RESEARCH ARTICLE
Independency of Coding for Affective Similarities
and for Word Co-occurrences in Temporal
Perisylvian Neocortex
a n o p e n a c c e s s
j o u r n a l
Patrick Dupont1
Antonietta Gabriella Liuzzi1*
, Karen Meersmans1*
, Simon De Deyne3
,
, Gerrit Storms2
, and Rik Vandenberghe1,4
1Laboratory for Cognitive Neurology, Department of Neurosciences, Leuven Brain Institute, KU Leuven, Leuven, Belgium
2Laboratory of Experimental Psychology, KU Leuven, Leuven, Belgium
3Computational Cognitive Science Lab, University of Melbourne, Melbourne, Australia
4Neurology Department, University Hospitals Leuven, Leuven, Belgium
*Antonietta Gabriella Liuzzi and Karen Meersmans share first authorship.
Keywords: fMRI, representational similarity analysis, semantics, word embedding models
ABSTRACT
Word valence is one of the principal dimensions in the organization of word meaning.
Co-occurrence-based similarities calculated by predictive natural language processing models
are relatively poor at representing affective content, but very powerful in their own way. Here,
we determined how these two canonical but distinct ways of representing word meaning
relate to each other in the human brain both functionally and neuroanatomically. Noi
re-analysed an fMRI study of word valence. A co-occurrence-based model was used and the
correlation with the similarity of brain activity patterns was compared to that of affective
similarities. The correlation between affective and co-occurrence-based similarities was low
(r = 0.065), confirming that affect was captured poorly by co-occurrence modelling. In a
whole-brain representational similarity analysis, word embedding similarities correlated
significantly with the similarity between activity patterns in a region confined to the superior
temporal sulcus to the left, and to a lesser degree to the right. Affective word similarities
correlated with the similarity in activity patterns in this same region, confirming previous
findings. The affective similarity effect extended more widely beyond the superior temporal
cortex than the effect of co-occurrence-based similarities did. The effect of co-occurrence-
based similarities remained unaltered after partialling out the effect of affective similarities (E
vice versa). To conclude, different aspects of word meaning, derived from affective judgements
or from word co-occurrences, are represented in superior temporal language cortex in a
neuroanatomically overlapping but functionally independent manner.
INTRODUCTION
Over the past decade, advances in computational modelling of word meaning have enabled
novel ways of studying the representation of meaning in the human brain. Models of word
meaning have relied on different types of sources. Data can be acquired from study partici-
pants performing explicit tasks, such as free word association (De Deyne et al., 2019), feature
generation (De Deyne et al., 2008; McRae et al., 2005), or valence ratings (Moors et al., 2013;
Van Rensbergen et al., 2016). According to such studies, word valence, or how positive/
negative a word is, is one of the principal dimensions of the organisation of word meaning
Citation: Liuzzi, UN. G., Meersmans, K.,
Storms, G., De Deyne, S., Dupont, P., &
Vandenberghe, R. (2023). Independency
of coding for affective similarities and
for word co-occurrences in temporal
perisylvian neocortex. Neurobiology
of Language, 4(2), 257–279. https://doi
.org/10.1162/nol_a_00095
DOI:
https://doi.org/10.1162/nol_a_00095
Supporting Information:
https://doi.org/10.1162/nol_a_00095
Received: 26 Gennaio 2022
Accepted: 9 Dicembre 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Rik Vandenberghe
rik.vandenberghe@kuleuven.be
Handling Editor:
Steven Small
Copyright: © 2023
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
(Belyk et al., 2017; De Deyne & Storms, 2008; Kotz & Paulmann, 2011; Kousta et al., 2011;
Kuperman et al., 2014; Meersmans et al., 2020; Osgood et al., 1957; Pauligk et al., 2019;
Troche et al., 2017; Van Rensbergen et al., 2015; Vigliocco et al., 2014). Word valence is also
highly relevant for how brain patterns are organised in response to words (Belyk et al., 2017;
Meersmans et al., 2020, 2022; Pauligk et al., 2019). The effect of affective similarity (modelled
as the Euclidean distance between words in a three-dimensional space formed by valence,
dominance, and arousal ratings) is strong in the perisylvian language network and beyond
(Meersmans et al., 2020, 2022).
As another highly influential approach to the representation of word meaning, word mean-
ing models can make use of the distributional structure (Harris, 1954) present in the vast amount
of pre-existing language corpora. Words can then be represented as word embeddings, cioè.,
continuous, low-dimensional, real-value vectors based on co-occurrences (Devereux et al.,
2010; Hollis & Westbury, 2016; Mitchell et al., 2008; Pereira et al., 2013). Natural language
processing (PNL; Abnar et al., 2018; Grave et al., 2019; Honnibal, 2017) is then applied to
extract and detect regularities and statistical patterns of co-occurrence and transforms real-
world language into computer-friendly real-valued vectors representations. A limitation, how-
ever, is that word embedding models have a rather limited ability to model the affective content
of words, such as word polarity or other aspects of text sentiment (De Deyne et al., 2021; Tang
et al., 2016; Yu et al., 2018). As these models are strongly context dependent, words with oppo-
site polarity (per esempio., “bad” vs. “good”) but similar contexts may be mapped near to each other by
such models (Agrawal et al., 2018). Valence, arousal, and dominance can be derived from
co-occurrence-based modelling but only provided that seed words are present of which the
valence has been determined beforehand based on subjective ratings or lexica (“mots germes”;
Recchia & Louwerse, 2015; Vincze & Bestgen, 2011). Even then, models based on task-
generated data outperform co-occurrence-based models in predicting valence, arousal and
dominance (Vankrunkelsven et al., 2018).
Hence, two of the principal ways of modeling word content, based on affective judgements
versus word co-occurrences, capture different aspects of word meaning. The interest of this
special relationship between co-occurrence-based modelling and affective word content
can be gauged based on the number of high-quality prior studies from the NLP community
(Tang et al., 2016; Yu et al., 2018), from computational linguistics (Westbury et al., 2015), psy-
cholinguistics (Bestgen & Vincze, 2012; De Deyne et al., 2021), and experimental psychology
on this topic (Hollis & Westbury, 2016; Recchia & Louwerse, 2015; Vankrunkelsven et al.,
2018; Vincze & Bestgen, 2011; Westbury et al., 2015).
We examined how brain activity patterns relate to these two ways of representing word
meaning that have been proven to be powerful in their own right but nevertheless appear
to be fundamentally distinct. Given the distinctive nature of these representations, one might
predict that they would rely on separate brain regions. Affective similarities have been reported
to correlate strongly with the similarity of brain activity patterns in superior temporal language
cortex (Meersmans et al., 2020), whereas co-occurrence-based similarities have been associ-
ated with a neuroanatomically relatively diffuse effect (Mitchell et al., 2008).
Word co-occurrences are influenced by word associations. Word associations can also be
derived from other sources, such as experimentally using cued word generation (De Deyne
et al., 2019; Vankrunkelsven et al., 2018). Hence, cued-association-based similarities hold a
position in between co-occurrence-based and affective similarities. To gain further insight into
the mechanism behind the correlations between co-occurrence-based similarities and brain
activity patterns, we evaluated whether effects were comparable when similarities were based
Neurobiology of Language
258
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
on task-generated word association data. This allows us to determine whether the critical fac-
tor determining the co-occurrence-based word similarity effects are the predictive computa-
tional modelling algorithms behind the word similarities or the word associations as such.
We re-analysed a functional magnetic resonance imaging (fMRI) data set from a study orig-
inally designed to investigate the representation of affective similarity between nouns. Nouns
were presented in both auditory and in visual modality to further increase generalisability of
our findings and analyses were pooled across modalities and tasks. Affective similarities were
calculated from estimates of word valence, arousal and dominance, which were extrapolated
from a data set of behavioural affective norms (Moors et al., 2013). These extrapolations were
made based on a graph derived from a huge data set based on free word association (Van
Rensbergen et al., 2016; for further details see Materials and Methods). Secondly, for the noun
stimulus set used in this experiment, we re-calculated word similarities, using the word2vec
vectors provided through the spaCy Python library, and examined how the corresponding
brain activity patterns related to those obtained for the affective similarities. The primary word
embeddings are based on the continuous bag of words (cBOW) approach. In this architecture,
the target word is predicted based on the embeddings of its immediate context (per esempio., 3–4
neighbouring words, depending on the window size) derived from a large internet-based text
corpus, composed of Wikipedia entries and an extensive Webcrawl (>13 billion words;
Honnibal, 2017). This study was meant to uncover commonalities and differences in coding
mechanisms for similarities based on word associations/co-occurrences versus affective judge-
menti, rispettivamente.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
MATERIALS AND METHODS
Subjects
Twenty-two subjects (15 women and 7 men, mean age 21.9, 19–24 years of age, all neuro-
logically healthy and right-handed) participated. This sample size is in accordance with pre-
vious fMRI studies using the same approach and yielding replicable results (Bruffaerts et al.,
2013; Carota et al., 2021; Liuzzi et al., 2015, 2019; Pauligk et al., 2019). All subjects were free
of significant neurological or psychiatric history. Subjects provided informed consent before
participating, and the experiments were approved by the Ethics Committee Clinical Studies
UZ/KU Leuven.
Stimuli
Stimulus words were selected from the 12,400 cue words studied as part of the Dutch Small
World of Words (SWOW) insieme di dati (De Deyne et al., 2019).
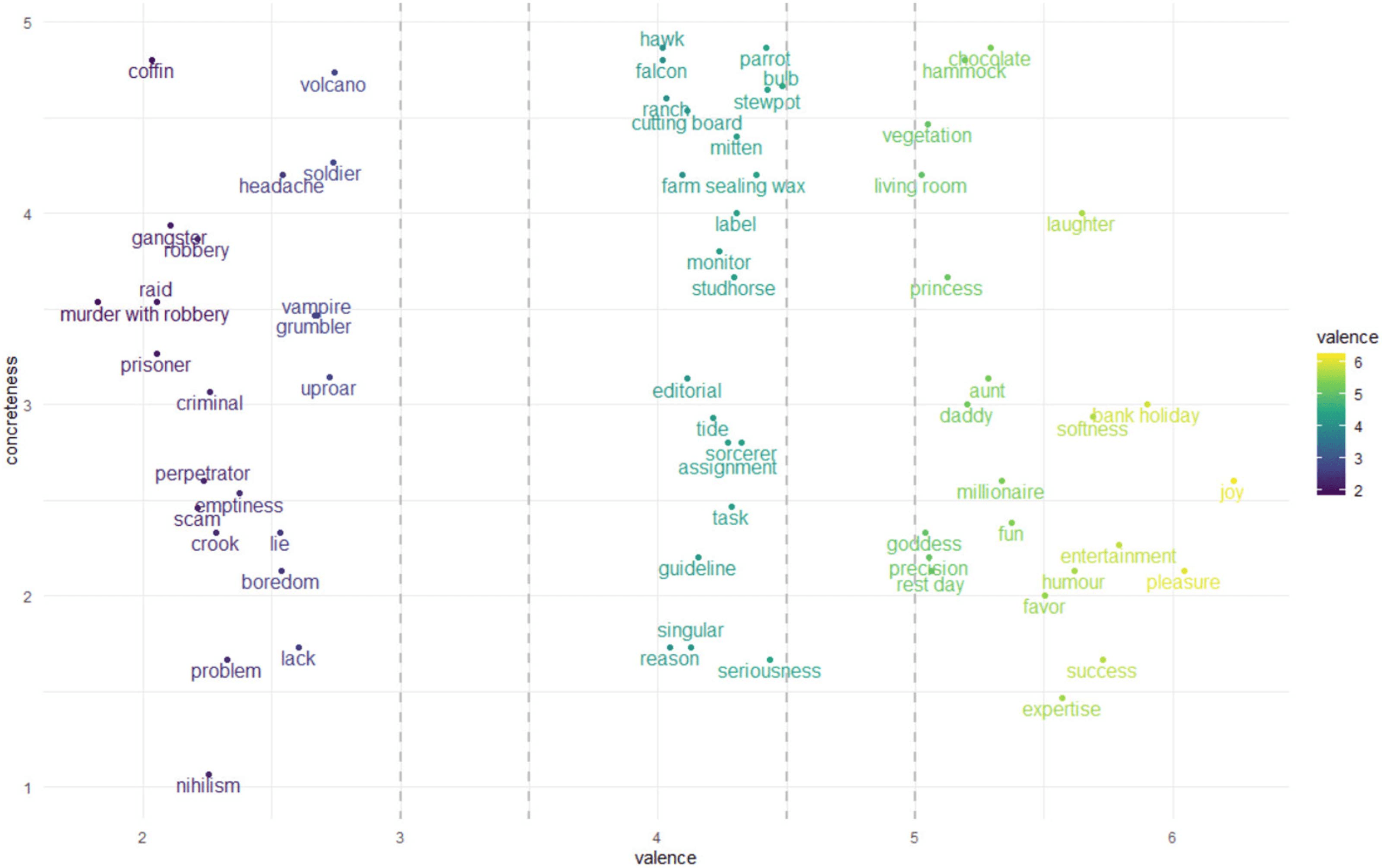
A total of 66 nouns were distributed evenly over three classes: positive, neutro, and neg-
ative valence (Figura 1). Words were selected in a semiautomated manner based on the fol-
lowing criteria: (1) to maximise the range in semantic similarity, questo è, contain word pairs that
range between highly similar and highly dissimilar; E (2) to match words between the three
valence classes for all relevant linguistic variables. The three groups were matched on age of
acquisition, concreteness, dominance, log10(frequency), orthographic neighbourhood density,
prevalence, and word length (Brysbaert et al., 2014; Keuleers et al., 2010, 2015; Marian et al.,
2012; Van Rensbergen et al., 2016). The word selection script did not converge to a solution
when arousal also needed to be matched between valence classes. Arousal was numerically
lower for the neutral valence (mean = 3.98, SD = 0.49) class than for the positive (mean =
4.23, SD = 0.79) or negative valence class (mean = 4.54, SD = 1.05) (one-way analysis of
Small World of Words:
Publicly available data set obtained
by means of online responses of
more than 70,000 individuals to
more than 12,000 parole, performed
for different native languages.
Neurobiology of Language
259
Affective similarities and word co-occurrences in temporal neocortex
Figura 1. Stimulus set. The two coordinates of each noun correspond to valence (x-axis) and concreteness (y-axis).
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
variance: F(2, 63) = 2.62, p = 0.08). This is a consequence of the well-established U-shaped
relationship between valence and arousal (Warriner et al., 2013).
The stimulus set is provided in Figure 1 and in Table S1 in the Supporting Information. IL
stimulus selection procedure yielded both concrete and abstract words without evident taxo-
nomic structure (as measured based on WordNet; Mugnaio, 1995).
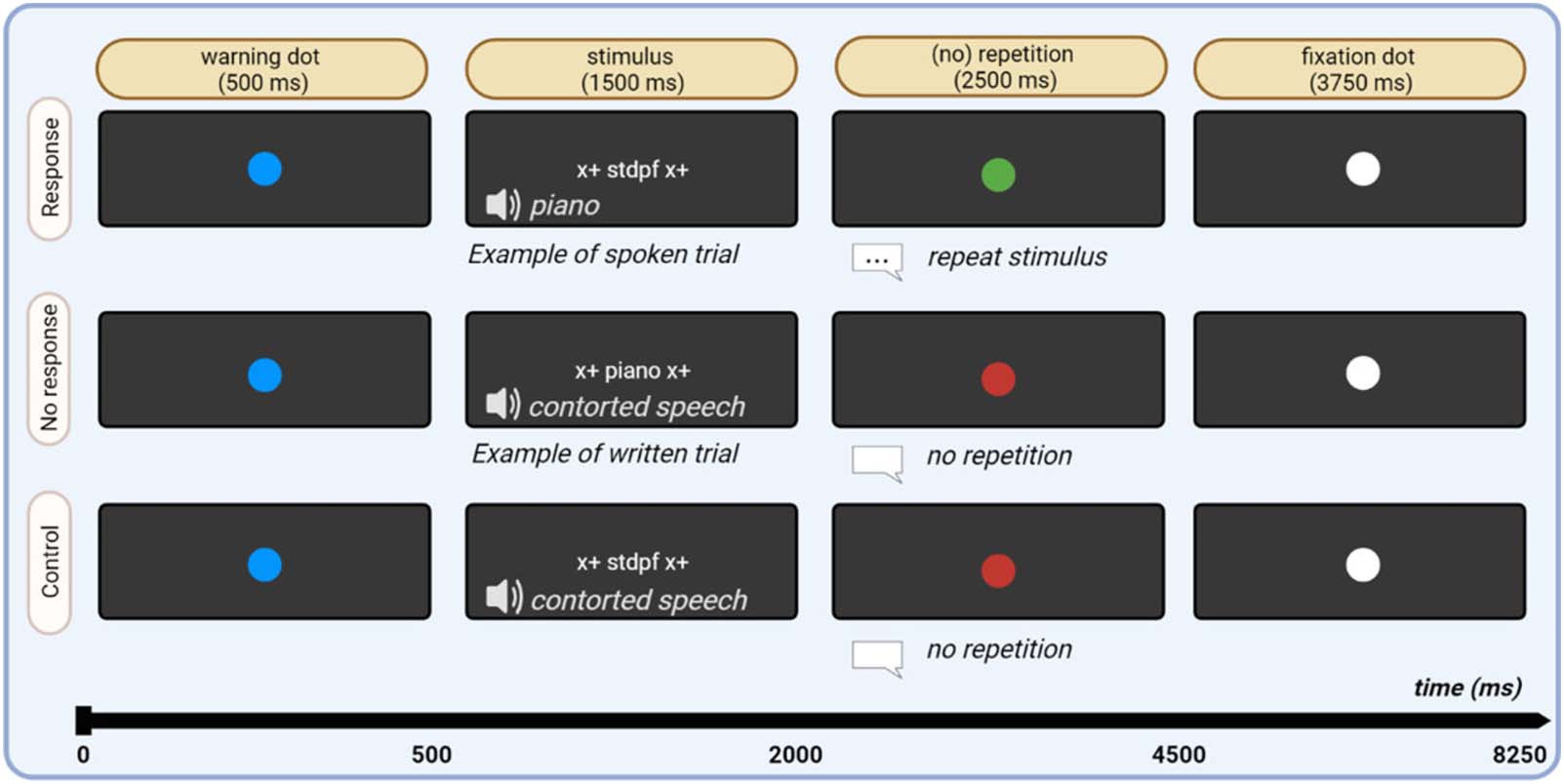
Experimental Design
The experiment was originally designed for the purpose of studying the effect of valence on
fMRI activity patterns. The experiment had a 3 × 2 × 2 factorial design with stimulus type
(positive, neutro, negative), modality (visual or auditory presentation), and overt articulation
(yes/no) as factors. After a warning dot (500 ms), stimulus words were presented in visual or
auditory modality (1,500 ms), followed by the presentation of a red or green dot (2,500 ms).
Presentation of a green dot indicated to the subjects that they had to repeat the word aloud
after stimulus presentation had ended. The red dot indicated that subjects had to remain silent.
Visual stimuli were presented in white on a black background. Auditory stimuli were pre-
sented through OptoActive II active noise cancelling headphones (Optoacoustics, 2022) A
minimise interference of scanner noise. The task was chosen to monitor subject engagement
in the scanner, while limiting the effect of attentional orienting and control that can occur
during explicit semantic tasks. To limit head motion, subjects were explicitly instructed to
Neurobiology of Language
260
Affective similarities and word co-occurrences in temporal neocortex
pronounce the words while restricting mouth movement to a minimum. Vocal responses were
recorded using the OptoActive II FOMRI III microphone (Optoacoustics, 2022), which enables
recording of responses in the fMRI environment in a sensitive manner. Head motion parame-
ters were determined during image analysis, and runs were rejected based on a prior criterion
(see below).
The initial 1,500 ms phase following word onset was identical between covert and overt
trials up to response cue onset. The covert and overt trials will be pooled in the current report.
Following the response phase, a white fixation dot was presented until the start of the next trial.
The interstimulus interval was 8.25 S. Every noun was presented once during each of the eight
runs (4× in visual and 4× in auditory modality; duration per run: 10.5 min). Visual stimuli were
accompanied by an auditory control (cioè., rotated spectrogram speech), while auditory stimuli
were accompanied by a visual control (cioè., consonant letter string). Audio controls were gen-
erated from the stimuli by rotating the spectrogram around 50% of the maximal frequency and
applying a low-pass filter at 95% of the maximal frequency. This procedure maintains certain
spectral and phonetic features and intonation, while at the same time rendering the word unin-
telligible (Scott et al., 2000). Visual controls were generated by replacing the vowels in the
original stimulus words by consonants and shuffling the letter order. During the control trials,
the visual and auditory control stimuli were presented simultaneously, and no response was
required (Figura 2).
Every run consisted of 77 trials (66 word trials + 11 control trials). The control trials were
included as event type in the general linear model but were not used further in the current
analyses. Every subject completed eight runs over two scan sessions. The word sets were pre-
sented in a pseudorandomised order and divided in a counterbalanced manner between the
two sessions. This resulted in a total of eight replications per word (4 spoken and 4 written
modality).
Word similarity matrices
The primary objective of the current analysis was to examine how similarities derived from
word co-occurrence-based models relate to similarities between fMRI activity patterns and
to identify the commonalities and differences of these effects with those obtained for affective
Figura 2. Experimental design.
Neurobiology of Language
261
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
Representational similarity analysis
(RSA):
Method where the similarity between
experimental stimuli (such as words)
is related to the similarity between
the activity patterns in response to
these stimuli.
similarities. The word co-occurrence model was derived directly from spaCy (Honnibal,
2017). SpaCy is an open-source Python library for NLP. It contains built-in word embeddings
(cioè., multidimensional vector representations of word meaning). Here, we use the word2vec
word embeddings from spaCy’s large Dutch model (https://spacy.io/models/nl#nl_core_news_lg),
which uses a cBOW approach on a combined corpus (web crawled text + Wikipedia). SpaCy
similarities were normally distributed (Figure 3D; Figure S1 in the Supporting Information). IL
resulting similarity matrix was used for representational similarity analysis (RSA).
In the affective similarity model, we used valence, dominance, and arousal ratings (Van
Rensbergen et al., 2016) to create a three-dimensional space. The pairwise Euclidean distance
d between stimulus words was calculated, normalised, and converted to a similarity metric by
1 − d. Valence, dominance, and arousal ratings were taken from a large data set of affective
estimates (Van Rensbergen et al., 2016). In this study, word valence, dominance, and arousal
were extrapolated for 14,000 words from a smaller data set of behavioural norms (n = 4,300
parole; Moors et al., 2013) using a k-nearest neighbour approach. The nearest neighbours were
determined based on word embedding similarities derived from a free association graph (De
Deyne et al., 2019). This graph is created from a large data set of continued associations
(>12,000 words; see below for more details) with the words as nodes in the graph and the
edges weighted by the association strength p(risposta|cue). In the affective ratings data set
by Moors et al. (2013), word valence, dominance, and arousal were rated on a 1–7 scale.
Only words for which these extrapolations were available were eligible for this study.
In a secondary analysis, for comparison with the spaCy based model, we included a behav-
ioural similarity model derived from the Small World of Words word association data set (De
Deyne et al., 2019). In this similarity model, word embeddings were derived from a large-scale
continued association task, during which subjects were instructed to provide three associates
for every cue (De Deyne et al., 2019). The SWOW data set comprises responses for over
12,000 cue words from 70,000+ subjects. From this data set, a graph is constructed using asso-
ciative strength (cioè., probability of the response given a cue) as edge weight. Word embed-
dings were extracted from the graph using a random walk algorithm, and semantic similarity
was calculated using cosine similarity (De Deyne et al., 2019; Liuzzi et al., 2019; Meersmans
et al., 2020). Valence (pleasantness; positive [holiday] − negative [murder], dominance
(power; strong [avalanche] − weak [sleep]), and arousal (activity; active [explosion] − passive
[silence]) are important dimensions in this graph (Van Rensbergen et al., 2015). Inoltre, UN
direct comparison between word embeddings and word association indicated that the latter is
better suited to capture affective information (De Deyne et al., 2021) The SWOW similarity
matrix was not normally distributed (Figure 3D and Figure S1 in the Supporting Information).
To evaluate and control for the possible contribution of phonological similarities to the
effects obtained, we calculated a phonological similarity matrix as 1 − the Levenshtein distance
between phonological transcriptions of the stimuli. As a separate phonological model, we also
calculated phonological neighbourhood density similarities as the difference in phonological
neighbourhood density between stimuli (cioè., the number of words that can be constructed by
substitution, aggiunta, or deletion of one phoneme). Phonological transcriptions and neighbour-
hood density estimates were retrieved from the CLEARPOND database for Dutch (Marian et al.,
2012; https://clearpond.northwestern.edu/dutchpond.php). These models were included to
evaluate whether the observed semantic effects are in part driven by phonology.
Since prior studies have emphasised the role of taxonomic structure and of experiential
similarities in the organization of brain activity pattern (Fernandino et al., 2022), we also cal-
culated word similarity based on taxonomy and word similarity derived from experiential
Neurobiology of Language
262
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
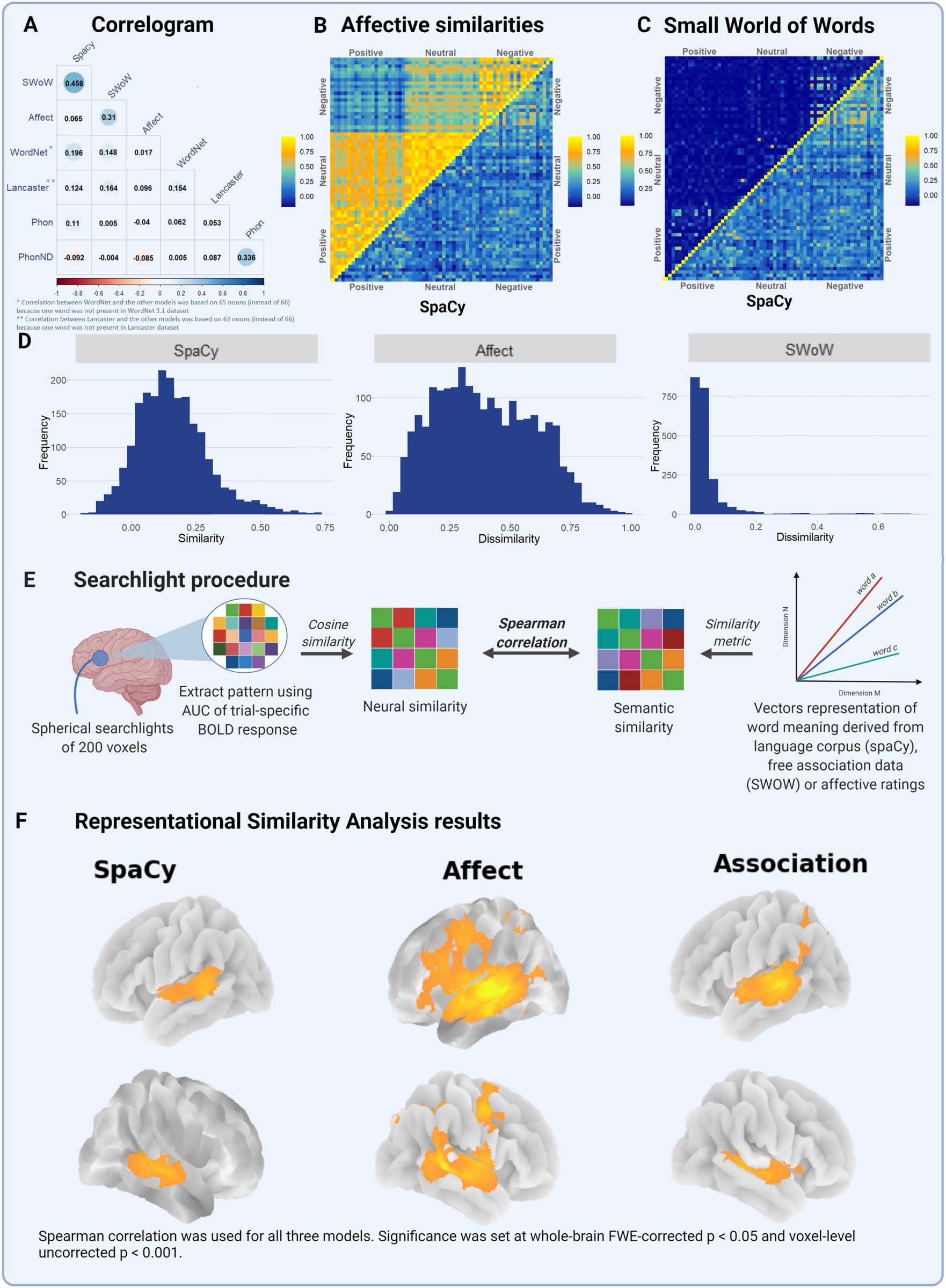
Figura 3. Overview of the word embedding models, the whole-brain representational similarity analysis (RSA) procedure, and the results. (UN)
Correlations between the different similarity matrices. (B) Affective similarity matrix (upper half of the matrix) next to the distributional similarity
matrix derived from spaCy (lower half of the matrix). (C) Compares the Small World of Words and spaCy matrices in the same manner. (D)
Distribution of the similarities from spaCy, affective ratings and Small World of Words (SWoW). (E) Provides a schematic overview of the
whole-brain RSA using a 200 voxel searchlight. (F) Results of the RSA for all three models.
Neurobiology of Language
263
Affective similarities and word co-occurrences in temporal neocortex
judgements for our set of nouns and tested the correlation with the dimensions of primary
interesse, the affective similarities, and the co-occurrence-based similarities. Taxonomic struc-
ture was modelled based on WordNet (https://wordnet.princeton.edu/). WordNet similarity
matrix was calculated based on Wu and Palmer (1994) similarity ( WPsim). WPsim calculates
the relatedness between two words by considering the depths of the two synonyms (cioè., syn-
sets) in the WordNet taxonomies, along with the depth of the least common subsumer (cioè., IL
most specific common hypernym). Experiential strength was based on the Lancaster sensori-
motor norms (Lynott et al., 2020). Lancaster similarity matrix was calculated by computing the
cosine similarity for all pairs of concept vectors (Vedi la tabella 1 for an overview of the models).
The taxonomic and experiential similarities were calculated from English translations of the
Dutch nouns. Three nouns were absent from the Lancaster norms and one noun was absent
from WordNet, hence these were left out (Table S1, for Dutch nouns and English translations).
Correlations between similarities from the different models
Correlations between the different word embedding models show the difference in overlap
between models (Figure 3A). The correlations between the co-occurrence model and the
association-based model was 0.458, showing the partial collinearity between the models. Questo
is in line with the view that co-occurrence-based models capture some of the meaning
encoded in word associations.
In contrasto, the affective similarity model correlated poorly with the co-occurrence-based
modello (r = 0.065). Affective similarity was better captured by the association-based model (r =
0.310) than by the co-occurrence-based models. This difference was formally evaluated using
the cocor package for R (Diedenhofen & Musch, 2015), indicating that the correlation
between the affective and association-based model was significantly stronger than the corre-
lation between the affective and co-occurrence-based model (Pearson and Filon’s z = −11.4;
P < 0.0001).
The correlations between the phonological (neighbourhood density) matrix and the seman-
tic matrices were overall very low (Figure 3A). The Spearman correlation of the co-occurrence-
based similarities with the taxonomy-based similarities was 0.196, and with the experiential
strength-based similarities 0.124. The Spearman correlation of the affective similarities with
the taxonomy-based similarities was 0.017, and with the experiential-strength-based similar-
ities 0.096. Despite the low or negative correlations with phonological distance and neigh-
bourhood density, taxonomy-based or experiential-strength-based similarity matrices, we also
calculated the RSA effect of co-occurrence-based and affective similarities after partialling out
each of these factors.
Image Acquisition
Functional and structural fMRI images were acquired on a 3T Philips Achieva with a 32-
channel head coil and equipped with the OptoActive II ANC headphones and FOMRI III
microphone. Functional images were acquired using a T2* sequence with 36 slices (multiband
acceleration factor = 2; repetition time = 1 s; echo time = 30 ms, field of view = 220 × 135 ×
220 mm3, voxel size = 2.75 × 2.75 × 3.75 mm3). The temporal signal-to-noise ratio was cal-
culated by dividing the mean of the residual time series by its standard deviation (Murphy
et al., 2007; Figure 4). Structural images were obtained using a T1- weighted 3D turbo-
field-echo (repetition time = 9.6 ms, echo time = 4.6 ms, in-plane resolution = 0.97 mm, slice
thickness = 1.2 mm) in every subject.
Neurobiology of Language
264
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
A
f
f
e
c
t
i
v
e
s
i
m
i
l
a
r
i
t
i
e
s
a
n
d
w
o
r
d
c
o
-
o
c
c
u
r
r
e
n
c
e
s
i
n
t
e
m
p
o
r
a
l
n
e
o
c
o
r
t
e
x
Table 1. Overview of the models.
SpaCy
Co-occurrence Wikipedia and webcrawl
Type
Data
N
300
cBOW
Procedure
Similarity
Cosine
Affective similarties
Task-based
Valence, dominance, and arousal
3 Human ratings extrapolated based
Euclidean
ratings
on SWOW similarities
Small World of
Task-based
Semantic associations
NA
Random walk
Cosine
Words
Phonological
similarities
Phonological
Transcriptions from CLEARPOND
NA Number of insertions, deletions or
substitutions needed to change
one string into the other
Levenshtein
Phonological
Phonological
Total number of words that differed
1 NA
neighbourhood
density
in the addition, deletion, or
substitution of a single phoneme
Absolute value of the
difference in phonological
neighbourhood density
Taxonomic structure
Taxonomy
WordNet v31
NA
Knowledge graph where words are
Wu & Palmer similarity
grouped into synsets
Experiential strength
Task-based
Lancaster sensorimotor norms
12 Human ratings for 39,707 concepts
Cosine similarity
across six perceptual modalities
and five action effectors
Note. N: number of dimensions in the model. cBOW: continuous bag of words. SWOW: Small World of Words.
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
2
6
5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Temporal signal-to-noise ratio, calculated by dividing the mean of the residual time
series by its standard deviation.
Image Preprocessing
Images were submitted to a preprocessing pipeline in SPM12 (Ashburner et al., 2021). Func-
tional images were realigned to correct for head motion, followed by slice timing correction.
Next, functional and structural images were coregistered and the anatomical images were seg-
mented into white matter, grey matter, and cerebrospinal fluid. The grey matter probability
maps are used to restrict the analysis to cortical voxels. The anatomical and functional images
were normalised to Montreal Neurological Institute (MNI) space. No smoothing was applied to
suit the multivariate nature of our analyses. During realignment, motion parameters were
generated. Runs where the framewise displacement (the sum of the absolute values of the
differentiated realignment parameters) exceeded 1 mm were excluded from further analysis.
Statistical Analysis
We applied RSA in a whole-brain searchlight procedure using the CosMoMVPA toolbox
(Oosterhof et al., 2016). Trials were pooled over tasks (overt articulation and covert trials), over
word classes (positive, neutral, negative) and over modalities (auditory and visual). Control
trials where no words were presented were not included. Figure 3E provides an overview of
the searchlight procedure.
Trial-specific activation maps were used as input for this analysis. These maps were gener-
ated by extracting the voxelwise blood oxygen level dependence (BOLD) response from the
normalised, unsmoothed images and calculating the area under the curve of the BOLD
response in every voxel from 2 to 8 s. This procedure is identical to that applied in previous
Neurobiology of Language
266
Affective similarities and word co-occurrences in temporal neocortex
multivariate pattern analysis (MVPA) studies (Bruffaerts et al., 2013; Liuzzi et al., 2015, 2017;
Meersmans et al., 2020, 2022). The number of voxels per searchlight was set a priori to 200
(corresponding to a sphere radius = 11 mm; voxel size equal to 2 × 2 × 2 mm3) and the rec-
ommended data centring was applied before the analysis. The searchlight analysis was limited
to subject-specific grey matter masks, created by thresholding the grey matter probability maps
at 0.3. In every searchlight, the activity pattern was extracted from the activation maps and
pairwise similarities were estimated using cosine similarity. The search sphere in CosMoMVPA
may contain voxels from opposite walls of a sulcus. During the analysis, subject-specific
voxelwise RSA maps were created containing the correlations between the activity similarities
of the searchlight centred on a specific voxel and the word similarity model. The distribution of
the task-based similarities (affective matrix and free association-based matrix) deviated from nor-
mality (Figure 3D; Figure S1). Therefore, Spearman correlation was used in the searchlight RSA for
all models to facilitate between-model comparison. These maps were r to z transformed and
smoothed before being submitted to a group-level t test using SPM12. At every voxel, the t test
evaluates whether the correlation is significantly different from zero across subjects. These t tests
were performed for every searchlight RSA separately. To find significant differences between pairs
of word similarity models, the same maps were submitted to a paired t test. Significance at the
group level was set to a whole-brain familywise error (FWE)-corrected threshold of p < 0.05 (with
uncorrected voxelwise p < 0.001).
Finally, in order to address the unique relation between a specific model (e.g., co-
occurrence) and the neural representation, partial correlation searchlight RSAs were
conducted. The procedure was identical to the one described above, with the only difference
that the correlation between a model representational dissimilarity matrix (RDM; e.g., co-
occurrence) and the neural RDM was controlled for another model (e.g., affective).
RESULTS
On average, subjects made 3.7 (range 0–14) repetition errors out of 264 overt articulation trials
(i.e., wrong word or no repetition) over the total of eight runs. Over all subjects, a total of eight
runs (4.5%) had to be excluded due to excessive head (framewise displacement > 1 mm).
Effect of Co-Occurrence-Based Similarities
Using searchlight RSA, similarities between words estimated using word2vec in spaCy corre-
lated significantly with the similarity of activity patterns in the lateral temporal neocortex sur-
rounding the superior temporal sulcus (STS) from the posterior end to the middle third, to the
left and also to the right (Figure 3F; Tavolo 2). Tavolo 3 provides the Spearman correlation and
95% confidence interval (CI) of the local maxima yielded by the group-level (SPM12) analysis of
the searchlight RSA for co-occurrence similarity. Since the analysis is based on a random-effects
modello, the Spearman correlations provided are the average of the Spearman correlations
obtained in each of the individuals. No other regions showed a significant similarity effect.
We verified the similarity between the word2vec based matrix (spaCy) and the fMRI activity
patterns after partialling out the effect of affective similarity and association-based similarities.
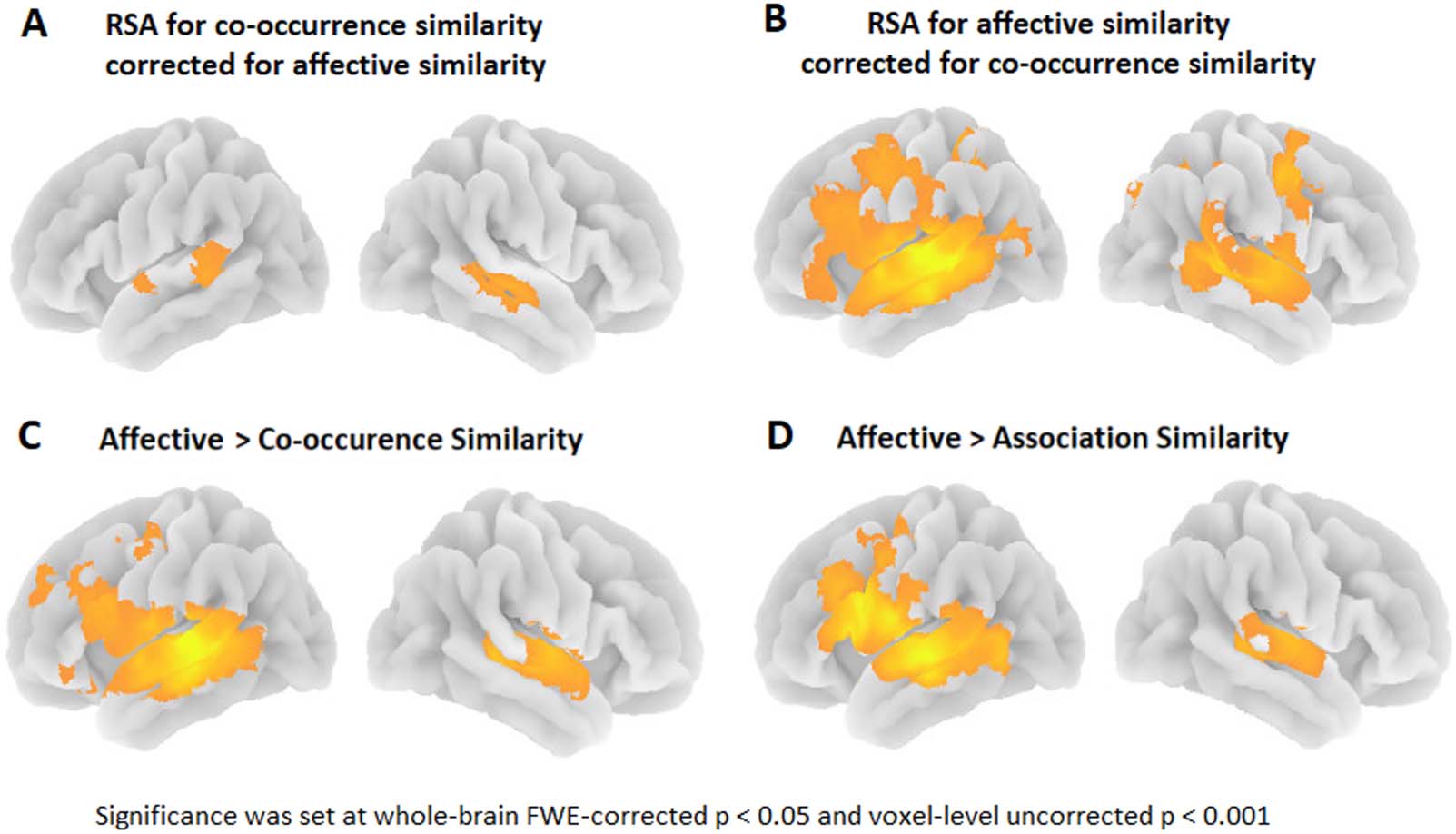
The RSA results remained essentially the same when controlling for affective similarities
(L pSTS peak coordinates: −57, −34, 2; T(21): 4.80; cluster size: 377 voxels; cluster level FWE-
corr p = 0.002; R pSTS peak coordinates: 48, −28, 2; T(21): 4.83; cluster size: 702 voxels; cluster
level FWE-corr p < 0.001; Figure 5A). This is logical given the very low correlation between the co-
occurrence-based model and the affective similarity model. When controlling for the association-
based similarities, no significant effects of the co-occurrence similarities were observed, in line
Neurobiology of Language
267
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
Table 2.
similarities.
Results of the whole-brain searchlight representational similarity analysis (RSA) with word2vec similarities (spaCy) and affective
RSA results for co-occurrence-based similarities
Label
L posterior STS
Size
782
FWE p
<0.001
R posterior STS
1,023
<0.001
RSA results for affective similarities
Label
Left and rightSTS, left MTG, precentral gyrus,
precuneus. Left middle and inferior FC, left FG
Size
13,319
FWE p
<0.001
RSA results for free association-based similarities
Label
Left STS, left rostral AG, left FG
Size
2,423
FWE p
<0.001
Right STS
1,210
<0.001
Peak coordinates
−34
−13
−1
−37
−28
−22
Peak coordinates
−25
−34
−64
Peak coordinates
−34
−43
−1
−37
−16
−28
−57
−54
−69
54
48
57
−69
−51
−36
−57
−42
−66
51
54
48
2
−4
−4
5
5
−1
5
5
−1
−1
−7
−4
−1
−1
5
t(21)
5.48
4.75
4.56
5.37
5.33
5.05
t(21)
8.60
7.54
7.27
t(21)
6.78
5.65
5.05
5.32
5.17
5.12
Note. Spearman correlation was used for all analyses. Significance was set at a whole-brain FWE-corrected threshold of p < 0.05 at the cluster level (after
applying an uncorrected voxelwise threshold p < 0.001). Size: cluster size in number of voxels (2 × 2 × 2 mm3), in MNI space. FWE p: FWE-corrected p-value at
the cluster-level. Abbreviations: STS: superior temporal sulcus; MTG: middle temporal gyrus; FC: frontal cortex; FG: fusiform gyrus; AG: angular gyrus; FWE:
familywise error.
with the moderate degree of collinearity between these two similarity models. As expected based
on the correlogram between the different matrices (Figure 3A), the results remained unchanged
when controlling for phonological similarity or similarity in phonological neighbourhood den-
sity using partial correlations (Table S2A and S2B). It also remained unchanged when control-
ling for taxonomy-based or for experiential similarities (Table S2C and S2D).
Comparison to the Effect of Affective Similarities
For the affective similarities, strong effects were obtained in the lateral temporal perisylvian
neocortex, and more distributed effects outside of lateral temporal cortex were also observed,
namely in left middle and inferior frontal regions and fusiform gyrus (Figure 3F; Table 3F–J),
which provides the Spearman correlation and 95% CI of the local maxima yielded by the
group-level (SPM12) analysis of the searchlight RSA for affective similarity. Using a paired
Neurobiology of Language
268
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
A
f
f
e
c
t
i
v
e
s
i
m
i
l
a
r
i
t
i
e
s
a
n
d
w
o
r
d
c
o
-
o
c
c
u
r
r
e
n
c
e
s
i
n
t
e
m
p
o
r
a
l
n
e
o
c
o
r
t
e
x
Table 3.
Correlational values and 95% confidence interval (CI) based on average correlation value across volunteers.
(A) RSA for co-occurrence similarity
Local maxima
−34
−57
5
Rho
0.026
Lower CI
−0.018
Upper CI
0.066
(F) RSA for affective similarity
Local maxima
−25
−69
5
Rho
0.034
Lower CI
−0.007
Upper CI
0.076
(B) RSA for co-occurrence similarity corrected for affective similarity
(G) RSA for affective similarity corrected for co-occurrence similarity
Local maxima
Rho
Lower CI
Upper CI
Local maxima
Rho
Lower CI
Upper CI
−57
−34
2
0.020
−0.021
0.063
−69
−25
5
0.034
−0.008
0.076
(C) RSA for co-occurrence similarity corrected for association–based similarity
(H) RSA for affective similarity corrected for association–based similarity
Local maxima
Rho
Lower CI
Upper CI
Local maxima
Rho
Lower CI
Upper CI
Not significant
−69
−25
5
0.030
−0.012
0.072
(D) RSA for co-occurrence similarity corrected for taxonomic similarity
(I) RSA for affective similarity corrected for taxonomic similarity
( WordNet)
( WordNet)
Local maxima
Rho
Lower CI
Upper CI
Local maxima
Rho
Lower CI
Upper CI
−57
−34
2
0.026
−0.016
0.068
−69
−25
5
0.033
−0.009
0.075
(E) RSA for co-occurrence similarity corrected for experiential similarity
( J) RSA for affective similarity corrected for experiential similarity
(Lancaster)
(Lancaster)
Local maxima
Rho
Lower CI
Upper CI
Local maxima
Rho
Lower CI
Upper CI
−57
−34
2
0.021
−0.021
0.064
−69
−25
5
0.033
−0.008
0.076
Note. Values are reported for the left posterior temporal peak local maxima based on the group-level (SPM12) of the (A) RSA for co-occurrence similarity, RSA for co-occurrence similarity (B)
corrected for affective similarity, (C) corrected for association-based similarity (D) corrected for taxonomic similarity, (E) corrected for experiential similarity. (F) RSA for affective similarity,
RSA for affective similarity (G) corrected for co-occurrence similarity, (H) corrected for association-based similarity, (I) corrected for taxonomic similarity, ( J) corrected for experiential
similarity.
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
2
6
9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
Figure 5. 3D rendering of the (A) semantic similarity effect for co-occurrence similarity corrected for affective similarity; (B) semantic sim-
ilarity effect for affective similarity corrected for co-occurrence similarity; (C) paired t test showing the correlation strength of affective similarity
stronger than co-occurrence similarity; (D) paired t test showing the correlation strength of affective similarity stronger than association-based
similarity. RSA: representational similarity analysis; FWE: familywise error.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
t-test across subjects, we evaluated significant differences between the models. Correlations
were significantly higher for the affective similarities compared to spaCy- or association-
derived similarities in bilateral superior and middle temporal gyrus and left middle and inferior
frontal gyrus (paired t test; FWE-corrected cluster-level p < 0.05 combined with uncorrected
voxel-level p < 0.001; Figure 5C and D). No regions were identified where spaCy- or
association-derived similarities had a stronger effect than affective similarities. The effects of
affective similarities remained unaltered when partialling out the effects of co-occurrence-
based (peak-coordinates: −69, −25, 5; t (21) = 8.68; cluster size = 13,143 voxels; cluster level
FWE-corr p < 0.001; Figure 5B) or association-based similarities. Similarly, results did not
change after partialling out phonological distance, phonological neighbourhood density,
taxonomy-based and experiential-based similarity (Table S3).
Effect of Free Word Association-Based Similarities
Co-occurrence-based models are influenced by word associations. We also evaluated the
effect of association-based similarities derived from SWOW. In a secondary analysis, we
repeated the RSA using a behavioural similarity matrix derived from free association data
(SWOW data set). The results of the SWOW similarity matrix overlapped with those for the
word2vec matrix in STS but extended more posteriorly into the rostral angular gyrus and infe-
riorly into the fusiform gyrus (LH: 2,423 voxels, peak coordinate −57, −34, −1; RH: 1,210
voxels, peak coordinate 51, −37, −1; Table 2; Figure 3F).
DISCUSSION
There was a striking correspondence between word co-occurrence-based similarities
(word2vec) and similarities in activity patterns in the neocortex surrounding the left STS,
and this was also true for affective similarities. Despite this co-localisation in superior temporal
Neurobiology of Language
270
Affective similarities and word co-occurrences in temporal neocortex
neocortex, the co-occurrence and affective similarity effects in lateral temporal neocortex
were independent, in line with the low correlation between the two similarity matrices. In
contrast, the results obtained with the co-occurrence-based and SWOW word association
model showed a strong resemblance to each other, in line with the correlation between the
word similarities derived from these two types of word association models.
Comparison to Previous fMRI Studies Using Word2vec Modelling
The co-occurrence-based similarities were localised to a region confined to the lateral superior
temporal cortex. This contrasts with some of the earlier studies of co-occurrence-based simi-
larities showing a neuroanatomically distributed correlation between co-occurrence-based
word similarities and the similarity of brain activity patterns. In a landmark study, Mitchell
et al. (2008) described a predictive relation between co-occurrence data and neural activity.
Each object stimulus word was represented as a 25-dimensional vector, with each value cor-
responding to the normalised sentence-wide co-occurrence of that word with 25 sensorimotor
verbs (e.g., see, hear, eat; see also Anderson et al., 2013). While dimensionality reduction is an
integral part of the word2vec approach applied here, the dimensions retained are entirely data
driven based on the corpora analysed (300 dimensions in the spaCy model). The low-
dimensional sensorimotor approach could potentially explain the high congruence between
predicted and observed data in visual, motor, or gustatory cortex in the Mitchell et al. (2008)
study. Representations in lateral temporal cortex seem to be less dependent on sensorimotor
information, but instead rely on language-internal sources. These representations are also bet-
ter modelled in a data-driven space with more dimensions than when dimensions are defined
by the investigator. When the Mitchell et al. (2008) data set was analysed using word2vec,
good concordance was achieved between the model and the overall brain activity pattern
(Abnar et al., 2018). However, in that study the neuroanatomical network underlying this
result was not the main topic of interest and relatively poorly defined.
Another study using co-occurrence-based vectors also reported a more distributed pattern
than the focalised lateral temporal region we found (Pereira et al., 2018). Between-study differ-
ences in the similarity modelling can explain the difference in outcome. In Pereira et al. (2018)
the co-occurrence-based vectors form the basis for spectral clustering that divides the semantic
space in approximately 180 clusters that are then each represented by a representative word
selected by the examiners. Next the authors derive dimensions from the set of representative
words. During the fMRI experiment the words were embedded in sentences that also contained
other words from the same cluster. In contrast to Pereira et al. (2018) the current paper directly
imports the word embeddings as determined by spaCy, without further processing steps. It is
conceivable that the sequence of meaning-oriented procedures in Pereira et al. (2018) may lead
to a semantic representation that is richer in content than the representations based purely on
corpus-derived word embeddings. This may then explain the wider neuroanatomical distribu-
tion of the effects in Pereira et al. (2018) compared to the current findings.
In an approach similar to the current one, Wang et al. (2018) performed a RSA with three
similarity matrices: based on co-occurrences, based on semantic features, and based on sub-
jective ratings. All words were abstract. Co-occurrence-based similarities were correlated with
similarities of activity patterns in perisylvian language regions in temporal and frontal cortex
(based on a volume of interest encompassing the perisylvian language network in its entirety),
whereas the more semantically based similarities resulted in a more distributed pattern. In their
globality, the findings by Wang et al. (2018) indicate that the perisylvian language network
codes for co-occurrence-based similarities. The current findings confirm this observation in
Neurobiology of Language
271
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
a whole-brain RSA in a larger group of participants (22 vs. 6), a wider range of words and input
modalities, and without prior word selection step based on stability of responses. Another
finding in common with Wang et al. (2018) as well as other studies (e.g., Meersmans et al.,
2020) is the strength of the effect of affective similarities. As a major difference with Wang
et al., in a whole-brain search analysis, the co-occurrence-based-similarities were limited to
the cortex surrounding the STS in our study while they were found in ventral and middle pre-
motor cortex in Wang et al. (2018) (mostly in the ventral and middle premotor cortex
according to Figure 4B). Furthermore, the correlation with the co-occurrence-based model
in the cortex surrounding the STS was bilateral in the current study and left lateralised in the
study by Wang et al. (2018). The difference in localisation may be due to a variety of reasons, for
instance, the use of a broader range of words in our study compared to abstract words only in
Wang et al., and the use of auditory along with visual modality.
A fourth study (Fernandino et al., 2022) revealed that representational similarity for co-
occurrence-based similarities was weaker than for taxonomy-based similarities or, even more
so, similarities based on componential experiential semantics (Binder et al., 2016). For that
reason, we repeated the RSA after partialling out taxonomic and experiential similarities.
Results remained essentially the same. The difference between studies can be accounted for
by the difference in composition of the stimulus set. In the current study, words were equally
distributed between three valence classes and selected semiautomatically to maximise the
range of the pairwise semantic similarities controlling for a list of relevant variables. The stim-
ulus set did not contain an evident taxonomic structure and many of the experiential features
of Binder et al. (2016) were not applicable to the words we used. The difference between
studies demonstrates that RSA reflects the structure present in the word set. Comparisons
between the effects of different dimensions within a word set are valid but should not be
interpreted in absolute terms, and the outcome will depend on the structure present in the
experimental set of words. Had we composed a word set where words were evenly distributed
over the traditional semantic categories, it is likely that the effect of taxonomic structure would
have been much more prominent (Fernandino et al., 2022).
Comparison Between the Results for Co-Occurrence and Affective Similarity-Based Models
The lateral temporal region that showed co-occurrence-based similarities in our study also
showed strong affective similarity effects. When the affective similarity effect was partialled
out, the co-occurrence-based effect remained unchanged, as expected given the low correla-
tion between the co-occurrence-based similarities and the affective similarities (Figure 3).
Hence, the cortex surrounding the STS codes for co-occurrences and for affective similarities
likely through relatively separate operational mechanisms.
Affective information is particularly difficult to capture from co-occurrence-based word
embeddings (De Deyne et al., 2021; Tang et al., 2016; Yu et al., 2018): The correlation
between affective and co-occurrences models equalled 0.065, while the correlation between
affective and free word association-based models was 0.310. In the current study, the affective
model resulted in stronger and notably more extensive effects than the co-occurrence model in
the RSA. The effect of affective similarities was present in the STS, replicating previous findings
(Meersmans et al., 2020), but was more widespread in line with the distributed representation
of word meaning (Binder & Desai, 2011; Huth et al., 2016). The effect of co-occurrence-based
similarities cannot be accounted for by the affective similarity effect, as it was unaltered when
partialling out the affective similarity effect. Models that are derived from evaluative judge-
ments by human subjects capture word meaning to a fuller extent than those that are derived
Neurobiology of Language
272
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
from co-occurrences in text corpora, and this is a plausible explanation of the stronger and
more extensive effects. However, the effect of co-occurrence-based similarities remain after
partialling out the affective similarities, and this indicates that the two models represent differ-
ent information about words and not that one is simply a weaker version of the other.
The comparison with SWOW-based similarities is of interest because they occupy a position
somewhat in between co-occurrence-based similarities and affective similarities. SWOW is based
on a cued free association task. It shares with affective similarities the origin in subjective judge-
ments by human participants and with co-occurrence-based similarities its strong link with word
associations. This intermediary position between co-occurrence-based and affective-judgement-
based modelling is also evident from the correlogram (Figure 3A): SWOW-based similarities
correlate both with co-occurrence-based similarities and with affective similarities. When
SWOW word association similarities were partialled out, the co-occurrence-based effect was
no longer significant. Co-occurrence and free word association-based similarities share a substan-
tial amount of information (r = 0.458; Figure 3A), but association-based models are known to
better capture affective information. With the current correlational approach, it is impossible to
disentangle whether the co-occurrence-based or the word-association-based similarities are
driving the effect more strongly given the collinearity between the two matrices (Figure 3A).
Hypothetical Functional-Anatomical Model of Superior Temporal Cortex
Taken together, the findings from the co-occurrence-based and the affective model may be
understood in the context of the theory of a hybrid conceptual system with symbolic and
embodied representations (Andrews et al., 2009; Louwerse & Jeuniaux, 2010; Paivio, 1991).
The co-occurrence-based model yielded strong effects specifically in the left STS, the affective
similarity model yielded even stronger, but independent, effects in this region, and beyond. A
hybrid model could explain why both affective similarity and word2vec similarities correlate
significantly with the superior temporal neocortex, despite a relatively low correlation
between the two similarity matrices (Figure 3A). The two processes, one co-occurrence/
association-based and the other driven by valence as one of the principal dimensions of word
meaning, may operate in a same region in a relatively independent manner.
The lateral temporal region to the left corresponds to what is also known as Wernicke’s
area, although its exact anatomical boundaries are debated (Binder, 2015). Wernicke already
hypothesised about the role of co-occurrences in the formation of associations between
cortical responses (Gage & Hickok, 2005), a mechanism that has since been shown to be
important in processing at distinct linguistic levels (Blank & Davis, 2016; Forseth et al.,
2020; Schuster et al., 2021). The activated zone surrounding the STS has been previously
implicated in human word intelligibility in a very consistent manner (Davis & Johnsrude,
2003; Evans et al., 2014; Obleser & Kotz, 2010; Scott et al., 2000). Word intelligibility requires
integration of multiple, hierarchical levels, from the acoustic over the phonological to the
semantic level (Obleser, 2014) and can be manipulated bottom up (e.g., by filtering the acous-
tic signal) or top down (e.g., predictability based on sentence context; Obleser & Kotz, 2010).
Obviously, in connected speech, statistical regularities are extremely useful for predicting and
recognizing words efficiently despite the high word rate and any degradation of the acoustic
signal that may exist. Even for artificially isolated words, word identification may be guided by
a scaffold of learnt co-occurrences. Here, the terms context and prediction do not refer to
actual experimental context or predictions, but to probabilities learnt from language exposure.
In connected speech, these co-occurrence-based regularities may be very informative for pre-
dictive coding (Arnal et al., 2011; Blank & Davis, 2016).
Neurobiology of Language
273
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
The role of superior temporal cortex in coding word associations/co-occurrences was rel-
atively independent from its role in coding affective similarities. Estimating affective similarities
requires subjective ratings and can be based on human ratings or on dictionaries and lexica of
data, such as emoticon labels (Tang et al., 2016), labelled by experts. The current data confirm
that these affective dimensions, known to be important for meaning representation for decades
(Osgood et al., 1957), are also expressed in the perisylvian lateral temporal cortex (Meersmans
et al., 2020). Intuitively one could have expected a more prominent role of so-called emotion
processing regions rather than language regions for coding affective word similarities. How-
ever, the current findings confirm earlier and consistent observations of the coding of affective
similarities by superior lateral temporal regions (Meersmans et al., 2020). Words with positive
or negative valence are generally more abstract than neutral valence words. Abstract words
more heavily rely on the linguistic system for their meaning than concrete words (Borghi &
Binkofski, 2014; Kousta et al., 2011; Louwerse, 2011; Vigliocco et al., 2014). We have previ-
ously postulated that this may relate to the strong effect of affective similarities in superior
temporal perisylvian language cortex (Meersmans et al., 2020). Neurobiologically, this is a
correlate of the entwinement between affect and language (Wilce, 2009).
Our results do not imply that superior temporal cortex activity patterns are exclusively
explained by either association-based similarities or affective similarities for any set of nouns.
As mentioned before, results of RSA mirror the structure within the stimulus set employed.
Activity patterns in superior temporal language cortex certainly code for other dimensions
too, such as taxonomic structure or compenential experiential semantic structure (Fairhall &
Caramazza, 2013; Fernandino et al., 2022). This by no means contradicts our findings. Which
model yields the best correlations between word similarity and similarity in activity patterns
depends on the type of stimulus set and maybe also the task performed (Meersmans et al.,
2022). For a word set with a high proportion of concrete words and hence a clear taxonomic
structure and strong sensory embodiment, an experiential model may be superior to a co-
occurrence-based model (Fernandino et al., 2022). However such an observation should
not be generalised to all types of word sets. The current stimulus set has been selected semi-
automatically to maximise the range in pairwise similarities and to control for the proportion of
positive, neutral, and negative valence stimuli. It was not generated in order to have a repre-
sentative and consistent sample of traditional semantic categories distributed over the three
valence classes. As a consequence, the current stimulus set did not contain a clear taxonomic
structure and several of the 64 features of a prevailing componential experiential semantic
model (Binder et al., 2016) were not applicable to the current stimulus set. It is important to
put fMRI representational similarity results in the context of the type of word stimuli examined.
As a consequence, exclusionary claims towards the representation of dimensions of word
meaning in the lateral superior temporal language cortex are unwarranted. When the stimulus
set contains a clear categorical structure, categorical effects can be found in superior temporal
cortex (Carota et al., 2021; Devereux et al., 2013), and likewise for taxonomy-based structure
(Fernandino et al., 2022) or experiential sensorimotor strengths (Carota et al., 2021;
Fernandino et al., 2022). The structure within the experimental set of nouns will determine
which dimensions are most strongly represented by superior temporal cortex in response to
these nouns in a dynamic, context-dependent manner.
Limitations
The use of overt articulation during fMRI can potentially introduce motion artefacts. The state-
of-the-art voice recording system with noise cancellation enables the recording of responses in
a sensitive manner even when speech volume is low. Subjects were specifically instructed to
Neurobiology of Language
274
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
n
o
_
a
_
0
0
0
9
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
pronounce the words without moving their mouths excessively and without head motion.
Head motion was evaluated and runs where the a priori threshold for framewise displacement
(>1 mm) was exceeded were excluded from further analysis.
Trials with overt articulation and trials without overt responses were pooled for RSA. Questo
increases the generalisability of our findings. The subjects received the task instruction only
after the word stimulus had been shown so that the initial phase (1,500 ms duration) Di
conscious word processing is matched regardless of response or judgement. For repetition
semantic processing is not necessary (Jefferies et al., 2006) although in the intact brain con-
scious word perception will inevitably activate a certain level of semantic processing. This is
also evidenced by the empirical effects of affective and word association-based similarities
upon the brain activity pattern.
The word sets were selected to be evenly distributed across different word valence classes.
They may therefore not be representative for the total noun thesaurus which is not distributed
evenly across negative, neutro, and positive valence classes. This could introduce a bias in
favour of affective similarity modelling versus word co-occurrence-based modelling. How-
ever, the main finding is the independency of coding between the two types of similarities
rather than the strength of the effect of one versus the other approach.
The affective ratings used to create the three-dimensional affective similarity space were
extrapolated from a smaller set of behavioural ratings based on similarities derived from the
SWOW association data set. As such, the stronger overlap between the association-based and
affective models can be in part explained by this procedure. D'altra parte, this underlines
our central conclusion of independent coding of co-occurrence-based and affective similari-
ties: Using extrapolations could potentially have introduced a similarity-based confound into
the affective estimates.
Behaviourally, there was no correlation between the affective similarity matrix and the
co-occurrence-based similarity. This observation fits with the initial rationale of the current
experiment, namely that co-occurrence-based similarities capture affective similarities poorly
and that for affective similarities to be calculated requires subjective input. The word set
we chose was not specifically selected for an absence of correlation between affective and
co-occurrence-based similarities, nevertheless for other word sets such a correlation may be
found, although we would expect it in general to be relatively weak. The conclusion of func-
tional independence of the superior temporal representation of affective and co-occurrence-
based similarities is based on the absence of a behavioural correlation, the co-localisation as
well as the absence of any effect of partialling out the effects of co-occurrence or affective
similarities, rispettivamente. The absence of an effect of partialling out the effects of co-occurrence
or affective similarities follows mostly from the absence of a correlation between the two
matrices behaviourally. In the future, for word sets where affective and co-occurrence-based
similarities show a weak but significant correlation, it will be worthwhile to examine what
remains after partialling out the effect of the respective matrices.
The visual control stimuli consisted of consonant letter strings and the auditory control stim-
uli of rotated spectrograms. Hence, in the visual modality, the stimuli still contain linguistic
symbols but not in the auditory control stimuli. This is unlikely to affect the results, Quale
are based on pooled analysis of auditory and visual word trials.
The sensitivity of the RSA is difficult to determine accurately as there is no groundtruth. Questo
is the case for most functional imaging studies of cognition. The sensitivity depends on a num-
ber of factors and not only on the number of words examined. Other factors that determine the
sensitivity and specificity are the number of within-subject replications per word (8 in our
Neurobiology of Language
275
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
study), the range of similarities between the words in the word set (maximised in our study
using as semiautomated procedure), the number of individuals scanned, the intertrial interval
duration and the hemodynamic modelling procedure. These multiple factors together deter-
mine sensitivity and specificity.
When association-based similarities were partialled out, the representational similarity
effect for co-occurrence-based similarities disappeared. This emphasises the shared informa-
tion in both word association models. Co-occurrence-based models and free word association
models both tap into word meaning as derived from word context. With a correlational
approach the collinearity between the different measures requires caution for exclusionary
explanations. Hence the findings should not be interpreted in terms of a general superiority
of one type of models versus another in modelling word associations Neither is it possible to
empirically ascertain whether the correlation between the co-occurrence-based matrix and the
fMRI-activity-based similarity matrix reflects any correspondence in underlying computational
mechanisms. The correlation between the co-occurrence-based matrix and the fMRI-activity-
based similarity matrix may also be indirect via the associations between word meanings.
Hence the proposal that co-occurrence-based mechanisms may play a role in coding of word
meaning in lateral temporal cortex remains hypothetical at the current stage.
CONCLUSIONS
The results of this study lead us to hypothesise that the brain uses several behavioural and
distributional mechanisms to construct word meaning, in accordance with a hybrid framework
of embodied and symbolic representations. While experiential information is represented in a
more distributed fashion, corpus-based information correlates with patterns in a focalised
region surrounding the STS. The latter relies on statistical regularities that allow for predictive
coding.
FUNDING INFORMATION
Rik Vandenberghe, Onderzoeksraad, KU Leuven (https://dx.doi.org/10.13039/501100004497),
Award ID: C14/17/108. Rik Vandenberghe, Onderzoeksraad, KU Leuven (https://dx.doi.org/10
.13039/501100004497), Award ID: C14/21/109. Rik Vandenberghe, Fonds Wetenschappelijk
Onderzoek (https://dx.doi.org/10.13039/501100003130), Award ID: G094418N. Antonietta
Gabriella Liuzzi, Fonds Wetenschappelijk Onderzoek (https://dx.doi.org/10.13039
/501100003130), Award ID: 1247821N.
AUTHOR CONTRIBUTIONS
Antonietta Gabriella Liuzzi: Formal analysis: Supporting; Methodology: Supporting; Writing –
revisione & editing: Supporting. Karen Meersmans: Conceptualization: Lead; Data curation:
Lead; Formal analysis: Lead; Investigation: Lead; Methodology: Lead; Writing – original draft:
Lead; Writing – review & editing: Lead. Gerrit Storms: Conceptualization: Supporting; Writing –
revisione & editing: Supporting. Simon De Deyne: Formal analysis: Equal; Methodology: Equal;
Software: Supporting; Writing – review & editing: Supporting. Patrick Dupont: Methodology:
Equal; Software: Equal; Writing – review & editing: Equal. Rik Vandenberghe: Conceptualiza-
zione: Lead; Funding acquisition: Lead; Methodology: Equal; Supervision: Lead; Writing – original
bozza: Equal; Writing – review & editing: Lead.
DATA AVAILABILITY STATEMENT
Data are available on https://github.com/lcn-kul/affective-similarities.
Neurobiology of Language
276
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
REFERENCES
Abnar, S., Ahmed, R., Mijnheer, M., & Zuidema, W. (2018).
Experiential, distributional and dependency-based word embed-
dings have complementary roles in decoding brain activity. In
Proceedings of the 8th Workshop on Cognitive Modeling and
Linguistica computazionale (CMCL 2018) (pag. 57–66). Association
for Computational Linguistics. https://doi.org/10.18653/v1/ W18
-0107
Agrawal, A., An, A., & Papagelis, M. (2018). Learning emotion-
enriched word representations. In Proceedings of the 27th Inter-
national Conference on Computational Linguistics (pag. 950–961).
Associazione per la Linguistica Computazionale.
Anderson, UN. J., Bruni, E., Bordignon, U., Poesio, M., & Baroni, M.
(2013). Of words, eyes and brains: Correlating image-based
distributional semantic models with neural representations of
concepts. Negli Atti del 2013 Conferenza sull'Empirico
Metodi nell'elaborazione del linguaggio naturale (pag. 1960–1970). Asso-
ciation for Computational Linguistics.
Andrews, M., Vigliocco, G., & Vinson, D. (2009). Integrating expe-
riential and distributional data to learn semantic representations.
Psychological Review, 116(3), 463–498. https://doi.org/10.1037
/a0016261, PubMed: 19618982
Arnal, l. H., Wyart, V., & Giraud, UN. l. (2011). Transitions in neural
oscillations reflect prediction errors generated in audiovisual
speech. Nature Neuroscience, 14(6), 797–801. https://doi.org
/10.1038/nn.2810, PubMed: 21552273
Ashburner, J., Barnes, G., Chen, C.-C., Daunizeau, J., Flandin, G.,
Friston, K., Gitelman, D., Glauche, V., Henson, R., Hutton, C.,
Jafarian, A., Kiebel, S., Kilner, J., Litvak, V., Mattout, J., Moran,
R., Penny, W., Phillips, C., Razi, A., … Zeidman, P. (2021).
SPM12 manual. Wellcome Centre for Human Neuroimaging.
Belyk, M., Brown, S., Lim, J., & Kotz, S. UN. (2017). Convergence of
semantics and emotional expression within the IFG pars orbitalis.
NeuroImage, 156, 240–248. https://doi.org/10.1016/j
.neuroimage.2017.04.020, PubMed: 28400265
Bestgen, Y., & Vincze, N. (2012). Checking and bootstrapping
lexical norms by means of word similarity indexes. Behavior
Research Methods, 44, 998–1006. https://doi.org/10.3758
/s13428-012-0195-z, PubMed: 22396137
Binder, J. R. (2015). The Wernicke area: Modern evidence and a
reinterpretation. Neurology, 85(24), 2170–2175. https://doi.org
/10.1212/ WNL.0000000000002219, PubMed: 26567270
Binder, J. R., Conant, l. L., Humphries, C. J., Fernandino, L.,
Simons, S. B., Aguilar, M., & Desai, R. H. (2016). Toward a
brain-based componential semantic representation. Cognitive
Neuropsychology, 33(3–4), 130–174. https://doi.org/10.1080
/02643294.2016.1147426, PubMed: 27310469
Binder, J. R., & Desai, R. H. (2011). The neurobiology of semantic
memory. Trends in Cognitive Sciences, 15(11), 527–536. https://
doi.org/10.1016/j.tics.2011.10.001, PubMed: 22001867
Blank, H., & Davis, M. H. (2016). Prediction errors but not sharp-
ened signals simulate multivoxel fMRI patterns during speech
perception. PLOS Biology, 14(11), Article e1002577. https://doi
.org/10.1371/journal.pbio.1002577, PubMed: 27846209
Borghi, UN. M., & Binkofski, F. (2014). Words as social tools: An
embodied view on abstract concepts. Springer. https://doi.org
/10.1007/978-1-4614-9539-0
Bruffaerts, R., Dupont, P., Peeters, R., De Deyne, S., Storms, G., &
Vandenberghe, R. (2013). Similarity of fMRI activity patterns in
left perirhinal cortex reflects semantic similarity between words.
Journal of Neuroscience, 33(47), 18597–18607. https://doi.org
/10.1523/JNEUROSCI.1548-13.2013, PubMed: 24259581
Brysbaert, M., Stevens, M., De Deyne, S., Voorspoels, W., & Storms,
G. (2014). Norms of age of acquisition and concreteness for
30,000 Dutch words. Acta Psychologia, 150, 80–84. https://doi
.org/10.1016/j.actpsy.2014.04.010, PubMed: 24831463
Carota, F., Nili, H., Pulvermüller, F., & Kriegeskorte, N. (2021).
Distinct fronto-temporal substrates of distributional and taxo-
nomic similarity among words: Evidence from RSA of bold sig-
nals. NeuroImage, 224, Article 117408. https://doi.org/10.1016
/j.neuroimage.2020.117408, PubMed: 33049407
Davis, M. H., & Johnsrude, IO. S. (2003). Hierarchical processing in
spoken language comprehension. Journal of Neuroscience,
23(8), 3423–3431. https://doi.org/10.1523/ JNEUROSCI.23-08
-03423.2003, PubMed: 12716950
De Deyne, S., Navarro, D. J., Collell, G., & Perfors, UN. (2021).
Visual and affective multimodal models of word meaning in
language and mind. Cognitive Science, 45(1), Article e12922.
https://doi.org/10.1111/cogs.12922, PubMed: 33432630
De Deyne, S., Navarro, D. J., Perfors, A., Brysbaert, M., & Storms,
G. (2019). The “Small World of Words”: English word association
norms for over 12,000 cue words. Behavior Research Methods,
51(3), 987–1006. https://doi.org/10.3758/s13428-018-1115-7,
PubMed: 30298265
De Deyne, S., & Storms, G. (2008). Word associations: Network
and semantic properties. Behavior Research Methods, 40(1),
213–231. https://doi.org/10.3758/ BRM.40.1.213, PubMed:
18411545
De Deyne, S., Verheyen, S., Ameel, E., Vanpaemel, W., Dry, M. J.,
Voorspoels, W., & Storms, G. (2008). Exemplar by feature appli-
cability matrices and other Dutch normative data for semantic
concepts. Behavior Research Methods, 40(4), 1030–1048.
https://doi.org/10.3758/BRM.40.4.1030, PubMed: 19001394
Devereux, B. J., Clarke, A., Marouchos, A., & Tyler, l. K. (2013).
Representational similarity analysis reveals commonalities and
differences in the semantic processing of words and objects.
Journal of Neuroscience, 33(48), 18906–18916. https://doi.org
/10.1523/JNEUROSCI.3809-13.2013, PubMed: 24285896
Devereux, B. J., Kelly, C., & Korhonen, UN. (2010). Using fMRI acti-
vation to conceptual stimuli to evaluate methods for extracting
conceptual representations from corpora. Negli Atti del
NAACL HLT 2010 First Workshop on Computational Neurolin-
guistics (pag. 70–78). Associazione per la Linguistica Computazionale.
Diedenhofen, B., & Musch, J. (2015). Cocor: A comprehensive
solution for the statistical comparison of correlations. PLOS
ONE, 10(3), Article e0121945. https://doi.org/10.1371/journal
.pone.0121945, PubMed: 25835001
Evans, S., Kyong, J. S., Rosen, S., Golestani, N., Warren, J. E.,
McGettigan, C., Mourao-Miranda, J., Wise, R. J. S., & Scott,
S. K. (2014). The pathways for intelligible speech: Multivariate
and univariate perspectives. Cerebral Cortex 24(9), 2350–2361.
https://doi.org/10.1093/cercor/bht083, PubMed: 23585519
Fairhall, S. L., & Caramazza, UN. (2013). Brain regions that represent
amodal conceptual knowledge. Journal of Neuroscience, 33(25),
10552–10558. https://doi.org/10.1523/ JNEUROSCI.0051-13
.2013, PubMed: 23785167
Fernandino, L., Tong, J. Q., Conant, l. L., Humphries, C. J., &
Binder, J. R. (2022). Decoding the information structure underly-
ing the neural representation of concepts. Atti del
National Academy of Sciences, 119(6), Article e2108091119.
https://doi.org/10.1073/pnas.2108091119, PubMed: 35115397
Forseth, K. J., Hickok, G., Rollo, P. S., & Tandon, N. (2020). Lan-
guage prediction mechanisms in human auditory cortex. Nature
Neurobiology of Language
277
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
Communications, 11(1), Article 5240. https://doi.org/10.1038
/s41467-020-19010-6, PubMed: 33067457
Gage, N. M., & Hickok, G. (2005). Multiregional cell assemblies,
temporal binding and the representation of conceptual knowl-
edge in cortex: A modern theory by a “classical” neurologist,
Carl Wernicke. Cortex 41(6), 823–832. https://doi.org/10.1016
/S0010-9452(08)70301-0, PubMed: 16353368
Grave, E., Bojanowski, P., Gupta, P., Joulin, A., & Mikolov, T.
(2019). Learning word vectors for 157 languages. Negli Atti
of the Eleventh International Conference on Language Resources
and Evaluation (LREC 2018). European Language Resources
Association (ELRA).
Harris, Z. S. (1954). Distributional structure. WORD, 10(2–3),
146–162. https://doi.org/10.1080/00437956.1954.11659520
Hollis, G., & Westbury, C. (2016). The principals of meaning:
Extracting semantic dimensions from co-occurrence models of
semantics. Psychonomic Bulletin & Review, 23(6), 1744–1756.
https://doi.org/10.3758/s13423-016-1053-2 , PubMed:
27138012
Honnibal, M. (2017). spaCy2: Natural language understanding with
Bloom embeddings, convolutional neural networks and incre-
mental parsing. Sentometrics Research. https://spacy.io/
Huth, UN. G., de Heer, W. A., Griffith, T. L., Theunissen, F. E.., &
Gallant, J. l. (2016). Natural speech reveals the semantic maps
that tile human cerebral cortex. Nature, 532(7600), 453–458.
https://doi.org/10.1038/nature17637, PubMed: 27121839
Jefferies, E., Crisp, J., & Lambon Ralph, M. UN. (2006). The impact of
phonological or semantic impairment on delayed auditory repe-
tition: Evidence from stroke aphasia and semantic dementia.
Aphasiology, 20(9), 963–992. https://doi.org/10.1080
/02687030600739398
Keuleers, E., Brysbaert, M., & Nuovo, B. (2010). SUBTLEX-NL: A new
measure for Dutch word frequency based on film subtitles.
Behavior Research Methods, 42(3), 643–650. https://doi.org/10
.3758/BRM.42.3.643, PubMed: 20805586
Keuleers, E., Stevens, M., Mandera, P., & Brysbaert, M. (2015).
Word knowledge in the crowd: Measuring vocabulary size and
word prevalence in a massive online experiment. Quarterly Jour-
nal of Experimental Psychology, 68(8), 1665–1692. https://doi
.org/10.1080/17470218.2015.1022560, PubMed: 25715025
Kotz, S. A., & Paulmann, S. (2011). Emotion, lingua, and the
brain. Language and Linguistics Compass, 5(3), 108–125.
https://doi.org/10.1111/j.1749-818X.2010.00267.x
Kousta S.-T., Vigliocco, G., Vinson, D. P., Andrews, M., & Del
Campo, E. (2011). The representation of abstract words: Why
emotion matters. Journal of Experimental Psychology: General,
140(1), 14–34. https://doi.org/10.1037/a0021446, PubMed:
21171803
Kuperman, V., Estes, Z., Brysbaert, M., & Warriner, UN. B. (2014).
Emotion and language: Valence and arousal affect word recogni-
zione. Journal of Experimental Psychology: General, 143(3), 1065–
1081. https://doi.org/10.1037/a0035669, PubMed: 24490848
Liuzzi, UN. G., Bruffaerts, R., Dupont, P., Adamczuk, K., Peeters, R.,
De Deyne, S., Storms, G., & Vandenberghe, R. (2015). Left peri-
rhinal cortex codes for similarity in meaning between written
parole: Comparison with auditory word input. Neuropsychologia,
76, 4–16. https://doi.org/10.1016/j.neuropsychologia.2015.03
.016, PubMed: 25795039
Liuzzi, UN. G., Bruffaerts, R., Peeters, R., Adamczuk, K., Keuleers, E.,
De Deyne, S., Storms, G., Dupont, P., & Vandenberghe, R.
(2017). Cross-modal representation of spoken and written word
meaning in left pars triangularis. NeuroImage, 150, 292–307.
https://doi.org/10.1016/j.neuroimage.2017.02.032, PubMed:
28213115
Liuzzi, UN. G., Dupont, P., Peeters, R., Bruffaerts, R., De Deyne, S.,
Storms, G., & Vandenberghe, R. (2019). Left perirhinal cortex
codes for semantic similarity between written words defined from
cued word association. NeuroImage, 191, 127–139. https://doi
.org/10.1016/j.neuroimage.2019.02.011, PubMed: 30753925
Louwerse, M. M. (2011). Symbol interdependency in symbolic and
embodied cognition. Topics in Cognitive Science, 3(2), 273–302.
https://doi.org/10.1111/j.1756-8765.2010.01106.x, PubMed:
25164297
Louwerse, M. M., & Jeuniaux, P. (2010). The linguistic and embodied
nature of conceptual processing. Cognition, 114(1), 96–104.
https://doi.org/10.1016/j.cognition.2009.09.002, PubMed:
19818435
Lynott, D., Connell, L., Brysbaert, M., Brand, J., & Carney, J. (2020).
The Lancaster Sensorimotor Norms: Multidimensional measures
of perceptual and action strength for 40,000 English words.
Behavior Research Methods, 52(3), 1271–1291. https://doi.org
/10.3758/s13428-019-01316-z, PubMed: 31832879
Marian, V., Bartolotti, J., Chabal, S., & Shook, UN. (2012). Clearpond:
Cross-linguistic easy-access resource for phonological and ortho-
graphic neighborhood densities. PLOS ONE, 7(8), Article e43230.
https://doi.org/10.1371/journal.pone.0043230, PubMed:
22916227
McRae, K., Cree, G. S., Seidenberg, M. S., & McNorgan, C. (2005).
Semantic feature production norms for a large set of living and
nonliving things. Behavior Research Methods, 37(4), 547–559.
https://doi.org/10.3758/BF03192726, PubMed: 16629288
Meersmans, K., Bruffaerts, R., Jamoulle, T., Liuzzi, UN. G., De Deyne,
S., Storms, G., Dupont, P., & Vandenberghe, R. (2020). Represen-
tation of associative and affective semantic similarity of abstract
words in the lateral temporal perisylvian language regions.
NeuroImage, 217, Article 116892. https://doi.org/10.1016/j
.neuroimage.2020.116892, PubMed: 32371118
Meersmans, K., Storms, G., De Deyne, S., Bruffaerts, R., Dupont, P.,
& Vandenberghe, R. (2022). Orienting to different dimensions of
word meaning alters the representation of word meaning in early
processing regions. Cerebral Cortex, 32(15), 3302–3317. https://
doi.org/10.1093/cercor/bhab416, PubMed: 34963135
Mugnaio, G. UN. (1995). WordNet: A lexical database for English. Com-
munications of the ACM, 38(11), 39–41. https://doi.org/10.1145
/219717.219748
Mitchell, T. M., Shinkareva, S. V., Carlson, A., Chang, K.-M.,
Malave, V. L., Mason, R. A., & Just, M. UN. (2008). Predicting
human brain activity associated with the meanings of nouns. Sci-
ence, 320(5880), 1191–1195. https://doi.org/10.1126/science
.1152876, PubMed: 18511683
Moors, A., De Houwer, J., Hermans, D., Wanmaker, S., van Schie,
K., Van Harmelen A.-L., De Schryver, M., De Winne, J., &
Brysbaert, M. (2013). Norms of valence, arousal, dominance,
and age of acquisition for 4,300 Dutch words. Behavior Research
Methods, 45(1), 169–177. https://doi.org/10.3758/s13428-012
-0243-8, PubMed: 22956359
Murphy, K., Bodurka, J., & Bandettini, P. UN. (2007). How long to
scan? The relationship between fMRI temporal signal to noise ratio
and necessary scan duration. NeuroImage, 34(2), 565–574.
https://doi.org/10.1016/j.neuroimage.2006.09.032, PubMed:
17126038
Obleser, J. (2014). Putting the listening brain in context. Language
and Linguistics Compass, 8(12), 646–658. https://doi.org/10.1111
/lnc3.12098
Obleser, J., & Kotz S. UN. (2010). Expectancy constraints in degraded
speech modulate the language comprehension network. Cerebral
Cortex, 20(3), 633–640. https://doi.org/10.1093/cercor/bhp128,
PubMed: 19561061
Neurobiology of Language
278
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Affective similarities and word co-occurrences in temporal neocortex
Oosterhof, N. N., Connolly, UN. C., & Haxby, J. V. (2016).
CoSMoMVPA: Multi-modal multivariate pattern analysis of neu-
roimaging data in Matlab/GNU octave. Frontiers in Neuroinfor-
matics, 10, Article 27. https://doi.org/10.3389/fninf.2016.00027,
PubMed: 27499741
Optoacoustics. (2022). OptoActive II headphones and microphones
[Apparatus]. https://www.optoacoustics.com/medical/
Osgood, C. E., Suci, G. J., & Tannenbaum, P. H. (1957). The mea-
surement of meaning. University of Illinois Press Urbana.
Paivio, UN. (1991). Dual coding theory: Retrospect and current
status. Canadian Journal of Psychology/ Revue Canadienne de
Psychologie, 45(3), 255–287. https://doi.org/10.1037/h0084295
Pauligk, S., Kotz, S. A., & Kanske, P. (2019). Differential impact of
emotion on semantic processing of abstract and concrete words:
ERP and fMRI evidence. Scientific Reports, 9(1), Article 14439.
https://doi.org/10.1038/s41598-019-50755-3, PubMed:
31594966
Pereira, F., Botvinick, M., & Detre, G. (2013). Using Wikipedia
to learn semantic feature representations of concrete concepts
Intelligenza, 194,
in neuroimaging experiments. Artificial
240–252. https://doi.org/10.1016/j.artint.2012.06.005, PubMed:
23243317
Pereira, F., Lou, B., Pritchett, B., Ritter, S., Gershman, S. J.,
Kanwisher, N., Botvinick, M., & Fedorenko, E. (2018). Toward
a universal decoder of linguistic meaning from brain activation.
Nature Communications, 9(1), Article 963. https://doi.org/10
.1038/s41467-018-03068-4, PubMed: 29511192
Recchia, G., & Louwerse, M. M. (2015). Reproducing affective
norms with lexical co-occurrence statistics: Predicting valence,
arousal, and dominance. Quarterly Journal of Experimental
Psychology, 68(8), 1584–1598. https://doi.org/10.1080
/17470218.2014.941296, PubMed: 24998307
Schuster, S., Himmelstoss, N. A., Hutzler, F., Richlan, F., Kronbichler,
M., & Hawelka, S. (2021). Cloze enough? Hemodynamic effects
of predictive processing during natural reading. NeuroImage,
228, Article 117687. https://doi.org/10.1016/j.neuroimage.2020
.117687, PubMed: 33385553
Scott, S. K., Blank, C. C., Rosen, S., & Wise, R. J. (2000). Identifica-
tion of a pathway for intelligible speech in the left temporal lobe.
Brain, 123(12), 2400–2406. https://doi.org/10.1093/ brain/123
.12.2400, PubMed: 11099443
Tang, D., Wei, F., Qin, B., Yang, N., Liu, T., & Zhou, M. (2016).
Sentiment embeddings with applications to sentiment analysis.
IEEE Transactions on Knowledge and Data Engineering, 28(2),
496–509. https://doi.org/10.1109/TKDE.2015.2489653
Troche, J., Crutch, S. J., & Reilly, J. (2017). Defining a conceptual
topography of word concreteness: Clustering properties of emo-
zione, sensation, and magnitude among 750 English words.
Frontiers in Psychology, 8, Article 1787. https://doi.org/10.3389
/fpsyg.2017.01787, PubMed: 29075224
Vankrunkelsven, H., Verheyen, S., Storms, G., & De Deyne, S.
(2018). Predicting lexical norms: A comparison between a word
association model and text-based word co-occurrence models.
Journal of Cognition, 1(1), Article 45. https://doi.org/10.5334
/joc.50, PubMed: 31517218
Van Rensbergen, B., De Deyne, S., & Storms, G. (2016). Estimating
affective word covariates using word association data. Behavior
Research Methods, 48(4), 1644–1652. https://doi.org/10.3758
/s13428-015-0680-2, PubMed: 26511372
Van Rensbergen, B., Storms, G., & De Deyne, S. (2015). Examining
assortativity in the mental lexicon: Evidence from word associa-
zioni. Psychonomic Bulletin & Review, 22(6), 1717–1724. https://
doi.org/10.3758/s13423-015-0832-5, PubMed: 25893712
Vigliocco, G., Kousta S.-T., Della Rosa, P. A., Vinson, D. P.,
Tettamanti, M., Devlin, J. T., & Cappa, S. F. (2014). The neural
representation of abstract words: The role of emotion. Cerebral
Cortex, 24(7), 1767–1777. https://doi.org/10.1093/cercor
/bht025, PubMed: 23408565
Vincze, N., & Bestgen, Y. (2011). Une procédure automatique pour
étendre des normes lexicales par l’analyse des cooccurrences dans
des textes. Traitement Automatique des Langues, 52(3), 191–216.
Wang, X., Wu, W., Ling, Z., Xu, Y., Fang, Y., Wang, X., Binder, J. R.,
Men, W., Gao J.-H., & Bi, Y. (2018). Organizational principles of
abstract words in the human brain. Cerebral Cortex, 28(12),
4305–4318. https://doi.org/10.1093/cercor/ bhx283, PubMed:
29186345
Warriner, UN. B., Kuperman, V., & Brysbaert, M. (2013). Norms of
valence, arousal, and dominance for 13,915 English lemmas.
Behavior Research Methods, 45(4), 1191–1207. https://doi.org
/10.3758/s13428-012-0314-x, PubMed: 23404613
Westbury, C., Keith, J., Briesemeister, B. B., Hofmann, M. J., &
Jacobs, UN. M. (2015). Avoid violence, rioting, and outrage;
approach celebration, delight, and strength: Using large text
corpora to compute valence, arousal, and the basic emotions.
Quarterly Journal of Experimental Psychology, 68(8), 1599–1622.
https://doi.org/10.1080/17470218.2014.970204, PubMed:
26147614
Wilce, J. (2009). Language and emotion. Cambridge University Press.
Wu, Z., & Palmer, M. (1994). Verb semantics and lexical selection.
In Proceedings of the 32nd annual meeting of the Association
for Computational Linguistics (pag. 133–138). Associazione per
Computing Machinery. https://doi.org/10.3115/981732.981751
Yu, L.-C., Wang, J., Lai, K. R., & Zhang, X. (2018). Refining word
embeddings using intensity scores for sentiment analysis. IEEE/
ACM Transactions on Audio, Speech, Language Processing,
26(3), 671–681. https://doi.org/10.1109/TASLP.2017.2788182
Neurobiology of Language
279
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
4
2
2
5
7
2
0
7
9
0
2
6
N
o
_
UN
_
0
0
0
9
5
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3