The What and How of Observational Learning
Sara Torriero1, Massimiliano Oliveri1,2, Giacomo Koch1,3,
Carlo Caltagirone1,3, and Laura Petrosini1,4
Astratto
& Neuroimaging evidence increasingly supports the hypothesis
that the same neural structures subserve the execution, imagi-
nation, and observation of actions. We used repetitive trans-
cranial magnetic stimulation (rTMS) to investigate the specific
roles of cerebellum and dorsolateral prefrontal cortex (DLPFC)
in observational learning of a visuomotor task. Subjects observed
an actor detecting a hidden sequence in a matrix and then
performed the task detecting either the previously observed
sequence or a new one. rTMS applied over the cerebellum
before the observational training interfered with performance of
the new sequence, whereas rTMS applied over the DLPFC inter-
fered with performance of the previously observed one. When
rTMS applied over cerebellar or prefrontal site was delivered
after the observational training, no influence was observed on
the execution of the task. These results furnish new insights on
the neural circuitry involved in the single component of obser-
vational learning and allow us to hypothesize that cerebellum
and DLPFC interact in planning actions, the former by permitting
the acquisition of procedural competencies and the latter by
providing flexibility among already acquired solutions. &
INTRODUCTION
Observational learning is the ability to learn to perform
an action by seeing it done. It does not just involve
copying an action but, Piuttosto, requires that the observer
transform the observation into an action as similar as
possible to the actor’s in terms of the goal to be
achieved and the motor strategies to be applied. Obser-
vational learning plays a pivotal role in the acquisition of
social skills and in the transfer of cultural knowledge and
tool use and, così, becomes a fundamental process
through which infants learn about adults (Meltzoff,
1999; Byrne & Russon, 1998). The mechanisms that
couple observation and learning are very similar to those
involved in learning ‘‘by doing.’’ Experimental and
neuroimaging studies support the position that these
kinds of learning at least partially share common neural
substrates related to the cognitive stages of motor
controllo. Learning an action by observation and by mental
practice involves rehearsal of the same neural pathways.
Infatti, common brain areas and computational process-
ing are activated in preparing actions, in mentally rep-
resenting one’s own action, and in observing another’s
action (Gre`zes & Decety, 2001). Using measures of
human event-related brain potentials similar involve-
ment of medial frontal and motor cortex was demon-
strated in the monitoring of one’s own actions and the
actions of others (van Schie, Mars, Coles, & Bekkering,
1IRCCS Fondazione ‘‘Santa Lucia’’, 2University of Palermo, 3Uni-
versity of Rome Tor Vergata, 4University of Rome ‘‘La Sapienza’’
2004). Studies in monkeys and in humans show the
existence of a widespread ‘‘mirror system’’ in frontal,
parietal, and temporal regions (Gre`zes & Decety, 2002;
Buccino et al., 2001; Rizzolatti, Fogassi, & Gallese, 2001;
Iacoboni et al., 1999, 2001; Bonda, Petrides, Ostry, &
Evans, 1996; Grafton, Arbib, Fadiga, & Rizzolatti, 1996;
Rizzolatti et al., 1996). Così, it is plausible to retain that
during observation of action a diffuse neural network
subserving motor representation is already tuned to
imitate and learn the observed actions.
In this diffuse network, activation of the dorsolateral
prefrontal cortex (DLPFC) was shown during observation
of actions (Buccino et al., 2004). A recent functional mag-
netic resonance imaging study reported activation of the
DLPFC related to the self-selection of intended action
rather than the mere observation of action (Cunnington,
Windischberger, Robinson, & Moser, 2006). It was pro-
posed that the dorsolateral prefrontal areas could play
an orchestrating role in the processes that occur in the
mirror neuron system by selecting and recombining the
motor elements to be imitated (Iacoboni, 2005).
Other research underlines the additional
involve-
ment of bilateral cerebellar areas in mental tasks that
do not require overt movements. Examples are silent
word generation and counting, tennis training move-
menti, and the observation or imagination of grasping
movements (Battaglia et al., 2006; Kuhtz-Buschbeck
et al., 2003; Ross, Tkach, Ruggirei, Lieber, & Lapresto,
2003; Petersen, van Mier, Fiez, & Raichle, 1998; Thach,
1996, 1998; Decety & Ingvar, 1990). Positron emission
tomography data report cerebellar activation during
D 2007 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 19:10, pag. 1656–1663
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
T
w
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
N
T
io
C
/
UN
e
R
–
P
T
D
io
C
1
l
9
e
1
–
0
P
1
D
6
F
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
C
/
N
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
C
6
N
5
.
6
2
P
0
D
0
7
B
.
sì
1
G
9
tu
.
e
1
S
0
T
.
o
1
N
6
0
5
8
6
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
.
.
F
o
N
1
8
M
UN
sì
2
0
2
1
observation of meaningful and meaningless actions for
the purpose of imitation and, conversely, no cerebel-
lar activation during the mere observation of meaning-
less actions (Leslie, Johnson-Frey, & Grafton, 2004;
Chaminade, Meltzoff, & Decety, 2002; Gre`zes & Decety,
2001, 2002; Decety & Gre`zes, 1999; Decety et al., 1997).
Inoltre, experimental findings demonstrate that in
rats cerebellar lesions markedly impair the learning of
new procedures not only by actual execution of a task
(Leggio et al., 1999) but also by observation (Leggio et al.,
2000). Così, there are a number of indications that cere-
bellar circuits are involved in ‘‘motor thought’’ whether
they are accompanied by actual motor acts.
The aim of the present work was to analyze the role of
specific neural substrates in learning a sequential visuo-
spatial task by observation. Learning complex actions by
observation is not a unitary function because it requires
the acquisition of very different competencies. In partic-
ular, when a subject learns a sequential visuomotor task
by observation, they acquire both the specific sequence
of items and the procedural rules about how to perform
the task. In the present observational protocol, subjects
observed an actor detecting a sequence. Then, they ac-
tually performed the task producing the sequence they
had just viewed (old sequence) as well as a different se-
quence they had never seen (new sequence). Detection of
the sequences required putting into action the procedural
rules to solve the task (cioè., vertical or horizontal move-
ment was allowed but never diagonal movement) anche
as recalling from memory the series of correctly touched
items. Nevertheless, reproducing the old sequence mainly
required exploiting knowledge of the observed sequence,
whereas detecting the new sequence mainly required
exploiting procedural competencies linked to the rules
of the task.
The proposal that the cerebellar network is fully
engaged in the acquisition of the procedural compo-
nents of a task is consistent with the functions attributed
to it. As indicated by neuroimaging studies, cerebellar
activation is closely linked to the first phases of acquisi-
tion and decreases to lower levels after practice (Petersen
et al., 1998; Vaina, Belliveau, des Roziers, & Zeffiro,
1998), supporting the cerebellar role as a procedural
machine. Inoltre, during the implicit acquisition of a
sequential visuomotor task, cerebellar lesions induce a
procedural impairment that can be bypassed by exploit-
ing previously acquired declarative knowledge (Molinari
et al., 1997).
Conversely, the prefrontal circuits are primarily en-
gaged in the declarative components of a task. This is in
accordance with the well-known role of the frontal
cortex in encoding and retrieval of memory span (Rossi
et al., 2001, 2004) and in visuospatial working memory
(Koch et al., 2005; Hoshi et al., 2000).
By taking into account these different competencies
in learning by observation, we analyzed the interfering
effects evoked by low-frequency repetitive transcranial
magnetic stimulation (rTMS) on the cerebellum or
on the DLPFC in an observational visuomotor task. Fur-
thermore, given the crucial role of the cerebellar and
prefrontal networks in motor performance, it seemed
crucial to verify the effects of rTMS on actual execu-
tion of the same task. Because the rTMS technique
induces inhibition of the stimulated brain areas for sev-
eral minutes after the stimulation period (Oliveri, Koch,
Torriero, & Caltagirone, 2005), it was possible to distin-
guish the effects on the observation or actual execution
of the task.
METHODS
Subjects
Seventy-five right-handed healthy subjects (35 men and
40 women; mean age = 24.1 years) participated in the
experiments after providing written informed consent.
The Fondazione ‘‘Santa Lucia’’ ethical committee ap-
proved the study.
Experimental Setting
UN 10 (cid:1) 10 black matrix appeared on a computer touch
screen. The subjects were asked to find a hidden
sequence of ‘‘correct’’ squares arranged in advance by
the experimenter. The sequence was composed of 20 ad-
jacent spatial positions in the matrix, forming a com-
plex ‘‘snakelike’’ pattern without any semantically or
geometrically describable arrangement. The subjects
had to start by touching a gray square, representing
the first element of the sequence, which always re-
mained lit up. In the search for the second correct
position, they had to touch the black squares bordering
the gray square by moving vertically or horizontally, Ma
never diagonally, in the matrix. When the correct posi-
tion was discovered, the touched square turned gray;
conversely, when an incorrect position was touched, IL
square turned red. An alert sound signaled the touching
of an incorrect position. Each (correct or incorrect)
touched square was lit up for 500 msec and then became
black again; così, no trace of the performed sequence
remained on the screen. Subjects had to start the
performed sequence again each time they found a new
correct position in the matrix. They were helped by a
sound produced by the program that signaled them to
return to the first square of the sequence. In the search
for the following positions, they had to obey the same
rules. The task ended when the subjects found the 20th
correct position.
Experimental Protocol
Four different 20-item sequences, which did not differ
for degree of difficulty,1 were arranged. Subjects were
Torriero et al.
1657
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
T
w
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
N
T
io
C
/
UN
e
R
–
P
T
D
io
C
1
l
9
e
1
–
0
P
1
D
6
F
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
C
/
N
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
C
6
N
5
.
6
2
P
0
D
0
7
B
.
sì
1
G
9
tu
.
e
1
S
0
T
.
o
1
N
6
0
5
8
6
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
F
.
.
.
.
o
N
1
8
M
UN
sì
2
0
2
1
submitted to an observational training that consisted of
observing a subject (actor) while discovering one out of
four correct sequences by trial and error. Then, the ob-
servers were required to actually execute the task. They
executed both the previously observed ‘‘old’’ sequence
and a ‘‘new’’ sequence they had never observed. IL
presentation order between old and new sequences was
randomized and counterbalanced among subjects.
Repetitive Transcranial Magnetic Stimulation
rTMS was delivered by means of a MagStim rapid
magnetic stimulator, using a figure-eight coil (70 mm
in diameter) over the left lateral cerebellum and the
right DLPFC. For cerebellar stimulation, the coil was
positioned tangentially to the scalp, with the handle
pointing upward, 1 cm under and 3 cm to the left of the
inion (Torriero, Oliveri, Koch, Caltagirone, & Petrosini,
2004; Theoret, Haque, & Pascual-Leone, 2001). For the
DLPFC stimulation, the anterior end of the coil wings
junction was placed over the F4 site of the International
10/20 EEG system. The coil was applied tangentially to
the scalp surface with the handle directed backward and
angled at about 458 to the midline.
To verify the specificity of the effects of cerebellar and
prefrontal rTMS, a control region, presumably not in-
volved in the task, was stimulated. The coil was placed
over a mesial fronto-parietal site, corresponding to the
Cz scalp position of the 10/20 EEG system, with the
handle pointing backward.
Anatomical information about the brain areas stimulat-
ed was obtained by performing magnetic resonance im-
aging on a single experimental subject after marking the
cerebellar and frontal scalp sites with capsules containing
soy oil. Off-line rTMS was applied at 1-Hz frequency for
10 min (600 stimuli) at 90% intensity of the motor thresh-
old. The latter was defined as the lowest TMS intensity (COME
assessed with single-pulse TMS) able to induce a visible
muscle twitch of the contralateral hand in at least 50% Di
a sequence of 10 consecutive trials.
Experimental Groups
Seventy-five subjects were divided into seven groups.
The first group (actors, n = 20) was composed of actors
who served as models for the remaining groups of
observers. The second group (controls, n = 10) era
composed of subjects submitted to observational train-
ing before their own performance. These two groups
were not submitted to any rTMS. The subjects in the
following three groups were submitted to observational
training at the end of the rTMS train over the Cz (Cz +
Obs, n = 10), the left cerebellar hemisphere (Cb + Obs,
n = 11), or the right DLPFC (DLPFC + Obs, n = 10).
Immediately after the observation, they actually per-
formed the task. The subjects in the last two groups
were first submitted to observational training and im-
mediately afterward to an rTMS train over the left
cerebellar hemisphere (Obs + Cb, n = 7) or the right
DLPFC (Obs + DLPFC, n = 7). Subsequently, Essi
performed the task.
Data Analysis
The total number of incorrect positions (total errors)
touched on the screen in detecting the correct sequence
was analyzed. To further evaluate the qualitative aspects
of errors, the incorrect span, calculated as the maximum
number of consecutive touching of incorrect squares,
and the perseverative errors, calculated as the sum of
consecutive touching of the same square or of a fixed
sequence of squares, were analyzed.
Logarithmic transformations significantly improved
the distribution of total errors and incorrect span. Even
if the resulting distribution was not Gaussian (Kolmogorov-
Smirnov test, P < .001), the number of outliers was
strongly reduced, and good variance homogeneity was
obtained (before transformation, Levene’s test, p < .009;
after transformation, Levene’s test, p > .05).
Two-way analyses of variance (ANOVAs) with repeated
measures were used followed by multiple comparisons
with post hoc Duncan’s test.
RESULTS
To verify whether observational training influenced the
learning of the visuo-spatial task, we compared the per-
formances of the actors with the performances of ob-
server controls who executed the previously observed
(old) sequence and a never observed (new) sequence.
When the observers reproduced the previously ob-
served sequence, they made significantly fewer errors
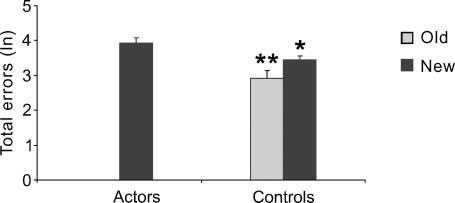
than the actors [one-way ANOVA: F(1,28) = 12.40, p =
.001]. È interessante notare, a beneficial effect of observational
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
T
w
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
N
T
io
C
/
UN
e
R
–
P
T
D
io
C
1
l
9
e
1
–
0
P
1
D
6
F
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
C
/
N
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
C
6
N
5
.
6
2
P
0
D
0
7
B
.
sì
1
G
9
tu
.
e
1
S
0
T
.
o
1
N
6
0
5
8
6
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
.
.
F
o
N
1
8
M
UN
sì
2
0
2
1
Figura 1. Behavioral results. Total errors are shown as natural
logarithms. Vertical bars indicate SEM. Gray columns = old
sequence; black columns = new sequence. Controls made
significantly fewer errors than actors in performing both new
and old sequence. *P < .05; **p < .001.
1658
Journal of Cognitive Neuroscience
Volume 19, Number 10
training was found when the observers performed a
never observed sequence [one-way ANOVA: F(1,28) =
4.19, p = .05] (Figure 1). In any case, by comparing
the performances of the observers in the old and new
sequences, a significant difference was found [one-way
ANOVA: F(1,9) = 6.21, p = .03]. These findings indicate
that observing an actor performing a sequential visuo-
motor task is advantageous in learning both the rules
and the specific sequence of items, even if the beneficial
effect is sequence specific.
A two-way ANOVA (3 groups (cid:1) 2 sequences) comparing
the performances of the Cb + Obs and DLPFC + Obs
groups against the (no-TMS) controls revealed highly sig-
nificant sequence [F(1,28) = 28.41, p = .00001] and in-
teraction effects [F(2,28) = 6.56, p = .004]. Post hoc
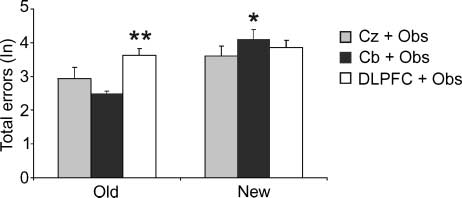
comparisons indicated that the Cb + Obs group made sig-
nificantly more errors than the controls in detecting the
new sequence ( p = .01), but performed comparably to
the controls in reproducing the old sequence ( p = .50).
On the contrary, the DLPFC + Obs group made signifi-
cantly more errors than the controls in reproducing the
old sequence ( p = .006) and performed comparably to
the controls in detecting the new sequence ( p = .11)
(Table 1).
To account for generic effects of rTMS, the perfor-
mances of the Cb + Obs and DLPFC + Obs groups were
compared with the performances of the Cz + Obs group.
A two-way ANOVA (3 groups (cid:1) 2 sequences) revealed
significant sequence [F(1,28) = 30.78, p = .00001] and
interaction effects [F(2,28) = 6.02, p = .007]. Once more,
post hoc comparisons indicated that the Cb + Obs group
made significantly more errors than the Cz + Obs group
in detecting the new sequence ( p = .05) but performed
comparably to the Cz + Obs group in reproducing the
old sequence ( p = .41). Again, the DLPFC + Obs group
made significantly more errors than the Cz + Obs group
in reproducing the old sequence ( p = .01) but per-
formed comparably to the Cz + Obs group in detecting
the new sequence ( p = .34) (Figure 2, Table 1).
Given the peculiar role played by prefrontal as well as
cerebellar regions in sequential behaviors (Hauser, 1999;
Schmahmann & Sherman, 1997), additional analyses were
performed on incorrect span and perseverations.
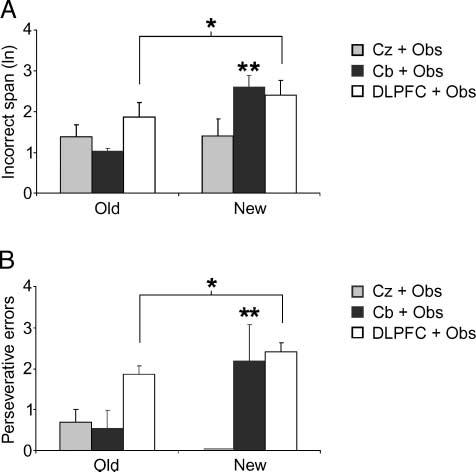
A two-way ANOVA (2 groups (cid:1) 2 sequences) compar-
ing the incorrect span of the Cz + Obs and Cb + Obs
groups showed significant sequence [F(1,19) = 7.07,
p = .01] and interaction [F(1,19) = 6.41, p = .02] effects.
Post hoc analyses revealed a significant difference be-
tween groups in detecting the new sequence ( p = .01).
Table 1. Number of Total Errors (Mean ± SEM )
Figure 2. Total errors in rTMS groups. Total errors are shown
as natural logarithms. Vertical bars indicate SEM. Gray columns =
Cz + Obs group; black columns = Cb + Obs; white columns =
DLPFC + Obs group. In comparison with rTMS of control site
(Cz + Obs), cerebellar rTMS (Cb + Obs) produced a selective
increase in total errors in performing the new sequence, whereas
DLPFC rTMS (DLPFC + Obs) produced an increase in total errors
in reproducing the old sequence. *p < .05; **p < .01.
A two-way ANOVA (2 groups (cid:1) 2 sequences) comparing
the Cz + Obs and DLPFC + Obs groups showed a signif-
icant group effect [F(1,18) = 5.44, p = .03] (Figure 3A).
A two-way ANOVA (2 groups (cid:1) 2 sequences) comparing
the perseverative errors of the Cz + Obs and Cb + Obs
groups showed a significant interaction effect [F(1,19) =
7.91, p = .01]. Post hoc analyses revealed a significant
difference between groups in detecting the new sequence
( p = .002). A two-way ANOVA (2 groups (cid:1) 2 sequences)
comparing the Cz + Obs and DLPFC + Obs groups
showed a significant group effect [F(1,18) = 6.60, p =
.02] (Figure 3B).
Finally, the effects of rTMS applied before the actual
execution of the task were analyzed to verify whether
the previously obtained results were because of influ-
ences on task execution rather than on observational
training. Thus, the Obs + Cb, Obs + DLPFC, and control
groups’ performances in the two sequences were com-
pared. A two-way ANOVA (3 groups (cid:1) 2 sequences)
showed a significant main effect of sequence [F(1,21) =
9.40, p = .005]. Once more, this result confirms that the
improving effect of observational training is sequence
specific. The ANOVA failed to reveal any significant
group or interaction effects (Table 1).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
0
p
1
d
6
f
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
c
/
n
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
c
6
n
5
.
6
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
0
t
.
o
1
n
6
0
5
8
6
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
DISCUSSION
The present research demonstrates that rTMS is able to
influence learning by observation of a visuospatial task by
exerting different effects on the various components of
o
n
1
8
M
a
y
2
0
2
1
Actors
63.5 ± 10.5
Old
New
Controls
Cz + Obs
Cb + Obs
DLPFC + Obs
Obs + Cb
Obs + DLPFC
30.0 ± 7.6
22.0 ± 4.8
18.2 ± 4.6
45.9 ± 10.3
30.8 ± 10.0
27.7 ± 7.3
35.21 ± 4.4
38.3 ± 4.3
89.8 ± 24.1
59.0 ± 13.1
34.2 ± 7.1
38.0 ± 16.1
Torriero et al.
1659
Schmahmann & Sherman, 1997; Appollonio, Grafman,
Schwartz, Massaquoi, & Hallett, 1993; Grafman et al.,
1992; el-Awar et al., 1991). The rTMS influence was spe-
cifically linked to the observation phase and was com-
pletely lacking in the execution phase. In fact, cerebellar
and prefrontal rTMS applied when the observational
training was over did not evoke any effect in reproducing
the old sequence or in detecting a new sequence.
The interfering effects of cerebellar rTMS on observa-
tional learning could be because of impairment in plan-
ning intentional strategies (i.e., in the ability to access
and effectively use a procedural rule already acquired or
in difficulty in acquiring a new one). Indeed, clinical re-
ports on cerebellar patients describe severe problems in
learning (Gomez-
cognitive planning and procedural
Beldarrain, Garcia-Monco, Rubio, & Pascual-Leone, 1998;
Molinari et al., 1997; Appollonio et al., 1993; Pascual-
Leone et al., 1993; Grafman et al., 1992). The present find-
ings allow extending to humans the experimentally dem-
onstrated notion that the cerebellum is involved in
learning a visuospatial task not only when acquisition is
achieved by actually executing a sequence of procedural
acts but also when procedure acquisition is achieved by
observation (Petrosini et al., 2003; Graziano et al., 2002).
The disruption of observational learning after cerebellar
rTMS demonstrates the need for functionally intact cere-
bellar structures in the critical phase of the observation/
execution matching process, supporting their crucial role
in the acquisition phases of observational learning. In fact,
the procedural competencies were fully maintained (and
then put into action) when cerebellar rTMS was performed
after observational training. This finding is in agreement
with neuroimaging evidence demonstrating high cere-
bellar activation during the acquisition phase of learning
and decreased cerebellar activation after prolonged prac-
tice (Petersen et al., 1998; Vaina et al., 1998).
The present results also support the role of the DLPFC
in learning by observation. In fact, DLPFC rTMS pro-
voked the worsening of performances mainly in repro-
ducing the old sequence and an increase in perseverative
errors and incorrect span in performing both sequences.
The interfering effect of DLPFC rTMS on both sequences
emphasizes the sequence-specific effect found when
rTMS was delivered over the cerebellar region. Many
studies report activation of the DLPFC when free selec-
tion between different response alternatives is required
and when there is a high attentional demand (Hadland,
Rushworth, Passingham, Jahanshahi, & Rothwell, 2001;
Jueptner et al., 1997). Moreover, it was recently demon-
strated that the DLPFC is specifically activated in con-
junction with the selection of an item from memory to
guide a response (Rowe, Toni, Josephs, Frackowiak, &
Passingham, 2000). In the present visuomotor task, re-
producing the old sequence required selection from
competing actions, retrieval from long-term memories
of the sequence of positions previously learned, and
on-line maintenance and updating in working memory
Figure 3. Incorrect span and perseverative errors. Incorrect span is
shown as a natural logarithm. Vertical bars indicate SEM. Gray
columns = Cz + Obs group; black columns = Cb + Obs; white
columns = DLPFC + Obs group. (A) Incorrect span was significantly
longer in detecting the new sequence after cerebellar rTMS (Cb + Obs)
in comparison with the span after control site rTMS (Cz + Obs);
DLPFC rTMS (DLPFC + Obs) provoked a lengthening of the incorrect
span, regardless of the kind of sequences performed. (B) Cb + Obs
showed more perseverative errors than Cz + Obs only in detecting
the new sequence, whereas DLPFC + Obs showed more perseverative
errors regardless of the kind of sequences performed. Note that
Cz + Obs group showed no perseverative errors at all in detecting
the new sequence. *p < .05; **p < .01.
the task depending on which neuronal structures are
inhibited. In fact, when the observational training fol-
lowed rTMS of the left lateral cerebellum, we found clear
interference with detection of a new sequence, different
from the previously observed one. Conversely, when
observational training followed rTMS of the right DLPFC,
we found interference with the reproduction of the old
previously observed sequence. The effect of rTMS re-
sulted in an increase in total errors and lengthening of
incorrect span and an enhancement of perseverative er-
rors. The remarkable result regarding perseverative er-
rors was found after cerebellar rTMS when the new
sequence had to be detected and after DLPFC rTMS
when both sequences were performed. Perseverations
are distinctive symptoms linked to prefrontal dysfunction
(Hauser, 1999), and interestingly, they are also elicited
by cerebellar damage (Mandolesi, Leggio, Graziano,
Neri, & Petrosini, 2001; Schmahmann & Sherman, 1997;
Botez-Marquard & Botez, 1993). This finding is in line
with the view that cerebellar lesions might provoke
‘‘frontal-like’’ cognitive deficits, supported by clinical re-
ports of severe problems in initiation/perseveration and
cognitive planning in cerebellar patients (Hauser, 1999;
1660
Journal of Cognitive Neuroscience
Volume 19, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
0
p
1
d
6
f
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
c
/
n
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
c
6
n
5
.
6
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
0
t
.
o
1
n
6
0
5
8
6
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
of the progressively touched items. All of these compe-
tencies have been attributed to the prefrontal circuits
(Mottaghy, 2006; Faw, 2003; Manes et al., 2002).
In conclusion, the present study provides new insights
on the neural circuitry involved in observational learn-
ing. It was recently suggested (Iacoboni, 2005; Buccino
et al., 2004) that imitative learning of novel actions is
supported by a core circuit for imitation comprising the
fronto-parietal mirror neuron system that interacts with
DLPFC and motor preparation areas. Other research un-
derlined the additional activation of bilateral cerebellar
areas when the observation of action has the goal of
imitation. Such cerebellar activation was not docu-
mented when observation was not aimed toward imita-
tion (Chaminade et al., 2002; Decety & Gre`zes, 1999).
The interplay between cerebellar and prefrontal areas,
supported also by their important anatomofunctional
connections (Middleton & Strick, 2001), allows for spec-
ulation that the cerebellum and prefrontal cortex inter-
act in planning actions—that is, the former permits
acquisition of efficient procedural competencies and
the latter provides flexibility among already acquired
and stored solutions (Pochon et al., 2001; Spence,
Hirsch, Brooks, & Grasby, 1998; Hyder et al., 1997; Frith,
Friston, Liddle, & Frackowiak, 1991).
Acknowledgments
We thank Dr. Laura Serra for magnetic resonance imaging-
guided localization of target sites.
Reprint requests should be sent to Sara Torriero, Laboratorio
di Neurologia Clinica e Comportamentale, IRCCS Fondazione
‘‘Santa Lucia,’’ via Ardeatina 306, 00178 Rome, Italy, or via
e-mail: s.torriero@libero.it.
Note
1. The number of errors made by four groups of five sub-
jects in detecting one of the four sequences was calculated. A
one-way ANOVA with sequence as main effect failed to reveal
any significant difference in number of errors among groups
[F(3,16) = 0.29, p = .83], demonstrating the same degree of
difficulty in the four sequences.
REFERENCES
Appollonio, I. M., Grafman, J., Schwartz, V., Massaquoi, S.,
& Hallett, M. (1993). Memory in patients with cerebellar
degeneration. Neurology, 43, 1536–1544.
Battaglia, F., Quartarone, A., Ghilardi, M. F., Dattola, R.,
Bagnato, S., Rizzo, V., et al. (2006). Unilateral cerebellar
stroke disrupts movement preparation and motor imagery.
Clinical Neurophysiology, 117, 1009–1016.
Bonda, E., Petrides, M., Ostry, D., & Evans, A. (1996).
Specific involvement of human parietal systems and the
amygdala in the perception of biological motion. Journal
of Neuroscience, 1, 3737–3744.
Botez-Marquard, T., & Botez, M. I. (1993). Cognitive behavior
in heredodegenerative ataxias. European Neurology, 33,
351–357.
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L.,
Gallese, V., et al. (2001). Action observation activates
premotor and parietal areas in a somatotopic manner:
An fMRI study. European Journal of Neuroscience, 13,
400–404.
Buccino, G., Vogt, S., Ritzl, A., Fink, G. R., Zilles, K., Freund,
H. J., et al. (2004). Neural circuits underlying imitation
learning of hand actions: An event-related fMRI study.
Neuron, 42, 323–334.
Byrne, R. W., & Russon, A. E. (1998). Learning by imitation:
A hierarchical approach. Behavioral and Brain Sciences,
21, 667–684.
Chaminade, T., Meltzoff, A. N., & Decety, J. (2002). Does
the end justify the means? A PET exploration of the
mechanisms involved in human imitation. Neuroimage,
15, 318–328.
Cunnington, R., Windischberger, C., Robinson, S., &
Moser, E. (2006). The selection of intended actions and
the observation of others’ actions: A time-resolved fMRI
study. Neuroimage, 29, 1294–1302.
Decety, J., & Gre`zes, J. (1999). Neural mechanisms subserving
the perception of human actions. Trends in Cognitive
Sciences, 3, 172–178.
Decety, J., Gre`zes, J., Costes, N., Perani, D., Jeannerod, M.,
Procyk, E., et al. (1997). Brain activity during observation
of actions. Influence of action content and subject’s
strategy. Brain, 120, 1763–1777.
Decety, J., & Ingvar, D. H. (1990). Brain structures
participating in mental simulation of motor behavior:
A neuropsychological interpretation. Acta Psychologica,
73, 13–34.
El-Awar, M., Kish, S., Oscar-Berman, M., Robitaille, Y., Schut, L.,
& Freedman, M. (1991). Selective delayed alternation
deficits in dominantly inherited olivopontocerebellar
atrophy. Brain and Cognition, 16, 121–129.
Faw, B. (2003). Pre-frontal executive committee for perception,
working memory, attention, long-term memory, motor
control, and thinking: A tutorial review. Consciousness
and Cognition, 12, 83–139.
Frith, C. D., Friston, K., Liddle, P. F., & Frackowiak, R. S.
(1991). Willed action and the prefrontal cortex in man:
A study with PET. Proceedings of the Royal Society of
London, Series B, Biological Sciences, 244, 241–246.
Gomez-Beldarrain, M., Garcia-Monco, J. C., Rubio, B., &
Pascual-Leone, A. (1998). Effect of focal cerebellar lesion
on procedural learning in the serial reaction time task.
Experimental Brain Research, 120, 25–30.
Grafman, J., Litvan, I., Massaquoi, S., Stewart, M., Sirigu, A.,
& Hallett, M. (1992). Cognitive planning deficit in patients
with cerebellar atrophy. Neurology, 42, 493–1496.
Grafton, S. T., Arbib, M. A., Fadiga, L., & Rizzolatti, G.
(1996). Localization of grasp representations in humans
by positron emission tomography. 2. Observation compared
with imagination. Experimental Brain Research, 112,
103–111.
Graziano, A., Leggio, G. M., Mandolesi, L., Neri, P.,
Molinari, M., & Petrosini, L. (2002). Learning power
of single behavioral units in acquisition of a complex
spatial behavior: An observational learning study in
cerebellar-lesioned rats. Behavioral Neuroscience, 116,
116–125.
Gre`zes, J., & Decety, J. (2001). Functional anatomy of
execution, mental simulation, observation, and verb
generation of actions: A meta-analysis. Human Brain
Mapping, 12, 1–19.
Gre`zes, J., & Decety, J. (2002). Does visual object afford
action? Evidence from a neuroimaging study.
Neuropsychologia, 40, 212–222.
Torriero et al.
1661
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
0
p
1
d
6
f
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
c
/
n
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
c
6
n
5
.
6
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
0
t
.
o
1
n
6
0
5
8
6
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
Hadland, K. A., Rushworth, M. F. S., Passingham, R. E.,
Jahanshahi, M., & Rothwell, J. C. (2001). Interference
with performance of a response selection task that has
no working memory component: An rTMS comparison
of the dorsolateral prefrontal and medial frontal cortex.
Journal of Cognitive Neuroscience, 13, 1097–1108.
Hauser, M. D. (1999). Perseveration, inhibition and the
prefrontal cortex: A new look. Current Opinion in
Neurobiology, 9, 214–222.
Hoshi, Y., Oda, I., Wada, Y., Ito, Y., Yutaka, Y., Oda, M.,
et al. (2000). Visuospatial imagery is a fruitful strategy for
the digit span backward task: A study with near-infrared
optical tomography. Brain Research, Cognitive Brain
Research, 9, 339–342.
Middleton, F. A., & Strick, P. L. (2001). Cerebellar projections
to the prefrontal cortex of the primate. Journal of
Neuroscience, 15, 700–712.
Molinari, M., Leggio, M. G., Solida, A., Corra, R., Misciagna, S.,
Silveri, M. C., et al. (1997). Cerebellum and procedural
learning: Evidence from focal cerebellar lesions. Brain,
120, 1753–1762.
Mottaghy, F. M. (2006). Interfering with working memory in
humans. Neuroscience, 139, 85–90.
Oliveri, M., Koch, G., Torriero, S., & Caltagirone, C. (2005).
Increased facilitation of the primary motor cortex following
1 Hz repetitive transcranial magnetic stimulation of the
contralateral cerebellum in normal humans. Neuroscience
Letters, 16, 188–193.
Hyder, F., Phelps, E. A., Wiggins, C. J., Labar, K. S., Blamire,
Pascual-Leone, A., Grafman, J., Clark, K., Stewart, M.,
A. M., & Shulman, R. G. (1997). ‘‘Willed action’’: A functional
MRI study of the human prefrontal cortex during a
sensorimotor task. Proceedings of the National Academy
of Sciences, U.S.A., 94, 6989–6994.
Iacoboni, M. (2005). Neural mechanisms of imitation.
Current Opinion in Neurobiology, 15, 632–637.
Iacoboni, M., Koski, L. M., Brass, M., Bekkering, H., Woods,
R. P., Dubeau, M. C., et al. (2001). Reafferent copies of
imitated actions in the right superior temporal cortex.
Proceedings of the National Academy of Sciences, U.S.A.,
20, 13995–13999.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta,
J. C., & Rizzolatti, G. (1999). Cortical mechanisms of
human imitation. Science, 24, 2526–2528.
Jueptner, M., Stephan, K. M., Frith, C. D., Brooks, D. J.,
Frackowiak, R. S. J., & Passingham, R. E. (1997). Anatomy
of motor learning: I. Frontal cortex and attention to
action. Neurophysiology, 77, 1313–1324.
Koch, G., Oliveri, M., Torriero, S., Carlesimo, G. A., Turriziani,
P., & Caltagirone, C. (2005). rTMS evidence of different
delay and decision processes in a fronto-parietal neuronal
network activated during spatial working memory.
Neuroimage, 24, 34–39.
Kuhtz-Buschbeck, J. P., Mahnkopf, C., Holzknecht, C., Siebner,
H., Ulmer, S., & Jansen, O. (2003). Effector-independent
representations of simple and complex imagined finger
movements: A combined fMRI and TMS study. European
Journal of Neuroscience, 18, 3375–3387.
Leggio, M. G., Molinari, M., Neri, P., Graziano, A., Mandolesi, L.,
& Petrosini, L. (2000). Representation of actions in rats:
The role of cerebellum in learning spatial performances
by observation. Proceedings of the National Academy of
Sciences, U.S.A., 97, 2320–2325.
Leggio, M. G., Neri, P., Graziano, A., Mandolesi, L., Molinari, M.,
& Petrosini, L. (1999). Cerebellar contribution to spatial
event processing: Characterization of procedural learning.
Experimental Brain Research, 127, 1–11.
Leslie, K. R., Johnson-Frey, S. H., & Grafton, S. T.
(2004). Functional imaging of face and hand imitation:
Towards a motor theory of empathy. Neuroimage, 21,
601–607.
Mandolesi, L., Leggio, M. G., Graziano, A., Neri, P., &
Petrosini, L. (2001). Cerebellar contribution to spatial
event processing: Involvement in procedural and working
memory components. European Journal of Neuroscience,
14, 2011–2022.
Manes, F., Sahakian, B., Clark, L., Rogers, R., Antoun, N.,
Aitken, M., et al. (2002). Decision-making processes
following damage to the prefrontal cortex. Brain, 125,
624–639.
Meltzoff, A. N. (1999). Origins of the theory of mind,
cognition and communication. Journal of Communication
Disorders, 32, 251–269.
Massaquoi, S., Lou, J. S., et al. (1993). Procedural learning
in Parkinson’s disease and cerebellar degeneration. Annals
of Neurology, 34, 594–602.
Petersen, S. E., van Mier, H., Fiez, J. A., & Raichle, M. E. (1998).
The effects of practice on the functional anatomy of task
performance. Proceedings of the National Academy of
Sciences, U.S.A., 3, 853–860.
Petrosini, L., Graziano, A., Mandolesi, L., Neri, P., Molinari, M.,
& Leggio, M. G. (2003). Watch how to do it! New advances
in learning by observation. Brain Research, Brain
Research Reviews, 42, 252–264.
Pochon, J. B., Levy, R., Poline, J. B., Crozier, S., Lehericy, S.,
Pillon, B., et al. (2001). The role of dorsolateral prefrontal
cortex in the preparation of forthcoming actions: An fMRI
study. Cerebral Cortex, 11, 260–266.
Rizzolatti, G., Fadiga, L., Matelli, M., Bettinardi, V.,
Paulesu, E., Perani, D., et al. (1996). Localization of
grasp representations in humans by PET: 1 Observation
versus execution. Experimental Brain Research, 111,
246–252.
Rizzolatti, G., Fogassi, L., & Gallese, V. (2001). Neurophysiological
mechanisms underlying the understanding and imitation
of action. Nature Reviews Neuroscience, 2, 661–670.
Ross, J. S., Tkach, J., Ruggirei, P. M., Lieber, M., & Lapresto, E.
(2003). The mind’s eye: Functional MR imaging evaluation of
golf motor imagery. AJNR. American Journal of
Neuroradiology, 24, 1036–1044.
Rossi, S., Cappa, S. F., Babiloni, C., Pasqualetti, P., Miniussi, C.,
Carducci, F., et al. (2001). Prefrontal cortex in long-term
memory: An ‘‘interference’’ approach using magnetic
stimulation. Nature Neuroscience, 4, 948–952.
Rossi, S., Miniussi, C., Pasqualetti, P., Babiloni, C., Rossini,
P. M., & Cappa, S. F. (2004). Age-related functional
changes of prefrontal cortex in long-term memory:
A repetitive transcranial magnetic stimulation study.
Journal of Neuroscience, 24, 7939–7944.
Rowe, J. B., Toni, I., Josephs, O., Frackowiak, R. S., &
Passingham, R. E. (2000). The prefrontal cortex: Response
selection or maintenance within working memory?
Science, 288, 1656–1660.
Schmahmann, J. D., & Sherman, J. C. (1997). Cerebellar
cognitive affective syndrome. International Review of
Neurobiology, 41, 433–440.
Spence, S. A., Hirsch, S. R., Brooks, D. J., & Grasby, P. M. (1998).
Prefrontal cortex activity in people with schizophrenia
and control subjects. Evidence from positron emission
tomography for remission of ‘‘hypofrontality’’ with
recovery from acute schizophrenia. British Journal of
Psychiatry, 172, 316–323.
Thach, W. T. (1996). On the specific role of the cerebellum
in motor learning and cognition: Clues from PET activation
and lesion studies in man. Behavioral and Brain Sciences,
19, 411–431.
1662
Journal of Cognitive Neuroscience
Volume 19, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
0
p
1
d
6
f
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
c
/
n
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
c
6
n
5
.
6
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
0
t
.
o
1
n
6
0
5
8
6
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Thach, W. T. (1998). What is the role of the cerebellum in
motor learning and cognition? Trends in Cognitive Sciences,
2, 331–337.
Theoret, H., Haque, J., & Pascual-Leone, A. (2001).
Increased variability of paced finger tapping accuracy
following repetitive magnetic stimulation of the
cerebellum in humans. Neuroscience Letters, 306,
29–32.
Torriero, S., Oliveri, M., Koch, G., Caltagirone, C., &
Petrosini, L. (2004). Interference of left and right
cerebellar rTMS with procedural learning. Journal
of Cognitive Neuroscience, 16, 1605–1611.
Vaina, L. M., Belliveau, J. W., des Roziers, E. B., & Zeffiro, T. A.
(1998). Neural systems underlying learning and
representation of global motion. Proceedings of the
National Academy of Sciences, U.S.A., 13, 12657–12662.
Van Schie, H. T., Mars, R. B., Coles, M. G., & Bekkering, H.
(2004). Modulation of activity in medial frontal and motor
cortices during error observation. Nature Neuroscience,
7, 549–554.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
0
p
1
d
6
f
5
/
6
1
9
1
/
9
1
3
6
0
1
/
0
1
2
6
5
o
6
c
/
n
1
2
7
0
5
0
6
7
4
1
3
9
0
/
1
j
0
o
1
c
6
n
5
.
6
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
0
t
.
o
1
n
6
0
5
8
6
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Torriero et al.
1663