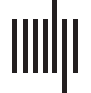RESEARCH ARTICLE
Cortical Tracking of Continuous Speech
Under Bimodal Divided Attention
Zilong Xie1*
, Christian Brodbeck2*
, and Bharath Chandrasekaran3
1School of Communication Science and Disorders, Florida State University, Tallahassee, FL, USA
2Department of Psychological Sciences, University of Connecticut, Storrs, CT, USA
3Department of Communication Science and Disorders, Università di Pittsburgh, Pittsburgh, PAPÀ, USA
*Joint first author.
Keywords: acoustic processing, continuous speech, crossmodal, divided attention, EEG, linguistico
processing
ABSTRACT
Speech processing often occurs amid competing inputs from other modalities, Per esempio,
listening to the radio while driving. We examined the extent to which dividing attention between
auditory and visual modalities (bimodal divided attention) impacts neural processing of
natural continuous speech from acoustic to linguistic levels of representation. We recorded
electroencephalographic (EEG) responses when human participants performed a challenging
primary visual task, imposing low or high cognitive load while listening to audiobook stories as
a secondary task. The two dual-task conditions were contrasted with an auditory single-task
condition in which participants attended to stories while ignoring visual stimuli. Behaviorally, IL
high load dual-task condition was associated with lower speech comprehension accuracy
relative to the other two conditions. We fitted multivariate temporal response function encoding
models to predict EEG responses from acoustic and linguistic speech features at different
representation levels, including auditory spectrograms and information-theoretic models of
sublexical-, word-form-, and sentence-level representations. Neural tracking of most acoustic and
linguistic features remained unchanged with increasing dual-task load, despite unambiguous
behavioral and neural evidence of the high load dual-task condition being more demanding.
Compared to the auditory single-task condition, dual-task conditions selectively reduced neural
tracking of only some acoustic and linguistic features, mainly at latencies >200 ms, while earlier
latencies were surprisingly unaffected. These findings indicate that behavioral effects of bimodal
divided attention on continuous speech processing occur not because of impaired early sensory
representations but likely at later cognitive processing stages. Crossmodal attention-related
mechanisms may not be uniform across different speech processing levels.
INTRODUCTION
Speech processing often occurs amid competing inputs from other sensory modalities, for
esempio, listening to the radio while driving. In such situations, listeners must allocate atten-
tion across modalities to effectively select the most relevant information within a modality.
This raises the question of whether and how dividing attention between modalities (per esempio., audi-
tion and vision; bimodal divided attention) affects the processing of natural continuous speech.
Resource-based theoretical frameworks have been invoked to scaffold the understanding of
mechanisms governing crossmodal attention (Wahn & König, 2017). Two contrastive resource-
a n o p e n a c c e s s
j o u r n a l
Citation: Xie, Z., Brodbeck, C., &
Chandrasekaran, B. (2023). Cortical
tracking of continuous speech
under bimodal divided attention.
Neurobiology of Language, 4(2),
318–343. https://doi.org/10.1162
/nol_a_00100
DOI:
https://doi.org/10.1162/nol_a_00100
Received: 1 novembre 2022
Accepted: 11 Gennaio 2023
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Zilong Xie
zx22c@fsu.edu
Bharath Chandrasekaran
b.chandra@pitt.edu
Handling Editor:
Jonathan Peelle
Copyright: © 2023
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
based accounts (modality-specific vs. supramodal) yield different hypotheses regarding the
effects of bimodal divided attention on continuous speech processing. Per the modality-
specific account, each sensory modality is allocated a limited pool of attentional resources,
and these pools of attentional resources operate independently of each other (Alais et al.,
2006; Arrighi et al., 2011; Duncan et al., 1997; Keitel et al., 2013; Parks et al., 2011; Porcu
et al., 2014). In contrasto, per the supramodal account, different sensory modalities share a cen-
tral, limited pool of attentional resources. The availability of resources to one modality is
inversely related to the amount of resources used by other modalities (Broadbent, 1958;
Ciaramitaro et al., 2017; Klemen et al., 2009; Macdonald & Lavie, 2011; Molloy et al., 2015).
Empirical evidence regarding bimodal divided attention effects on speech processing primarily
comes from experimenter-constrained tasks (per esempio., Gennari et al., 2018; Kasper et al., 2014; Mattys
et al., 2009, 2014; Mattys & Palmer, 2015; Mattys & Wiget, 2011). Many studies have shown the
detrimental effects of bimodal divided attention on the acoustic processing of simplified, controlled
speech stimuli (per esempio., syllable or single words; Gennari et al., 2018; Mattys et al., 2014; Mattys &
Palmer, 2015; Mattys & Wiget, 2011), which is consistent with the supramodal account of attention.
Speech processing entails mapping acoustic features into linguistic representations of increasing
complexity (Brodbeck & Simone, 2020; Hickok & Poeppel, 2007), raising the question of how
bimodal divided attention affects linguistic representations beyond acoustic processing. Behavioral
studies with simple speech stimuli indicate that reduced acoustic processing under bimodal divided
attention may lead to compensatory changes manifested by increased reliance on higher-order
linguistic knowledge during auditory lexical perception (Mattys et al., 2009). Tuttavia, to date,
there is a lack of a systematic and holistic analysis of divided attention-related changes across
different levels (acoustic to linguistic) of natural continuous speech processing, which is distinctly
different from processing simple speech stimuli (Gaston et al., 2023; Hamilton & Huth, 2020).
Here, we assessed electroencephalography (EEG) to provide a systematic and holistic anal-
ysis of the acoustic and linguistic processing of continuous speech (Brodbeck & Simone, 2020;
Gillis et al., 2022). The continuous speech paradigm uses the multivariate temporal response
function approach (Crosse et al., 2016; Ding & Simone, 2012) to predict neural responses from
a combination of hypothesis-driven acoustic and linguistic properties of continuous speech.
The predictive power of each speech property is used to quantify the corresponding processing
levels (Brodbeck & Simone, 2020; Gillis et al., 2022). The spectro-temporal acoustic properties
included an acoustic spectrogram and an acoustic onset spectrogram. The linguistic properties
included measures of informativeness (surprisal and entropy) based on the information-
theoretic framework (Brodbeck et al., 2018). Prior work suggests that both acoustic and
linguistic representations are strongly modulated by selective attention, within the auditory
modality and across modalities. Attentional effects are disproportionality more robust on the
linguistic representations than acoustic-based representations (Brodbeck et al., 2018, 2020).
Here we integrated the continuous speech paradigm with an audiovisual dual-task para-
digm to examine the effects of bimodal divided attention on the acoustic and linguistic pro-
cessing of continuous speech. In the dual-task paradigm, participants performed a challenging
primary visuospatial task that imposed low or high cognitive load while listening to audiobook
stories as a secondary task. The two dual-task conditions were contrasted with an auditory
single-task condition in which participants attended to the story while ignoring visual stimuli.
We hypothesized that compared to the auditory single-task condition, dual-task conditions
would lead to reduced acoustic and linguistic representations of continuous speech, particolarmente
at high cognitive load. Tuttavia, we hypothesized that linguistic representations may be
affected to a relatively greater extent based on evidence from the literature on selective atten-
zione. These hypotheses are aligned with the supramodal account of crossmodal attention.
Neurobiology of Language
319
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
MATERIALS AND METHODS
Experimental Design
Bimodal divided attention was manipulated via a dual-task paradigm. Specifically, participants
performed a primary visuospatial n-back task of varying (high or low) cognitive load (Jaeggi
et al., 2007) while listening to continuous speech as a secondary task. We designated the
visual task as the primary task to maximize the chance of observing the bimodal divided atten-
tion effects on continuous speech processing. The cognitive load of the dual-task paradigm
was manipulated via 3- and 0-back tasks on the visuospatial stimuli ( blue squares;
Figure 1A and 1B). The dual-task conditions were contrasted with an auditory single-task
condition (Figure 1C), in which participants explicitly attended to the auditory stimuli while
ignoring the visual stimuli. To obtain a behavioral measure for the auditory task, participants
were instructed to respond to two multiple-choice comprehension questions on the story
segments at the end of each trial. Detailed task instructions are presented in the section on
Experimental Procedure.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Trial design. In the two dual-task conditions, the primary task was to respond to the visual stimuli and the secondary task was to
attend to auditory stimuli (story segments of about 60 S). (UN) In the high load condition, participants responded only when the current blue
square matched the one 3 positions back (examples highlighted in red squares). (B) In the low load condition, participants responded only
when the current blue square matched the first square in each trial (highlighted in the red square). (C) In the auditory single-task condition,
participants were instructed to attend to the auditory stimulus and ignore the visual stimuli. At the end of each trial, participants responded to
two multiple-choice comprehension questions for the story segments. ISI: interstimulus interval.
Neurobiology of Language
320
Continuous speech processing under bimodal divided attention
Each task condition consisted of 15 trials of visual stimuli paired with 15 unique story seg-
ments and was presented in a separate block. The order of the story segments was fixed and
identical across participants in order to maintain the continuity of the storyline. The order of
task conditions was counterbalanced across participants. Each trial of visual stimuli ended
later than the corresponding story segment. Such offset gaps were not significantly different
across task conditions, F(2, 42) = 0.01, p = 0.99. The experiment was controlled with E-Prime
Version 2.0.10 (Schneider et al., 2002).
Participants
Adult native American English speakers (N = 18) were recruited from the Austin, Texas, com-
munity. Data from one participant were excluded due to technical problems. Data from
another participant were excluded because their story comprehension accuracy was lower
for the auditory single-task condition (66.67%) than the two dual-task conditions (73.37%
for low load and 76.67% for high load). We interpreted this result as that this participant
did not understand or follow the task instructions. The final sample consisted of 16 partici-
pants (18 A 23 yr old; 11 females, 5 males; 14 right-handed, 2 left-handed). The sample size
was selected based on prior work examining the effects of bimodal attention on the neural
processing of speech stimuli (per esempio., Gennari et al., 2018; Kasper et al., 2014). Previous studies
have shown that music training can influence speech processing (per esempio., Bidelman & Alain,
2015). Therefore, we recruited only participants without significant formal music training
(≤four years of continuous training, not currently practicing). All participants had normal
air and bone-conduction audiometric thresholds, defined as ≤20 dB hearing level for octave
frequencies from 0.25 A 8 kHz. The thresholds were measured via an Equinox 2.0 PC-based
audiometer (Interacoustics, 2023). Additional inclusion criteria are as follows: no history of
psychological or neurological disorders, no use of neuropsychiatric medication, and having
normal or corrected-to-normal vision. Before the experiment, all participants provided
written, informed consent. Participants received monetary compensation for their participa-
zione. The Institutional Review Board at the University of Texas at Austin approved the exper-
imental protocols.
Stimuli and Apparatus
The stimuli were composed of visual and auditory materials. The visual stimuli (Figura 1) were
blue squares at one of eight loci around a white fixation cross in the center of a black screen,
adapted from Jaeggi et al. (2007). The duration for individual squares was 500 ms, and the
interval between consecutive squares was 2,500 ms. Twenty-three squares were included in
a trial, lasting 69 S. The stimuli were displayed on a VIEWPixx/EEG LCD monitor (VPixx Tech-
nologies, 2022) with a scanning LED-backlight design (29.1 cm [height] × 52.2 cm [width];
display resolution: 1920 × 1080; refresh rate: 120 Hz) at an ∼110 cm distance from partici-
pants’ eyes.
The auditory stimuli were English audiobook recordings selected from a classic work of
fiction, Alice’s Adventures in Wonderland (Carroll, 1865/2010, Chapters 1–7). The audiobook
was narrated by an adult male American English speaker at a sampling rate of 22.05 kHz. IL
chapters were divided into 45 segments (each ∼60 s long). Each segment began where the
story ended in the previous segment. In each segment, silent periods of more than 500 ms
were shortened to 500 ms. The story stimuli were presented diotically via insert earphones
(ER-3; Etymotic, 2023) to the participants at a 70 dB sound pressure level. A trial of visual
stimuli (23 blue squares) was presented concurrently with each story segment, with the
Neurobiology of Language
321
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
segment beginning later (3 s after the onset of the visual trial) and ending earlier relative to the
visual trial.
Experimental Procedure
High and low load dual-task
The cognitive load of the dual-task conditions was manipulated via the visual task. For the
high load condition, the visual task required participants to respond when the current blue
square matched the one three positions back in the sequence (cioè., 3-back task, Figure 1A).
For the low load condition, the visual task required participants to respond when the current
blue square matched the first square in the sequence (cioè., 0-back task, Figure 1B). We random-
ized the location of the first square across trials. Matched squares were treated as targets, E
unmatched ones were non-targets. Note that targets could appear only starting from the fourth
square in the sequence for a given trial in the 3-back task. In other words, targets would be
among the last 20 squares in the sequence on a given trial. We designed the 0-back task to
match that. Six of the last 20 squares were set as targets for both task conditions, and the
remaining 14 were non-targets. The target locations were randomized across trials.
Participants responded to the targets by pressing buttons on a game controller. Participants
were told that speed and accuracy were equally important. Participants were required to rest
their fixations on a white cross in the middle of the screen. To encourage engagement, accu-
racy feedback on the visual task was displayed after their responses. The number of button
presses was not significantly different between 3- and 0-back visual tasks (T (15) = 0.96, p =
0.36). After the visual task, participants responded to two multiple-choice comprehension
questions for the auditory stories. The questions were designed such that a superficial famil-
iarity with the story was insufficient for answering them (per esempio., “How many miles does Alice
think she has fallen?" (1) 2,000; (2) 3,000; (3) 4,000; (4) 5,000). Participants had unlimited time
to respond to the story questions. No feedback was provided after their responses.
Critically, to manipulate the priority of the auditory and visual tasks, participants were
instructed to focus primarily on the visual task and attend to the auditory stimulus as a second-
ary task. They were explicitly told that their data could not be used if their performance on the
visual task was poor.
Auditory single-task
In this condition, participants were instructed to focus on the story segments and ignore the
visual stimuli (Figure 1C). Participants were required to keep their eyes open and rest their
fixations on a white cross in the middle of the screen. At the end of each trial, participants
responded to two multiple-choice questions to assess their comprehension of the story seg-
menti. Participants had unlimited time to respond to questions. Visual feedback about the
accuracy of the story question was displayed following their responses.
Exit questionnaire
After participants completed the experiment, they completed a brief exit questionnaire related
to their familiarity with the auditory stimuli. Specifically, participants indicated if they had read
the Alice’s Adventures in Wonderland or watched a film adaptation, and if yes, approximately
how long ago. Participants also indicated to what extent reading the book or watching the
movie helped their task performance on a 10-point scale (1 = not at all, 10 = quite a bit). Four
out of 16 participants had read the book, E 10 out of 16 participants had watched the movie.
We examined whether reading (vs. not reading) the book and watching (vs. not watching) IL
Neurobiology of Language
322
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
movie modulate the effect of task condition (cioè., auditory-single task and dual-task conditions)
on behavioral auditory task (story) accuracy, using a three-way mixed analysis of variance
(ANOVA). We did not find significant main effects of reading the book (p = 0.14, ηp
2 = 0.17)
or watching the movie (p = 0.96, ηp
2 = 0.0003), or significant interactions between any vari-
ables (all ps > 0.37, ηp
2 < 0.08). These results suggest that prior familiarity with the auditory
stimuli did not appear to significantly contribute to task performance.
Electrophysiological Data Acquisition and Preprocessing
Acquisition
The experiment was conducted in a dark, acoustically shielded booth. Participants were
seated in a comfortable chair during tasks. EEG data were recorded using the Easycap
(2023) recording cap with 64 actiCAP active electrodes (Brain Products, 2023a) at a sampling
rate of 5 kHz. The electrode locations were determined according to the extended 10–20 sys-
tem (Oostenveld & Praamstra, 2001), with a common ground at the Fpz electrode site and TP9
as the reference. The electrode impedances were below 20 kΩ.
The EEG data were acquired using BrainVision actiCHAmp amplifier linked to BrainVision
Pycorder software Version 1.0.7 (Brain Products, 2023b).
Preprocessing
The EEG data were preprocessed offline in MNE-Python (Gramfort et al., 2013), and the
deconvolution analysis was implemented with the Eelbrain package (Brodbeck et al., 2021).
The data were re-referenced to the average of the electrodes TP9 and TP10, and then band-
pass filtered from 1 to 15 Hz using a zero-phase overlap-add finite impulse response filter
(hamming window) with default settings in MNE-Python. Independent component analysis
was applied to EEG data combined across the three task conditions in individual participants
using the extended-infomax algorithm. Artifact-related components (mainly ocular artifacts)
were identified according to the topographical distribution and time course via visual inspec-
tion and then removed. After that, the EEG data were segmented into epochs that were time-
locked to the onsets of corresponding story segments, and then downsampled to 100 Hz. The
maximum possible duration of the epochs was set to 61 s.
Assessing Neural Tracking of Visual and Auditory Stimuli
To assess the neural representation of speech, we used the multivariate temporal response
function (mTRF) approach (Crosse et al., 2016; Ding & Simon, 2012). In this approach, the
EEG signal is predicted using time-delayed multiple regression. We first generated several
visual, acoustic, and linguistic models (see below for detailed descriptions). Each model
was used to define several predictor variables, each implementing a specific linking hypoth-
esis for predicting brain activity from the corresponding model. We then tested the predictive
power of different combinations of predictor variables to evaluate which acoustic and linguis-
tic models are associated with predictive power for the EEG data. Each predictor variable thus
operationalizes a hypothesis that EEG responses are modulated by a given property of the
speech signal, which would indicate neural representations arising from a corresponding
acoustic or linguistic model. Figure 2 displays an example of each predictor variable.
Visual model
Because the visual stimuli were temporally sparse, visual responses were modeled analo-
gously to a visual evoked potential. The visual predictor was a one-dimensional time
Neurobiology of Language
323
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
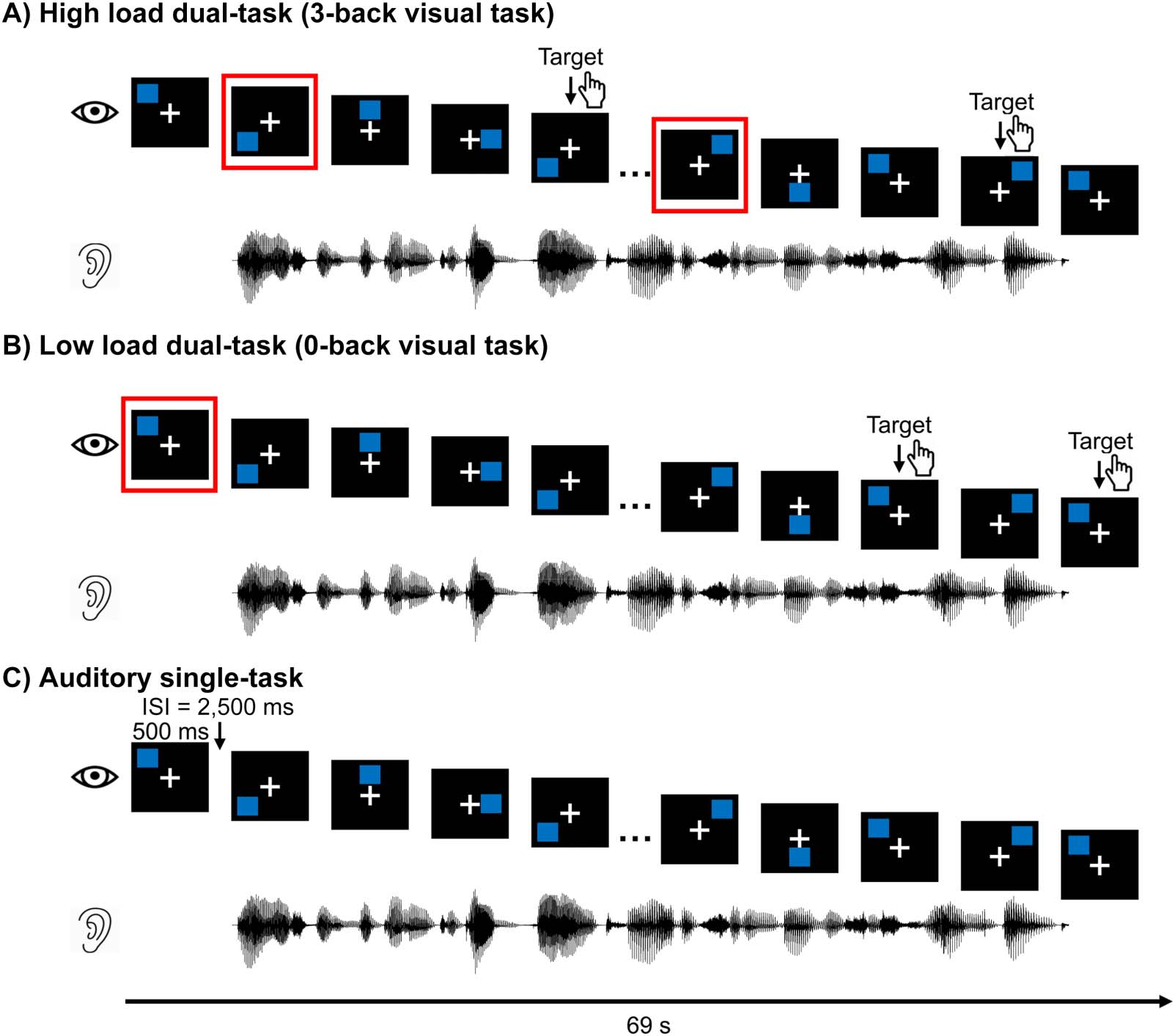
Figure 2. An excerpt of raw electroencephalography (EEG) responses from all 64 electrodes (top row) and the predictor variables (subsequent
rows) used to model the EEG responses. Note that visual predictors consist of a separate one-dimensional array with impulses for onsets and
offsets of the blue squares. They are combined into a single predictor in this example for illustration purposes.
series with an impulse at the onsets and offsets of the blue squares. We did not separate pre-
dictors for targets and non-targets because this study was not intended to explore differences in
neural processing of visual targets and non-targets, and thus there were not enough targets to
estimate stable responses.
Neurobiology of Language
324
Continuous speech processing under bimodal divided attention
Acoustic model
The acoustic model was designed to assess EEG responses related to representations of acous-
tic spectro-temporal features. All acoustic predictors were derived from 256-band gammatone-
based spectrograms of the speech stimuli, with cut-off frequencies from 0.02 to 5 kHz. The
256-band spectrograms were downsampled to 1 kHz and log transformed using the formula
log(1 + x), where x refers to the downsampled spectrograms. A spectrogram predictor was then
created by summing the 256-band spectrograms in eight logarithmically spaced frequency
bands. In addition, an onset spectrogram predictor was defined to detect and control for
representations of acoustic edges. These were generated using an auditory edge detection
model (Brodbeck et al., 2020; Fishbach et al., 2001) and applied to each frequency band of
the 256-band spectrograms. The onsets across these 256 bands were also summed into eight
logarithmically spaced frequency bands to generate 8-band onset spectrogram predictors.
Linguistic models
Linguistic processing was assessed using information-theoretic models. These models assume
that listeners maintain predictive models of speech, which can be linked to brain activity
through surprisal and entropy measures (Brodbeck et al., 2018). Previous work suggests that
listeners maintain multiple such predictive models, differing in complexity, in parallel
(Brodbeck et al., 2022). The predictive models were all defined over phoneme sequences,
determined for each stimulus via forced alignment using the Montreal Forced Aligner (MFA;
McAuliffe et al., 2017). The predictors based on the specific information-theoretic models
(described in subsequent sections) all consisted of time series with an impulse of variable size
at each phoneme onset. In order to provide a control for responses related to linguistic
segmentation, two additional predictors were included: A word onset predictor with a unit
(value of 1) impulse at the onset of each word-initial phoneme and a phoneme onset predictor
with a unit impulse at the onset of all other phonemes.
Sublexical model. The sublexical model assumes that listeners predict upcoming phonemes or
speech sounds based on a local context, consisting of only a few preceding sounds. To imple-
ment such a model, all sentences from the SUBTLEX-US corpus (Keuleers et al., 2010) were
transcribed into phoneme sequences without word boundary markers, and a 5-gram model
(Heafield, 2011) was trained on these phoneme sequences. This model was then applied to
the experimental stimuli to compute a probability distribution over phonemes at each pho-
neme position, conditional on the four preceding phonemes. This distribution was used to
calculate a sublexical surprisal predictor (the surprisal of encountering phoneme phk at posi-
tion k in the stimulus is −log2 (p(phk|context)) and a sublexical entropy predictor (the entropy
at phoneme position phk reflects the uncertainty about what the next phoneme, phk+1, will be
−
p(phk+1 = ph|context)log2 p(phk+1 = ph|context )). Surprisal is a measure of the
amount of new information provided by a stimulus; a response to sublexical surprisal is thus
evidence that listeners integrate information on a sublexical level. A response to entropy addi-
tionally suggests that listeners create a probabilistic expectation about future input before
encountering the phoneme (Pickering & Gambi, 2018). A response to either of those predictors
would provide evidence that listeners maintain a sublexical language model.
Pphonemes
ph
Word-form model. The word-form model aims to predict the surface form of the word that is
currently being heard, but without access to any information preceding the word. To imple-
ment this model, a phonological lexicon was generated by combining pronunciations from the
MFA English dictionary and the Carnegie Mellon University Pronouncing Dictionary (https://
Neurobiology of Language
325
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
www.speech.cs.cmu.edu/cgi-bin/cmudict). The word-form model was implemented based on
the cohort model of word recognition (Brodbeck et al., 2018; Marslen-Wilson, 1987). Each
word was assigned a prior probability based on its count frequency in the SUBTLEX corpus
(Keuleers et al., 2010), substituting a count of 1 for missing words. For each word in the speech
stimuli, the cohort model was then implemented by starting with the complete lexicon and, for
each subsequent phoneme of the word, incrementally removing words that were not compat-
ible with that phoneme in that position. The changing probability distribution over the lexicon
was then used to define two predictors, each with a value at each phoneme position: phoneme
surprisal (log inverse of the posterior probability of the phoneme given the preceding pho-
nemes) and cohort entropy (Shannon entropy over all words currently in the cohort,
Plexicon
word p(word|context)log2 p(word|context). This model implements the hypothesis that lis-
teners recognize words using a probabilistic model that takes into account all the information
since the last word boundary (i.e., where the word started), but that does not further take into
account any context when considering possible word forms as candidates.
Sentence model. The sentence model is very similar to the word-form model, with the only
difference being that the prior expectation for each word is modulated by the sentence con-
text. To implement this, a lexical 5-gram model (Heafield, 2011) was trained on the whole
SUBTLEX-US database (Keuleers et al., 2010). This 5-gram model was used to set the prior
probability for each word in the lexicon based on the preceding four words before applying
the procedure described for the word-form model above. The same two linking hypotheses
were used to define predictor variables (phoneme surprisal and cohort entropy). The sentence
model implements the hypothesis that listeners use a wider context including multiple words,
when modulating their phoneme-by-phoneme perception and expectations.
Estimation of neural tracking
We used forward encoding mTRF models to predict EEG responses from the predictors
described above. The mTRF models were fitted to the EEG responses at individual electrodes
using the boosting algorithm implemented in the Eelbrain toolbox (Brodbeck et al., 2021). The
predictive power of the mTRF models was evaluated by how accurately they could predict
EEG responses from novel trials of the same condition. This was quantified through the
z-transformed Pearson’s correlation coefficient between predicted and actual EEG responses
(i.e., prediction accuracy). Higher prediction accuracy indicates better neural tracking of the
corresponding predictor.
The mTRFs were estimated separately for each subject and condition using a fivefold cross-
validation strategy. First, the trials were divided into five test sets. For each test set, EEG
responses were predicted from the average of four mTRF models, estimated from the remaining
four data sets (among those 4 data sets, each served as validation set once and as training data
during the remaining 3 fits, thus leading to 4 different mTRF models). Thus, predictions of EEG
responses for each test set were made with mTRF models that were estimated without ever
using that test set. The mTRFs were generated from a basis of 50 ms Hamming windows with
stimulus-EEG lag from −100 to 500 ms (window center). The mTRFs were estimated jointly for
all predictors with coordinate descent to minimize the ‘1 error. After each step, the change in
error was evaluated in the validation set, and if there was an increase in error, the TRF for the
predictor responsible for the change was frozen (in its state before the change). This continued
until the whole mTRF was frozen. A single measure of prediction accuracy (Fisher
z-transformed correlation between predicted and measured response, see above) was
Neurobiology of Language
326
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
calculated after concatenating the predicted responses from the five test sets. Because predic-
tions were based on cross-validation, the expected correlation by chance is z = 0. For analysis
of the response functions, the mTRFs were averaged across all the test sets. For the visual pre-
dictor, the TRFs to onsets and offsets were combined for an effective response function with
lags from −100 to 1,000 ms relative to visual stimulus onset (because the visual stimulus
always lasted exactly 500 ms).
To estimate the neural tracking of a given predictor (or a combination of predictors), we
calculated the change in prediction accuracy (i.e., Δz) when the predictor(s) of interest was
(were) removed from the full model that included all the visual, acoustic, and linguistic pre-
dictors. This procedure tests for variability in the responses that can be uniquely attributed to
the predictor(s) under investigation and cannot be explained by any other predictors. Such a
strong test is warranted because different properties of natural, connected speech tend to be
correlated in time. Note that the analysis of the mTRFs themselves could not implement such
strict control, and thus we cannot exclude the possibility that response functions include com-
ponents that are confounded with other, correlated speech features. For this reason, we focus
our interpretation on tests of predictive power more than mTRF comparisons.
Statistical Analysis
All statistical analyses, if unspecified, were implemented in R ( Version 4.2.1; R Core Team,
2022).
First, we examined the effect of task condition (auditory single-task, or low or high load
dual-task) on behavioral performance, and neural visual, acoustic, and linguistic processing
separately. A paired t test (two-sided), or one- or two-way repeated-measures ANOVA, which-
ever was appropriate, was performed with an alpha level of 0.05. The reported p values of
those analyses were adjusted using the false discovery rate (FDR) method (Benjamini &
Hochberg, 1995). We also calculated effect sizes (Cohen’s d for t tests and partial eta squared
(ηp
2) for ANOVAs) and Bayes factors (BF). The BFs were computed using appropriate functions
from the R package “BayesFactor” ( Version 0.9.12.4.4; Morey et al., 2022). Post hoc analysis,
if necessary, was performed using paired t tests (two-sided). FDR-corrected p values, effect
sizes (Cohen’s d ), and BFs were reported. More analysis details are provided in the following
paragraphs.
Behavioral performance was quantified by three measures, including the accuracy and
reaction time (RT) for the visual task and the accuracy for the auditory task. Visual accuracy
was calculated as the difference in hit rates (i.e., correctly responding to a target) and false
alarm rates (i.e., identifying a non-target as being a target). Visual RT was calculated as the
median RT for hits only (Jaeggi et al., 2007; Snodgrass & Corwin, 1988). Auditory accuracy
was calculated as the percentage of correctly answered story questions.
The extent of neural visual processing was determined using a mass-univariate analysis,
comparing the predictive power (z) between the full model and a model missing the visual
predictor. For this, we averaged the prediction accuracy for visual predictors across task con-
ditions at individual electrodes and tested whether the averaged difference in prediction accu-
racy (Δz) was greater than zero using a mass-univariate, one-sample t test (one-sided). This
was implemented in the Eelbrain package. The mass-univariate test was a cluster-based per-
mutation test, using a t value equivalent to uncorrected p ≤ 0.05 as the cluster-forming thresh-
old. A corrected p value was computed for each cluster based on the cluster-mass statistic in a
null distribution from 10,000 permutations (or a complete set of all possible permutations, in
cases where this was fewer than 10,000; Maris & Oostenveld, 2007). We reported the largest
Neurobiology of Language
327
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
t value from the cluster (i.e., tmax) as an estimate of effect size (Brodbeck et al., 2018). Neural
acoustic and linguistic processing were analyzed in the same manner.
We followed each of these analyses by examining the extent to which task conditions mod-
ulated neural tracking of individual predictors, or subsets of predictors. To this end we used the
significant clusters from the mass-univariate analysis as region of interest (ROI) to extract Δz
values averaged across the electrodes in the cluster, but for each condition separately. Regard-
ing neural acoustic processing, we examined the spectrogram and onset spectrogram predic-
tors separately. Regarding neural linguistic processing, we conducted three sets of analyses to
examine individual linguistic predictors. First, a two-way repeated-measures ANOVA was per-
formed to examine the effects of context levels (sublexical, word-form, and sentence) and task
condition on prediction accuracy. Second, a two-way repeated-measures ANOVA was per-
formed to examine the effects of predictor type (entropy and surprisal) and task condition
on prediction accuracy. Third, a one-way repeated-measures ANOVA was performed to
examine the effect of task condition on the prediction accuracy of word onsets. Further, if a
significant effect of task condition was observed from any of those analyses, we conducted
follow-up analyses to examine whether task conditions eliminated neural tracking of the cor-
responding predictor(s) by testing whether the prediction accuracy at individual task condi-
tions was greater than zero using one-sample t tests (one-sided).
Finally, we examined the effect of task conditions on the mTRFs from predictors which
showed significant task condition effects on prediction accuracy. The predictors include visual
predictors, onset spectrogram, three context levels (sublexical, word-form, and sentence), and
two predictor types (entropy and surprisal). We calculated the global field power (GFP) of
mTRFs across the corresponding ROI from the above analyses of prediction accuracy. We then
compared the GFP of mTRFs between task conditions using mass-univariate paired t tests (two-
sided). The mTRF analyses were implemented in the Eelbrain package with default parameters
except for the analysis time window. For visual predictors, we concatenated the mTRFs for
visual onsets and offsets to analyze the response to visual stimuli as a whole. For the onset
spectrogram, we averaged the mTRFs across the eight frequency bands. The analysis time
window was 0 to 1,000 ms for visual predictors and 0 to 450 ms for auditory predictors.
RESULTS
Table 1 summarizes the key findings regarding the effect of task condition on behavioral
performance, and neural visual, acoustic, and linguistic processing.
Divided Attention and Visual Load Impair Behavioral Performance
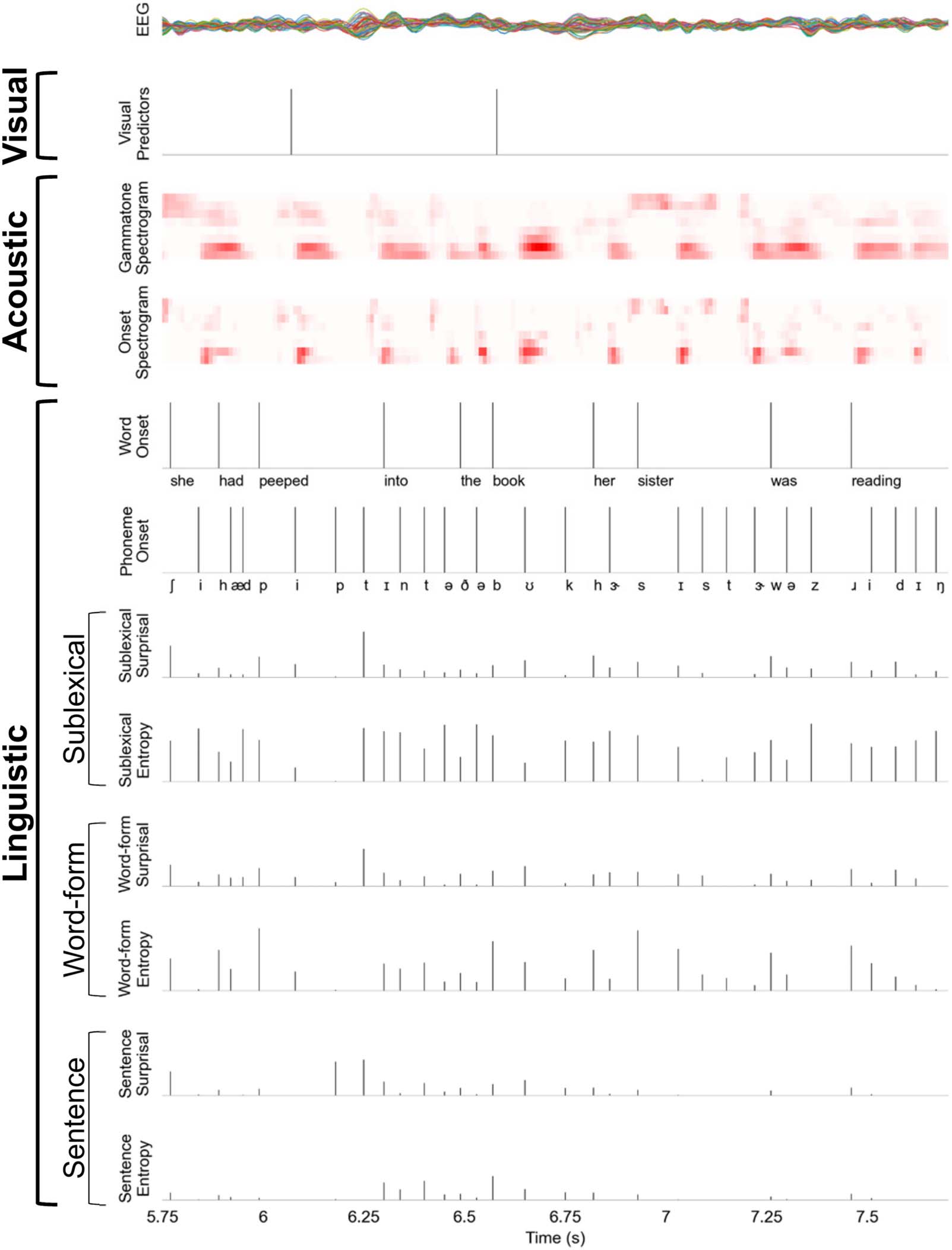
Figure 3A and 3B display the accuracy and RT of the visual task for individual participants.
Compared to the low load (0-back) condition, the high load (3-back) condition was associated
with lower accuracy (low load: mean = 99.54% (SD = 0.82) vs. high load: mean = 63.31%
(SD = 21.85), t(15) = 6.60, p < 0.001, Cohen’s d = 1.65, BF = 2.59 × 103) and slower RT (low
load: mean = 453.11 ms (SD = 67.54) vs. high load: mean = 785.24 ms (SD = 233.41), t(15) =
−5.33, p < 0.001, Cohen’s d = 1.33, BF = 330.30). These results confirmed that the manipu-
lation of cognitive load in the visual task was successful.
Figure 3C displays the auditory task accuracy for individual participants. The mean accu-
racy was 88.96% (SD = 5.93) in the auditory single-task condition, 84.58% (SD = 11.86) in the
low load dual-task condition, and 65.63% (SD = 12.75) in the high load dual-task condition.
The effect of task condition was significant (F(2, 30) = 36.59, p < 0.001; ηp
2 = 0.71, BF = 6.75 ×
106). Post hoc analysis revealed that auditory task accuracy was significantly lower in the high
Neurobiology of Language
328
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
Type
Behavioral
Table 1.
Task effects on continuous speech processing
Measure
Key result
Visual accuracy
Visual reaction time
Auditory accuracy
Low load > High load
Low load < High load Auditory single-task = Low load > High load
Neural (Δz)
Visual
Acoustic
Auditory single-task < Low load < High load Gammatone spectrogram No significant task effect Onset spectrogram Auditory single-task > Low load = High load
Linguistic
Sublexical, word-form,
and sentence context
Auditory single-task > Low load = High load
Entropy and surprisal
Entropy: Auditory single-task > Low load = High load
Surprisal: Auditory single-task > Low load = High load
Word onset
No significant task effect
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
load dual-task condition compared to the other two conditions: versus auditory single-task,
T(15) = 7.38, P < 0.001, Cohen’s d = 1.84, BF = 8.31 × 103; and versus low load dual-task,
t(15) = 6.34, p < 0.001, Cohen’s d = 1.58, BF = 1.70 × 103. The auditory task accuracy was not
significantly different between auditory single-task and low load dual-task conditions (t(15) =
1.75, p = 0.10, Cohen’s d = 0.44, BF = 0.88).
Further, we examined the relationship between visual and auditory task performance dur-
ing the dual-task conditions. The change in auditory accuracy (i.e., (low load − high load)/low
load) was negatively correlated with the change in visual RT (i.e., (high load − low load)/low
load) (Spearman’s ρ = −0.46, uncorrected p = 0.038, one-sided; Figure 3D), such that listeners
who slowed down more on the visual task from low to high load conditions tended to have a
smaller drop in auditory accuracy. The change in auditory accuracy was not significantly
correlated with the change in visual accuracy (Spearman’s ρ = −0.29, uncorrected p =
0.28, one-sided).
These results demonstrate that divided (vs. selective) attention and increasing visual load
impair behavioral visual and auditory performance.
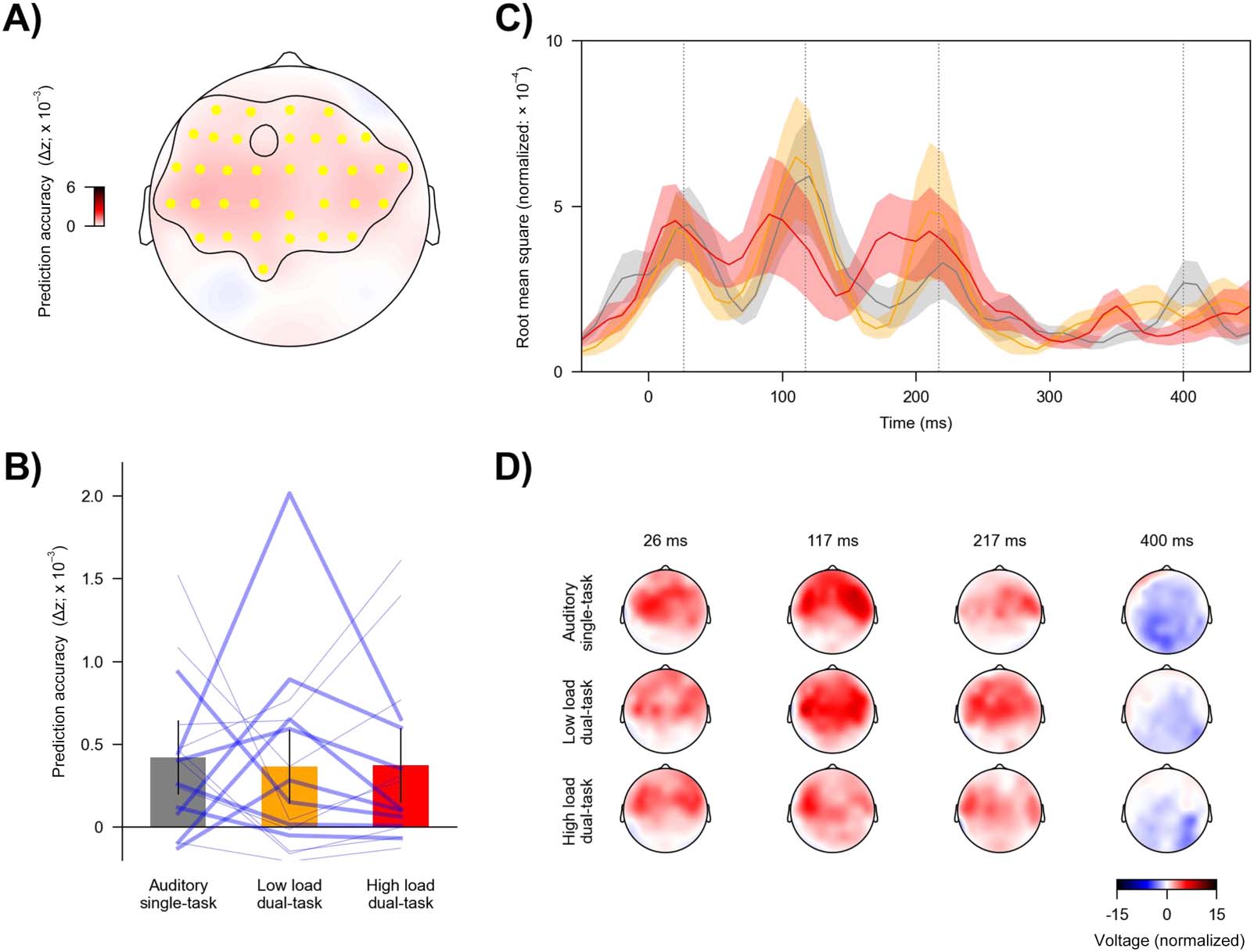
Neural Tracking of Visual Stimuli Is Strongly Modulated by Divided Attention and Visual Load
To assess neural tracking of visual stimuli, we focused on the predictive power of visual pre-
dictors while controlling for all speech-related predictors (acoustic and linguistic). Adding pre-
dictors for visual stimuli to a model including only speech predictors significantly improved its
predictive power (prediction accuracy averaged across task conditions; tmax = 12.93, p <
0.001), providing evidence for neural tracking of visual stimuli. The cluster-based test resulted
in a single significant cluster that spread across all electrodes, with the largest effects on pari-
etal and occipital electrodes (Figure 4A).
Neurobiology of Language
329
Continuous speech processing under bimodal divided attention
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Behavioral performance on visual and auditory tasks. (A) Accuracy on the low load
(0-back) and high load (3-back) visual tasks, which was calculated as the difference in hit rates
(i.e., correctly responding to a target) and false alarm rates (i.e., identifying a non-target as being
a target). (B) Reaction time on the low load (0-back) and high load (3-back) visual tasks, which was
calculated for hits only. (C) Accuracy on the auditory task, which was calculated as the percentage
of correctly answered story questions. Individual lines in A–C denote individual participants (n =
16). (D) Correlation between the change in auditory accuracy (i.e., (low load − high load)/low load)
and the change in visual RT (i.e., (high load − low load)/low load). The gray line is the linear regres-
sion line. n.s. (not significant): p > 0.05, *** P < 0.001.
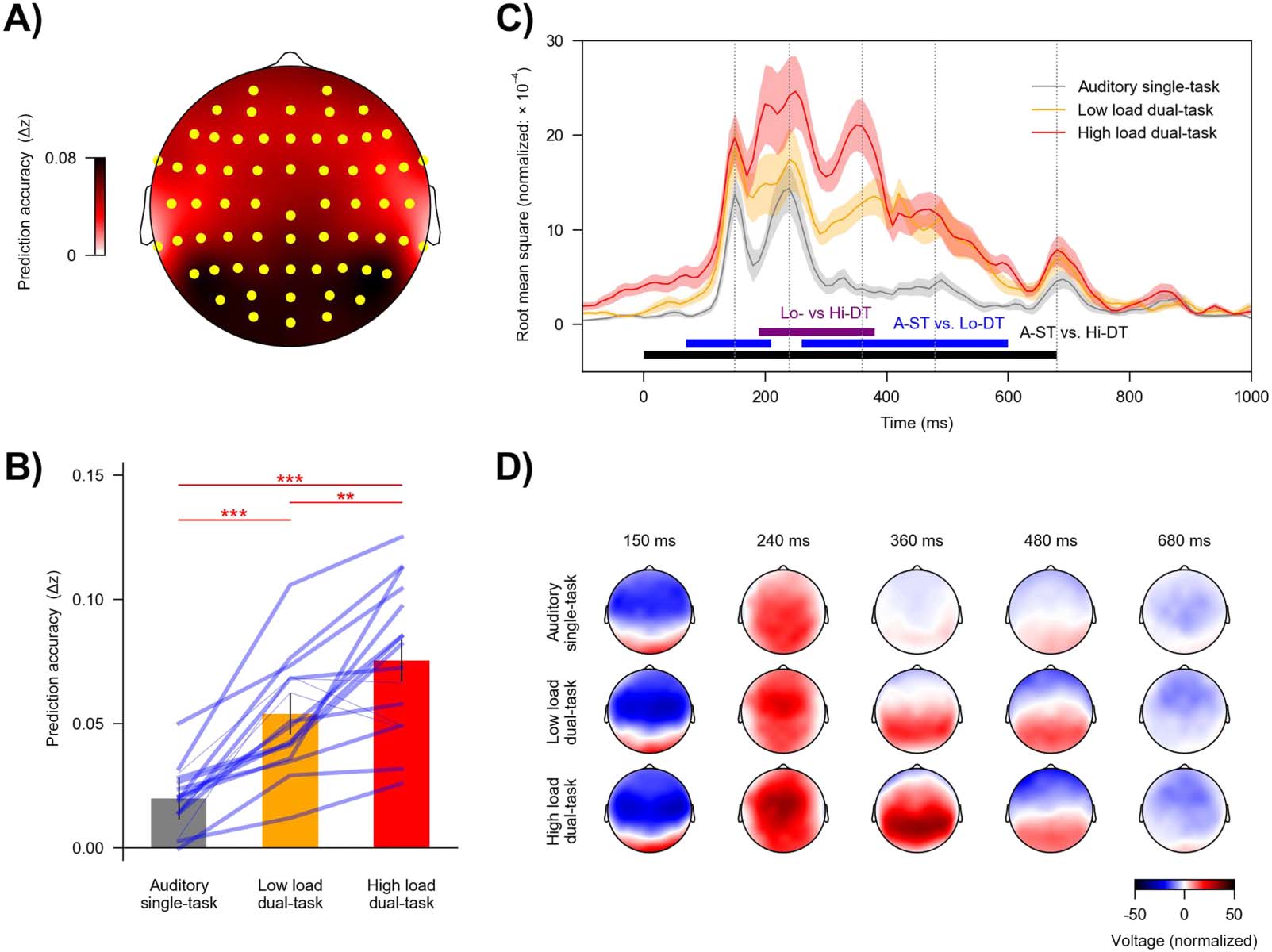
Importantly, the predictive power of the visual predictors was modulated by task condition
(F(2, 30) = 46.10, p < 0.001, ηp
2 = 0.76, BF = 6.09 × 107). As shown in Figure 4B, the high load
dual-task condition was associated with the highest predictive power (mean = 0.075, SD =
0.029), followed by the low load dual-task condition (mean = 0.053, SD = 0.022), and lowest
for the auditory single-task condition (mean = 0.020, SD = 0.012): high load dual-task versus
auditory single-task, t(15) = 9.52, p < 0.001, Cohen’s d = 2.38, BF = 1.50 × 105; high load
dual-task versus low load dual-task, t(15) = 3.34, p = 0.005, Cohen’s d = 0.84, BF = 10.7; low
load dual-task versus auditory single-task, t(15) = 6.64, p < 0.001, Cohen’s d = 1.66, BF =
2.72 × 103. Together, these results suggest that neural tracking of visual stimuli was succes-
sively enhanced with increasing load of the visual task.
We analyzed mTRFs to further clarify how the difference in model predictive power was
reflected in brain responses. Visual mTRFs can be conceptualized as evoked responses to the
visual stimuli. Consistent with results for prediction accuracy, the mTRFs were also modulated
by the task condition (Figure 4C). The high load dual-task condition showed larger mTRF
amplitudes than the auditory single-task condition from 0 to 680 ms (p < 0.001) and the
low load dual-task condition from 190 to 380 ms (p < 0.001). The mTRF amplitudes for the
Neurobiology of Language
330
Continuous speech processing under bimodal divided attention
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Neural tracking of visual stimuli across task conditions. Visual stimuli were associated with a robust response, which further
increased with task relevance and load. (A) Topography showing the increase in prediction accuracy (Δz) due to visual predictors, which
was significantly above zero in a single cluster encompassing the highlighted (yellow) electrodes. (B) Prediction accuracy across task condi-
tions. Blue lines denote individual participants: Thicker lines indicate higher prediction accuracy for the high vs. low load condition, and
thinner lines indicate lower accuracy for the high versus low load condition. Red asterisks denote p values for comparison between conditions.
Error bars denote the 95% within-subject confidence interval (Loftus & Masson, 1994). (C) Global field power of the visual temporal response
functions (TRFs). Visual stimuli lasted from 0 to 500 ms. Shaded areas denote within-subject standard errors around the mean (for color labels
see panel B). Horizontal lines denote time windows in which TRFs were significantly different between conditions. (D) Topographies of
selected times in panel C (grey vertical lines). A-ST: auditory single-task, Lo-DT: low load dual-task, Hi-DT: high load dual-task. ** p <
0.01, *** p < 0.001.
low load dual-task condition were larger than the auditory single-task condition from 70 to
210 ms (p = 0.009) and from 260 to 600 ms (p < 0.001).
Divided Attention, but Not Visual Load, Reduces Late Neural Tracking of Acoustic Features
The acoustic predictors significantly contributed to model prediction beyond the visual and
linguistic predictors in a cluster that spread across almost all electrodes, with maxima at tem-
poral sites (tmax = 12.00, p < 0.001; Figure 5A). As expected, these results provide evidence for
robust neural tracking of acoustic properties of speech.
The prediction accuracy for acoustic predictors was modulated by task condition (F(2, 30) =
14.83, p < 0.001, ηp
2 = 0.50, BF = 581.38; Figure 5B). Post hoc analysis showed that the pre-
diction accuracy significantly dropped in the two dual-task conditions compared to the audi-
tory single-task condition (vs. low load dual-task, t(15) = 3.84, p = 0.002, Cohen’s d = 0.96,
BF = 25.60; vs. high load dual-task, t(15) = 4.78, p < 0.001, Cohen’s d = 1.20, BF = 130.60).
The prediction accuracy was not significantly different between the dual-task conditions
(t(15) = 0.77, p = 0.45, Cohen’s d = 0.19, BF = 0.33). These results suggest that neural tracking
Neurobiology of Language
331
Continuous speech processing under bimodal divided attention
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
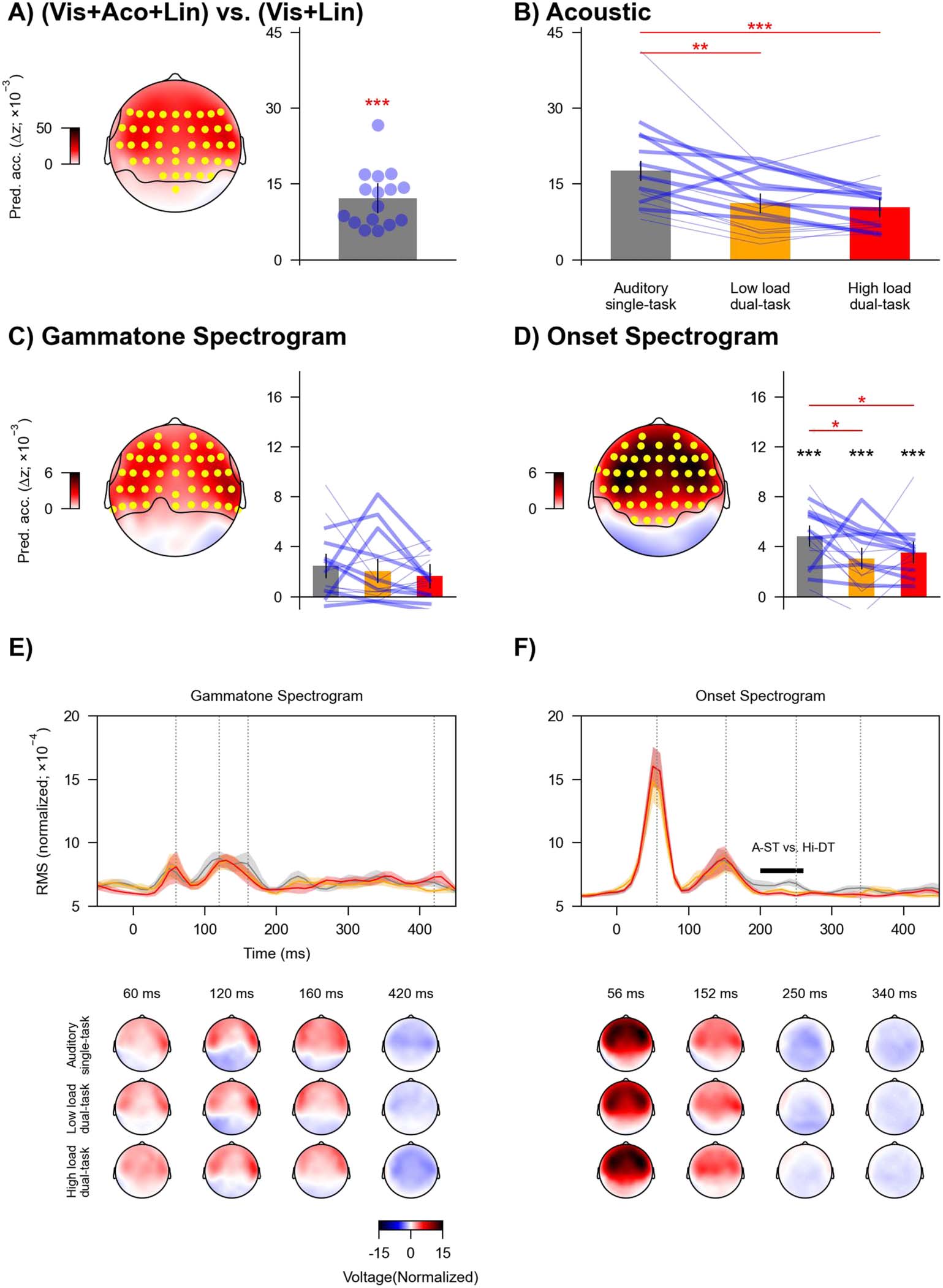
Figure 5. Neural tracking of acoustic information across task conditions. (A) Increase in prediction accuracy (Δz) due to acoustic predictors
of speech (gammatone and onset spectrogram), which was significantly above zero in a cluster encompassing the highlighted (yellow) elec-
trodes. Blue dots denote individual participants. (B) Prediction accuracy across task conditions for acoustic predictors, i.e., combined gam-
matone and onset spectrogram. (C) and (D) Prediction accuracy across task conditions for gammatone spectrogram and onset spectrogram
separately. Topographies highlight electrodes (yellow) with prediction accuracy that was significantly above zero. Black asterisks denote
p values for testing against (above) zero at individual conditions. In B–D, blue lines denote individual participants: Thicker lines indicate lower
accuracy for high vs. low load condition, and thinner lines indicate higher accuracy for high vs. low load condition. Red asterisks denote
p values for comparison between conditions. Error bars denote 95% confidence interval. (E) and (F) GFP (top) of mTRFs and related topog-
raphies (bottom) for gammatone and onset spectrogram. The mTRFs were averaged across the eight frequency bands. Shaded areas denote
within-subject standard errors around the mean. Horizontal lines denote time windows in which mTRFs were significantly different between
conditions. Topographies are shown for selected times indicated in GFPs (grey vertical lines). * p < 0.05, ** p < 0.01, *** p < 0.001.
Neurobiology of Language
332
Continuous speech processing under bimodal divided attention
of acoustic information was reduced when directing attention from one task (auditory) to two
tasks (visual and auditory).
Then, we assessed whether the effect of task condition could be attributed to specific acous-
tic predictors. The two acoustic predictors both independently contributed to overall model
prediction (gammatone spectrogram: tmax = 6.08, p < 0.001, Figure 5C; onset spectrogram:
tmax = 9.91, p < 0.001, Figure 5D). The effect of task condition on the prediction accuracy
was significant for onset spectrogram (F(2, 30) = 4.93, p = 0.033, ηp
2 = 0.25, BF = 4.17) but
not for gammatone spectrogram (F(2, 30) = 0.70, p = 0.59, ηp
2 = 0.04, BF = 0.26). Post hoc
analysis revealed that the prediction accuracy for onset spectrogram significantly dropped in
the two dual-task conditions compared to the auditory single-task condition (vs. low load
dual-task, t(15) = 2.61, p = 0.030, Cohen’s d = 0.65, BF = 3.14; vs. high load dual-task,
t(15) = 2.89, p = 0.030, Cohen’s d = 0.72, BF = 4.94). The prediction accuracy was not
significantly different between the dual-task conditions (t(15) = −0.79, p = 0.44, Cohen’s
d = 0.20, BF = 0.34).
Considering the modulation by task condition, we further examined whether divided atten-
tion eliminated neural tracking of onset spectrogram. The prediction accuracy at individual
task conditions was significantly above zero (all uncorrected ps < 0.001, Cohen’s d > 1.20,
BF > 256.40; Figure 5D), suggesting that directing attention from one task to two tasks did not
eliminate the neural tracking of acoustic onsets.
Finalmente, we examined the effect of task condition on the mTRFs for the onset spectrogram
(Figure 5F). mTRFs to a continuous stimulus like the auditory spectrogram can be conceived of
as evoked responses to an elementary event in the stimulus (cioè., the impulse response). IL
mTRF amplitudes in the auditory single-task condition were larger compared to the high load
dual-task condition from 200 A 260 ms (p = 0.003). Further, a visual inspection of the mTRFs
from individual subjects revealed two relatively reliable peaks at about 56 (P1) E 152
(P2) ms. Latencies of these peaks were not significantly different across conditions (56 ms:
F(2, 30) = 0.65, uncorrected p = 0.94, ηp
2 = 0.004; 152 ms: F(2, 30) = 0.62, uncorrected
p = 0.54, ηp
2 = 0.04).
In sum, acoustic tracking was very similar across conditions, with only a slight reduction in
the tracking of acoustic onsets in the divided attention tasks, compared to the single task.
This difference was likely explained by a reduction in a relatively late response component,
starting at 200 ms.
Divided Attention, but Not Visual Load, Reduces Late Tracking of Linguistic Information
The linguistic predictors significantly contributed to model prediction beyond the visual and
acoustic predictors (tmax = 4.95, P < 0.001; Figure 6A). The cluster-based test indicated that the
effect of linguistic predictors was primarily observed for temporal-central electrodes. These
results provide evidence for neural tracking of linguistic properties of speech.
The prediction accuracy for linguistic predictors was modulated by task condition (F(1.41,
21.15) = 6.66, p = 0.029, ηp
2 = 0.31, BF = 10.82; Figure 6B). The prediction accuracy signif-
icantly dropped in the two dual-task conditions compared to the auditory single-task condition
(vs. low load dual-task, t(15) = 2.83, p = 0.029, Cohen’s d = 0.71, BF = 4.49; vs. high load
dual-task, t(15) = 2.61, p = 0.029, Cohen’s d = 0.65, BF = 3.16). The prediction accuracy was
not significantly different between the two dual-task conditions (t(15) = −1.80, p = 0.091,
Cohen’s d = 0.45, BF = 0.95). These results suggest that neural tracking of linguistic informa-
tion was reduced when directing attention from one task to two tasks.
Neurobiology of Language
333
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
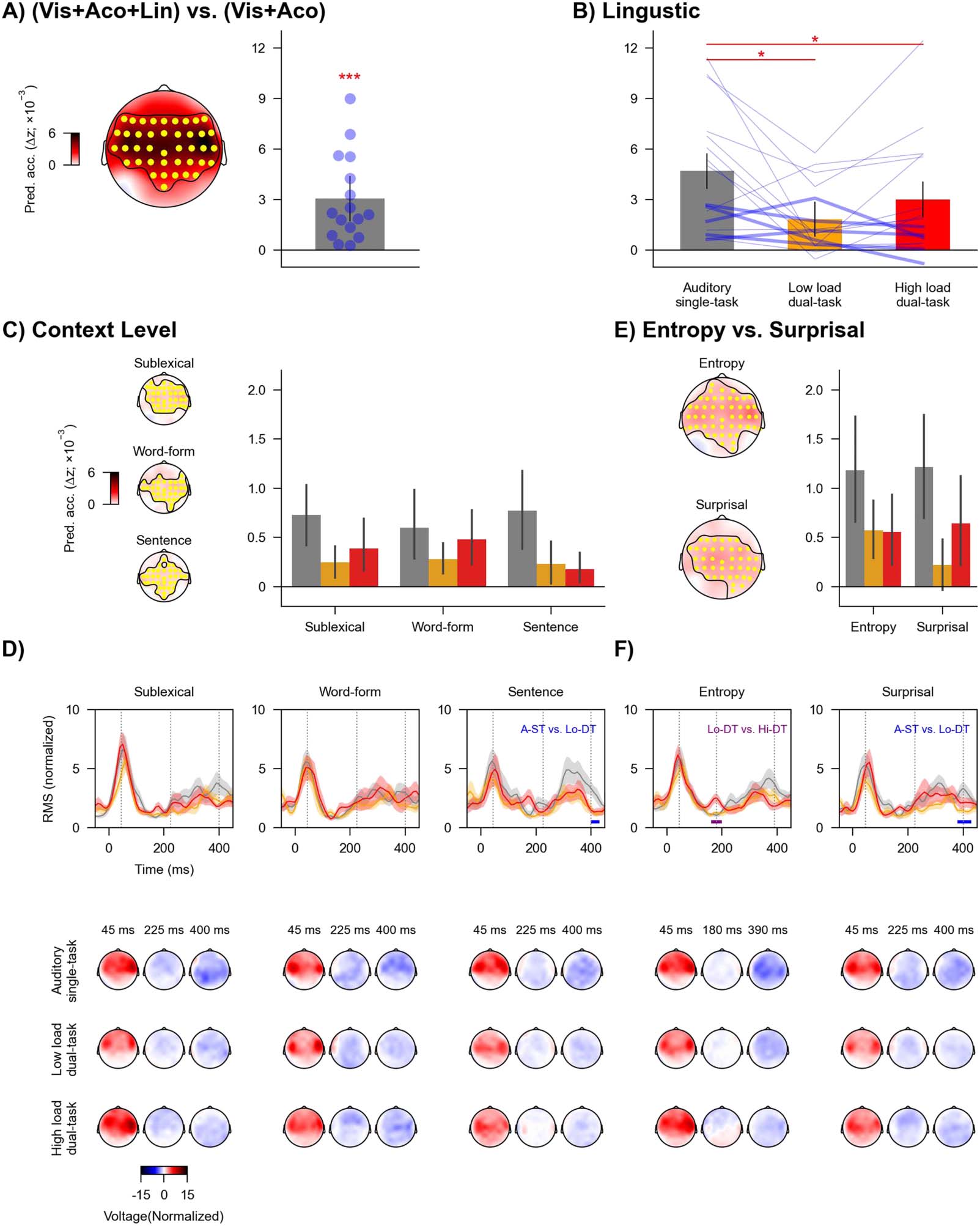
Figure 6. Neural tracking of linguistic information across task conditions. (A) Increase in prediction accuracy (Δz) due to linguistic predictors
of speech (word onsets, phoneme onsets, sublexical surprisal and entropy, word-form surprisal and entropy, and sentence surprisal and
entropy), which was significantly above zero across highlighted (yellow) electrodes in the topography. Blue dots denote individual participants.
(B) Prediction accuracy for combined linguistic predictors across conditions. Blue lines denote individual participants: Thicker lines indicate
lower accuracy for high versus low load condition, and thinner lines indicate higher accuracy for high versus low load condition. Red asterisks
denote p values for comparison between conditions. (C) Prediction accuracy for three context levels (sublexical, word-form, and sentence)
across conditions. Each level includes entropy and surprisal predictors. (D) Global field power (top) of mTRFs and related topographies (bot-
tom) for each context level. The mTRF GFPs were averaged across entropy and surprisal. (E) Prediction accuracy for entropy and surprisal.
Each predictor includes the three context levels. (F) GFP of mTRFs and related topographies for entropy and surprisal. In B, C, and E, error bars
denote 95% confidence interval. In D and F, shaded areas denote standard errors around the mean. Horizontal lines denote time windows in
which the mTRFs were significantly different between task conditions. Topographies are shown for selected times indicated in GFPs (grey
vertical lines). * p < 0.05, *** p < 0.001.
Neurobiology of Language
334
Continuous speech processing under bimodal divided attention
Next, we conducted three sets of analyses to assess whether the effect of task condition
could be attributed to specific linguistic properties.
Task effects appear to be similar across different context levels
The first analysis focused on the three context levels (sublexical, word-form, and sentence).
Each level independently contributed significantly to model prediction (sublexical: tmax =
5.22, p < 0.001; word-form: tmax = 3.92, p < 0.001; sentence: tmax = 4.98, p < 0.001;
Figure 6C). A two-way repeated-measures ANOVA showed that the interaction between con-
text level and task condition was not significant (F(2.55, 38.18) = 1.19, p = 0.40, ηp
2 = 0.073,
BF = 0.19). The main effect of context level was not significant (F(2, 30) = 0.32, p = 0.77, ηp
2 =
0.021, BF = 0.08). But the main effect of task condition was significant (F(1.2, 18.01) = 8.46,
p = 0.021, ηp
2 = 0.36, BF = 1.40 × 103). Post hoc analysis showed that the prediction accuracy
was significantly reduced from the auditory single-task condition to the low load (t(15) = 2.90,
p = 0.016, Cohen’s d = 0.73, BF = 5.08) and high load (t(15) = 4.27, p = 0.002, Cohen’s d =
1.07, BF = 54.63) dual-task conditions. But the prediction accuracy was not significantly dif-
ferent between the low and high load dual-task condition (t(15) = 1.02, p = 0.32, Cohen’s d =
0.26, BF = 0.40). Further, we found similar patterns of results when restricting the two-way
repeated-measures ANOVA analysis to the dual-task conditions. In sum, patterns of task con-
dition effects observed for linguistic predictors appeared to be similar across the different
linguistic models.
Considering the modulation by context level and task condition, we further examined
whether divided attention eliminated neural tracking of any of these predictors. The prediction
accuracies for all predictors at individual task conditions were significantly above zero (all
uncorrected ps < 0.03, Cohen’s d > 0.53, BF > 1.51).
Regarding mTRFs, the effect of task condition was not significant for sublexical or
word-form context but was for sentence context (Figure 6D). The mTRF amplitude of sentence
context in the auditory single-task condition was larger compared to the low load dual-task
condition from 400 A 430 ms (p = 0.036). Topographies suggest that this is due to a broadly
distributed more negative component in the single task condition.
Initial response peaks to linguistic features appear relatively early. This is consistent with
previous results (Brodbeck et al., 2022) and might be partly because forced alignment, Quale
was used to determine phoneme timing, does not take into account coarticulation effects.
Some information about upcoming phonetic features might thus have systematically preceded
the estimates of phoneme onset times we used.
Neural tracking of surprisal might increase with visual load
The second analysis focused on entropy and surprisal. The two predictors independently con-
tributed significantly to model prediction (entropy: tmax = 5.51, P < 0.001; surprisal: tmax =
3.91, p = 0.001; Figure 6E). A two-way repeated-measures ANOVA showed that the interac-
tion between predictor type (entropy vs. surprisal) and task condition was not significant
(F(1.32, 19.86) = 1.29, p = 0.40, ηp
2 = 0.079, BF = 0.31). The main effect of predictor type
was not significant (F(1, 15) = 0.31, p = 0.65, ηp
2 = 0.02, BF = 0.23). But the main effect of task
condition was significant (F(1.35, 20.2) = 9.85, p = 0.011, ηp
2 = 0.40, BF = 890.10). Post hoc
analysis showed that, when averaging across surprisal and entropy, the prediction accuracy
was significantly reduced from the auditory single-task condition to the low load (t(15) =
3.28, p = 0.008, Cohen’s d = 0.82, BF = 9.56) and high load dual-task conditions (t(15) = 4.11,
p = 0.003, Cohen’s d = 1.03, BF = 40.91). Numerically, the prediction accuracy was improved
Neurobiology of Language
335
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
from the low load to high load dual-task condition, but this difference was not significant (t(15) =
1.27, p = 0.22, Cohen’s d = 0.32, BF = 0.51).
Because of theoretical predictions of enhanced reliance on linguistic representations during
higher visual task load (see Introduction and Discussion), we further restricted the two-way
repeated-measures ANOVA to the dual-task conditions. The interaction between predictor
type and task condition was significant (F(1, 15) = 5.75, uncorrected p = 0.03 (FDR-corrected
p = 0.063), ηp
2 = 0.28, BF = 1.23). There was no significant main effect of predictor type
(F(1, 15) = 1.92, p = 0.33, ηp
2 = 0.11, BF = 0.40) or task condition (F(1, 15) = 1.62, p = 0.36,
ηp
2 = 0.097, BF = 0.79). Post hoc analysis showed that for entropy, the prediction accuracy
was not different between the dual-task conditions (t(15) = 0.10, p = 0.92, Cohen’s d =
0.03, BF = 0.26). But for surprisal, the prediction accuracy was significantly improved from
the low load to high load dual-task condition (t(15) = 2.20, uncorrected p = 0.044, Cohen’s
d = 0.55, BF = 1.66).
Considering the modulation by predictor type and task condition, we further examined
whether divided attention eliminated neural tracking of entropy or surprisal. The prediction
accuracies for both predictors at individual task conditions were significantly above zero
(all uncorrected ps < 0.01, Cohen’s d > 0.66, BF > 3.36), except for the surprisal predictors
at the low load dual-task condition (uncorrected p = 0.059, Cohen’s d = 0.41, BF = 0.78).
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 7. Neural tracking of word onsets across task conditions. (UN) Topography showing the increase in prediction accuracy (Δz) due to
word onsets, which was significantly above zero across highlighted (yellow) electrodes. (B) Prediction accuracy across conditions. Blue lines
denote individual participants: Thicker lines indicate higher accuracy for the high vs. low load condition, and thinner lines indicate lower
accuracy for the high vs low load condition. Error bars denote 95% within-subject confidence interval (Loftus & Masson, 1994). (C) Globale
field power of mTRFs. Shaded areas denote within-subject standard error around the mean. (D) Topographies of selected times in panel C (grigio
vertical lines).
Neurobiology of Language
336
Continuous speech processing under bimodal divided attention
Regarding mTRFs, the mTRF amplitude of entropy (Figure 6F) in the low load dual-task
condition was smaller than the high load dual-task condition from 160 A 200 ms (uncorrected
p = 0.037). The mTRF amplitude of surprisal in the low load dual-task condition was smaller
compared to the auditory single-task condition from 380 A 430 ms (uncorrected p = 0.012).
We did not observe a significant effect of task load, although the mTRF to surprisal during high
visual load was numerically stronger than low load from 200 ms onward.
Divided attention or visual load does not affect neural tracking of word onsets
The third set of analysis focused on word onset. This predictor independently contributed
significantly to model prediction (tmax = 4.51, P < 0.001; Figure 7A). But the effect of task
condition on prediction accuracy was not significant (F(2, 30) = 0.07, p = 0.93, ηp
2 = 0.005,
BF = 0.17; Figure 7B).
Taken together, results suggest that directing attention from one task to two tasks may reduce
but does not eliminate the neural tracking of linguistic features of speech. However, increasing
visual load does not lead to a further reduction. On the contrary, an increasing load of the dual
task might even be associated with enhanced neural tracking of phoneme surprisal. However,
this effect should be interpreted with care because the effect was not significant when analyzing
all linguistic predictors as a group or after correction for multiple comparisons.
DISCUSSION
We examined the extent to which bimodal divided attention influences acoustic and linguistic
representations of natural continuous speech. Compared to unimodal auditory speech
processing, the visual tasks affected acoustic onsets (Figure 5D and 5F; but not the acoustic
spectrogram, Figure 5C and 5E) and linguistic representations related to predictive processing
(Figure 6C to 6F; but not to lexical segmentation, Figure 7). Surprisingly, we did not find evi-
dence of further reduction (at any processing level) with increased visual (dual) task load,
despite unambiguous behavioral and neural evidence for the high load task being more
demanding (Figure 3 and Figure 4).
Locus of Effects of Bimodal Divided Attention on Continuous Speech Processing
We noted a potential dissociation between the impact of the dual-task on behavioral perfor-
mance in the speech comprehension task and a relative lack of impact on neural speech pro-
cessing. Behaviorally, the dual-task load clearly impacted listeners’ ability to answer auditory
comprehension questions. However, neural tracking of acoustic and linguistic speech features
was affected only at late response components and remained largely unchanged with varying
dual-task load. This neural and behavioral dissociation suggests that bimodal divided attention
largely impacts only late, post-perceptual processes of continuous speech processing. The sig-
nificant and unchanged responses related to predictive processing using the sentence context
suggest that listeners could track multi-word sequences regardless of dual-task load. We posit
that the decreased behavioral performance originates from higher-order cognitive processes
that are not adequately described by probabilistic word-sequence models, such as semantic
integration and memory formation.
Our hypothesis that the dual-task interference to continuous speech processing originates
from higher-order processes is consistent with the load theory of attention by Lavie and
colleagues (Lavie, 2005; Lavie & Tsal, 1994). The load theory proposes a distinction between
perceptual load and cognitive load (load on cognitive control, e.g., working memory).
Compared with perceptual load, cognitive load influences only later stages of stimulus
Neurobiology of Language
337
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
n
o
_
a
_
0
0
1
0
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Continuous speech processing under bimodal divided attention
processing, e.g., memory or behavioral responses (Lavie, 2005; Murphy et al., 2016; however,
see for example Sörqvist et al., 2012). Our dual-task paradigm manipulated the visual working
memory load and thus can be considered as a form of cognitive load.
Previous behavioral research has suggested that increased dual-task load is associated with
reduced acoustic sensitivity during speech recognition (Mattys et al., 2014; Mattys & Palmer,
2015; Mattys & Wiget, 2011). In our data, the dual-task did not alter early acoustic responses
and only had subtle effects on later (>200 ms) responses (see Figure 5E and 5F). This suggests
that the effect of bimodal divided attention may not be on basic acoustic representations
per se, but on secondary acoustic analysis stages or on how these representations are accessed
and used by higher-order processes.
Implications for Resource-Based Accounts
A common framework for understanding effects under dual-task paradigms is resource based
(Wahn & König, 2017). When two tasks draw from a limited pool of shared resources,
increased load in one task is associated with poorer performance in another task. Such a
hypothesis is often referred to as the supramodal account of crossmodal attention (Broadbent,
1958; Ciaramitaro et al., 2017; Klemen et al., 2009; Macdonald & Lavie, 2011; Molloy et al.,
2015). In contrasto, if the increased load in one task does not affect a corresponding decrease in
another, the two modalities can draw on separate resource pools. Such a hypothesis is con-
sistent with a modality-specific account of crossmodal attention (Alais et al., 2006; Arrighi
et al., 2011; Duncan et al., 1997; Keitel et al., 2013; Parks et al., 2011; Porcu et al., 2014).
Here, we observed a reduction in neural tracking of speech acoustic and linguistic features
under bimodal divided attention, consistent with previous studies demonstrating detrimental
effects of bimodal divided attention for simplified speech stimuli such as syllables (Gennari
et al., 2018), parole (Kasper et al., 2014), and sentences (Salo et al., 2015). In conjunction with
the co-occurring improved neural tracking of visual stimuli, this finding appears to suggest a
tradeoff between attending to the auditory versus visual modalities. Hence, these results
appear to align with the supramodal hypothesis of the dual-task effects that the auditory
and visual tasks of our study draw on a limited pool of shared resources (Broadbent, 1958;
Ciaramitaro et al., 2017; Klemen et al., 2009; Macdonald & Lavie, 2011; Molloy et al., 2015).
Tuttavia, a supramodal hypothesis of the dual-task effects does not seem to fit other key
results from our study. Primo, the impact of bimodal divided attention is specific to certain
features of the speech signals: we found bimodal attention effects for acoustic onsets but
not auditory spectrogram representations, and for predictive linguistic processing, indexed
through information-theoretic variables, but not lexical segmentation, indexed through the
word-onset predictors. In each case, the impact of divided attention is not a generally reduced
representation but is restricted to only specific response components in the response time
courses (the mTRFs).
Inoltre, a resource-based account would suggest that when the visual load is further
increased, available resources for speech representations should further decrease, which is not
what we observed. Invece, adding a visual task exacted a cost on neural speech representa-
zioni, but this cost did not scale with the task load. In contrast to these neural effects, task load
did affect behavioral performance on the auditory task. These divergent results may require an
explanation involving different resource pools (Wahn & König, 2017). Per esempio, there may
be a resource pool for sensory processing, which is sensitive to divided attention but not task
load, and thus is relatively modality-specific. There may be a second resource pool, che è
Neurobiology of Language
338
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
sensitive to task load and affects higher-order story comprehension, and thus is relatively
supramodal.
Selective Versus Divided Attention on Speech Processing
Previous studies on continuous speech processing have shown that selective attention within
and between modalities strongly modulates neural processing of both acoustic and linguistic
features of continuous speech, and the attentional effects seem to be even stronger for linguis-
tic processing (Brodbeck et al., 2018; Broderick et al., 2018; Ding et al., 2018; Kiremitçi et al.,
2021; Vanthornhout et al., 2019; Yahav & Golumbic, 2021). Specifically, neural tracking of
acoustic features is reduced and delayed but not eliminated for unattended speech, ma il
tracking of linguistic features is virtually abolished. A parsimonious null hypothesis, consistent
with the notion of a shared resource pool, is that speech representations during divided atten-
tion ought to be halfway between attended and ignored speech. Our results suggest that this is
not the case.
Primo, certain speech features (per esempio., acoustic spectrogram and word onsets) that have been
shown in prior work to be modulated by selective attention are insensitive to bimodal divided
Attenzione. Secondo, unlike prior work demonstrating differential selective attention effects on
the relative balance of acoustic versus linguistic processing, we did not observe a greater
reduction in linguistic processing than acoustic processing with the manipulation of divided
Attenzione. The neural tracking of both feature classes is reduced but not eliminated. Third, for
those features showing modulation by divided attention, we did not observe any delay in the
neural responses as reflected in the mTRFs (Figura 5 and Figure 6). Fourth, the effect of divided
attention emerged largely at later stages (after ∼200 ms) with the earlier latencies relatively
unaffected. Così, the effect of bimodal divided attention on neural continuous speech process-
ing appears to be feature-specific and occurs relatively late in processing.
These differences indicate that selective and divided attention are subserved by distinct
mechanisms. Relative to selective attention, bimodal divided attention tasks may be associated
with additional recruitment of frontal regions that interact with sensory cortices (Gennari et al.,
2018; Johnson & Zatorre, 2006; Loose et al., 2003). A stronger engagement of frontal regions
has been associated with poorer task performance (Gennari et al., 2018; Johnson & Zatorre,
2006). These neural findings appear to align with the argument that the costs of bimodal
divided attention may come from limitations of executive control to coordinate processes
related to two tasks rather than a competition for shared sensory resources (Katus & Eimer,
2019; Loose et al., 2003). The differential effects of selective and divided attention on contin-
uous speech processing suggest that the costs of selective attention are more likely to originate
from “filter” mechanisms (Broadbent, 1958; Lachter et al., 2004) that pass task-relevant signals
but block task-irrelevant others, rather than from the re-allocation of shared resources. Never-
theless, future studies are needed to elucidate mechanisms underlying differences in continu-
ous speech processing between selective and divided attention.
Increased Responses to Surprisal With Dual-Task Load
We found that increasing visual load increased responses to phoneme surprisal, but not
entropy. Statistically, this effect was seen only after excluding the auditory single-task condi-
tion and should thus be interpreted with care, but it is consistent with several extant findings.
The dissociation between entropy and surprisal is consistent with recent evidence that these
two processes may reflect different neural processes (Gaston et al., 2023). Neural responses
associated with surprisal may reflect prediction errors that signal the difference between
Neurobiology of Language
339
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
predicted and observed phonemes. Such prediction error signals may be boosted when atten-
tion is directed at the speech stimuli (per esempio., auditory single-task; Auksztulewicz & Friston, 2015;
Smout et al., 2019). Così, we may observe elevated responses to surprisal in the auditory
single-task condition compared with the dual-task conditions (Figure 6E). Further, IL
increased response to surprisal with dual-task load might reflect a shift toward more reliance
on linguistic representations during speech processing when resources for auditory processing
were constrained under divided attention of higher load (Mattys et al., 2009; Mattys & Wiget,
2011).
Neural Tracking of Word Onsets Was Not Affected by Divided Attention
Tracking of word onsets might reflect lexical segmentation (Sanders et al., 2002; Sanders &
Neville, 2003) E, along with other linguistic features, is strongly affected by selective atten-
zione (Brodbeck et al., 2018). It has been suggested that neural responses to word onsets reflect
the dynamic allocation of attention to time windows that contain word onsets (Astheimer &
Sanders, 2009). Tuttavia, our results indicate that tracking of word onsets is robust to manip-
ulations of attentional load by adding a visual task and increasing dual-task load. This suggests
that the word-onset attention effect may draw on a relatively unshared resource pool, or that
the word-onset responses reflect a more mechanistic aspect of lexical segmentation.
Study Limitations
One difference between the auditory single-task condition and the dual-task conditions is that
only the dual-task conditions required button-press responses (approximately 6 responses per
trial). Così, the additional button responses across conditions could be a potential confound
that may affect the comparison between the auditory single-task condition and the dual-task
conditions. Tuttavia, this difference is unlikely to be the cause of the observed results
(Tavolo 1). This is because artifacts from motor activity would be expected to cause a uniform
decrease in predictive power of the different stimulus representations in the dual-task condi-
tions compared to the auditory single-task condition. Invece, our data showed that the
decrease in predictive power in the dual-task (vs. auditory single-task) conditions was only
evident for some representations (per esempio., onset spectrogram, entropy, and surprisal, but not for
gammatone spectrogram and word onset).
The auditory stimulus material, Alice’s Adventures in Wonderland, is a relatively well-
known book. This raises the question whether prior knowledge of the auditory stimuli could
have made it easier for participants to answer the comprehension questions without paying
close attention to the story. This may affect how participants coordinate attention between
the auditory and visual tasks, thus contributing to task performance. Tuttavia, results on
the exit questionnaire suggest that prior familiarity with the auditory stimuli did not appear
to significantly contribute to behavioral performance on the auditory task. Further, as indicated
in the methods section, the questions were designed such that a superficial familiarity with the
auditory stimuli is insufficient for successful performance.
CONCLUSION
This study demonstrates a potential dissociation between the impact of dual-task load on
behavioral speech comprehension performance and a relative lack of impact on time-locked
neural representations of continuous speech. The behavioral effects of bimodal divided
attention on continuous speech processing occur not because of impaired early sensory rep-
resentations but likely at later cognitive processing stages.
Neurobiology of Language
340
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
ACKNOWLEDGMENTS
We thank Rachel Reetzke, Elise LeBovidge, and Jacie McHaney for their assistance with par-
ticipant recruitment and data collection. This work was supported by grants from the National
Institute on Deafness and Other Communication Disorders-National Institutes of Health and
the National Science Foundation. The content is solely the responsibility of the authors and
does not necessarily represent the official views of the National Institutes of Health or the
National Science Foundation.
FUNDING INFORMATION
Bharath Chandrasekaran, National Institute on Deafness and Other Communication Disorders
(https://dx.doi.org/10.13039/100000055), Award ID: R01DC013315. Christian Brodbeck,
National Science Foundation (https://dx.doi.org/10.13039/100000001), Award ID: BCS-
1754284.
AUTHOR CONTRIBUTIONS
Zilong Xie: Conceptualization: Lead; Data curation: Equal; Formal analysis: Equal; Investiga-
zione: Lead; Methodology: Lead; Project administration: Lead; Software: Equal; Supervision:
Lead; Validation: Lead; Visualization: Equal; Writing – original draft: Equal; Writing – review
& editing: Equal. Christian Brodbeck: Data curation: Equal; Formal analysis: Equal; Method-
ology: Equal; Software: Lead; Visualization: Equal; Writing – original draft: Equal; Writing –
revisione & editing: Equal. Bharath Chandrasekaran: Conceptualization: Equal; Data curation:
Supporting; Formal analysis: Supporting; Funding acquisition: Lead; Investigation: Supporting;
Methodology: Supporting; Project administration: Supporting; Resources: Lead; Software:
Supporting; Supervision: Lead; Writing – review & editing: Equal.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
.
/
l
REFERENCES
Alais, D., Morrone, C., & Burr, D. (2006). Separate attentional
resources for vision and audition. Proceedings of the Royal Soci-
ety B: Biological Sciences, 273(1592), 1339–1345. https://doi.org
/10.1098/rspb.2005.3420, PubMed: 16777721
Arrighi, R., Lunardi, R., & Burr, D. (2011). Vision and audition do
not share attentional resources in sustained tasks. Frontiers in
Psychology, 2, Article 56. https://doi.org/10.3389/fpsyg.2011
.00056, PubMed: 21734893
Astheimer, l. B., & Sanders, l. D. (2009). Listeners modulate tem-
porally selective attention during natural speech processing. Bio-
logical Psychology, 80(1), 23–34. https://doi.org/10.1016/j
.biopsycho.2008.01.015, PubMed: 18395316
Auksztulewicz, R., & Friston, K. (2015). Attentional enhancement
of auditory mismatch responses: A DCM/MEG study. Cerebral
Cortex, 25(11), 4273–4283. https://doi.org/10.1093/cercor
/bhu323, PubMed: 25596591
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society: Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Bidelman, G. M., & Alain, C. (2015). Musical training orchestrates
coordinated neuroplasticity in auditory brainstem and cortex to
counteract age-related declines in categorical vowel perception.
Journal of Neuroscience, 35(3), 1240–1249. https://doi.org/10
.1523/JNEUROSCI.3292-14.2015, PubMed: 25609638
Brain Products. (2023UN). ActiCAP [Electrodes]. https://www
.brainproducts.com
Brain Products. (2023B). ActiCHAmp and Pycoder ( Version 1.0.7)
[Amplifier and software]. https://www.brainproducts.com
Broadbent, D. E. (1958). Perception and communication. Pergamon
Press. https://doi.org/10.1037/10037-000
Brodbeck, C., Bhattasali, S., Heredia, UN. UN. C., Resnik, P., Simone, J. Z.,
& Lau, E. (2022). Parallel processing in speech perception with local
and global representations of linguistic context. Elife, 11, Article
e72056. https://doi.org/10.7554/eLife.72056, PubMed: 35060904
Brodbeck, C., Das, P., Gillis, M., Kulasingham, J. P., Bhattasali, S.,
Gaston, P., Resnik, P., & Simone, J. Z. (2021). Eelbrain: A Python
toolkit for time-continuous analysis with temporal response func-
zioni. bioRxiv. https://doi.org/10.1101/2021.08.01.454687
Brodbeck, C., Hong, l. E., & Simone, J. Z. (2018). Rapid transforma-
tion from auditory to linguistic representations of continuous
speech. Current Biology, 28(24), 3976–3983. https://doi.org/10
.1016/j.cub.2018.10.042, PubMed: 30503620
Brodbeck, C., Jiao, A., Hong, l. E., & Simone, J. Z. (2020). Neural
speech restoration at the cocktail party: Auditory cortex recovers
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurobiology of Language
341
Continuous speech processing under bimodal divided attention
masked speech of both attended and ignored speakers. PLOS
Biology, 18(10), Article e3000883. https://doi.org/10.1371
/journal.pbio.3000883, PubMed: 33091003
Brodbeck, C., & Simone, J. Z. (2020). Continuous speech processing.
Current Opinion in Physiology, 18, 25–31. https://doi.org/10
.1016/j.cophys.2020.07.014, PubMed: 33225119
Broderick, M. P., Anderson, UN. J., Di Liberto, G. M., Crosse, M. J., &
Lalor, E. C. (2018). Electrophysiological correlates of semantic
dissimilarity reflect the comprehension of natural, narrative
speech. Current Biology, 28(5), 803–809. https://doi.org/10
.1016/j.cub.2018.01.080, PubMed: 29478856
Carroll, l. (2010). Alice’s adventures in wonderland [Audiobook].
Librivox. (Originally published 1865). https://librivox.org/alices
-adventures-in-wonderland-by-lewis-carroll-5
Ciaramitaro, V. M., Chow, H. M., & Eglington, l. G. (2017).
Cross-modal attention influences auditory contrast sensitivity:
Decreasing visual load improves auditory thresholds for amplitude-
and frequency-modulated sounds. Journal of Vision, 17(3),
Article 20. https://doi.org/10.1167/17.3.20, PubMed: 28355632
Crosse, M. J., Di Liberto, G. M., Bednar, A., & Lalor, E. C. (2016).
The multivariate temporal response function (mTRF) toolbox: UN
MATLAB toolbox for relating neural signals to continuous stimuli.
Frontiers in Human Neuroscience, 10, Article 604. https://doi.org
/10.3389/fnhum.2016.00604, PubMed: 27965557
Ding, N., Pan, X., Luo, C., Su, N., Zhang, W., & Zhang, J. (2018).
Attention is required for knowledge-based sequential grouping:
Insights from the integration of syllables into words. Journal of
Neuroscience, 38(5), 1178–1188. https://doi.org/10.1523
/JNEUROSCI.2606-17.2017, PubMed: 29255005
Ding, N., & Simone, J. Z. (2012). Neural coding of continuous
speech in auditory cortex during monaural and dichotic listen-
ing. Journal of Neurophysiology, 107(1), 78–89. https://doi.org
/10.1152/jn.00297.2011, PubMed: 21975452
Duncan, J., Martens, S., & Ward, R. (1997). Restricted attentional
capacity within but not between sensory modalities. Nature,
387(6635), 808–810. https://doi.org/10.1038/42947, PubMed:
9194561
Easycap. (2023). Easycap [Recording cap]. www.easycap.de
Etymotic. (2023). ER-3 [Earphones]. https://www.etymotic.com
Fishbach, A., Nelken, I., & Yeshurun, Y. (2001). Auditory edge
detection: A neural model for physiological and psychoacousti-
cal responses to amplitude transients. Journal of Neurophysiol-
ogy, 85(6), 2303–2323. https://doi.org/10.1152/jn.2001.85.6
.2303, PubMed: 11387378
Gaston, P., Brodbeck, C., Phillips, C., & Lau, E. (2023). Auditory word
comprehension is less incremental in isolated words. Neurobiology
of Language, 4(1), 29–52. https://doi.org/10.1162/nol_a_00084
Gennari, S. P., Millman, R. E., Hymers, M., & Mattys, S. l. (2018).
Anterior paracingulate and cingulate cortex mediates the effects
of cognitive load on speech sound discrimination. NeuroImage,
178, 735–743. https://doi.org/10.1016/j.neuroimage.2018.06
.035, PubMed: 29902588
Gillis, M., Van Canneyt, J., Francart, T., & Vanthornhout, J. (2022).
Neural tracking as a diagnostic tool to assess the auditory path-
modo. Hearing Research, 426, Article 108607. https://doi.org/10
.1016/j.heares.2022.108607, PubMed: 36137861
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A., Strohmeier,
D., Brodbeck, C., Goj, R., Jas, M., Brooks, T., Parkkonen, L., &
Hämäläinen, M. (2013). MEG and EEG data analysis with
MNE-Python. Frontiers in Neuroscience, 7, Article 267. https://
doi.org/10.3389/fnins.2013.00267, PubMed: 24431986
Hamilton, l. S., & Huth, UN. G. (2020). The revolution will not be
controlled: Natural stimuli in speech neuroscience. Language,
Cognition and Neuroscience, 35(5), 573–582. https://doi.org/10
.1080/23273798.2018.1499946, PubMed: 32656294
Heafield, K. (2011). KenLM: Faster and smaller language model
queries. In Proceedings of the sixth workshop on statistical
machine translation (pag. 187–197). Associazione per il calcolo-
linguistica nazionale.
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8(5),
393–402. https://doi.org/10.1038/nrn2113, PubMed: 17431404
Interacoustics. (2023). Equinox 2.0 [Audiometer]. https://www
.interacoustics.com/audiometers/equinox
Jaeggi, S. M., Buschkuehl, M., Etienne, A., Ozdoba, C., Perrig,
W. J., & Nirkko, UN. C. (2007). On how high performers keep cool
brains in situations of cognitive overload. Cognitive, Affective, &
Behavioral Neuroscience, 7(2), 75–89. https://doi.org/10.3758
/CABN.7.2.75, PubMed: 17672380
Johnson, J. A., & Zatorre, R. J. (2006). Neural substrates for dividing
and focusing attention between simultaneous auditory and visual
events. NeuroImage, 31(4), 1673–1681. https://doi.org/10.1016/j
.neuroimage.2006.02.026, PubMed: 16616520
Kasper, R. W., Cecotti, H., Touryan, J., Eckstein, M. P., &
Giesbrecht, B. (2014). Isolating the neural mechanisms of inter-
ference during continuous multisensory dual-task performance.
Journal of Cognitive Neuroscience, 26(3), 476–489. https://doi
.org/10.1162/jocn_a_00480, PubMed: 24047391
Katus, T., & Eimer, M. (2019). The sources of dual-task costs in
multisensory working memory tasks. Journal of Cognitive Neuro-
science, 31(2), 175–185. https://doi.org/10.1162/jocn_a_01348,
PubMed: 30277430
Keitel, C., Maess, B., Schröger, E., & Müller, M. M. (2013). Early
visual and auditory processing rely on modality-specific atten-
tional resources. NeuroImage, 70, 240–249. https://doi.org/10
.1016/j.neuroimage.2012.12.046, PubMed: 23287527
Keuleers, E., Brysbaert, M., & Nuovo, B. (2010). SUBTLEX-NL: A new
measure for Dutch word frequency based on film subtitles.
Behavior Research Methods, 42(3), 643–650. https://doi.org/10
.3758/BRM.42.3.643, PubMed: 20805586
Kiremitçi, I., Yilmaz, Ö., Çelik, E., Shahdloo, M., Huth, UN. G., &
Çukur, T. (2021). Attentional modulation of hierarchical speech
representations in a multitalker environment. Cerebral Cortex,
31(11), 4986–5005. https://doi.org/10.1093/cercor/ bhab136,
PubMed: 34115102
Klemen, J., Büchel, C., & Rose, M. (2009). Perceptual load interacts
with stimulus processing across sensory modalities. European
Journal of Neuroscience, 29(12), 2426–2434. https://doi.org/10
.1111/j.1460-9568.2009.06774.x, PubMed: 19490081
Lachter, J., Forster, K. I., & Ruthruff, E. (2004). Forty-five years after
Broadbent (1958): Still no identification without attention. Psy-
chological Review, 111(4), 880–913. https://doi.org/10.1037
/0033-295X.111.4.880, PubMed: 15482066
Lavie, N. (2005). Distracted and confused?: Selective attention
under load. Trends in Cognitive Sciences, 9(2), 75–82. https://
doi.org/10.1016/j.tics.2004.12.004, PubMed: 15668100
Lavie, N., & Tsal, Y. (1994). Perceptual load as a major determinant
of the locus of selection in visual attention. Perception & Psycho-
physics, 56(2), 183–197. https://doi.org/10.3758/BF03213897,
PubMed: 7971119
Loftus, G. R., & Masson, M. E. (1994). Using confidence intervals in
within-subject designs. Psychonomic Bulletin & Review, 1(4),
476–490. https://doi.org/10.3758/ BF03210951, PubMed:
24203555
Loose, R., Kaufmann, C., Auer, D. P., & Lange, K. W. (2003).
Human prefrontal and sensory cortical activity during divided
Neurobiology of Language
342
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Continuous speech processing under bimodal divided attention
attention tasks. Human Brain Mapping, 18(4), 249–259. https://
doi.org/10.1002/hbm.10082, PubMed: 12632463
Macdonald, J. S. P., & Lavie, N. (2011). Visual perceptual load
induces inattentional deafness. Attention, Perception, & Psycho-
physics, 73(6), 1780–1789. https://doi.org/10.3758/s13414-011
-0144-4, PubMed: 21611856
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical testing
of EEG- and MEG-data. Journal of Neuroscience Methods,
164(1), 177–190. https://doi.org/10.1016/j.jneumeth.2007.03
.024, PubMed: 17517438
Marslen-Wilson, W. D. (1987). Functional parallelism in spoken
word-recognition. Cognition, 25(1–2), 71–102. https://doi.org
/10.1016/0010-0277(87)90005-9, PubMed: 3581730
Mattys, S. L., Barden, K., & Samuel, UN. G. (2014). Extrinsic cogni-
tive load impairs low-level speech perception. Psychonomic
Bulletin & Review, 21(3), 748–754. https://doi.org/10.3758
/s13423-013-0544-7, PubMed: 24189992
Mattys, S. L., Brooks, J., & Cooke, M. (2009). Recognizing speech
under a processing load: Dissociating energetic from informa-
tional factors. Cognitive Psychology, 59(3), 203–243. https://doi
.org/10.1016/j.cogpsych.2009.04.001, PubMed: 19423089
Mattys, S. L., & Palmer, S. D. (2015). Divided attention disrupts per-
ceptual encoding during speech recognition. Journal of the
Acoustical Society of America, 137(3), 1464–1472. https://doi
.org/10.1121/1.4913507, PubMed: 25786957
Mattys, S. L., & Wiget, l. (2011). Effects of cognitive load on speech
recognition. Journal of Memory and Language, 65(2), 145–160.
https://doi.org/10.1016/j.jml.2011.04.004
McAuliffe, M., Socolof, M., Mihuc, S., Wagner, M., & Sonderegger,
M. (2017). Montreal Forced Aligner: Trainable text-speech align-
ment using Kaldi. In INTERSPEECH 2017 (pag. 498–502). ISCA
Web. https://doi.org/10.21437/Interspeech.2017-1386
Molloy, K., Griffith, T. D., Chait, M., & Lavie, N. (2015). Inatten-
tional deafness: Visual load leads to time-specific suppression of
auditory evoked responses. Journal of Neuroscience, 35(49),
16046–16054. https://doi.org/10.1523/ JNEUROSCI.2931-15
.2015, PubMed: 26658858
Morey, R. D., Rouder, J. N., Jamil, T., Urbanek, S., Forner, K., & Ly,
UN. (2022). BayesFactor: Computation of Bayes factors for com-
mon designs ( Version 0.9.12–4.4) [Software]. https://CRAN.R
-project.org/package=BayesFactor
Murphy, G., Groeger, J. A., & Greene, C. M. (2016). Twenty years of
load theory—Where are we now, and where should we go next?
Psychonomic Bulletin & Review, 23(5), 1316–1340. https://doi
.org/10.3758/s13423-015-0982-5, PubMed: 26728138
Oostenveld, R., & Praamstra, P. (2001). The five percent electrode
system for high-resolution EEG and ERP measurements. Clinical
Neurophysiology, 112(4), 713–719. https://doi.org/10.1016
/S1388-2457(00)00527-7, PubMed: 11275545
Parks, N. A., Hilimire, M. R., & Corballis, P. M. (2011). Steady-state
signatures of visual perceptual load, multimodal distractor filter-
ing, and neural competition. Journal of Cognitive Neuroscience,
23(5), 1113–1124. https://doi.org/10.1162/jocn.2010.21460,
PubMed: 20146614
Pickering, M. J., & Gambi, C. (2018). Predicting while compre-
hending language: A theory and review. Psychological Bulletin,
144(10), 1002–1044. https://doi.org/10.1037/ bul0000158,
PubMed: 29952584
Porcu, E., Keitel, C., & Müller, M. M. (2014). Visual, auditory and
tactile stimuli compete for early sensory processing capacities
within but not between senses. NeuroImage, 97, 224–235.
https://doi.org/10.1016/j.neuroimage.2014.04.024, PubMed:
24736186
R Core Team. (2022). R: A language and environment for statistical
computing. R Foundation for Statistical Computing. https://www
.R-project.org/
Salo, E., Rinne, T., Salonen, O., & Alho, K. (2015). Brain activations
during bimodal dual tasks depend on the nature and combina-
tion of component tasks. Frontiers in Human Neuroscience, 9,
Article 102. https://doi.org/10.3389/fnhum.2015.00102, PubMed:
25767443
Sanders, l. D., & Neville, H. J. (2003). An ERP study of continuous
speech processing: IO. Segmentation, semantics, and syntax in native
speakers. Cognitive Brain Research, 15(3), 228–240. https://doi.org
/10.1016/S0926-6410(02)00195-7, PubMed: 12527097
Sanders, l. D., Newport, E. L., & Neville, H. J. (2002). Segmenting
nonsense: An event-related potential index of perceived onsets in
continuous speech. Nature Neuroscience, 5(7), 700–703. https://
doi.org/10.1038/nn873, PubMed: 12068301
Schneider, W., Eschman, A., & Zuccolotto, UN. (2002). E-Prime:
User’s guide. Reference guide. Getting started guide. Psychology
Software Tools.
Smout, C. A., Tang, M. F., Garrido, M. I., & Mattingley, J. B. (2019).
Attention promotes the neural encoding of prediction errors.
PLOS Biology, 17(2), Article e2006812. https://doi.org/10.1371
/journal.pbio.2006812, PubMed: 30811381
Snodgrass, J. G., & Corwin, J. (1988). Pragmatics of measuring rec-
ognition memory: Applications to dementia and amnesia. Journal
of Experimental Psychology: General, 117(1), 34–50. https://doi
.org/10.1037/0096-3445.117.1.34, PubMed: 2966230
Sörqvist, P., Stenfelt, S., & Rönnberg, J. (2012). Working memory
capacity and visual–verbal cognitive load modulate auditory–
sensory gating in the brainstem: Toward a unified view of atten-
zione. Journal of Cognitive Neuroscience, 24(11), 2147–2154.
https://doi.org/10.1162/jocn_a_00275, PubMed: 22849400
Vanthornhout, J., Decruy, L., & Francart, T. (2019). Effect of task and
attention on neural tracking of speech. Frontiers in Neuroscience,
13, Article 977. https://doi.org/10.3389/fnins.2019.00977,
PubMed: 31607841
VPixx Technologies. (2022). VIEWPixx [monitor]. https://vpixx.com
/products/viewpixx/
Wahn, B., & König, P. (2017). Is attentional resource allocation
across sensory modalities task-dependent? Advances in Cognitive
Psychology, 13(1), 83–96. https://doi.org/10.5709/acp-0209-2,
PubMed: 28450975
Yahav, P. H.-S., & Golumbic, E. Z. (2021). Linguistic processing of
task-irrelevant speech at a cocktail party. Elife, 10, Article e65096.
https://doi.org/10.7554/eLife.65096, PubMed: 33942722
Neurobiology of Language
343
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
3
1
8
2
0
7
9
0
3
4
N
o
_
UN
_
0
0
1
0
0
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3