The Neural Time Course of Semantic Ambiguity
Resolution in Speech Comprehension
Lucy J. MacGregor1, Jennifer M. Rodd2, Rebecca A. Gilbert1,
Olaf Hauk1, Ediz Sohoglu1,3, and Matthew H. Davis1
Astratto
■ Semantically ambiguous words challenge speech compre-
hension, particularly when listeners must select a less frequent
(subordinate) meaning at disambiguation. Using combined mag-
netoencephalography (MEG) and EEG, we measured neural re-
sponses associated with distinct cognitive operations during
semantic ambiguity resolution in spoken sentences: (io) initial acti-
vation and selection of meanings in response to an ambiguous
word and (ii) sentence reinterpretation in response to subsequent
disambiguation to a subordinate meaning. Ambiguous words
elicited an increased neural response approximately 400–800 msec
after their acoustic offset compared with unambiguous control
words in left frontotemporal MEG sensors, corresponding to
sources in bilateral frontotemporal brain regions. This response
may reflect increased demands on processes by which multiple
alternative meanings are activated and maintained until later
selection. Disambiguating words heard after an ambiguous word
were associated with marginally increased neural activity over
bilateral temporal MEG sensors and a central cluster of EEG elec-
trodes, which localized to similar bilateral frontal and left tem-
poral regions. This later neural response may reflect effortful
semantic integration or elicitation of prediction errors that guide
reinterpretation of previously selected word meanings. Across
participants, the amplitude of the ambiguity response showed
a marginal positive correlation with comprehension scores, sug-
gesting that sentence comprehension benefits from additional
processing around the time of an ambiguous word. Better com-
prehenders may have increased availability of subordinate mean-
ing, perhaps due to higher quality lexical representations and
reflected in a positive correlation between vocabulary size and
comprehension success. ■
INTRODUCTION
Most common words are semantically ambiguous (for a re-
view, see Rodd, Gaskell, & Marslen-Wilson, 2002), come
that their meaning depends on context. Per esempio,
“ace” can refer to a playing card or a tennis serve that an
opponent is unable to return. Così, the ability to make
sense of—resolve—ambiguity is a fundamental part of
speech comprehension. When listeners (or readers) en-
counter an ambiguous word (per esempio., “ace”), semantic prim-
ing studies suggest that they automatically activate the
multiple meanings of that word in parallel (irrespective
of context) Ma, within a few hundred milliseconds, settle
on a single preferred meaning (Seidenberg, Tanenhaus,
Leiman, & Bienkowski, 1982; Swinney, 1979). Initial mean-
ing selection operates on the information available at
that time (Cai et al., 2017; Rodd, Cutrin, Kirsch, Millar,
& Davis, 2013; Duffy, Morris, & Rayner, 1988; for a review,
see Vitello & Rodd, 2015), which will be particularly chal-
lenging if disambiguating context is absent or delayed
until after the ambiguous word. If a subsequent context
supports a subordinate (less frequent, thus more unex-
pected) Senso, then a later process of reinterpretation
is often necessary for accurate comprehension.
1University of Cambridge, 2University College London, 3Università
of Sussex
Individual differences in comprehension success have
been associated with abilities at accessing, selecting, E
reinterpreting ambiguous word meanings (Henderson,
Snowling, & Clarke, 2013; Szabo Wankoff & Cairns, 2009;
Gernsbacher, Varner, & Faust, 1990). Damage to the anteri-
or temporal lobe, a region known to be associated with se-
mantic processing in general (Patterson, Nestor, & Rogers,
2007), has been shown to impair the processing of ambigu-
ous word meanings (Zaidel, Zaidel, Oxbury, & Oxbury,
1995), but it is still unclear how variation in comprehension
ability relates to variation in the associated neural processes.
The aim of the current study is to understand the neural
mechanisms that support two stages of successful ambiguity
resolution (initial meaning activation/selection and sub-
sequent reinterpretation) and to explore the relationship
between behavioral and neural responses to ambiguity.
The cortical network supporting ambiguity resolution in
sentences was first reported in a fMRI study by Rodd, Davis,
and Johnsrude (2005). Listeners were presented with high-
ambiguity sentences containing multiple ambiguities (per esempio.,
“there were DATES and PEARS on the kitchen table”), E
the associated BOLD activation was contrasted with that
produced by low-ambiguity control sentences (per esempio., “there
was beer and cider on the kitchen shelf”). Additional activa-
tion during comprehension of high-ambiguity sentences
was observed in bilateral inferior frontal gyrus (IFG),
© 2019 Istituto di Tecnologia del Massachussetts. Published under a
Creative Commons Attribution 4.0 Internazionale (CC BY 4.0) licenza.
Journal of Cognitive Neuroscience 32:3, pag. 403–425
https://doi.org/10.1162/jocn_a_01493
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
T
F
.
o
N
0
5
M
UN
sì
2
0
2
1
particularly in pars triangularis and opercularis, and in left
posterior temporal regions, including posterior middle tem-
poral gyrus (pMTG), posterior inferior temporal gyrus
(pITG), and fusiform. These activations were observed in
the absence of explicit awareness of the ambiguities and
when listeners were given no explicit task, suggesting in-
volvement of these regions when comprehension occurs
automatically as in natural speech comprehension. This ba-
sic observation that semantic ambiguity resolution involves
frontotemporal regions is now well established, having been
replicated using fMRI for spoken (Vitello, Warren, Devlin, &
Rodd, 2014; Rodd, Johnsrude, & Davis, 2012; Tahmasebi
et al., 2012; Rodd, Longe, Randall, & Tyler, 2010) and written
(Mason & Just, 2007; Zempleni, Renken, Hoeks, Hoogduin,
& Stowe, 2007) sentences and shown to have a consistent
localization across individuals (Vitello et al., 2014). This fron-
totemporal response to ambiguity has proven useful in
translational work, Per esempio, as a neural marker of
residual semantic processing of speech at different levels
of sedation (Davis et al., 2007) and as evidence for intact
speech comprehension, which has prognostic value for
patients diagnosed as being in a vegetative state (Coleman
et al., 2007, 2009).
Tuttavia, attempts to attribute specific cognitive oper-
ations like initial meaning activation/selection and sub-
sequent reinterpretation to distinct cortical regions have
been less successful. One experimental approach has been
to compare neural responses to sentences containing am-
biguous words with varying meaning frequencies; come
sentences are expected to load on different processes in
ambiguity resolution. Per esempio, initial meaning selec-
tion is assumed to be more difficult for sentences contain-
ing ambiguous words with meanings that have similar
frequencies (balanced) than for words with a more dom-
inant meaning (biased). Conversely, reinterpretation is as-
sumed to be more difficult or more likely when sentences
are disambiguated to a subordinate (less frequent and
therefore less expected) Senso. In this way, BOLD re-
sponses due to differences in meaning frequency can be
related to processes at the time of ambiguity (initial mean-
ing activation/selection) or disambiguation (subsequent
reinterpretation). Using this approach, responses to sub-
ordinate meanings have been attributed to reinterpreta-
tion processes in the left (Vitello et al., 2014) or bilateral
(Mason & Just, 2007; Zempleni et al., 2007) IFG, some-
times extending into superior and middle frontal areas
(Mason & Just, 2007). Tuttavia, pMTG/pITG has also
been implicated in reinterpretation, with studies observ-
ing greater activation for subordinate meanings in left
( Vitello et al., 2014) or bilateral (Zempleni et al., 2007)
posterior temporal regions, though null results are also
reported (Mason & Just, 2007). Initial meaning selection
has also been associated with responses in the left IFG
(Mason & Just, 2007), but other studies have failed to
observe greater activation for balanced compared with
biased ambiguous words, and hence, evidence for selec-
tion processes is currently lacking ( Vitello et al., 2014).
An alternative approach to separating neural responses
during initial meaning selection from those involved in
subsequent reinterpretation has explored differences used
in the timing of frontotemporal responses. Rodd et al.
(2012) used a rapid fMRI acquisition sequence to measure
the time course of the BOLD response to ambiguous sen-
tences in which the timing of disambiguation was varied.
They assumed that additional BOLD responses associated
with reinterpretation (relative to unambiguous control
sentences) would occur later for ambiguous sentences in
which disambiguation occurred after an additional delay.
Hence, they contrasted delayed disambiguation sentences,
like “The ecologist thought that the PLANT by the river
should be closed down” with immediate disambiguation
sentences, like “The scientist thought that the FILM on
the water was from the pollution” (AMBIGUOUS and
disambiguation words highlighted). BOLD responses to
immediate and delayed ambiguity resolution showed
differences in timing in the left IFG and in posterior tem-
poral areas (fusiform, pITG, and pMTG) consistent with
reinterpretation. Inoltre, BOLD responses were also
observed in the IFG for sentences in which the disambig-
uating information occurred before the ambiguous word
(“The hunter thought that the HARE in the field was actu-
ally a rabbit”). Because these sentences should not require
reinterpretation Rodd and colleagues concluded that the
IFG is also involved in meaning selection.
Taken together, an emerging picture of the differential
contribution of inferior frontal and posterior temporal
brain regions to semantic ambiguity resolution is that
meaning selection may be underpinned by IFG and rein-
terpretation by IFG and posterior temporal areas together.
Tuttavia, there is a lack of consistent findings in relevant
experiments perhaps due to the challenge of associating
a slow BOLD response, which has a rise time of around
5 sec ( Josephs & Henson, 1999; Boynton, Engel, Glover,
& Heeger, 1996), with distinct neurocognitive processes
that operate over a shorter time period. This leads to
two problems. Primo, during the comprehension of a single
sentence lasting less than 5 sec, the measured BOLD re-
sponse to different neurocognitive events will inevitably
sovrapposizione, making it difficult to tease apart initial meaning
activation/selection and subsequent reinterpretation.
Secondo, given that meaning selection is thought to occur
within a few hundred milliseconds (Seidenberg et al., 1982;
Swinney, 1979), the associated neural response may be
transient and not detected in the BOLD signal.
Several studies have utilized more temporally sensitive
measures of cognition to investigate the processing of
ambiguous words. During natural reading, the duration
of fixation times have been shown to be longer for ambig-
uous words in the absence of biasing context compared
with unambiguous controls (Frazier & Rayner, 1990; al-
though for evidence that reading times for ambiguous
words with biased meanings do not differ from unambig-
uous controls, see Duffy et al., 1988; Rayner & Duffy,
1986). ERP studies with word-by-word presentation have
404
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
.
/
F
o
N
0
5
M
UN
sì
2
0
2
1
shown a sustained frontal negativity for ambiguous words
presented in a semantically neutral context compared with
unambiguous words (Hagoort & Brown, 1994) and for
ambiguous words in a semantically neutral but syntactically
constraining context compared with unambiguous controls
(Lee & Federmeier, 2006, 2009, 2012; Federmeier, Segal,
Lombrozo, & Kutas, 2000). These findings suggest that
processing of ambiguous words is more effortful than pro-
cessing words with single meanings. ERP studies using
word-by-word visual presentation have also looked for ef-
fects potentially associated with reinterpretation (Gunter,
Wagner, & Friederici, 2003; Hagoort & Brown, 1994). In
these studies, N400 responses have been observed in re-
sponse to disambiguating words that resolve an ambiguity
to its subordinate meaning. Tuttavia, these studies did not
control for both the presence/absence of ambiguity and the
word form itself. Hence, differences in word form and
meaning might also be responsible for these neural effects.
In this study, we used combined magnetoencephalogra-
phy (MEG) and EEG, which provides the temporal resolu-
tion required to distinguish neural responses at different
time points during sentences and to relate these responses
to distinct neurocognitive processes. Our volunteers lis-
tened to spoken sentences (see Figure 1A) that manipulated
the presence/absence of an ambiguous word (AMBIGUITY)
and subsequent disambiguation (disambiguation, per esempio.,
“The man thought that one more ACE/SPRINT might be
enough to win the tennis/game.”). These sets of sentences
enable us to specify conditions and time points in which we
expect either initial meaning access and selection or reinter-
pretation to occur.
In the absence of biasing context, an ambiguous word
(ACE) should require additional meaning access and se-
lection processes relative to a matched, unambiguous
control word (SPRINT). This comparison of sentences
with and without an ambiguous word (cioè., the main ef-
fect of ambiguity) provides the first experimental contrast
in our study. Neural activity during and after the ambig-
uous word will reflect processes involved in initial mean-
ing activation and selection that are more strongly taxed
by ambiguous than control (unambiguous) parole. These
processes should occur before subsequent context words
that drive reinterpretation.
Given that the words that precede the ambiguous
word are relatively uninformative, initial meaning access
and selection should result in most listeners settling on
the dominant (playing card) meaning of the ambiguous
word. The subsequent presentation of a sentence-final
word (tennis) that is incompatible with the dominant
meaning of ACE disambiguates the ambiguous word
to its subordinate meaning. For listeners to avoid mis-
interpretation, resource-demanding reinterpretation
processes should be triggered by the sentence-final word
(tennis) but not by an alternative final word (game) Quello
is consistent with both meanings (Rodd, Johnsrude, &
Davis, 2010; Kambe, Rayner, & Duffy, 2001; Duffy et al.,
1988). Because this reinterpretation process will only
occur if the sentence-final word (tennis) occurs in a sen-
tence that contains the ambiguous word (ACE), the neu-
ral correlates of reinterpretation can be detected using
the interaction between ambiguous words and subordi-
nate reintererptation, time-locked to the sentence-final
word.
For both the meaning access/selection (main effect) E
reinterpretation (interaction) contrasts, we measured
evoked MEG/EEG responses relative to the offset of the
critical words. This is a time point at which listeners have
heard sufficient phonetic information to recognize the
words and are therefore engaged in processing meaning.
We used an active comprehension task on noncritical trials
(relatedness judgment) during MEG/EEG scanning to en-
sure attentive listening throughout without contaminating
neural measures obtained during critical trials.
In addition to our analyses of main effects and inter-
actions, we were also interested in relating neural responses
to individual differences in sentence comprehension. Noi
therefore administered a postscanning behavioral task to
provide a trial-by-trial measure of the comprehension of
critical sentences that required reinterpretation of an
ambiguous word. We were interested in whether more
successful ambiguity resolution would be associated with
greater neural engagement or reduced processing effort
at the time of ambiguity or reinterpretation. We were also
interested in whether there was a relationship between
comprehension and verbal and nonverbal abilities (as mea-
sured using standard vocabulary and fluid reasoning tests).
METHODS
Stimuli
Sets of 80 spoken sentences were constructed according
ad a 2 × 2 factorial design in which we manipulated (1)
the presence/absence of an ambiguous word (ambiguity:
ambiguous vs. controllo) E (2) the presence of one of
two sentence-final words, which in the ambiguous sen-
tences either disambiguated the ambiguous word so it re-
solved to a subordinate meaning or left it unresolved
(disambiguation: resolved vs. unresolved). Because iden-
tical sentence-final words also completed the unambigu-
ous control sentences, we also use the terms resolved/
unresolved to refer to the equivalent control conditions
(see Figure 1A and Table 1). Ambiguous words occurred
midsentence after a neutral context that did not bias in-
terpretation toward either meaning of the ambiguous
word (mean word offset of 1423 msec after sentence
onset; see Figure 1B) and were followed by additional
neutral context words. In the “ambiguous-resolved” sen-
tences, the sentence-final word disambiguated the am-
biguous word toward a subordinate meaning (mean
word onset and offset were 1068 E 1506 msec after
the offset of the ambiguous word; see Figure 1B). In
the “ambiguous-unresolved” sentences, the sentence-
final word was necessarily more general so that both
MacGregor et al.
405
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
T
.
o
N
0
5
M
UN
sì
2
0
2
1
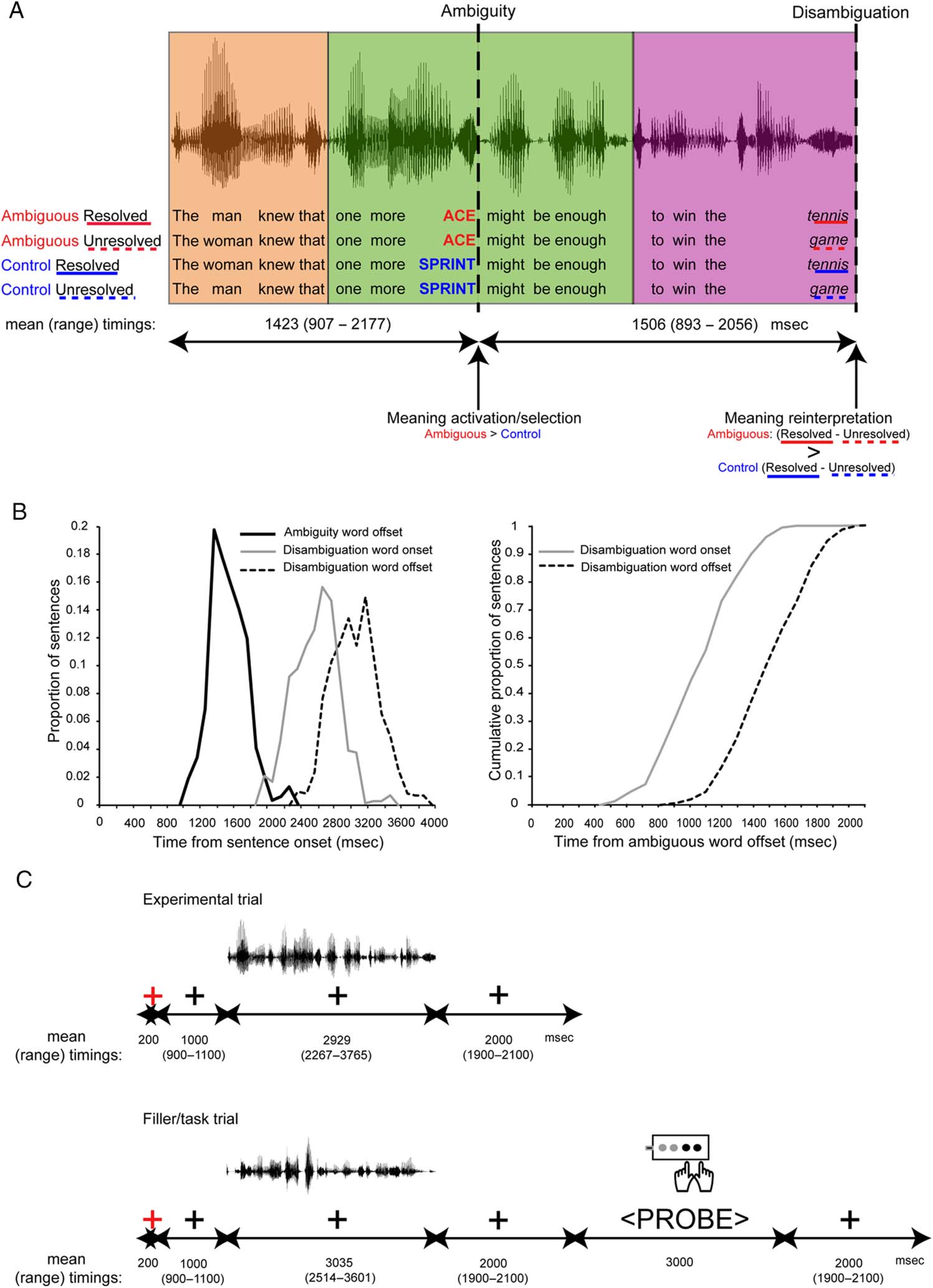
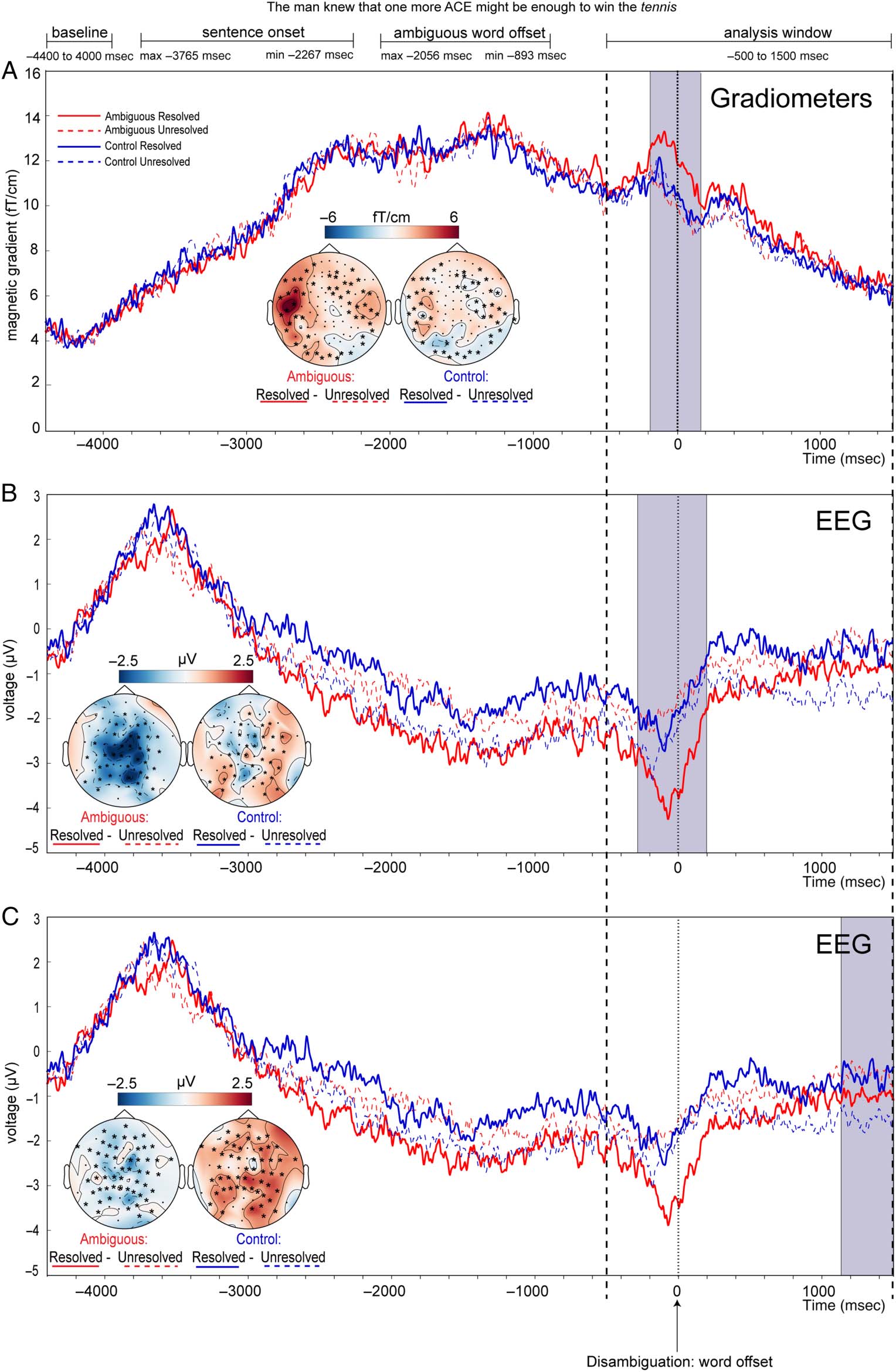
Figura 1. Stimulus and
experimental examples and
timings. (UN) Example quartet of
the spoken sentence stimuli
showing the four experimental
conditions, which were designed
to investigate neural processes
occurring at two critical time
points during semantic ambiguity
resolution: (1) ambiguity and (2)
disambiguation. MEG responses
were measured time-locked to
the offsets of critical words. Al
time of ambiguity, responses to
ambiguous words (red) were
predicted to be larger than
unambiguous control words
(blue), reflecting more effortful
semantic selection processes.
At the time of disambiguation,
responses to disambiguating
words that resolved the
ambiguity to a subordinate
Senso (red solid underline)
were predicted to be larger than
control words that left the
ambiguity unresolved (red dotted
underline) and control words that
completed the unambiguous
sentence with each of the words
used in the ambiguous sentences
(blue solid/dotted underline),
reflecting the greater probability
of reinterpretation processes.
Each sentence was combined
from three fragments
(highlighted with background
colour) from different recordings
such that linguistically identical
fragments were acoustically
identical across conditions, E
so that the splice points occurred
at least one word before and one
word after the ambiguous/control
word. (B) Frequency distributions
of the time durations (msec)
between critical words at
ambiguity and at disambiguation,
shown as proportions across all
640 sentences (cioè., Tutto
conditions). Durations are
categorized into 100-msec time
bins. The left panel displays the distribution of timings of ambiguity word offsets and of disambiguation word onsets and offsets relative to sentence
onsets. The right panel shows the cumulative distribution of timings of the onsets and offsets of the disambiguation words relative to the ambiguity
word offsets. The offsets of the disambiguation words occur more than 800 msec after ambiguity word offset for all sentences (cioè., at a time beyond the
duration of the analysis window for the ambiguity words), and the onsets of the disambiguation words occur more than 800 msec after ambiguity word
offsets for 81% of sentences. (C) Structure and timings (mean and range) of the components of the experimental trials (top) and the filler/task trials
(bottom).
meanings of the ambiguous word remained plausible.
Identical sentence-final words also completed the control
unambiguous sentences. Sentence transcriptions and
stimulus properties can be downloaded from https://
osf.io/3jhtb/.
number of syllables, and number of phonemes (Baayen,
Piepenbrock, & Gulikers, 1995). Sentence-final words
Quello, in the ambiguous sentences either did or did not
resolve the ambiguities, were also matched on the same
factors (Tavolo 2).
The critical 80 ambiguous and 80 unambiguous control
words were matched on mean frequency of occurrence,
Analysis of a large database of meaning dominance rat-
ings for single ambiguous words (Gilbert, Betts, Jose, &
406
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
T
/
.
o
N
0
5
M
UN
sì
2
0
2
1
Tavolo 1. Examples of Four Stimulus Sets Heard by a Single Participant
Condition
Lead In
Ambiguity Control
Continuation
Sentence-final Word
Ambiguous-resolved
The man knew that
Ambiguous-
unresolved
Control-resolved
one more
The woman hoped
that one more
The woman hoped
that one more
ACE
ACE
might be enough to
win the
might be enough to
win the
SPRINT
might be enough to
win the
Control-unresolved
The man knew that
SPRINT
might be enough to
one more
win the
tennis
game
tennis
game
D
o
w
N
l
o
UN
D
e
D
Ambiguous-resolved
Dan looked over all the
ARTICLES
and found that most of
broken
them were
Ambiguous-
unresolved
Rob went through all the
ARTICLES
and found that most of
useless
them were
Control-resolved
Rob went through all the
HAMMERS
and found that most of
broken
them were
Control-unresolved
Dan looked over all the
HAMMERS
and found that most of
useless
them were
Ambiguous-resolved
The couple thought
Ambiguous-
unresolved
that this
The man heard that this
JAM
JAM
was worse than the one
motorway
on the
was worse than the one
television
on the
Control-resolved
The man heard that this
STORM
was worse than the one
motorway
on the
Control-unresolved
The couple thought
STORM
was worse than the one
television
that this
on the
Ambiguous-resolved
His grandfather joked
Ambiguous-
unresolved
that this
His uncle claimed
that this
LEEK
LEEK
was the biggest he
had ever
was the biggest he
had ever
cooked
found
Control-resolved
His uncle claimed
TROUT
was the biggest he
cooked
that this
had ever
Control-unresolved
His grandfather joked
TROUT
was the biggest he
found
that this
had ever
Rodd, 2017) created using standard word association
metodi (Twilley, Dixon, Taylor, & Clark, 1994) con-
firmed that the ambiguous-resolved condition sentences
utilized the subordinate meaning of the ambiguous
parole, with the exception of a small number of sentences
(mean dominance = 0.23, SD = 0.21, max = 0.76, min =
0). The ambiguous-resolved condition sentences were
also tested using the word association method to ensure
that disambiguation to the subordinate meaning occured
only at the sentence-final word (not earlier): Participants
who did not take part in the MEG experiment were pre-
sented with the ambiguous-resolved condition sentences
without the final word, followed by the isolated ambiguous
word, and asked to generate a word that was related to the
ambiguous word as used in the sentence. Dominance rat-
ings of the ambiguous words in context were comparable
to those taken from the database of isolated ambiguous
parole (mean dominance = 0.25, SD = 0.16, max = 0.53,
min = 0). Meaning dominance ratings can be downloaded
from https://osf.io/3jhtb/.
MacGregor et al.
407
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
/
F
.
o
N
0
5
M
UN
sì
2
0
2
1
Tavolo 2. Descriptive Statistics, Mean (SD), for Key Properties of the Four Keywords
Keyword
Sentence-final Word
Property
N
Frequency (log-transformed)
No. syllables
No. phonemes
Ambiguous
80
1.61 (0.51)
1.28 (0.48)
3.85 (1.13)
Control
80
1.3 (0.61)
1.32 (0.47)
4.05 (1.15)
Resolved
80
1.42 (0.64)
1.98 (0.89)
5.40 (2.08)
Unresolved
80
1.83 (0.70)
1.98 (0.95)
5.06 (1.92)
IL 80 ambiguous words and their matched unambigu-
ous control words were used to create 80 stimulus sets.
Within each set, there were two lead-in contexts (per esempio.,
“The man knew…” and “The woman hoped…”), Quale
were crossed with the ambiguous/control words and the
sentence-final ambiguity-resolving/unresolving words, così
resulting in eight stimulus versions. For each set, the eight
versions were separated into two lists—List A and List B,
each containing one sentence from each of the four
conditions such that each ambiguous/control word and
sentence-final word occurred twice, but following a differ-
ent lead-in context. Participants heard stimuli from either
List A or List B (320 stimuli in total), which meant that
although they heard each ambiguous word twice—in a re-
solved and an unresolved sentence—each followed a
different lead-in context (see Figure 1A and Table 1 for
examples of stimulus sets heard by one participant).
The stimuli were spoken by a native speaker of
Southern British English (author M. H. D.) and digitally
recorded (44.1 kHz sampling rate) in a sound-proofed
booth. For each stimulus set, all eight versions of the sen-
tences were recorded, then six segments were extracted
from the recordings, corresponding to the lead in por-
zione (two versions), the target word (ambiguous, unam-
biguous) plus surrounding words, and the sentence-final
word plus surrounding words (see shading in Figure 1A).
The six segments were then concatenated to make the
eight sentence versions, which were carefully checked
to ensure no splices were audible. The procedure of
splicing and then recombining segments meant that,
across conditions, the critical sections of each sentence
(per esempio., ambiguous word, disambiguation) were acoustically
identical. The exact point for splicing was chosen to en-
sure that the recombined stimuli sounded natural (per esempio.,
by selecting silent periods during plosives). Stimuli were
normalized within and between conditions for root-
mean-square (RMS) amplitude using Praat software (from
www.praat.org).
In addition to the experimental stimuli, 20 sets of filler
sentences were constructed with similar lexicosyntactic
structures and properties as the experimental stimuli.
There were four sentence versions per set in which the
ambiguous/control words were crossed with the sentence-
final ambiguity-resolving/unresolving words (80 fillers in
total); as with the experimental stimuli, the ambiguous/
control words and sentence-final words occurred twice
but with a different lead in for each repetition. RMS am-
plitudes of the fillers were adjusted to match the mean
RMS amplitude of the experimental files. Participants
heard all filler sentences. For each of the filler sentences,
probe words were selected for visual presentation in the
relatedness judgment task, which was included to probe
for comprehension and to ensure attentive listening.
Probe words were either strongly related (50% of probes)
or unrelated (50% of probes) to the meaning of the
sentence meaning. The probes were never related to
the unintended meaning of the ambiguous words.
Cloze Probability Test
Following a suggestion from a reviewer, we ran a sen-
tence completion test on our four experimental sentence
types to test whether there were differences in cloze
probability across the four conditions. Data were col-
lected from 77 participants (aged 20–39 years, born and
residing in the United Kingdom, who had learned English
as their first language and had no hearing difficulties)
over the Internet using jsPsych (de Leeuw, 2015) E
JATOS (Lange, Kuhn, & Filevich, 2015), following recruit-
ment via Prolific (Palan & Schitter, 2018; Peer, Samat,
Brandimarte, & Acquisti, 2016). Data from five partici-
pants were excluded (see below) and were replaced to
meet our a priori goal of analyzing 72 set di dati (giving
us a cloze probability resolution for each item of 1.4%).
The same sets of 80 sentences from the MEG study
were used in this test, except that the final words of
each sentence were not presented. Così, for each of
IL 80 experimental items, there were four possible
sentences created by crossing the two lead-in versions
with the two key words (ambiguous or control). A
avoid excess stimulus repetition, each participant was
tested on only two of four sentence variants (cioè., Essi
heard each lead-in version only once, with one variant
presented with an ambiguous word and the other with
a control word; 160 experimental item trials in total).
We counterbalanced whether ambiguous or control
words were presented first for specific items, and which
lead-in variant was paired with an ambiguous word
408
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
T
.
o
N
0
5
M
UN
sì
2
0
2
1
resulting in four experimental versions. Although we
aimed at testing 18 participants in each of the four ex-
perimental versions, because of accidental overrecruitment
in one version, we collected data from 19 participants
in one version, 18 in two versions, E 17 in another
version.
Participants were told that they would hear sentences
in which the ending had been cut off, and their task was
to complete the sentence with the word or words that
first came to mind. In each trial, a spoken sentence was
presented up until the splice point at which the resolved
and unresolved sentences diverged acoustically (Vedere
Figure 1B; cioè., a silent period between the key word
and the sentence-final word). This allowed us to avoid
presenting coarticulatory or other cues that could con-
strain or bias listeners’ choice of sentence-final words.
Tuttavia, because the splice point often occurred two
or more words before the end of the sentence, we also
presented the remaining words before the sentence-final
word as written text. Per esempio, for the item “ace,” lis-
teners would hear: “The man knew that one more ACE
might be enough” (lead-in 1, ambiguous/control key
word) and see: “to win the…,” followed by a text entry
box for a sentence completion response. For splice
points occurring in the middle of a word, these words
were also presented at the start of the text segment to
avoid confusion. Splice points occurred at the same place
for all four sentences for each item, and hence, the text
presented on the screen was the same for all four ver-
sions of each sentence. In addition to the cloze task, COME
in the MEG/EEG experiment (see below), participants
completed the Mill Hill Vocabulary Test (Raven, Raven,
& Court, 1998).
Sentence continuations from each participant were
scored for whether or not they matched the critical
resolved/unresolved sentence. We took only the first
word from each response. These first-word responses
were checked for spelling errors and corrected when
the intended word was obvious (six responses were ex-
cluded for being nonwords and therefore uninterpret-
able). We also checked whether the first-word response
was a repetition of the final word(S) in the cutoff sen-
tence and corrected where necessary (per esempio., sentence:
“The man asked about the nuggets and was told they
were…”, risposta: “were chicken.”). Data sets from five
participants were excluded (and replaced) because (1)
they produced nine or more (5+%) nonresponses or
unusable/uninterpretable responses and/or (2) Essi
scored less than 33% correct on the vocabulary test
(cioè., 2.5 SDs below the sample mean from the main
MEG study). From the 11,520 trials (72 participants ×
160 sentences), 47 missing and uninterpretable re-
sponses were removed, resulting in 11,473 responses
for inclusion in the analysis. A response was scored as a
match if it was (1) an exact match, (2) an inflected form
of the target word (per esempio., “tastes” responses matched the
target word “taste”), O (3) a longer or contracted form of
the target word (per esempio., “gymnasium” responses matched
the target word “gym”). Responses were combined over
participants, lead-in variants and versions.
For each of the 80 experimental items, we calculated
the proportions of responses that matched the resolved
sentence-final words (per esempio., tennis and game) for sen-
tences containing the ambiguous and control words
(per esempio., ACE and SPRINT). The resulting cloze probabili-
ties for the critical words in our sentences were low
overall (Vedi la tabella 3; cloze probabilities for all stimuli
can be downloaded from https://osf.io/3jhtb/), confirm-
ing that—as intended—the sentence-final words were
only weakly constrained by the preceding context. As
the distributions of cloze probabilities for the four con-
ditions were highly skewed, with high frequencies of 0
and near-0 cloze probabilities (cioè., cases where partici-
pants never or very rarely responded with the resolved/
unresolved sentence-final word), we log-transformed
the cloze probabilities to make these distributions more
normal. Before this transformation, any probabilities of
0 were changed to a lower bound probability (½ divided
by the total number of responses for that condition) A
avoid undefined values that result from taking the na-
tural log of 0.
To quantify the degree of experimental control
achieved in our materials, log-transformed cloze proba-
bilities were entered into a Bayesian repeated-measures
ANOVA with default priors ( JASP Team, 2019; Morey &
Rouder, 2015; Rouder, Morey, Speckman, & Province,
2012). This analysis allows us to test for reliable dif-
ferences in cloze probabilities between conditions as in
a conventional ANOVA but importantly to also assess
evidence for the null hypothesis (cioè., that our sentence
Tavolo 3. Descriptive Statistics for Cloze Proportions across the Four Sentence Conditions Shown by Key Word (Ambiguous or
Control) and Sentence-final Word Response (Matching the Resolved Word or Matching the Unresolved Word)
Keyword
Sentence-final Word Response
Mean Cloze
Ambiguous (per esempio., ACE)
Resolved (per esempio., tennis)
Unresolved (per esempio., game)
Control (per esempio., SPRINT)
Resolved
Unresolved
0.03
0.09
0.06
0.08
SD
0.06
0.15
0.11
0.14
Median Cloze
Range
0.00
0.03
0.00
0.01
0.00–0.34
0.00–0.61
0.00–0.53
0.00–0.60
MacGregor et al.
409
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
T
.
o
N
0
5
M
UN
sì
2
0
2
1
materials were well-matched as intended). We included
within-item factors for word type (ambiguous or control)
and sentence-final word response type (resolved or un-
resolved word). Model comparisons provide very strong
evidence for a difference between resolved and unre-
solved words (BF10 = 43.217)—indicating, as expected,
that the more specific resolved words (per esempio., tennis) were
less predicted than the more generic unresolved words
(per esempio., game). Model comparisons provide moderate evi-
dence for the null hypothesis that there is no difference
between cloze probabilities following ambiguous (per esempio.,
ACE) and control (per esempio., SPRINT) parole, (BF10 = 0.130).
Most importantly, Tuttavia, model comparisons also pro-
vide moderate evidence for the null hypothesis that the
interaction between ambiguity and resolved/unresolved
final words is absent (BF10 = 0.258). Based on standard
interpretations of Bayes factors (Lee & Wagenmakers,
2014), this suggests that it is approximately four times
more likely that the interaction is absent than present.
This therefore makes us confident that any interaction in
MEG/EEG response amplitude at the sentence-final word
will not be attributable to differences in cloze probabilities.
Participants
Twenty right-handed native British English speakers with
normal hearing and no record of neurological diseases
took part in the study for financial compensation.
Ethical approval was issued by Cambridge Psychology
Research Ethics Committee (University of Cambridge),
and informed written consent was obtained from all vol-
unteers. No participants had taken part in any of the pre-
tests described or had previously heard the sentences
used. Data from four participants were excluded because
of high noise in MEG or EEG (greater than 50% of trials
were rejected during data processing; see Methods); we
report data from 16 participants (10 women), aged 20–
39 years (mean = 26.5 years, SD = 6 years).
Experimental Procedure
Experimental stimuli from List A or List B were presented
auditorily (through in-ear headphones connected via tub-
ing to a pair of Etymotic drivers, www.etymotic.com) In
four blocks (80 stimuli in each block; 320 stimuli in total)
interspersed with the fillers (20 stimuli in each block; 80
stimuli in total) using E-Prime 2 software (Psychology
Software Tools). The four sentences from each stimulus
set appeared in separate blocks to avoid repetition of
the key words within a block. Across participants, the or-
der of blocks within the list was counterbalanced accord-
ing to a Latin square design, such that each condition
appeared before and after the other conditions for an
equal number of times. Each participant heard a different
pseudorandomized version for each block. Within a block,
there were no more than three sequential presentations of
an ambiguous stimulus and no more than two sequential
presentations of stimuli of a particular condition. There
were no more than two sequential presentations of
fillers/task trials and no more than 10 trials between two
fillers/task trials.
Figure 1C, D shows the structure of the experiment. IL
start of an experimental trial was signaled to the listener by
a red fixation cross (200 msec) visually presented on the
screen, during which they were encouraged to blink if
necessary. The fixation turned black during a silent period
(jittered 1000 ± 100 msec) and remained on the screen
throughout the duration of the spoken sentence (2267–
3765 msec) and for a postsentence silent period (jittered
2000 ± 100 msec). The first part of a filler/task trial followed
an identical structure, but spoken sentences were always
followed by a relatedness judgment task in which single
words were presented visually (3000 msec), followed by a
black fixation cross (jittered 2000 ± 100 msec), and partic-
ipants had to respond whether the word was related or un-
related to the meaning of the sentence they had just heard.
Behavioral Measures
Participants also performed a number of behavioral tasks,
allowing us to assess individual differences in compre-
hension skill, verbal knowledge, and nonverbal ability.
Following the MEG/EEG recording, we tested partici-
pants’ comprehension of the critical sentences in which
an ambiguous word was resolved to a subordinate mean-
ing. Participants listened to the 80 ambiguous-resolved
sentences they had heard during the MEG/EEG session,
each followed by auditory presentation of the ambiguous
word from that sentence. They were asked to explain the
meaning of that word, as it was used in the preceding
sentence, by typing in a synonym or a definition. They
were not explicitly told that the words to which they
had to respond were ambiguous. These responses were
subsequently scored by a native English speaker, naïve to
the purpose of the experiment, who indicated whether
participants generated the subordinate or dominant
meaning of these words.
Participants’ vocabulary knowledge was tested using the
34-question multiple-choice Mill Hill Vocabulary Test
(Raven et al., 1998). We also measured participants’ non-
verbal ability with the Cattell 2a Culture Fair Test (Cattell
& Cattell, 1960), composed of four multiple choice subtests
in which participants (1) complete a sequence of drawings,
(2) select the odd one out from a set of drawings, (3) com-
plete a pattern, E (4) identify which drawing fulfils the
criteria of an example. Following scoring of the individual
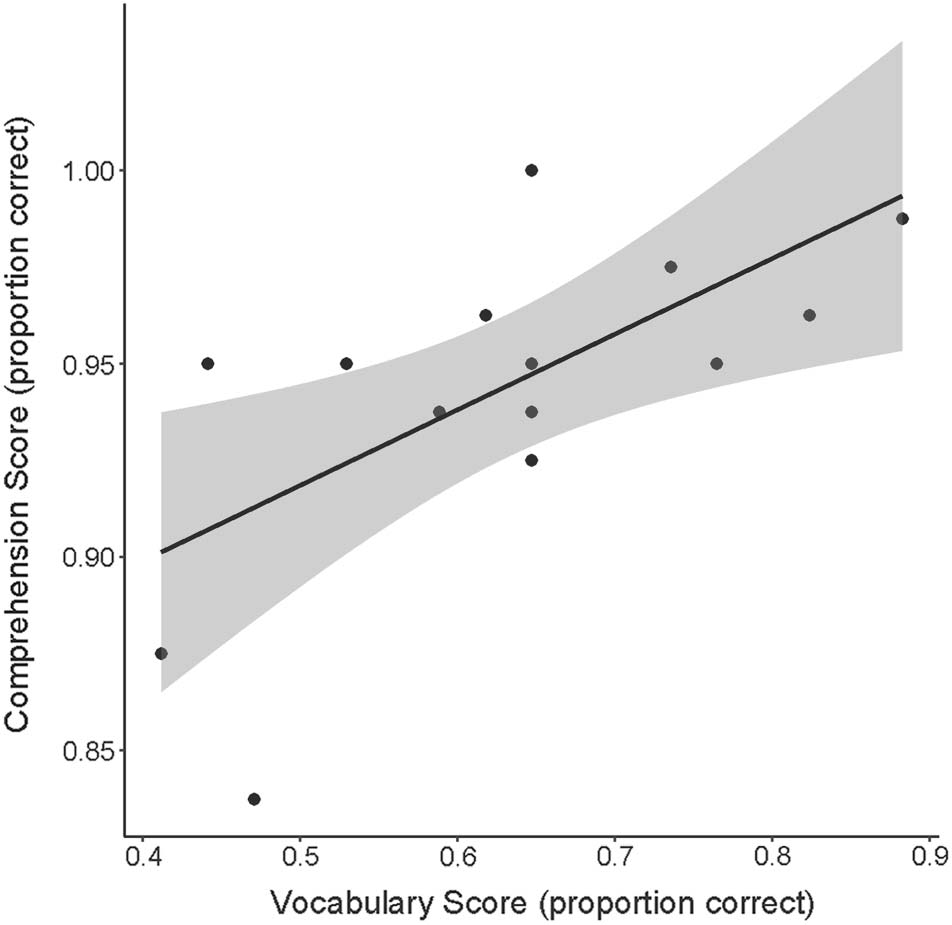
behavioral tests, we assess across-participant correlations
between test scores using Pearson correlations.
MEG and EEG Data Acquisition and Preprocessing
Magnetic fields were recorded (sampling rate 1000 Hz,
bandpass filter 0.03–330 Hz) using a 306-channel Vector-
view system (Elekta Neuromag), which contained one
410
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
/
.
F
o
N
0
5
M
UN
sì
2
0
2
1
magnetometer and two orthogonal gradiometer sensors
at 102 locations within a helmet. Electric potentials were
simultaneously recorded from 70 Ag/AgCl electrodes po-
sitioned according to the 10–10 system and embedded
within an elasticated cap (Easy Cap). Additional elec-
trodes positioned on the nose and one cheek were used
as a reference and the ground, rispettivamente. Vertical and
horizontal electrooculograms were monitored with elec-
trodes placed above and below the left eye and either side
of the eyes, rispettivamente. Electrocardiogram was recorded
with electrodes placed at the upper left and lower right
area of the torso. Head position relative to the sensor ar-
ray was recorded (using the Elekta Neuromag cHPI pro-
tocol with sampling rate of 200 Hz) by using five head
position indicator (HPI) coils that emitted sinusoidal
magnetic fields (293–321 Hz). Before the recording, IL
positions of the HPI coils and 70 EEG electrodes relative
to three anatomical fiducials (nasion, left and right pre-
auricular points) were digitally recorded using a 3-D digi-
tizer (Fastrak Polhemus). Approximately 80 additional
head points over the scalp were also digitized to allow
the offline reconstruction of the head model and coregis-
tration with individual MRI images.
MEG and EEG Data Processing
To minimize the contribution of magnetic sources from
outside the head as well as any artifacts closer to the
MEG sensor array, the data from the 306 MEG sensors were
processed using the signal space separation method (SSS;
Taulu & Kajola, 2005) and its temporal extension (tSSS;
Taulu & Simola, 2006), as implemented in Maxfilter 2.2 soft-
ware (Elekta Neuromag): MEG sensors that generated
poor quality data were identified and data-interpolated
magnetic interference from nonneural sources was sup-
pressed (tSSS buffer of 10 msec and correlation threshold
Di .98). Within-block movement in head position (COME
measured by HPI coils with HPI step set to 10 msec) were
compensated and data interpolated to adjust for head
movement between blocks (interpolation to the first
block). Finalmente, data were downsampled to 250 Hz.
Subsequent preprocessing was performed using MNE
Python version 0.14 (Gramfort et al., 2013, 2014). For
each participant, continuous data from the four record-
ing blocks were concatenated and visually inspected,
and bad EEG channels were identified. To identify compo-
nents associated with eye blinks and cardiac activity and re-
duce their contribution to the data, an independent
component analysis (ICA; FastICA method) was performed
on the raw data (filtered 1–45 Hz, data from bad EEG
channels excluded). Before fitting and applying the ICA,
the data were whitened (decorrelated and scaled to unit
variance—“z-standardized”—also called sphering transfor-
mazione) by means of a PCA. The number of PCA compo-
nents entering the ICA decomposition was selected such
that a cumulative variance of 0.9 was explained. Bad EEG
channels were interpolated after ICA using spherical spline
interpolation (Perrin, Pernier, Bertrand, & Echallier, 1989),
continuous data were filtered (fourth order Butterworth,
0.1–40 Hz), and EEG data were rereferenced to the average
over all EEG channels suitable for source analysis. Lungo
epochs were created around the offset of the critical words
at the two time points of interest (ambiguity: −2800 to
2500 msec; disambiguation: −4400 to 1500 msec), E
each data point was baseline-corrected using mean
amplitude in the silent period before the sentence onset
(ambiguity: −2800 to −2400 msec; disambiguation:
−4400 to −4000 msec).
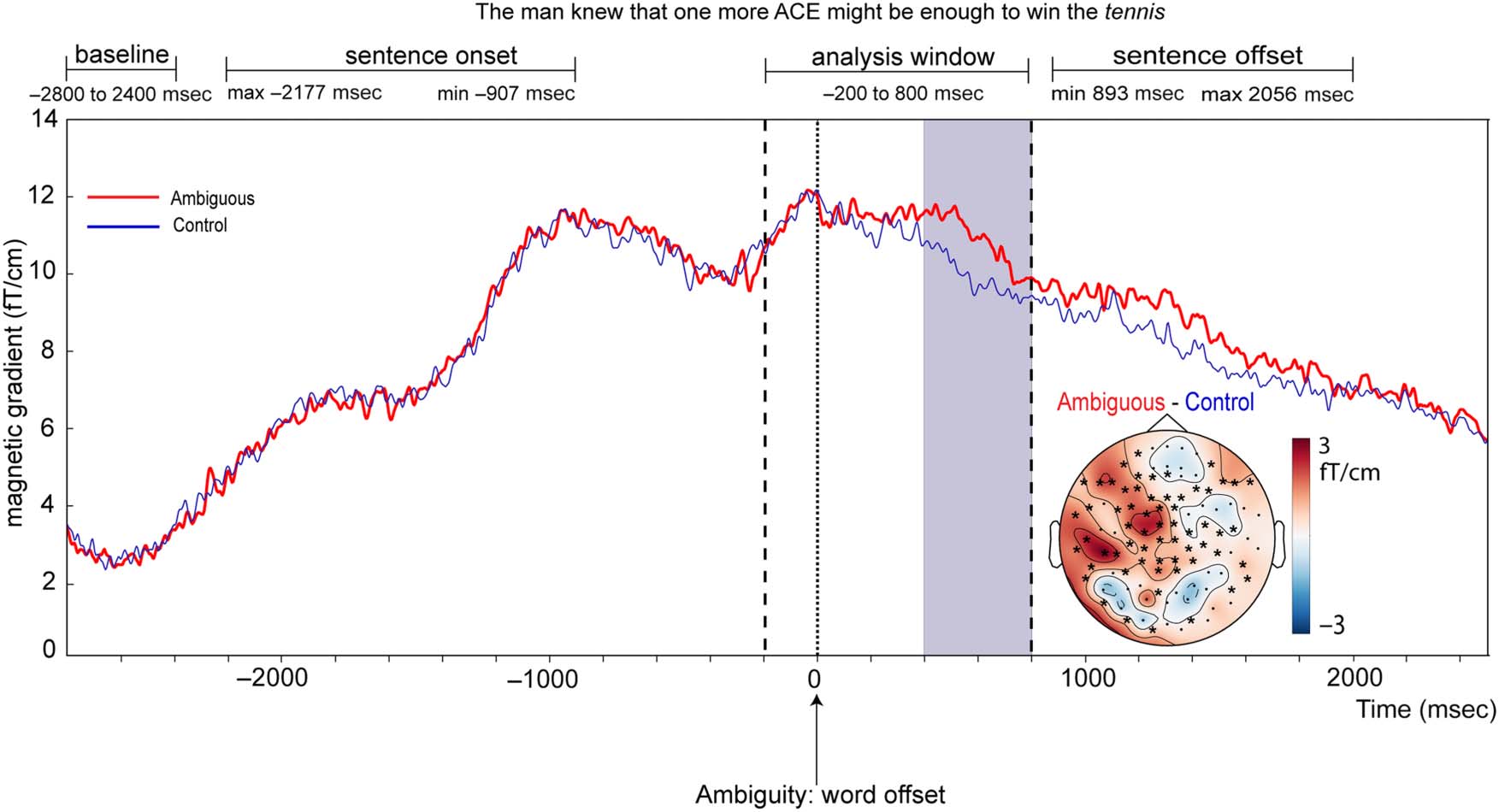
We chose to time-lock MEG and EEG responses to
word offset because, at this point, listeners would have
sufficient phonological information to recognize the
critical words. Because many of our critical words were
monosyllabic, word recognition was unlikely to occur be-
fore this time point (Marslen-Wilson, 1987). Subsequent
processing and analyses were performed on shorter
epochs before and after these word offsets (ambiguity:
−200 to 800 msec; disambiguation: −500 to 1500 msec).
These time windows were chosen in advance based on
our expectations regarding the timing of neural responses
associated with initial meaning selection and reinter-
pretation and on the known timing of the critical words
in our stimuli (Figure 1B). In all sentences, there was at
least 800 msec between the ambiguous word offset and
disambiguation word offset (Figure 1B, right, dotted line),
and in 81% of sentences, there was at least 800 msec
between ambiguous word offset and disambiguation
word onset (Figure 1B, right, solid line); così, we could
be confident that effects before 800 msec should be
attributable to initial meaning activation and selection trig-
gered by the ambiguity rather than subsequent reinterpre-
tation triggered by the disambiguating word. Epochs were
rejected when peak-to-peak amplitudes within the epoch
exceeded the following thresholds: 1000 fT/cm in gradio-
meters, 3500 fT in magnetometers, E 120 μV in EEG
(mean rejection rates: targets 13.3% trials, sentence-final
parole 21.1% trials), and the remaining epochs were aver-
aged across conditions.
Sensor Space Analysis
Before analysis, between-participant differences in head po-
sitions within the helmet were calculated and compensated.
To do this, we calculated the mean sensor array across par-
ticipants and then identified the participant closest to this
average (according to both translation and rotation parame-
ters). MEG data from all participants were transformed to
this common sensory array using the “-trans” option in
MaxFilter 2.2 software (Elekta Neuromag). Data were then
analyzed separately for gradiometers, magnetometers, E
EEG. Before the gradiometer analysis, for every participant
and condition, data from each of the 102 sensor pairs were
combined by taking the RMS of the two amplitudes:
rms gð Þ ¼
. This is a standard procedure in MEG
ffiffiffiffiffiffiffiffiffiffiffiffiffiffi
g11 þ g22
2
q
MacGregor et al.
411
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
.
T
/
o
N
0
5
M
UN
sì
2
0
2
1
analysis, which removes information about the direction of
the two orthogonal gradients at each location. The direc-
tions of the gradients vary across locations with respect to
the brain, and thus are not meaningful for the purposes of
our experimental questions. Before EEG analysis, the data
were rereferenced to the average of left and right mastoid
recordings to allow data to be more comparable to most
previous research on language (note that average referen-
cing is required for combined MEG/EEG source analysis).
Between-condition differences were assessed using
nonparametric cluster-based permutation tests (Maris &
Oostenveld, 2007) to correct for multiple comparisons
in time and space. Using this method, conditions were
compared, and a t value was calculated for every time
point and every sensor. All samples with t values greater
than a threshold equivalent to p < .05 (t = 1.753, one-
tailed; t = 2.131, two-tailed) were selected and clustered
based on temporal and spatial adjacency, and then cluster-
level test statistics were calculated by summing all t values
in a cluster. To evaluate significance, the maximum
cluster-level test statistic was compared against a null dis-
tribution generated by permutations: The participant-
specific averages were randomly permuted within each
participant (5000 times), and the Monte Carlo method
was used to create an approximation of the distribu-
tion of the test statistics under the null hypothesis. The
Monte Carlo p value is the proportion of cluster-level test
statistics from the permutation distribution that is larger
than the observed cluster-level test statistic. Clusters in
which the p value was smaller than the critical alpha level
of .05 support the conclusion that the two conditions are
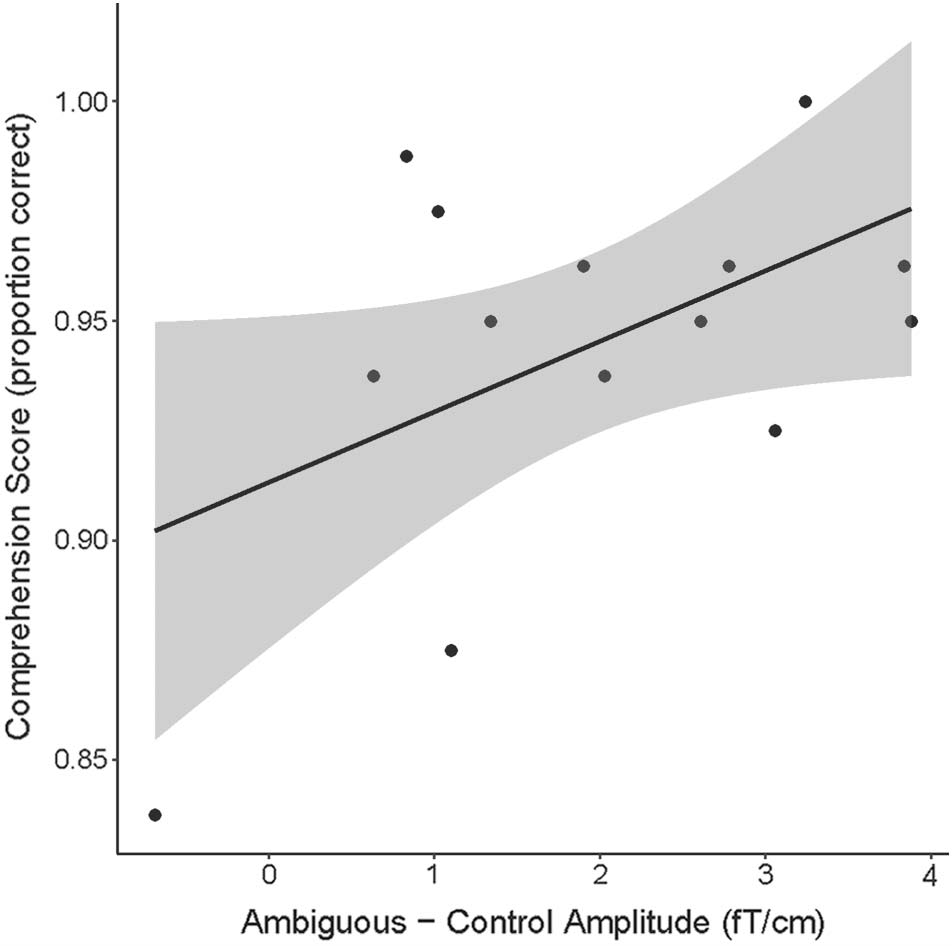
significantly different. Across participants, we tested for
correlations between the amplitude of neural responses
and behavioral scores, using the mean amplitude across
the significant sensor time points within the cluster.
Analyses focused on responses at the time of ambigu-
ity and at the time of disambiguation (Figure 1A). To
identify neural processes associated with initial meaning
activation or selection at the time of ambiguity, we tested
for a directional main effect of ambiguity; that is, whether
ambiguous words elicit greater neural responses than the
unambiguous control words. To identify neural processes
associated with reinterpretation at the time of disambig-
uation, we tested for a directional interaction between
ambiguity and disambiguation. The interaction allowed
us to avoid confounds due to differences in the informa-
tiveness of the sentence-final words within each stimulus
set (e.g., tennis necessarily has a more specific meaning
than game). Specifically, disambiguating sentence-final
words that resolve the ambiguity to a subordinate mean-
ing should elicit greater activity than sentence-final words
that leave the ambiguity unresolved, and this difference in
activation should be greater than the difference between
responses to the acoustically identical sentence-final words
in an unambiguous sentence. For the gradiometer analy-
ses, we performed one-tailed tests because the data had
been rectified using RMS transformation and so values
were all positive and monotonically linked to underlying
neural activity. We could therefore be confident that am-
biguous words would lead to increased signal compared
with control words. For magnetometer and EEG analyses,
we performed two-tailed tests because we did not have
specific predictions regarding the polarity of these effects.
Correlation analyses assessing individual differences in
comprehension were all two-tailed because, even for
comparisons in which we can be confident of observing
greater activity for ambiguous than for control items (e.g.,
ambiguous vs. control items for gradiometers), we could
not anticipate whether more successful ambiguity resolu-
tion would be associated with greater neural engagement
or reduced processing effort (see Taylor, Rastle, & Davis,
2013, 2014, for discussion).
Source Estimation
To estimate the neural sources underpinning the observed
sensor data, we used SPM 12 (Welcome Trust Centre for
Neuroimaging). Data from all three neurophysiological
measurement modalities (EEG and MEG magnetometers
and gradiometers) were integrated using multimodal
source inversion, which has been shown to give more pre-
cise localization than that obtained by considering each mo-
dality in isolation (Henson, Mouchlianitis, & Friston, 2009).
With such an approach, sensor types with higher estimated
levels of noise contribute less to the resulting source solu-
tions. For each participant, high-resolution structural MRI
images (T1-weighted) were obtained using a GRAPPA 3-D
MPRAGE sequence (resolution time = 2250 msec, echo
time = 2.99 msec, flip angle = 9%, and acceleration
factor = 2) on a 3T Tim Trio MR scanner (Siemens) with
1 × 1 × 1 mm isotropic voxels. For each individual, the
structural MRI image was normalized to the standard
Montreal Neurological Institute (MNI) template brain.
The inverse normalization parameters were then used
to spatially transform canonical meshes for the cortex
(8196 vertices) and scalp and skull (2562 vertices) to
the individual space of each participant’s MRI. Sensor lo-
cations and the scalp meshes were aligned using the
three fiducial points measured during digitization with
those identified on the MRI scan and with the digitized
head shape. Forward models to specify how any given
source configuration appears at the sensors were created
separately for MEG using a single-shell model and for
EEG using a boundary element model (following the
recommendations specified in Litvak et al., 2011).
Source inversion was performed using the distributed
L2-minimum-norm method (no depth weighting), which
attempts to minimize overall source power while assum-
ing all currents are equally likely to be active (Dale et al.,
2000). An additional constraint was imposed (SPM “group
inversion”, as recommended in Litvak et al., 2011), whereby
responses for all participants should be explained by the
same set of sources, which has been shown to improve
group-level statistical power (Litvak & Friston, 2008). In
412
Journal of Cognitive Neuroscience
Volume 32, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
4
-
0
p
3
d
f
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
c
3
n
/
_
a
1
_
8
0
6
1
1
4
1
9
0
3
2
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
3
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
0
5
M
a
y
2
0
2
1
on individual participant inversions. Thus, this method ex-
ploits the availability of repeated measurements (from dif-
ferent participants) to constrain source reconstruction.
Importantly, however, the method does not bias activation
differences between conditions to a given source. Source
power (equivalent to the sum of squared amplitude) in
the 0.1–40 Hz range was calculated from the resulting
solutions and converted into 3-D images. Significant ef-
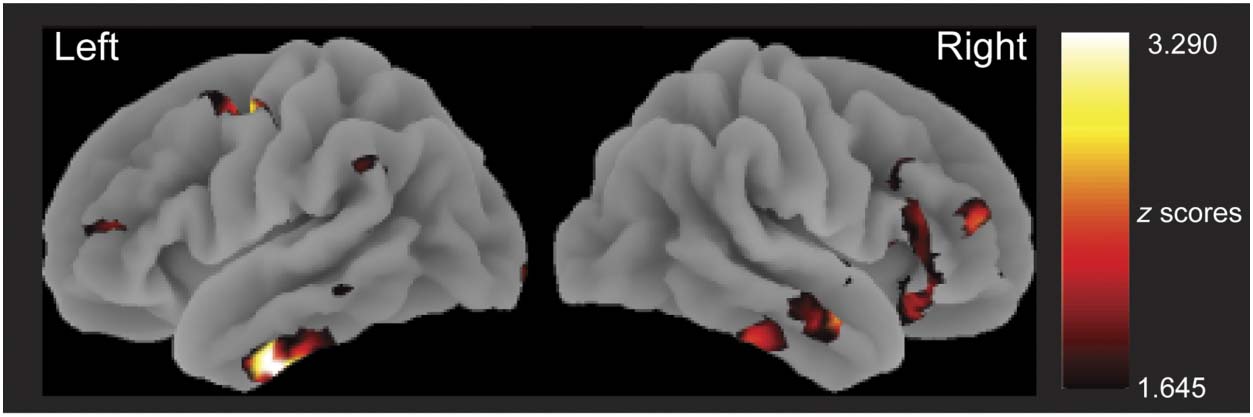
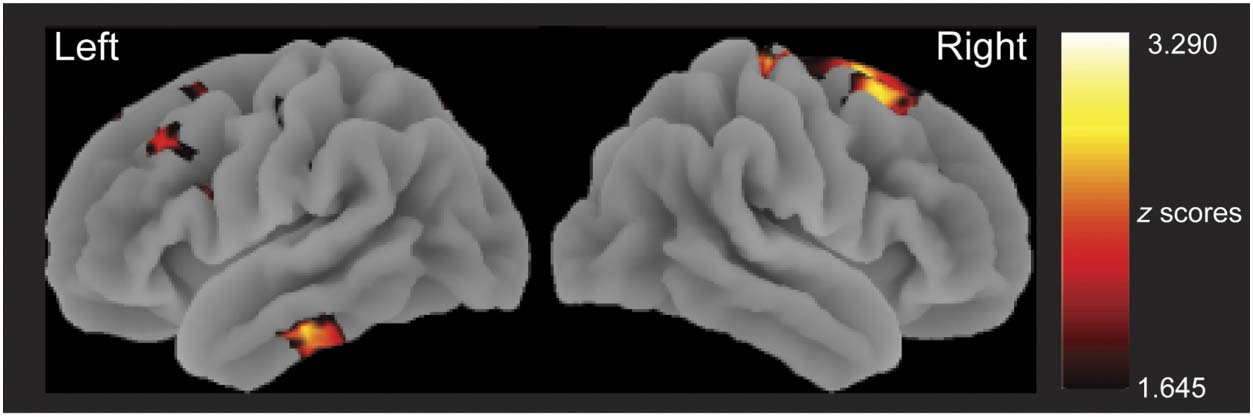
fects from sensor space were localized by taking the
mean 3-D source power estimates across the relevant
time windows and mapping the data onto MNI space
brain templates. Between-condition differences were cal-
culated, and statistical significance in each voxel was
assessed with a series of one-sample t tests at the group
level (i.e., mean signal divided by cross-participant varia-
bility). Because the aim of the source reconstruction was to
localize significant sensor space effects, results are displayed
with an uncorrected voxel-wise threshold ( p < .05, Gross
et al., 2013)
Figure 2. Positive correlation between participants’ vocabulary
score and their score on the post-MEG comprehension test for the
ambiguous-resolved sentences. Shaded areas show the 95% confidence
interval of the regression line.
brief, the procedure involves (1) realigning and con-
catenating sensor-level data across participants, (2) esti-
mating a single source solution for all participants, and
(3) using the resulting group solution as a Bayesian prior
RESULTS
Behavioral Results
On the semantic relatedness judgment task, participants
scored highly overall (mean proportion correct = 0.93,
SD = 0.05), indicating they had listened attentively to
the sentence stimuli. Overall, participants performed well
on the post-MEG/EEG comprehension test, indicating
successful disambiguation of the ambiguous-resolved
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
4
-
0
p
3
d
f
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
c
3
n
/
_
a
1
_
8
0
6
1
1
4
1
9
0
3
2
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
3
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
Figure 3. Evoked response at the time of ambiguity for gradiometers. Responses illustrate significantly greater activation for ambiguous (red line)
compared with unambiguous control words (blue line) corresponding to a cluster in the data from gradiometer pairs (RMS transformed)
beginning approximately 400 msec after word offset, which was prominent over left frontotemporal sensors (analysis time window of −200 to
800 msec relative to word offset). Responses are averaged over all sensors contributing to the significant cluster (highlighted on the topographic
plot). Topographic plot shows the distribution over the scalp of the between-condition difference (Ambiguous–Control), averaged over the
maximal temporal extent of the cluster (highlighted in purple).
MacGregor et al.
413
in response to ambiguous compared with unambiguous
control words observed in a single sensor time cluster
from approximately 400 to 800 msec after word offset
and most pronounced over left frontotemporal sensors
(cluster: 392–800 msec, p = .034, one-tailed; Figure 3).
Across participants, the amplitude of this response (aver-
aged over significant sensor time points) showed a margin-
ally significant positive correlation with comprehension
scores, r(14) = .51, p = .052, two-tailed (Figure 4).
These analyses included responses to all trials irrespective
of whether the sentence was correctly interpreted. To fur-
ther explore the relationship between MEG responses and
successful comprehension, we reanalyzed the data exclud-
ing trials from sentences that were incorrectly understood
in the post-MEG comprehension test (one participant was
excluded because of a failure in recording the comprehen-
sion data). The MEG response at the time of ambiguity re-
mained statistically reliable (cluster: 372–800 msec, p =
.025, one-tailed) and the cross-participant correlation with
comprehension remained marginally significant, r(14) =
0.45, p = .092, two-tailed. Because, on average, only 6%
of sentences were misunderstood, there were insufficient
trial numbers to explore comprehension failures in more
detail.
To confirm that the ambiguity response occurred before
the presentation of any disambiguating information, we car-
ried out a post hoc analysis in which we excluded those trials
in which the sentence for at least one condition had less
than an 800-msec delay between target word offset and
the onset of disambiguating words. That is, we excluded
those items for which our analysis window could include a
response to the onset of disambiguation words. This re-
sulted in the exclusion of the sentences for 19/80 ambiguous
words (76/320 sentences per participant); in the remaining
sentences, the onset of the disambiguating word started af-
ter the end of the analysis time window (defined a priori as
−200 to 800 msec relative to target word offset). Reanalysis
of this subset of trials still showed a significant ambiguity ef-
fect (cluster: 304–800 msec, p = .38, one-tailed) confirming
that these effects are due to ambiguous words and not sub-
sequent disambiguation.
Source localization of the significant neural response to
ambiguous words showed cortical generators in fronto-
temporal regions bilaterally (Figure 5; numbered source
clusters are reported in Table 4). On the left, increased
Figure 4. Positive correlation between the amplitude of the MEG effect
at the time of ambiguity (Ambiguous–Control) and comprehension
scores across participants. Shaded areas show the 95% confidence
interval of the regression line.
sentences (mean = 0.94, SD = 0.04 proportion correct;
scores for one participant were inadvertently not recorded
resulting in n = 15 for analyses of comprehension scores).
Nonverbal IQ scores were above average for the general
population (mean = 130.3, SD = 15.8 normalized scores).
On average, participants knew around two thirds of the
words in the vocabulary test (mean = 0.63, SD = 0.12
proportion correct). Correlational analysis revealed a pos-
itive correlation between sentence comprehension and
vocabulary scores, r(15) =.638, p = .0105 (Figure 2).
There were no reliable correlations between any of the
other behavioral measures.
MEG/EEG Responses at the Time of Ambiguity
Statistical analysis in sensor space revealed significant
effects for gradiometers only (there were no significant
clusters for magnetometers or EEG). At the offset of the
ambiguous word, there was significantly greater activity
Figure 5. Source localization of
the ambiguity-associated
response shown in sensor space
analysis in Figure 3. Results
show activations displayed at
p < .05 (uncorrected) for clarity.
414
Journal of Cognitive Neuroscience
Volume 32, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
3
e
4
-
0
p
3
d
f
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
c
3
n
/
_
a
1
_
8
0
6
1
1
4
1
9
0
3
2
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
3
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
0
5
M
a
y
2
0
2
1
Table 4. Peak Voxel Locations (in MNI Space) and Summary Statistics from Source Analysis of the Response to Ambiguity
Coordinates (mm)
Cluster
Voxels (n)
Region
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
577
204
281
226
218
236
76
463
78
69
27
33
48
33
44
47
34
Left inferior temporal gyrus
Left precentral gyrus
Right middle temporal gyrus
Left calcarine sulcus
Left calcarine sulcus
Right middle frontal gyrus
Right inferior temporal gyrus
Right inferior frontal gyrus (pars triangularis)
Right inferior frontal gyrus (pars orbitalis)
Right inferior frontal gyrus (pars orbitalis)
Right inferior frontal gyrus (pars triangularis)
Right middle frontal gyrus (orbital)
Right superior frontal gyrus (orbital)
Right superior frontal gyrus (orbital)
Left middle frontal gyrus
Left supramarginal gyrus
Left inferior frontal gyrus (pars opercularis)
Right inferior frontal gyrus (pars opercularis)
Right calcarine sulcus
Left inferior temporal gyrusa
Right superior temporal gyrus
Left occipital pole
Left occipital pole
x
−52
−36
58
−12
−2
38
54
36
48
36
54
34
26
24
−36
−58
−40
42
10
−62
58
−16
−32
y
−14
−4
−6
−100
−96
46
−32
20
28
26
26
56
48
56
44
−44
16
18
−96
−40
0
−96
−92
z
−36
48
−22
−8
4
12
−26
26
−10
−18
14
−2
−4
−4
10
32
22
10
2
−14
−2
−18
−18
Z
3.41
3.12
2.75
2.71
1.97
2.56
2.49
2.24
2.23
2.19
2.07
2.1
2.01
1.71
2.05
2.02
2.01
1.93
1.85
1.81
1.8
1.75
1.68
Regions are labeled using the AAL atlas (Tzourio-Mazoyer et al., 2002). Activations are thresholded voxel-wise at p < .5 (uncorrected) and cluster-wise
at k > 25 voxels.
aThis cluster borders the left middle temporal gyrus and in the Harvard–Oxford atlas is labeled as such.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
T
F
.
power for ambiguous compared with unambiguous
control words was seen in the anterior portion of the
ITG extending posteriorly and on the border with MTG
(Cluster 1, Cluster 15). On the right, there was an area of
activation in homologous regions of the ITG (Cluster 6),
which extended into the MTG (Cluster 3), and a small
cluster in superior temporal gyrus (Cluster 16). There was
also a cluster in supramarginal gyrus (Cluster 11). Frontally,
there was a large right-lateralized cluster of activation
in the IFG pars triangularis (Cluster 7), extending into
IFG pars orbitalis (Cluster 8), and IFG pars opercularis
(Cluster 13), and in the middle frontal and superior frontal
gyri (Clusters 5 E 9). On the left, similar clusters of ac-
tivation were seen in IFG pars opercularis (Cluster 12) E
middle frontal gyrus (Cluster 10).
MEG/EEG Responses at the Time of Disambiguation
At the sentence-final word, nonparametric cluster-based
permutation analysis revealed marginally significant inter-
actions between ambiguity and disambiguation for gra-
diometers and for EEG. These arise from sensor-time
clusters at around the time of word offset (Figura 6).
For gradiometers, the interaction corresponded to a
cluster in the left and right hemisphere, lasting from
approximately 200 msec before to 200 msec after the
sentence-final word (cluster: −196 to 156 msec, p =
.078, one-tailed). For EEG, the interaction corresponded
to a cluster for a central cluster of electrodes from over a
similar latency range (cluster: −276 to 212 msec, p =
.081, two-tailed). As predicted, these two marginal effects
o
N
0
5
M
UN
sì
2
0
2
1
MacGregor et al.
415
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
F
T
/
o
N
0
5
M
UN
sì
2
0
2
1
Figura 6. Evoked responses at the time of disambiguation for gradiometers (UN) and EEG (B and C). Responses illustrate marginally significant
interactions between ambiguity and disambiguation. For the gradiometers (UN), responses illustrate significantly greater activation for sentence-final
words that resolved the ambiguity (red solid line) minus words that left the ambiguity unresolved (red dotted line) compared with the activation
difference between identical sentence-final words (blue solid and blue dotted lines) that completed the unambiguous sentences (analysis time
window of −500 to 1500 msec relative to word offset). This effect corresponds to a cluster in the data from gradiometer pairs (RMS transformed),
which is prominent around word offset and is visually similar to a cluster in the data from EEG (B). There is a second cluster for EEG data (C),
corresponding to a significantly greater difference in activation between the sentence-final words for unambiguous sentences than the difference
when these words completed ambiguous sentences. Responses are averaged over all sensors contributing to the significant cluster (highlighted
on the topographic plot). Topographic plots show the distribution over the scalp of the between-condition differences (resolved–unresolved),
averaged over the maximal temporal extent of the clusters (highlighted in purple), for ambiguous and control conditions separately.
416
Journal of Cognitive Neuroscience
Volume 32, Numero 3
Figura 7. Source localization of
the disambiguation-associated
response shown in sensor space
analysis in Figure 6. Results
show activations displayed at
P < .05 (uncorrected) for
clarity.
reflect greater activation for sentence-final words that re-
solved the ambiguity to a subordinate meaning compared
with words that left the ambiguity unresolved; no equiva-
lent difference was observed for resolved/unresolved
words that completed the unambiguous sentences. The
EEG data also showed a marginally significant interaction
for a sensor time cluster in a later time window (cluster:
1144–1500 msec, p = .083, two-tailed), but as can be seen
in Figure 6C, the effect was driven by a greater difference
between sentence-final words in the unambiguous control
sentences than the ambiguous sentences. Because the di-
rection of this interactional effect is inconsistent with any
specific functional contribution to reinterpretation, we do
not consider it further.
To fully characterize the interaction of interest, we also
performed post hoc simple-effect analyses. For the am-
biguous sentences, sentence-final words that resolved
the ambiguity elicited greater activity than those which
left the ambiguity unresolved corresponding to clusters in
the gradiometer (cluster: −236 to 336 msec, p = .002, one-
tailed) and EEG data (cluster: −196 to 236 msec, p = .047,
two-tailed). There was no significant effect for the unam-
biguous sentences (i.e., those that contain a control word
rather than an ambiguous word). There was also greater
Table 5. Peak Voxel Locations (in MNI Space) and Summary Statistics from Source Analysis of the Response to Disambiguation
Coordinates (mm)
Cluster
Voxels (n)
Region
1
2
3
4
5
6
7
8
9
10
11
12
1357
Right supplementary motor area
387
253
150
Right superior frontal gyrus
Right precentral gyrus
Left inferior temporal gyrus
Left fusiform gyrus
Left middle frontal gyrus
Right middle frontal gyrus
Right middle frontal gyrus
68
Left lateral occipital cortex
Left precuneus
101
Left inferior frontal gyrus (pars opercularis)
51
65
73
78
27
25
Left precentral gyrus
Left superior frontal gyrus
Left superior frontal gyrus
Right supramarginal gyrus
Right supramarginal gyrus
Left supramarginal gyrus
Left supplementary motor area
x
12
24
12
−52
−42
−36
34
34
−8
−4
−50
−36
−6
−18
44
50
−50
−4
y
6
12
−22
−24
−32
22
44
36
−74
−72
12
−16
36
10
−32
−30
−28
−4
z
62
60
70
−26
−18
40
30
28
52
44
22
48
46
52
42
48
28
60
Z
2.95
2.93
2.75
2.82
1.74
2.35
2.16
1.98
2.13
1.91
2.12
2.09
1.97
1.97
1.96
1.96
1.92
1.9
Regions are labeled using the AAL atlas (Tzourio-Mazoyer et al., 2002). Activations are thresholded voxel-wise at p < .05 (uncorrected) and cluster-
wise at k > 25 voxels.
MacGregor et al.
417
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
F
/
T
o
N
0
5
M
UN
sì
2
0
2
1
activation for words resolving the ambiguity relative to
acoustically identical words that completed an unambiguous
sentence (gradiometers: cluster −128 to 212 msec, p =
.014, one-tailed; cluster −436 to −100 msec, p = .059,
one-tailed; EEG: cluster −172 to 226 msec, p = .028, two-
tailed) but no difference between the sentence-final words
that left the ambiguity unresolved compared with the
same words that completed an unambiguous sentence.
To identify the source of the disambiguation effect, we
performed source localization on the time window −196
A 156 msec, covering the overlapping time period of ef-
fects in the MEG gradiometer and EEG analyses. Noi
looked for regions with increased power for words that
resolved the ambiguity than for words that left the am-
biguity unresolved, compared with the equivalent differ-
ence in power between identical words that completed
unambiguous sentences. Results (Figura 7 and Table 5)
show generators in left frontotemporal regions, including
regions that overlap with those active at the time of am-
biguity such as the ITG, extending to fusiform (Cluster 2).
There was also a cluster in IFG pars opercularis (Cluster 6)
and smaller frontal clusters in superior frontal gyrus
(Clusters 8 E 9), middle frontal gyrus (Cluster 3),
precentral gyrus (Cluster 7), and SMA (Cluster 13). On
the right, there was a large cluster in SMA, extending to
superior frontal gyrus and precentral gyrus (Cluster 1)
and in the middle frontal gyrus (Cluster 4). We also saw
bilateral clusters in the supramarginal gyrus (Clusters 10
E 11).
DISCUSSION
Using MEG/EEG, we investigated the spatiotemporal dy-
namics of semantic ambiguity resolution by recording neu-
ral responses time-locked to the offset of an ambiguous
word and to a subsequent disambiguating word that re-
solved the ambiguity to a subordinate meaning. Building
on previous fMRI research, we capitalized on the high
temporal resolution of MEG/EEG to distinguish between
the neurocognitive processes of initial meaning access/
selection versus reinterpretation. These are functionally
distinct processes that in our sentences occur just a few
hundred milliseconds apart. We feel confident that we
have distinguished these neurocognitive effects for two
reasons. Primo, an increased neural response associated
with the processing of ambiguous words occurred before
the presentation of disambiguating information that
triggers reinterpretation. Secondo, neural manifestations
of these processes were assessed with two orthogonal sta-
tistical contrasts: Initial ambiguity processing was assessed
through a main effect, whereas reinterpretation was as-
sessed with an interaction.
At the time of ambiguity, we observed significantly
greater MEG responses for ambiguous words versus un-
ambiguous control words (Figura 3). The effect remained
significant when we excluded trials in which the onset of
the sentence-final word that triggers reanalysis occurred
within the analysis window. Così, this neural effect of
ambiguity was observed before the presentation or pro-
cessing of disambiguating information. Inoltre, IL
amplitude of the MEG response at the time of ambiguity
correlated positively with individual differences in compre-
hension skill, as measured by our post-MEG comprehen-
sion test for ambiguous-resolved sentences (Figura 4),
although this effect was only marginally significant. Com-
prehension also correlated positively with vocabulary
scores across participants (Figura 2). We discuss the cog-
nitive processes associated with these neural responses in
the next section. In a subsequent section, we then turn to
neural responses at the time of disambiguation; we ob-
served marginally greater MEG and EEG response ampli-
tudes at the offset of sentence-final words that resolved
an ambiguous word to a subordinate meaning (Figura 6).
Source estimation localized ambiguity responses to bi-
lateral frontotemporal regions (Figura 5) and disambigu-
ation responses to bilateral frontal and left temporal
regions (Figura 7). Given the overlapping neural localiza-
tion of the two cognitively distinct processes involved in
ambiguity resolution, we will discuss these findings from
source localization in a final section of the discussion,
drawing on comparisons with the fMRI literature to
inform our functional interpretation of these neural
responses.
Functional Significance of Neural Responses
to Ambiguity
We take the increased neural response after the offset of
ambiguous words to reflect more effortful processing of
words with more than one meaning compared with
matched single-meaning control words. More specifically,
we relate the effect to the increased demands of meaning
access and selection when multiple possible meanings
are known. This neural effect is consistent with fMRI
studies, as well as data from eye-tracking and ERP studies
on the processing of visually presented ambiguous words
in sentences that we reviewed in the introduction.
Although we described the observed response to am-
biguity as a neural correlate of initial meaning activation
or selection, which we distinguish from subsequent re-
interpretation, this still leaves details of its functional con-
tribution unspecified. È, thus far, unclear whether the
ambiguity response reflects processes involved in either
(io) accessing and maintaining multiple meanings or (ii)
selecting a single meaning of an ambiguous word (per esempio.,
by boosting or suppressing one or other meaning).
Both these processes should be more engaged and/or
more demanding for words with multiple meanings and
hence plausibly observed in our comparison of responses
to ambiguous and control words. Critical for distinguishing
these two processes is the time course over which lis-
teners select a single meaning of an ambiguous word for
sentences in which prior context does not constrain the
likely meaning (as in the present experiment). Tuttavia,
418
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
F
/
.
o
N
0
5
M
UN
sì
2
0
2
1
conventional univariate analysis of MEG/EEG data cannot
provide information on whether and when both meanings
of an ambiguous word are active.
Several sources of experimental evidence have been
used to infer the time course of meaning selection of
ambiguous words in neutral context sentences. For ex-
ample, cross-modal priming studies from Seidenberg
et al. (1982) and Swinney (1979) are consistent with ini-
tial access to multiple meanings followed by selection of
a single, dominant meaning. Swinney (1979) provides
evidence for selective access by three syllables after word
offset, in the time range of 750–1000 msec (P. 657),
whereas Seidenberg et al. (1982) suggest it can occur
sooner, within 200 msec of word offset (both studies in-
dicate activation of both meanings at word offset but do
not test additional time points). Because both sets of data
include a speeded response task with latencies between
500 E 1000 msec, of which around 150 msec can be
accounted for in motor response planning, the minimum
time course over which multiple meanings are main-
tained before selection occurs is in the region of 550–
950 msec. Tuttavia, it is difficult to infer the specific
timing of selection from these studies, in part because
meaning activation is only measured indirectly (by lexical
decision or naming RTs to targets related to one or other
ambiguous word meaning) at discrete points in time and
not to the ambiguous word itself. Nonetheless, in the
context of this study, these findings would suggest that
meaning selection takes place before disambiguating in-
formation is presented for the majority of our sentences.
Tuttavia, successful comprehension of most of our
critical sentences ultimately depends on selecting a lower
frequency or subordinate meaning. Therefore, initial selec-
tion of a single dominant meaning (if that also entails full
suppression of alternative meanings) would make rein-
terpretation even more difficult. Yet, our post-MEG/EEG
comprehension test showed that, on average, listeners
were able to understand more than 90% of delayed dis-
ambiguation sentences, indicating that reinterpretation
was for the most part successful. Therefore, even if full
suppression occurs, listeners can still semantically reana-
lyze the sentence when they encounter a disambiguating
word that conflicts with the previously selected meaning
(perhaps using phonological or working memory). Alter-
natively, full suppression of alternative meanings may
not occur, and multiple meanings of ambiguous words re-
main accessible and to some degree active, at least up to
the point of disambiguation. This proposal is consistent
with RT data from a self-paced reading task showing that
multiple meanings can be maintained over even longer de-
lays until disambiguation (Miyake, Just, & Carpenter,
1992).
One parsimonious description of longer term mainte-
nance of multiple meanings is through a graded constraint
satisfaction process in which listeners make progressively
stronger commitments over time as evidence for alterna-
tives increases (MacDonald, Pearlmutter, & Seidenberg,
1994). By this account, neural activity after an ambiguous
word reflects the activation of multiple alternative inter-
pretations in a representational space that also provides
a mechanism for meaning maintenance such that subse-
quent context can guide selection. In this account, there
is therefore no separation of the neural resources required
for initial activation and maintenance in working memory,
and meaning selection. At face value, this appears consis-
tent with source localization results that we discuss below.
One hallmark of this constraint satisfaction account is
that individual differences in sentence comprehension
arise from experience-dependent learning of the proba-
bilities and regularities that underlie language rather than
in some external, capacity-limited system (such as work-
ing memory; see MacDonald & Christiansen, 2002, for
theoretical elaboration along with recurrent neural
network implementation). The present data provide
tentative findings concerning the relationship between
individual differences in comprehension and neural
responses to semantically ambiguous words. For a sen-
tence in which preceding context does not provide any
specific information to constrain word meaning, the acti-
vation and maintenance of multiple semantic alternatives
is optimal. Hence, additional activation associated with
ambiguous words should be associated with more suc-
cessful comprehension. In line with this proposal, we ob-
served a positive correlation (albeit, only marginally
significant using a two-tailed test) between the amplitude
of the ambiguity-related MEG response and comprehen-
sion success in individual participants. The positive rela-
tionship remained when we excluded sentences
containing ambiguous words that specific participants
did not interpret correctly in the post-MEG/EEG compre-
hension test. This association is therefore not explained
by reduced responses to sentences for which listeners
failed to correctly retrieve the subordinate meaning.
Così, better comprehenders show greater neural pro-
cessing effort in response to ambiguous words.
We explain this correlation between neural responses
and comprehension as indicating that successful com-
prehension of sentences containing ambiguous words
requires additional processes for activation and mainte-
nance of alternative meanings. These result in increased
availability of the appropriate meaning, which is re-
quired when subsequent context resolves the ambiguity
to a subordinate meaning. È interessante notare, better compre-
henders not only have increased availability of subordi-
nate meanings but also achieved higher vocabulary
scores. It might be that higher quality lexical represen-
tations are required both for access to low-frequency
meanings of unambiguous words (for the more difficult
items in the vocabulary test) and for accessing subordi-
nate meanings of ambiguous words (as in our MEG/EEG
study). Nonetheless, given the small number of par-
ticipants and marginally significant results in this study,
this correlation between neural activity and successful
comprehension requires replication and extension. For
MacGregor et al.
419
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
.
F
/
o
N
0
5
M
UN
sì
2
0
2
1
esempio, we might use more difficult sentences to di-
rectly compare neural activity associated with successful
and unsuccessful ambiguity resolution or consider other
predictors of individual variation to relate ambiguity res-
olution (specifically) and spoken language comprehen-
sion (more generally). This study showed no association
between nonverbal IQ and comprehension, but our par-
ticipants did not show as much variation in cognitive
abilities as we might expect in the wider population.
More systematic exploration with a larger group of indi-
viduals with greater variability in comprehension and
measures of other cognitive factors (such as phonological
short-term or working memory) would be valuable.
Functional Interpretation of Neural Responses
to Disambiguation
In addition to neural activity at the time of the ambiguous
word we observed a potential neural marker of reinterpre-
tation during the presentation of sentence-final words that
favor the subordinate meaning of a previous ambiguous
word. Importantly, reinterpretation effects observed at sen-
tence offset in both MEG and EEG were apparent as an in-
teraction between the presence of an ambiguous word and
a sentence-final word that mandated access to an initially
nonpreferred, subordinate meaning. This statistical interac-
tion rules out the possibility that these effects are responses
simply to the presence of an ambiguous word or a more in-
formative sentence-final word (the potentially disambiguat-
ing words necessarily referred to more specific concepts and
had a lower cloze probability). Consistent with this con-
clusion, post hoc simple effects showed that the neural
response to a sentence-final word was affected by the pres-
ence of an ambiguous word earlier in a sentence only when
the sentence-final word disambiguated the ambiguity (E
not if the sentence-final word left the ambiguity unresolved).
Allo stesso modo, response differences between ambiguous and
control words were only apparent at sentence offset if the
sentence-final word served to resolve the ambiguity (Ma
not if the sentence-final word did not conflict with the dom-
inant meaning of the ambiguous word). Although the neural
responses associated with reinterpretation in MEG (gradi-
ometers) and EEG were only marginally significant in analy-
ses correcting for time and sensors, the same pattern of
neural difference was observed in both modalities and in
overlapping tine windows. This similarity gives us greater
confidence in the reliability of these observations.
The approximate timing and sensor topography of
neural responses to reinterpretation are broadly con-
sistent with interpretation as an N400 effect (Kutas &
Hillyard, 1980). Although the N400 has been frequently
observed in the EEG and MEG literature on language pro-
cessing and known to be associated with the processing
of meaning, as yet there is no consensus on an under-
lying functional account or computational mechanisms
(for a review, see Kutas & Federmeier, 2011). For exam-
ple, cognitive accounts suggest it may reflect the ease of
accessing information in semantic memory (Kutas &
Federmeier, 2000) or of integrating semantic information
into context ( Van Berkum, 2009). Computationally, Esso
may be more generally characterized as a semantic pre-
diction error signal (Rabovsky & McRae, 2014), linked to
changes in a probabilistic representation of sentence
Senso (Rabovsky, Hansen, & McClelland, 2018).
ERP N400 responses have previously been observed in
response to disambiguating words that resolve an ambi-
guity to its subordinate meaning (Gunter et al., 2003;
Hagoort & Brown, 1994), although as discussed in the in-
troduction, there are several differences between these
previous studies and ours. Primo, in previous work, sen-
tences were visually presented word-by-word, whereas
our sentences were presented auditorily as connected
speech. Secondo, previous studies did not control for both
the presence/absence of ambiguity and the word form it-
self. We showed a statistical interaction between these
two factors for sentence-final words that trigger reinter-
pretation effects. Unlike previous studies, this interaction
cannot be due to simple differences in word form or
meaning between the critical words in our sentences.
One possibility raised by a reviewer was that the neural
interaction generating this N400-like response to reinter-
pretation could arise from differences in cloze probability
between sentence-final words in our critical conditions.
Tuttavia, a sentence completion test on our materials
showed that cloze probabilities were low overall (IL
median cloze probability was zero in both conditions that
contained the resolved word, close to zero for the unre-
solved words, and did not differ between ambiguous and
control words). We did not include highly constrained sen-
tences or semantic anomalies that are typical of N400 stud-
ies. More importantly, Anche se, a Bayesian analysis of cloze
probability values provided moderately strong evidence that
there was not an interaction between ambiguity and reinter-
pretion (cioè., this analysis provides evidence that the sen-
tences in our critical conditions were matched for cloze
probability). Hence, we can conclude that our N400-like
effect of reinterpretation is not due to variation in the ease
of meaning access due to cloze probability, but rather due to
sentence-final words triggering reinterpretation. None-
theless, future work to determine the functional nature of
the neural response to reinterpretation would benefit from
comparing this response to the semantic error response
evoked by a sentence-final anomalous word. Anomalous
words should trigger an N400-like response but would not
result in reinterpretation, and hence, differences between
anomalous words and words driving reinterpretation may
be informative.
The Role of Frontotemporal Regions in
Ambiguity Resolution
With regard to the anatomical questions that motivated
this study, our source localization provides evidence that
frontal and temporal lobe regions are activated both in
420
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
.
F
T
o
N
0
5
M
UN
sì
2
0
2
1
response to ambiguous words in a neutral context (before
presentation of disambiguating information; Figura 5)
and subsequently in response to a disambiguating word
that resolves the ambiguity to a subordinate meaning
(Figura 7). Previous fMRI evidence has similarly demon-
strated the involvement of frontotemporal regions in
ambiguity resolution (Musz & Thompson-Schill, 2017;
Vitello et al., 2014; Rodd et al., 2005, 2012; Rodd, Longe,
et al., 2010; Mason & Just, 2007; Zempleni et al., 2007).
Tuttavia, unlike in fMRI, timing information from MEG/
EEG allows us to confidently attribute our ambiguity and
disambiguation responses specifically to initial processing
of the ambiguous word and also to subsequent reinterpre-
tation of the ambiguous word. Initial meaning activation/
selection of an ambiguous word was identified through a
statistical main effect, whereas subsequent reinterpretation
at a disambiguating word was identified through a statistical
interaction. Inoltre, responses associated with initial
meaning activation/selection and subsequent reinterpreta-
tion could be separated in time; the neural response to
ambiguity occurred before the onset of disambiguating
words that trigger reinterpretation. Così, these are two in-
dependent effects, and overlap of the neural sources can
inform our understanding of the underlying mechanisms.
As we reviewed in the introduction, previous fMRI
studies on ambiguity resolution have associated activa-
tion in IFG regions on the left ( Vitello et al., 2014) or bi-
laterally (Rodd et al., 2012; Mason & Just, 2007; Zempleni
et al., 2007) with reinterpretation, and in one study, acti-
vation extended into superior and middle frontal areas
(Mason & Just, 2007) in line with the left IFG and supe-
rior and middle frontal clusters shown here. Only two
previous fMRI studies on ambiguity resolution tentatively
associated initial meaning selection with activation in IFG
(Rodd et al., 2012; Mason & Just, 2007). Consistent with
previous conclusions and our findings that IFG is active
both during initial meaning selection and subsequent rein-
terpretation, one dominant proposal regarding the func-
tional role of the left IFG is its involvement in selecting
between competing semantic representation ( Jefferies,
2013; Thompson-Schill, D’Esposito, Aguirre, & Farah,
1997) or resolving conflict arising from competing stimu-
lus representations of any format (Novick, Trueswell, &
Thompson-Schill, 2005; for the suggestion that IFG activa-
tion is involved in selection [or conflict resolution] Piuttosto
than simply reflecting increased competition between se-
mantic representations, see Grindrod, Bilenko, Myers, &
Blumstein, 2008).
An alternative account of IFG contributions to lan-
guage, the unification account (Hagoort, 2005, 2013),
proposes a more general role for the IFG in combining
individual words into coherent sentence- and discourse-
level representations. These are processes that we might
also expect to be taxed as the number of meanings in-
creases and multiple meanings are accessed, maintained,
or predicted. Although we cannot offer any evidence to
adjudicate between these views, we argued above that
meaning selection of the ambiguous words in our study
is likely not completed during the time window before
disambiguation. This seems to favor a more graded
rather than absolute form of selection, perhaps consis-
tent with a constraint satisfaction or unification account.
Previous fMRI studies on ambiguity resolution asso-
ciated activation in the left MTG and ITG/fusiform to re-
interpretation ( Vitello et al., 2014; Rodd et al., 2005,
2012; Rodd, Longe, et al., 2010; Zempleni et al., 2007).
In line with this, localization of the MEG/EEG response
to a disambiguating word indicated a source in the left
ITG and fusiform, which we attribute to reinterpretation.
Notably, we also observed neural sources of the MEG
response to an ambiguous word in MTG and ITG bilater-
alleato, which could be linked to initial meaning activation or
selection. Posterior temporal regions have often been
proposed to contribute to meaning access for isolated
parole (see Lau, Phillips, & Poeppel, 2008; Hickok &
Poeppel, 2007). These regions would plausibly show
greater activation when listeners access multiple meanings
of ambiguous words: Primo, when ambiguity is initially
encountered and again at a disambiguating word inconsis-
tent with the previously preferred meaning, which triggers
an increase in activation of an alternative. We also note
that left posterior MTG activation has previously been
observed in response to syntactically ambiguous words,
using fMRI (Snijders et al., 2009) and MEG (Tyler,
Cheung, Devereux, & Clarke, 2013), although a recent
meta-analysis suggests that these posterior temporal re-
gions are recruited more for semantic rather than syntactic
processing (Rodd, Vitello, Woollams, & Adank, 2015).
We earlier characterized the MEG/EEG reinterpretation
effect as resembling an N400. In line with this proposal,
we note there is some overlap between source localization
of the reinterpretation response to the left ITG and IFG,
and regions proposed to underpin the classic N400 effect,
which have been explored using fMRI and MEG/EEG (Lau,
Weber, Gramfort, Hamalainen, & Kuperberg, 2016; Lau,
Gramfort, Hämäläinen, & Kuperberg, 2013; Maess,
Herrmann, Hahne, Nakamura, & Friederici, 2006; Halgren
et al., 2002). The N400 is likely to reflect a combination
of neural processes originating in multiple cortical region
sources but across a number of studies; it has been pro-
posed that the effect may originate in posterior temporal
regions before being observed in more anterior portions
of the temporal lobe and IFG (for a review, see Lau et al.,
2008).
È interessante notare, the ITG/fusiform activations we observed
at the time of disambiguation and in response to ambi-
guity extended to more anterior and inferior temporal
regions than has been seen in previous fMRI studies of
ambiguity resolution. Anterior temporal activations have
been less consistently observed in fMRI, perhaps because
standard EPI acquisitions give relatively poor signal in
these regions ( Visser, Jefferies, & Lambon Ralph, 2010;
Devlin et al., 2000, although see Musz & Thompson-
Schill, 2017, for evidence of anterior inferior temporal
MacGregor et al.
421
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
/
.
F
o
N
0
5
M
UN
sì
2
0
2
1
representations of ambiguous words shown by multivariate
pattern analysis fMRI). Tuttavia, damage to the anterior
temporal lobe has long been associated with impaired se-
mantic processing in general (Patterson et al., 2007) E
of semantically ambiguous words in particular (per esempio., mea-
sured by patients’ ability to produce alternative interpreta-
tions of unresolved ambiguous sentences (Zaidel et al.,
1995). Così, the inferior temporal activation we ob-
served when listeners initially encounter an ambiguous
word and when disambiguating information is heard is
largely consistent with other evidence for semantic con-
tributions of these basal temporal regions.
One other point to consider is that both the frontal
and temporal neural sources of responses to ambiguity
and disambiguation appear to be somewhat bilateral.
Previous fMRI studies have reported significant activation
in right frontal regions (Mason & Just, 2007; Zempleni
et al., 2007; Rodd et al., 2005), although reports of right
temporal lobe responses are more limited, and a meta-
analysis of fMRI studies of semantic and syntactic pro-
cessing demands reveals fewer and less reliable findings
of right than left frontotemporal activity (Rodd et al.,
2015). Tuttavia, in the absence of statistical comparison
of left- and right-sided activity in fMRI or MEG/EEG, we
hesitate to draw strong conclusions from these obser-
vations (see Peelle, 2012, for arguments that lateralized
effects in thresholded statistical maps provide little or
no evidence for functional lateralization). Inoltre,
other evidence is consistent with bilateral contributions
to ambiguity resolution, Per esempio, from behavioral
studies using lateralized word presentations (Faust &
Gernsbacher, 1996; Burgess & Simpson, 1988) and neuro-
psychological studies (Tompkins, Baumgaertner, Lehman,
& Fassbinder, 2000; Swaab, Brown, & Hagoort, 1998;
Hagoort, 1993). Although functional imaging evidence
can potentially play an important role in determining the
differential contributions of the left and right hemisphere
to ambiguity resolution, published studies, including the
present work, have yet to report hemispheric dissociations
sufficient to conclude that the left and right hemispheres
make distinct functional contributions to initial meaning
activation and selection.
Conclusions
Taken together with previous fMRI research, our observa-
tions suggest that both temporal and frontal regions play
an important role both in initial meaning activation and
selection for ambiguous words, as well as later reinter-
pretation triggered by a disambiguating word. Previous
research has tried to fractionate frontal and temporal re-
gions based on the time course of activation during de-
layed disambiguation sentences (Rodd et al., 2012) or by
comparing responses to ambiguous words with balanced
and biased meaning frequencies ( Vitello et al., 2014;
Mason & Just, 2007; Zempleni et al., 2007). Tuttavia,
source localization results from MEG/EEG suggest that
frontal and temporal regions play a coordinated role both
in the initial interpretation of ambiguous words pre-
sented in neutral sentence contexts and subsequently
when interpretations need to be revised. This proposal
could be taken to challenge traditional divisions between
temporal lobe contributions to semantic representation
and frontal contributions to working memory or selec-
zione (see Musz & Thompson-Schill, 2017, for a recent
statement along these lines).
Rather than the traditional fractionation of temporal
and frontal responses, we instead propose a graded, con-
straint satisfaction account that elides a simple distinction
between semantic representations and processing. In this
account, neural activity after an ambiguous word reflects
the activation of multiple alternative interpretations in a
representational space that also supports neural mecha-
nisms for meaning maintenance and eventual selection.
During this time period, selection can be construed as
stronger, but not exclusive, activation of a particular
Senso, which can only be confirmed when disambigu-
ating information is presented. At this point, successful
meaning integration and interpretation may require re-
interpretation, which can be realized in terms of a re-
weighting of the activation levels of different meanings.
Future work to assess the representational dynamics of
these frontal and temporal responses (per esempio., using represen-
tational similarity or other, similar multivariate methods;
Kriegeskorte, Mur, & Bandettini, 2008) might provide addi-
tional evidence for this account.
Ringraziamenti
This work was supported by the Medical Research Council (SUAG/
008 RG91365 to M. H. D. and SUAG058 G101400 to O. H.). IL
authors thank Jane Warren for helping in creating the stimuli,
Maarten van Casteren for helping during data collection, E
Isobel Davis helping with scoring and analyzing behavioral data.
Reprint requests should be sent to Lucy J. MacGregor, MRC
Cognition and Brain Sciences Unit, University of Cambridge,
15 Chaucer Road, Cambridge, CB2 7EF, or via e-mail: lucy.
macgregor@mrc-cbu.cam.ac.uk.
REFERENCES
Baayen, H. R., Piepenbrock, H., & Gulikers, l. (1995). The CELEX
lexical database (release 2). Distributed by the Linguistic Data
Consortium. University of Pennsylvania, Philadelphia, PAPÀ.
Boynton, G. M., Engel, S. A., Glover, G. H., & Heeger, D. J. (1996).
Linear systems analysis of functional magnetic resonance
imaging in human V1. Journal of Neuroscience, 16, 4207–4221.
Burgess, C., & Simpson, G. B. (1988). Cerebral hemispheric
mechanisms in the retrieval of ambiguous word meanings.
Brain and Language, 33, 86–103.
Cai, Z. G., Gilbert, R. A., Davis, M. H., Gaskell, M. G., Farrar, L.,
Adler, S., et al. (2017). Accent modulates access to word
Senso: Evidence for a speaker-model account of spoken
word recognition. Cognitive Psychology, 98, 73–101.
Cattell, R. B., & Cattell, UN. K. S. (1960). Handbook for the
individual or group culture fair intelligence test. Champaign,
IL: Testing.
422
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
F
T
/
o
N
0
5
M
UN
sì
2
0
2
1
Coleman, M. R., Davis, M. H., Rodd, J. M., Robson, T., Ali, A.,
Hagoort, P., & Brown, C. (1994). Brain responses to lexical
Owen, UN. M., et al. (2009). Towards the routine use of brain
imaging to aid the clinical diagnosis of disorders of
consciousness. Brain, 132, 2541–2552.
Coleman, M. R., Rodd, J. M., Davis, M. H., Johnsrude, IO. S.,
Menon, D. K., Pickard, J. D., et al. (2007). Do vegetative
patients retain aspects of language comprehension? Evidence
from fMRI. Brain, 130, 2494–2507.
Dale, UN. M., Liu, UN. K., Fischl, B. R., Buckner, R. L., Belliveau,
J. W., Lewine, J. D., et al. (2000). Dynamic statistical
parametric mapping: Combining fMRI and MEG for high-
resolution imaging of cortical activity. Neuron, 26, 55–67.
Davis, M. H., Coleman, M. R., Absalom, UN. R., Rodd, J. M.,
Johnsrude, IO. S., Matta, B. F., et al. (2007). Dissociating
speech perception and comprehension at reduced levels of
awareness. Proceedings of the National Academy of
Scienze, U.S.A., 104, 16032–16037.
ambiguity resolution and parsing. In C. Clifton, Jr., l. Frazier,
& K. Rayner, (Eds.), Perspectives on sentence processing
(pag. 45–80). Hillsdale, NJ: Lawrence Erlbaum.
Halgren, E., Dhond, R. P., Christensen, N., Van Petten, C.,
Marinkovic, K., Lewine, J. D., et al. (2002). N400-like
magnetoencephalography responses modulated by semantic
context, word frequency, and lexical class in sentences.
Neuroimage, 17, 1101–1116.
Henderson, L., Snowling, M., & Clarke, P. (2013). Accessing,
integrating, and inhibiting word meaning in poor
comprehenders. Scientific Studies of Reading, 17, 177–198.
Henson, R. N., Mouchlianitis, E., & Friston, K. J. (2009). MEG
and EEG data fusion: Simultaneous localisation of face-
evoked responses. Neuroimage, 47, 581–589.
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8, 393–402.
de Leeuw, J. R. (2015). jsPsych: A JavaScript library for
JASP Team. (2019). JASP (Version 0.10.0.0) [Computer
creating behavioral experiments in a web browser. Behavior
Research Methods, 47, 1–12.
Devlin, J. T., Russell, R. P., Davis, M. H., Price, C. J., Wilson, J.,
Moss, H. E., et al. (2000). Susceptibility-induced loss of signal:
Comparing PET and fMRI on a semantic task. Neuroimage,
11, 589–600.
Duffy, S. A., Morris, R. K., & Rayner, K. (1988). Lexical ambiguity
and fixation times in reading. Journal of Memory and
Language, 27, 429–446.
Faust, M. E., & Gernsbacher, M. UN. (1996). Cerebral mechanisms
for suppression of inappropriate information during sentence
comprehension. Brain and Language, 53, 234–259.
Federmeier, K. D., Segal, J. B., Lombrozo, T., & Kutas, M.
(2000). Brain responses to nouns, verbs and class-ambiguous
words in context. Brain, 123, 2552–2566.
Frazier, L., & Rayner, K. (1990). Taking on semantic
commitments: Processing multiple meanings vs. multiple
sensi. Journal of Memory and Language, 29, 181–200.
software].
Jefferies, E. (2013). The neural basis of semantic cognition:
Converging evidence from neuropsychology, neuroimaging
and TMS. Cortex, 49, 611–625.
Josephs, O., & Henson, R. N. UN. (1999). Event-related functional
magnetic resonance imaging: Modelling, inference and
optimization. Philosophical Transactions of the Royal Society
of London, Series B: Biological Sciences, 354, 1215–1228.
Kambe, G., Rayner, K., & Duffy, S. UN. (2001). Global context
effects on processing lexically ambiguous words: Evidence
from eye fixations. Memory and Cognition, 29, 363–372.
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008).
Representational similarity analysis—Connecting the
branches of systems neuroscience. Frontiers in Systems
Neuroscience, 2, 4.
Kutas, M., & Federmeier, K. D. (2000). Electrophysiology
reveals semantic memory use in language comprehension.
Trends in Cognitive Sciences, 4, 463–470.
Gernsbacher, M. A., Varner, K. R., & Faust, M. E. (1990).
Kutas, M., & Federmeier, K. D. (2011). Thirty years and
Investigating differences in general comprehension skill.
Journal of Experimental Psychology: Learning Memory and
Cognition, 16, 430–445.
Gilbert, R. A., Betts, H. N., Jose, R., & Rodd, J. M. (2017).
New UK-based dominance norms for ambiguous words.
Paper presented at the Poster session presented at the
Experimental Psychology Society Meeting.
Gramfort, A., Luessi, M., Larson, E., Engemann, D., Strohmeier,
D., Brodbeck, C., et al. (2013). MEG and EEG data analysis
with MNE-Python. Frontiers in Neuroscience, 7, 267.
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A.,
Strohmeier, D., Brodbeck, C., et al. (2014). MNE software for
processing MEG and EEG data. Neuroimage, 86, 446–460.
Grindrod, C. M., Bilenko, N. Y., Myers, E. B., & Blumstein, S. E.
(2008). The role of the left inferior frontal gyrus in implicit
semantic competition and selection: An event-related fMRI
study. Brain Research, 1229, 167–178.
Gross, J., Baillet, S., Barnes, G. R., Henson, R. N., Hillebrand, A.,
Jensen, O., et al. (2013). Good practice for conducting and
reporting MEG research. Neuroimage, 65, 349–363.
Gunter, T. C., Wagner, S., & Friederici, UN. D. (2003). Working
memory and lexical ambiguity resolution as revealed by ERPs:
A difficult case for activation theories. Journal of Cognitive
Neuroscience, 15, 643–657.
Hagoort, P. (1993). Impairments of lexical semantic processing
in aphasia—Evidence from the processing of lexical
ambiguities. Brain and Language, 45, 189–232.
Hagoort, P. (2005). On Broca, brain, and binding: A new
framework. Trends in Cognitive Sciences, 9, 416–423.
Hagoort, P. (2013). MUC (memory, unification, controllo) E
beyond. Frontiers in Psychology, 4, 416.
counting: Finding meaning in the N400 component of the
event-related brain potential (ERP). Annual Review of
Psychology, 62, 621–647.
Kutas, M., & Hillyard, S. UN. (1980). Reading senseless sentences:
Brain potentials reflect semantic incongruity. Scienza, 207,
203–205.
Lange, K., Kuhn, S., & Filevich, E. (2015). “Just another tool for
online studies” ( JATOS): An easy solution for setup and
management of web servers supporting online studies.
PLoS One, 10, e0130834.
Lau, E. F., Gramfort, A., Hämäläinen, M. S., & Kuperberg, G. R.
(2013). Automatic semantic facilitation in anterior temporal
cortex revealed through multimodal neuroimaging. Journal
of Neuroscience, 33, 17174–17181.
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network
for semantics: (De)constructing the N400. Nature Reviews
Neuroscience, 9, 920–933.
Lau, E. F., Weber, K., Gramfort, A., Hamalainen, M. S., &
Kuperberg, G. R. (2016). Spatiotemporal signatures of lexical-
semantic prediction. Cerebral Cortex, 26, 1377–1387.
Lee, C. L., & Federmeier, K. D. (2006). To mind the mind: An
event-related potential study of word class and semantic
ambiguity. Brain Research, 1081, 191–202.
Lee, C. L., & Federmeier, K. D. (2009). Wave-ering: An ERP
study of syntactic and semantic context effects on ambiguity
resolution for noun/verb homographs. Journal of Memory
and Language, 61, 538–555.
Lee, C. L., & Federmeier, K. D. (2012). Ambiguity’s aftermath:
How age differences in resolving lexical ambiguity affect
subsequent comprehension. Neuropsychologia, 50,
869–879.
MacGregor et al.
423
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
T
F
/
o
N
0
5
M
UN
sì
2
0
2
1
Lee, M., & Wagenmakers, E. (2014). Bayesian model
comparison. In Bayesian cognitive modeling: A practical
course (pag. 101–117). Cambridge: Cambridge University
Press.
Litvak, V., & Friston, K. (2008). Electromagnetic source
reconstruction for group studies. Neuroimage, 42,
1490–1498.
Litvak, V., Mattout, J., Kiebel, S., Phillips, C., Henson, R., Kilner,
J., et al. (2011). EEG and MEG data analysis in SPM8.
Computational Intelligence and Neuroscience, 2011,
852961.
MacDonald, M. C., & Christiansen, M. H. (2002). Reassessing
working memory: Comment on Just and Carpenter (1992)
and Waters and Caplan (1996). Psychological Review, 109,
35–54.
MacDonald, M. C., Pearlmutter, N. J., & Seidenberg, M. S.
(1994). The lexical nature of syntactic ambiguity resolution
[corrected]. Psychological Review, 101, 676–703.
Maess, B., Herrmann, C. S., Hahne, A., Nakamura, A., & Friederici,
UN. D. (2006). Localizing the distributed language network
responsible for the N400 measured by MEG during auditory
sentence processing. Brain Research, 1096, 163–172.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190.
Marslen-Wilson, W. D. (1987). Functional parallelism in spoken
word recognition. Cognition, 25, 71–102.
Mason, R. A., & Just, M. UN. (2007). Lexical ambiguity in sentence
comprehension. Brain Research, 1146, 115–127.
Miyake, A., Just, M. A., & Carpenter, P. UN. (1992). Working
memory constraints on the maintenance of multiple
interpretations of lexical ambiguities. Bulletin of the
Psychonomic Society, 30, 482.
Morey, R. D., & Rouder, J. N. (2015). BayesFactor: Computation
of Bayes factors for common designs (R Package Version
0.9.12-2) [Computer software]. Available at: https://cran.r-
project.org/package=BayesFactor.
Musz, E., & Thompson-Schill, S. l. (2017). Tracking competition
and cognitive control during language comprehension with
multi-voxel pattern analysis. Brain and Language, 165, 21–32.
Novick, J. M., Trueswell, J. C., & Thompson-Schill, S. l. (2005).
Cognitive control and parsing: Reexamining the role of
Broca’s area in sentence comprehension. Cognitive, Affective
& Behavioral Neuroscience, 5, 263–281.
Palan, S., & Schitter, C. (2018). Prolific.ac—A subject pool for
online experiments. Journal of Behavioral and
Experimental Finance, 17, 22–27.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic
knowledge in the human brain. Nature Reviews
Neuroscience, 8, 976–987.
Peelle, J. E. (2012). The hemispheric lateralization of speech
processing depends on what “speech” is: A hierarchical
perspective. Frontiers in Human Neuroscience, 6, 309.
Peer, E., Samat, S., Brandimarte, L., & Acquisti, UN. (2016).
Beyond the Turk: An empirical comparison of alternative
platforms for online behavioral research. Journal of
Experimental Psychology, 70, 153–163.
Perrin, F., Pernier, J., Bertrand, O., & Echallier, J. F. (1989).
Spherical splines for scalp potential and current-density
mapping. Electroencephalography and Clinical
Neurophysiology, 72, 184–187.
Rabovsky, M., Hansen, S. S., & McClelland, J. l. (2018).
Modelling the N400 brain potential as change in a
probabilistic representation of meaning. Nature Human
Behaviour, 2, 693–705.
Rabovsky, M., & McRae, K. (2014). Simulating the N400 ERP
component as semantic network error: Insights from a
feature-based connectionist attractor model of word
Senso. Cognition, 132, 68–89.
Raven, J., Raven, J. C., & Court, J. H. (1998). Manual for Raven’s
progressive matrices and vocabulary scales. Sezione 5, Mill
Hill vocabulary scale (1998 ed). Oxford: Oxford Psychologists
Press.
Rayner, K., & Duffy, S. UN. (1986). Lexical complexity and fixation
times in reading: Effects of word frequency, verb complexity,
and lexical ambiguity. Memory and Cognition, 14, 191–201.
Rodd, J. M., Cutrin, B. L., Kirsch, H., Millar, A., & Davis, M. H.
(2013). Long-term priming of the meanings of ambiguous
parole. Journal of Memory and Language, 68, 180–198.
Rodd, J. M., Davis, M. H., & Johnsrude, IO. S. (2005). The neural
mechanisms of speech comprehension: fMRI studies of
semantic ambiguity. Cerebral Cortex, 15, 1261–1269.
Rodd, J. M., Gaskell, G., & Marslen-Wilson, W. D. (2002).
Making sense of semantic ambiguity: Semantic competition
in lexical access. Journal of Memory and Language, 46,
245–266.
Rodd, J. M., Johnsrude, IO. S., & Davis, M. H. (2010). The role of
domain-general frontal systems in language comprehension:
Evidence from dual-task interference and semantic
ambiguity. Brain and Language, 115, 182–188.
Rodd, J. M., Johnsrude, IO. S., & Davis, M. H. (2012). Dissociating
frontotemporal contributions to semantic ambiguity
resolution in spoken sentences. Cerebral Cortex, 22,
1761–1773.
Rodd, J. M., Longe, O. A., Randall, B., & Tyler, l. K. (2010). IL
functional organisation of the frontotemporal language
system: Evidence from syntactic and semantic ambiguity.
Neuropsychologia, 48, 1324–1335.
Rodd, J. M., Vitello, S., Woollams, UN. M., & Adank, P. (2015).
Localising semantic and syntactic processing in spoken and
written language comprehension: An activation likelihood
estimation meta-analysis. Brain and Language, 141, 89–102.
Rouder, J. N., Morey, R. D., Speckman, P. L., & Province,
J. M. (2012). Default Bayes factors for ANOVA designs.
Journal of Mathematical Psychology, 56, 356–374.
Seidenberg, M. S., Tanenhaus, M. K., Leiman, J. M., & Bienkowski,
M. (1982). Automatic access of the meanings of ambiguous
words in context: Some limitations of knowledge-based
processing. Cognitive Psychology, 14, 489–537.
Snijders, T. M., Vosse, T., Kempen, G., Van Berkum, J. J.,
Petersson, K. M., & Hagoort, P. (2009). Retrieval and
unification of syntactic structure in sentence comprehension:
An fMRI study using word-category ambiguity. Cerebral
Cortex, 19, 1493–1503.
Swaab, T. Y., Brown, C., & Hagoort, P. (1998). Understanding
ambiguous words in sentence contexts: Electrophysiological
evidence for delayed contextual selection in Broca’s aphasia.
Neuropsychologia, 36, 737–761.
Swinney, D. UN. (1979). Lexical access during sentence
comprehension: (Re)consideration of context effects.
Journal of Verbal Learning and Verbal Behavior, 18,
645–659.
Szabo Wankoff, L., & Cairns, H. S. (2009). Why ambiguity
detection is a predictor of early reading skill.
Communication Disorders Quarterly, 30, 183–192.
Tahmasebi, UN. M., Davis, M. H., Wild, C. J., Rodd, J. M.,
Hakyemez, H., Abolmaesumi, P., et al. (2012). Is the link
between anatomical structure and function equally strong at
all cognitive levels of processing? Cerebral Cortex, 22,
1593–1603.
Taulu, S., & Kajola, M. (2005). Presentation of electromagnetic
multichannel data: The signal space separation method.
Journal of Applied Physics, 97, 124905.
Taulu, S., & Simola, J. (2006). Spatiotemporal signal space
separation method for rejecting nearby interference in MEG
424
Journal of Cognitive Neuroscience
Volume 32, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
.
T
F
o
N
0
5
M
UN
sì
2
0
2
1
measurements. Physics in Medicine and Biology, 51,
1759–1768.
Taylor, J. S., Rastle, K., & Davis, M. H. (2013). Can cognitive
models explain brain activation during word and pseudoword
reading? A meta-analysis of 36 neuroimaging studies.
Psychological Bulletin, 139, 766–791.
Taylor, J. S., Rastle, K., & Davis, M. H. (2014). Interpreting
response time effects in functional imaging studies.
Neuroimage, 99, 419–433.
Thompson-Schill, S. L., D’Esposito, M., Aguirre, G. K., & Farah,
M. J. (1997). Role of left inferior prefrontal cortex in retrieval
of semantic knowledge: A reevaluation. Atti del
National Academy of Sciences, U.S.A., 94, 14792–14797.
Tompkins, C. A., Baumgaertner, A., Lehman, M. T., & Fassbinder, W.
(2000). Mechanisms of discourse comprehension impairment
after right hemisphere brain damage: Suppression in lexical
ambiguity resolution. Journal of Speech, Language, E
Hearing Research, 43, 62–78.
Twilley, l. C., Dixon, P., Taylor, D., & Clark, K. (1994).
University-of-Alberta norms of relative meaning frequency for
566 homographs. Memory and Cognition, 22, 111–126.
Tyler, l. K., Cheung, T. P., Devereux, B. J., & Clarke, UN. (2013).
Syntactic computations in the language network: Characterizing
dynamic network properties using representational similarity
analysis. Frontiers in Psychology, 4, 271.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neuroimage, 15, 273–289.
Van Berkum, J. J. (2009). The neuropragmatics of ’simple’
utterance comprehension: An ERP review. In K. Y. U.
Sauerland (Ed.), Semantics and pragmatics: From
experiment to theory (pag. 276–316). Basingstoke, United
Kingdom: Palgrave Macmillan.
Visser, M., Jefferies, E., & Lambon Ralph, M. UN. (2010).
Semantic processing in the anterior temporal lobes: UN
meta-analysis of the functional neuroimaging literature.
Journal of Cognitive Neuroscience, 22, 1083–1094.
Vitello, S., & Rodd, J. M. (2015). Resolving semantic ambiguities
in sentences: Cognitive processes and brain mechanisms.
Language & Linguistic Compass, 9, 391–405.
Vitello, S., Warren, J. E., Devlin, J. T., & Rodd, J. M. (2014). Roles
of frontal and temporal regions in reinterpreting semantically
ambiguous sentences. Frontiers in Human Neuroscience,
8, 530.
Zaidel, D. W., Zaidel, E., Oxbury, S. M., & Oxbury, J. M.
(1995). The interpretation of sentence ambiguity in
patients with unilateral focal brain surgery. Brain and
Language, 51, 458–468.
Zempleni, M. Z., Renken, R., Hoeks, J. C. J., Hoogduin, J. M., &
Stowe, l. UN. (2007). Semantic ambiguity processing in
sentence context: Evidence from event-related fMRI.
Neuroimage, 34, 1270–1279.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
3
e
4
–
0
P
3
D
F
2
0
/
1
3
3
2
1
/
4
3
1
/
4
o
0
C
3
N
/
_
UN
1
_
8
0
6
1
1
4
1
9
0
3
2
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
9
0
3
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
F
.
T
o
N
0
5
M
UN
sì
2
0
2
1
MacGregor et al.
425