statistics, 6, 65–70.
Kapnoula, E. C., Packard, S., Gupta, P., & McMurray, B. (2015).
Immediate lexical integration of novel word forms. Cognition,
134, 85–99.
Krause, C. M., Grönholm, P., Leinonen, A., Laine, M., Säkkinen,
A.-L., & Söderholm, C. (2006). Modality matters: The effects
of stimulus modality on the 4- to 30-Hz brain electric
oscillations during a lexical decision task. Brain Research,
1110, 182–192.
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network
for semantics: (De)constructing the N400. Nature Reviews
Neuroscience, 9, 920–933.
Levelt, W. J. M., Roelofs, A., & Meyer, A. S. (1999). A theory of
lexical access in speech production. Behavioral and Brain
Sciences, 22, 1–38.
Lindsay, S., & Gaskell, M. G. (2013). Lexical integration of novel
words without sleep. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 39, 608–622.
Marinkovic, K., Rosen, B. Q., Cox, B., & Kovacevic, S.
(2012). Event-related theta power during lexical-semantic
retrieval and decision conflict is modulated by alcohol
intoxication: Anatomically constrained MEG. Frontiers in
Psychology, 3, 121.
Marr, D. (1970). A theory for cerebral neocortex. Proceedings of
the Royal Society of London, Series B, Biological Sciences,
176, 161–234.
McClelland, J. L., McNaughton, B. L., & O’Reilly, R. C. (1995).
Why there are complementary learning systems in the
hippocampus and neocortex: Insights from the successes and
failures of connectionist models of learning and memory.
Psychological Review, 102, 419–457.
Meeuwissen, E. B., Takashima, A., Fernández, G., & Jensen, O.
(2011). Evidence for human fronto-central gamma activity
during long-term memory encoding of word sequences. PLoS
One, 6, e21356.
Nelson, D. L., McEvoy, C. L., & Schreiber, T. A. (1998). The
University of South Florida word association, rhyme, and
word fragment norms. Retrieved from www.usf.edu/
FreeAssociation
Oldfield, R. C. (1971). The assessment and analysis of handedness:
The Edinburgh inventory. Neuropsychologia, 9, 97–113.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational
Intelligence and Neuroscience, 2011, 156869.
Prabhakaran, R., Blumstein, S. E., Myers, E. B., Hutchison, E., &
Britton, B. (2006). An event-related fMRI investigation of
phonological–lexical competition. Neuropsychologia, 44,
2209–2221.
Squire, L. R., & Alvarez, P. (1995). Retrograde amnesia and
memory consolidation: A neurobiological perspective. Current
Opinion in Neurobiology, 5, 169–177.
Szmalec, A., Page, M. P. A., & Duyck, W. (2012). The
development of long-term lexical representations through
Hebb repetition learning. Journal of Memory and Language,
67, 342–354.
Takashima, A., Bakker, I., van Hell, J. G., Janzen, G., &
McQueen, J. M. (2014). Richness of information about
novel words influences how episodic and semantic memory
networks interact during lexicalization. Neuroimage, 84,
265–278.
Takashima, A., Bakker, I., van Hell, J. G., Janzen, G., &
McQueen, J. M. (2017). Interaction between episodic and
semantic memory networks in the acquisition and
consolidation of novel spoken words. Brain and Language,
167, 44–60.
Tamminen, J., & Gaskell, M. G. (2013). Novel word integration
in the mental lexicon: Evidence from unmasked and masked
semantic priming. Quarterly Journal of Experimental
Psychology, 66, 1001–1025.
Tse, D., Langston, R. F., Kakeyama, M., Bethus, I., Spooner,
P. A., Wood, E. R., et al. (2007). Schemas and memory
consolidation. Science, 316, 76–82.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, O., Delcroix, N., et al. (2002). Automated anatomical
labeling of activations in SPM using a macroscopic anatomical
parcellation of the MNI MRI single-subject brain. Neuroimage,
15, 273–289.
van der Ven, F., Takashima, A., Segers, E., & Verhoeven, L.
(2015). Learning word meanings: Overnight integration and
study modality effects. PLoS One, 10, e0124926.
van Kesteren, M. T. R., Ruiter, D. J., Fernández, G., & Henson,
R. N. (2012). How schema and novelty augment memory
formation. Trends in Neurosciences, 35, 211–219.
Bakker-Marshall et al.
633
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
2
1
1
7
8
7
4
6
5
/
j
o
c
n
_
a
_
0
1
2
4
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3