Temporal Expectations Prepare Visual
Working Memory for Behavior
Wen Jin1,2, Anna C. Nobre1,2, and Freek van Ede1,3
Astratto
■ Working memory enables us to retain past sensations in
service of anticipated task demands. How we prepare for antici-
pated task demands during working memory retention remains
poorly understood. Here, we focused on the role of time—
asking how temporal expectations help prepare for ensuing
memory-guided behavior. We manipulated the expected probe
time in a delayed change-detection task and report that temporal
expectation can have a profound influence on memory-guided
behavioral performance. EEG measurements corroborated the
utilization of temporal expectations: demonstrating the involve-
ment of a classic EEG signature of temporal expectation—the
contingent negative variation—in the context of working memory.
We also report the influence of temporal expectations on 2 EEG
signatures associated with visual working memory—the lateral-
ization of 8- to 12-Hz alpha activity, and the contralateral delay
activity. We observed a dissociation between these signatures,
whereby alpha lateralization (but not the contralateral delay
activity) adapted to the time of expected memory utilization.
These data show how temporal expectations prepare visual
working memory for behavior and shed new light on the electro-
physiological markers of both temporal expectation and working
memory. ■
INTRODUCTION
Working memory (Baddeley, 1992) enables us to retain past
sensations with the purpose of guiding adaptive future
behavior. In considering this purpose-oriented nature of
working memory (de Vries, Slagter, & Olivers, 2020;
van Ede, 2020; Nobre & Stokes, 2019; van Ede, Chekroud,
Stokes, & Nobre, 2019; Myers, Stokes, & Nobre, 2017;
Chatham & Badre, 2015; D’Esposito & Postle, 2015; Carlisle,
Arita, Pardo, & Woodman, 2011; Rainer, Rao, & Mugnaio, 1999;
Chelazzi, Mugnaio, Duncan, & Desimone, 1993), it is relevant
to consider not only what we encode and retain in working
memory (Lee, Kravitz, & Baker, 2013; Serences, Ester, Vogel,
& Awh, 2009; McNab & Klingberg, 2008; Vogel, McCollough,
& Machizawa, 2005; Griffin & Nobre, 2003) but also when
we expect to utilize the contents in working memory in
service of ensuing behavior.
Despite a growing realization of the important role of tem-
poral expectations in guiding behavior (Nobre & van Ede,
2018), most studies to date have investigated temporal ex-
pectations in service of relatively pure sensory (Rohenkohl,
Cravo, Wyart, & Nobre, 2012; Vangkilde, Coull, & Bundesen,
2012; Jaramillo & Zador, 2011; Lakatos, Karmos, Mehta,
Ulbert, & Schroeder, 2008; Lange & Röder, 2006; Coull &
Nobre, 1998; Jones & Boltz, 1989) or motor (Heideman,
Quinn, Woolrich, van Ede, & Nobre, 2020; Heideman,
1Oxford Centre for Human Brain Activity, Wellcome Centre for
Integrative Neuroimaging, 2University of Oxford, 3Institute for
Brain and Behavior Amsterdam, Vrije Universiteit Amsterdam
van Ede, & Nobre, 2018; Los, Kruijne, & Meeter, 2017; van
Elswijk, Kleine, Overeem, & Stegeman, 2007; Praamstra,
2006; Janssen & Shadlen, 2005; Schoffelen, Oostenveld, &
Fries, 2005) compiti. In everyday situations, Tuttavia, we often
rely on detailed contents of working memory to inform and
guide behavior (van Ede et al., 2019; Myers et al., 2017;
Chatham & Badre, 2015; Olivers, Peters, Houtkamp, &
Roelfsema, 2011; Chelazzi et al., 1993). Yet, less is under-
stood about the ways in which temporal expectations are
utilized while we anticipate using detailed information in
working memory to guide our behavior.
Complementing other recent studies on the role of tem-
poral expectations in working memory (Boettcher, Gresch,
Nobre, & van Ede, 2020; Zokaei, Board, Manohar, & Nobre,
2019; Wilsch, Henry, Herrmann, Herrmann, & Obleser,
2018; Olmos-Solis, van Loon, Los, & Olivers, 2017; van
Ede, Niklaus, & Nobre, 2017; Wilsch, Henry, Herrmann,
Maess, & Obleser, 2015), we sought to investigate the
behavioral consequences and neural signatures of temporal
expectation in the context of a well-studied change-
detection task of visual working memory, with lateralized
encoding displays (Vogel et al., 2005; Vogel & Machizawa,
2004). We investigated the consequences of temporal
expectations on memory-guided behavior in this task, by
manipulating the time at which the contents of working
memory would likely be probed.
EEG measurements enabled us to address additional rele-
vant questions regarding key electrophysiological signatures
of temporal expectations and of visual working memory
retention after lateralized displays. These signatures involved
© 2020 Istituto di Tecnologia del Massachussetts. Published under a
Creative Commons Attribution 4.0 Internazionale (CC BY 4.0) licenza.
Journal of Cognitive Neuroscience 32:12, pag. 2320–2332
https://doi.org/10.1162/jocn_a_01626
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
C
2
l
1
e
2
–
2
P
3
D
2
F
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
C
0
N
_
/
UN
1
_
8
0
6
1
2
6
2
2
6
6
0
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
6
N
2
0
6
7
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
T
.
/
o
N
0
5
M
UN
sì
2
0
2
1
the contingent negative variation or CNV (Boettcher, Stokes,
Nobre, & van Ede, 2020; Cravo, Rohenkohl, Santos, & Nobre,
2017; Praamstra, Kourtis, Kwok, & Oostenveld, 2006;
Miniussi, Wilding, Coull, & Nobre, 1999; Weinberg, 1972;
Walter, Cooper, Aldridge, McCallum, & Inverno, 1964), IL
lateralization of 8- to 12-Hz alpha activity (Hakim, Adam,
Gunseli, Awh, & Vogel, 2019; van Ede, 2018; Myers, Walther,
Wallis, Stokes, & Nobre, 2015; Wallis, Stokes, Cousijn,
Woolrich, & Nobre, 2015; Lozano-Soldevilla, ter Huurne,
Cools, & Jensen, 2014; van Dijk, van der Werf, Mazaheri,
Medendorp, & Jensen, 2010; Sauseng et al., 2009), E
the contralateral delay activity or CDA (Hakim et al.,
2019; Schmidt & Zelinsky, 2017; Luria, Balaban, Awh, &
Vogel, 2016; Kuo, Stokes, & Nobre, 2012; Carlisle et al.,
2011; van Dijk et al., 2010; Vogel et al., 2005; Vogel &
Machizawa, 2004). Specifically, we addressed whether the
CNV also supports temporal expectation during working
memory and whether alpha lateralization and the CDA—
two spatially selective markers of attention and/or reten-
tion in visual working memory—are each sensitive to
temporal expectation and adapt to the time of expected
memory utilization.
We hypothesized that if temporal expectations are uti-
lized to prepare for upcoming visual-working-memory-
guided behavior, then performance to early probes should
be better when probes were expected to appear early as
opposed to late. We further hypothesized that the utiliza-
tion of temporal expectations should be traceable in the
CNV, yielding a more negative frontal potential ahead of
the early probe when the probe is expected early as opposed
to late. Finalmente, we reasoned that if working memory reten-
tion is modulated by temporal expectation, this would be
reflected in its related neural markers. Accordingly, we tested
for modulations of alpha lateralization and the CDA to test
whether they tracked modulation in working memory reten-
zione. In other words, we investigated whether these
signatures depend not only on what information is retained
in working memory (left vs. right items) but also on when
this information is expected to become required for guiding
behavior.
METHODS
Participants
Twenty-five healthy human volunteers (7 men, mean age =
26, range: of 18–35) participated in the study. Sample size
was set based on prior studies from the laboratory with
similar outcome variables (van Ede et al., 2017, 2019). All par-
ticipants had normal or corrected-to-normal vision. Data
from one participant had to be excluded from the analysis
because of below-chance task performance (49.8% correct
responses, compared with M = 76.02, SE = 1.96% in the re-
maining sample). All participants provided written informed
consent before participation and were paid £15 per hour.
The study was approved by the central university research
ethics committee of the University of Oxford.
Task and Procedure
Participants performed a delayed visual change-detection
working memory task with lateralized encoding displays,
in which we manipulated temporal expectations regarding
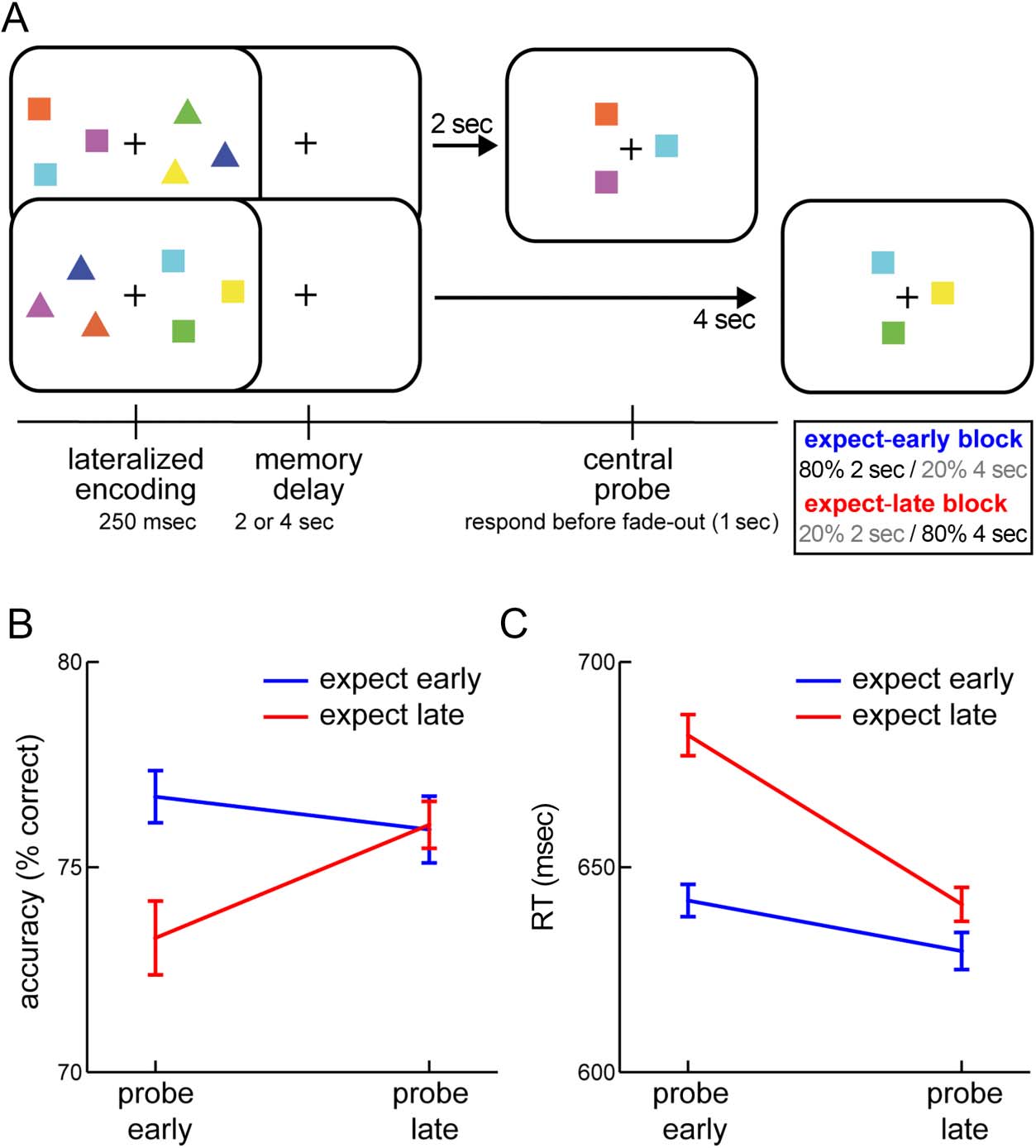
expected probe time across blocks (Figure 1A).
Participants sat approximately 90 cm in front of a computer
monitor (100-Hz refresh rate). At memory encoding, one
side of the display always contained three colored squares,
whereas the other side contained three colored triangles.
Each shape had a unique color from a pool of six possible
colors (red, green, blue values: [255, 107, 64], [255, 255,
64], [64, 255, 107], [64, 64, 255], [64, 255, 255], E [255,
64, 255]). Individual shapes were approximately 1° visual
angle in width and height and were positioned at a random
point along the outer edge of an invisible circle (6.4° diam-
eter; anchored at 6.4° to the left and right of fixation).
Before each of two experimental sessions, participants were
explicitly instructed to remember one set of shapes (Dire
piazze) while ignoring the other set (triangles). Which
set of shapes was relevant was counterbalanced across ses-
sions. Irrelevant shapes served to balance the visual encod-
ing display (and to require item selection at encoding), come
that any lateralized patterns of neural activity could be
attributed to the selection and retention of the relevant
memory items (rather than bottom–up visual responses
to the display). The positioning of the relevant and irrele-
vant shapes to the left and right locations on the screen
was randomly varied across trials.
Encoding displays containing the 2 × 3 shapes were pre-
sented for 250 msec, followed by a memory delay period of
either 2 O 4 sec in which only the fixation cross (2° in width
and height) remained on the screen. After the memory delay,
a probe display was presented that always contained the
three relevant shapes, but of which two of the shapes (from
the relevant shape set) had either swapped colors (change)
or not (no change). Change and no-change trials were equally
likely and randomly intermixed. Probe displays were lpre-
sented centrally, but otherwise preserved the spatial configu-
ration of the relevant shape set. Participants responded using
the keyboard in front of them and pressed “M” (for “match”)
if they thought the color configuration had remained the
same, or “C” (for “change”) if they thought it had changed.
To encourage holding the memory items ready for
guiding behavior, we instructed participants to make their
change/no-change report within 1 sec after probe onset. A
this end, we gradually faded the visibility of the probe by
adjusting its transparency levels from 0–100% over a
1-sec period, starting at probe onset. Participants were
explicitly told that responses that would be received after
the probe had disappeared would be counted as incorrect.
The program provided feedback on performance as
soon as a response was collected or the maximum response
time had been reached. The fixation cross changed (for
300 msec) to green if a correct response was made, or to
red otherwise. The next trial began after an intertrial inter-
val between 750 E 1000 msec.
Jin, Nobre, and van Ede
2321
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
C
2
l
1
e
2
–
2
P
3
D
2
F
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
C
0
N
_
/
UN
1
_
8
0
6
1
2
6
2
2
6
6
0
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
6
N
2
0
6
7
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
.
/
T
o
N
0
5
M
UN
sì
2
0
2
1
The critical manipulation in the current study involved
the expected probe time. In “expect-early” blocks, 80% Di
trials (32/40 per block) had a memory delay of 2 sec,
whereas 20% of trials (8/40 per block) had a delay of
4 sec. In contrasto, in “expect-late” blocks, 80% of trials
had a memory delay of 4 sec, whereas only 20% of trials
had a delay of 2 sec. Expect-early and expect-late blocks
were randomly interleaved. Participants were explicitly
informed what block type they were in before the block
started, as signaled by a preblock instruction screen.
Participants completed two consecutive sessions of the
task of approximately 45 min each, con un 10- to 15-min
break in between them. The two sessions each contained
10 blocks (5 per block type) Di 40 trials, yielding 800 trials
per participant.
Visual Localizer
In between each block of the main working memory task,
we included visual localizer modules that served to find the
posterior electrodes that best captured visual processing of
the stimuli that we used in the main task. In each localizer
display, we presented a single set of three colored shapes
(three squares or three triangles), either to the left or to the
right of fixation. Participants were instructed to maintain
fixation on the central cross, without any further task.
Individual localizer displays were presented for 250 msec,
followed by an interstimulus interval of 400–600 msec.
Each localizer module contained 80 stimulus displays,
yielding 1,600 localizer displays across the two sessions
(800 per side).
The rationale for including localizers was motivated by the
use of similar localizers in several of our prior studies (per esempio.,
van Ede et al., 2017; van Ede, Jensen, & Maris, 2010).
Localizers provided an independent means to find left and
right electrodes that were maximally sensitive to visual pro-
cessing of the contralateral (vs. ipsilateral) visual field—and
to do so on a participant-by-participant basis. Having found
these participant-specific electrodes that were maximally
sensitive to visual processing of the contralateral visual field,
we were then able to use these electrodes to focus our anal-
ysis on lateralized signatures of visual processing associated
with working memory retention—alpha lateralization and
the CDA. By focusing on these independently selected elec-
trodes, we were thus able to increase the sensitivity of our
analysis. Because the localizer-based electrode selection
served only as a “proxy” for the set of relevant posterior elec-
trodes (there was no strict guarantee that these lateralized
working memory signatures would occur in precisely the
same electrodes), we additionally present the topographies
of these lateralized EEG modulations.
Analysis of Behavioral Data
We considered two measures of performance: accuracy
and RT. For accuracy, we calculated the percentage of
trials with a correct response. Incorrect response trials in-
cluded trials in which the wrong button was pressed or in
which no button press was recorded in the 1-sec re-
sponse window during which the probe gradually faded.
Participants were explicitly told that responses occurring
after the probe had disappeared would be counted as in-
correct (and were given negative feedback immediately
after the end of the response window was reached).
For RTs, we only considered trials in which a response
was collected (95.84 ± 1.22% of trials).
To quantify the effects of temporal expectations on
working-memory-guided performance, we used a 2 × 2
repeated-measures ANOVA with the factors Block Type
(expect early/late) and Memory Delay (probe early/late)
and used follow-up paired-samples t tests to test for tem-
poral expectations effects at each memory delay sepa-
rately. For this, we directly compared expect-early and
expect-late conditions, separately for performance after
early and late probes. For these post hoc t tests, we addi-
tionally report Bonferroni-corrected p values that we
denote as “pBonferroni.”
EEG Acquisition
EEG was acquired using Synamps amplifiers and
Neuroscan acquisition software. We used 61 electrodes
that we placed according to the international 10–10 posi-
tioning system (Chatrian, Lettich, & Nelson, 1985).
During acquisition, the left mastoid served as the refer-
ence. A right mastoid measurement was included for
off-line referencing to the average of both mastoids. Noi
positioned the ground electrode on the left upper arm.
Vertical and horizontal EOG were measured using elec-
trodes placed to the lateral side of each eye (horizontal
EOG), as well as above and below the left eye (vertical
EOG). Data were acquired with a hardware filter between
0.1 E 200 Hz, digitized at 1000 Hz, and stored for off-
line analysis.
EEG Analysis
Data were analyzed in using the FieldTrip toolbox
(Oostenveld, Fries, Maris, & Schoffelen, 2011) in MATLAB
(The MathWorks). We used independent component anal-
ysis to correct for ocular contributions to the EEG. Dopo
independent component analysis correction, we dis-
carded trials with exceptionally high variance on the basis
of visual inspection (using the function ft_rejectvisual
with the summary method). On average, 686 ± 15 (fuori
Di 800) trials were retained for analysis per participant.
We focused on two complementary sets of analyses—
focusing on a global signature of temporal expectation
(the CNV) and on the spatially specific memory-retention
signatures that lateralized according to the side of the
memory items (alpha lateralization and the CDA).
2322
Journal of Cognitive Neuroscience
Volume 32, Numero 12
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
C
2
l
1
e
2
–
2
P
3
D
2
F
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
C
0
N
_
/
UN
1
_
8
0
6
1
2
6
2
2
6
6
0
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
6
N
2
0
6
7
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
.
/
T
o
N
0
5
M
UN
sì
2
0
2
1
Analysis of CNV
Data were baseline corrected by subtracting the average
potential in the 250-msec window preceding encoding
onset, and averaged across trials. We used a predefined
cluster of frontal EEG electrodes, centered on electrode
Fz together with its immediate neighbors: (FCz, AFz, Fz,
F1, F2), as in the study of Boettcher, Stokes, et al. (2020).
Data were averaged across the selected electrodes and com-
pared between trials in which the probe was expected to
occur early (expect-early blocks) or late (expect-late
blocks). To increase visualization of this slow potential,
we smoothed the CNV time courses with a Gaussian kernel
with an SD of 25 msec.
Analysis of Lateralized Alpha and CDA
To increase sensitivity for the analysis of neural lateraliza-
zione, we applied a surface-Laplacian transform (Perrin,
Pernier, Bertrand, & Echallier, 1989) that increases separa-
bility of nearby sources of activity (here, activity in left and
right visual electrodes; note that we deliberately did not
apply the same transform to the more global CNV analysis,
as the CNV is relatively widespread).
We used clusters of left and right posterior electrodes
that were selected on a participant-by-participant basis,
based on the data from our independent visual localizer
modules (see Visual Localizer section)—as in our prior study
(van Ede et al., 2017). In short, we compared responses to
localizer displays with left versus right items, and compared
8- to 12-Hz alpha activity in the 150- to 400-msec poststim-
ulus window. Our choice to focus on the 150- to 400-msec
window was based on our prior experience with similar
localizers (per esempio., van Ede et al., 2010, 2017), which have indi-
cated this as the approximate window in which the induced
alpha response to a sensory stimulus is most pronounced.
By starting 150 msec after stimulus onset, we minimize the
contribution of the initial evoked response, which leads to
an increase in power across a wide range of frequencies,
thereby obscuring any stimulus-induced alpha decreases
in the first 150 msec after stimulus onset. We then selected
the posterior electrodes that showed the clearest difference
between left and right localizer stimuli based on visual
inspection. We selected electrodes from among the fol-
lowing potential posterior electrodes: left: (P7, P5, P3,
P1, PO7, PO3, O1); right: (P8, P6, P4, P2, PO8, PO4, O2).
We selected no less than four and no more than six elec-
trodes per side. The number of participants (out of 24)
for which a given electrode was included were P7 (21),
P5 (24), P3 (21), P1 (2), PO7 (24), PO3 (23), O1 (11),
P8 (18), P6 (23), P4 (22), P2 (4), PO8 (24), PO4 (24),
and O2 (15).
For the analysis of the CDA, like for the CNV, we applied
a baseline correction using a 250-msec pre-encoding
baseline and smoothed the time courses with a Gaussian
kernel with an SD of 25 msec. We considered the data in
the selected left and right posterior electrode clusters, as a
function of whether the to-be-memorized shapes had
been presented to the left or right of fixation at encoding.
Per electrode cluster, the CDA was calculated by sub-
tracting conditions where the memorized items had been
contralateral versus ipsilateral to the electrode cluster
under investigation. CDA waveforms were subsequently
averaged between the left and right electrode clusters
to yield a single CDA waveform per temporal expectation
condition.
For the analysis of alpha lateralization, we first applied
a time–frequency analysis using a short-time Fourier
transform of Hanning-tapered data, as implemented in
FieldTrip (Oostenveld et al., 2011). We estimated spectral
power between 5 E 25 Hz (in steps of 1 Hz) and used a
300-msec sliding time window that we advanced over the
data in steps of 30 msec. As for the CDA, time–frequency
maps of power were contrasted between trials in which
the memory items were contralateral versus ipsilateral to
each electrode cluster. For normalization purposes, we
expressed this difference in spectral power as a percent-
age change: ((contra − ipsi) / (contra + ipsi)) × 100. A
visualize the time courses of alpha lateralization in
expect-early and expect-late conditions, we extracted
the activity in the predefined 8- to 12-Hz alpha band.
Topographical maps of alpha lateralization and CDA
were obtained by applying the same procedures to all
symmetrical electrode pairs and plotting the results in
the right electrode of each pair.
Avoiding Neural Contamination by Early Probes
Our EEG analyses focused on the early memory delay
period where temporal expectation effects are known to
be most pronounced (per esempio., Heideman, Rohenkohl, et al.,
2018; Nobre & van Ede, 2018; Nobre, 2001). This allowed
us to compare neural activity between all trials in expect-
early and expect-late blocks (regardless of whether the
probe actually appeared after 2 O 4 sec). To avoid contam-
ination by any probe-related activity into the presented
analysis—which would be problematic given the larger
number of early probes in the expect-early condition—
we only considered data for which we could be sure that
there was no contamination by the probe (as in van Ede
et al., 2017). For the CNV and CDA analyses, we substituted
the data from all trials with a 2-sec memory delay with “Not
a Number” from the time of probe onset (2250 msec after
encoding onset). For the time–frequency analysis, we did
the same, but started filling the contaminated data with Not
a Number 150 msec earlier, to deal with the fact that we had
used a 300-msec window sliding time window (yielding
150-msec smearing to each side).
Statistical Approach to EEG Data
We compared expect-early and expect-late conditions using
cluster-based permutation analyses (Maris & Oostenveld,
Jin, Nobre, and van Ede
2323
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
C
2
l
1
e
2
–
2
P
3
D
2
F
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
C
0
N
_
/
UN
1
_
8
0
6
1
2
6
2
2
6
6
0
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
6
N
2
0
6
7
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
.
T
o
N
0
5
M
UN
sì
2
0
2
1
2007), as implemented in the FieldTrip toolbox (Oostenveld
et al., 2011). We used the default clustering settings, con
10,000 permutations. We applied these analyses on the data
extracted from the predefined electrode clusters. All topo-
graphical analyses served only to confirm the physiological
plausibility (van Ede & Maris, 2016) of the identified
patterns and were not subjected to further inferential sta-
tistical tests.
RESULTS
Participants performed a visual working memory task in
which we manipulated the time at which information in
working memory would become relevant for guiding
behavior (Figure 1A). Participants were instructed to remem-
ber the squares or the triangles and to judge whether the
configuration of these relevant shapes had changed or not
between the encoding display and the probe display. In
expect-early blocks, 80% of trials had a memory delay of 2
sec, whereas 20% of trials had a delay of 4 sec. In contrasto,
in expect-late blocks, 80% of trials had a memory delay of
4 sec, whereas only 20% of trials had a delay of 2 sec.
Temporal Expectations Are Utilized during Visual
Working Memory: Behavior and CNV
To assess whether participants utilized the temporal pre-
dictability of the probe array to facilitate their working-
memory-guided behavior, we investigated behavioral
accuracy and RTs when the probe array occurred early
or late (memory delay) as a function of whether it was ex-
pected to occur early or late (block type).
For accuracy (Figure 1B) we observed a significant inter-
action between the time of probing (Memory Delay) E
the expected probe time (Block Type), F(1, 23) = 4.329,
p = .049, ηp
2 = .158), without a main effect of Memory
Delay ( p = .202) or Block Type ( p = .095). When visual
working memory was probed early, participants were sub-
stantially better when they also expected the probe to oc-
cur early as opposed to when they expected the probe to
occur late (M = 76.7 vs. 73.3% correct; T(23) = 2.522, p =
.019, pBonferroni = 0.038, d = 0.515). In contrasto, Quando
memory was probed late, there was no longer an effect
of temporal expectation (M = 75.9 vs. 76.0% correct; p =
.926). This lack of effect after the long memory delay is in
line with ample prior studies on temporal expectation in the
domains of perception and action (Nobre & van Ede, 2018
Figura 1. Temporal expectations
shape working-memory-guided
behavior. (UN) Visual working
memory task with a standard
temporal expectation
manipulation. Participants
were instructed to selectively
remember the squares or the
triangles. After a memory delay
Di 2 O 4 sec, a central probe
was presented and participants
judged whether the probe
configuration remained the same
(4-sec example) or changed
(2-sec example) from the
encoded configuration of the
relevant shapes. Across blocks,
probes were either most likely
(80%) to occur early (2 sec) O
late (4 sec), but would occur at
the unexpected time in 20% Di
the trials. (B) Percentage correct
responses as a function of
expected probe time and actual
memory delay. (C) RT as a
function of expected probe time
and actual memory delay. Error
bars indicate ± 1 SEM calculated
across participants (n = 24), after
within-subject correction
(Cousineau, 2005).
2324
Journal of Cognitive Neuroscience
Volume 32, Numero 12
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
C
2
l
1
e
2
–
2
P
3
D
2
F
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
C
0
N
_
/
UN
1
_
8
0
6
1
2
6
2
2
6
6
0
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
6
N
2
0
6
7
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
F
T
/
o
N
0
5
M
UN
sì
2
0
2
1
for review) and is likely attributed to the fact that, once the
short delay has passed, participants can update their ex-
pectations according to the equated temporal conditional
probability at the later interval.
Importantly, this pattern of data shows that the worse
performance when the probe unexpectedly comes early
cannot be because of a weaker memory trace per se
because performance in these expect-late blocks recovers
when probed at the late interval (despite the fact that now
more time has passed since encoding). Invece, this is likely
attributed to a suboptimal readiness (or “coding format”)
of the memory array, when prompted to perform the
working memory task earlier than expected (see also
van Ede et al., 2017).
For RT (Figure 1C), we observed a qualitatively similar
pattern. We found a significant interaction between Memory
Delay and Block Type, F(1, 23) = 15.9324, p = 5.745e−4,
ηp
2 = .409) that was again constituted by a particularly robust
benefit of matching expectation for early probes (M = 630.4
vs. 666.0 ms; T(23) = −6.271, p = 2.126e−6, pBonferroni =
4.253e−6, d = −1.280), although a trend for faster responses
in expect-early blocks persisted after late probes (M = 620.1
vs. 628.7 ms; T(23) = −2.087, p = .048, pBonferroni = 0.096, d =
−0.426). RTs additionally showed a main effect of Memory
Delay, F(1, 23) = 15.461, p = 6.659e−4, ηp
2 = .683) E
Block Type, F(1, 23) = 37.064, p = 3.287e−6, ηp
2 = .650),
with reactions being generally faster after late probes and in
expect-early blocks.
EEG measurements provided complementary evidence
that participants prepared for the time of expected memory
utilization while engaging in visual working memory reten-
zione. Focusing on the initial memory interval—in which
expect-early and expect-late blocks are known to differ most
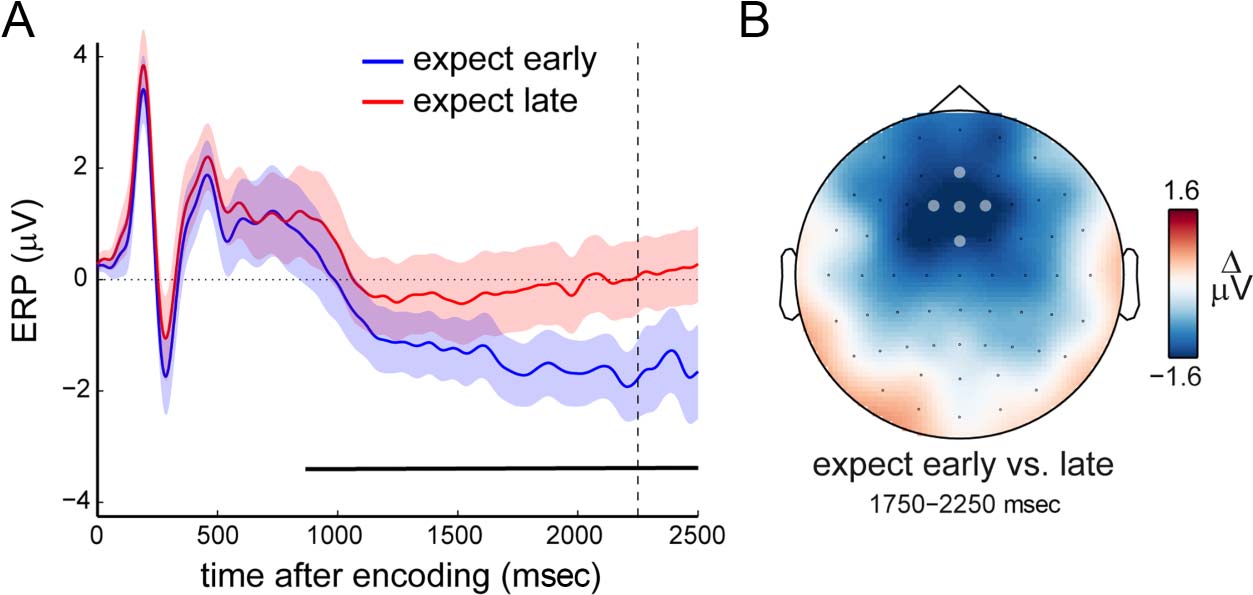
(Nobre & van Ede, 2018; Nobre, 2001)—we observed the
gradual emergence of a frontal scalp potential that was
more negative when memory was expected to be probed
early versus late (Figure 2A; black line; cluster p < .0001).
This potential likely reflects the CNV (Boettcher, Stokes,
et al., 2020; Cravo et al., 2017; Praamstra et al., 2006;
Miniussi et al., 1999; Weinberg, 1972; Walter et al., 1964),
as also evidenced by its clear frontal topography (Figure 2B).
Together, these behavioral and EEG data each make
clear that participants take the expected time of prospec-
tive memory use into account while engaging in visual
working memory. They show that temporal expectations
are utilized to benefit performance at the time of expected
memory use and that such expectations are associated with
a CNV during visual working memory retention.
Notably, in other work from our laboratory, we recently
showed how the CNV can be sensitive to the contents of
working memory, even when carefully controlling response
demands between conditions (Boettcher, Stokes, et al.,
2020). It thus remains a theoretical possibility that the
CNV modulation in the current study too may be related
to differences in working memory processes between
temporal expectation conditions, in addition to changes
in response readiness.
Owing to the lateralized nature of our displays, we were
also in the position to investigate patterns of lateralized
neural activity according to the encoded location of the
relevant memory items. We focused on two human EEG sig-
natures commonly associated with visual working memory
after such displays: the lateralization of posterior 8- to 12-Hz
alpha activity (Hakim et al., 2019; van Ede, 2018; Myers,
Walther, et al., 2015; Lozano-Soldevilla et al., 2014; van Dijk
et al., 2010; Sauseng et al., 2009) and the CDA (Hakim et al.,
2019; Schmidt & Zelinsky, 2017; Luria et al., 2016; Carlisle
et al., 2011; van Dijk et al., 2010; Vogel et al., 2005; Vogel &
Machizawa, 2004).
Timed Reorganization of Alpha Lateralization in
Anticipation of the Expected Probe
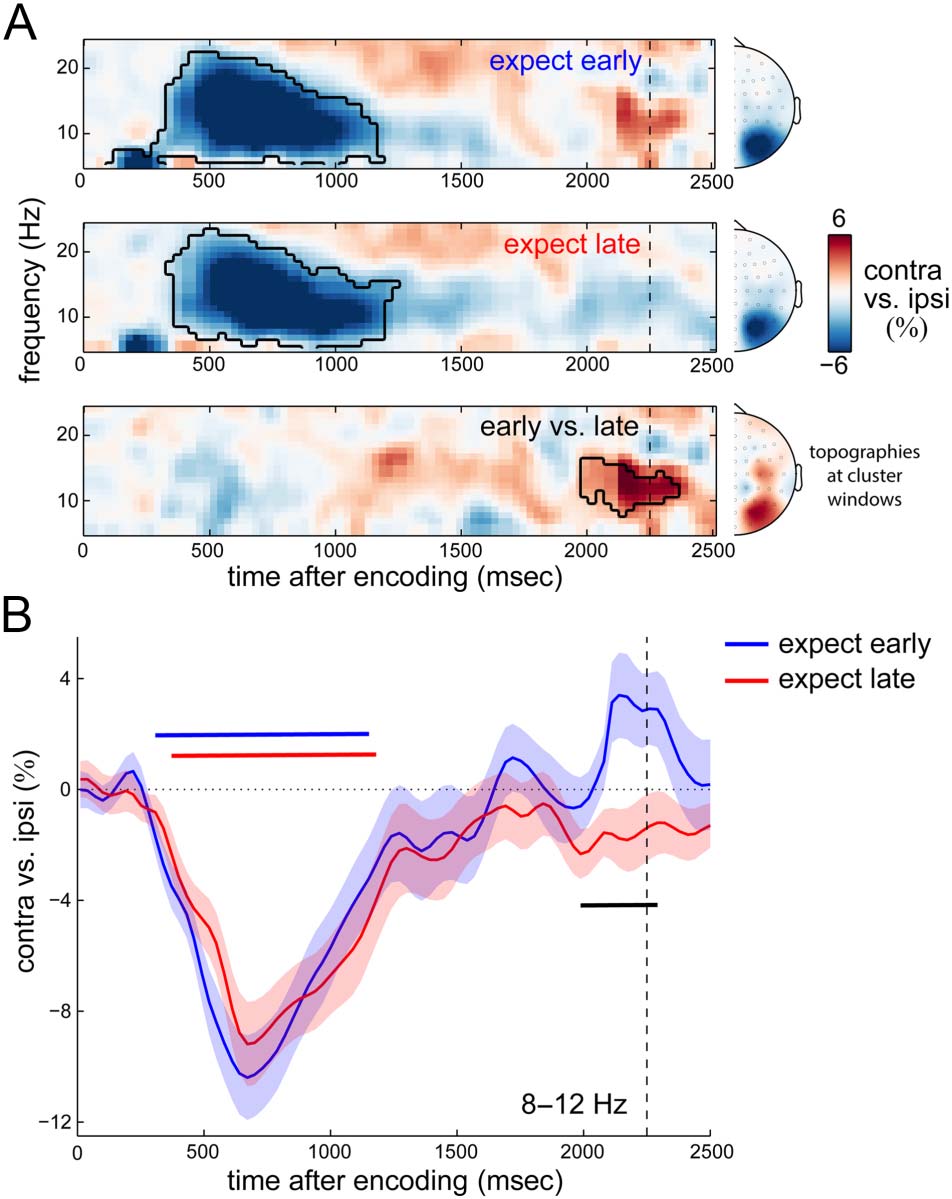
We first focused on alpha lateralization. Figure 3A shows the
time- and frequency-resolved lateralization of neural activity
Figure 2. A CNV potential
associated with temporal
expectations during working
memory. (A) Event-related
potentials in predefined frontal
electrodes (marked in B;
[FCz, AFz, Fz, F1, F2];
as in Boettcher, Stokes,
et al., 2020), as a function
of expected probe time.
Dashed vertical line indicates
the time at which an early
probe could occur. Black
horizontal line indicates
significant difference between
conditions after a cluster-based
permutation analysis (Maris &
Oostenveld, 2007). Shaded areas indicate ± 1 SEM, calculated across participants (n = 24). (B) Topography associated with the potential
difference shown in A.
Jin, Nobre, and van Ede
2325
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
t
/
o
n
0
5
M
a
y
2
0
2
1
expect-late blocks (Figure 3A, bottom) revealed a signifi-
cant difference in spectral lateralization (cluster p = .017)
centered on alpha band activity at the anticipated time of
the probe in expect-early blocks. This difference in laterali-
zation also has a posterior topography. Figure 3B shows the
associated time courses of 8- to 12-Hz alpha lateralization
in expect-early and expect-late blocks and confirms the
temporally tuned spatial reorganization of alpha activity
by temporal expectation (cluster pearly = .0002, cluster
plate = .0002, cluster pearly vs. late = .0095).
We note that this difference in alpha lateralization
between expect-early and expect-late blocks cannot be
explained by differences in visual input (probe onset).
Time windows for analysis were adjusted to remove any data
points that could be contaminated by probe onset (see
Methods section for details). Because the clusters of the
modulation of alpha lateralization started at 1990 msec,
this effect was also unlikely to result from our correction
procedure that was applied from 2100 msec onward.
Moreover, although our correction procedure resulted in
an imbalance of usable expect-early and expect-late trials
(from 2100 msec onward), the mean lateralization values
that we compared between conditions should not be sys-
tematically biased toward lower or higher values by the
number of available trials.
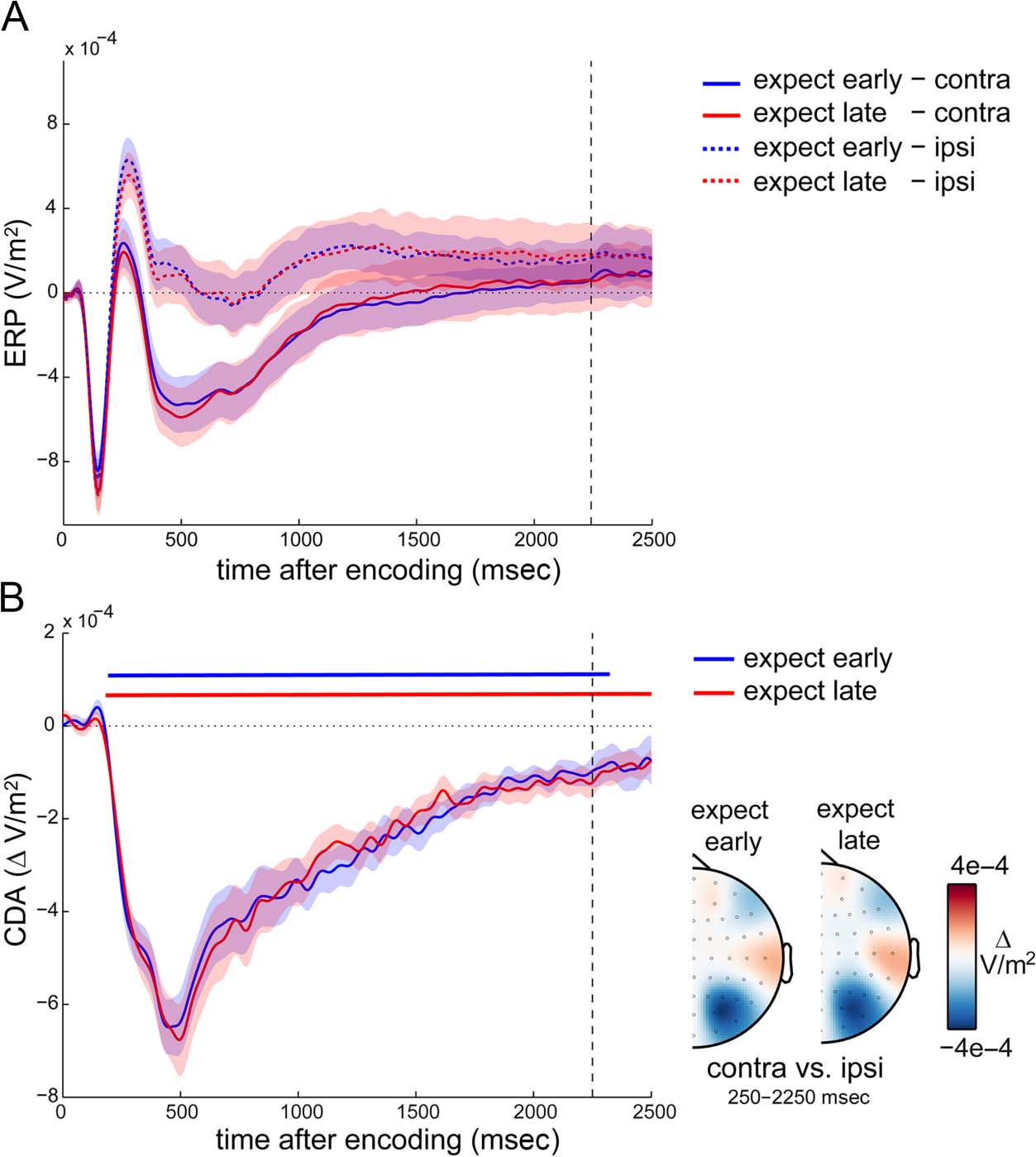
No Effect of Temporal Expectation on the CDA
In a similar spirit to the analysis of alpha lateralization, we
also investigated the effects of temporal expectation on a
second lateralized measure implicated in visual working
memory retention after lateralized encoding displays:
the CDA (e.g., Hakim et al., 2019; Schmidt & Zelinsky,
2017; Luria et al., 2016; Carlisle et al., 2011; van Dijk et al.,
2010; Vogel et al., 2005; Vogel & Machizawa, 2004).
Figure 4A shows the ERPs in contra- and ipsilateral posterior
EEG electrodes separately for expect-early and expect-late
blocks, whereas Figure 4B depicts the associated contra-
minus-ipsi difference waves that constitute the CDA.
Although, for simplicity, we refer to any lateralized posterior
ERP activity as the CDA, we note that the early parts of this
component are likely to reflect, or receive contributions
from, the N2pc (Luck & Hillyard, 1994) and/or related
components (e.g., Töllner, Müller, & Zehetleitner, 2012;
Robitaille & Jolicoeur, 2006).
As with the initial alpha lateralization, in both block types,
we observed a clear difference between contra- and ipsilat-
eral traces (cluster pearly < .0001, cluster plate < .0001) that
were each also characterized by a posterior topography
(Figure 4B). However, unlike the alpha lateralization, no
reversal in the CDA was observed around the time of
expected memory use in the expect-early blocks, and no
difference was observed when directly comparing the
CDA in expect-early and expect-late blocks (no clusters
found; all univariate p before multiple-comparison correc-
tion ≥ .0774).
Figure 3. Temporal expectation is associated with a temporally specific
spatial reorganization of alpha activity the time of expected memory
utilization. (A) Time–frequency maps of neural lateralization (contra- vs.
ipsilateral to the location of the relevant memory shapes at encoding)
in selected posterior electrodes (see Methods section), separately
for expect-early (top) and expect-late (middle) conditions, as well as
their difference (bottom). Black outlines indicate significant clusters.
Topographies show lateralization collapsed over the time–frequency
points of the respective clusters. (B) Time courses of lateralization
in the predefined 8- to 12-Hz alpha band. Horizontal lines indicate
significant clusters (black for the difference between temporal
expectation conditions). Dashed vertical lines indicate the time at
which an early probe could occur.
in selected posterior electrodes (see Methods section for
details on our independent electrode selection), relative
to the location of the relevant memory array. In line with
prior studies (e.g., Hakim et al., 2019; Lozano-Soldevilla
et al., 2014; van Dijk et al., 2010; Sauseng et al., 2009), we
observed robust lateralization of alpha power over the
posterior scalp reflecting a relative attenuation in contra-
lateral (vs. ipsilateral) electrodes. This lateralization was
particularly pronounced in the first second of the memory
delay and was highly similar in expect-early and expect-late
blocks (Figure 3A, top and middle; cluster pearly < .0001;
cluster plate < .0001). Later in the memory delay, however,
the pattern of alpha lateralization appeared to depend on
temporal expectation. Alpha lateralization in expect-early
blocks reversed sign around the time of expected memory
use, whereas it remained unchanged in the expect-late
blocks. A direct comparison between expect-early and
2326
Journal of Cognitive Neuroscience
Volume 32, Number 12
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
/
t
o
n
0
5
M
a
y
2
0
2
1
Figure 4. No effect of temporal
expectation on the CDA. (A)
Event-related potentials in
selected posterior electrodes
contra- and ipsilateral to the
side of the relevant memory
items at encoding. (B) CDA
waveforms (contra- vs. ipsilateral
ERPs in (A) as a function
of temporal expectation,
together with the associated
topographies). Dashed vertical
lines indicate the time at which
an early probe could occur.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
0
5
M
a
y
2
0
2
1
DISCUSSION
The current data provide a clear demonstration that
temporal expectations are utilized during visual working
memory retention, with consequences for the accuracy
and speed of the upcoming memory-guided behavior.
The demonstration of a CNV as well as the modulation
in alpha lateralization—that were each sensitive to the
time of expected memory utilization—provided direct ev-
idence that temporal expectations were used during the
memory delay. These demonstrations advance the litera-
tures on both working memory and on temporal expecta-
tions in relevant ways.
Regarding working memory, we show that time plays a
key role in preparing for a memory-guided behavior. In
understanding how working memory bridges past to future,
it is thus relevant to consider not only what items (McNab &
Klingberg, 2008; Vogel et al., 2005; Griffin & Nobre, 2003)
and which features (Niklaus, Nobre, & van Ede, 2017;
Lee et al., 2013; Serences et al., 2009) to retain and prioritize
in working memory. It is also relevant to consider when
our memories will become relevant for guiding a prospec-
tive behavior (see also Zokaei et al., 2019; de Vries, van
Driel, & Olivers, 2017; Olmos-Solis et al., 2017; van Ede
et al., 2017; Wasserman, Grosch, & Nevin, 1982; Perkins,
Lydersen, & Beaman, 1973). Working-memory-guided
behavior was impaired when items were unexpectedly
probed early, but recovered when probed late. Impaired
performance could therefore not be because of a weaker
memory trace per se (if anything, a weaker trace would
be expected after the late interval, because more time
would have passed since encoding). Rather, when unex-
pectedly probed early, participants appeared not to be as
ready to use their memories to guide performance.
Temporal expectations may thus play a key role in shaping
the readiness to utilize the contents of working memory for
guiding behavior—possibly mediated by reconfiguring the
“coding format” of memory content in anticipation of its
expected use (de Vries et al., 2020; Nobre & Stokes, 2019;
Christophel, Iamshchinina, Yan, Allefeld, & Haynes, 2018;
van Loon, Olmos-Solis, Fahrenfort, & Olivers, 2018; Myers
et al., 2017; Lewis-Peacock, Drysdale, Oberauer, & Postle,
2012; Olivers et al., 2011). Notably, whereas prior studies
on “memory readiness” all focused on the prioritization of
individual memory items, in our case, this dynamic change
in readiness applied to the utilization of multiple items in
memory collectively.
Regarding temporal expectation, our data make clear
that the benefits of temporal expectations are not limited
to the type of “pure” perception and action tasks in which
they are traditionally investigated: during “blank inter-
vals” while awaiting a sensory stimulus to detect, identify,
Jin, Nobre, and van Ede
2327
or discriminate, or awaiting a go-signal to execute a sim-
ple motor act (such as a speeded button press). Here, we
have investigated temporal expectations while partici-
pants retained multiple items in working memory. Prior
studies have shown that temporal orienting effects—of
the type studied here—can be abolished in the face of
concurrent working memory demands (Capizzi, Correa,
& Sanabria, 2013; Capizzi, Sanabria, & Correa, 2012). In
these dual-task studies, however, temporal expectations
did not regard the working memory task itself. Rather, work-
ing memory demands served to increase “cognitive load” for
the primary temporal expectation task. When temporal ex-
pectations do concern the working memory task and predict
the time of upcoming working memory utilization, temporal
expectations can have profound effects on working-memory-
guided behavior, as our data make clear. By showing that
temporal expectations can influence the accuracy of
working-memory-guided behavior, our data further suggest
that this influence reflects more than a mere change in
response readiness.
Relation to Other Recent Studies on Temporal
Expectations in Working Memory
Our findings build on recent demonstrations for a role of
temporal expectations in working memory. For example,
temporal expectations regarding the time of stimulus
presentation can facilitate the encoding of sensory infor-
mation into working memory ( Wilsch et al., 2015, 2018).
In contrast, in the current work, temporal expectations
were manifested during memory retention, showing that
temporal expectations can also facilitate how information
that is already in working memory guides behavior.
In other recent studies from our own laboratory (Zokaei
et al., 2019; van Ede et al., 2017), we manipulated temporal
expectations to investigate the flexibility of prioritization
among memory contents (building on, e.g., de Vries et al.,
2020; Rerko & Oberauer, 2013; Lewis-Peacock et al., 2012;
Griffin & Nobre, 2003; Oberauer, 2002). In these tasks,
probes were equally likely to occur early or late, and we
instead varied which memory item was most likely to be
probed at each probe time. We now show that temporal
expectations can also influence memory-guided behavior
in contexts where multiple items in memory collectively
become relevant for guiding behavior. The current task
further allowed us to investigate the CNV and CDA, two
EEG signatures that were not studied in our prior work,
which focused instead on alpha lateralization (van Ede
et al., 2017) and pupil size (Zokaei et al., 2019).
Another study manipulated the time at which visual
templates in working memory would become relevant for
guiding visual search and measured micro-saccadic biases
to template-matching distractors before the search display
(Olmos-Solis et al., 2017). Biases depended on temporal
expectation, consistent with bringing the memory template
into an active state at the time at which it was expected to
guide search (see also de Vries, van Driel, Karacaoglu, &
Olivers, 2018; van Loon, Olmos-Solis, & Olivers, 2017;
Myers, Rohenkohl, et al., 2015). This is akin to holding all
memory content ready for the delayed change detection
in the current task.
Complementing the abovementioned studies, here, we
investigated temporal expectations in the context of a later-
alized change detection task that has been widely used to
study visual working memory and its electrophysiological
correlates (e.g., Hakim et al., 2019; Fukuda, Kang, &
Woodman, 2016; Luria et al., 2016; Vogel et al., 2005; Vogel
& Machizawa, 2004). To our knowledge, expected probe
time has not previously been investigated in the context of
this popular task. We were able to replicate the lateralized
alpha and CDA electrophysiological markers associated with
this change detection task, putting us in the position to ask
whether and how these signatures are also modulated by
temporal expectation.
Alpha Lateralization and CDA Are Differentially
Sensitive to Temporal Expectation
One of the more intriguing findings of the current study
was the differential sensitivity of alpha lateralization versus
the CDA to temporal expectation. Although alpha laterali-
zation underwent significant reorganization in anticipation
of the probe array, the CDA appeared unaffected. Thus, the
two markers likely capture different information during
working memory retention (for related findings, see also
Hakim et al., 2019; Fukuda et al., 2016; Myers, Walther,
et al., 2015).
What might alpha lateralization reflect in our task?
Relevant memory items were lateralized in the encoding
display but were always probed centrally. Any differences
in lateralization must therefore be attributed to differences
in spatial attention and/or retention of the relevant memory
items. We propose that alpha likely reflects shifting of spatial
attention in our task—first to the relevant side of the display
during encoding and early maintenance (regardless of
temporal expectation), followed by a spatial shift to the
center of the screen in anticipation of the central probe
(contingent on temporal expectation). We further speculate
that an overshoot in this spatial shift—possibly reflecting a
physiological “rebound” from the state that preceded this
shift—may account for the apparent “reversal” of lateraliza-
tion. We note, however, that the reversal in the expect-early
condition was not significant by itself but instead contrib-
uted to the significant difference in alpha lateralization
between expect-early and expect-late conditions. This
particular effect thus has to be interpreted with some
caution. Moreover, whether this shift just before the time
of the expected probe includes a concomitant spatial
transformation of the contents of memory ( Woodman,
Vogel, & Luck, 2012) or a pure attention shift in anticipa-
tion of the central task demands remains an interesting
question to address in future research.
The CDA, in contrast, showed no modulation by tempo-
ral expectation, thus providing further evidence against
2328
Journal of Cognitive Neuroscience
Volume 32, Number 12
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
the earlier suggestion that these two signatures reflect a
single underlying neural process (van Dijk et al., 2010)
and for its diminished sensitivity to changes in the focus
of spatial attention (Hakim et al., 2019; Fukuda et al.,
2016). What, then, did the CDA reflect in our task? In our
data, the CDA gradually dissipated with time (regardless
of temporal expectations). On the basis of these data, we
speculate that the CDA reflects a lingering trace resulting
from the spatially biased encoding, which then dissipates
gradually; at least in cases where it is known in advance that
the memory contents will not be probed at the same spatial
location where they were encoded. Whether the CDA
would have remained sustained and/or been sensitive to
temporal expectation if items were probed items at their
encoded locations (thereby not requiring any spatial
transformation of the encoded memory content) remain
relevant questions for future studies.
Conclusion
The current work provides the clear demonstration of the
utility of temporal expectations for guiding upcoming
working-memory-guided behavior, as reflected in behav-
ioral performance and the CNV—extending the literatures
on both working memory and on temporal expectations in
relevant ways. This helps place time on the map as an
important, yet underexplored, variable in working memory
tasks. As a bonus, we have also presented new findings
regarding two key electrophysiological signatures associ-
ated with visual working memory in tasks with lateralized
displays. We hope our findings will motivate relevant new
experimental manipulations that will help further elucidate
the role of these signatures in working memory—taking
into account both the space and the time of prospective
memory use.
Acknowledgments
This research was funded by a Wellcome Trust Senior Investigator
Award (104571/Z/14/Z) and a James S. McDonnell Foundation
Understanding Human Cognition Collaborative Award (220020448)
to A. C. N., a Marie Skłodowska–Curie Fellowship from the
European Commission (ACCESS2WM) to F. v. E., and by the
NIHR Oxford Health Biomedical Research Centre. The Wellcome
Centre for Integrative Neuroimaging is supported by core funding
from the Wellcome Trust (203139/Z/16/Z). The funders had no
role in study design, data collection and analysis, decision to
publish, or preparation of the article.
Reprint requests should be sent to Freek van Ede, Institute for
Brain and Behavior Amsterdam, Department of Experimental
and Applied Psychology, Vrije Universiteit Amsterdam, The
Netherlands, or via e-mail: freek.van.ede@vu.nl.
Author Contributions
Wen Jin: Conceptualization; Data curation; Formal analysis;
Funding acquisition; Investigation; Methodology, Project adminis-
tration; Resources; Software; Supervision; Validation; Visualiza-
tion; Writing - Original Draft; Writing - Review & Editing. Anna C.
Nobre: Conceptualization; Data curation; Formal analysis;
Funding acquisition; Investigation; Methodology; Project admin-
istration; Resources; Software; Supervision; Validation; Visualiza-
tion; Writing - Original Draft, Writing - Review & Editing. Freek
van Ede: Conceptualization; Data curation; Formal analysis;
Funding acquisition; Investigation; Methodology; Project admin-
istration; Resources; Software; Supervision; Validation; Visualiza-
tion; Writing - Original Draft, Writing - Review & Editing.
REFERENCES
Baddeley, A. (1992). Working memory. Science, 255, 556–559.
DOI: https://doi.org/10.1126/science.1736359, PMID: 1736359
Boettcher, S. E. P., Gresch, D., Nobre, A. C., & van Ede, F. (2020).
Output planning at the input stage: Action imprinting for future
memory-guided behaviour. Neuron. DOI: https://doi.org/
10.2139/ssrn.3550240
Boettcher, S. E. P., Stokes, M. G., Nobre, A. C., & van Ede, F.
(2020). One thing leads to another: Anticipating visual
object identity based on associative-memory templates.
Journal of Neuroscience, 40, 4010–4020. DOI: https://doi.
org/10.1523/JNEUROSCI.2751-19.2020, PMID: 32284338,
PMCID: PMC7219293
Capizzi, M., Correa, Á., & Sanabria, D. (2013). Temporal
orienting of attention is interfered by concurrent working
memory updating. Neuropsychologia, 51, 326–339. DOI:
https://doi.org/10.1016/j.neuropsychologia.2012.10.005,
PMID: 23085375
Capizzi, M., Sanabria, D., & Correa, Á. (2012). Dissociating
controlled from automatic processing in temporal preparation.
Cognition, 123, 293–302. DOI: https://doi.org/10.1016/j.
cognition.2012.02.005, PMID: 22397820
Carlisle, N. B., Arita, J. T., Pardo, D., & Woodman, G. F. (2011).
Attentional templates in visual working memory. Journal of
Neuroscience, 31, 9315–9322. DOI: https://doi.org/10.1523/
JNEUROSCI.1097-11.2011, PMID: 21697381, PMCID:
PMC3147306
Chatham, C. H., & Badre, D. (2015). Multiple gates on working
memory. Current Opinion in Behavioral Sciences, 1, 23–31.
DOI: https://doi.org/10.1016/j.cobeha.2014.08.001, PMID:
26719851, PMCID: PMC4692183
Chatrian, G. E., Lettich, E., & Nelson, P. L. (1985). Ten percent
electrode system for topographic studies of spontaneous and
evoked EEG activities. American Journal of EEG Technology,
25, 83–92. DOI: https://doi.org/10.1080/00029238.1985.
11080163
Chelazzi, L., Miller, E. K., Duncan, J., & Desimone, R. (1993).
A neural basis for visual search in inferior temporal cortex.
Nature, 363, 345–347. DOI: https://doi.org/10.1038/363345a0,
PMID: 8497317
Christophel, T. B., Iamshchinina, P., Yan, C., Allefeld, C., &
Haynes, J.-D. (2018). Cortical specialization for attended
versus unattended working memory. Nature Neuroscience,
21, 494–496. DOI: https://doi.org/10.1038/s41593-018-0094-4,
PMID: 29507410
Coull, J. T., & Nobre, A. C. (1998). Where and when to pay
attention: The neural systems for directing attention to spatial
locations and to time intervals as revealed by both PET and
fMRI. Journal of Neuroscience, 18, 7426–7435. DOI: https://
doi.org/10.1523/JNEUROSCI.18-18-07426.1998, PMID: 9736662,
PMCID: PMC6793260
Cousineau, D. (2005). Confidence intervals in within-subject
designs: A simpler solution to Loftus and Masson’s method.
Tutorials in Quantitative Methods for Psychology, 1, 42–45.
DOI: https://doi.org/10.20982/tqmp.01.1.p042
Cravo, A. M., Rohenkohl, G., Santos, K. M., & Nobre, A. C. (2017).
Temporal anticipation based on memory. Journal of Cognitive
Jin, Nobre, and van Ede
2329
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
Neuroscience, 29, 2081–2089. DOI: https://doi.org/10.1162/
jocn_a_01172, PMID: 28777060, PMCID: PMC5884434
D’Esposito, M., & Postle, B. R. (2015). The cognitive neuroscience
of working memory. Annual Review of Psychology, 66, 115–142.
DOI: https://doi.org/10.1146/annurev-psych-010814-015031,
PMID: 25251486, PMCID: PMC4374359
de Vries, I. E. J., Slagter, H. A., & Olivers, C. N. L. (2020).
Oscillatory control over representational states in working
memory. Trends in Cognitive Sciences, 24, 150–162. DOI:
https://doi.org/10.1016/j.tics.2019.11.006, PMID: 31791896
de Vries, I. E. J., van Driel, J., Karacaoglu, M., & Olivers, C. N. L.
(2018). Priority switches in visual working memory are supported
by frontal delta and posterior alpha interactions. Cerebral Cortex,
28, 4090–4104. DOI: https://doi.org/10.1093/cercor/bhy223,
PMID: 30215669, PMCID: PMC6188546
de Vries, I. E. J., van Driel, J., & Olivers, C. N. L. (2017). Posterior α
EEG dynamics dissociate current from future goals in working
memory-guided visual search. Journal of Neuroscience, 37,
1591–1603. DOI: https://doi.org/10.1523/JNEUROSCI.2945-
16.2016, PMID: 28069918, PMCID: PMC5299573
Fukuda, K., Kang, M.-S., & Woodman, G. F. (2016). Distinct neural
mechanisms for spatially lateralized and spatially global visual
working memory representations. Journal of Neurophysiology,
116, 1715–1727. DOI: https://doi.org/10.1152/jn.00991.2015,
PMID: 27440249, PMCID: PMC5144708
Griffin, I. C., & Nobre, A. C. (2003). Orienting attention to
locations in internal representations. Journal of Cognitive
Neuroscience, 15, 1176–1194. DOI: https://doi.org/10.1162/
089892903322598139, PMID: 14709235
Hakim, N., Adam, K. C. S., Gunseli, E., Awh, E., & Vogel, E. K.
(2019). Dissecting the neural focus of attention reveals distinct
processes for spatial attention and object-based storage in
visual working memory. Psychological Science, 30, 526–540.
DOI: https://doi.org/10.1177/0956797619830384, PMID:
30817220, PMCID: PMC6472178
Heideman, S. G., Quinn, A. J., Woolrich, M. W., van Ede, F., &
Nobre, A. C. (2020). Dissecting beta-state changes during timed
movement preparation in Parkinson’s disease. Progress in
Neurobiology, 184, 101731. DOI: https://doi.org/10.1016/j.
pneurobio.2019.101731, PMID: 31778771, PMCID: PMC6977086
Heideman, S. G., Rohenkohl, G., Chauvin, J. J., Palmer, C. E.,
van Ede, F., & Nobre, A. C. (2018). Anticipatory neural dynamics
of spatial-temporal orienting of attention in younger and older
adults. Neuroimage, 178, 46–56. DOI: https://doi.org/10.1016/j.
neuroimage.2018.05.002, PMID: 29733953, PMCID:
PMC6057272
Heideman, S. G., van Ede, F., & Nobre, A. C. (2018). Temporal
alignment of anticipatory motor cortical beta lateralisation
in hidden visual-motor sequences. European Journal of
Neuroscience, 48, 2684–2695. DOI: https://doi.org/10.1111/
ejn.13700, PMID: 28921756, PMCID: PMC6220967
Janssen, P., & Shadlen, M. N. (2005). A representation of the
hazard rate of elapsed time in macaque area LIP. Nature
Neuroscience, 8, 234–241. DOI: https://doi.org/10.1038/
nn1386, DOI: https://doi.org/10.1038/nn0305-396b, PMID:
15657597
Jaramillo, S., & Zador, A. M. (2011). The auditory cortex mediates
the perceptual effects of acoustic temporal expectation. Nature
Neuroscience, 14, 246–251. DOI: https://doi.org/10.1038/
nn.2688, PMID: 21170056, PMCID: PMC3152437
Jones, M. R., & Boltz, M. (1989). Dynamic attending and responses
to time. Psychological Review, 96, 459–491. DOI: https://doi.
org/10.1037/0033-295X.96.3.459, PMID: 2756068
Kuo, B.-C., Stokes, M. G., & Nobre, A. C. (2012). Attention
modulates maintenance of representations in visual short-term
memory. Journal of Cognitive Neuroscience, 24, 51–60. DOI:
https://doi.org/10.1162/jocn_a_00087, PMID: 21736457,
PMCID: PMC3480577
Lakatos, P., Karmos, G., Mehta, A. D., Ulbert, I., & Schroeder, C. E.
(2008). Entrainment of neuronal oscillations as a mechanism of
attentional selection. Science, 320, 110–113. DOI: https://doi.
org/10.1126/science.1154735, PMID: 18388295
Lange, K., & Röder, B. (2006). Orienting attention to points in
time improves stimulus processing both within and across
modalities. Journal of Cognitive Neuroscience, 18, 715–729.
DOI: https://doi.org/10.1162/jocn.2006.18.5.715, PMID:
16768372
Lee, S.-H., Kravitz, D. J., & Baker, C. I. (2013). Goal-dependent
dissociation of visual and prefrontal cortices during working
memory. Nature Neuroscience, 16, 997–999. DOI: https://
doi.org/10.1038/nn.3452, PMID: 23817547, PMCID:
PMC3781947
Lewis-Peacock, J. A., Drysdale, A. T., Oberauer, K., & Postle, B. R.
(2012). Neural evidence for a distinction between short-term
memory and the focus of attention. Journal of Cognitive
Neuroscience, 24, 61–79. DOI: https://doi.org/10.1162/
jocn_a_00140, PMID: 21955164, PMCID: PMC3222712
Los, S. A., Kruijne, W., & Meeter, M. (2017). Hazard versus
history: Temporal preparation is driven by past experience.
Journal of Experimental Psychology: Human Perception and
Performance, 43, 78–88. DOI: https://doi.org/10.1037/
xhp0000279, PMID: 27808547
Lozano-Soldevilla, D., ter Huurne, N., Cools, R., & Jensen, O.
(2014). GABAergic modulation of visual gamma and alpha
oscillations and its consequences for working memory
performance. Current Biology, 24, 2878–2887. DOI: https://
doi.org/10.1016/j.cub.2014.10.017, PMID: 25454585
Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during visual
search: Evidence from human electrophysiology. Journal
of Experimental Psychology: Human Perception and
Performance, 20, 1000–1014. DOI: https://doi.org/10.1037/
0096-1523.20.5.1000
Luria, R., Balaban, H., Awh, E., & Vogel, E. K. (2016). The
contralateral delay activity as a neural measure of visual
working memory. Neuroscience & Biobehavioral Reviews,
62, 100–108. DOI: https://doi.org/10.1016/j.neubiorev.2016.
01.003, PMID: 26802451, PMCID: PMC4869985
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience Methods,
164, 177–190. DOI: https://doi.org/10.1016/j.jneumeth.2007.
03.024, PMID: 17517438
McNab, F., & Klingberg, T. (2008). Prefrontal cortex and
basal ganglia control access to working memory. Nature
Neuroscience, 11, 103–107. DOI: https://doi.org/10.1038/
nn2024, PMID: 18066057
Miniussi, C., Wilding, E. L., Coull, J. T., & Nobre, A. C. (1999).
Orienting attention in time: Modulation of brain potentials.
Brain, 122, 1507–1518. DOI: https://doi.org/10.1093/brain/
122.8.1507, PMID: 10430834
Myers, N. E., Rohenkohl, G., Wyart, V., Woolrich, M. W., Nobre,
A. C., & Stokes, M. G. (2015). Testing sensory evidence against
mnemonic templates. eLife, 4, e09000. DOI: https://doi.org/
10.7554/eLife.09000.018, DOI: https://doi.org/10.7554/
eLife.09000, PMID: 26653854, PMCID: PMC4755744
Myers, N. E., Stokes, M. G., & Nobre, A. C. (2017). Prioritizing
information during working memory: Beyond sustained
internal attention. Trends in Cognitive Sciences, 21, 449–461.
DOI: https://doi.org/10.1016/j.tics.2017.03.010, PMID:
28454719, PMCID: PMC7220802
Myers, N. E., Walther, L., Wallis, G., Stokes, M. G., & Nobre, A. C.
(2015). Temporal dynamics of attention during encoding
versus maintenance of working memory: Complementary
views from event-related potentials and alpha-band
oscillations. Journal of Cognitive Neuroscience, 27, 492–508.
DOI: https://doi.org/10.1162/jocn_a_00727, PMID: 25244118,
PMCID: PMC4678590
2330
Journal of Cognitive Neuroscience
Volume 32, Number 12
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
t
/
o
n
0
5
M
a
y
2
0
2
1
Niklaus, M., Nobre, A. C., & van Ede, F. (2017). Feature-based
attentional weighting and spreading in visual working memory.
Scientific Reports, 7, 42384. DOI: https://doi.org/10.1038/
srep42384, PMID: 28233830, PMCID: PMC5324041
Nobre, A. C. (2001). Orienting attention to instants in time.
Neuropsychologia, 39, 1317–1328. DOI: https://doi.org/
10.1016/S0028-3932(01)00120-8
Nobre, A. C., & Stokes, M. G. (2019). Premembering experience:
A hierarchy of time-scales for proactive attention. Neuron, 104,
132–146. DOI: https://doi.org/10.1016/j.neuron.2019.08.030,
PMID: 31600510, PMCID: PMC6873797
Nobre, A. C., & van Ede, F. (2018). Anticipated moments: Temporal
structure in attention. Nature Reviews Neuroscience, 19, 34–48.
DOI: https://doi.org/10.1038/nrn.2017.141, PMID: 29213134
Oberauer, K. (2002). Access to information in working
memory: Exploring the focus of attention. Journal of
Experimental Psychology: Learning, Memory, and
Cognition, 28, 411–421. DOI: https://doi.org/10.1037/0278-
7393.28.3.411
Olivers, C. N. L., Peters, J., Houtkamp, R., & Roelfsema, P. R.
(2011). Different states in visual working memory: When it
guides attention and when it does not. Trends in Cognitive
Sciences, 15, 327–334. DOI: https://doi.org/10.1016/j.tics.2011.
05.004, PMID: 21665518
Olmos-Solis, K., van Loon, A. M., Los, S. A., & Olivers, C. N. L.
(2017). Oculomotor measures reveal the temporal dynamics of
preparing for search. Progress in Brain Research, 236, 1–23.
DOI: https://doi.org/10.1016/bs.pbr.2017.07.003, PMID:
29157407
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational
Intelligence and Neuroscience, 2011, 156869. DOI: https://
doi.org/10.1155/2011/156869, PMID: 21253357, PMCID:
PMC3021840
Perkins, D., Lydersen, T., & Beaman, D. (1973). Acquisition
under mixed-delay and multiple-delay matching-to-sample.
Psychological Reports, 32, 635–640. DOI: https://doi.org/
10.2466/pr0.1973.32.2.635
Perrin, F., Pernier, J., Bertrand, O., & Echallier, J. F. (1989).
Spherical splines for scalp potential and current density mapping.
Electroencephalography and Clinical Neurophysiology, 72,
184–187. DOI: https://doi.org/10.1016/0013-4694(89)90180-6
Praamstra, P. (2006). Prior information of stimulus location:
Effects on ERP measures of visual selection and response
selection. Brain Research, 1072, 153–160. DOI: https://doi.
org/10.1016/j.brainres.2005.11.098, PMID: 16406014
Praamstra, P., Kourtis, D., Kwok, H. F., & Oostenveld, R. (2006).
Neurophysiology of implicit timing in serial choice reaction-
time performance. Journal of Neuroscience, 26, 5448–5455.
DOI: https://doi.org/10.1523/JNEUROSCI.0440-06.2006,
PMID: 16707797, PMCID: PMC6675318
Rainer, G., Rao, S. C., & Miller, E. K. (1999). Prospective coding for
objects in primate prefrontal cortex. Journal of Neuroscience,
19, 5493–5505. DOI: https://doi.org/10.1523/JNEUROSCI.19-
13-05493.1999, PMID: 10377358, PMCID: PMC6782318
Rerko, L., & Oberauer, K. (2013). Focused, unfocused, and
defocused information in working memory. Journal of
Experimental Psychology: Learning, Memory, and Cognition,
39, 1075–1096. DOI: https://doi.org/10.1037/a0031172, PMID:
23421511
Robitaille, N., & Jolicoeur, P. (2006). Fundamental properties of
the N2pc as an index of spatial attention: Effects of masking.
Canadian Journal of Experimental Psychology, 60, 101–111.
DOI: https://doi.org/10.1037/cjep2006011, PMID: 17133886
Rohenkohl, G., Cravo, A. M., Wyart, V., & Nobre, A. C. (2012).
Temporal expectation improves the quality of sensory
information. Journal of Neuroscience, 32, 8424–8428. DOI:
https://doi.org/10.1523/JNEUROSCI.0804-12.2012, PMID:
22699922, PMCID: PMC4235252
Sauseng, P., Klimesch, W., Heise, K. F., Gruber, W. R., Holz, E.,
Karim, A. A., et al. (2009). Brain oscillatory substrates of
visual short-term memory capacity. Current Biology, 19,
1846–1852. DOI: https://doi.org/10.1016/j.cub.2009.08.062,
PMID: 19913428
Schmidt, J., & Zelinsky, G. J. (2017). Adding details to the
attentional template offsets search difficulty: Evidence
from contralateral delay activity. Journal of Experimental
Psychology: Human Perception and Performance, 43,
429–437. DOI: https://doi.org/10.1037/xhp0000367, PMID:
28240928
Schoffelen, J.-M., Oostenveld, R., & Fries, P. (2005). Neuronal
coherence as a mechanism of effective corticospinal interaction.
Science, 308, 111–113. DOI: https://doi.org/10.1126/science.
1107027, PMID: 15802603
Serences, J. T., Ester, E. F., Vogel, E. K., & Awh, E. (2009).
Stimulus-specific delay activity in human primary visual cortex.
Psychological Science, 20, 207–214. DOI: https://doi.org/
10.1111/j.1467-9280.2009.02276.x, PMID: 19170936, PMCID:
PMC2875116
Töllner, T., Müller, H. J., & Zehetleitner, M. (2012). Top–down
dimensional weight set determines the capture of visual
attention: Evidence from the PCN component. Cerebral Cortex,
22, 1554–1563. DOI: https://doi.org/10.1093/cercor/bhr231,
PMID: 21903593
van Dijk, H., van der Werf, J., Mazaheri, A., Medendorp, W. P.,
& Jensen, O. (2010). Modulations in oscillatory activity with
amplitude asymmetry can produce cognitively relevant event-
related responses. Proceedings of the National Academy of
Sciences, U.S.A., 107, 900–905. DOI: https://doi.org/10.1073/
pnas.0908821107, PMID: 20080773, PMCID: PMC2818898
van Ede, F. (2018). Mnemonic and attentional roles for states of
attenuated alpha oscillations in perceptual working memory:
A review. European Journal of Neuroscience, 48, 2509–2515.
DOI: https://doi.org/10.1111/ejn.13759, PMID: 29068095,
PMCID: PMC6220786
van Ede, F. (2020). Visual working memory and action:
Functional links and bi-directional influences. Visual Cognition.
DOI: https://doi.org/10.1080/13506285.2020.1759744
van Ede, F., Chekroud, S. R., Stokes, M. G., & Nobre, A. C. (2019).
Concurrent visual and motor selection during visual working
memory guided action. Nature Neuroscience, 22, 477–483.
DOI: https://doi.org/10.1038/s41593-018-0335-6, PMID:
30718904, PMCID: PMC6420070
van Ede, F., Jensen, O., & Maris, E. (2010). Tactile expectation
modulates pre-stimulus β-band oscillations in human
sensorimotor cortex. Neuroimage, 51, 867–876. DOI: https://
doi.org/10.1016/j.neuroimage.2010.02.053, PMID: 20188186
van Ede, F., & Maris, E. (2016). Physiological plausibility can
increase reproducibility in cognitive neuroscience. Trends in
Cognitive Sciences, 20, 567–569. DOI: https://doi.org/
10.1016/j.tics.2016.05.006, PMID: 27233147
van Ede, F., Niklaus, M., & Nobre, A. C. (2017). Temporal
expectations guide dynamic prioritization in visual working
memory through attenuated alpha oscillations. Journal of
Neuroscience, 37, 437–445. DOI: https://doi.org/10.1523/
JNEUROSCI.2272-16.2016, PMID: 28077721, PMCID:
PMC5242399
van Elswijk, G., Kleine, B. U., Overeem, S., & Stegeman, D. F.
(2007). Expectancy induces dynamic modulation of corticospinal
excitability. Journal of Cognitive Neuroscience, 19, 121–131.
DOI: https://doi.org/10.1162/jocn.2007.19.1.121, PMID:
17214569
Vangkilde, S., Coull, J. T., & Bundesen, C. (2012). Great
expectations: Temporal expectation modulates perceptual
processing speed. Journal of Experimental Psychology:
Jin, Nobre, and van Ede
2331
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
.
f
t
o
n
0
5
M
a
y
2
0
2
1
Human Perception and Performance, 38, 1183–1191.
DOI: https://doi.org/10.1037/a0026343, PMID: 22250866
van Loon, A. M., Olmos-Solis, K., Fahrenfort, J. J., & Olivers,
C. N. L. (2018). Current and future goals are represented in
opposite patterns in object-selective cortex. eLife, 7,
e38677. DOI: https://doi.org/10.7554/eLife.38677, PMID:
30394873, PMCID: PMC6279347
van Loon, A. M., Olmos-Solis, K., & Olivers, C. N. L. (2017). Subtle
eye movement metrics reveal task-relevant representations
prior to visual search. Journal of Vision, 17, 13. DOI: https://
doi.org/10.1167/17.6.13, PMID: 28637052
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity predicts
individual differences in visual working memory capacity.
Nature, 428, 748–751. DOI: https://doi.org/10.1038/
nature02447, PMID: 15085132
Vogel, E. K., McCollough, A. W., & Machizawa, M. G. (2005).
Neural measures reveal individual differences in controlling
access to working memory. Nature, 438, 500–503. DOI:
https://doi.org/10.1038/nature04171, PMID: 16306992
Wallis, G., Stokes, M., Cousijn, H., Woolrich, M., & Nobre, A. C.
(2015). Frontoparietal and cingulo-opercular networks play
dissociable roles in control of working memory. Journal of
Cognitive Neuroscience, 27, 2019–2034. DOI: https://doi.
org/10.1162/jocn_a_00838, PMID: 26042457
Walter, W. G., Cooper, R., Aldridge, V. J., McCallum, W. C.,
& Winter, A. L. (1964). Contingent negative variation: An
electric sign of sensori-motor association and expectancy in
the human brain. Nature, 203, 380–384. DOI: https://doi.org/
10.1038/203380a0, PMID: 14197376
Wasserman, E. A., Grosch, J., & Nevin, J. A. (1982). Effects of
signaled retention intervals on pigeon short-term memory.
Animal Learning & Behavior, 10, 330–338. DOI: https://doi.
org/10.3758/BF03213719
Weinberg, H. (1972). The contingent negative variation:
Its relation to feedback and expectant attention.
Neuropsychologia, 10, 299–306. DOI: https://doi.org/
10.1016/0028-3932(72)90021-8
Wilsch, A., Henry, M. J., Herrmann, B., Herrmann, C. S., &
Obleser, J. (2018). Temporal expectation modulates the cortical
dynamics of short-term memory. Journal of Neuroscience, 38,
7428–7439. DOI: https://doi.org/10.1523/JNEUROSCI.2928-
17.2018, PMID: 30012685, PMCID: PMC6596135
Wilsch, A., Henry, M. J., Herrmann, B., Maess, B., & Obleser, J.
(2015). Alpha oscillatory dynamics index temporal
expectation benefits in working memory. Cerebral Cortex,
25, 1938–1946. DOI: https://doi.org/10.1093/cercor/bhu004,
PMID: 24488943
Woodman, G. F., Vogel, E. K., & Luck, S. J. (2012). Flexibility in
visual working memory: Accurate change detection in the face
of irrelevant variations in position. Visual Cognition, 20, 1–28.
DOI: https://doi.org/10.1080/13506285.2011.630694, PMID:
22287933, PMCID: PMC3266348
Zokaei, N., Board, A. G., Manohar, S. G., & Nobre, A. C. (2019).
Modulation of the pupillary response by the content of visual
working memory. Proceedings of the National Academy
of Sciences, U.S.A., 116, 22802–22810. DOI: https://doi.org/
10.1073/pnas.1909959116, PMID: 31636213, PMCID:
PMC6842592
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
2
-
2
p
3
d
2
f
0
/
2
3
0
2
1
/
4
1
1
2
5
/
7
2
3
o
2
c
0
n
_
/
a
1
_
8
0
6
1
2
6
2
2
6
6
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
2
0
6
7
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
/
.
o
n
0
5
M
a
y
2
0
2
1
2332
Journal of Cognitive Neuroscience
Volume 32, Number 12