Stimulus Onset Asynchrony Affects Weighting-related
Event-related Spectral Power in Self-motion Perception
Ben Townsend , Joey K. Legere, Martin v. Mohrenschildt, and Judith M. Shedden
Astratto
■ Self-motion perception relies primarily on the integration of
the visual, vestibular, proprioceptive, and somatosensory sys-
tems. There is a gap in understanding how a temporal lag
between visual and vestibular motion cues affects visual–
vestibular weighting during self-motion perception. The beta
band is an index of visual–vestibular weighting, in that robust
beta event-related synchronization (ERS) is associated with
visual weighting bias, and robust beta event-related desynchro-
nization is associated with vestibular weighting bias. The present
study examined modulation of event-related spectral power dur-
ing a heading judgment task in which participants attended to
either visual (optic flow) or physical (inertial cues stimulating
the vestibular, proprioceptive and somatosensory systems)
motion cues from a motion simulator mounted on a MOOG
Stewart Platform. The temporal lag between the onset of visual
and physical motion cues was manipulated to produce three lag
conditions: simultaneous onset, visual before physical motion
onset, and physical before visual motion onset. There were
two main findings. Primo, we demonstrated that when the
attended motion cue was presented before an ignored cue,
the power of beta associated with the attended modality was
greater than when visual–vestibular cues were presented simul-
taneously or when the ignored cue was presented first. Questo era
the case for beta ERS when the visual-motion cue was attended
A, and beta event-related desynchronization when the physical-
motion cue was attended to. Secondo, we tested whether the
power of feature-binding gamma ERS (demonstrated in audio-
visual and visual–tactile integration studies) increased when
the visual–vestibular cues were presented simultaneously versus
with temporal asynchrony. We did not observe an increase in
gamma ERS when cues were presented simultaneously, suggesting
that electrophysiological markers of visual–vestibular binding
differ from markers of audiovisual and visual–tactile integration.
All event-related spectral power reported in this study were
generated from dipoles projecting from the left and right motor
areas, based on the results of Measure Projection Analysis. ■
INTRODUCTION
The visual, vestibular, proprioceptive, and somatosensory
systems collect information about how an organism moves
through its environment, and integrate this information in
associated brain areas, such as medial superior temporal
area and ventral intraparietal area (for a review, Vedere
DeAngelis & Angelaki, 2012), to produce a smooth, unified
perception of self-motion. One complicating factor in this
integration process is that each of these cues to motion is
perceived on different timelines. Per esempio, self-motion
information from the visual system is perceived faster than
self-motion information from the vestibular system (per esempio.,
RTs are ∼220 msec for light and ∼440 msec for galvanic
vestibular stimulation; Barnett-Cowan & Harris, 2009);
Tuttavia, our perception of self-motion is a function of
multisensory integration. Understanding how the tempo-
ral factors of visual and vestibular perception affect multi-
sensory integration has been of interest to researchers in
many fields of science and engineering. Per esempio,
understanding this construct has been a major focus for
McMaster University, Hamilton, Ontario, Canada
transfer of training research and for setting policies by
flight training administration authorities.
Given the different temporal trajectories of information
processing between sensory systems, the temporal inte-
gration of multisensory stimuli has long been of interest
to researchers. Per esempio, in audiovisual integration,
direction-incongruent stimuli give rise to the ventriloquist
effect, in which the two stimuli are perceived as having the
same source despite a spatially separated origin (Alais &
Burr, 2004). This effect disappears when the synchrony
of the audiovisual stimuli exceeds ∼300 msec (Slutsky &
Recanzone, 2001). We still do not fully understand the
potential effect of temporal asynchrony on visual–
vestibular integration and self-motion perception, espe-
cially in the context of driving and flight motion-simulator
research. Tuttavia, a recent study demonstrated that
changes in the velocity of a visual or physical self-motion
cue are most quickly detected when the stimuli are
aligned, compared with a 100-msec timing difference
(Kenney et al., 2020). Inoltre, Rodriguez and Crane
(2021) demonstrated that visual-inertial (per esempio., visual–
vestibular) heading perception is also sensitive to tempo-
ral misalignments of less than 250 msec between the
motion cues.
© 2023 Istituto di Tecnologia del Massachussetts. Published under a
Creative Commons Attribution 4.0 Internazionale (CC BY 4.0) licenza.
Journal of Cognitive Neuroscience 35:7, pag. 1092–1107
https://doi.org/10.1162/jocn_a_01994
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
C
N
_
UN
_
0
1
9
9
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multisensory integration is also affected by attention
allocation (Macaluso et al., 2016). Attention can be volun-
tarily allocated toward a stimulus, a sensory modality, or a
specific region of space to achieve task goals (Li, Piëch, &
Gilbert, 2004). Tuttavia, processing can also be involun-
tarily captured by sensory events, even when the attention
capturing signals are unrelated to the current goal-
directed activity (Öhman, Flykt, & Esteves, 2001). EEG is
a useful tool to explore the online processes related to the
interaction between attention and multisensory integra-
zione. The high temporal resolution of EEG has been effec-
tive in testing hypotheses related to synchronization of
neural oscillations as a mechanism for the integration of
information across sensory modalities (Senkowski,
Schneider, Foxe, & Engel, 2008). Synchronization of neu-
ral oscillations (event-related spectral power [ERSP]) È
quantified by measuring power of event-related synchro-
nizations (ERSs) and desynchronizations (ERDs) within
particular frequency bands (per esempio., theta, alpha, beta,
gamma). One hypothesis about interpretation of neural
oscillations is that distinct spectral timelines index differ-
ent local cortical networks involved in sensory processing,
attention allocation, and multisensory integration (Siegel,
Donner, & Engel, 2012). Most studies that support the
spectral timelines hypothesis are based on audiovisual
or visuotactile integration (for a review, see Keil &
Senkowski, 2018). Per esempio, Senkowski, Talsma,
Grigutsch, Herrmann, and Woldorff (2007) showed that
the closer in time the audiovisual stimuli were presented
together, the more feature binding-related gamma ERS
was elicited early after stimulus onset. This finding also
supports Singer and Gray’s (1995) temporal correlation
hypothesis, which suggests that oscillations within the
gamma band facilitate integration across sensory modali-
ties. As far as we know, there are few published studies
exploring how the onset timing of multisensory stimuli
affects EEG correlates of visual–vestibular integration.
Townsend, Legere, O’Malley, von Mohrenschildt, E
Shedden (2019) used a high-fidelity motion simulator
and a high-density EEG array to observe ERSP in response
to simultaneous-onset visual- and physical-motion stimuli.
To examine the effect of attention allocation to visual ver-
sus physical motion, in a blocked design, participants
made heading judgments to visual (or physical) cues only,
while ignoring the other modality. For each trial, headings
of the motion cues were either spatially congruent (per esempio.,
heading was the same for visual and physical) or incongru-
ent (per esempio., visual and physical headings differed). Impor-
tantly, in all conditions, the visual and physical cues to
self-motion were presented simultaneously. Measure Pro-
jection Analysis (MPA) identified cortical regions in the
premotor and sensory motor areas (Brodmann’s areas
[BAs] 6 E 4) associated with motor processing. ERSP
analysis within these areas revealed sensitivity of theta-
(4–7 Hz), alpha- (8–12 Hz), and beta- (13–30 Hz) band
oscillations to attended visual versus physical self-motion
stimuli. Specifically, attending to the visual-motion
stimulus (while ignoring the physical-motion stimulus)
evoked earlier theta ERS and alpha ERD, whereas attention
to the physical-motion stimulus (while ignoring the visual-
motion stimulus) evoked longer-lasting and more power-
ful beta ERD. Complimentary research suggests that theta
ERS is an index of heading processing (Townsend, Legere,
von Mohrenschildt, & Shedden, 2022; for a review, Vedere
Buzsáki & Moser, 2013), and alpha ERD/ERS is associated
with focal attention and cognitive load (for a review, Vedere
Klimesch, 2012). Most important for the present article,
previous research has indicated that beta ERD/ ERS
indexed visual–vestibular weighting (Townsend et al.,
2019, 2022). Per esempio, when attention was focused
on the visual-motion stimulus (while ignoring physical-
motion cues), beta ERS was stronger, whereas when atten-
tion was focused on the physical-motion stimulus (while
ignoring visual-motion cues), beta ERD was stronger
(Townsend et al., 2019). The purpose of the present article
was to further examine visual–vestibular weighting by
manipulating the timing of onset of the self-motion cues.
Previous research has demonstrated that the beta band
is an index of visual–vestibular weighting, and that atten-
tion allocation plays a key role in how weighting is distrib-
uted among multisensory inputs (Townsend et al., 2019,
2022). Those studies, Tuttavia, did not investigate the
impact stimulus onset timing has on the process of
visual–vestibular weighting within self-motion perception.
Previous research has shown that discrepancies in the
onset timing of audiovisual stimuli can affect multisensory
weighting (Fister, Stevenson, Nidiffer, Barnett, & Wallace,
2016; Sheppard, Raposo, & Churchland, 2013). We need a
better understanding about how the interaction of atten-
tion allocation and temporal misalignment affect the
underlying cortical activity associated with visual–
vestibular integration during self-motion perception. IL
goals of the present study were twofold. The first goal was
to examine the effect of attention allocation and temporal
asynchrony on induced ERSP, specifically the power and
time course of beta oscillations associated with visual–
vestibular weighting. The second goal was to examine
induced gamma oscillations. Previous multisensory
research (per esempio., Senkowski et al., 2007) demonstrated more
powerful feature-binding gamma ERS when audiovisual
multisensory cue onsets were presented closer in time.
The present study extends this work by asking whether
feature-binding reflected by gamma ERS is similar for
visual–vestibular integration.
Participants attended to either physical (ignoring visual)
or visual (ignoring physical) motion cues (blocked design)
and discriminated between left and right self-motion head-
ing (random presentation within a block). There were
three SOA conditions: (1) visual motion onset 100 msec
before physical motion onset, (2) physical motion onset
100 msec before visual motion onset, E (3) simulta-
neous visual and physical motion onset. Given previous
research (Townsend et al., 2019, 2022), we hypothesized
that beta ERD would be most powerful when participants
Townsend et al.
1093
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
C
N
_
UN
_
0
1
9
9
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
attended to the physical-motion cues, and beta ERS would
be most powerful when participants attended to visual-
motion cues. This pattern, Tuttavia, would be modulated
by the temporal lag conditions, such that beta ERD in
response to attention to physical motion would be
enhanced if the attended physical-motion cue was pre-
sented before the ignored visual-motion cue, and beta
ERS in response to attention to visual motion would be
enhanced if the attended visual-motion cue was presented
before the ignored physical-motion cue. Inoltre, if
gamma ERS is most powerful during conditions of tempo-
ral synchrony (Senkowski et al., 2007), the present study
may provide evidence that gamma ERS is an index of
general processes related to multisensory binding and
integration across multiple sensory systems. If this is not
the case, feature binding-related gamma ERS may only
be specific to processes such as audiovisual and visual–
tactile integration.
METHODS
Participants
Thirty-six participants (20 women) were recruited from
the McMaster University psychology participant pool and
the McMaster community. The sample size was sufficient
based on a power analysis of data from our previous study
(Townsend et al., 2019; 37 sample size, 0.73 effect size,
0.05 error probability, 0.95 power, four measurements)
conducted by G*Power Software (Faul, Erdfelder,
Buchner, & Lang, 2009). Ages ranged from 17 A 23 years
(M = 18 years, SD = 1.30 years). Those recruited from the
participant pool were compensated with course credits.
All participants self-reported normal or corrected-to-
normal visual acuity and reported no major problems
with vertigo, motion sickness, or claustrophobia. Questo
experiment was approved by the Hamilton Integrated
Research Ethics Board and complied with the Canadian
tri-council policy on ethics.
Stimuli
Visual Motion Stimuli
Visual motion stimuli were presented on a 43-in. LCD
panel, 51 In. in front of the participant, subtending a visual
angle of 41°. The panel had a refresh rate of 60 Hz and a
resolution of 1920 × 1080 (1080P).
The visual display, which contributed to the perception
of self-motion, was composed of a fixation cross in the cen-
ter of the display and two tracks on a gray surface. Each
track consisted of a series of yellow dashes perpendicular
to the length of the track, drawn in perspective to a vanish-
ing point so that the track appeared to extend into the dis-
tance. One track veered right, whereas the other veered
left, both at 35°, starting at the lower center of the display.
Both tracks together subtended a horizontal visual angle
of 33.69°. A horizon line was created by a gray surface upon
which the tracks laid, and a blue sky with white clouds
above, accentuating the perception of traveling along a
track into the distance. The perception of self-motion
along the track was created via a first-person viewpoint ani-
mation that simulated a forward trajectory to align with the
acceleration and perceived velocity that result from the
physical-motion cues (see Figure 1B and C for two tempo-
ral snapshots). The duration of the visual-motion stimulus
on each trial was 700 msec, which included a 200-msec
acceleration period followed by 500 msec at a fixed veloc-
ità. This was followed by a 960-msec pause in the final posi-
tion at the end of the track. At the completion of the trial
(1660 msec), the visual stimulus was reset to the starting
position of the tracks.
Physical Motion Stimuli
A motion simulator provided physical-motion stimuli. IL
motion simulator cabin was supported by a MOOG
Stewart platform with six-degrees-of-freedom motion
(Moog series 6DOF2000E). Participants were seated in a
bucket-style car seat fixed to the cabin floor.
Each physical-motion stimulus consisted of the cabin
moving in a forward linear translation, 35° left or right
for 330 msec at 0.01 G. This forward acceleration was
presented as a precomputed parabolic movement of the
platform. This surge was followed by a corresponding
1330 msec washout (see Figure 1A). During the washout
period, the cabin is slowly moved to the original position
below threshold for detecting the direction of movement.
Figure 1A also illustrates motion noise above 60 Hz, Quale
is because of mechanical vibrations of the simulator. Noi
also presented very small movements in random direc-
tions other than the forward motion that simulated the
feel and sound of wheels on the road, and which also
helped to mask mechanical vibrations and direction of
washout motion. As can be seen in the figure, the mechan-
ical vibrations and injected noise have very low energy,
which is experienced as a rumbling accompanying the per-
ception of forward motion. The acceleration intensity was
selected based on preliminary testing to achieve a clear
perception of forward motion within the spatial restric-
tions of the movement of the platform while minimizing
compensating movements of the head, neck, or upper
body (Townsend et al., 2019). Physical forward accelera-
tions were well above vestibular thresholds of .009 G, COME
discussed by Kingma (2005). The motion force, S(T), era
described by:
8
>><
>>:
s tð Þ
A1 0 ≤ t ≤ tp
−A2 tp ≤ tb
A2 tb ≤ t ≤ te
0 else
where t represents time in seconds, tp represents present
time, tb represents the breakpoint, and te represents the
end time. A1 describes the initial forward acceleration,
−A2 describes the initial (backward) acceleration of the
1094
Journal of Cognitive Neuroscience
Volume 35, Numero 7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
C
N
_
UN
_
0
1
9
9
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
C
N
_
UN
_
0
1
9
9
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. (UN) An example of the profile of physical motion measured during a single trial by an accelerometer (red line); the high-frequency
component represents the high sensitivity of the accelerometer (sensitive to 0.0001 G, sampling at 1 Kh). Note that the frequencies above 60 Hz
represent mechanical vibrations of the motion system and simulator. The x axis represents time, and the y axis represents acceleration (g = m/sec2).
The acceleration profile is similar for 35° left and 35° right physical-motion trials. (B) The visual display before the onset of motion; at this point, IL
participant does not know whether visual motion will indicate travel along the left or right track. (C) A still screen capture of the dynamic visual
motion display at approximately 1 sec after visual onset of a left visual motion trial.
washout, and A2 describes the deceleration of the
washout. Acceleration was measured using an Endevco
accelerometer (model number 752A13), calibrated to
approximately 1-mV/g sensitivity.
Procedure
The entire session was between 1.5 E 2 hr in duration.
The timeline of the session included collection of demo-
graphic information, followed by completion of one
practice block (30 trials; ∼2 min), application of EEG elec-
trodes (25 min), completion of four experimental blocks
(60 min), and participant clean up and debriefing (15 min).
There were 796 experimental trials divided into four
blocks of 199 trials each. Participants fixated on the fixation
cross for the duration of each trial; a blink break was
provided every 15 trials. The attend-visual (AV) E
attend-physical (AP) tasks were blocked to avoid task
switching effects. The task required participants to direct
attention to the visual-motion stimulus and ignore the
physical-motion cues (AV task) or to direct attention to
the physical-motion stimulus and ignore the visual-motion
cues (AP task). They responded with a button press to
indicate whether the direction of the attended-modality
motion was left or right heading.
Given the importance of collecting enough clean data
with correct responses in each attention condition for
EEG analyses, and given that participants have a more dif-
ficult time ignoring the visual while attending the physical
stimulus (Townsend et al., 2019), we collected three AP
blocks compared with one AV block. Presentation order
was controlled so that the AV block was presented as the
Townsend et al.
1095
first, second, or third of the four blocks. Inoltre, A
ensure that participants maintained attention to the
intended modality (especially during AP blocks), each
block contained eight catch trials in which the ignored
modality heading was incongruent with the attended
modality heading.
SOA was manipulated to produce simultaneous (S),
visual-first ( V1st), and physical-first (P1st) conditions. In
the simultaneous condition, visual and physical motion
cues were onset at the same time. In the V1st condition,
the visual motion stimulus was onset 100 msec before the
physical motion, and in the P1st condition, the physical
motion stimulus began 100 msec before the visual motion.
The duration of 100 msec was selected as the SOA based
on previous research that demonstrated a window in
which temporal alignment of visual–vestibular cues
speeds up the perception of self-motion (Kenney et al.,
2020; O’Malley, Townsend, von Mohrenschildt, &
Shedden, 2015). This research provided evidence that a
temporal misalignment of 100 msec delayed the responses
to the self-motion cues, relative to visual–vestibular cues
that were closer in temporal alignment. Così, the benefits
of multisensory integration were weakened, which was the
case regardless of which motion cue was being attended.
There were an equal number of left and right heading
trials in each block, randomly presented.
EEG Data Acquisition
EEG data were collected using the BioSemi ActiveTwo
electrophysiological system (www.biosemi.com) con
128 sintered Ag/AgCl scalp electrodes. Four additional
electrodes recorded eye movements (two placed laterally
from the outer canthi and two below the eyes on the upper
cheeks). Continuous signals were recorded using an open
pass band from direct current to 150 Hz and digitized at
1024 Hz.
EEG Preprocessing
All processing was performed in MATLAB 2014a (IL
MathWorks) using functions from EEGLAB (Delorme &
Makeig, 2004) on the Shared Hierarchical Academic
Research Computing Network (www.sharcnet.ca). EEG
data were band-pass filtered between 1 E 50 Hz, E
epoched from 1000 msec prestimulus to 2000 msec post-
stimulus. Each epoch was baseline corrected using the
whole-epoch mean (Groppe, Makeig, & Kutas, 2009).
Channels with a standard deviation exceeding 200 μV were
interpolated after referencing (on average, 0.97 channels
interpolated per participant, with a total of 35 channels
interpolated). Bad epochs were rejected if they had voltage
spikes exceeding 500 μV or violated EEGLAB’s joint prob-
ability functions (Delorme, Sejnowski, & Makeig, 2007).
Single-subject EEG data were submitted to an extended
adaptive mixture independent component (IC) analysis
(Palmer, Kreutz-Delgado, & Makeig, 2012) with an n −
(1 + interpolated channels) principal components analy-
sis reduction (Makeig, Campana, Jung, & Sejnowski, 1995).
Decomposing an EEG signal into ICs allows for analysis
of each individual signal produced by the brain that would
otherwise be indistinguishable. Dipoles were then fit to
each IC using the fieldtrip plugin for EEGLAB following
adaptive mixture IC analysis (Oostenveld, Fries, Maris, &
Schoffelen, 2011). ICs for which dipoles were located out-
side the brain, or explained less than 85% of the weight
variance, were excluded from further analysis. On average,
20.47 ICs per participant were excluded from analysis.
ERSP Measure Projection Analysis
ERSP was computed for each of the remaining ICs. Fifty
log-spaced frequencies between 3 E 50 Hz were com-
puted, with three cycles per wavelet at the lowest fre-
quency up to 25 at the highest. MPA was used to cluster
ICs across participants using the Measure Projection
Toolbox for MATLAB (Bigdely-Shamlo, Mullen, Kreutz-
Delgado, & Makeig, 2013). MPA is a method of categoriz-
ing the location and consistency of EEG measures, ad esempio
ERSP, across single-subject data into 3-D domains. Each
domain is a subset of ICs that are identified as having
spatially similar dipole models, as well as similar cortical
activity (measure-similarity). MPA fits the selected ICs into
a 3-D model of the brain, composed of a cubic space grid
with 8-mm spacing according to normalized Montreal
Neurological Institute space. The MPA toolbox identified
cortical regions of interest by incorporating the probabilis-
tic atlas of human cortical structures provided by the
Laboratory of Neuroimaging project (Shattuck et al.,
2008). Voxels that fell outside of the brain model (muscle
artifacts, eccetera.) were excluded from the analysis.
We then calculated local convergence values, using an
algorithm based on Bigdely-Shamlo et al. (2013), Quale
deals with the multiple comparisons problem. Local con-
vergence calculates the measure-similarity of dipoles
within a given domain and compares them with random-
ized dipoles. A pairwise IC similarity matrix was created by
estimating the signed mutual information between IC-pair
ERSP measure vectors, assuming a Gaussian distribution,
to compare dipoles. As explained in detail by Bigdely-
Shamlo et al. (2013), signed mutual information was esti-
mated to improve the spatial smoothness of the obtained
MPA significance value beyond determining similarity of
dipoles through correlation. Bootstrap statistics were used
to obtain a significance threshold for convergence at each
location of our 3-D brain model. Following past literature,
we set the raw voxel significance threshold to p < .001
(Chung, Ofori, Misra, Hess, & Vaillancourt, 2017; Bigdely-
Shamlo et al., 2013).
Our analyses focused on two relevant domains: the right
motor area, with the greatest concentration of dipoles
consistent with right premotor and SMA (BA 6), and the
left motor area, with the greatest concentration of dipoles
consistent with left premotor and SMA (BA 6). For the
1096
Journal of Cognitive Neuroscience
Volume 35, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
right motor area, each participant contributed, on average,
2.33 (±1.53) ICs, with each participant contributing at
least one IC, with a range from 1–7 ICs. For the left
motor area, each participant contributed, on average,
2.19 (±1.51) ICs. There were five participants who did
not contribute to this domain. The range of contributed
ICs was 0–6.
ERSPs were computed for each experimental condition
within each domain calculated by MPA. Bootstrap statistics
were used to assess differences in ERSP between condi-
tions to uncover main effects of task and SOA. Differences
at each power band were computed by projecting the
ERSP for each condition to each voxel in the domain. This
projection was weighted by dipole density per voxel and
then normalized by the total domain voxel density for each
participant. Analysis of projected source measures were
separated into discrete spatial domains by threshold-
based affinity propagation clustering based on a similarity
matrix of pairwise correlations between ERSP measure
values for each position. Following Chung et al. (2017),
we used the maximal exemplar-pair similarity, which
ranges from 0–10 to set a value of 0.8 (Chung et al., 2017;
Ofori, Coombes, & Vaillancourt, 2015; Bigdely-Shamlo
et al., 2013).
RESULTS
Behavioral Results
Behavioral data were analyzed with two 2 × 3 repeated-
measures ANOVAs for measures of judgment accuracy
and RT. Outliers were defined as trials with RTs greater
than 3 SDs above or below the mean in each condition
and were eliminated from all further analyses. The
Greenhouse–Geisser correction was applied to all effects
that violated Mauchley’s test of sphericity. All behavioral
results are illustrated in Figure 2.
Accuracy
Participants were more accurate at discriminating direc-
tion in the attend-visual task (M = 99%, SE = .003) than
the attend-physical task (M = 95%, SE = .01), F(1, 35) =
10.50, p = .003, ηp
2 = .23. Moreover, there was a main
effect of SOA on accuracy (Greenhouse–Geisser cor-
rected), F(1.69, 59.02) = 5.77, p = .03, ηp
2 = .14, and a
Task ×SOA interaction, F(2, 70) = 5.00, p = .009, ηp
2 =
.13. Bonferroni-corrected pairwise comparisons sup-
ported the observation that the SOA effects were apparent
during the attend-physical task only; there were no signif-
icant differences in accuracy between any of the SOA
conditions during the attend-visual task. More specifically,
participants were more accurate in the attend-physical
physical-first (AP(P1st)) condition (M = 95.9%, SE = .01)
than the attend-physical visual-first (AP( V1st)) condition
(M = 94.10%, SE = 0.02; p = .007).
Response Time
Participants were faster at discriminating direction in the
attend-visual task (M = 1018 msec, SE = 90.20) than
the attend-physical task (M = 1409 msec, SE = 78.72),
F(1, 35) = 39.43, p < .001, ηp
2 = .53. There was a main
effect of SOA, F(2, 70) = 519.35, p < .001, η p
2 = .94, such
that responses were fastest in the V1st conditions (M =
1189 msec, SE = 6.10), followed by the simultaneous
conditions (M = 1317 msec, SE = 5.88), and slowest in
the P1st conditions (M = 1451 msec, SE = 6.16). There
was a trend toward a Task × SOA interaction on RTs
(Greenhouse–Geisser corrected), F(1.52, 53.22) = 3.48,
p = .05, η p
2 = .9, such that Bonferroni-corrected pairwise
comparisons revealed RT differences across conditions in
both attend-physical and attend-visual tasks. During the
attend-visual task, responses were faster for the visual-first
(AV( V1st)) trials (M = 899 msec, SE = 92.99) compared
Figure 2. Behavioral data. (A) Boxplots for accuracy data showing post hoc simple effects within each task. (B) Boxplots for RT data showing
post hoc simple effects within each task. All p values were corrected for multiple comparisons using Bonferroni correction (*p < .05, **p < .001).
AVV = attend-visual visual first; AVS = attend-visual simultaneous, attend-visual physical first; APV = attend-physical visual first; APS = attend-physical
simultaneous; APP = attend-physical physical first.
Townsend et al.
1097
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
with simultaneous (AV(S)) trials (M = 1020 msec, SE =
90.19; p < .001), which were in turn faster than physical-
first (AV(P1st)) trials (M = 1135 msec, SE = 88.36; p <
.001). Likewise, during the attend-physical task, responses
were faster for the AP( V1st) trials (M = 1269 msec, SE =
77.69) compared with simultaneous (AP(S)) trials (M =
1406 msec, SE = 79.04; p < .001), which were in turn faster
than AP(P1st) trials (M = 1552 msec, SE = 80.12; p < .001).
Thus, two important observations are that (1) participants
are faster overall when attending to visual motion, but
importantly, (2) both attend-visual and attend-physical
conditions are highly sensitive to which stimulus was pre-
sented first. Exploring the ERSP results provides insights
into how the temporal order of stimuli may be affecting
multisensory integration and thus leading to differences
in accuracy and RTs.
Oscillatory Power
Effects of SOA in Attend-Visual Task
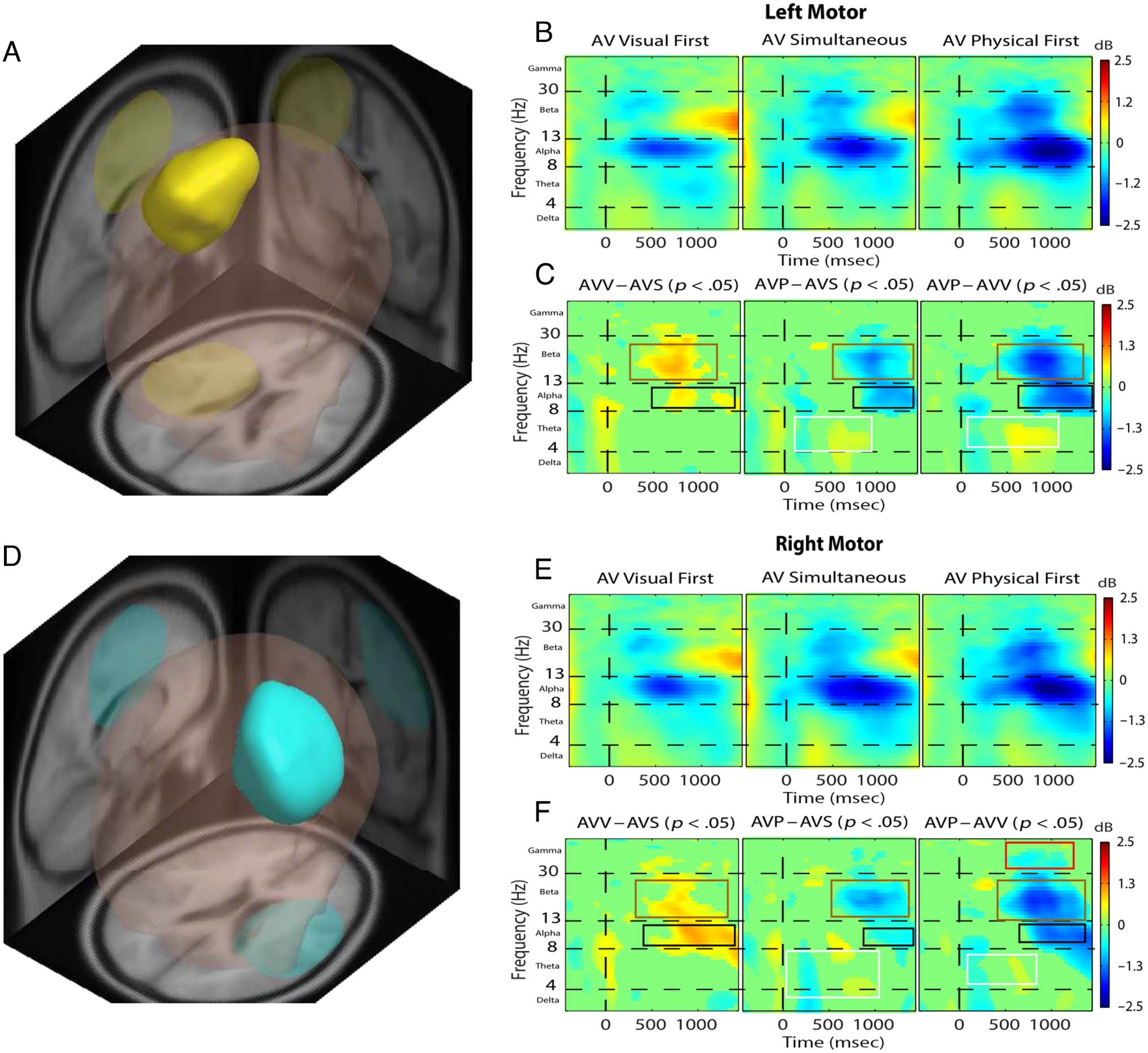
Figure 3 presents a comparison of the left and right
motor areas to illustrate the effect of the timing of
the stimulus onset on the cortical activity during the
attend-visual conditions in both MPA domains. All ERSP
represents a difference in oscillatory power compared
with baseline (pretrial) cortical activity, where an ERS
represents more spectral power than baseline and an
ERD represents less spectral power than baseline. The
1000-msec baseline EEG was recorded during the ISI
before each trial, while the simulator was stationary and
participants were fixating on the fixation cross. Figure 3A
shows the left motor area, with the highest dipole den-
sity in the premotor and SMA (BA 6), and Figure 3D
shows the right motor area, with the highest dipole den-
sity in the premotor and SMA (BA 6). In Panel B (left
motor) and E (right motor), we show the associated
ERSP plots for the AV( V1st), AV(S), and AV(P1st) condi-
tions. The ERSP plots are followed by bootstrapped
comparisons (α = .05) between each possible pair of
conditions for left (Panel C) and right (Panel F) motor
areas. The following sections will describe observations
of the activity changes associated with experimental con-
ditions across frequency bands theta, alpha, beta, and
gamma. All of the comparisons outlined in the following
sections were significant at p < .05.
Theta-band latency differences. The AV(P1st) condition
elicited theta ERS significantly later than the AV(S) and
AV( V1st) conditions. Specifically, in both the left and right
motor areas (Panels C and F, respectively), AV(S) elicited
greater theta ERS from ∼100 msec to 200 msec post
stimulus and AV(P1st) elicited greater theta ERS later in the
trial, from ∼500 msec to 950 msec post stimulus. Likewise,
AV(V1st) elicited greater theta ERS from stimulus onset to
300 msec post stimulus and AV(P1st) elicited greater theta
ERS from ∼500 msec to 1000 msec post stimulus.
In the left and right
Alpha-band power differences.
motor areas (C and F, respectively), AV(P1st) elicited
the strongest alpha ERD, compared with AV(S) (∼750–
1500 msec poststimulus) and AV( V1st) (∼600–1500 msec
poststimulus), and AV(S) elicited stronger alpha ERD
than AV( V1st) (∼550–1500 msec poststimulus). Thus, in
general, alpha ERD AV(P1st) > AV(S) > AV( V1st).
Beta-band power differences. Much like the results in
the alpha band, we found that the earlier the physical motion
was presented, the stronger the elicited beta-band ERD
power. In the left and right motor areas (C and F, respec-
tively), AV(P1st) elicited the strongest beta ERD, compared
with AV(S) (∼500–1500 msec poststimulus) and AV(V1st)
(∼400–1500 msec poststimulus), and AV(S) elicited stronger
alpha ERD than AV(V1st) (∼300–1000 msec poststimulus).
Così, in general, beta ERD AV(P1st) > AV(S) > AV(V1st).
Gamma-band power differences. AV( V1st) elicited a
more powerful gamma ERS than AV(P1st) from ∼600–
1200 msec poststimulus in the right motor area (F).
Effects of SOA in Attend-Physical Task
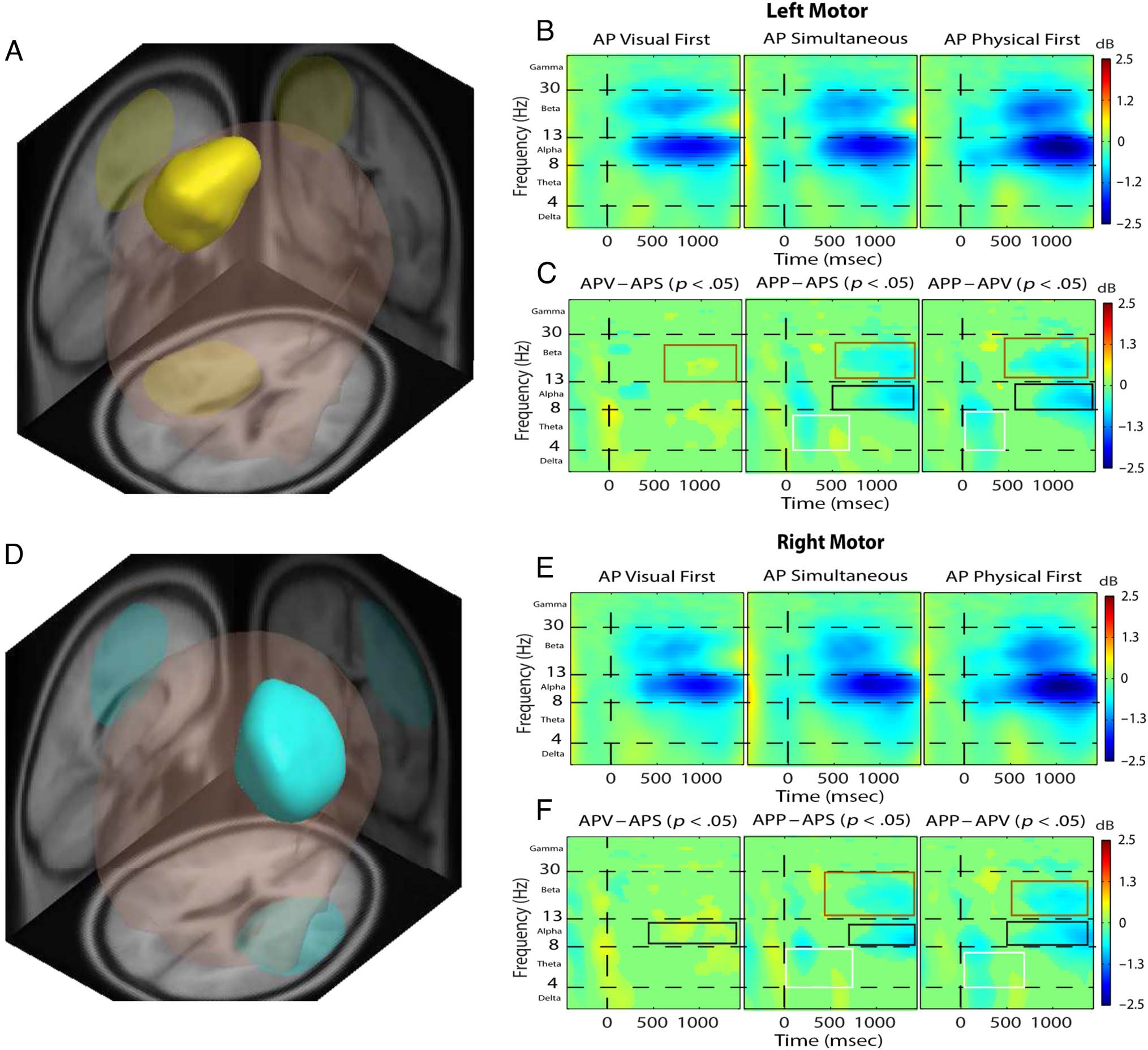
Figura 4 presents a comparison of the same left and right
motor areas as Figure 3 to illustrate the effect of stimulus
onset timing on the cortical activity during the attend-
physical conditions in both MPA domains. All of the com-
parisons outlined in the following sections were significant
at p < .05.
Theta-band latency differences. The AP(P1st) condition
elicited theta ERS significantly later than the AP(S) and
AP( V1st) conditions. Specifically, in both the left and right
motor areas (C and F, respectively), AP(S) elicited greater
theta ERS from stimulus onset to ∼300 msec post stimulus
and AP(P1st) elicited greater theta ERS later in the trial,
from ∼500 msec to 600 msec post stimulus. Likewise,
AP( V1st) elicited greater theta ERS from stimulus onset
to ∼400 msec post stimulus and AP(P1st) elicited greater
theta ERS from ∼500 msec to 600 msec post stimulus.
In the left and right
Alpha-band power differences.
motor areas (C and F, respectively), AP(P1st) elicited
the strongest alpha ERD, compared with AP(S) (∼700–
1500 msec poststimulus) and AP( V1st) (∼600–1500 msec
poststimulus), and AP(S) elicited stronger alpha ERD
than AP( V1st) (∼600–1500 msec poststimulus). Thus, in
general, alpha ERD AP(P1st) > AP(S) > AP(V1st).
In the left and right motor
Beta-band power differences.
areas (C and F, rispettivamente), AP(P1st) elicited the strongest
beta ERD, compared with AP(S) (∼550–1500 msec post-
stimulus) and AP( V1st) (∼500–1500 msec poststimulus),
and AP(S) elicited stronger alpha ERD than AP( V1st)
(∼800–1200 msec poststimulus). Così, in general, beta
ERD AP(P1st) > AP(S) > AP(V1st).
1098
Journal of Cognitive Neuroscience
Volume 35, Numero 7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
C
N
_
UN
_
0
1
9
9
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
C
N
_
UN
_
0
1
9
9
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 3. Attend-visual task. Left motor area (UN, B, and C) and right motor area (D, E, and F) identified by MPA and respective ERSP analysis. IL
ERSP plots show time (msec) across the x axis and frequency of the EEG signal along the y axis. Panels B (left) and E (right) show the associated ERSP
plots for the attend-visual visual first (AV( V1st)), attend-visual simultaneous (AVS), and attend-visual physical first (AV(P1st)) conditions. Panels C (left
motor area) and F (right motor area) show the bootstrapped comparisons ( P < .05) between each possible pair of conditions. ERS power is depicted
in yellow/red, ERD power is depicted in blue, and green shows no difference in spectral power compared with baseline. MPA motor areas: (A and D)
3-D representations of the brain with the yellow region representing the left motor area and the blue region representing the right motor area. The
greatest concentration of dipoles in left and right regions was consistent with premotor and SMAs (BA 6). (B and E) ERSP plots for each condition.
(C and F) Bootstrapped comparisons examine each possible pair of conditions; frequency and time of significant comparisons are shown by the
colored boxes. Both left and right motor areas show similar conditional differences. Theta: AV( V1st) and AV(S) elicits theta ERS significantly earlier
than AV(P1st) (white boxes). Alpha: AV(P1st) elicits stronger alpha ERD than AV(S) and AV( V1st), and AV(S) elicits strong alpha ERD than AV( V1st)
(black boxes). Beta: AV(P1st) elicits stronger beta ERD than AV(S) and AV( V1st), and AV(S) elicits stronger beta ERD than AV( V1st) (brown boxes).
Gamma: Differences in gamma existed only in the right motor area: The AV( V1st) condition elicited significantly stronger gamma ERS than AV(P1st)
(red boxes).
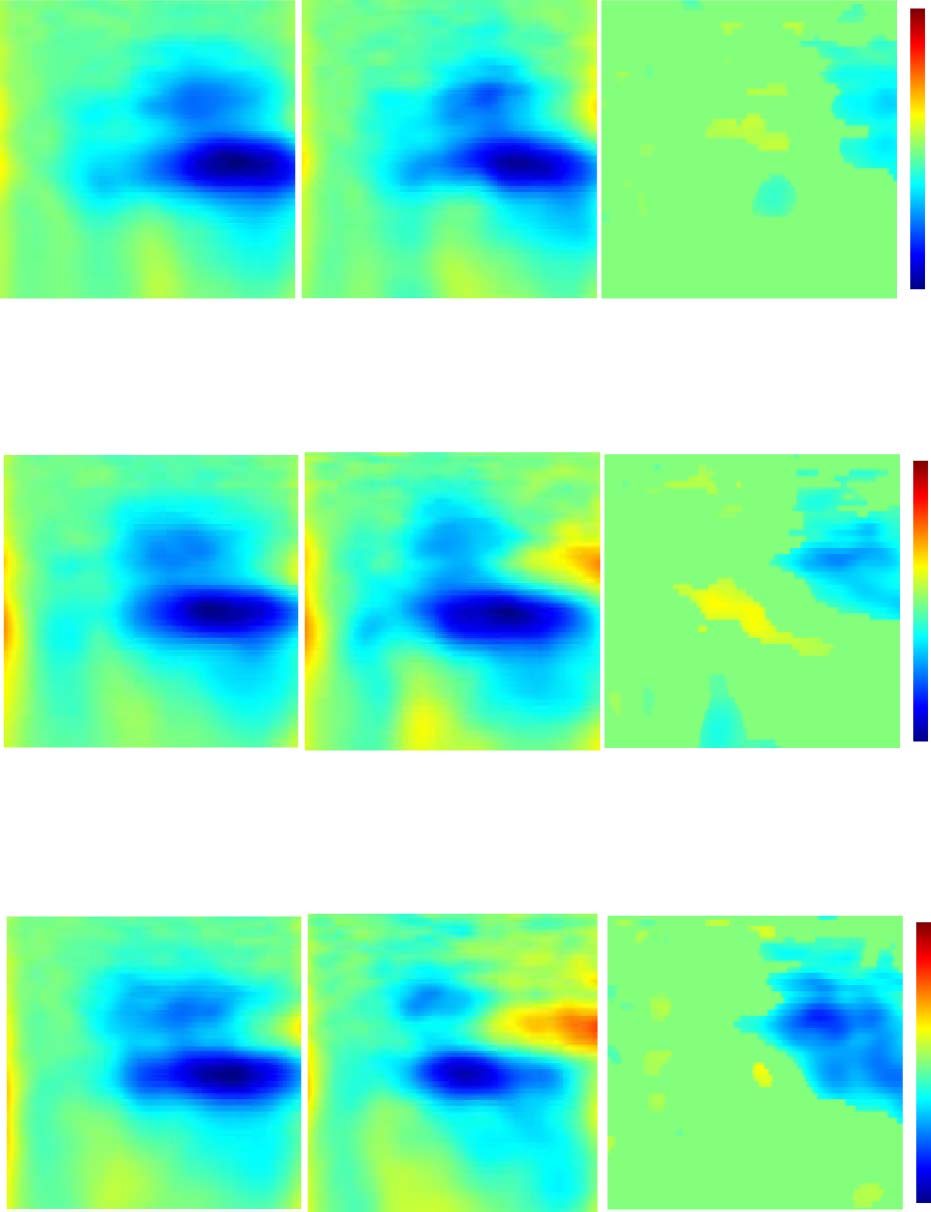
Effects of Attention Allocation across SOA Conditions
Figure 5 presents the same right motor area as Figures 3
and 4 to illustrate the interaction of stimulus onset timing
and attention allocation. We compared cortical activity
between conditions of attention allocation at each level
of the SOA condition (i.e., AV(S) vs. AP(S), AV( V1st) vs.
AP( V1st), and AV(P1st) vs. AP(P1st)). Similar results were
found in the left motor area. All of the comparisons out-
lined in the following sections were significant at p < .05.
Townsend et al.
1099
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. (Attend-physical task). Left motor area (A, B, and C) and right motor area (D, E, and F) identified by MPA and respective ERSP analysis. The
ERSP plots show time (msec) across the x axis and frequency of the EEG signal along the y axis. Panels B (left) and E (right) show the associated ERSP
plots for the attend-physical visual first (AP( V1st)), attend-physical simultaneous (APS), and attend-physical physical first (AP(P1st)) conditions. Panels
C (left motor area) and F (right motor area) show the bootstrapped comparisons ( p < .05) between each possible pair of conditions. ERS power is
depicted in yellow/red, ERD power is depicted in blue, and green shows no difference in spectral power compared with baseline. MPA motor areas:
(A) and (D) show 3-D representations of the brain, with the yellow region representing the left motor area and the blue region representing the right
motor area. The greatest concentration of dipoles in the left and right regions were consistent with premotor and SMAs (BA 6). (B and E) ERSP plots
for each condition. (C and F) Bootstrapped comparisons examine each possible pair of conditions; frequency and time of significant comparisons
are shown by the colored boxes. Both left and right motor areas show similar conditional differences. Theta: AP( V1st) and AV(S) elicits theta ERS
significantly earlier than AP(P1st) (white boxes). Alpha: AP(P1st) elicits stronger alpha ERD than AP(S) and AP( V1st), and AP(S) elicits strong alpha ERD
than AP( V1st) (black boxes). Beta: AP(P1st) elicits stronger beta ERD than AP(S) and AP( V1st), and AP(S) elicits stronger beta ERD than AP( V1st)
(brown boxes).
Theta-band power differences. AV(S) elicited a more
powerful theta ERS than AP(S) from ∼250 msec to
400 msec post stimulus (C).
In the right motor area
Alpha-band power differences.
(A), AV(S) elicited a stronger alpha ERD, compared with
AP(S) (∼50–550 msec poststimulus) (C). AP(V1st) elicited
greater alpha ERD than AV( V1st) from ∼800 msec – end
of trial (D).
In the right motor area
Beta-band power differences.
(A), AP(P1st) elicited a stronger beta ERD than AV(P1st)
from ∼550–1500 msec poststimulus (B), AV(S) elicited
a stronger beta ERS than AP(S) from ∼800 msec – end
1100
Journal of Cognitive Neuroscience
Volume 35, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Right motor area identified by MPA and respective ERSP analysis. The ERSP plots show time (msec) across the x axis and frequency of the
EEG signal along the y axis. (B), (C), and (D) show the associated ERSP plots for the attend-physical and attend-visual conditions at each level of the
SOA condition, and the bootstrapped comparisons ( p < .05) between each pair of conditions. ERS power is depicted in yellow/red, ERD power is
depicted in blue, and green shows no difference in spectral power compared with baseline. MPA right motor area: (A) 3-D representations of the
brain with the blue region representing the right motor area. The greatest concentration of dipoles in right region was consistent with premotor and
SMAs (BA 6). (B, C, and D) Bootstrapped comparisons examine each possible pair of conditions; frequency and time of significant comparisons are
shown by the colored boxes. Theta: AV(S) elicits stronger theta ERS than AP(S) (C; white box). Alpha: AV(S) elicits stronger alpha ERD than AP(S)
(C), and AP( V1st) elicits stronger alpha ERD than AV( V1st) (D; black boxes). Beta: AP(P1st) elicits stronger beta ERD than AV(P1st) (B), AV(S) elicits
stronger beta ERS than AP(S) (C), and AV( V1st) elicits stronger beta ERS than AP( V1st) (D; brown boxes).
of trial (C), and AV( V1st) elicited more powerful beta ERS
than AP(V1st) from ∼700 msec – end of trial (D).
DISCUSSION
Behavioral research has demonstrated a temporal binding
window for visual–vestibular integration, in which multi-
sensory integration affects heading perception, temporal
order judgements, and attention allocation (e.g.,
Rodriguez & Crane, 2021; Shayman et al., 2018). Research
exploring the cortical processes underlying this temporal
window is currently scarce. To better understand the
online processes related to multisensory temporal bind-
ing, we must look to literature focused on the integration
of other senses, such as audiovisual, or visuotactile inte-
gration. Studies such as Senkowski et al. (2007) have dem-
onstrated that the closer audiovisual stimuli are presented
temporally, the more powerful the elicited feature-binding
gamma ERS response. Past multisensory research has
demonstrated a Gaussian integration window, in which
integration breaks at a temporal asynchrony specific to
the senses being integrated (e.g., Rodriguez & Crane,
2021). The present study explored how EEG oscillations
related to attention and multisensory weighting in self-
motion perception (theta, alpha, and beta; Townsend
et al., 2019, 2022), and multisensory feature binding
(gamma; Senkowski et al., 2007) were affected by varying
conditions of SOA. All differences in cortical activity dis-
cussed are projected from the motor area (likely including
integrative areas such as ventral intraparietal area and
medial superior temporal area) based on the MPA, which
identified ROIs across participants.
The Effects of Timing Onset within an
Attended Modality
Recent research by Townsend et al. (2019, 2022) showed
that theta, alpha, and beta oscillations reveal brain net-
works involved in the perception of self-motion. More-
over, the power of these individual oscillations changed
Townsend et al.
1101
dynamically depending on which sensory inputs were
attended to. Taken together, our two previous studies
demonstrated that the beta band is most sensitive to
changes in visual–vestibular weighting. Specifically, these
studies showed that a strong beta ERS is an electrophysi-
ological signature of heavy visual weighting, and a strong
beta ERD is a signature of vestibular weighting.
The current study revealed changes in the same spectral
bands as the previously mentioned studies and contrib-
uted additional key insights to the understanding of self-
motion perception. One robust result that we observed
was when presenting an attended motion cue before an
ignored cue, the power of the beta oscillation associated
with weighting bias toward the attended modality (ERS for
visual and ERD for vestibular) was greater than during
simultaneous presentation of the attended and ignored
cues. This result suggests that the power of weighting-
related beta oscillations during self-motion perception is
also sensitive to the timing of the onset, and not just atten-
tion allocation. Regardless of which modality is being
attended to, the earlier the attended motion cue is pre-
sented in relation to the ignored cue, the more powerful
the weighting-related ERSP. The inverse was true when
the ignored cues were presented before the attended
cues. Beta ERS was less powerful in the AV(P1st) condition
versus AV(S), and beta ERD was less powerful in the
AP( V1st) condition versus AP(S).
The beta cycle has long been thought to reflect an initi-
ation and termination of motor output (for a review, see
Kilavik, Zaepffel, Brovelli, MacKay, & Riehle, 2013). Con-
trary to this hypothesis, Townsend et al. (2019, 2022) dem-
onstrated a beta rebound during passive full-body motion
that was induced by attention, and suggested that beta
oscillations during motor processing may actually reflect
perceptual weighting of the visual, vestibular, and propri-
oceptive systems. The beta rebound may reflect the
inhibition of processing the physical-motion stimuli, con-
sidering visual–vestibular integration is a subadditive
process. Subadditive inhibition typically occurs during
integration when there is a discrepancy in the reliability
of multiple sensory inputs (Angelaki, Gu, & DeAngelis,
2009). The Townsend et al. (2022) study showed that
participants performed the heading discrimination task
at 99% accuracy in both visual- and physical-motion only
conditions (the same motion stimuli as the current study).
Considering there were likely no significant differences in
reliability between the two sensory inputs, we believe that
the temporal advantage caused by the SOA led to strong
inhibitory responses during integration. Our behavioral
and EEG results fall in line with Townsend et al. (2019,
2022). Similar to our previous research, the average of
participants’ accuracy on the heading discrimination task
ranged between conditions from 98–100%. We believe the
oscillatory differences in the beta band between the
stimulus onset timing conditions may be a product of
the perceptual weights being changed because of the
SOA. For example, the processing of the visual stimulus
during the AV( V1st) condition began 100 msec before
the processing of the physical-motion stimulus. This
perceptual head start could have increased the weighting
in favor of the visual stimulus, more so than in the AV(S)
condition. A similar weighting bias may have taken place
during the attend-physical conditions, as we found similar
results (but in beta ERD). These power differences in ERSP
did not result in differences in accuracy, however (attend-
visual 99% accuracy, attend-physical 95% accuracy). We
believe that the tasks may not have been sensitive enough
to capture correlations between behavioral differences
and oscillatory power.
RTs, on the other hand, were affected by the SOA. Keep-
ing in mind that RTs were measured from the onset of the
to-be-attended stimulus, RTs were fastest when the visual-
motion cues were presented first regardless of whether
visual or physical cues were attended. In contrast, RTs
were slowest when the physical-motion cues were pre-
sented first, regardless of which cue was attended. The
visual system is dominant over the vestibular system, as
reported in many studies (e.g., Angelaki et al., 2009),
and it is not surprising that we see this RT effect with
100-msec SOAs. Visual cues also lead to faster perceptual
processing compared with vestibular cues (Barnett-Cowan
& Harris, 2013), and the visual cue would have provided
stronger priming than the vestibular cue when attention
was directed to the opposite cue. Thus, RTs benefited
more when the visual-motion cue was presented first.
The present study clearly demonstrates that the timing
of stimulus onset is a critical component of the visual–
vestibular weighting process and is indexed by dynamic
changes in the beta band.
The Interaction of Stimulus Timing and
Attentional Selection
Not only did we find that the timing of stimulus onsets
affected ERSP, we also found an interaction between the
timing of onsets and attention allocation. This result has
a direct application to pilot training; for example, current
policies of Transport Canada and Federal Aviation Admin-
istration require physical cues to motion to precede visual
cues to motion during pilot simulator training. Pilots are
trained to attend to visual instruments and ignore vestib-
ular inputs caused by forces such as turbulence, to avoid
spatial disorientation (Braithwaite, 1997). One question
that arises from this practice is how the temporal
asynchrony and selective attention interact to affect pilots’
multisensory processing. We compared the visual- versus
the physical-motion conditions at each SOA condition. Our
comparison of AP(S) versus AV(S) was a replication of a
condition in Townsend et al. (2019), and we found similar
results in the present study, the most important observa-
tion being stronger beta ERS in attend-visual conditions
and stronger beta ERD in attend-physical conditions. This
comparison acted as a baseline, whereas the other two
comparisons presented novel findings.
1102
Journal of Cognitive Neuroscience
Volume 35, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The comparisons AP(P1st) versus AV(P1st) (contrasting
attention conditions when the physical stimulus onset first),
and AP(V1st) versus AV(V1st) (contrasting attention condi-
tions when the visual stimulus onset first) demonstrated
an interaction of attention allocation and SOA in the
beta band. When the physical-motion cue was presented
100 msec before the visual cue, there were fewer ERSP
differences between AP(P1st) versus AV(P1st), compared with
the baseline comparison. Most notably, the typical beta
rebound elicited by attention to the visual-motion cue was
not present in the AV(P1st) condition. Based on the findings
of Townsend et al. (2019, 2022), the lack of a beta rebound
in the AV(P1st) condition suggests that presenting the
physical-motion cue before the visual-motion cue resulted
in greater weighting of vestibular signals than if the motion
cues were presented simultaneously. This finding is rele-
vant to simulator training for pilots. If the vestibular cue
to motion is presented before the visual cue, it may disrupt
the operator’s ability to down-weight potentially disorient-
ing vestibular cues that pilots are trained to ignore.
The lack of a beta rebound in the AV(P1st) condition
resulted in relatively little difference in ERSP between
AP(P1st) versus AV(P1st). However, when the visual-motion
cue was presented 100 msec before the physical-motion
cue, there was a robust beta ERS in the AV( V1st) condition
versus a beta ERD in the AP( V1st) condition. This analysis
revealed that visual–vestibular weighting is more sensitive
to changes in the onset timing of the visual cues to motion
than the vestibular cues. This finding is supported by
Barnett-Cowan and Harris (2013), who demonstrated that
perception of visual stimuli is faster than perception of
vestibular stimuli. Considering the visual cue naturally
has a temporal advantage (during simultaneous presenta-
tion), it is likely that the vestibular cue would need to be
presented more than 100 msec before the visual cue to
create the robust ERSP differences that were demon-
strated between the conditions of attention allocation
when the visual cue was presented first.
Feature-binding Gamma ERS in
Visual–Vestibular Integration
We examined gamma ERS under varying conditions of
SOA to test the temporal correlation hypothesis (Engel,
Fries, & Singer, 2001; Singer & Gray, 1995) in the context
of visual–vestibular integration. This hypothesis posits that
synchronization of gamma-band oscillations is a key mech-
anism for integration across distributed cortical networks.
Evidence supporting this hypothesis has been demon-
strated in multiple studies (e.g., Senkowski et al., 2007;
Sakowitz, Quiroga, Schürmann, & Başar, 2001) that typi-
cally focus on audiovisual integration. For example,
Senkowski et al. (2007) presented human participants
with audiovisual stimuli with varying degrees of temporal
asynchrony and required them to attend to one modality-
specific stimuli while ignoring the other. They found that
gamma ERS was not significantly different between
modalities but, for both modalities, significantly stronger
gamma ERS was elicited when temporal asynchrony was
25 msec or less, compared with longer SOAs. In the pres-
ent study, the temporal correlation hypothesis predicts
that the simultaneous conditions (AP(S) and AV(S)) elicit
stronger gamma ERS compared with the V1st and P1st
conditions. Our results do not support this hypothesis.
The present study only found differences in the gamma
band when comparing the AV( V1st) and AV(P1st) condi-
tions, such that AV( V1st) elicited stronger gamma ERS than
AV(P1st). We are currently unaware of any literature
directly explaining this finding. We offer two possible con-
clusions for our results. First, visual–vestibular integration
does not rely on gamma ERS to synchronize modality-
specific information across cortical networks. This facilita-
tion of gamma ERS could be specific to superadditive
integration processes (e.g., audiovisual integration; Dias,
McClaskey, & Harris, 2021) as opposed to subadditive inte-
gration processes (e.g., visual–vestibular integration;
Angelaki et al., 2009). Or second, visual–vestibular integra-
tion has a broader temporal window than 100 msec for
gamma facilitation (compared with the Senkowski et al.,
2007, temporal window of 25 msec), and therefore our
experimental design was not sensitive enough to detect
differences in gamma ERS because of SOA. A broader tem-
poral window for visual–vestibular integration would be
consistent with behavioral research (Rodriguez & Crane,
2021) and research demonstrating that perception for ves-
tibular inputs being relatively slower than other senses
(Barnett-Cowan & Harris, 2013). More research needs to
be conducted to better understand the role of stimulus
timing in visual–vestibular feature binding.
Limitation and Future Directions
Our heading discrimination task required participants to
push a button as quickly as possible to make a heading
judgment. It is possible that the preparation and execution
of thumb movements during the button press contributed
to the recorded EEG signal in the motor areas. Pilot studies
revealed that participants had a tendency to only attend to
visual cues to motion unless they were told that some
physical-motion cues were spatially incongruent to
visual-motion cues. Collecting RTs during the heading
judgment task was important to ensure that participants
attended to the correct motion cues to elicit the appropri-
ate cortical activity. Our previous research (Townsend
et al., 2019, 2022) demonstrated that RT data were diag-
nostic of attention allocation, such that visual headings
were judged faster than physical headings.
The somatosensory system detects pressure and stretch
on the skin, muscles, and joints during self-motion
(Lackner, 1992). The forces generated by acceleration that
produce vestibular or proprioceptive cues would be
strong signals of self-motion perception; however, forces
generated by the acceleration of our motion simulator
would have also stimulated receptors in the back, seat,
Townsend et al.
1103
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
and feet of the seated participants. Although there is
evidence from patients with spinal lesions that the somato-
sensory system does not contribute significantly to our per-
ception of self-motion (Walsh, 1961), we cannot completely
rule out the somatosensory system’s contribution to our
EEG signal projecting from the motor areas.
Functional neuroimaging studies exploring the neural
correlates of visual motion perception typically use optic
flow to elicit cortical responses to vection, or the illusion
of inertial motion generated by visual-only stimuli.
Some studies have compared coherent optic flow to
control stimuli such as random (incoherent) dot motions
(e.g., Cardin & Smith, 2010), static dot patterns (e.g.,
Deutschländer et al., 2004), or spatially scrambled versions
of the original self-motion stimulus (e.g., Barry et al.,
2014). In these studies, participants are not physically
moved, so researchers commonly rely on self-report data
to determine whether participants experienced the
vection illusion. We did not collect self-report data to
determine whether participants experienced vection
from our visual-motion cues in the present study. There-
fore, we cannot be completely certain that our visual-
motion stimuli would have elicited vection on their own.
However, a large body of research has shown that visually
induced vection is strengthened when paired with vestib-
ular stimulation (e.g., Gallagher, Dowsett, & Ferrè, 2019;
Weech & Troje, 2017; Johnson, Sunahara, & Landolt,
1999). Our visual- and physical-motion stimuli were
developed to combine for an immersive experience of
self-motion that is similar to environments used in aviation
and driving research and training.
Our research can be applied to the clinical space to bet-
ter understand pathologies of self-motion perception and
visual–vestibular integration. Patients with pathologies
such as Mal de Débarquement Syndrome ( Van Ombergen,
Van Rompaey, Maes, Van de Heyning, & Wuyts, 2016),
Persistent Postural-Perceptual Dizziness (Popkirov, Staab,
& Stone, 2018), and Parkinson’s disease ( Yakubovich
et al., 2020) show lower thresholds for self-motion percep-
tion. For example, a recent study has shown that, com-
pared with healthy, age-matched controls, Parkinson’s
disease patients perform worse on heading judgment
tasks because of overweighting of impaired visual-motion
cues (Yakubovich et al., 2020). If we can establish electro-
physiological biomarkers of the healthy versus impaired
self-motion perception, we will develop a better under-
standing of the integration and motor impairments that
are common in pathologies such as Parkinson’s disease.
Identification of these biomarkers in the prediagnostic
phase of the disease could lead to a greater time window
for possible preventative measures and earlier treatments
(Noyce, Lees, & Schrag, 2016).
Conclusion
The present study examined cortical activity elicited in
response to self-motion cues that varied in attention
allocation and stimulus onset synchrony. There were two
main findings. First, SOA produced robust differences in
cortical activity during attention to both visual and physical
motion. The electrophysiological signatures of visual
(strong beta ERS) versus vestibular (strong beta ERD)
weighting bias were enhanced when the attended motion
cue was presented 100 msec before the ignored cue.
When comparing across conditions of attention allocation,
presenting the visual-motion cue first created more robust
conditional differences than when physical-motion cues
were presented first. These results demonstrate that the
timing of visual–vestibular stimuli plays a critical role in
multisensory weighting during self-motion perception,
and that this weighting process is more sensitive to tem-
poral changes in visual stimuli compared with vestibular
stimuli. Second, contrary to the findings of several audio-
visual and visuotactile studies, the temporal synchrony of
visual- and physical-motion cues did not elicit gamma ERS
beyond baseline. It is possible that the 100-msec SOA was
not long enough to elicit these hypothesized differences.
It could also be the case that visual–vestibular integration
does not elicit processes indexed by gamma ERS.
Reprint requests should be sent to Ben Townsend, Department
of Psychology, Neuroscience and Behaviour, McMaster Univer-
sity, 1280 Main St. West, Hamilton, Ontario, Canada L8S 4 L8, or
via e-mail: townsepb@mcmaster.ca.
Data Availability Statement
The data and code for all analyses are available online at
https://github.com/ bentownsend11/Stimulus-onset
-asynchrony-affects-attention-related-ERSP-in-self-motion
-perception.
Author Contributions
Ben Townsend: Conceptualization; Formal analysis; Inves-
tigation; Methodology; Project administration; Visualiza-
tion; Writing—Original draft; Writing—Review & editing.
Joey K. Legere: Formal analysis; Software. Martin v.
Mohrenschildt: Funding acquisition; Methodology;
Resources; Software; Supervision. Judith M. Shedden:
Conceptualization; Funding acquisition; Methodology;
Project administration; Resources; Supervision; Writing—
Review & editing.
Funding Information
Funding for this study was provided to JMS and MvM by
The Natural Sciences and Engineering Research Council
of Canada, grant numbers: RGPGP-2014-00051 and
RGPIN-2020-07245; and the Canada Foundation for Inno-
vation (https://dx.doi.org/10.13039/501100000196), grant
number: 2009M00034. These funding sources had no
1104
Journal of Cognitive Neuroscience
Volume 35, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
involvement in the study design, the collection, analysis
and interpretation of data, in the writing of the report,
and in the decision to submit the article for publication.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .675; W/M = .125;
M/ W = .15; W/ W = .05.
REFERENCES
Alais, D., & Burr, D. (2004). The ventriloquist effect results from
near-optimal bimodal integration. Current Biology, 14,
257–262. https://doi.org/10.1016/j.cub.2004.01.029, PubMed:
14761661
Angelaki, D., Gu, Y., & DeAngelis, G. (2009). Multisensory
integration: Psychophysics, neurophysiology, and
computation. Current Opinion in Neurobiology, 19,
452–458. https://doi.org/10.1016/j.conb.2009.06.008,
PubMed: 19616425
Barnett-Cowan, M., & Harris, L. R. (2009). Perceived timing of
vestibular stimulation relative to touch, light and sound.
Experimental Brain Research, 198, 221–231. https://doi.org
/10.1007/s00221-009-1779-4, PubMed: 19352639
Barnett-Cowan, M., & Harris, L. R. (2013). Vestibular perception
is slow: A review. Multisensory Research, 26, 387–403.
https://doi.org/10.1163/22134808-00002421, PubMed:
24319930
Neuroscience, 16, 130–138. https://doi.org/10.1038/nn.3304,
PubMed: 23354386
Cardin, V., & Smith, A. T. (2010). Sensitivity of human visual and
vestibular cortical regions to egomotion-compatible visual
stimulation. Cerebral Cortex, 20, 1964–1973. https://doi.org
/10.1093/cercor/bhp268, PubMed: 20034998
Chung, J. W., Ofori, E., Misra, G., Hess, C. W., & Vaillancourt, D. E.
(2017). Beta-band activity and connectivity in sensorimotor
and parietal cortex are important for accurate motor
performance. Neuroimage, 144, 164–173. https://doi.org/10
.1016/j.neuroimage.2016.10.008, PubMed: 27746389
DeAngelis, G. C., & Angelaki, D. E. (2012). Visual–vestibular
integration for self-motion perception. In M. M. Murray &
M. T. Wallace (Eds.), The neural bases of multisensory
processes. CRC Press/Taylor & Francis. Available from: https://
www.ncbi.nlm.nih.gov/books/NBK92839/.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Delorme, A., Sejnowski, T., & Makeig, S. (2007). Enhanced
detection of artifacts in EEG data using higher-order statistics
and independent component analysis. Neuroimage, 34,
1443–1449. https://doi.org/10.1016/j.neuroimage.2006.11
.004, PubMed: 17188898
Deutschländer, A., Bense, S., Stephan, T., Schwaiger, M.,
Dieterich, M., & Brandt, T. (2004). Rollvection versus
linearvection: Comparison of brain activations in PET.
Human Brain Mapping, 21, 143–153. https://doi.org/10.1002
/hbm.10155, PubMed: 14755834
Dias, J. W., McClaskey, C. M., & Harris, K. C. (2021). Audiovisual
speech is more than the sum of its parts: Auditory–visual
superadditivity compensates for age-related declines in
audible and lipread speech intelligibility. Psychology and
Aging, 36, 520–530. https://doi.org/10.1037/pag0000613,
PubMed: 34124922
Engel, A. K., Fries, P., & Singer, W. (2001). Dynamic predictions:
Oscillations and synchrony in top–down processing. Nature
Reviews Neuroscience, 2, 704–716. https://doi.org/10.1038
/35094565, PubMed: 11584308
Faul, F., Erdfelder, E., Buchner, A., & Lang, A. G. (2009).
Statistical power analyses using G*Power 3.1: Tests for
correlation and regression analyses. Behavior Research
Methods, 41, 1149–1160. https://doi.org/10.3758/BRM.41.4
.1149, PubMed: 19897823
Fister, J. K., Stevenson, R. A., Nidiffer, A. R., Barnett, Z. P., &
Wallace, M. T. (2016). Stimulus intensity modulates
multisensory temporal processing. Neuropsychologia,
88, 92–100. https://doi.org/10.1016/j.neuropsychologia.2016
.02.016, PubMed: 26920937
Barry, R. J., Palmisano, S., Schira, M. M., De Blasio, F. M.,
Gallagher, M., Dowsett, R., & Ferrè, E. R. (2019). Vection in virtual
Karamacoska, D., & MacDonald, B. (2014). EEG markers of
visually experienced self-motion (vection). In Frontiers of
Human Neuroscience Conference Abstract: Australasian
Society for Psychophysiology, Inc. https://doi.org/10.3389
/conf.fnhum.2014.216.00013
Bigdely-Shamlo, N., Mullen, T., Kreutz-Delgado, K., & Makeig, S.
(2013). Measure projection analysis: A probabilistic approach
to EEG source comparison and multi-subject inference.
Neuroimage, 72, 287–303. https://doi.org/10.1016/j
.neuroimage.2013.01.040, PubMed: 23370059
Braithwaite, M. G. (1997). The British Army Air Corps in-flight
spatial disorientation demonstration sortie. Aviation, Space,
and Environmental Medicine, 68, 342–345. PubMed:
9096833
Buzsáki, G., & Moser, E. I. (2013). Memory, navigation and theta
rhythm in the hippocampal-entorhinal system. Nature
reality modulates vestibular-evoked myogenic potentials.
European Journal of Neuroscience, 50, 3557–3565. https://doi
.org/10.1111/ejn.14499, PubMed: 31233640
Groppe, D. M., Makeig, S., & Kutas, M. (2009). Identifying
reliable independent components via split-half comparisons.
Neuroimage, 45, 1199–1211. https://doi.org/10.1016/j
.neuroimage.2008.12.038, PubMed: 19162199
Johnson, W. H., Sunahara, F. A., & Landolt, J. P. (1999).
Importance of the vestibular system in visually induced
nausea and self-vection. Journal of Vestibular Research,
9, 83–87. https://doi.org/10.3233/ VES-1999-9202, PubMed:
10378179
Keil, J., & Senkowski, D. (2018). Neural oscillations orchestrate
multisensory processing. Neuroscientist, 24, 609–626.
https://doi.org/10.1177/1073858418755352, PubMed:
29424265
Townsend et al.
1105
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Kenney, D. M., O’Malley, S., Song, H. M., Townsend, B.,
von Mohrenschildt, M., & Shedden, J. M. (2020). Velocity
influences the relative contributions of visual and vestibular
cues to self-acceleration. Experimental Brain Research,
238, 1423–1432. https://doi.org/10.1007/s00221-020-05824-9,
PubMed: 32367145
Kilavik, B. E., Zaepffel, M., Brovelli, A., MacKay, W. A., & Riehle,
A. (2013). The ups and downs of beta oscillations in
sensorimotor cortex. Experimental Neurology, 245, 15–26.
https://doi.org/10.1016/j.expneurol.2012.09.014, PubMed:
23022918
Kingma, H. (2005). Thresholds for perception of direction of
linear acceleration as a possible evaluation of the otolith
function. BMC Ear, Nose and Throat Disorders, 5, 5. https://
doi.org/10.1186/1472-6815-5-5, PubMed: 15972096
Klimesch, W. (2012). Alpha-band oscillations, attention, and
controlled access to stored information. Trends in Cognitive
Sciences, 16, 606–617. https://doi.org/10.1016/j.tics.2012.10
.007, PubMed: 23141428
Lackner, J. R. (1992). Multimodal and motor influences on
orientation: Implications for adapting to weightless and
virtual environments. Journal of Vestibular Research, 2,
307–322. https://doi.org/10.3233/ VES-1992-2405, PubMed:
1342405
Li, W., Piëch, V., & Gilbert, C. D. (2004). Perceptual learning
and top–down influences in primary visual cortex. Nature
Neuroscience, 7, 651–657. https://doi.org/10.1038/nn1255,
PubMed: 15156149
Macaluso, E., Noppeney, U., Talsma, D., Vercillo, T., Hartcher-
O’Brien, J., & Adam, R. (2016). The curious incident of
attention in multisensory integration: Bottom–up vs.
top–down. Multisensory Research, 29, 557–583. https://doi
.org/10.1163/22134808-00002528
Makeig, S., Bell, A., Jung, T. P., & Sejnowski, T. J. (1995).
Independent component analysis of electroencephalographic
data. Advances in Neural Information Processing Systems,
8, 145–151.
Noyce, A. J., Lees, A. J., & Schrag, A. E. (2016). The prediagnostic
phase of Parkinson’s disease. Journal of Neurology,
Neurosurgery & Psychiatry, 87, 871–878. https://doi.org/10
.1136/jnnp-2015-311890, PubMed: 26848171
O’Malley, S., Townsend, B., von Mohrenschildt, M., & Shedden,
J. M. (2015). The integration of physical acceleration cues
with visual acceleration cues. Canadian Journal of
Experimental Psychology-Revue, 69, 349.
Ofori, E., Coombes, S. A., & Vaillancourt, D. E. (2015). 3D
cortical electrophysiology of ballistic upper limb movement
in humans. Neuroimage, 115, 30–41. https://doi.org/10.1016/j
.neuroimage.2015.04.043, PubMed: 25929620
Öhman, A., Flykt, A., & Esteves, F. (2001). Emotion drives
attention: Detecting the snake in the grass. Journal
of Experimental Psychology: General, 130, 466–478.
https://doi.org/10.1037/0096-3445.130.3.466, PubMed:
11561921
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
Palmer, J. A., Kreutz-Delgado, K., & Makeig, S. (2012). AMICA:
An adaptive mixture of independent component analyzers
with shared components. Swartz Center for Computatonal
Neursoscience, University of California San Diego (Technical
Report).
Popkirov, S., Staab, J. P., & Stone, J. (2018). Persistent
postural-perceptual dizziness (PPPD): A common,
characteristic and treatable cause of chronic dizziness.
Practical Neurology, 18, 5–13. https://doi.org/10.1136
/practneurol-2017-001809, PubMed: 29208729
Rodriguez, R., & Crane, B. T. (2021). Effect of timing delay
between visual and vestibular stimuli on heading perception.
Journal of Neurophysiology, 126, 304–312. https://doi.org/10
.1152/jn.00351.2020, PubMed: 34191637
Sakowitz, O. W., Quiroga, R. Q., Schürmann, M., & Başar, E.
(2001). Bisensory stimulation increases gamma-responses
over multiple cortical regions. Cognitive Brain Research, 11,
267–279. https://doi.org/10.1016/S0926-6410(00)00081-1,
PubMed: 11275488
Senkowski, D., Schneider, T. R., Foxe, J. J., & Engel, A. K.
(2008). Crossmodal binding through neural coherence:
Implications for multisensory processing. Trends in
Neurosciences, 31, 401–409. https://doi.org/10.1016/j.tins
.2008.05.002, PubMed: 18602171
Senkowski, D., Talsma, D., Grigutsch, M., Herrmann, C. S., &
Woldorff, M. G. (2007). Good times for multisensory
integration: Effects of the precision of temporal synchrony as
revealed by gamma-band oscillations. Neuropsychologia, 45,
561–571. https://doi.org/10.1016/j.neuropsychologia.2006.01
.013, PubMed: 16542688
Shattuck, D. W., Chiang, M.-C., Barysheva, M., McMahon, K. L.,
de Zubicaray, G. I., Meredith, M., et al. (2008). Visualization
tools for high angular resolution diffusion imaging. In
Proceedings of the medical image computing and
computer-assisted intervention. MICCAI International
Conference Medical Image Computing Computer-Assisted
Intervention (Vol. 5242, pp. 298–305). https://doi.org/10
.1007/978-3-540-85990-1_36
Shayman, C. S., Seo, J. H., Oh, Y., Lewis, R. F., Peterka, R. J.,
& Hullar, T. E. (2018). Relationship between vestibular
sensitivity and multisensory temporal integration. Journal of
Neurophysiology, 120, 1572–1577. https://doi.org/10.1152/jn
.00379.2018, PubMed: 30020839
Sheppard, J. P., Raposo, D., & Churchland, A. K. (2013).
Dynamic weighting of multisensory stimuli shapes decision-
making in rats and humans. Journal of Vision, 13, 4. https://
doi.org/10.1167/13.6.4, PubMed: 23658374
Siegel, M., Donner, T. H., & Engel, A. K. (2012). Spectral
fingerprints of large-scale neuronal interactions. Nature
Reviews Neuroscience, 13, 121–134. https://doi.org/10.1038
/nrn3137, PubMed: 22233726
Singer, W., & Gray, C. M. (1995). Visual feature integration and
the temporal correlation hypothesis. Annual Review of
Neuroscience, 18, 555–586. https://doi.org/10.1146/annurev
.ne.18.030195.003011, PubMed: 7605074
Slutsky, D. A., & Recanzone, G. H. (2001). Temporal and spatial
dependency of the ventriloquism effect. NeuroReport, 12,
7–10. https://doi.org/10.1097/00001756-200101220-00009,
PubMed: 11201094
Townsend, B., Legere, J. K., von Mohrenschildt, M., & Shedden,
J. M. (2022). Beta-band power is an index of multisensory
weighting during self-motion perception. Neuroimage:
Reports, 2, 100102. https://doi.org/10.1016/j.ynirp.2022
.100102
Townsend, B., Legere, J. K., O’Malley, S., von Mohrenschildt, M.,
& Shedden, J. M. (2019). Attention modulates event-related
spectral power in multisensory self-motion perception.
Neuroimage, 191, 68–80. https://doi.org/10.1016/j
.neuroimage.2019.02.015, PubMed: 30738208
Van Ombergen, A., Van Rompaey, V., Maes, L. K., Van de
Heyning, P. H., & Wuyts, F. L. (2016). Mal de debarquement
syndrome: A systematic review. Journal of Neurology, 263,
843–854. https://doi.org/10.1007/s00415-015-7962-6, PubMed:
26559820
Walsh, E. G. (1961). Role of the vestibular apparatus in the
perception of motion on a parallel swing. Journal of
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
1106
Journal of Cognitive Neuroscience
Volume 35, Number 7
Physiology, 155, 506–513. https://doi.org/10.1113/jphysiol
.1961.sp006643, PubMed: 13782902
Weech, S., & Troje, N. F. (2017). Vection latency is reduced by
bone-conducted vibration and noisy galvanic vestibular
stimulation. Multisensory Research, 30, 65–90. https://doi.org
/10.1163/22134808-00002545
Yakubovich, S., Israeli-Korn, S., Halperin, O., Yahalom, G.,
Hassin-Baer, S., & Zaidel, A. (2020). Visual self-motion cues
are impaired yet overweighted during visual–vestibular
integration in Parkinson’s disease. Brain Communications,
2, fcaa035. https://doi.org/10.1093/braincomms/fcaa035,
PubMed: 32954293
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
0
9
2
2
1
4
0
2
4
0
/
/
j
o
c
n
_
a
_
0
1
9
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Townsend et al.
1107