Taking Attention Out of Context: Frontopolar Transcranial
Magnetic Stimulation Abolishes the Formation of
New Context Memories in Visual Search
Artyom Zinchenko1, Markus Conci1, Paul C. J. Taylor1,
Hermann J. Müller1,2, and Thomas Geyer1
Astratto
■ This study investigates the causal contribution of the left
frontopolar cortex (FPC) to the processing of violated expec-
tations from learned target–distractor spatial contingencies
during visual search. The experiment consisted of two phases:
learning and test. Participants searched for targets presented
either among repeated or nonrepeated target–distractor con-
figurations. Prior research showed that repeated encounters of
identically arranged displays lead to memory about these
arrays, which then can come to guide search (contextual cue-
ing effect). The crucial manipulation was a change of the
target location, in a nevertheless constant distractor layout, at
the transition from learning to test. In addition to this change,
we applied repetitive transcranial magnetic stimulation (rTMS)
over the left lateral FPC, over a posterior control site, or no
rTMS at all (baseline; between-group manipulation) to see
how FPC rTMS influences the ability of observers to adapt
context-based memories acquired in the training phase. IL
learning phase showed expedited search in repeated relative
to nonrepeated displays, with this context-based facilitation be-
ing comparable across all experimental groups. For the test
phase, the recovery of cueing was critically dependent on the
stimulation site: Although there was evidence of context adap-
tation toward the end of the experiment in the occipital and
no-rTMS conditions, observers with FPC rTMS showed no
evidence of relearning at all after target location changes.
This finding shows that FPC plays an important role in the reg-
ulation of prediction errors in statistical context learning, così
contributing to an update of the spatial target–distractor con-
tingencies after target position changes in learned spatial
arrays. ■
INTRODUCTION
Prediction errors are key features in current accounts of
perception, apprendimento, and action. At the heart of these so-
called predictive coding theories (Friston, 2010; Rao &
Ballard, 1999) is the idea that the brain is essentially an
inference machine that actively tries to explain its sensa-
zioni. In this regard, hierarchically higher brain areas
modulate (“predict”) the tuning properties of lower level
areas via feedback processing. Lower level areas, in turn,
compute the differences between top–down expecta-
tions and the actual sensory input, with this difference
providing a prediction error signal. In this scheme, pre-
diction errors serve as important learning signals that
may cause a reevaluation and thus an update of current
expectations.
In this study, we examined the contribution of the
frontopolar cortex (FPC) to the regulation of prediction
errors in a selective attention, questo è, visual search, task.
In recent years, contributions of the anterior pFC have
1Ludwig-Maximilians-Universität München, 2Birkbeck, Università
of London
© 2018 Istituto di Tecnologia del Massachussetts
been shown in a variety of “easy” feature singleton and
“difficult” conjunction search tasks (see Pollmann,
Weidner, Müller, & von Cramon, 2006, for an overview).
The critical finding in these studies is that of increased
anterior pFC activations following a change of the tar-
get’s perceptual dimension (per esempio., when an orientation-
defined target is followed by a color-defined target)
relative to a dimension repeat trial (per esempio., a color target
is followed by another color target). According to predic-
tive coding theories, the enhanced prefrontal activations
following dimension change trials may reflect a larger
prediction error, relative to dimension repeat trials,
because expectations about the target’s (repeated)
dimensional identity are violated (see Summerfield,
Trittschuh, Monti, Mesulam, & Egner, 2008, for the idea
that BOLD signal amplitudes index top–down expec-
tations). Inoltre, it has been suggested that the
anterior pFC regulates prediction errors by attentionally
weighting/amplifying the target’s perceptual dimension
to determine the presence and location of the search
target at an overall salience or master map of the visual
array (Weidner, Pollmann, Müller, & von Cramon, 2002).
Although these findings may suggest a contribution of
Journal of Cognitive Neuroscience 31:3, pag. 442–452
doi:10.1162/jocn_a_01358
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
the pFC to predictive coding, Per esempio, by shifting
attention toward the (changed) perceptual dimension
of the current target and thus increasing the efficacy of
perceptual inference (per esempio., Kok, Rahnev, Jehee, Lau, &
de Lange, 2012), more recent studies show that the
anterior pFC, besides its effects on dimension-based
Attenzione, is also involved in the control of visuospatial
attention following target location changes in learned
display arrangements (Pollmann & Manginelli, 2009;
Pollmann et al., 2007).
Pollmann and Manginelli (2009) measured fMRI
(BOLD) activations while having their observers perform
a relatively difficult letter search task, divided into a learn-
ing phase and a test phase. Unbeknown to observers, In
half of the trials, the target was presented within a con-
stant spatial layout of distractor elements, enabling them
to associate the target with the constant distractor con-
figuration (see Chun & Jiang, 1998). Following initial
apprendimento, in the subsequent test phase, a change of the
target position was introduced in an otherwise constant
distractor array. This target location change was imple-
mented to test whether and how context memory adapts
to this change and which neuronal structures support the
adaptation. Pollmann and Manginelli (2009) observed a
postchange reduction of contextual cueing in terms of
search RTs, accompanied by a significant increase in
BOLD activations in the left FPC, in addition to various
other regions of the frontoparietal attention network
(Corbetta & Shulman, 2002). Although fMRI activations
were initially reduced for repeated relative to nonre-
peated displays, the processing of repeated (relative to
nonrepeated) displays led to stronger activations after
target position changes in the subsequent test phase.
Importantly, the magnitude of postchange FPC activa-
tions for repeated displays was positively correlated with
the strength of the contextual cueing effect in the initial
learning phase, although there was no systematic rela-
tionship between postchange contextual cueing and
FPC activations. Based on these findings, Pollmann and
Manginelli (2009) hypothesized that FPC serves a dual
function in the processing of changes in learned, contex-
tual cueing arrays: first, to detect changes in these arrays,
and second, to inform other structures of the frontopar-
ietal network about display changes. Via the latter, FPC
would effectively help make the subtle target position
change more salient, thus supporting frontoparietal
structures in the reallocation of attention to the changed
target position. According to this view, FPC would facili-
tate the adaptation of contextual cueing by initiating
attention shifts toward the changed target position.
Tuttavia, given that the adaptation process is typically
not very efficient (see below), there is at least one alter-
native interpretation of (increased) FPC activations in the
processing of changed contextual cueing displays, namely,
that these activations reflect additional cognitive (execu-
tive) processes required for the suppression of the cur-
rent sensory input, questo è, the changed target position.
In other words, FPC would contribute to the enhanced
processing of initially learned (old) distractor arrange-
menti, rather than facilitating the processing of changed
(new) target locations in these arrangements. IL
current study used rTMS to evaluate the two alternative
accounts of FPC function in a context memory adapta-
tion task.
FPC as Facilitator of Old or New Target Positions?
The ability to form new associations between target
positions and the spatial pattern formed by the repeated
distractor locations is strongly affected by already existing
context memory representations (Zellin, von Mühlenen,
Müller, & Conci, 2014; Zellin, Conci, von Mühlenen, &
Müller, 2011, 2013; Conci & Müller, 2012; Conci, Sun,
& Müller, 2011; Makovski & Jiang, 2010). Collectively,
these studies show that learning of target–distractor
associations is typically limited to a single target location
within a given arrangement (Zellin et al., 2011) and that
adaptation of contextual cueing after a target location
change is rather inefficient, with a persisting atten-
tional bias toward the old target position (Zellin et al.,
2013, 2014; Conci & Müller, 2012; Conci et al., 2011;
Manginelli & Pollmann, 2009). One possible reason why
contextual cueing exhibits resistance to adapt to changed
target locations is that context learning is resource de-
manding, relying on visual working memory resources
(per esempio., Annac et al., 2013; Jungé, Scholl, & Chun, 2007;
Jiang & Leung, 2005). Così, it may cease after a number
of repeated encounters of the same arrangements, as the
target–distractor relations are typically stable over time,
so that there would be no additional processing advan-
tages if resource-dependent learning were to continue.
A related idea is to conceive contextual cueing as a pre-
dictive coding mechanism (per esempio., Zinchenko, Conci,
Müller, & Geyer, 2018; Conci, Zellin, & Müller, 2012) Quello
takes into account the distribution or reliability of sta-
tistical signals (cioè., of repeated search arrays) in con-
text learning. For instance, over the course of learning,
observers may come to form hypotheses about regu-
larities (or their absence) in the displays encountered,
which then influence the way they process target loca-
tion changes. Così, a search scenario with only 50% Rif-
peated displays may not be experienced as a “statistically
rich” sensory environment, causing observers to “turn
off” further learning. In terms of perceptual inference,
the turning off of learning would be equivalent to the
maintenance of the already established context mem-
ory. From this perspective, subsequent to a target location
change, observers may still exhibit a strong tendency to
expect the target at the old, questo è, learned, target posi-
zione (Vedere, per esempio., Manginelli & Pollmann, 2009, or Zellin
et al., 2014), accompanied by a suppression of prediction
errors signaling a mismatch between the current and
learned target positions.
Zinchenko et al.
443
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
In summary, Pollmann and Manginelli (2009) ob-
served increased FPC activations following target loca-
tion changes in learned search arrays. Although these
activations may reflect prediction errors due to FPC be-
ing involved in the matching of learned predictions and
sensory input, what role the FPC plays in the regulation
of these errors remains an open issue. Conceivably,
FPC might help minimize prediction errors by facil-
itating the processing of the new, questo è, relocated,
target position, thus supporting the (in any case slow)
adaptation of context memories to the new distractor-
to-target spatial relations (“FPC+” hypothesis). Alter-
natively, FPC might contribute to “resolving” prediction
errors through suppression of the changed sensory
input (“FPC−” hypothesis), as a result of which the
priority assigned to the initially learned target position
would tend to be maintained. In short, FPC might be
involved either in reinforcing processing of the already
learned target location while suppressing the changed
input (FPC−) or in facilitating adaptation to a new
target location in an otherwise unchanged array of
distractors (FPC+).
Rationale of This Study
On this background, this study was designed to investi-
gate the role of the FPC in the adaptation of contex-
tual cueing. Specifically, we applied low-frequency
rTMS to examine the contribution of the left FPC in
the processing of repeated distractor configurations
after target location changes. rTMS may reduce and/or
inhibit excitability in the underlying cortex (Filipović,
Rothwell, & Bhatia, 2010; Hoffman & Cavus, 2002;
Wassermann & Lisanby, 2001). For this reason, we
applied rTMS to FPC after an initial learning phase and
before a subsequent test phase to examine its effects on
context adaptation. According to the FPC− hypothesis,
prediction errors are minimized by suppressing the
changed target location (thus enhancing the initial
target location). A transient disruption of this inhib-
itory process should, in turn, facilitate context adap-
tazione, and as a result, the changed target location
would be incorporated more effectively into the ex-
isting context representation. By contrast, the FPC+
hypothesis would postulate a relatively rapid adapta-
zione, facilitating learning of the changed input. In this
case, rTMS-induced interference of FPC should weaken
adaptation, thus supporting the maintenance of exist-
ing target–distractor associations and impeding posi-
tional biasing toward the changed target locations.
rTMS over the FPC was compared against rTMS over
a control (posterior) region as well as a no-TMS base-
line condition to ensure that variations of the adapta-
tion of contextual cueing were causally linked to FPC
functioning rather than being associated with non-
specific effects that may have resulted from the rTMS
methodology.
METHODS
Participants
Forty-two right-handed participants naive to the purpose
of the study took part in the experiment (mean age =
26.5 years, 24 women). The sample size was determined
on the basis of previous contextual cueing studies (per esempio.,
Zellin et al., 2013; Geyer, Zehetleitner, & Müller, 2010;
Schankin & Schubö, 2009; Chun & Jiang, 1998), aiming
for 85% power to detect an f(U ) effect size of 1.0 (partial
η2 = .5, three groups, four measurements; interaction
of Context (repeated, novel) × Epoch (1–4)) with an
alpha level of .05 and a nonsphericity correction of 1.
All participants reported normal or corrected-to-normal
vision and were randomly assigned to one of three
groups (con 14 observers each): FPC rTMS, occipital
rTMS, and no rTMS. The groups did not differ with re-
gard to their mean age and sex: FPC rTMS, mean age =
26.2 years, SD = 2.7 years, nine women; occipital rTMS,
mean age = 26.9 years, SD = 3.8 years, eight women;
no rTMS, mean age = 26.6 years, SD = 4.1 years, seven
women (all ts < 0.7, ps > .5). The study was approved
by the ethics committee of the Department Psychology
of Ludwig-Maximilians-Universität München. Observers
provided written informed consent before the experiment
and received A20 (∼$23.50) for their participation. All col-
lected data is available through Open Science Framework
(https://osf.io/zhk2j/ ).
Apparatus and Stimuli
The experimental routine was programmed in MATLAB
with Psychtoolbox extensions (Brainard, 1997; Pelli,
1997) and was run on an Intel PC under the Windows 7
operating system. Participants were seated in a dimly lit
room in front of a 23-in. LCD monitor (ASUS; refresh
rate = 60 Hz, display resolution = 1920 × 1080 pixels)
at a viewing distance of 80 cm (controlled by a chin rest).
The search displays consisted of 12 gray items (lumi-
nance = 1.0 cd/m 2; 1 target and 11 distractors)
presented against a black background (0.11 cd/m2). Tutto
stimuli extended 0.35° of visual angle in both width and
height. The items were arranged on four (invisible) con-
centric circles around the display center (with a radius of
1.74°, 3.48°, 5.22°, and 6.96° for Circles 1 through 4,
rispettivamente). The target was always positioned on the
third circle. There were overall 24 possible target loca-
zioni, eight of which (two in each quadrant) were used
for repeated displays with constant distractor layouts
throughout the experiment. Another eight target loca-
zioni (two in each quadrant) were used for nonrepeated
displays with random distractor arrangements. Another
set of eight additional target locations (two in each
quadrant) was used for repeated displays in the test
phase. The “T” target was rotated randomly by 90° either
to the left or the right. IL 11 remaining items were
L-shaped distractors rotated randomly at orthogonal
444
Journal of Cognitive Neuroscience
Volume 31, Numero 3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Example of repeated
and nonrepeated search layouts
in the learning and test phases
of the experiment. In repeated
displays, the target position
changed from learning to test,
whereas the distractor
arrangement remained
constant. In nonrepeated
displays, the target position was
constant (across learning and
test), but the distractor
arrangement varied from trial to
trial. See Methods for further
details.
orientations (0°, 90°, 180°, or 270°). Figura 1 presents
example display layouts.
Trial Sequence
A trial started with the presentation of a fixation cross
(0.10° × 0.10°, luminance = 1.0 cd/m2) for 500 msec,
followed by a blank interval of 200 msec, after which
the search display was presented. Observers were in-
structed to respond as quickly and accurately as possible
to the orientation of the target “T” (left vs. right). Each
search display stayed on the screen until a manual re-
sponse was elicited. If the “T” was rotated to the right
(left), observers responded by pressing the right (left)
arrow button on a computer keyboard with their right
(left) index finger. Following a response error, a red
minus sign appeared on screen for 1000 msec. Each trial
was followed by a blank intertrial interval of 1000 msec.
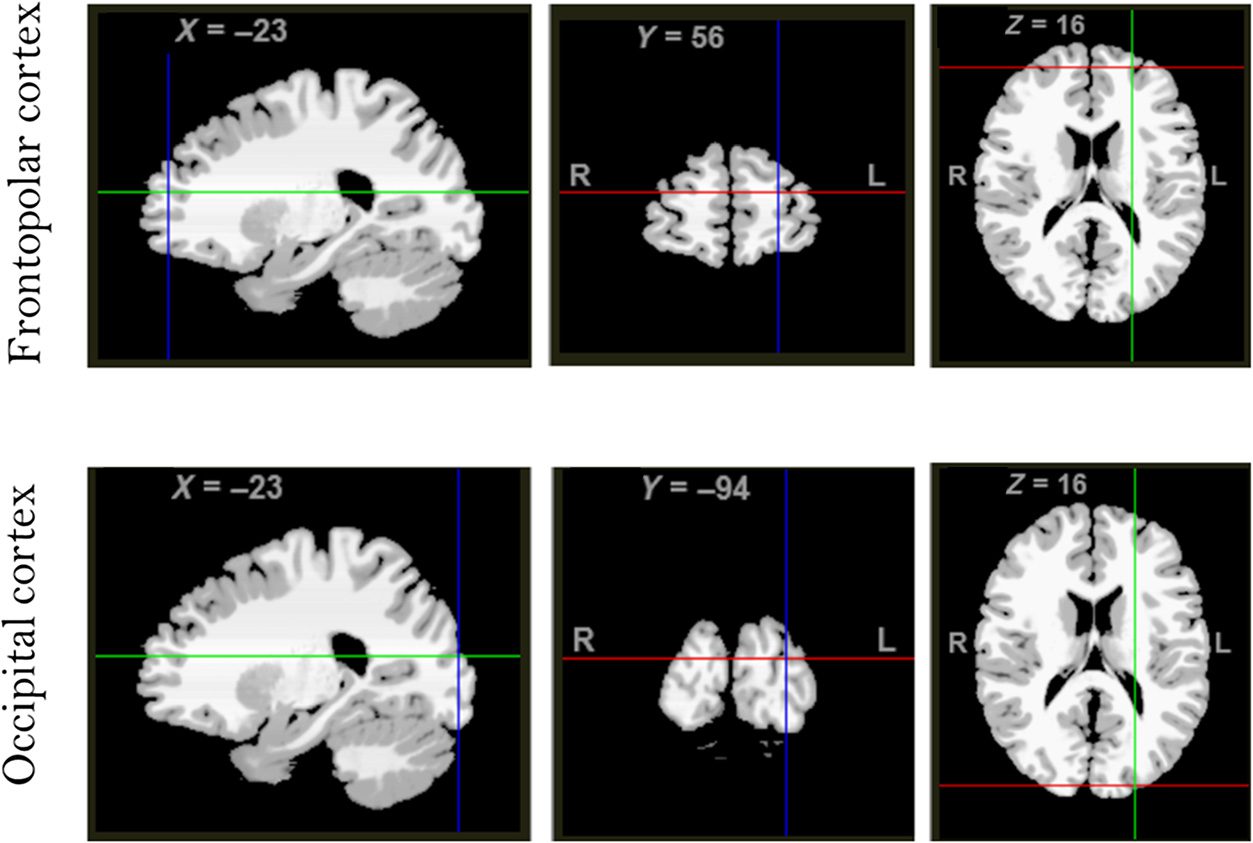
Neurological Institute (MNI) coordinates of the stimula-
tion sites based on observers’ 3T structural magnetic res-
onance images. We used MRI-guided TMS positioning
and placed the TMS coil on the locations of the ob-
servers’ scalp corresponding to the critical cortical MNI
coordinates reported by Pollmann and Manginelli
(2009). For the frontopolar TMS, the MNI coordinates
were x = −23, y = 56, z = 16, and for the occipital
TMS, the coordinates were x = −23, y = −94, z = 16
(Guarda la figura 2 for MNI coordinates overlaid on a reference
brain image from Lacadie, Fulbright, Rajeevan, Constable,
& Papademetris, 2008, using Yale BioImage Suite
Package). The control occipital region was selected by
preserving the x and z coordinates of the frontopolar
test region while only changing the y coordinate to target
occipital regions, thus ensuring that the laterality of
the stimulation region was perfectly matched across
the TMS stimulation conditions.
Transcranial Magnetic Stimulation
Design and Procedure
We applied rTMS (frequency = 1 Hz, intensity = 110%
active motor threshold, duration = 10 min, cioè., 600
pulses) using a figure-8 coil with an internal diameter
Di 7.0 cm (MagPro R30 machine with MC-B70 Butterfly
Coil, Magventure, Denmark). TMS was applied offline in
between the two experimental phases, either to the left
lateral FPC or to a control site over the occipital region.
The latter condition controls for the nonspecific clicking
sound and tactile sensation of TMS (Marzi et al., 1998).
Coil positioning was realized by neuronavigation via
frameless infrared stereotactic registration (Brainsight,
Rogue Research, Canada) to determine the Montreal
In all three groups (FPC rTMS, occipital rTMS, no rTMS),
the repeated display condition comprised eight layouts
associated with distinct target locations in the two phases
of the experiment. For each display, target–distractor
configurations were randomly generated at the beginning
of the experiment. These arrangements were repeatedly
presented on randomly selected trials throughout the
search task, with the restriction that each repeated dis-
play was shown only once per block. Displays in the non-
repeated condition were generated online at the start of
a given nonrepeated trial. In each group, a repeated
arrangement was presented on half of the trials, and a
Zinchenko et al.
445
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. MRI-guided TMS
positioning over frontopolar
and occipital stimulation sites
with corresponding MNI
coordinates.
nonrepeated arrangement was presented on the other
half. Trial order was randomized within each block. A
equate target location repetition effects between the
two types of displays, the target appeared equally
often at each of 24 possible locations throughout the
experiment: Eight locations were used for repeated
displays, eight were used for nonrepeated displays, E
the remaining eight locations were used for repeated
displays after the target location change in the test
phase. Inoltre, item density was controlled for
each display by presenting three search items in a
given quadrant (see Annac, Conci, Müller, & Geyer,
2017; Geyer et al., 2010). The orientation of the target
in a given repeated display was random in each trial,
whereas the orientations of the distractor elements
were held constant across trials (cf. Chun & Jiang,
1998). Figura 1 depicts an example search display for
the repeated and nonrepeated context conditions in
the learning and test phases. The experiment was
divided into two phases of 16 blocks each, yielding a
total of 256 trials in each phase. Participants had the
opportunity to take a short break between trial blocks
or continue directly with the next block. RTs were
collapsed across eight consecutive blocks to achieve
reliable estimates of contextual cueing performance
(cf. Chun & Jiang, 1998).
Participants in the no-rTMS group performed the
visual search task without any TMS stimulation and took
a 10-min break during the interval between the learning
and test phases. In the TMS groups, the task structure
was identical, except that participants received rTMS
(1 Hz, 600 pulses, 110% active motor threshold) during
the 10-min interval between the learning and test
phases. There was no significant difference in mean
stimulation intensity between the FPC rTMS and occip-
ital rTMS groups, T(27) = 1.18, p > 2 (mean intensity =
50.1% maximal stimulator output for frontopolar group
vs. 47.5% maximal stimulator output for the occipital
group). Before the start of the experiment, participants
performed one practice block of 24 trials (data not re-
corded). Observers were randomly assigned to each
rTMS group/condition.
Recognition Test
At the end of the experiment, observers performed a
yes–no recognition test, permitting us to assess whether
they had acquired any explicit memory of the repeated
configurations presented in the search task of the exper-
iment (a “standard” procedure in contextual cueing ex-
periments; cf. Chun & Jiang, 1998). A tal fine, eight
repeated displays from the search task and eight newly
composed displays were presented, and observers were
asked to indicate whether or not they had seen a given
display previously by pressing the left or the right mouse
button, rispettivamente (nonspeeded responses). The eight
repeated displays, along with the eight newly generated
displays, were presented in random order for four times
(cioè., in four separate blocks), yielding a total of 72 recog-
nition trials. Observers’ responses in the recognition task
were nonspeeded, and no error feedback was provided.
RESULTS
For the RT analyses, error trials and “extreme” RTs below
200 msec and above 4000 msec were excluded from the
dati. This outlier criterion led to the removal of <3% of
all trials. Mean contextual cueing effects (nonrepeated
RTs − repeated RTs) and the associated differences in
error rates were calculated and further submitted to a
2 × 2 mixed-design ANOVA with the within-group factors
Experimental phase (two levels: learning, test) and Session
446
Journal of Cognitive Neuroscience
Volume 31, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
c
n
_
a
_
0
1
3
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
half (two levels: first half of a given session (=8 blocks),
second half (=8 blocks). Greenhouse–Geisser corrected
values are reported in case Mauchley’s test of sphericity
was significant ( p < .05). In case of significant inter-
actions, Bonferroni-corrected pairwise t tests were used
for further comparisons. Data analyses were performed
with R version 3.4.3 (R Core Team, 2017).
Error Rates
We found no significant main effects or interactions
for observers’ error rates in all experimental groups
(all ps > .05). Overall, participants’ mean error rates
were very low (no-rTMS group = 2.4%, FPC rTMS
group = 1.3%, Occipital rTMS group = 1.9%).
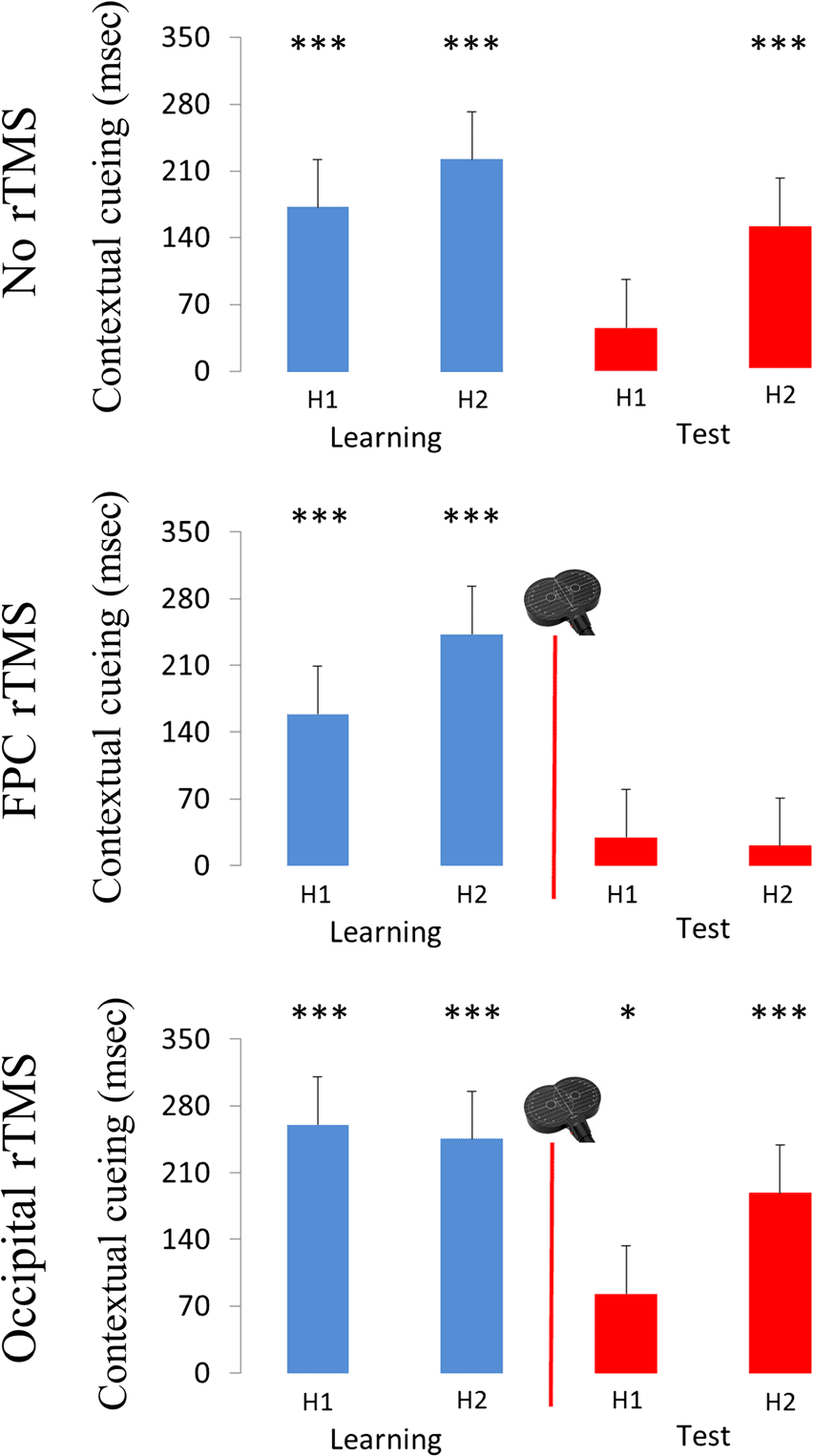
Figura 3. Mean contextual cueing effects in milliseconds [RT
nonrepeated − RT repeated] and associated mean standard errors
(SEM ) for Halves 1 (H1) E 2 (H2) in the learning and test phases,
separately for the no-rTMS, FPC rTMS, and occipital rTMS groups. IL
asterisks represent significance levels of *p < .05 and ***p < .001,
when testing contextual cueing effects against the zero baseline.
RTs
No-rTMS Group
For the no-rTMS group (see top part of Figure 3), we
(only) found a main effect of Experimental half: Across
the two experimental phases, contextual cueing was
overall smaller in the first half of whatever phase
compared with the second half (116 msec vs. 191 msec),
F(1, 13) = 8.21, p = .013, ηp
2 = .39. We also observed
a marginally significant interaction of Half × Phase,
F(1, 13) = 3.47, p = .085, ηp
2 = .21: Although the cue-
ing effect was comparable across the halves of Phase 1
(Half 1 = 188 msec vs. Half 2 = 229 msec), F(1, 13) =
1.98, p = .182, ηp
2 = .13, it was smaller in the first
half relative to the second half of Phase 2 (46 msec
vs. 152 msec), F(1, 13) = 7.25, p = .018, ηp
2 = .36. Note
that the cueing effect was reliable (i.e., different from
zero) in all halves of both the learning and test phases
(all ps < .05, ts > 4), except for Half 1 of Phase 2, T(27) =
1.11, p > .28 (see also Figure 3). Prossimo, we compared the
contextual cueing effects in Half 2 of the learning phase
(cioè., when the effect was firmly established immediately
before target location change) to the cueing effects in
Halves 1 E 2 of the test phase (thus providing measures
of contextual cueing in relatively early vs. relatively late
stages after the change and after some exposure to the
relocated target). These comparisons revealed contextual
cueing to be reduced in the first half of the test phase
relative to the second half of learning phase (46 msec
vs. 229 msec), T(27) = 2.82, P < .01, and then to recover
again in the second half of the test phase (nonsignificant
difference of 152 msec vs. 229 msec), t(27) = 1.35, p =
.188. This pattern of results confirms previous findings
that that cueing is compromised after target location
changes but then recovers to some extent with extended
experience on the relocated displays (Zellin et al., 2014).
FPC rTMS Group
In the FPC rTMS group (see middle part of Figure 3), the
main effect of phase was significant: The cueing effect
was overall larger in the learning phase (200 msec) than
in the test phase (25 msec), F(1, 13) = 13.23, p = .003,
ηp
2 = .5. In addition, we found an interaction of phase
and half, F(1, 13) = 6.6, p = .023, ηp
2 = .34. Follow-up
analyses for each of the two phases revealed that the
main effect of half was significant in the learning phase,
F(1, 13) = 9.76, p = .008, ηp
2 = .43, but not in the test
phase after TMS, F(1, 13) = 0.08, p = .779, ηp
2 = .01. In
the learning phase, the cueing effect was smaller in the
first compared with the second half: 158 msec versus
242 msec, t(54) = −2.08, p < .05. In the test phase,
by contrast, the cueing effect was markedly reduced
and comparable between Halves 1 and 2: 29 vs. 21 msec
( p > .7; neither of these effects was significantly different
from zero: both ts < 1, ps > .4). Finalmente, the contextual
cueing effect in Half 2 of the learning phase was larger
Zinchenko et al.
447
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
than in Halves 1 E 2 of the test phase (242 msec vs.
29 msec, T(27) = 4.76, P < .001; 242 msec vs. 21 msec,
t(27) = 4.52, p < .001). This pattern indicates that TMS
over the FPC effectively hinders the adaptation of con-
textual cueing to a changed target location within a
repeated distractor layout.
Occipital rTMS Group
With rTMS of an occipital (control) region (see bottom
part of Figure 3), the main effect of phase was again sig-
nificant: The cueing effect was larger in the learning
phase (252 msec) relative to the test phase (135 msec),
F(1, 13) = 6.58, p = .023, ηp
2 = .34). The interaction of
Phase × Half was also significant, F(1, 13) = 17.61, p =
.001, ηp
2 = .58. Follow-up analyses for the two phases
revealed that although cueing was comparable between
Halves 1 and 2 of the learning phase, F(1, 13) = 0.2, p =
.662, ηp
2 = .02 (260 vs. 244 msec), the effect was smaller
in Half 1 relative to Half 2 of the test phase, F(1, 13) =
7.99, p = .014, ηp
2 = .38 (83 vs. 188 msec). Of note, the
cueing effect was significant in Halves 1 and 2 both in
the learning and test phases (all ps < .05, ts > 2.5).
Finalmente, we compared the contextual cueing effects in
Halves 1 E 2 of the test phase with the firmly established
effect in Half 2 of the preceding learning phase: Contextual
cueing was significantly reduced in the first half of the
test phase (82 msec vs. 244 msec), T(27) = 3.04, P < .01,
but then exhibited a reasonable (albeit nonperfect) adap-
tation to the changed target locations—in the second
half (188 msec vs. 244 msec), t(27) = 1.18, p > .2. Così,
the results of the occipital rTMS group mirror those of
the “baseline” no-rTMS group (in which participants
had not received any intervention at all).
Omnibus Analysis
To further test and directly compare the effect patterns
among the three groups, we entered the mean contex-
tual cueing effects into a 3 × 2 × 2 mixed-design
ANOVA with the between-subject factor rTMS group
(three levels: no rTMS, FPC rTMS, occipital rTMS) E
the within-subject factors Phase (two levels: apprendimento,
test) and Half (two levels: first half, second half ). Questo
ANOVA revealed a significant main effect of Phase:
cueing was overall reduced after the target location
changes in the test phase relative to initial learning phase
(87 msec vs. 216), F(1, 39) = 20.16, P < .001, ηp
2 = .34.
The main effect of Half was also significant: Cueing was
overall more marked in the second half relative to the
first half of whatever phase (179 msec vs. 124 msec), F(1,
39) = 11.29, p = .002, ηp
2 = .22, of each session.
Furthermore and most importantly, the three-way inter-
action was significant, F(2, 39) = 10.02, p < .001, ηp
2 =
.34. As can be seen from Figure 3, contextual cueing was
overall comparable among the three groups in the initial
learning phase, and it was substantially reduced immedi-
ately after the introduction of the target location change
in the test phase (compare Half 2 of the learning phase
with Half 1 of the test phase) for all groups. The signifi-
cant three-way interaction then arises from the fact that a
recovery of contextual cueing, indicative of successful
adaptation to the target location change, in Half 2 of
the test phase was evident only for the no-rTMS (“base-
line”) and occipital rTMS (“control”) groups, whereas
there was no indication of a recovery—or relearning of
the changed target location—whatsoever
in the FPC
rTMS group.1 This result was supported by an additional
series of correlation analyses that tested whether the
strength of contextual cueing before relocation (in the
learning phase) was related to the degree of adaptation
(or the lack of it) after the target location change in the
test phase. None of these correlations were significant
(no-rTMS group: r = −.01, t(54) = −0.09, p > .9; FPC
rTMS group: r = .19, T(54) = 1.44, p > 1.5; occipital
rTMS group: r = .06, T(54) = 0.46, p > .6), indicating that
variations in contextual cueing that emerged in the test
phase are causally related to the specific TMS condition
(rather than the magnitude of cueing in initial learning).
Recognition Test Performance
Explicit recognition performance was analyzed by com-
paring observers’ hit rates (repeated display correctly
judged as repeated in the recognition test) with their cor-
responding false alarm rates (nonrepeated display incor-
rectly judged as repeated) by means of a 2 (Tipo di
risposta: hit, false alarm; within-subject variable) × 3
(Group: no rTMS, FPC rTMS, occipital rTMS; between-
subject variable) mixed-design ANOVA. On average, ob-
servers correctly recognized repeated displays in 51% Di
the trials (hit rate), whereas they falsely classified non-
repeated displays as repeated in 48% of trials (false alarm
rate). This hit rate was not significantly different from the
false alarm rate, F(1, 39) = 1.75, p > .19, ηp
2 = .043, E
there was also no difference across the three groups
(interaction of Group × Response Type: F(2, 39) =
2.5, p > .09, ηp
2 = .114). The main effect of Group
was also not significant, F(2, 39) = 1.45, p > .2, ηp
2 =
.067. This suggests that observers could not explicitly
recognize repeated display arrangements.
DISCUSSION
Objects in our visual environment typically appear in a
certain context that remains invariant across repeated en-
counters (per esempio., a mail box in the front yard; see Brady,
Konkle, Oliva, & Alvarez, 2009; Palmer, 1975). Our visual
system has the capability to extract and learn such
context–object associations, which in turn facilitate pro-
cessing of task critical target objects. In recent years,
numerous studies have shown that visual search can
448
Journal of Cognitive Neuroscience
Volume 31, Numero 3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
substantially benefit from learned distractor-to-target spa-
tial relations (see Goujon, Didierjean, & Thorpe, 2015;
Wolfe & Horowitz, 2017, for reviews). Tuttavia, statistical
context learning should also exhibit a degree of flexibility
to incorporate changes that may occur in our sensory en-
vironment. Specifically, context memory should be able
to adapt to target position changes, ensuring that atten-
tion is effectively allocated to targets at their novel loca-
zioni, rather than prioritizing no longer relevant
positions in the environment. In a recent fMRI investiga-
zione, Pollmann and Manginelli (2009) suggested that FPC
may play an important role in this type of context adapta-
zione. Specifically, they suggested that FPC detects discrep-
ancies between the current sensory input and associated
context-based expectations and then informs other
(frontoparietal) attention structures about these incon-
sistencies, which could eventually lead to a shift of
visuospatial attention to the changed target position
(“FPC+” hypothesis). Although such a facilitatory FPC
influence may eventually lead to context adaptation, UN
number of studies have reported rather inefficient,
slow, and resource-demanding adaptation to changed
distractor-to-target spatial relations (Annac et al., 2017;
Zellin et al., 2013, 2014). Based on these findings, Questo
study was designed to test an alternative account of FPC
functions in context adaptation, namely, that FPC contrib-
utes to the regulation of prediction errors by “explaining
away,” that is, suppressing the changed target position
and thus effectively supporting already acquired contex-
tual memory representations (FPC− hypothesis). Noi
hypothesized that if FPC facilitates the processing of
changed target locations (FPC+), then interference with
FPC by means of TMS would weaken context adaptation
even further, as there would be no guidance from FPC on
the reallocation of attention after target position changes.
Alternatively, if FPC supports attention guidance by
previously established context memories (FPC−), Poi
we would expect to find improved adaptation, because
attentional (mis)guidance would be reduced during to
the inactivation of this region. The results clearly
support the “FPC+” hypothesis. Stimulation of FPC in-
between a learning phase and a test phase did not yield
any context adaptation. This is in contrast with the no-
TMS and occipital rTMS conditions, in which there was
a relatively high degree of adaptation, approaching
(Anche se, within the time on task allowed, not quite
reaching) the level of the initial learning effect.
Our findings provide important novel insights into the
role of the FPC: They affirm a causal involvement of the
left FPC in the adaptation of learned target–distractor
arrays and thus in statistical context (Rif)learning in visual
search. Pollmann and Manginelli (2009) used fMRI to
show that neural activity in the left FPC increases selec-
tively after target location changes in repeated displays.
They hypothesized that these activations arise because
FPC signals a perceptual change to frontoparietal brain
areas, providing the basis for these attention-guiding
networks to adapt to the change and to update context
memory. The current study provides a causal test of this
hypothesis. Participants were unable to adapt their con-
text memory, even after hundreds of repetitions, Quando
normal FPC functioning was disrupted, questo è, fronto-
polar TMS hinders context adaptation.
Other studies lend further support to his account of
FPC function. For instance, Badre and Wagner (2005)
studied the role of FPC in proactive interference, questo è,
when a memory relating to a past experience hinders
the subsequent acquisition of new memories. Par-
ticipants in their study were asked to maintain a set of
target words and indicate whether a subsequently pre-
sented “probe” word was from the target set. Badre
and Wagner found that probe words not included in
the current target set but included in the target set on
the preceding trial resulted in impaired performance
and in increased activation in the left FPC—suggesting
that the FPC is involved in the resolution of proactive
interference when information that was previously held
in memory interferes with performance on the current
trial. In a different study, Badre and D’Esposito (2007)
further examined the involvement of the left FPC in a
varied/competing mapping task. Participants’ task was
to compare two test items in terms of one of four dimen-
sions (texture, orientation, shape, or size), questo è, Essi
had to make a decision whether the two items were
the same or different in terms of the relevant dimen-
sion—which was cued on each trial by the color of a
square frame around the test items (at the start of a trial).
Crucially, Badre and D’Esposito (2007) varied the map-
ping between a given colored square and a given dimen-
sion. In the consistent condition, a given context color
did reliably indicate a single response dimension across
all trials/blocks. In the inconsistent condition, by contrast,
the assignment of a given context color to a response di-
mension was variable across blocks (per esempio., in some blocks,
a green frame meant that observers had to compare the
two test items in terms of their shape [while ignoring
their other properties], whereas in other blocks it meant
they had to compare the texture of the test items), così
engendering some competition between the color-to-
dimension mappings. Badre and D’Esposito (2007)
found increased activation of the left FPC in the inconsis-
tent condition, when a color cue was predictive of multi-
ple target dimensions on different blocks. This indicates
that the FPC was involved in the resolution or reorgani-
zation of cue–response associations in the inconsistent
mapping task. The current data are in line with such an
interpretation: The left anterior FPC might also be
involved in the detection of changed target–distractor
spatial associations and their corresponding reorganiza-
zione, questo è, adaptation, in a visual search statistical
learning task.
The present findings may also indicate that the FPC is
responsible for the acquisition of rules and environmen-
tal regularities irrespective of previously established
Zinchenko et al.
449
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
C
N
_
UN
_
0
1
3
5
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
ricordi. For instance, previous observations showed
that rTMS over the left (Costa et al., 2011) and right
(Costa et al., 2013) FPCs led to impaired visual–spatial
prospective memories. Because contextual cueing re-
quires the detection of repeated target–distractor arrays,
the FPC may be involved in the detection and sub-
sequent acquisition of such associations. In line with this
idea, Strange, Henson, Friston, and Dolan (2001) used
fMRI to show that the bilateral FPC mediates the learn-
ing of rules: They observed increased activation in
the bilateral FPC when a new rule had to be learned,
and this activation decreased with improving task
performance, possibly representing rule acquisition.
Recentemente, Paniukov and Davis (2018) showed that the
FPC continues to integrate information about rules of
a given task even after the correct rule was extracted.
This indicates that FPC is involved in a constant monitor-
ing of evidence to support rule-based representations.
Seger and Cincotta (2006) and Liu, Braunlich, Wehe,
and Seger (2015) found that the FPC was more active
during rule learning compared with rule application in
variants of a classification task. Based on this, it was sug-
gested that FPC may be involved in the acquisition and
maintenance of rule-based learning. Crucially, this ac-
count may be in line with the findings of Pollmann and
Manginelli (2009) that FPC activations were lower for
repeated relative to nonrepeated arrays during initial
context learning and higher after target relocation.
Specifically, once a given spatial regularity is learned,
activity in the FPC is reduced (though not completely
gone; see Paniukov & Davis, 2018), but it is increased
again after target is relocated and new associations
have to be acquired (Strange et al., 2001).
Così, FPC appears to be involved not only in the re-
learning after a target location change but also in initial
learning of target–distractor constellations, before any
target location changes. For instance, stimulation of the
FPC may interfere with the efficient use of contextual
cues, as disruption of the FPC may result in an inability
or difficulty to acquire spatial target–distractor associa-
zioni. This prediction receives some support from the
findings of Nydam, Sewell, and Dux (2018), who applied
cathodal transcranial direct current stimulation over the
left pFC while participants performed a standard con-
textual cueing task—which resulted in a reduction of
contextual cueing during the first half of experiment.
Similar results have been obtained with other perceptual
decision tasks, including visual search (per esempio., Raja
Beharelle, Polanía, Hare, & Ruff, 2015; Reinhart &
Woodman, 2015). È interessante notare, using anodal transcranial
direct current stimulation over the FPC, these studies
found an increased proportion of responses toward
novel, questo è, unlearned, stimuli (Raja Beharelle et al.,
2015) or improved learning of the target’s perceptual
attributes in the context of a visual search task (Reinhart
& Woodman, 2015). Overall, this work points to a
functional role of FPC in setting up context-based
memories that, once established, require maintaining
the balance between acquired memories and the cur-
rently available stimulus information. Assuming the
ubiquity of statistical learning, this would be equivalent
to an adaptation of existing context memories with re-
gard to the new stimulus characteristics. In this view,
FPC would modulate both initial context learning and
later context adaptation.
Conclusione
Several previous visual search studies (Pollmann et al.,
2006, 2007; Weidner et al., 2002; Pollmann, Weidner,
Müller, & von Cramon, 2000) have shown that the left
FPC shows transient signal increases when the target’s
defining perceptual dimension changes across successive
trials. These activation increments were interpreted as
enhanced requirements for attention to shift to the target
dimension, thus requiring an up-modulation of salience
signals, which represent the conspicuity of the target in
relation to the surrounding distractor items. Our results
extend these previous findings by showing that the FPC
is not only sensitive to the repetition of certain target
dimensions (colore, orientation, eccetera.) but also monitors
the spatial array of learned, invariant search layouts
within which the target is embedded. Although we have
not measured visuospatial attention directly, the present
finding would generally support the view that the detec-
tion of changes in previously learned search layouts leads
to an FPC biasing signal to guide attention more effi-
ciently toward the changed target location in an attempt
to overcome the detrimental effects resulting from pre-
viously learned context representations.
Ringraziamenti
This research was supported by a project grant from the German
research foundation (Deutsche Forschungsgemeinschaft,
DFG, grant GE 1889/4-1 to T. G. and M. C. and TA 857/3-1 A
P. T.) and BMBF grant 801210010-20.
Reprint requests should be sent to Artyom Zinchenko, Depart-
ment Psychologie, Lehrstuhl für Allgemeine und Experimentelle
Psychologie, Ludwig-Maximilians-Universität München,
Leopoldstraße 13 80802 München, Germany, or via e-mail:
artyom.zinchenko@psy.lmu.de.
Note
1. Note that three further comparisons, questo è, Experiment ×
Phase × Half mixed-design ANOVAs, comparing each experi-
mental condition with each of the other conditions, revealed
a significant three-way interaction of experiment, phase, E
half in the analysis of the FPC rTMS and occipital rTMS groups,
F(1, 26) = 21.4, P < .001, ηp
2 = .45, as well as the FPC rTMS and
no-rTMS groups, F(1, 26) = 9.86, p = .004, ηp
2 = .27). By
contrast, the three-way interaction was nonsignificant in the
occipital rTMS and no-rTMS groups, F(1, 26) = 1.49, p = .233,
ηp
2 = .05.
450
Journal of Cognitive Neuroscience
Volume 31, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
c
n
_
a
_
0
1
3
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Annac, E., Conci, M., Müller, H. J., & Geyer, T. (2017). Local
item density modulates adaptation of learned contextual
cues. Visual Cognition, 25, 262–277.
Annac, E., Manginelli, A. A., Pollmann, S., Shi, Z., Müller, H. J., &
Geyer, T. (2013). Memory under pressure: Secondary-task effects
on contextual cueing of visual search. Journal of Vision, 13, 6.
Badre, D., & D’Esposito, M. (2007). Functional magnetic
resonance imaging evidence for a hierarchical organization of
the prefrontal cortex. Journal of Cognitive Neuroscience, 19,
2082–2099.
Badre, D., & Wagner, A. D. (2005). Frontal lobe mechanisms
that resolve proactive interference. Cerebral Cortex, 15,
2003–2012.
Brady, T. F., Konkle, T., Oliva, A., & Alvarez, G. A. (2009).
Detecting changes in real-world objects: The relationship
between visual long-term memory and change blindness.
Communicative & Integrative Biology, 2, 1–3.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436.
Chun, M. M., & Jiang, Y. V. (1998). Contextual cueing: Implicit
learning and memory of visual context guides spatial
attention. Cognitive Psychology, 36, 28–71.
Conci, M., & Müller, H. J. (2012). Contextual learning of
multiple target locations in visual search. Visual Cognition,
20, 746–770.
Conci, M., Sun, L., & Müller, H. J. (2011). Contextual remapping
in visual search after predictable target-location changes.
Psychological Research, 75, 279–289.
Conci, M., Zellin, M., & Müller, H. J. (2012). Whatever after next?
Adaptive predictions based on short- and long-term memory
in visual search. Frontiers in Psychology, 3, 409.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
Costa, A., Oliveri, M., Barban, F., Bonnì, S., Koch, G., Caltagirone,
C., et al. (2013). The right frontopolar cortex is involved in
visual-spatial prospective memory. PLoS One, 8, e56039.
Costa, A., Oliveri, M., Barban, F., Torriero, S., Salerno, S.,
Lo Gerfo, E., et al. (2011). Keeping memory for intentions:
A cTBS investigation of the frontopolar cortex. Cerebral
Cortex, 21, 2696–2703.
Filipović, S. R., Rothwell, J. C., & Bhatia, K. (2010). Slow (1 Hz)
repetitive transcranial magnetic stimulation (rTMS) induces
a sustained change in cortical excitability in patients with
Parkinson’s disease. Clinical Neurophysiology, 121, 1129–1137.
Friston, K. (2010). The free-energy principle: A unified brain
theory? Nature Reviews Neuroscience, 11, 127–138.
Geyer, T., Zehetleitner, M., & Müller, H. J. (2010). Contextual
cueing of pop-out visual search: When context guides the
deployment of attention. Journal of Vision, 10, 20.
Goujon, A., Didierjean, A., & Thorpe, S. (2015). Investigating
implicit statistical learning mechanisms through contextual
cueing. Trends in Cognitive Sciences, 19, 524–533.
Hoffman, R. E., & Cavus, I. (2002). Slow transcranial magnetic
stimulation, long-term depotentiation, and brain
hyperexcitability disorders. American Journal of Psychiatry,
159, 1093–1102.
Jiang, Y. V., & Leung, A. W. (2005). Implicit learning of
ignored visual context. Psychonomic Bulletin & Review,
12, 100–106.
Jungé, J. A., Scholl, B. J., & Chun, M. M. (2007). How is spatial
context learning integrated over signal versus noise? A
primacy effect in contextual cueing. Visual Cognition, 15,
1–11.
Kok, P., Rahnev, D., Jehee, J. F. M., Lau, H. C., & de Lange,
F. P. (2012). Attention reverses the effect of prediction
in silencing sensory signals. Cerebral Cortex, 22,
2197–2206.
Lacadie, C. M., Fulbright, R. K., Rajeevan, N., Constable, R. T.,
& Papademetris, X. (2008). More accurate Talairach
coordinates for neuroimaging using non-linear registration.
Neuroimage, 42, 717–725.
Liu, Z., Braunlich, K., Wehe, H. S., & Seger, C. A. (2015). Neural
networks supporting switching, hypothesis testing, and rule
application. Neuropsychologia, 77, 19–34.
Makovski, T., & Jiang, Y. V. (2010). Contextual cost: When a
visual-search target is not where it should be. Quarterly
Journal of Experimental Psychology, 63, 216–225.
Manginelli, A. A., & Pollmann, S. (2009). Misleading contextual
cues: How do they affect visual search? Psychological
Research, 73, 212–221.
Marzi, C. A., Miniussi, C., Maravita, A., Bertolasi, L., Zanette, G.,
Rothwell, J. C., et al. (1998). Transcranial magnetic
stimulation selectively impairs interhemispheric transfer of
visuo-motor information in humans. Experimental Brain
Research, 118, 435–438.
Nydam, A. S., Sewell, D. K., & Dux, P. E. (2018). Cathodal
electrical stimulation of frontoparietal cortex disrupts
statistical learning of visual configural information. Cortex,
99, 187–199.
Palmer, S. E. (1975). The effects of contextual scenes on the
identification of objects. Memory & Cognition, 3, 519–526.
Paniukov, D., & Davis, T. (2018). The evaluative role of
rostrolateral prefrontal cortex in rule-based category learning.
Neuroimage, 166, 19–31.
Pelli, D. G. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442.
Pollmann, S., Mahn, K., Reimann, B., Weidner, R., Tittgemeyer,
M., Preul, C., et al. (2007). Selective visual dimension
weighting deficit after left lateral frontopolar lesions. Journal
of Cognitive Neuroscience, 19, 365–375.
Pollmann, S., & Manginelli, A. A. (2009). Early implicit
contextual change detection in anterior prefrontal cortex.
Brain Research, 1263, 87–92.
Pollmann, S., Weidner, R., Müller, H. J., & von Cramon, D. Y.
(2000). A fronto-posterior network involved in visual
dimension changes. Journal of Cognitive Neuroscience, 12,
480–494.
Pollmann, S., Weidner, R., Müller, H. J., & von Cramon, D. Y.
(2006). Neural correlates of visual dimension weighting.
Visual Cognition, 14, 877–897.
R Core Team. (2017). R: A language and environment for
statistical computing. Vienna: R Foundation for Statistical
Computing. https://www.R-project.org/.
Raja Beharelle, A., Polanía, R., Hare, T. A., & Ruff, C. C. (2015).
Transcranial stimulation over frontopolar cortex elucidates
the choice attributes and neural mechanisms used to
resolve exploration–exploitation trade-offs. Journal of
Neuroscience, 35, 14544–14556.
Rao, R. P. N., & Ballard, D. H. (1999). Predictive coding in the
visual cortex: A functional interpretation of some extra-
classical receptive-field effects. Nature Neuroscience, 2,
79–87.
Reinhart, R. M. G., & Woodman, G. F. (2015). Enhancing
long-term memory with stimulation tunes visual attention
in one trial. Proceedings of the National Academy of
Sciences, U.S.A., 112, 625–630.
Schankin, A., & Schubö, A. (2009). Cognitive processes
facilitated by contextual cueing: Evidence from event-related
brain potentials. Psychophysiology, 46, 668–679.
Seger, C. A., & Cincotta, C. M. (2006). Dynamics of frontal,
striatal, and hippocampal systems during rule learning.
Cerebral Cortex, 16, 1546–1555.
Zinchenko et al.
451
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
c
n
_
a
_
0
1
3
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Strange, B. A., Henson, R. N. A., Friston, K. J., & Dolan, R. J.
(2001). Anterior prefontal cortex mediates rule learning in
humans. Cerebral Cortex, 11, 1040–1046.
Summerfield, C., Trittschuh, E. H., Monti, J. M., Mesulam, M.-M.,
& Egner, T. (2008). Neural repetition suppression reflects
fulfilled perceptual expectations. Nature Neuroscience, 11,
1004–1006.
Wassermann, E. M., & Lisanby, S. H. (2001). Therapeutic
application of repetitive transcranial magnetic stimulation:
A review. Clinical Neurophysiology, 112, 1367–1377.
Weidner, R., Pollmann, S., Müller, H. J., & von Cramon, D. Y.
(2002). Top–down controlled visual dimension weighting:
An event-related fMRI study. Cerebral Cortex, 12, 318–328.
Wolfe, J. M., & Horowitz, T. S. (2017). Five factors that
guide attention in visual search. Nature Human
Behaviour, 1, 0058.
Zellin, M., Conci, M., von Mühlenen, A., & Müller, H. J. (2011).
Two (or three) is one too many: Testing the flexibility of
contextual cueing with multiple target locations. Attention,
Perception, & Psychophysics, 73, 2065–2076.
Zellin, M., Conci, M., von Mühlenen, A., & Müller, H. J.
(2013). Here today, gone tomorrow—Adaptation to
change in memory-guided visual search. PLoS One, 8,
e59466.
Zellin, M., von Mühlenen, A., Müller, H. J., & Conci, M.
(2014). Long-term adaptation to change in implicit
contextual learning. Psychonomic Bulletin & Review,
21, 1073–1079.
Zinchenko, A., Conci, M., Müller, H. J., & Geyer, T. (2018).
Predictive visual search: Role of environmental regularities
in the learning of context cues. Attention, Perception, &
Psychophysics, 80, 1096–1109.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
4
2
1
7
8
8
3
5
6
/
j
o
c
n
_
a
_
0
1
3
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
452
Journal of Cognitive Neuroscience
Volume 31, Number 3