The Association between Gray Matter Volume and
Reading Proficiency: A Longitudinal Study of
Beginning Readers
Janosch Linkersdörfer1,2, Alina Jurcoane1,3, Sven Lindberg1,2,
Jochen Kaiser3, Marcus Hasselhorn1,2,3, Christian J. Fiebach1,3,4,
and Jan Lonnemann1,2
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
Astratto
■ Neural systems involved in the processing of written lan-
guage have been identified by a number of functional imaging
studies. Structural changes in cortical anatomy that occur in the
course of literacy acquisition, Tuttavia, remain largely unknown.
Here, we follow elementary school children over their first
2 years of formal reading instruction and use tensor-based mor-
phometry to relate reading proficiency to cortical volume at
baseline and follow-up measurement as well as to intraindividual
longitudinal volume development between the two measure-
ment time points. A positive relationship was found between
baseline gray matter volume in the left superior temporal gyrus
and subsequent changes in reading proficiency. Inoltre, UN
negative relationship was found between reading proficiency at
the second measurement time point and intraindividual cortical
volume development in the inferior parietal lobule and the pre-
central and postcentral gyri of the left hemisphere. These results
are interpreted as evidence that reading acquisition is associated
with preexisting structural differences as well as with experience-
dependent structural changes involving dendritic and synaptic
pruning. ■
INTRODUCTION
Reading is a relatively recent human cultural invention
that has to be taught explicitly and practiced intensively
to be mastered. As a consequence, no genetically pre-
disposed neural systems for reading exist. Invece, Imparare-
ing to read is thought to involve the use of a distributed
set of brain regions originally serving other purposes
(Dehaene, 2004).
The neural basis of reading has been investigated
extensively in functional imaging studies (for meta-
analyses, Vedere, per esempio., Bolger, Perfetti, & Schneider, 2005;
Jobard, Crivello, & Tzourio-Mazoyer, 2003; Turkeltaub,
Eden, Jones, & Zeffiro, 2002). Most consistently, these
studies identified three neural systems in the left hemi-
sphere to play an important role in the processing of
written language. The first system includes parts of the
posterior superior temporal gyrus and the inferior parie-
tal lobule and is assumed to support the mapping of
orthographic elements of written words (graphemes)
onto corresponding phonological elements (phonemes);
1Center for Individual Development and Adaptive Education of
Children at Risk (IDeA), Frankfurt am Main, Germany, 2German
Institute for International Educational Research, Frankfurt am
Main, Germany, 3Goethe University, Frankfurt am Main, Germany,
4Radboud University, Nijmegen, The Netherlands
© 2014 Istituto di Tecnologia del Massachussetts
a second system in the inferior frontal gyrus has been
associated with articulatory processes and the active
analysis of phonological elements (per esempio., Schlaggar &
McCandliss, 2007); and finally, a system in occipitotem-
poral regions, including parts of the extrastriate fusiform
and inferior temporal gyri, is considered to facilitate the
direct mapping between familiar visual words or frequent
letter combinations within words and corresponding se-
mantic and phonological representations (per esempio., McCandliss,
Cohen, & Dehaene, 2003; Cohen et al., 2000).
A common developmental hypothesis is that, in the
beginning of formal reading instruction, children pri-
marily make use of the temporoparietal system to decode
written words in a laborious letter-by-letter (or syllable-
by-syllable) maniera. Through continuous practice and
in tight coupling with the temporoparietal system, IL
occipitotemporal system is increasingly incorporated,
eventually resulting in automatic and fluent reading
performance (Pugh et al., 2001). These assumptions
have been supported by functional imaging studies
showing greater activity in children relative to adults in
the temporoparietal system (Church, Coalson, Lugar,
Petersen, & Schlaggar, 2008; Booth et al., 2001) E
longitudinal increases in activity over the first years of
reading instruction in the occipitotemporal system in
reading or reading-related tasks (Ben-Shachar, Dougherty,
Deutsch, & Wandell, 2011).
Journal of Cognitive Neuroscience 27:2, pag. 308–318
doi:10.1162/jocn_a_00710
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
C
4
N
5
_
4
UN
/
_
j
0
o
0
C
7
N
1
0
_
UN
P
_
D
0
0
B
7
sì
1
G
0
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
/
.
F
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Despite the large body of functional imaging research,
only a few studies have examined neurostructural cor-
relates of reading, questo è, how different levels of reading
skill relate to differences in cortical gray matter structure.
These studies have focused mainly on comparisons be-
tween control participants and persons suffering from
developmental dyslexia, a severe difficulty in learning to
read accurately and fluently. Although both reductions
and increases of gray matter properties have been reported
in dyslexic readers, the results of two recent coordinate-
based meta-analyses of voxel-based morphometry (VBM)
studies point to gray matter reductions in bilateral tem-
poroparietal (Richlan, Kronbichler, & Wimmer, 2013;
Linkersdörfer, Lonnemann, Lindberg, Hasselhorn, &
Fiebach, 2012), left occipitotemporal, and bilateral cere-
bellar (Linkersdörfer et al., 2012) regions as the most con-
sistent neurostructural correlates of dyslexia.
Studies investigating neurostructural correlates of
reading in nonimpaired populations are even more
sparse and so far yielded mixed results. More specifically,
a positive association between reading performance and
measures of gray matter volume, density, or thickness
has been demonstrated in bilateral (Blackmon et al.,
2010) or left (He et al., 2013; Steinbrink et al., 2008)
temporoparietal and left (Zhang et al., 2013) or right
(Hoeft et al., 2007) occipitotemporal regions. In addi-
zione, negative correlations have been reported for left
occipitotemporal and bilateral inferior frontal regions
(Blackmon et al., 2010) and the left posterior cingulate
cortex (He et al., 2013). Inoltre, as all of these
studies employed a cross-sectional design (cioè., participants
were studied at only one point in time), they cannot
answer the question of whether reading-related differences
in cortical anatomy arise in the course of reading instruc-
tion and represent the result of different levels of reading
proficiency or whether they can be regarded as a pre-
existing condition possibly causing differences in reading
proficiency. One VBM study (Carreiras et al., 2009) found
higher bilateral temporoparietal gray matter density in
late-literates (persons who acquired literacy as adults)
compared with illiterates, which supports the hypothesis
of experience-dependent structural changes. It remains
unclear, Tuttavia, whether structural changes associated
with reading acquisition in adulthood resemble those that
occur when reading is learned in childhood. Cortical gray
matter structure has been shown to undergo massive
changes throughout childhood and adolescence, domi-
nated mainly by neuronal and synaptic pruning processes,
which result in cortical thinning (per esempio., Huttenlocher &
Dabholkar, 1997). Pruning is thought to reflect experience-
dependent plastic changes, which is supported by studies
reporting an association between cortical thinning and
better cognitive performance (per esempio., Squeglia, Jacobus,
Sorg, Jernigan, & Tapert, 2013; Sowell et al., 2004).
The present study examined possible gray matter changes
associated with the acquisition of written language by follow-
ing elementary school children longitudinally in their
first 2 years of reading instruction. For this longitudinal
sample, we collected both standardized reading tests
and structural magnetic resonance imaging (MRI) dati.
To relate childrenʼs reading proficiency to cortical volume
differences at the baseline and follow-up measurements as
well as to differences in the intraindividual cortical volume
development between the two measurement time points,
we used tensor-based morphometry (TBM; Vedere, per esempio.,
Ashburner & Friston, 2003). In contrast to VBM, Quale
identifies regional neurostructural differences by seg-
mentation of tissue classes and statistical analysis of the
resulting tissue probability maps after nonlinear transfor-
mation to a common space, TBM derives information from
high-resolution deformation fields obtained from these
nonlinear transformations. In cross-sectional TBM studies,
participantsʼ structural images are nonlinearly transformed
to a common anatomical template; in longitudinal studies,
structural images of the same participant are nonlinearly
transformed to each other. From the resulting deformation
tensor fields, the Jacobian matrix is computed for each
voxel, whose determinant quantifies the fractional volume
expansion and contraction needed to align the two images.
These relative local volume differences between a partici-
pantʼs image and the template, questo è, the group average,
in a cross-sectional analysis or between images from the
same participant at different time points in a longitudinal
analysis are then, after transformation to a common space,
used in the statistical analysis. TBM is considered to offer
more precise and robust results than VBM, especially in
longitudinal analyses (Anderson et al., 2012; Hua et al.,
2009; Lee et al., 2007).
METHODS
Participants
Twenty-eight normally developing children participated
in the MRI procedure at both measurement time points.
The study was approved by the ethics committee of the
chamber of physicians of the state of Hessen, Germany,
and informed consent was obtained from all participating
children and their parents. Six children were excluded
from the analysis because of movement artifacts (Vedere
below), resulting in a final sample of 22 children (10 girls).
All children were in their first year of elementary school at
the first measurement time point (mean age = 7.5 years,
SD = 0.4 years) and in their second year of elementary
school at the second measurement time point (mean age =
8.4 years, SD = 0.4 years), with approximately 1 year in
between (mean time between measurements = 349 days,
SD = 45 days).
Cognitive Assessment
At both measurement time points, reading proficiency
was assessed by means of two standardized reading tests
targeting comprehension and fluency at the single word
Linkersdörfer et al.
309
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
C
4
N
5
_
4
UN
/
_
j
0
o
0
C
7
N
1
0
_
UN
P
_
D
0
0
B
7
sì
1
G
0
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
.
F
/
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
level. Comprehension ability was measured with the word
comprehension subtest of a German reading test with
norms for grades 1–6 (ELFE 1–6; Lenhard & Schneider,
2006). In this test, children see a picture and select the
appropriate written word out of four alternatives. Dis-
tractor words are phonologically and orthographically
similar to the target word and matched for length in sylla-
bles. The test consists of 72 items, and children solve as
many of them as possible in 3 min. Reading fluency was
measured with the fluency subtest of a further German
reading test with norms for grades 1–6 (SLRT-II; Moll &
Landerl, 2010). In this test, children read out loud as many
words as possible from a list of 156 existing words and a
second list of 156 pseudowords, with a time limit of 1 min
per list. Standardized test scores (T scores) for the two
reading tests were averaged to yield a composite score
representing a robust measure of the childʼs reading pro-
ficiency at the respective measurement time point. In
aggiunta, to assess intraindividual changes in reading
proficiency between the two measurement time points,
a change score was computed for each child by subtract-
ing the composite score at the first measurement time
point from the composite score at the second measure-
ment time point. As a measure of general intelligence,
Ravenʼs Colored Progressive Matrices (Bulheller & Häcker,
2002) were administered at the second measurement time
point. This test is an untimed power test designed for chil-
dren aged 4–11 years and consists of 36 colored diagram-
matic puzzles, each with a missing part that has to be
chosen from six alternatives. The number of correctly
solved items was used as a measure of intelligence in the
analysis. All cognitive measures were taken on a separate
day preceding the MRI scanning session.
Image Acquisition
Children were accustomed to the scanning procedure
and trained to lie still using a custom-built MRI simulator
(mock scanner) on the day of the cognitive assessment.
Structural MRI data were acquired with a 3-T Siemens
Magnetom Tim Trio scanner using a standard transmit–
receive head coil. The same protocol was used for each par-
ticipant and both measurement time points. T1-weighted
images were acquired using an optimized Modified Driven
Equilibrium Fourier Transform (Deichmann, Schwarzbauer,
& Turner, 2004) sequence (repetition time = 7.92 sec, echo
time = 2.48 sec, flip angle = 16°, FOV = 256 × 224 mm,
176 sagittal slices, voxel size = 1 × 1 × 1 mm). During
the scanning procedure, the children were instructed to
lie as still as possible and were able to watch a movie.
Image Processing and TBM
Individual structural images from both measurement
time points were visually inspected with regards to signal-
to-noise ratio and screened for movement artifacts by
two independent researchers. On the basis of these
inspections, six children had to be excluded from the anal-
ysis because of excessive movement artifacts. Image pre-
processing included removal of non-brain tissue using the
hybrid watershed/surface deformation procedure (Ségonne
et al., 2004) implemented in the Freesurfer software pack-
age (Fischl, 2012) and correction for field inhomogeneity
with the N4ITK algorithm (Tustison et al., 2010). Dopo
these steps, the images were submitted to two process-
ing pipelines: a cross-sectional pipeline assessing inter-
individual differences in cortical volume at the first and
second measurement time points, rispettivamente, and a lon-
gitudinal pipeline assessing interindividual differences in
intraindividual cortical volume development between the
two measurement time points. Processing was carried
out using the Advanced Normalisation Tools software
package (Avants et al., 2011).
Cross-sectional processing involved the creation of
group-specific optimal average templates for each of
the two measurement time points by iteratively refining
nonlinear coregistrations of all images from the respec-
tive measurement time point (see Klein et al., 2010). Dopo
a linear transformation, each individual image from one
measurement time point was then nonlinearly trans-
formed to the template from the same measurement time
point using the Symmetric Normalization algorithm
(Avants, Epstein, Grossman, & Gee, 2008) implemented
in the Advanced Normalisation Tools. For the resulting
deformation tensor field, the determinant of the Jacobian
matrix was computed, which quantifies the fractional
volume expansion and contraction required to match
the template at each voxel. The Jacobian maps were sub-
jected to a log transformation to render the values more
normally distributed (Avants & Gee, 2004). To allow for
group analysis of the Jacobian maps in a common refer-
ence space, the linear transformation between the res-
pective group template and the Montreal Neurological
Institute (MNI) space was computed and used to trans-
form the Jacobian map of each participant into MNI space.
Before statistical analysis, the Jacobian maps were
smoothed with a 4-mm FWHM Gaussian filter.
Longitudinal processing involved registration of each
participantʼs image from the second measurement time
point (follow-up scan) to his or her image from the first
measurement time point (baseline scan). To avoid poten-
tial bias resulting from asymmetric interpolation (Vedere, per esempio.,
Fox, Ridgway, & Schott, 2011), the linear transforma-
tion between the follow-up and the baseline image was
split halfway between the two images according to the
approach described by Yushkevich et al. (2010). IL
symmetric linear transformations from the respective
images to the halfway space were then used to initialize
the nonlinear registration of the follow-up to the base-
line image. Jacobian determinants of the resulting defor-
mation field, in this case indicating the voxel-wise
fractional volume expansion and contraction between the
two measurement time points in the respective participant,
were computed and log transformed. The Jacobian map of
310
Journal of Cognitive Neuroscience
Volume 27, Numero 2
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
C
4
N
5
_
4
UN
/
_
j
0
o
0
C
7
N
1
0
_
UN
P
_
D
0
0
B
7
sì
1
G
0
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
T
/
.
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
each participant was finally transformed into MNI space by
combining (UN) the linear transformation from the halfway
space to the baseline image space with (B) the linear and
nonlinear transformations from the baseline image space
to the baseline group template space and (C) the linear
transformation from the baseline group template space to
the MNI space, which had been computed in the cross-
sectional processing pipeline. Finalmente, the Jacobian maps
were smoothed with a 4-mm FWHM Gaussian filter.
Statistical Analysis
Whole-brain voxel-wise regression analysis was performed
on the smoothed Jacobian images using the SPM8 software
package (Wellcome Trust Centre for Neuroimaging, Uni-
versity College London, London, United Kingdom). Primo,
cortical volume differences at the first measurement time
point (T1) were related to (UN) the composite score for read-
ing proficiency at T1, (B) the composite score for reading
proficiency at the second measurement time point (T2),
E (C) the change in reading proficiency between T1
and T2 as variable of interest. Participantʼs intelligence
score, genere, and age at T1 in days were included in the
model as nuisance variables. Secondo, cortical volume dif-
ferences at T2 were related to the composite score for
reading proficiency at T2, and participantʼs intelligence
score, genere, and age at T2 in days were included in the
model as nuisance variables. Finalmente, differences in the
intraindividual cortical volume development between T1
and T2 were related to the composite score for reading
proficiency at T2, and participantʼs intelligence score,
genere, age at T1, and time interval between T1 and T2,
both measured in days, were included in the model as
nuisance variables.
For all group-level statistics, a gray matter tissue prob-
ability map, adjusted for the age and gender distribution
of the sample, was generated using the Template-O-Matic
toolbox (Wilke, Holland, Altaye, & Gaser, 2008), thresh-
olded at p = .25 and binarized to yield an explicit gray
matter mask that was applied to the statistical analysis.
Clusters of voxels were considered significant and reported
when exceeding a voxel level threshold of p < .001 (uncor-
rected) and a cluster size threshold of p < .05, corrected for
multiple comparison using Gaussian random field theory
(Friston, Worsley, Frackowiak, Mazziotta, & Evans, 1994).
Analysis of behavioral performance and post hoc anal-
yses of the TBM results were carried out using the statis-
tical package R, version 2.15.1 (R Project for Statistical
Computing, Vienna, Austria; R Core Team, 2012).
RESULTS
Behavioral Performance
Reading proficiency in our sample, measured by compos-
ite scores combining comprehension and fluency perfor-
mance, varied between T scores of 42.25 and 69 (mean =
57.10, SD = 7.13) at T1 and between 39 and 66.5 (mean =
53.92, SD = 6.79) at T2, indicating that childrenʼs reading
proficiency varied over the whole range of normal com-
petencies, with a tendency toward above-average per-
formance. We observed high correlations between the
different reading (sub)tests (i.e., comprehension, fluency-
subtest words, fluency-subtest pseudowords) at both
measurement time points (T1: r = .7–.83; T2: r = .6–.66;
all ps < .005), supporting our decision to integrate them
into a single composite score. Moreover, reading perfor-
mance over the two measurement time points was highly
correlated (composite score, r = .8; comprehension, r = .73;
fluency-subtest words, r = .69; fluency-subtest pseudo-
words, r = .75; all ps < .001). Reading proficiency change
scores varied between T scores of −11.75 and 5 (mean =
−3.18, SD = 4.44). The mean decrease in reading profi-
ciency scores between T1 and T2 might, at least in part,
be attributable to the fact that reading proficiency scores
in our sample were relatively high at T1; thus, subsequent
measures were more likely to result in lower values, a statis-
tical phenomenon known as “regression toward the mean”
(e.g., Bland & Altman, 1994). A moderate negative relation-
ship between the reading composite score at T1 and the
reading proficiency change score (r = −.39, p = .08) sup-
ports this assumption. Intelligence scores (IQ) ranged from
88 to 135 (mean = 107.23, SD = 11.33), indicating, again,
normal to above-average abilities in our sample. At both
measurement time points, reading proficiency did not cor-
relate significantly with intelligence (measurement time
point 1: r = .08, p = .726; measurement time point 2: r =
.05, p = .837) or sex (coded as 0 for girls and 1 for boys;
measurement time point 1: point biserial r = −.4, p =
.07; measurement time point 2: point biserial r = −.31,
p = .165).
TBM
A cluster showing a positive relationship between gray
matter volume at T1 and change in reading proficiency
scores between T1 and T2 could be found in the left
superior temporal gyrus (cluster extent = 1469 voxels,
cluster level p = .034 corrected, peak t in cluster = 5.93;
cf. Table 1 and Figure 1). The analyses relating volume
differences at T1 to reading proficiency at T1 or T2 and
the analysis relating volume differences at T2 to reading
proficiency at T2 did not result in significant findings. To
exclude the possibility that the significant cluster indi-
cating a larger volume of left superior temporal areas in
children with higher reading proficiency change scores
was solely based on differences in the baseline level of
reading proficiency, we computed the partial correlation
between the mean Jacobian value, averaged over all
voxels contained in the cluster, and the reading change
score, while controlling for reading proficiency at T1
and the same variables as in the whole-brain analysis
Linkersdörfer et al.
311
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. Significant Cluster Showing a Positive Association between Gray Matter Volume at the First Measurement Time Point and
Change in Reading Proficiency
MNI Coordinates of
Local Maxima
Region
L superior temporal gyrus
aSubpeak within cluster.
x
−64
−64
y
−38
−27
z
10
13
t
5.93
5.55
Cluster Size ( Voxels)
Cluster p Value
1469
a
.034
a
(i.e., intelligence score, sex, age at T1). The results of this
post hoc analysis were significant (r partial = 0.83, p <
.001), indicating that gray matter volume in the superior
temporal cluster at T1 was indeed related to changes in
reading proficiency, even when accounting for baseline
level in reading proficiency. Furthermore, to investigate
whether cortical volume in the superior temporal cluster
was related differently to developmental changes in the
separate reading (sub)tests (i.e., comprehension, fluency-
subtest words, fluency-subtest pseudowords) summarized
in the reading change score, partial correlations between
change scores for the separate (sub)tests and the mean
Jacobian value, averaged over all voxels contained in the clus-
ter, were computed, while, again, controlling for the same
variables as in the whole-brain analysis. The results show
significant positive correlations for all reading (sub)tests
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
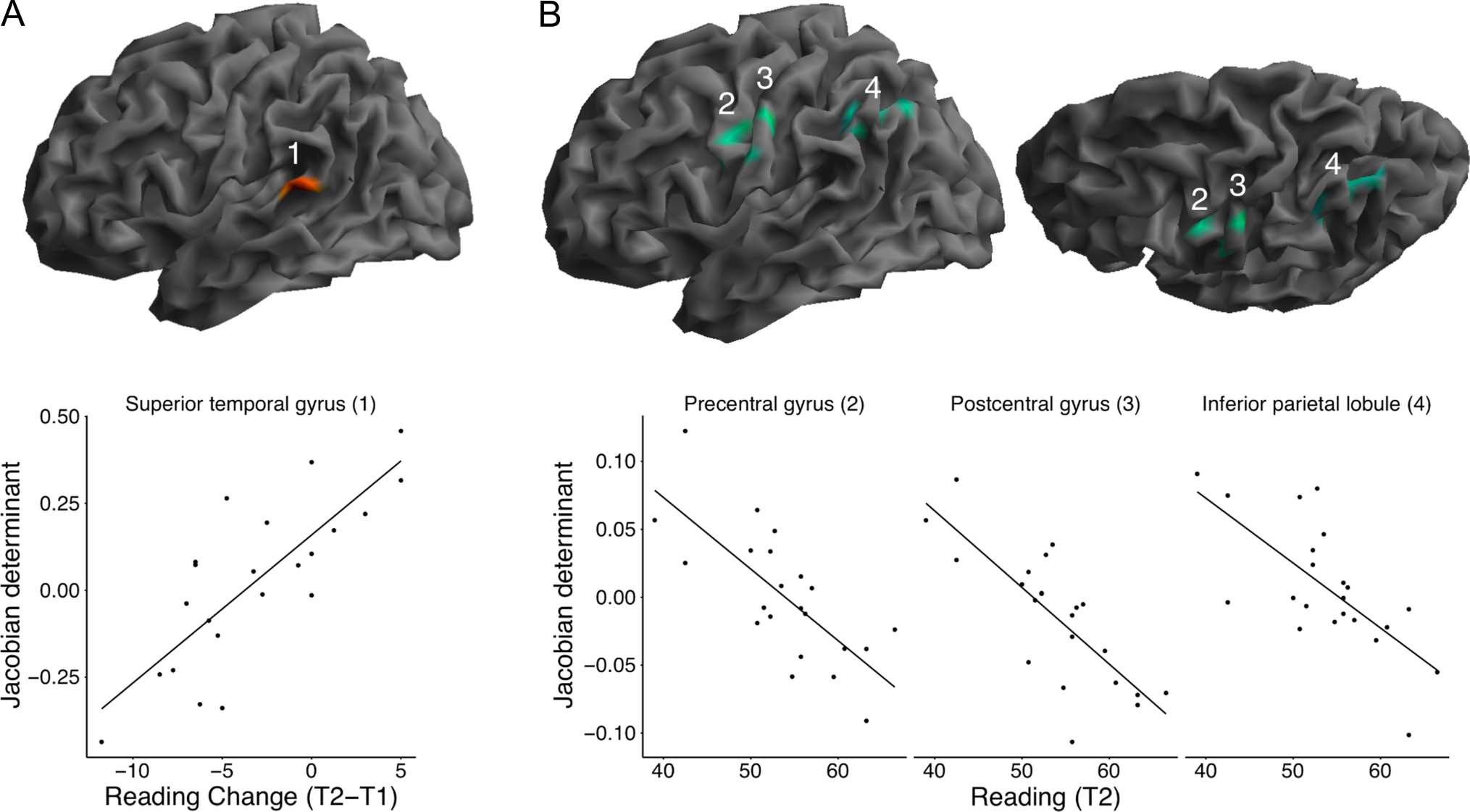
Figure 1. Associations between cortical volume and reading proficiency. (A, top) Surface rendering of the left brain hemisphere displaying the
cluster (red) in the superior temporal gyrus showing a positive association between gray matter volume at the first measurement time point (T1) and
change in reading proficiency between the first and second measurement time points (T2). (A, bottom) Relationship between reading proficiency
change (change score subtracting the composite score of standardized reading tests at T1 from the composite score at T2) and the gray matter
volume at T1 (log-transformed Jacobian determinants, averaged over all voxels in the cluster; positive values indicate relatively larger volume, and
negative values indicate relatively smaller volume compared with the group average). (B, top) Surface renderings showing the three left-hemispheric
clusters (green) that display a negative association between gray matter volume change between T1 and T2 and reading proficiency at T2, in the
precentral (2) and postcentral (3) gyri and in the inferior parietal lobule (3). (B, bottom) Scatterplots illustrating the relationship between reading
proficiency (composite score of standardized reading tests at T2) and gray matter volume development between T1 and T2 (log-transformed
Jacobian determinants, averaged over all voxels in the respective clusters; positive values indicate relative volume expansion, and negative values
represent relative volume contraction). For purposes of visualization, significant effects (cf. Tables 1 and 2) are displayed at p < .0025.
312
Journal of Cognitive Neuroscience
Volume 27, Number 2
Table 2. Significant Clusters Showing a Negative Association between Gray Matter Volume Development and Reading Proficiency at
the Second Measurement Time Point
MNI Coordinates of
Local Maxima
x
−47
−45
−52
−56
−59
−36
−44
−49
y
−7
−2
0
−18
−11
−62
−55
−48
z
37
28
24
42
29
40
37
35
t
−5.60
−4.94
−4.02
−5.61
−4.82
−5.15
−4.83
−5.08
Cluster Size ( Voxels)
Cluster p Value
743
a
a
603
a
978
a
a
.018
a
a
.049
a
.004
a
a
Region
L precentral gyrus
L postcentral gyrus
L angular gyrus
L supramarginal gyrus
aSubpeak within cluster.
(r partial = .59–.7, all ps < .005), indicating that the differ-
ent subtests were equally associated with gray matter vol-
ume in the cluster.
Relating differences in the intraindividual cortical vol-
ume development between the two measurement time
points to reading proficiency at T2 resulted in three sig-
nificant clusters showing a negative relationship (Table 2;
Figure 1). The largest cluster (cluster extent = 978 vox-
els, cluster level p = .004 corrected, peak t in cluster =
−5.15) was located in the left inferior parietal lobule,
including mainly the supramarginal and angular gyri.
The second cluster was found in the left precentral gyrus
(cluster extent = 743 voxels, cluster level p = .018
corrected, peak t in cluster = −5.6), and the third cluster
was located in the left postcentral gyrus (cluster extent =
603 voxels, cluster level p = .049 corrected, peak t in
cluster = −5.61). No significant clusters showing a posi-
tive relationship between reading proficiency and vol-
ume development were found. Possible differences in
the relationship between cortical development in the
three clusters and performance in the separate reading
(sub)tests (i.e., comprehension, fluency-subtest words,
fluency-subtest pseudowords), which were summarized
in the composite score for reading proficiency, were
examined by computing partial correlations between
T scores for the separate reading (sub)tests at T2 and
mean Jacobian values for the separate clusters, aver-
aged over all voxels contained in the respective cluster,
including the same nuisance variables as in the whole-
brain analysis (i.e., intelligence score, gender, age at
T1, and time interval between T1 and T2). The results
show significant negative correlations for all reading
(sub)tests and clusters (r partial = −.62 to −.75, all
ps < .005), indicating that the different (sub)tests were
equally associated with volume development in the three
clusters.
DISCUSSION
In this study, we used TBM to investigate the relationship
between reading proficiency and cortical gray matter
volume in elementary school children during their first
2 years of formal reading instruction. Higher baseline
gray matter volume in the left superior temporal cortex
was positively related to subsequent changes in reading
proficiency. Moreover, longitudinal volume reductions in
the inferior parietal lobule and the precentral and post-
central gyri of the left hemisphere were related to higher
reading proficiency at the follow-up measurement.
The significant cluster showing a positive relationship
between gray matter volume at the first measurement
time point and change in reading proficiency between
the two measurement time points was located in the left
posterior superior temporal gyrus, including parts of the
planum temporale. These areas, which lie directly poste-
rior to the primary auditory cortex, are considered as part
of “Wernickeʼs area” and have been implicated in differ-
ent language functions (for an overview, see, e.g., Price,
2012). The planum temporale, because of its marked left-
ward asymmetry, has traditionally been considered as an
auditory association area specific to speech perception
(Geschwind & Levitsky, 1968). More recent research,
however, has shown that this region is equally activated
by speech and tone stimuli (Binder et al., 2000; Binder,
Frost, Hammeke, Rao, & Cox, 1996). Furthermore, the
planum temporale, especially its posterior portions near
the TPJ, seems to be involved in both speech perception
and speech production (Hickok, Okada, & Serences,
2009; Wise et al., 2001) but also in the audio-motor inter-
action between nonspeech sounds and motor actions
(Hickok, Buchsbaum, Humphries, & Muftuler, 2003).
Therefore, current models of speech processing view
the planum temporale as a sensorimotor integration area,
Linkersdörfer et al.
313
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
which relays information between auditory cortical re-
gions and anterior systems involved in motor control of
the vocal tract and which therefore plays an important
role in the acquisition of spoken language (Hickok,
Houde, & Rong, 2011). The association between the
gray matter volume of this area in beginning readers
and subsequent changes in their reading proficiency, as
demonstrated by our study, suggests that the planum
temporale might also be involved in the acquisition of
written language. This assumption is supported by func-
tional imaging studies reporting overlapping left planum
temporale activation for listening to and reading words
(Buchsbaum et al., 2005; Nakada, Fujii, Yoneoka, &
Kwee, 2001). Therefore, this region might facilitate the
multisensory integration of visual input and auditory–
motor representations.
The left inferior parietal cluster, which exhibited a neg-
ative association between cortical volume development
and reading proficiency at the second measurement time
point, was located in the supramarginal and angular gyri,
areas that correspond well to the anatomical characteri-
zation of the temporoparietal part of the phonological
reading system (e.g., Démonet, Taylor, & Chaix, 2004;
Rumsey et al., 1997). The temporoparietal system is
assumed to support the transformation of orthographic
elements of visual words (graphemes) into associated
phonological elements (phonemes; Booth et al., 2002;
Shaywitz et al., 2002). Thus, structural changes in this re-
gion over the first years of reading instruction, as demon-
strated by our study, are in line with developmental
accounts of reading acquisition, suggesting that children
rely mainly on phonological recoding strategies in the
beginning stages of learning to read (Ehri, 1999; Frith,
1985; Chall, 1983). Furthermore, our results are in agree-
ment with the neuroanatomical model of reading devel-
opment by Pugh et al. (2001), which suggests a predominant
role of the temporoparietal system in beginning readers, a
claim that has been supported by findings from functional
imaging studies showing greater activity in children rela-
tive to adults in these regions in response to reading or
reading-related tasks (Church et al., 2008; Booth et al.,
2001).
Two additional clusters showing negative associations
between volume development and reading proficiency
were found in the precentral and postcentral gyri of the
left hemisphere. Neural populations in proximity to the
peaks of these clusters have been attributed to the sen-
sorimotor control of movements of the lips, tongue, jaw,
and larynx (e.g., Grabski et al., 2012; Bohland &
Guenther, 2006; Pulvermüller et al., 2006) and have been
shown to produce speech through highly coordinated
sequences of spatial patterns of activity that hierarchically
represent the articulatory features of phonemes (Bouchard,
Mesgarani, Johnson, & Chang, 2013). Furthermore, activity
in these speech production areas, especially in the left
hemisphere, has also been demonstrated for the passive
perception of speech sounds (Pulvermüller et al., 2006;
Wilson, Saygin, Sereno, & Iacoboni, 2004), supporting
theories postulating a motor involvement in speech per-
ception (Galantucci, Fowler, & Turvey, 2006; Liberman,
Cooper, Shankweiler, & Studdert-Kennedy, 1967). These
findings have been interpreted as evidence for the exis-
tence of cell assemblies integrating articulatory and auditory
neurons to represent the phonological properties of
speech sounds, which might develop in response to an
increased tuning to the phonemic structure of the native lan-
guage in the early stages of learning to speak (Pulvermüller
& Fadiga, 2010). The formation of such an auditory–motor
network might be facilitated by the multisensory integra-
tion function of the planum temporale discussed above.
The left precentral gyrus has further been implicated as
part of the neural reading network by meta-analyses of
functional imaging studies investigating nonimpaired
adults (Bolger et al., 2005; Turkeltaub et al., 2002) and
children (Houdé, Rossi, Lubin, & Joliot, 2010). Thus, neural
representations of articulatory features of phonemes also
seem to play an important role in the processing of written
language.
Indeed, learning to read an alphabetic script seems to
increase childrenʼs awareness of the phonological struc-
ture of spoken language through the incorporation of
orthographic information, ultimately leading to a reorgani-
zation of existing phonological representations (Nation &
Hulme, 2011; Pattamadilok, Knierim, Kawabata Duncan,
& Devlin, 2010; Castro-Caldas, Petersson, Reis, Stone-
Elander, & Ingvar, 1998). The observed structural changes
in the precentral and postcentral gyri of the left hemi-
sphere, which co-occur with similar changes in the left infe-
rior parietal lobule, might therefore be interpreted as the
neural correlate of such reorganizational processes. In
beginning readers, the intensive use of phonological recod-
ing strategies implemented by neural circuits in the left
inferior parietal cortex may lead to the mapping of visual
(i.e., orthographic) features of words onto preexisting pho-
nological representations in left hemisphere circuits sub-
serving speech perception and production (Pattamadilok
et al., 2010). The formation of a higher-order neural
assembly that integrates orthographic and phonological
information may promote the reciprocal refinement of
grapheme–phoneme conversion knowledge and articulatory–
auditory representations of phonemes and might possibly
be further strengthened by the coactivation of inferior
parietal with precentral and postcentral neurons because
of lip movements and subvocal articulation, typically found
in children in the beginning stages of learning to read (e.g.,
Huey, 1968).
Previous studies that related reading performance to
measures of gray matter volume, gray matter density,
or gray matter thickness in normal reading participants
in a cross-sectional manner, that is, at only one point in
time, reported mostly positive associations in temporo-
parietal (He et al., 2013; Blackmon et al., 2010; Steinbrink
et al., 2008) and occipitotemporal regions (Zhang et al.,
2013; Hoeft et al., 2007). In contrast to these studies, no
314
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
association between gray matter volume at baseline or
follow-up measurement and reading proficiency at the
respective (or the following) measurement time point
could be found in this study. However, intraindividual
changes in reading proficiency between the two mea-
surement time points were positively related to baseline
gray matter volume in the superior temporal gyrus, in-
cluding parts of the planum temporale, that is, similar
areas as reported by some of the studies cited above (i.e.,
He et al., 2013; Blackmon et al., 2010; Steinbrink et al.,
2008). Furthermore, it resonates with studies reporting
neurostructural differences between normal reading par-
ticipants and participants suffering from developmental
dyslexia, a specific impairment affecting the acquisition of
reading skills. Most specifically, a recent meta-analysis of
VBM studies (Richlan et al., 2013) found a cluster of reliable
gray matter reductions in dyslexic readers in superior
temporal regions only slightly more inferior to the superior
temporal cluster reported in this study. Moreover, dyslexic
readers have been reported to not show the typical leftward
asymmetry of the planum temporale (e.g., Hynd, Semrud-
Clikeman, Lorys, Novey, & Eliopulos, 1990; Larsen, Høien,
Lundberg, & Odegaard, 1990), although reports have been
inconsistent (for a review, see Heim & Keil, 2004). Struc-
tural abnormalities of the planum temporale and sur-
rounding areas have further been associated with other
developmental speech and language disorders like specific
language impairment (e.g., Badcock, Bishop, Hardiman,
Barry, & Watkins, 2012) or stuttering (e.g., Foundas et al.,
2004). We therefore interpret the significant positive rela-
tionship between left superior temporal gray matter
volume and subsequent changes in reading proficiency as
evidence for preexisting neurostructural differences, which
might be associated with differences in general linguistic
abilities. These abilities may be of increasing importance
in the course of long-term reading development (e.g.,
Dickinson, Golinkoff, & Hirsh-Pasek, 2010), which might
explain why, in contrast to previous studies investigating
neurostructural correlates of reading in older participants,
no association between interindividual differences in gray
matter volume in this area and reading proficiency mea-
sured at the respective time point could be found in the
beginning readers investigated in this study.
Our results further demonstrated a negative relation-
ship between longitudinal cortical volume development
and reading proficiency at the follow-up measurement
in inferior parietal and precentral and postcentral regions.
Whereas better readers on average exhibited volume
reductions, poor readers on average exhibited volume
growth in these areas. These findings seem inconsistent
with previous findings of positive correlations between
measures of cortical volume and reading proficiency in left
inferior parietal regions, as reported by some of the stud-
ies cited above (i.e., He et al., 2013; Blackmon et al., 2010).
Furthermore, the only previous study that reports volume
changes directly related to reading acquisition found
higher gray matter density in temporoparietal regions in
persons who acquired literacy as adults when compared
with illiterate adults (Carreiras et al., 2009). It remains
unclear, however, whether the processes of neural plas-
ticity underlying learning in adulthood are the same as
those underlying learning in childhood. Although changes
in cortical anatomy can be regarded as the product of a
mixture of developmental and experiential processes at
any point in development, the influence of these processes
may vary over time (Galván, 2010). We thus hypothesize
that the observed discrepancies may be because of the
age and/or the developmental stage of the participants
examined in this study: Cortical development is character-
ized by an excessive increase in brain connectivity through
dendritic branching and the formation of new synapses in
infancy and early childhood, followed by a phase domi-
nated by dendritic and synaptic pruning in late childhood
and adolescence (e.g., Huttenlocher & Dabholkar, 1997).
This pruning phase is thought to reflect the experience-
dependent strengthening of frequently used and weaken-
ing or elimination of sparsely used connections; it might
mark a shift from relatively diffuse cortical representations
of cognitive functions in early childhood toward a more
accurate, efficient, and faster, that is, more adult like, pro-
cessing in specialized neural networks, which are further
refined throughout life (Huttenlocher, 2002). We suggest
that the observed cortical volume reductions in better
readers over the first 2 years of elementary school rep-
resent the formation of a more mature and fine-tuned,
left-lateralized cortical network for the processing of pho-
nological features of speech sounds.
This assumption is in line with recent studies that re-
port an association between cortical thinning and cogni-
tive performance. For example, Sowell et al. (2004) found
a negative association between longitudinal cortical thick-
ness development and vocabulary knowledge in elemen-
tary school children in the left dorsal frontal and parietal
lobes. Similarly, thinner parietal cortices were reported to
be associated with better performance in verbal learning
and memory, visuospatial functioning, and spatial plan-
ning and problem solving (Squeglia et al., 2013). Further-
more, thinner, that is, more mature, cortical gray matter
seems to be associated with more mature patterns of
brain activity and increased functional selectivity in cogni-
tive tasks (Wendelken, OʼHare, Whitaker, Ferrer, & Bunge,
2011; Lu et al., 2009). Thus, pruning processes might be of
special importance in younger children at the beginning of
reading development and may lay out the basic neural
foundation of the reading network. In later stages of read-
ing development, where learning builds on largely defined
neural and cognitive systems and overall plasticity is re-
duced, the reading network may be consolidated through
increased network-internal connectivity. This might be
accompanied by increased dendritic branching and syn-
apse formation (e.g., Draganski & May, 2008), which may
explain the findings of higher cortical volume in better
readers in studies examining older participants. This, how-
ever, we suggest may be a plasticity-related process that is
Linkersdörfer et al.
315
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
secondary to the cortical volume changes we observed in
the early stages of reading acquisition. The observed differ-
ences in volume development between good and poor
readers might be due to interindividual differences in
the coordination of pruning and growth processes (cf.
Yeatman, Dougherty, Ben-Shachar, & Wandell, 2012).
Although in better readers, pruning might be more prom-
inent and result in the faster formation of more mature and
efficient neural reading circuits, growth processes might
dominate development in the poor readers resulting in
more diffuse cortical representations. These experience-
dependent neurostructural changes in the beginning of
reading instruction might be transient and vanish in the
course of development.
To conclude, this study investigated neurostructural
correlates of reading in beginning readers by following
elementary school children over their first 2 years of
reading instruction. A positive relationship was found be-
tween left superior temporal gray matter volume at the
baseline measurement and subsequent changes in read-
ing proficiency. This finding may represent preexisting
neurostructural differences that might be associated with
long-term potential for the development of reading and
other language skills. In addition, a negative relationship
was found between cortical volume development and
reading proficiency at the follow-up measurement in cor-
tical areas in the inferior parietal lobule and precentral
and postcentral gyri of the left hemisphere. These results
might represent experience-dependent neuroanatomical
changes due to synaptic and dendritic pruning and, thus,
highlight the importance of developmental studies using
longitudinal designs, especially in children in the begin-
ning stages of learning to read.
Acknowledgments
This research was funded by the Hessian initiative for the devel-
opment of scientific and economic excellence (LOEWE). C. J. F.
is furthermore supported by an Emmy–Noether Grant from the
German Research Foundation (DFG FI-848/3) and by a VIDI
Grant from the Netherlands Organization for Scientific Research
(45209006). We would like to thank all the participating children
and their families for their support.
Reprint requests should be sent to Janosch Linkersdörfer,
German Institute for International Educational Research,
Schloßstraße 29, 60486 Frankfurt am Main, Germany, or via
e-mail: j.linkersdoerfer@idea-frankfurt.eu.
REFERENCES
Anderson, V. M., Schott, J. M., Bartlett, J. W., Leung, K. K.,
Miller, D. H., & Fox, N. C. (2012). Gray matter atrophy rate as
a marker of disease progression in AD. Neurobiology of
Aging, 33, 1194–1202.
Ashburner, J., & Friston, K. J. (2003). Morphometry. In
R. Frackowiak (Ed.), Human brain function (pp. 707–722).
London: Elsevier Press.
Avants, B. B., Epstein, C. L., Grossman, M., & Gee, J. C. (2008).
Symmetric diffeomorphic image registration with cross-
correlation: Evaluating automated labeling of elderly and
neurodegenerative brain. Medical Image Analysis, 12,
26–41.
Avants, B. B., & Gee, J. C. (2004). Geodesic estimation for large
deformation anatomical shape averaging and interpolation.
Neuroimage, 23(Suppl. 1), S139–S150.
Avants, B. B., Tustison, N. J., Song, G., Cook, P. A., Klein, A., &
Gee, J. C. (2011). A reproducible evaluation of ANTs similarity
metric performance in brain image registration. Neuroimage,
54, 2033–2044.
Badcock, N. A., Bishop, D. V. M., Hardiman, M. J., Barry, J. G., &
Watkins, K. E. (2012). Co-localisation of abnormal brain
structure and function in specific language impairment.
Brain and Language, 120, 310–320.
Ben-Shachar, M., Dougherty, R. F., Deutsch, G. K., & Wandell,
B. A. (2011). The development of cortical sensitivity to visual
word forms. Journal of Cognitive Neuroscience, 23,
2387–2399.
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S.,
Springer, J. A., Kaufman, J. N., et al. (2000). Human temporal
lobe activation by speech and nonspeech sounds. Cerebral
Cortex, 10, 512–528.
Binder, J. R., Frost, J. A., Hammeke, T. A., Rao, S. M., & Cox,
R. W. (1996). Function of the left planum temporale in
auditory and linguistic processing. Brain, 119, 1239–1247.
Blackmon, K., Barr, W. B., Kuzniecky, R., Dubois, J., Carlson, C.,
Quinn, B. T., et al. (2010). Phonetically irregular word
pronunciation and cortical thickness in the adult brain.
Neuroimage, 51, 1453–1458.
Bland, J. M., & Altman, D. G. (1994). Regression towards the
mean. British Medical Journal, 308, 1499.
Bohland, J. W., & Guenther, F. H. (2006). An fMRI investigation
of syllable sequence production. Neuroimage, 32, 821–841.
Bolger, D. J., Perfetti, C. A., & Schneider, W. (2005). Cross-
cultural effect on the brain revisited: Universal structures plus
writing system variation. Human Brain Mapping, 25,
92–104.
Booth, J. R., Burman, D. D., Meyer, J. R., Gitelman, D. R.,
Parrish, T. B., & Mesulam, M. M. (2002). Functional anatomy
of intra- and cross-modal lexical tasks. Neuroimage, 16, 7–22.
Booth, J. R., Burman, D. D., Van Santen, F. W., Harasaki, Y.,
Gitelman, D. R., Parrish, T. B., et al. (2001). The development
of specialized brain systems in reading and oral-language.
Child Neuropsychology, 7, 119–141.
Bouchard, K. E., Mesgarani, N., Johnson, K., & Chang, E. F.
(2013). Functional organization of human sensorimotor
cortex for speech articulation. Nature, 495, 327–332.
Buchsbaum, B. R., Olsen, R. K., Koch, P. F., Kohn, P.,
Kippenhan, J. S., & Berman, K. F. (2005). Reading, hearing,
and the planum temporale. Neuroimage, 24, 444–454.
Bulheller, S., & Häcker, H. O. (2002). Coloured Progressive
Matrices (CPM). Deutsche bearbeitung und normierung
nach J. C. Raven. Frankfurt, Germany: Pearson Assessment.
Carreiras, M., Seghier, M. L., Baquero, S., Estévez, A., Lozano, A.,
Devlin, J. T., et al. (2009). An anatomical signature for literacy.
Nature, 461, 983–986.
Castro-Caldas, A., Petersson, K. M., Reis, A., Stone-Elander, S., &
Ingvar, M. (1998). The illiterate brain. Learning to read and
write during childhood influences the functional organization
of the adult brain. Brain, 121, 1053–1063.
Chall, J. S. (1983). Stages of reading development. New York:
McGraw Hill.
Church, J. A., Coalson, R. S., Lugar, H. M., Petersen, S. E., &
Schlaggar, B. L. (2008). A developmental fMRI study of
reading and repetition reveals changes in phonological and
visual mechanisms over age. Cerebral Cortex, 18, 2054–2065.
Cohen, L., Dehaene, S., Naccache, L., Lehéricy, S., Dehaene-
Lambertz, G., Hénaff, M. A., et al. (2000). The visual word
316
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
form area: Spatial and temporal characterization of an initial
stage of reading in normal subjects and posterior split-brain
patients. Brain, 123, 291–307.
Dehaene, S. (2004). Evolution of human cortical circuits for
reading and arithmetic: The “neuronal recycling” hypothesis.
In S. Dehaene, J. R. Duhamel, M. Hauser, & G. Rizzolatti
(Eds.), From monkey brain to human brain (pp. 133–158).
Cambridge, MA: MIT Press.
Deichmann, R., Schwarzbauer, C., & Turner, R. (2004).
Optimisation of the 3D MDEFT sequence for anatomical
brain imaging: Technical implications at 1.5 and 3 T.
Neuroimage, 21, 757–767.
Démonet, J. F., Taylor, M. J., & Chaix, Y. (2004). Developmental
dyslexia. Lancet, 363, 1451–1460.
integration for speech processing. Journal of Neurophysiology,
101, 2725–2732.
Hoeft, F., Ueno, T., Reiss, A. L., Meyler, A., Whitfield-Gabrieli, S.,
Glover, G. H., et al. (2007). Prediction of childrenʼs reading
skills using behavioral, functional, and structural neuroimaging
measures. Behavioral Neuroscience, 121, 602–613.
Houdé, O., Rossi, S., Lubin, A., & Joliot, M. (2010). Mapping
numerical processing, reading, and executive functions in the
developing brain: An fMRI meta-analysis of 52 studies
including 842 children. Developmental Science, 13, 876–885.
Hua, X., Leow, A. D., Levitt, J. G., Caplan, R., Thompson, P. M.,
& Toga, A. W. (2009). Detecting brain growth patterns in
normal children using tensor-based morphometry. Human
Brain Mapping, 30, 209–219.
Dickinson, D. K., Golinkoff, R. M., & Hirsh-Pasek, K. (2010).
Huey, E. (1968). The psychology and pedagogy of reading.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
Speaking out for language: Why language is central to reading
development. Educational Researcher, 39, 305–310.
Draganski, B., & May, A. (2008). Training-induced structural
changes in the adult human brain. Behavioural Brain
Research, 192, 137–142.
Ehri, L. C. (1999). Phases of development in learning to read
words. In J. Oakhill & R. Beard (Eds.), Reading development
and the teaching of reading: A psychological perspective
(pp. 79–108). Oxford: Blackwell Science.
Fischl, B. (2012). FreeSurfer. Neuroimage, 62, 774–781.
Foundas, A. L., Bollich, A. M., Feldman, J., Corey, D. M., Hurley,
M., Lemen, L. C., et al. (2004). Aberrant auditory processing
and atypical planum temporale in developmental stuttering.
Neurology, 63, 1640–1646.
Fox, N. C., Ridgway, G. R., & Schott, J. M. (2011). Algorithms,
atrophy and Alzheimerʼs disease: Cautionary tales for clinical
trials. Neuroimage, 57, 15–18.
Friston, K. J., Worsley, K. J., Frackowiak, R. S. J., Mazziotta, J. C.,
& Evans, A. C. (1994). Assessing the significance of focal
activations using their spatial extent. Human Brain
Mapping, 1, 210–220.
Frith, U. (1985). Beneath the surface of developmental dyslexia.
In K. E. Patterson, J. C. Marshall, & M. Coltheart (Eds.),
Surface dyslexia (pp. 301–330). London: Routledge and
Kegan Paul.
Galantucci, B., Fowler, C. A., & Turvey, M. T. (2006). The motor
theory of speech perception reviewed. Psychonomic Bulletin
& Review, 13, 361–377.
Galván, A. (2010). Neural plasticity of development and
learning. Human Brain Mapping, 31, 879–890.
Geschwind, N., & Levitsky, W. (1968). Human brain: Left–right
asymmetries in temporal speech region. Science, 161,
186–187.
Grabski, K., Lamalle, L., Vilain, C., Schwartz, J.-L., Vallée, N.,
Tropres, I., et al. (2012). Functional MRI assessment of
orofacial articulators: Neural correlates of lip, jaw, larynx, and
tongue movements. Human Brain Mapping, 33, 2306–2321.
He, Q., Xue, G., Chen, C., Chen, C., Lu, Z.-L., & Dong, Q.
(2013). Decoding the neuroanatomical basis of reading
ability: A multivoxel morphometric study. Journal of
Neuroscience, 33, 12835–12843.
Heim, S., & Keil, A. (2004). Large-scale neural correlates of
developmental dyslexia. European Child and Adolescent
Psychiatry, 13, 125–140.
Hickok, G., Buchsbaum, B., Humphries, C., & Muftuler, T.
(2003). Auditory–motor interaction revealed by fMRI:
Speech, music, and working memory in area Spt. Journal of
Cognitive Neuroscience, 15, 673–682.
Hickok, G., Houde, J., & Rong, F. (2011). Sensorimotor
integration in speech processing: Computational basis and
neural organization. Neuron, 69, 407–422.
Cambridge, MA: MIT Press.
Huttenlocher, P. R. (2002). Neural plasticity: The effects of
environment on the development of the cerebral cortex.
Cambridge, MA: Harvard University Press.
Huttenlocher, P. R., & Dabholkar, A. S. (1997). Regional
differences in synaptogenesis in human cerebral cortex.
Journal of Comparative Neurology, 387, 167–178.
Hynd, G. W., Semrud-Clikeman, M., Lorys, A. R., Novey, E. S., &
Eliopulos, D. (1990). Brain morphology in developmental
dyslexia and attention deficit disorder/hyperactivity. Archives
of Neurology, 47, 919–926.
Jobard, G., Crivello, F., & Tzourio-Mazoyer, N. (2003). Evaluation
of the dual route theory of reading: A meta-analysis of 35
neuroimaging studies. Neuroimage, 20, 693–712.
Klein, A., Ghosh, S. S., Avants, B., Yeo, B. T. T., Fischl, B.,
Ardekani, B., et al. (2010). Evaluation of volume-based
and surface-based brain image registration methods.
Neuroimage, 51, 214–220.
Larsen, J. P., Høien, T., Lundberg, I., & Odegaard, H. (1990).
MRI evaluation of the size and symmetry of the planum
temporale in adolescents with developmental dyslexia. Brain
and Language, 39, 289–301.
Lee, A. D., Leow, A. D., Lu, A., Reiss, A. L., Hall, S., Chiang, M.-C.,
et al. (2007). 3D pattern of brain abnormalities in Fragile X
syndrome visualized using tensor-based morphometry.
Neuroimage, 34, 924–938.
Lenhard, W., & Schneider, W. (2006). Ein Leseverständnistest
für Erst-bis Sechstklässler. Göttingen, Germany: Hogrefe.
Liberman, A. M., Cooper, F. S., Shankweiler, D. P., & Studdert-
Kennedy, M. (1967). Perception of the speech code.
Psychological Review, 74, 431–461.
Linkersdörfer, J., Lonnemann, J., Lindberg, S., Hasselhorn, M., &
Fiebach, C. J. (2012). Grey matter alterations co-localize with
functional abnormalities in developmental dyslexia: An ALE
meta-analysis. PloS One, 7, e43122.
Lu, L. H., Dapretto, M., OʼHare, E. D., Kan, E., McCourt, S. T.,
Thompson, P. M., et al. (2009). Relationships between brain
activation and brain structure in normally developing
children. Cerebral Cortex, 19, 2595–2604.
McCandliss, B. D., Cohen, L., & Dehaene, S. (2003). The visual
word form area: Expertise for reading in the fusiform gyrus.
Trends in Cognitive Sciences, 7, 293–299.
Moll, K., & Landerl, K. (2010). SLRT-2—Lese-und
Rechtschreibtest. Bern, Switzerland: Hans Huber.
Nakada, T., Fujii, Y., Yoneoka, Y., & Kwee, I. L. (2001). Planum
temporale: Where spoken and written language meet.
European Neurology, 46, 121–125.
Nation, K., & Hulme, C. (2011). Learning to read changes
childrenʼs phonological skills: Evidence from a latent variable
longitudinal study of reading and nonword repetition.
Developmental Science, 14, 649–659.
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Hickok, G., Okada, K., & Serences, J. T. (2009). Area Spt in the
human planum temporale supports sensory–motor
Pattamadilok, C., Knierim, I. N., Kawabata Duncan, K. J., &
Devlin, J. T. (2010). How does learning to read affect
Linkersdörfer et al.
317
speech perception? Journal of Neuroscience, 30,
8435–8444.
Price, C. J. (2012). A review and synthesis of the first 20 years of
PET and fMRI studies of heard speech, spoken language
and reading. Neuroimage, 62, 816–847.
Pugh, K. R., Mencl, W. E., Jenner, A. R., Katz, L., Frost, S. J.,
Lee, J. R., et al. (2001). Neurobiological studies of reading and
reading disability. Journal of Communication Disorders,
34, 479–492.
Pulvermüller, F., & Fadiga, L. (2010). Active perception:
Sensorimotor circuits as a cortical basis for language. Nature
Reviews Neuroscience, 11, 351–360.
Pulvermüller, F., Huss, M., Kherif, F., Moscoso del Prado Martin,
F., Hauk, O., & Shtyrov, Y. (2006). Motor cortex maps
articulatory features of speech sounds. Proceedings of the
National Academy of Sciences, U.S.A., 103, 7865–7870.
R Core Team. (2012). R: A language and environment for
statistical computing. Vienna, Austria: R Foundation for
Statistical Computing.
Richlan, F., Kronbichler, M., & Wimmer, H. (2013). Structural
abnormalities in the dyslexic brain: A meta-analysis of voxel-
based morphometry studies. Human Brain Mapping, 34,
3055–3065.
Rumsey, J. M., Nace, K., Donohue, B., Wise, D., Maisog, J. M.,
& Andreason, P. (1997). A positron emission tomographic
study of impaired word recognition and phonological
processing in dyslexic men. Archives of Neurology, 54,
562–573.
Schlaggar, B. L., & McCandliss, B. D. (2007). Development of
neural systems for reading. Annual Review of Neuroscience,
30, 475–503.
Ségonne, F., Dale, A. M., Busa, E., Glessner, M., Salat, D., Hahn,
H. K., et al. (2004). A hybrid approach to the skull stripping
problem in MRI. Neuroimage, 22, 1060–1075.
Shaywitz, B. A., Shaywitz, S. E., Pugh, K. R., Mencl, W. E.,
Fulbright, R. K., Skudlarski, P., et al. (2002). Disruption of
posterior brain systems for reading in children with
developmental dyslexia. Biological Psychiatry, 52,
101–110.
Sowell, E. R., Thompson, P. M., Leonard, C. M., Welcome, S. E.,
Kan, E., & Toga, A. W. (2004). Longitudinal mapping of
cortical thickness and brain growth in normal children.
Journal of Neuroscience, 24, 8223–8231.
Squeglia, L. M., Jacobus, J., Sorg, S. F., Jernigan, T. L., & Tapert,
S. F. (2013). Early adolescent cortical thinning is related
to better neuropsychological performance. Journal of the
International Neuropsychological Society, 19, 962–970.
Steinbrink, C., Vogt, K., Kastrup, A., Müller, H.-P., Juengling,
F. D., Kassubek, J., et al. (2008). The contribution of white
and gray matter differences to developmental dyslexia:
Insights from DTI and VBM at 3.0 T. Neuropsychologia, 46,
3170–3178.
Turkeltaub, P. E., Eden, G. F., Jones, K. M., & Zeffiro, T. A.
(2002). Meta-analysis of the functional neuroanatomy of
single-word reading: Method and validation. Neuroimage,
16, 765–780.
Tustison, N. J., Avants, B. B., Cook, P. A., Zheng, Y., Egan, A.,
Yushkevich, P. A., et al. (2010). N4ITK: Improved N3 bias
correction. IEEE Transactions on Medical Imaging, 29,
1310–1320.
Wendelken, C., OʼHare, E. D., Whitaker, K. J., Ferrer, E., &
Bunge, S. A. (2011). Increased functional selectivity over
development in rostrolateral prefrontal cortex. Journal of
Neuroscience, 31, 17260–17268.
Wilke, M., Holland, S. K., Altaye, M., & Gaser, C. (2008).
Template-O-Matic: A toolbox for creating customized
pediatric templates. Neuroimage, 41, 903–913.
Wilson, S. M., Saygin, A. P., Sereno, M. I., & Iacoboni, M. (2004).
Listening to speech activates motor areas involved in speech
production. Nature Neuroscience, 7, 701–702.
Wise, R. J., Scott, S. K., Blank, S. C., Mummery, C. J., Murphy, K.,
& Warburton, E. A. (2001). Separate neural subsystems within
“Wernickeʼs area.” Brain, 124, 83–95.
Yeatman, J. D., Dougherty, R. F., Ben-Shachar, M., & Wandell,
B. A. (2012). Development of white matter and reading skills.
Proceedings of the National Academy of Sciences, U.S.A.,
109, E3045–3053.
Yushkevich, P. A., Avants, B. B., Das, S. R., Pluta, J., Altinay, M.,
Craige, C., et al. (2010). Bias in estimation of hippocampal
atrophy using deformation-based morphometry arises from
asymmetric global normalization: An illustration in ADNI 3 T
MRI data. Neuroimage, 50, 434–445.
Zhang, M., Li, J., Chen, C., Mei, L., Xue, G., Lu, Z., et al. (2013).
The contribution of the left mid-fusiform cortical thickness to
Chinese and English reading in a large Chinese sample.
Neuroimage, 65, 250–256.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
3
/
0
2
8
/
2
3
0
0
0
8
6
/
1
1
8
8
8
2
o
3
c
4
n
5
_
4
a
/
_
j
0
o
0
c
7
n
1
0
_
a
p
_
d
0
0
b
7
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
318
Journal of Cognitive Neuroscience
Volume 27, Number 2