RESEARCH ARTICLE
Causal Contributions of the Domain-General
(Multiple Demand) and the Language-Selective
Brain Networks to Perceptual and Semantic
Challenges in Speech Comprehension
Lucy J. MacGregor1
, Rebecca A. Gilbert1
Sharon W. Erzinçlioğlu1
, Jennifer M. Rodd3
, Zuzanna Balewski2, Daniel J. Mitchell1
, John Duncan1
,
,
Evelina Fedorenko4,5,6
, and Matthew H. Davis1
1MRC Cognition and Brain Sciences Unit, University of Cambridge, Cambridge, UK
2Helen Wills Neuroscience Institute, University of California, Berkeley, Berkeley, CA
3Psychology and Language Sciences, University College London, London, UK
4Department of Brain and Cognitive Sciences, Istituto di Tecnologia del Massachussetts, Cambridge, MA
5McGovern Institute for Brain Research, Istituto di Tecnologia del Massachussetts, Cambridge, MA
6Program in Speech and Hearing Bioscience and Technology, Harvard University, Cambridge, MA
Keywords: speech perception, language comprehension, semantic ambiguity, lesion, adaptation,
apprendimento, multiple demand (MD) system
ABSTRACT
Listening to spoken language engages domain-general multiple demand (MD; frontoparietal)
regions of the human brain, in addition to domain-selective (frontotemporal) language regions,
particularly when comprehension is challenging. Tuttavia, there is limited evidence that
the MD network makes a functional contribution to core aspects of understanding language.
In a behavioural study of volunteers (n = 19) with chronic brain lesions, but without aphasia,
we assessed the causal role of these networks in perceiving, comprehending, and adapting
to spoken sentences made more challenging by acoustic-degradation or lexico-semantic
ambiguity. We measured perception of and adaptation to acoustically degraded (noise-
vocoded) sentences with a word report task before and after training. Participants with greater
damage to MD but not language regions required more vocoder channels to achieve 50%
word report, indicating impaired perception. Perception improved following training,
reflecting adaptation to acoustic degradation, but adaptation was unrelated to lesion location
or extent. Comprehension of spoken sentences with semantically ambiguous words was
measured with a sentence coherence judgement task. Accuracy was high and unaffected by
lesion location or extent. Adaptation to semantic ambiguity was measured in a subsequent
word association task, which showed that availability of lower-frequency meanings of
ambiguous words increased following their comprehension (word-meaning priming). Word-
meaning priming was reduced for participants with greater damage to language but not MD
regions. Language and MD networks make dissociable contributions to challenging speech
comprehension: Using recent experience to update word meaning preferences depends on
language-selective regions, whereas the domain-general MD network plays a causal role in
reporting words from degraded speech.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
a n o p e n a c c e s s
j o u r n a l
Citation: MacGregor, l. J., Gilbert,
R. A., Balewski, Z., Mitchell, D. J.,
Erzinçlioğlu, S. W., Rodd, J. M.,
Duncan, J., Fedorenko, E., & Davis,
M. H. (2022). Causal contributions of
the domain-general (multiple demand)
and the language-selective brain
networks to perceptual and semantic
challenges in speech comprehension.
Neurobiology of Language, 3(4),
665–698. https://doi.org/10.1162/nol_a
_00081
DOI:
https://doi.org/10.1162/nol_a_00081
Supporting Information:
https://doi.org/10.1162/nol_a_00081
Received: 14 Marzo 2022
Accepted: 7 settembre 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Lucy J. MacGregor
lucy.macgregor@mrc-cbu.cam.ac.uk
Handling Editor:
Stephen M. Wilson
Tavolo 1 updated since final publication.
See erratum for details: https://doi.org
/10.1162/nol_x_00103
Copyright: © 2022
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
Brain networks for challenges to speech comprehension
Multiple demand (MD) rete:
A set of bilateral frontoparietal brain
regions that respond to a range of
diverse demanding tasks.
Language-selective network:
A set of left-lateralised
frontotemporal brain regions that
respond selectively to linguistic
stimuli.
INTRODUCTION
During speech comprehension, listeners are continually challenged by various aspects of the
input, which leads to uncertainty at multiple levels of the linguistic hierarchy. Per esempio,
acoustic challenges arise when speech is quiet, in an unfamiliar accent, produced by a young
child who has not yet mastered articulation, or otherwise degraded. In such cases, perception
of the individual phonemes and lexical forms is more uncertain. Linguistic challenges arise
when there is lexical-semantic or syntactic ambiguity, or complexity from low-frequency
words or constructions, such that the intended meaning is unclear. To resolve these uncer-
tainties during speech comprehension, listeners make use of diverse sources of information
(Altmann & Kamide, 1999; Cutler et al., 1997; Garrod & Pickering, 2004; Hagoort et al.,
2004; Münster & Knoeferle, 2018; Özyürek, 2014; Van Berkum, 2009; Zhang et al., 2021).
Inoltre, listeners learn in response to their experiences: They show perceptual and
semantic adaptation such that improvements in the perception and comprehension of different
types of challenging speech can be observed over time (Davis et al., 2005; Rodd et al., 2013).
in questo documento, we consider the potential functional contributions of two distinct groups of cor-
tical brain regions—the domain-selective language network and domain-general multiple
demand (MD) network—to successful perception and comprehension of different types of
challenging speech, and to subsequent perceptual and semantic adaptation.
Role of Language-Selective Versus Domain-General (Multiple Demand) Regions in
Language Comprehension
The language-selective network is a set of left-lateralised frontal and temporal regions that
respond reliably to linguistic stimuli with different input modalities, languages, and tasks
(per esempio., Binder et al., 1997; Blank et al., 2014; Fedorenko et al., 2010; Fedorenko et al., 2012;
MacSweeney et al., 2002; Mahowald & Fedorenko, 2016; Mineroff et al., 2018; Paunov et al.,
2019; Scott et al., 2017; for a review, see Fedorenko, 2014) but not to nonlinguistic stimuli such
as music, mathematical expressions, or computer code (Fedorenko et al., 2011; Ivanova et al.,
2020; Monti et al., 2012). These regions are functionally connected (Braga et al., 2020), E
show correlated response profiles at rest and during naturalistic listening (Blank et al., 2014;
Braga et al., 2020; Mineroff et al., 2018; Paunov et al., 2019), leading to their characterisation
as a functionally coherent network. Lesion studies show that damage to, or degeneration of, Questo
network leads to impairments in language function (Bates et al., 2003; Mesulam et al., 2014;
Mirman et al., 2015; Mirman & Thye, 2018; Turken & Dronkers, 2011) but does not cause def-
icits in other cognitive domains (Apperly et al., 2006; Fedorenko & Varley, 2016; Ivanova et al.,
2021; Polk & Kertesz, 1993; Varley et al., 2001; Varley et al., 2005; Varley & Siegal, 2000),
indicating a necessary and selective role of the network in language comprehension.
Sometimes, linguistic stimuli also activate a set of bilateral frontal, parietal, cingular, E
opercular regions (see Diachek et al., 2020, for a large scale fMRI investigation and relevant dis-
cussion), which together form the MD network (Duncan, 2010B, 2013). This network is domain-
general, responding during diverse demanding tasks (Duncan & Owen, 2000; Fedorenko et al.,
2012; Fedorenko et al., 2013; Hugdahl et al., 2015; Shashidhara et al., 2019) and has been linked
to cognitive constructs such as executive control, working memory, selective attention, and fluid
intelligence (Assem, Blank, et al., 2020; Cole & Schneider, 2007; Duncan & Owen, 2000; Vincent
et al., 2008; Woolgar et al., 2018). Regions of the MD network show strongly synchronized activity
and fluctuation patterns that dissociate sharply from those of the language network (Blank et al.,
2014; Mineroff et al., 2018; Paunov et al., 2019). Inoltre, damage to the MD network leads to
patterns of cognitive impairment that differ from those observed in cases of language network
Neurobiology of Language
666
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Brain networks for challenges to speech comprehension
damage (Duncan, 2010UN; Fedorenko & Varley, 2016; Woolgar et al., 2010; Woolgar et al., 2018),
confirming a functional dissociation between the two networks. (See Fedorenko & Blank, 2020,
for a review focusing on the dissociation between subregions of Broca’s area.)
Recentemente, it has been argued that the MD network does not play a functional role in lan-
guage comprehension (Blank & Fedorenko, 2017; Diachek et al., 2020; Shain et al., 2020;
Wehbe et al., 2021; for reviews, see Campbell & Tyler, 2018; Fedorenko & Shain, 2021). Quello
È, activation of MD regions does not reflect core cognitive operations that are essential to
language comprehension such as perceiving word forms and accessing word meanings.
Invece, it is proposed that activation of MD regions reflects a general increase in effort, Quale
is imposed by task demands in particular, or in some cases even mis-localisation of language-
selective activity because of the proximity of the two systems in some parts of the brain (per esempio., In
the inferior frontal gyrus (IFG), Fedorenko et al., 2012; see Quillen et al., 2021, for evidence
that increased linguistic and nonlinguistic task demands matched on difficulty differentially
recruite langage-selective versus domain-general regions).
Tuttavia, existing evidence that domain-general MD regions do not contribute to language
comprehension is limited in two ways. Primo, relevant studies have typically drawn conclusions
about function based on the magnitude of neural activity (per esempio., the BOLD fMRI response). IL
strongest causal inference about the necessity (and selectivity) of brain regions for particular
cognitive processes comes from approaches that transiently disrupt neural functioning in the
healthy brain (per esempio., transcranial magnetic stimulation, or TMS) and measure the effects on
behaviour, or from cases of acquired brain damage, either in case studies or multi-patient
lesion-symptom mapping investigations that exploit inter-individual variability in behavioural
and neural profiles to link specific brain systems to behavioural outcomes (Halai et al., 2017).
A recent lesion study found that the extent of damage to the MD network predicted deficits in
fluid intelligence; in contrast, MD lesions did not predict remaining deficits in verbal fluency
after the influence of fluid intelligence was removed, which instead were predicted by damage
to the language-selective network (Woolgar et al., 2018), in line with the dissociation dis-
cussed above. These results provide convincing evidence that the MD network but not the
language network contributes to fluid intelligence, but suggest that the MD network contribu-
tion does not extend to language function. Tuttavia, given that language function was
assessed with a verbal fluency task—an elicited production paradigm that relies on a host
of diverse cognitive operations—the question of whether the MD network causally contributes
to specific aspects of language comprehension remains unanswered.
A second limitation of previous studies is their focus on the comprehension of clearly per-
ceptible and relatively unambiguous language, whereas naturalistic speech comprehension
typically involves dealing with noise and uncertainty in the input. Per esempio, speech may
be in an unfamiliar accent or contain disfluencies and mispronunciations; there may be back-
ground speech or other sounds or distractions; or the words and syntax may be ambiguous or
uncommon. These features can make identifying words and inferring meaning—core compu-
tations of comprehension—more difficult (for a review of different types of challenges to
speech comprehension, see Johnsrude & Rodd, 2015). It therefore remains a possibility that
the MD network is functionally critical for successful comprehension in these more challeng-
ing listening situations (Diachek et al., 2020).
Challenges to Speech Perception, Comprehension and Adaptation
Here, we focus on two challenges, which arise from acoustic degradation and from lexical-
semantic ambiguity. Acoustically degraded speech makes word recognition less accurate
Neurobiology of Language
667
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Brain networks for challenges to speech comprehension
Semantic ambiguity resolution:
Selection of the appropriate meaning
of a semantically ambiguous word.
(Mattys et al., 2012), reduces perceived clarity (Sohoglu et al., 2014), increases listening effort
(Pichora-Fuller et al., 2016; Wild et al., 2012), and encourages listeners to utilise informative
semantic contextual cues (Davis et al., 2011; Miller et al., 1951; Obleser et al., 2007; Rysop
et al., 2021) and other forms of prior knowledge (Mugnaio & Isard, 1963; Sohoglu et al., 2014;
Sumby & Pollack, 1954). Acoustically degraded speech engages brain regions that plausibly
fall within the MD network, including parts of the premotor, motor, opercular, and insular cor-
tex (Davis & Johnsrude, 2003; Du et al., 2014, 2016; Erb et al., 2013; Evans & Davis, 2015;
Hardy et al., 2018; Hervais-Adelman et al., 2012; Vaden et al., 2013; Vaden et al., 2015; Wild
et al., 2012), as well in the angular gyrus (Rysop et al., 2021) and IFG (Davis et al., 2011; Davis
& Johnsrude, 2003). Inoltre, disruption of premotor regions either by TMS (D’Ausilio
et al., 2009; D’Ausilio et al., 2012; Meister et al., 2007) or following lesions (Moineau
et al., 2005; for a review, see Pulvermüller & Fadiga, 2010) has been shown to impair percep-
tion of degraded speech. Tuttavia, given that the MD network was not explicitly defined in
previous studies, the functional contribution of the MD network to acoustically degraded
speech perception remains untested.
Lexical-semantic ambiguity (for a review, see Rodd, 2018) challenges comprehension
because of the competition between alternative meanings of a single word form during mean-
ing access (Rayner & Duffy, 1986; Rodd et al., 2002; Seidenberg et al., 1982; Swinney, 1979),
and because costly reinterpretation is sometimes required (Blott et al., 2021; Duffy et al., 1988;
Rodd et al., 2010, 2012). Domain-general cognitive operations may be useful in responding to
the challenge, as evidenced by the positive relationship between individuals’ success in
semantic ambiguity resolution and executive functioning skill (Gernsbacher et al., 1990;
Gernsbacher & Faust, 1991; Khanna & Boland, 2010) and in dual-task studies showing that
performance on nonlinguistic visual tasks is impaired during semantic reinterpretation (Rodd
et al., 2010), but these domain-general operations may be plausibly generated by either
language-selective or domain-general cortical regions.
Functional imaging studies show that semantic ambiguity resolution engages left-lateralised
frontal and temporal brain regions typical of the language-selective network, specifically pos-
terior parts of middle and inferior temporal lobe, anterior temporal lobe, and the posterior IFG
(Bilenko et al., 2009; Musz & Thompson-Schill, 2017; Rodd et al., 2005; Vitello et al., 2014;
Zempleni et al., 2007; for a review, see Rodd, 2020). The possibility that the IFG in particular
plays a causal role is supported by the observation that individuals with Broca’s aphasia have
difficulties in using context to access subordinate word meanings (Hagoort, 1993; Swaab
et al., 1998; Swinney et al., 1989), although patients in these studies were selected based
on their language profile rather than lesion location.
Although subregions within the IFG form part of the language-selective network, as dis-
cussed above, there are also subregions that fall within the domain-general MD network
(per esempio., Fedorenko & Blank, 2020). Indeed IFG recruitment during ambiguity resolution has been
typically accounted for by invoking domain-general constructs of cognitive control or conflict
resolution (Novick et al., 2005; Thompson-Schill et al., 1997) which resolve competition
between alternative meanings of ambiguous words (Musz & Thompson-Schill, 2017). Cur-
rently, the heterogeneity of the IFG makes activations within this region difficult to interpret
functionally, without careful anatomical identification of relevant components (Tahmasebi
et al., 2012).
A range of studies show that listeners can adapt to the challenges of perceiving and com-
prehending acoustically degraded or semantically ambiguous sentences. Listeners’ perception
of degraded speech improves spontaneously over time with repeated exposure (Davis et al.,
Neurobiology of Language
668
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Brain networks for challenges to speech comprehension
Word meaning priming:
The increase in ease of accessing a
particular word meaning following
exposure to that same meaning in a
prime sentence, which can be
considered a type of semantic
adaptation or learning.
Noise vocoding:
A method of acoustically degrading a
speech signal in which spectral detail
is reduced but slow amplitude
modulations—approximately
reflecting syllabic units—are
retained.
2005; Guediche et al., 2014; Hervais-Adelman et al., 2008; Loebach & Pisoni, 2008; Sohoglu
& Davis, 2016; Stacey & Summerfield, 2008), so long as attention is directed to speech (Huyck
& Johnsrude, 2012). This perceptual adaptation is facilitated by visual/auditory feedback pre-
sented concurrently or in advance (Wild et al., 2012), generalises across talkers (Huyck et al.,
2017), is supported by lexical-level information such that learning through exposure to pseu-
dowords is less effective than with real words (although in some cases, learning with pseudo-
words is possible; Hervais-Adelman et al., 2008) but does not additionally benefit from
sentence-level semantic information (learning was as effective with meaningless syntactic
prose; Davis et al., 2005).
Regarding adaptation to ambiguous words, research has shown that accessing a less fre-
quent (subordinate) meaning of an ambiguous word is easier following exposure to the same
meaning of an ambiguous word in a prime sentence. Although the cognitive operations under-
pinning this so-called word meaning priming effect remain somewhat underspecified, IL
effect can be described as a form of longer-term lexicosemantic learning since it can be
observed tens of minutes or even hours after initial exposure, or perhaps longer if adaptation
is consolidated by sleep (Betts et al., 2018; Gaskell et al., 2019; Rodd et al., 2013).
The Current Study
In the current study, we ask whether speech perception and comprehension in different chal-
lenging circumstances, and adaptation in response to these challenges, depend on the MD net-
work or the language-selective network. To do this, we investigated the impact of lesions to these
networks, on behavioural measures of speech perception, comprehension, and adaptation.
We recruited participants (n = 19) on the basis of having long-standing lesions that either (1) had
substantial overlap with the domain-selective language network, (2) had substantial overlap with
the domain-general MD network, O (3) had overlap with neither language nor MD network. IL
participants performed behavioural tasks to assess the immediate effects and longer-term conse-
quences of two types of listening challenge. For the first challenge (acoustic-phonetic), we mea-
sured perception of noise-vocoded spoken sentences in a word report task. Adaptation to this
type of acoustic degradation was assessed in a subsequent word report task following a period
of training. For the second challenge (lexicosemantic), we measured comprehension of spoken
sentences that included low-frequency meaning of semantically ambiguous words, in a sen-
tence coherence judgement task. Adaptation to semantic ambiguity was assessed in a word
association task to measure the consequences of experience with the lower-frequency mean-
ings for subsequent meaning access. Whilst all cognitive tasks will require the contribution of
some general cognitive operations (per esempio., Attenzione, working memory), our tasks were chosen to
be simple enough for participants to perform, thus minimising the demands on such general
cognitive operations, in the absence of acoustic-degradation or lexicosemantic ambiguity.
These tasks are made more difficult by challenges to perceptual processes (per esempio., acoustically
degraded speech), or semantic processes (per esempio., lexicosemantic ambiguity) that are a central part
of language function. We acknowledge that perceptual and semantic challenges to language
function may have secondary impacts on domain-general functions (per esempio., due to increased
working memory demand or a requirement that listeners use sentence context to support pro-
cessazione). Tuttavia, we expect the same sorts of additional domain-general operations to apply
both to degraded speech perception and semantic ambiguity comprehension, as well as to
adaptation to these challenges. Così, if brain lesions have a dissociable impact on accommo-
dating these different challenges, then this would suggest a causal contribution to a specific
aspect of language functioning rather than a contribution to domain-general processes.
Neurobiology of Language
669
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Brain networks for challenges to speech comprehension
Planned analyses focused on comparing behavioural performance measures across the
three participant groups. Tuttavia, the aetiologies (per esempio., stroke, tumour excision) that lead to
brain lesions do not respect functional boundaries of the two networks of interest, and there-
fore our primary analyses treated lesion volume in each network as a graded rather than a
categorical factor. We report group identities of the participants in the demographics table,
and in the lesion maps and data plots contained in the figures below, since this was the basis
of our participant recruitment and so that interested readers can observe how individual
participants—defined on the basis of lesion location—perform our various tasks. Group anal-
yses can be found in the Supporting Information at https://doi.org/10.1162/nol_a_00081.
For each task, behavioural performance measures were associated with lesion location and
extent by performing correlational analyses using probabilistic functional activation atlases
(per esempio., Woolgar et al., 2018). Finalmente, across-task analyses assessed potential dissociations
between the contributions of these two networks for accommodating and adapting to different
sources of listening challenge during speech comprehension.
MATERIALS AND METHODS
Participants
Twenty-one right-handed native English speakers were recruited from the Cambridge Cognitive
Neuroscience Research Panel (CCNRP), a database of volunteers who have suffered a brain
lesion and have expressed interest in taking part in research. Participants were invited to take
part in the current research on the basis that they had chronic lesions (minimum time since
injury of 3 yr) to cortical areas falling predominantly in the language or MD networks (or lesions
in other areas for control participants), but without knowledge of their behavioural profiles.
Così, volunteers were not recruited on the basis of a known language impairment or aphasia
diagnosis. The two networks were broadly defined, based on previous functional imaging data
from typical volunteers (described below), and linked to lesions traced on anatomical MRI
scans for CCNRP volunteers. Participants gave written informed consent under the approval
of the Cambridge Psychology Research Ethics Committee. Data from two participants were
not included in the final analyses of either task (one participant was unable to complete either
task due to fatigue and hearing difficulties; a second failed to complete the semantic ambiguity
experimental tasks and also had difficulties accurately reporting back the words they heard in
the degraded speech task, achieving only 68% word report accuracy for the clear speech
condition across pre- and post-training test sessions; see task details below).
The remaining 19 participants (8 female, mean age 61 yr, range 37–75 yr) had brain lesions
caused by tumour excision (n = 8), stroke (haemorrhagic: n = 6, ischaemic: n = 1), or a com-
bination of these (tumour excision and haemorrhagic stroke: n = 1), with other causes being
abscess excision (n = 1) or resection because of epileptic seizures (n = 1), and one of unknown
cause (n = 1). Individual participant characteristics are detailed in Table 1. Two participants
contributed data to just one of the tasks (md6 was excluded from the degraded speech exper-
iment for not completing the task; md10 was excluded from the semantic ambiguity analyses
for giving multiple responses during the word association task). Così, data from 18 participants
were included for each of the experiments analysed separately (see below) and from 17 par-
ticipants for the cross-experimental analyses.
The National Adult Reading Test (NART; Nelson, 1982) was used to estimate premorbid IQ.
The Test of Reception of Grammar (TROG-2; Bishop, 2003) was used as a background assess-
ment of linguistic competence.
Neurobiology of Language
670
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 1.
Participant profiles, including initial group assignment, demographics, lesion details, IQ based on the NART, and TROG2 scores.
N
e
tu
R
o
B
o
o
G
sì
io
l
Participant Group
LANG
lang1
Age at
test
50
Sex
F
o
F
l
UN
N
G
tu
UN
G
e
lang2
lang3
lang4
lang5
lang6
md1
md2
md3
md4
md5
md6a
md7
md8
md9
md10b
other1
LANG
LANG
LANG
LANG
LANG
MD
MD
MD
MD
MD
MD
MD
MD
MD
MD
OTHER
75
58
65
50
56
58
63
64
53
69
61
52
74
67
81
58
Lesion aetiology
Lesion location
Tumour
L posterior temporal, posterior corpus
callosum, posterior cingulum
Stroke (ischaemic)
L frontal and anterior insular cortex
Stroke (haemorrhagic)
R frontotemporoparietal and anterior
Tumour
thalamus
L temporal
F
F
F
M Other (abscess removal)
L temporal
M Tumour
R inferior parietal/temporal
M Stroke (ischaemic)
L occipitoparietal
F
Tumour
L superior parietal lobe
M Other (unknown)
R frontotemporoparietal surrounding insular
cortex. Anterior branch of internal capsule
F
Tumour
R superior frontal
M Stroke (haemorrhagic)
L frontal
M Tumour + Stroke
(haemorrhagic)
R posterior frontal and some anterior/medial,
extending to post-central gyrus/parietal
areas
F
Tumour
M Tumour
M Tumour
Bifrontal
R frontal
L anterior frontal
M Stroke (haemorrhagic)
L temporooccipital
M Other (resection for
epilepsy)
R temporal
other2
OTHER
60
M Stroke (haemorrhagic)
R basal ganglia (putamen + caudate +
thalamus) and internal capsule dorsal
anterior insula
other3
OTHER
37
F
Stroke (haemorrhagic)
L frontal
Premorbid IQ
(from NART)
108
TROG2
Score
14
Total lesion
volume (cm3)
29.49
86
123
117
101
112
97
123
115
106
101
123
112
120
126
81
NA
97
105
8
18
16
14
12
15
17
15
14
4
13
16
16
18
14
16
17
15
64.06
139.70
21.90
19.71
42.38
78.21
37.06
68.59
114.68
113.15
131.27
59.02
27.68
24.05
13.26
17.84
25.37
12.52
Note. NART = National Adult Reading Test (Nelson, 1982); TROG2 = Test of Reception of Grammar (Bishop, 2003); lang/LANG = language; md/MD = multiple demand; L = left; R = right.
a Participant md6 was excluded from the degraded speech task analyses.
b Participant md10 was excluded from the semantic ambiguity task analyses.
6
7
1
B
R
UN
io
N
N
e
T
w
o
R
k
S
F
o
R
C
H
UN
l
l
e
N
G
e
S
T
o
S
P
e
e
C
H
C
o
M
P
R
e
H
e
N
S
io
o
N
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
N
o
_
UN
_
0
0
0
8
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Brain networks for challenges to speech comprehension
Lesion Analysis
Lesion analysis followed procedures developed in previous research (Woolgar et al., 2018).
Each participant had a structural MRI image (T1-weighted spoiled gradient echo MRI scans
con 1 × 1 × 1 mm resolution) which included lesion tracing as part of previous participation
in the CCNRP. From these images, we estimated the volume of lesion that overlapped with
the language network, the MD network, or elsewhere. The two networks were defined from
probabilistic fMRI activation maps constructed from large numbers of healthy participants
(Language: n = 220, MD: n = 63), who performed tasks developed to localise language pro-
cessing and domain-general executive processing (see Blank et al., 2014; Fedorenko, 2014;
Fedorenko et al., 2013; Mahowald & Fedorenko, 2016). The activation maps for the language
network contrasted data from participants reading or listening to sentences versus lists of pseu-
dowords (neural responses in the language network are modality-independent; Fedorenko
et al., 2010; Scott et al., 2017); those for the MD network contrasted data from participants
performing a hard versus easy visuospatial working memory task (remembering 8 vs. 4 loca-
zioni, rispettivamente, in un 3 × 4 grid). The visuo-spatial task captures all major components of the
MD network defined by overlap of multiple demands (Assem, Glasser, et al., 2020). Further-
more, defining the network with a non-auditory, non-language task makes relating the impact
of damage to the network on spoken language functions potentially more noteworthy than
using a task that targets auditory or language processing. Each individual participant’s activa-
tion map for the relevant contrast (sentences > pseudowords, hard > easy spatial working
memory) was thresholded at a p < 0.001 uncorrected level, binarised and normalised before
the resulting images were combined in template space. Thus, the language and MD networks
are functionally defined for each individual separately before being combined (for discussion
of the benefits of using an individual subject approach, see Fedorenko, 2021). The resulting
probabilistic activation overlap maps (Figure 1A) contain information in each voxel about
the proportion of participants who show a significant effect (at p < 0.001) for the contrast of
interest. Following Woolgar et al. (2018), we thresholded the probabilistic map for each
network at 5%, thus retaining voxels in which activation was present for at least 5% of the
contributing participants.
We then calculated the lesion volume falling into each network (defined in the probabilistic
map) for each of the 19 participants (Figure 1B). Participants were initially assigned to one of
three broad groups (LANG, MD or OTHER) based on the proportion and volume of their
lesions falling into language and MD regions as well as the overall proportion of each network
that was damaged (Figure 1C; see Supporting Information for further details of group assign-
ment). However, since assignment of participants to groups is based on arbitrary lesion volume
thresholds and because the group allocation for several participants was not clear-cut (e.g.,
lang2, lang3, md5 in Figure 1B) our main analyses correlate behavioural performance mea-
sures with lesion volume in the two key networks, thereby avoiding these arbitrary choices.
We detail the group assignments in describing the participants and results so that the interested
reader can track information about individual participants. Group analyses are included in the
Supporting Information.
Statistical Analysis
Analyses were performed using R statistical software ( Version 3.6.1; R Core Team, 2019). For
each task, the primary analyses assessed whether more extensive damage to the language and
MD networks were associated with more impaired performance on the behavioural tasks, with
one-tailed Pearson’s r correlation coefficients. We compared the strength of different
Neurobiology of Language
672
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
Figure 1. Language and multiple demand (MD) networks, participant lesion maps, and lesion volumes. (A) The language and MD networks
against which we compared participants’ lesions. The images show probabilistic activation maps of the language network and the MD network
based on fMRI data from large numbers of neurotypical participants (language: n = 220; MD: n = 63), which have been thresholded to show
regions active in at least 5% of participants during the relevant functional task and plotted onto a volume rendering of the brain. (B) Volume of
lesion falling into each network for each of the 19 participants in the present study. Solid line depicts an equal volume of each network affected
by the lesion. Different colours/shapes indicate assignment of the participants into the LANGUAGE (LANG), MD, and OTHER Groups upon
which recruitment was based (for categorical group analyses, see Supporting Information). (C) Lesion overlap across participants depicted on
volume renderings of the brain and on midline sagittal slices viewed as if from the left or right (Montreal Neurological Institute (MNI) space x
coordinates of −8 and +8, cross-hairs show the location of y = 0, z = 0 in these slices). Images are shown separately for participants originally
assigned to each of the three groups (see Supporting Information for group analyses). Two participants assigned to the MD group (md6, md10)
contributed data to tasks for only one type of challenge and therefore images are shown separately for the two challenge types. Brighter colours
reflect greater lesion overlap across participants.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
correlations within task (i.e., comparing the impact of damage to language and MD networks
on a given behavioural measure) and between task (i.e., comparing the impact of damage to a
given network on different behavioural measures) with two-tailed Meng’s z tests (Meng et al.,
1992) using the ‘cocor’ package (Diedenhofen & Musch, 2015). The between-task compari-
sons focused on the 17 participants for whom we had data for both the degraded speech and
the lexical-semantic ambiguity tasks.
Neurobiology of Language
673
Brain networks for challenges to speech comprehension
Challenge 1. Acoustically Degraded Speech Perception and Adaptation
The first challenge increased speech comprehension difficulty at the acoustic-phonetic level of
the input, by acoustic degradation of spoken sentences with noise vocoding (Shannon et al.,
1995). Noise vocoding reduces the spectral detail in the speech signal but retains the slow
amplitude modulations, which approximately reflect syllabic units, and the broadband spec-
tral changes that convey speech content. These low frequency modulations and broadband
spectral modulations have been shown to be most important for accurate speech perception
(Elliott & Theunissen, 2009; Shannon et al., 1995). We selected the particular numbers of
channels in the vocoder based on previous research, which established that intelligibility
(as measured by word report: How many words of the sentence a participant can accurately
report) increases with the logarithmic increase in number of channels (McGettigan et al.,
2014). In healthy adults with good hearing, for short sentences of 6–13 words long, intellig-
ibility is near 100% for 16-channel vocoded speech, near 0% for 1-channel vocoded speech,
and at an intermediate level for 4-channel vocoded speech (Peelle et al., 2013). We assessed
speech perception in terms of the logarithmic number of channels estimated as required to
achieve 50% word report accuracy of these sentences and assessed adaptation by comparing
performance before and after a training period.
Stimuli
The stimuli for the degraded speech task were 40 declarative sentences, varying in length
(6–13 words, M = 9, SD = 2.45) and duration (1.14 to 3.79 s, M = 2.12, SD = 0.60), which were
selected from coherent low ambiguity sentences used in previous studies (Davis et al., 2011).
Sentences were recorded by a female native speaker of British English and digitised at a
sampling rate of 22050 Hz. We created three degraded versions of the sentences, of decreas-
ing intelligibility, using 16, 8, and 4 channels in the vocoder. To do this, the frequency range
50–8000 Hz was divided into 16, 8, or 4 logarithmically spaced frequency channels. Each
channel was low-pass filtered at 30 Hz and half-wave rectified to produce an amplitude enve-
lope for each channel, which was then applied to white noise that was filtered in the same
frequency band. Finally, the channels were recombined to create the noise-vocoded version of
the sentence.
The 40 sentences were grouped into eight sets of five sentences such that each set con-
tained 45 words in total and were expected (based on previous word report data) to be approx-
imately equally intelligible. Each participant heard all eight sentence sets, but assignment of
sets to the different levels of degradation (clear, 16-, 8-, 4-channel vocoded) and to the pre-
and post-training test (described below) was counterbalanced across participants.
Procedure
The experiment started with four practice trials to familiarise the participants with the stimuli
and the word report task. Participants listened to four different sentences (not included in the
test set) at increasing levels of degradation (clear, 16, 8, 4) and after each sentence had to
repeat the sentence or as many words from the sentence as possible in the correct order.
The experiment then followed a test–train–test format (cf. Sohoglu & Davis, 2016) with the
40 experimental sentences (eight sets of five sentences; see Stimuli section above for details
of assignment of the sentences to the pre-test and post-test and to the different levels of deg-
radation). In the initial test, participants listened to 20 of the sentences, five at each level of
degradation (clear, 16, 8, 4; order randomised uniquely for each participant) and performed
the word report task. This was followed by a training period in which participants listened
Neurobiology of Language
674
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
passively to the same 20 sentences, each repeated four times at decreasing levels of degrada-
tion, whilst the written text of the sentence was presented visually on a computer screen.
Following the training, participants listened to the other (previously unheard) 20 sentences,
five at each level of degradation, and again performed the word report task.
Data processing and analysis
For each participant, we calculated the number and proportion of words correctly reported for each
sentence at each level of degradation (clear, 16-, 8-, 4-channel vocoded) and for the pre- and post-
training test. Words were scored as correct only if there was a perfect match with the spoken word
from the sentence (morphological variants were scored as incorrect, but homonyms, even if seman-
tically anomalous, were scored as correct). Words reported in the correct order were scored as cor-
rect even if intervening words were absent or incorrectly reported, but scored as incorrect if they
were reported in the wrong order. To verify that decreasing the number of vocoded channels
increased the challenge of speech perception and that training facilitated perception, we analysed
differences in proportion of words correctly reported between the sentences with different numbers
of channels (clear, 16, 8, 4) and pre- and post-test sentences, with a logistic mixed effects model
using the lme4 package (Bates et al., 2015). The model had a single categorical fixed effects predictor
for Training (pre-test or post-test) with deviation coding defining one planned contrast: pre-test = −1/2
versus post-test = 1/2. There was also a continuous fixed effect predictor of Log2Channels (log2
number of channels). The final model contained a by-subject random intercept, by-subject
slopes with Training and Log2Channel, and a by-item random intercept for sentence.
To quantify the relationship between acoustic degradation and speech perception perfor-
mance in single participants we also fit a logistic psychometric function to the word report
accuracy data separately for each participant, for averaged data, and for pre- and post-training
tests separately using the quickpsy package (Linares & López-Moliner, 2006). The parameters
of the logistic function were estimated using direct maximisation of the likelihood with the
following equation:
f x : α; β; γ; λ
ð
Þ
ð
Þ ¼ γ þ
1 − γ − λ
(cid:1) (cid:3)
1 þ e − x
α
β
During the fitting, we treated clear speech as equivalent to 32-channel vocoded speech and
converted the number of channels vocoded at each level of degradation into their log equiv-
alents (χ). From each fit, we obtained alpha (α), the number of channels estimated to give 50%
accuracy on the word report task. This value, referred to as threshold number of channels was
used for the subsequent analyses of the impact of lesion on performance (cf. McGettigan
et al., 2014). Lower alpha values indicate that fewer channels were required to reach this
threshold and thus reflect better performance or more accurate perception. Beta (β) corre-
sponds to the slope or steepness of the curve. Gamma (γ) is the guess rate, which was fixed
to 0 for this open set speech task. Lambda (λ) is the lapse rate, or expected proportion of errors
as the number of channels reaches the highest levels. Lambda represents the upper horizontal
asymptote and was fixed at 1 minus the proportion of correct word report observed for clear
speech for each participant separately. This was required as some participants did not achieve
100% word report for clear speech.
Challenge 2. Semantically Ambiguous Speech Comprehension and Adaptation
The second challenge increased speech comprehension difficulty at the lexical-semantic level,
by the inclusion of semantically ambiguous words, in sentence contexts that in most cases
Neurobiology of Language
675
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
supported the lower frequency meaning. We assessed speech comprehension in terms of the
speed and accuracy of judging the coherence of these sentences, which were interspersed
with sentences without ambiguities and anomalous sentences. The coherence judgement task
appeared well-suited for assessing competence at semantic ambiguity resolution for several
reasons. Firstly, to respond accurately listeners must understand the whole sentence and not
just identify one (or more) unusual words. For example, a sentence might initially make sense
but then become anomalous only at the end (“It was a rainy day and the family were thinking
to the banana”) or might initially seem odd but would eventually make sense (“It was a terrible
hand and the gambler was right to sit it out.”). Secondly, because most of the meanings that
we used in the sentences were the less frequent meanings, accurate performance relies on
listeners utilising contextual cues to select the appropriate meaning rather than the higher-
frequency, more accessible meaning. The use of lower-frequency word meanings also maxi-
mised our chance to observe word meaning priming effects, as described below. Thirdly,
participants make a speeded judgement giving a continuous measure of performance in
addition to accuracy.
To assess the increase in availability of low frequency word meanings in response to expe-
rience, we measured changes to meaning preferences in a word association task. This task
provides a direct measure of how participants interpret ambiguous word forms in the absence
of any sentence context. Specifically, using two counterbalanced sentence sets, we measured
the increase in proportion of word association responses that were consistent with the (low
frequency) meaning used in the sentence context for ambiguous words that had been heard
(primed) compared to those that had not (unprimed). Counterbalanced assignment of sen-
tences to primed and unprimed conditions for different participants ensured that differences
in meaning frequency or dominance did not confound assessment of the word-meaning
priming effect (for further discussion of word-meaning priming, see Rodd et al., 2013).
Stimuli
The stimuli for the coherence judgement task were 120 declarative sentences, selected from
two previous studies (Davis et al., 2011; Rodd et al., 2005). Of these, 40 were high-ambiguity
coherent sentences, 40 low-ambiguity coherent sentences, and 40 anomalous sentences. The
high-ambiguity sentences each contained two ambiguous words that were disambiguated
within the sentence (e.g., “The PITCH of the NOTE was extremely high.” The ambiguous
words were not repeated across the set of 40 sentences.). Prior dominance ratings (Gilbert &
Rodd, 2022) indicated that in most of the sentences, the context biased the interpretation of the
ambiguous words towards their subordinate (less frequent) meanings (mean dominance =
0.31; SD = 0.25). The low-ambiguity sentences were matched with the high-ambiguity sen-
tences across the set for number of words, number of syllables, syntactic structure and natu-
ralness but contained words with minimal ambiguity (e.g., “The pattern on the rug was quite
complex.”). These 80 coherent sentences were separated into two lists (List A and List B), each
containing 20 high-ambiguity and 20 low-ambiguity sentences. Participants were presented
with sentences from either list (List A or List B) and thus were exposed to half of the ambiguous
words in this part of the experiment. Each list also contained all 40 anomalous sentences (i.e.,
the same sentences were presented to all participants) which had been created from the
low-ambiguity sentences by randomly substituting content words matched for syntactic class,
frequency of occurrence, and numbers of syllables (e.g., “There were tweezers and novices in
her listener heat.”). Thus, the anomalous sentences had identical phonological, lexical, and
syntactic properties but lacked coherent meaning (see Table 2 for psycholinguistic properties
of the 3 sentence types).
Neurobiology of Language
676
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
Table 2. Descriptive characteristics of the three sentence types.
Sentence type
High ambiguity
Low ambiguity
Anomalous
N
40
40
40
Mean (range)
number of
words
9.6 (6–18)
9.6 (6–18)
9.0 (6–13)
Note. Cells are empty where data are not applicable.
Mean (range)
number of
syllables
11.8 (7–22)
Mean (range)
duration in
seconds
2.2 (1.1–4.0)
Mean (range)
naturalness rating
on 9-point scale
6.1 (3.6–7.9)
Mean (range)
imageability rating
on 9-point scale
5.4 (2.1–9.0)
12.2 (8–23)
2.2 (1.6–4.3)
6.4 (3.4–7.9)
5.0 (2.0–8.0)
11.6 (6–20)
2.3 (1.3–3.5)
The stimuli for the word association task were the 80 ambiguous target words from the 40
high-ambiguity sentences. Given that participants had only heard half of the high-ambiguity
sentences in the sentence coherence judgement task (List A or List B), for 40 of the ambiguous
words, the subordinate meaning was primed (previously heard in a supportive sentence
context) and for the other 40 the subordinate meaning was not primed.
Sentences and single words were recorded individually by a male native speaker of British
English (M.H.D) and sentences were equated for root mean square amplitude across
conditions.
Procedure
The task consisted of two phases. In the first phase, participants listened to 80 sentences (20
high-ambiguity, 20 low-ambiguity, 40 anomalous) and had to judge as quickly and as accu-
rately as possible the coherence of each sentence. They indicated their response by pressing a
green button if the sentence made sense and a red button if it did not. Participants were given
examples (not included in the test set) to encourage them to listen to the sentence in its entirety
before making the judgement.
Following the coherence judgement task, participants completed other behavioural tasks
(not relevant to the current investigation) for 20–30 min before moving to the second phase:
a word association task. In this phase, participants heard 80 ambiguous words presented in
isolation, of which half had been presented in phase 1 (primed) and half were new (unprimed;
counterbalanced across participants). For each word, participants had to repeat it and then say
the first related word that came to mind. Responses were audio recorded and later coded as
consistent with the subordinate meaning (e.g., “NOTE-music”) or inconsistent with the subor-
dinate meaning (“NOTE-write”).
Data processing and analysis
There were 1,440 experimental trials (18 participants × 80 items). We excluded trials with very
fast responses (more than 300 ms before the offset of the sentence), which were assumed to
arise from accidental key presses or anticipatory responses. This resulted in the exclusion of
two anomalous sentence trials and one low-ambiguity sentence trial.
For each participant, we first assessed whether they could discriminate the coherent sen-
tences (high-ambiguity and low-ambiguity) from the incoherent sentences better than would
be expected by chance, by calculating d-prime values for the high-ambiguity and low-
ambiguity sentences separately:
d prime ¼ z Hits
ð
Þ − z False Alarms
ð
Þ
Neurobiology of Language
677
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
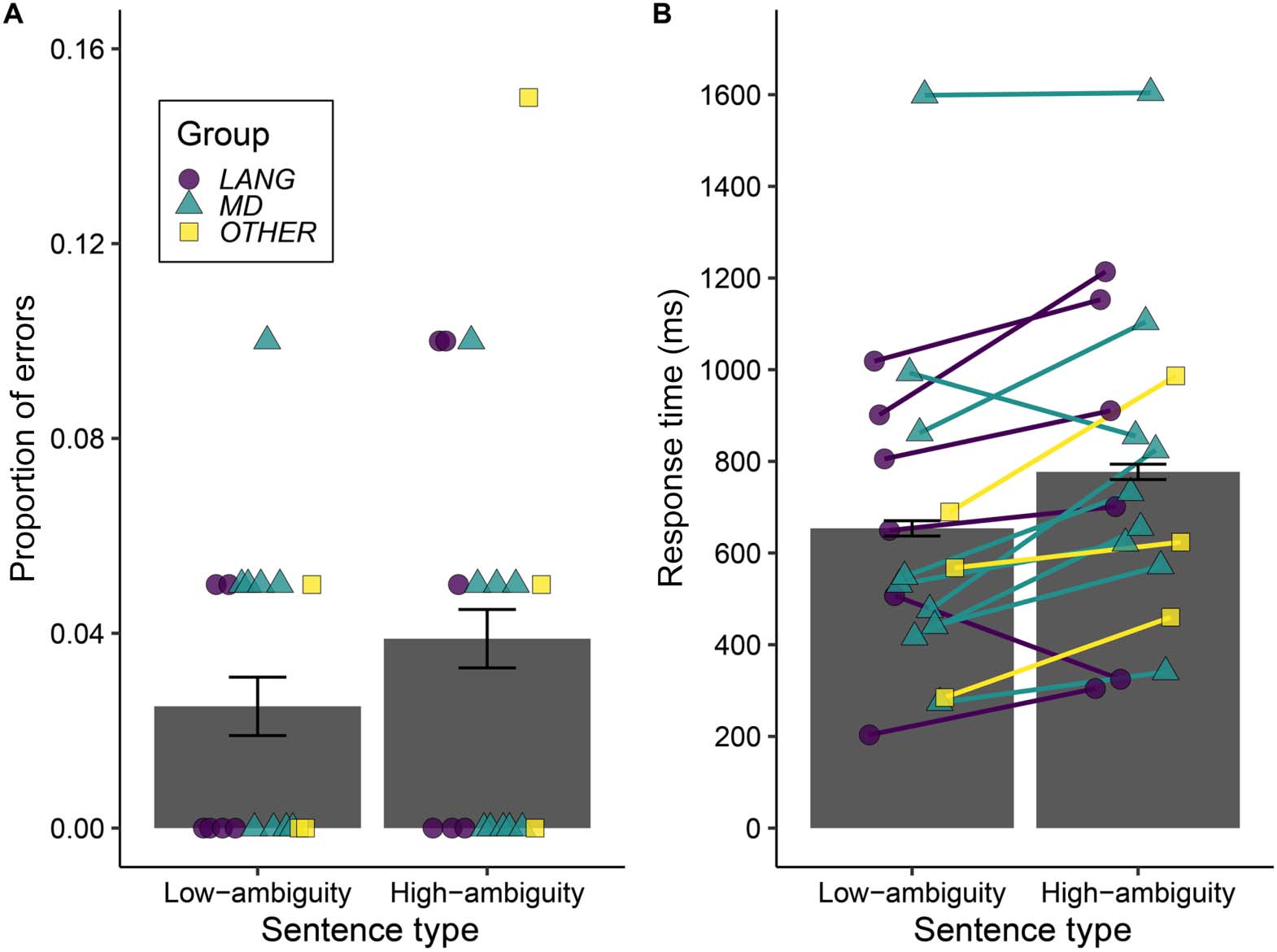
Hits correspond to the proportion of coherent sentences correctly judged as coherent. False
alarms correspond to the proportion of incoherent sentences incorrectly judged as coherent.
To allow for calculation of the z-scores, hit rates of 1 were adjusted by 1 − 1/2N (i.e., to a value
of 0.975) and false alarm rates of 0 were adjusted by 1/2N (Macmillan & Kaplan, 1985; i.e., to
a value of 0.0125).
As the false alarm rate was necessarily identical for high-ambiguity and low-ambiguity con-
ditions (we only included a single set of incoherent sentences), differences in accuracy
between the high-ambiguity and low-ambiguity sentences can be assessed using error rates
when participants judged these coherent sentences to be anomalous. Therefore, for the main
accuracy analyses we excluded the 40 anomalous sentence trials, leaving 719 trials (1 trial
was excluded based on a fast response time; see above).
The response time analyses focused on ambiguous and unambiguous sentence trials. Of the
720 total experimental trials (18 participants × 40 items), we excluded trials incorrectly judged
as incoherent (23 trials: 14 ambiguous, 9 unambiguous). For exclusions of trials based on
response times, we followed the general principle of minimal trimming with model criticism
(Baayen & Milin, 2010). We excluded trials with very fast response times (less than 300 ms
before offset; as for the accuracy analysis), which were assumed to reflect accidental key
presses (1 trial) as well as trials with very slow response times (three trials with responses longer
than 4,000 ms after sentence offset) because we were interested in speeded responses. Further
exclusions were considered after first determining whether any transformation of the depen-
dent variable was required to meet assumptions of the linear mixed-effects models, of homo-
geneity of residual variance and normally distributed residuals. Model diagnostic plots
(quantile–quantile and histogram plots of the residuals) for the raw, log10-transformed and
inverse transformed response time data showed that log10 transformation best met the
assumptions. Examination of the plots for outliers indicated that no further trimming was
necessary, thus there were 693 correctly judged coherent trials included in the analyses.
We analysed differences in accuracy and response times between the high-ambiguity and
low-ambiguity sentence trials with a logistic mixed effects model (accuracy) or a linear mixed
effects model (log-10 response times) using the lme4 package (Bates et al., 2015). The models
had a single categorical fixed effects predictor for Sentence Type (High-ambiguity or Low-
ambiguity) with deviation coding defining one planned contrast: High-ambiguity = 1/2 versus
Low-ambiguity = −1/2. The final models each contained a by-subject and by-item random
intercept.
The correlational analyses used the model residuals (comparing predictions to the data) to
estimate the ambiguity response time effect (difference between responses for high-ambiguity
and low-ambiguity sentence trials) for each participant. A positive residual difference indicates
that the participant’s ambiguity effect was larger than predicted by the model (response
times were slower than estimated for the high-ambiguity condition and/or faster than estimated
for the low-ambiguity condition). A negative residual difference means that their response time
effect was smaller than predicted by the model (response times were faster than estimated for
the high-ambiguity condition and/or slower than estimated for the low-ambiguity condition).
For the word association task, each response was independently coded for consistency with
the subordinate meaning used in the priming sentence by two of the authors (LM and ZB), who
were blind to the experimental condition (primed/unprimed) of the responses. For example, the
word “ball” came from the sentence “The ball was organised by the pupils to celebrate the end of
term,” so responses such as “party” and “dance” were coded as consistent whereas responses
such as “kick” and “round” were coded as inconsistent. The consistency scores for the unprimed
Neurobiology of Language
678
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
words give a baseline measure of the preference for the dominant meaning. Response codes
from the first author were used with the exception of one participant for whom data were lost
and only the codings from the second rater were available; inter-rater reliability for the remain-
der of the responses was high (94% agreement from 1,360 responses, Cohen’s Kappa = 0.862).
We analysed differences in the proportions of responses consistent with the subordinate
meaning between primed and unprimed words (word meaning priming) with a logistic mixed
effects model with a categorical fixed effect predictor for Priming Type (Primed or Unprimed)
with deviation coding defining one planned contrast: Primed = 1/2 versus Unprimed = −1/2.
There was also a continuous fixed effect predictor of Meaning Dominance (Gilbert & Rodd,
2022) and the associated interactions. The final model contained a by-subject and by-item
random intercept and a by-subject random slope for Dominance.
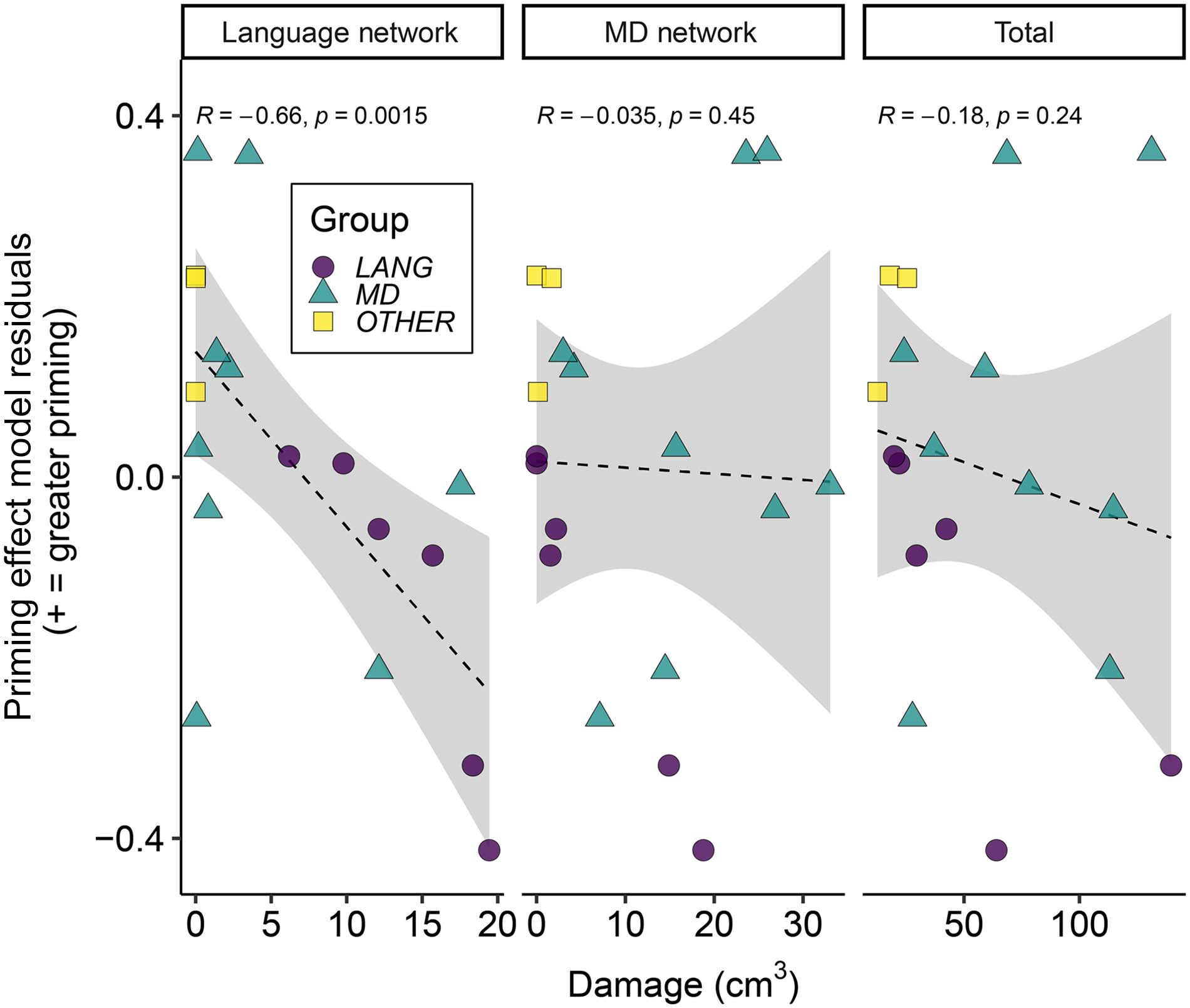
In the main correlational analyses we used the model residuals (comparing predictions to
the data) to estimate word priming effects (difference between response values for primed
and unprimed words) for each participant. A positive residual difference indicates that the
participant’s priming effect was larger than predicted by the model (proportion of responses
consistent with the subordinate meaning was underestimated for the primed condition and/or
overestimated for the unprimed condition). A negative residual difference means that their
priming effect was smaller than predicted by the model (proportion of responses consistent
with the subordinate meaning was overestimated for the primed condition and/or underesti-
mated for the unprimed condition).
RESULTS
Challenge 1. Acoustically Degraded Speech Perception and Adaptation
Word report task
Figure 2A shows the mean proportion of words correctly reported for speech with different
numbers of channels, for the pre- and post-training tests. Word report accuracy was near ceil-
ing (100%) for the clear speech reflecting the participants’ ability to perform the task, but was
close to floor levels for the 4-channel vocoded condition, reflecting the challenge of the acous-
tic degradation. The mixed effect model confirmed that speech perception accuracy increased
as the log2 number of channels increased (model coefficient: β = 3.199, SE = 0.236, z =
13.556, p < 0.0001). Accuracy was greater following training (model coefficient: β = 1.262,
SE = 0.392, z = 3.217, p = 0.001), showing that participants were able to learn. There was no
interaction between the level of degradation and training.
The outputs of fitting the data with a logistic psychometric function are shown in Figure 2B.
Analyses to assess the impact of lesions on performance used the threshold number of channels
(the estimated number of channels required for 50% word report accuracy) with lower values
reflecting better perception (fewer channels needed to reach 50% accuracy). Figure 2C shows
the mean performance before and after training for the group and for individual participants.
Figure 3 shows the relationship between degraded speech perception performance and the
extent and location of lesions. Correlational analyses showed that the mean threshold number
of channels across pre- and post-training tests positively correlated with damage to the MD
network (r = 0.427, p = 0.039) but not with damage to the language network (r = −0.152,
p = 0.727), or with total damage (r = 0.216, p = 0.194). Comparisons of these correlations
demonstrated that poorer speech perception was numerically more strongly predicted by dam-
age to the MD network than to the language network, although this did not reach the p < 0.05
threshold of statistical significance (z = −1.954, p = 0.051). There was no evidence for MD
Neurobiology of Language
679
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Word report task results. (A) Word report accuracy scores for different levels of degradation and the pre- and post-training tests
separately. Bars show mean values across all 18 participants and error bars show ±1 SEM, adjusted to remove between-subject variance
(Morey, 2008). (B) Psychometric logistic function fits separately for the pre-training (solid) and post-training (dashed) data for the mean across
all 18 participants (black colour) and each participant separately (coloured by group). The horizontal line indicates the 50% word report
accuracy threshold. Vertical lines indicate the estimated threshold number of channels corresponding to the 50% word report accuracy thresh-
old for the mean fits across all 18 participants. (C) Estimated threshold number of channels (log scale) required for 50% accuracy in the word
report task for the pre- and post-training tests separately. Bars show mean values across all 18 participants and error bars show ±1 SEM,
adjusted to remove between-subject variance (Morey, 2008). Individual participant values are overlaid (colour and shape reflect participant
group; see Supporting Information).
Neurobiology of Language
680
Brain networks for challenges to speech comprehension
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Relationship between speech perception and lesion volume. Individual participant data
for estimated threshold number of channels (log scale) required for 50% word report accuracy for
the mean of pre- and post-training tests plotted separately against damage to the language network,
to the multiple demand (MD) network, and total damage (colour and shape reflect participant
group; see Supporting Information). Higher threshold number of channels indicates worse speech
perception performance. The dashed line shows the linear best fit, and grey shaded areas show 95%
confidence intervals.
network damage being more predictive of speech perception than total damage (z = −1.122,
p = 0.262). There were no correlations between perceptual learning of degraded speech (i.e.,
change in threshold from pre- to post-training) and the volume of brain damage in the lan-
guage network (r = −0.123, p = 0.727), MD network (r = 0.003, p = 0.504), or total damage
(r = −0.006, p = 0.491. There was also no evidence that MD damage was more predictive of
degraded speech perception than of degraded speech adaptation (z = −1.403, p = 0.161).
Challenge 2. Semantically Ambiguous Speech Comprehension and Adaptation
Sentence coherence judgement task
All participants showed d-prime values substantially above 0 indicating successful discrimina-
tion of the incoherent sentences from both the high-ambiguity (mean = 3.66, SD = 0.25,
range = 3.0–4.20) and the low-ambiguity sentences (mean = 3.75, SD = 0.37, range =
2.72–4.20). Figure 4 shows the mean error rates (Figure 4A) and the mean response times
(Figure 4B) for the different Sentence Types and for individual participants. Across all partici-
pants the proportions of correct responses were near ceiling (mean = 0.97, SD = 0.03, range =
0.92–1.0). The mixed effect model showed no effect of Sentence Type on accuracy (model
coefficient: β = −0.440, SE = 0.538, z = −0.817, p = 0.414), and hence we have no evidence
that sentences containing ambiguous words were less well understood.
Neurobiology of Language
681
Brain networks for challenges to speech comprehension
Sentence coherence task results. (A) Proportion of errors and (B) response times mea-
Figure 4.
sured from sentence offset for coherence judgements to low-ambiguity and high-ambiguity sen-
tences. In each case, bars reflect the mean values across all 18 participants and error bars show
±1 SEM, adjusted to remove between-subject variance (Morey, 2008). Individual participant values
are plotted (colour and shape reflect participant group; see Supporting Information).
The mixed effect model for response times confirmed that high-ambiguity sentences were
responded to more slowly than low-ambiguity sentences (β = 0.043, SE = 0.019, t(2.319) =
2.319, p = 0.023), showing that ambiguous words increased the challenge of sentence com-
prehension. However, there was no correlation between individual ambiguity response time
effects (model residual difference measure) and extent of damage to the language network (r =
−0.102, p = 0.656), the MD network (r = −0.038, p = 0.559) or overall damage (r = 0.076, p =
0.383).
Word association task
We excluded primed trials corresponding to words from sentences that were responded to
incorrectly in the coherence judgement task. This resulted in exclusion of 28 trials (words from
14 ambiguous sentences: 1.94% of data) across the 18 participants, leaving 1,412 observa-
tions. For unprimed words (i.e., for ambiguous words that were not presented to participants
in the coherence judgement task), the mean proportion of responses (across items and partic-
ipants) that were consistent with the subordinate meaning of the word was 0.29 (SD = 0.09).
This value, which gives a baseline measure of the preference for the dominant meaning,
indicates that the sentence-primed meanings were indeed the subordinate or less preferred
meanings (note that the value is similar to the one derived from an existing database (Gilbert
& Rodd, 2022; see Stimuli section above). Figure 5 shows the mean proportion of responses
consistent with the subordinate meaning for primed and unprimed words.
Neurobiology of Language
682
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Word association task results, showing proportion of responses consistent with the sub-
ordinate meaning for ambiguous words in the Unprimed and Primed conditions. Bars reflect the
mean values across all 18 participants and error bars show ±SEM, adjusted to remove between-
subject variance. Individual participant values are plotted (colour and shape reflect participant
group; see Supporting Information).
We observed a main effect of Priming (β = 0.352, SE = 0.137, z = 2.565, p = 0.010), which
reflects a higher proportion of responses consistent with the subordinate meaning for the
primed compared to unprimed words. This finding demonstrates a change in word meaning
preferences in response to recent experience of sentences containing ambiguous words. We
also observed a main effect of meaning Dominance of the word (β = 1.112, SE = 0.111, z =
10.054, p < 0.0001), reflecting an increase in proportion of responses consistent with the sub-
ordinate meaning as the dominance of that meaning increased (became less subordinate and
closer in frequency to the alternative dominant meaning). There was an interaction between
Dominance and Priming (β = −0.300, SE = 0.171, z = −2.124, p = 0.034), reflecting a stronger
Priming effect for meanings that were more subordinate.
Correlational analyses revealed a negative relationship between individual word meaning
priming effects (model residual difference measure) and the extent of damage to the language
network (r = −0.659, p = 0.001) but not the MD network (r = −0.035, p = 0.446) or total dam-
age (r = −0.180, p = 0.237). Comparisons between these correlations showed that word mean-
ing priming was more strongly predicted by damage to the language network than to the MD
network (z = −2.182, p = 0.0291); a comparison with the correlation with total damage did not
reach the conventional p < 0.05 threshold (z = −1.863, p = 0.062). There was also evidence
that damage to the language network was more predictive of individual participants’ word
meaning priming than the ambiguity response time effect (z = −2.6523, p = 0.008).
Figure 6 shows scatter plots of the correlations between word-meaning priming and damage
Neurobiology of Language
683
Brain networks for challenges to speech comprehension
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Relationship between word meaning priming and lesion volume. Individual participant
data for word meaning priming effects estimated from the model residuals, plotted separately
against damage to the language network and to the multiple demand (MD) network, and against
total damage (colour and shape reflect participant group; see Supporting Information). The dashed
line shows the linear best fit, and grey shaded areas show 95% confidence intervals.
to language network, MD network, and total damage. A further correlational analysis showed
no relationship between participants’ ambiguity response time effect and their word meaning
priming effect (both measured using the model residuals: r = −0.26, p = 0.298, two-tailed),
suggesting that reduced word meaning priming effect could not be explained simply as due
to poorer comprehension.
Dissociations Between Challenges to Speech Perception and Comprehension
Table 3 summarises the correlations between the lesion volume in each network and measures
of acoustically degraded speech perception, semantically ambiguous speech comprehension,
and associated adaptation to each of the challenges. Table 3A displays lesion–behaviour
correlations from the 18 participants tested for each challenge. Where significant lesion–
behaviour correlations are observed, additional comparisons between correlations with
behaviour for lesions to the two networks and within each challenge type are also shown.
Table 3B displays lesion–behaviour correlations for the 17 participants for whom we have data
for both types of challenge. This table shows comparisons between correlations for the two
types of challenge.
Neurobiology of Language
684
Brain networks for challenges to speech comprehension
Summary of correlations between the extent of damage to the multiple demand (MD) and language networks and performance on
Table 3.
the tasks measuring speech perception/comprehension and adaptation for the two types of challenge (acoustic degradation and semantic
ambiguity).
Lesion
Challenge
Acoustic degradation
Task
MD network
(1) Perception
r = 0.427, p = 0.039
Language network
r = −0.152, p = 0.727
MD vs. language network
z = −1.954, p = 0.051
n = 18a
(2) Adaptation
r = 0.003, p = 0.504
r = −0.123, p = 0.313
NA
(1) vs. (2)
z = −1.403, p = 0.161
NA
–
Semantic ambiguity
(3) Comprehension
r = −0.038, p = 0.559
r = −0.102, p = 0.656
NA
n = 18b
(4) Adaptation
r = −0.035, p = 0.446
r = −0.659, p = 0.001
z = −2.182, p = 0.0291
Acoustic degradation vs.
semantic ambiguity
n = 17a,b
(3) vs. (4)
(1) vs. (3)
(2) vs. (4)
(1) vs. (4)
(3) vs. (2)
NA
z = −2.652, p = 0.008
z = 1.644, p = 0.100
NA
NA
z = −1.678, p = 0.093
z = −1.042, r = 0.300
z = −2.160, r = 0.031
NA
NA
–
–
–
–
–
Note. Tasks measuring speech perception/comprehension and adaptation are labeled (1)–(4). Where significant lesion–behaviour correlations are observed,
additional comparisons of the strength of these correlations are also shown. Correlations are shown for data from the 18 participants for each type of challenge
and associated within-challenge (across-lesion, across-task) comparisons. Two participants (md6 and md10) performed tasks for only one challenge type.
Across-challenge comparisons are shown for data from the 17 participants for whom we have data for both types of challenge. Significant lesion–behaviour
correlations (in bold) are shown between the MD network and Task (1): acoustically degraded speech perception (turquoise), and between the language net-
work and Task (4): adaptation to semantic ambiguity (word meaning priming; purple). In the case of Task (1) the finding that an increase in lesion volume is
associated with worse behavioural outcomes is reflected in a positive correlation since higher perceptual thresholds indicate worse perception. In Task (4) the
finding that an increase in lesion volume is associated with worse behavioural outcomes is reflected in a negative correlation since less word meaning priming
indicates less adaptation. A dash indicates that analyses were not performed. NA = data are not applicable.
a Excluding md6.
b Excluding md10.
As detailed in the task-specific results (summarised in Table 3), acoustically degraded
speech perception was predicted by the degree of MD network damage and this correlation
was in the opposite direction to, but not significantly stronger than, the correlation with
language network damage volume (see Word report task section in Results). Conversely
semantically ambiguous speech adaptation (word meaning priming) was predicted by lan-
guage network damage and this correlation was significantly stronger than the nonsignificant
correlation with MD network damage volume (see Word association task section in Results).
This double dissociation provides evidence for causal associations between the integrity of
the MD network and abilities at degraded speech perception and between the language net-
work and word meaning priming. Further comparisons of the strength of correlations between
tasks within the same type of challenge (acoustic degradation or semantic ambiguity) showed
that damage to the language network was more predictive of impaired word meaning priming
than of comprehension of ambiguous sentences. This finding provides support for a specific
contribution of the language network to adaptation that is independent to its role in compre-
hension (at least as measured here), which was shown to be largely independent of language
or MD network lesions. However, there was no evidence that damage to the MD network
was more strongly predictive of degraded speech perception than of degraded speech
adaptation.
Neurobiology of Language
685
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
To further explore the specificity of the contribution of the MD and language networks to
acoustically degraded speech perception and word meaning priming respectively, we also
compared the strength of correlations between tasks using data from the 17 participants
who performed both the acoustic degradation and the semantic ambiguity tasks (Table 3).
There was no evidence that damage to the MD Network was more predictive of degraded
speech perception than of word meaning priming (z = −1.042, p = 0.300), or of comprehen-
sion of ambiguous sentences (z = 1.644, p = 0.100). Damage to the language network was
more predictive of impaired word meaning priming than of degraded speech perception (z =
−2.160, p = 0.031), although the comparison with degraded speech adaptation did not reach
the p < 0.05 threshold (z = −1.678, p = 0.093).
DISCUSSION
We report two main findings. First, we show that damage to the domain-general MD network,
but not the language-selective network, causes significant impairments to the perception of
acoustically degraded speech. Word report accuracy for noise-vocoded sentences decreased
as the number of channels in the vocoder decreased, reflecting an increased challenge to
speech perception. The degree of perceptual impairment (i.e., the number of channels
required for 50% correct word report) depends on the extent of damage to the MD network,
but not damage to the language network (Figure 3; Table 3). Word recognition improved
following a period of training, reflecting adaptation or perceptual learning for this form of
acoustic degradation, but the degree of learning was not reliably predicted by lesion location
or extent.
In contrast to these results with acoustically challenging speech, we found no evidence that
semantically challenging speech comprehension was dependent on the MD system: All par-
ticipants were highly accurate in judging the coherence of sentences and were no less accu-
rate when the sentences contained ambiguous words, indicating an intact ability to access the
typically less frequent (subordinate) word meanings used in our high-ambiguity sentences.
Although participants were slower to make judgements for sentences which include ambigu-
ous words, reflecting more effortful comprehension when words have multiple meanings,
there was no significant association between response time slowing for ambiguous sentences
and the extent of damage to the MD or language networks.
Our second main finding is that despite accurate comprehension of semantically ambiguous
speech, damage to the language network—but not to the MD network—caused a significant
reduction in updating of word meaning preferences following recent linguistic experience. As
shown in previous studies of individuals without brain lesions, our participants were (as a
group) more likely to generate a word associate related to the less frequent meaning of an
ambiguous word when they had encountered this meaning in an earlier sentence (word mean-
ing priming, as reported by Rodd et al., 2013). However, the magnitude of this word meaning
priming effect was predicted by the extent of damage to the language network, but not to the
MD network (Figure 6; Table 3), a dissociation that was supported by a statistically significant
difference between the strength of these two correlations. The reduction in word meaning prim-
ing was not explained by sentence comprehension difficulties as there was no correlation
between the magnitude of word meaning priming and increased response times when judging
the coherence of sentences containing semantically ambiguous words. Furthermore, across-
task comparisons showed that the damage to the language network was more predictive of
impaired word meaning priming than impaired comprehension of ambiguous sentences or
impaired perception of acoustically degraded speech.
Neurobiology of Language
686
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
Below, we discuss our two main findings in greater detail. First, we discuss possible cog-
nitive operations performed by the MD network that are required for the perception of acous-
tically degraded speech. We then turn to the linguistic challenge of resolving lexical-semantic
ambiguity. We discuss the functional contribution of the language-selective network in adap-
tation such that low frequency meanings of semantically ambiguous words become more
accessible following recent exposure. In a final section we consider the dissociation between
these different challenges to speech processing and explore implications for the neural basis of
speech perception, comprehension, and adaptation.
The MD Network Makes a Causal Contribution to Perception of Acoustically Degraded Speech
Recently it has been argued that the MD network does not play a functional role in language
comprehension (Blank & Fedorenko, 2017; Diachek et al., 2020; Shain et al., 2020; Wehbe
et al., 2021; for reviews, see Campbell & Tyler, 2018; Fedorenko, 2014). According to such an
account, activations observed during language comprehension within the MD network could
reflect a generic increase in effortful processing or contributions to specific task demands (such
as decision-making), rather than computations essential for language comprehension (e.g.,
identifying words; accessing word meanings). However, this line of research left open the pos-
sibility that MD contributions may be necessary when speech is acoustically degraded and
challenging to perceive (Diachek et al., 2020). Here we provide novel evidence that the
MD network indeed makes a causal contribution to perception of acoustically degraded
speech by assessing the impact of damage to MD regions on performance in a word report
task that indexes cognitive operations required for word identification.
Previous fMRI studies have shown that listening to acoustically challenging speech is asso-
ciated with an increase in activation in prefrontal and motor regions that plausibly fall within
the MD network (Adank, 2012; Davis & Johnsrude, 2003; Du et al., 2016; Erb et al., 2013;
Hardy et al., 2018; Hervais-Adelman et al., 2012; Rysop et al., 2021; Vaden et al., 2013;
Vaden et al., 2015; Wild et al., 2012). However, these studies did not explicitly define MD
regions or test the necessity of MD contributions, and hence this association has not been
firmly established. A substantial advance, then, comes from our finding that neural integrity
of the MD network supports more successful word report for degraded speech, which allows
us to conclude a causal role of MD regions in degraded speech perception.
The MD network has previously been linked to a diverse range of domain-general cognitive
constructs, including executive control, working memory, and fluid intelligence. These
constructs may reflect a combination of different cognitive operations, including setting and
monitoring of task goals; directing attention; and the storage, maintenance, integration, and
inhibition of information across different time scales. It is therefore of interest to consider
which of these operations, performed by the MD network, might be critical for the perception
of acoustically degraded speech. For example, focused attention may be particularly important
when the identities of specific phonemes or words are uncertain. Monitoring may be important
for tracking the accuracy of phoneme perception and word recognition over time.
Future work can tease apart these possible distinct cognitive operations, either by focusing
on potential contribution of distinct subnetworks within the broader MD network, or by
exploring correlations between these other functions of MD networks and perception of
degraded speech. Given strong evidence of inter-regional correlations during naturalistic lis-
tening paradigms (Assem, Blank, et al., 2020; Blank et al., 2014; Mineroff et al., 2018; Paunov
et al., 2019), we here treated the MD network as a functionally integrated system. However,
other research concerned with domain-general cognitive processes has proposed that the MD
Neurobiology of Language
687
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
network consists of at least two interconnected, but distinct subnetworks (one comprising lat-
eral frontal and parietal areas and the other cingular and opercular areas), which may contrib-
ute differently to cognition (Dosenbach et al., 2007; Dosenbach et al., 2008; Nomura et al.,
2010). In the context of effortful speech comprehension, Peelle (2018) proposes a three-way
distinction between frontoparietal, premotor, and cingular-opercular contributions to atten-
tion, working memory, and performance monitoring processes respectively. Consistent with
the proposed role of cingular-opercular regions are data showing that activation is associated
with better word recognition on subsequent trials (Vaden et al., 2013; Vaden et al., 2015),
which may reflect mechanisms for tracking the accuracy of phoneme perception and word
recognition over time. Although the present data cannot adjudicate between bi- or tripartite
views, further research using similar methods and data from a larger number of individuals
could potentially dissociate the effect of lesions of these three subnetworks and establish
underlying mechanisms. For example, we might predict that focal damage to cingular-
opercular regions would result in a greater impairment in degraded speech perception when
perceptual difficulty varies from trial-to-trial compared to cases in which trial difficulty is
grouped into blocks.
Replicating a range of previous behavioural findings (Davis et al., 2005; Hervais-Adelman
et al., 2008; Huyck & Johnsrude, 2012; Loebach & Pisoni, 2008; Peelle & Wingfield, 2005;
Sohoglu & Davis, 2016), we showed that listeners adapt to acoustically degraded speech over
time. This finding extends earlier observations of perceptual learning to individuals with
lesions to language-selective and domain-general regions. We found no evidence that damage
to either MD or language-selective networks led to reduced perceptual learning and hence
cannot make causal claims about the contribution of either network to this form of learning.
Future studies using similar methods would benefit from a larger number of participants, with
more variable and more extensive lesions.
The Language-Selective Network Makes a Causal Contribution to Adaptation to Semantically
Ambiguous Speech
Semantically ambiguous words introduce a substantial challenge to speech comprehension
because of the need to engage competition processes to select between alternative meanings
and the cognitive cost of reinterpretation when initial selection fails (Rodd et al., 2002; Rodd
et al., 2010, 2012). The presence of two or more ambiguous words in each of the
high-ambiguity sentences used in our study made comprehension especially challenging.
Nonetheless, we observed that comprehension—indicated by judging high-ambiguity sen-
tences to be coherent—was ultimately successful (although slower than for low-ambiguity
sentences) and that accuracy in judging coherence did not differ between high- and low-
ambiguity sentences. Neither response time differences nor the accuracy of coherence judge-
ments were associated with the degree of damage to MD or language-selective brain networks.
Thus, our study does not provide evidence for a specific causal role of either of these brain
networks for comprehension of sentences containing ambiguous words.
Despite intact comprehension of sentences containing semantically ambiguous words, we
observed differential effects of lesion location and extent on learning mechanisms involved in
adapting lexical-semantic processing after successful disambiguation. Previous research has
established that recent exposure to low-frequency (subordinate) meanings of ambiguous words
in a sentence context facilitates subsequent meaning access and selection of those meanings, a
process termed word-meaning priming (Betts et al., 2018; Gaskell et al., 2019; Gilbert et al.,
2018; Rodd et al., 2013; Rodd et al., 2016). Previous functional imaging studies have not
Neurobiology of Language
688
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
studied neural activity associated with word-meaning priming and hence the present results
make a novel contribution to understanding the neural basis of this adaptation process. We
here replicated the standard word meaning priming effect for the group of participants tested
overall, but showed that the magnitude of the priming effect was significantly reduced by dam-
age to the language-selective network but not to the MD network.
There is substantial anatomical overlap between the language network shown here to be
critical for updating of word-meaning preferences following successful disambiguation, and
the fronto-temporal brain regions previously shown to respond to semantic ambiguity resolu-
tion (Bilenko et al., 2009; Musz & Thompson-Schill, 2017; Rodd et al., 2005; Vitello et al.,
2014; Zempleni et al., 2007; for a review, see Rodd, 2020), consistent with shared neural
resources between semantic comprehension and subsequent adaptation. The absence of an
effect of language-network damage on immediate comprehension, coupled with the observed
impact of language-network damage on semantic adaptation may therefore reflect the rela-
tively high functioning of the volunteers, the limited severity of the language lesions, and/or
the relative insensitivity of the comprehension task to distinguishing between these relatively
unimpaired volunteers. It also remains possible that particular subregions of the language
network are differentially important for immediate comprehension compared to subsequent
adaptation. These issues could be explored in future work, which would clearly benefit from
larger numbers of participants. A larger sample would also allow the use of alternative
methods such as voxel-based lesion-symptom mapping to localise function more specifically
within the network, for example, by contrasting frontal and temporal lobe lesions.
One striking illustration of the longevity of learning is that word-meaning priming has pre-
viously been observed 24 hr after a single exposure to an ambiguous word; especially if there
is an intervening period of sleep (Gaskell et al., 2019). This latter finding, in combination with
a wider literature on the role of consolidation processes that facilitate the acquisition of new
lexical knowledge (Dumay & Gaskell, 2007; Gaskell & Dumay, 2003; Tamminen et al., 2010),
led Gaskell et al. (2019) to suggest that word meaning priming may involve a two-stage com-
plementary systems account of learning (McClelland, 2013), as proposed for the acquisition of
novel words (Davis & Gaskell, 2009). According to this account, short-term learning arises
from hippocampally mediated binding of associations between words in the sentences, while
these short-term changes are consolidated into long-term changes to word meaning prefer-
ences after sleep.
The present study constrains these complementary systems accounts of learning by reveal-
ing a causal contribution of language-selective cortical regions even for short-term adaptation
of familiar word meanings. Future work could further consider the interaction of hippocampal
and cortical regions in the learning and maintenance of meaning preferences over different
timescales and the relationship between learning novel vocabulary and updating of existing
lexical semantic knowledge (for recent meta-analyses of word form learning and consolida-
tion, see Schimke et al., 2021; Tagarelli et al., 2019). We note that previous research has
shown that individuals with aphasia (identified behaviourally) can learn novel vocabulary
but that learning is highly variable (Kelly & Armstrong, 2009; Tuomiranta et al., 2011,
2014). The present work similarly shows variability in the impact of cortical lesions on adapt-
ing the meanings of familiar words. However, our participants were not recruited on the basis
of language impairment and retained good comprehension both on a standardised measure of
sentence comprehension (TROG2) and on the experimental measure of ambiguity resolution
tested here. It might be that individuals with more extensive lesions to language selective
cortex, or more focal lesions of posterior temporal and inferior frontal regions that contribute
to ambiguity resolution would show a greater impairment to comprehension. Such a finding
Neurobiology of Language
689
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
would suggest that a common set of cortical regions support comprehension and learning of
ambiguous words in sentences. Further refinement of lesion definitions and tests of larger sam-
ples of individuals could also provide more detailed anatomical evidence concerning the rel-
ative contributions of language-selective and/or domain-general subregions of the IFG to
semantic ambiguity resolution. These regions lie in close proximity and thus may appear to
overlap in group studies (Fedorenko & Blank, 2020), which may explain why they have not
been dissociated in previous imaging research on semantic ambiguity resolution.
Neural Dissociation of Different Challenges to Speech Perception and Comprehension
Taken together, our findings provide a double dissociation indicating independent functional
contributions of the MD and language-selective networks to responding to and adapting to
different types of difficult-to-understand sentences. Specifically, we show that the challenge
of perceiving acoustically degraded sentences (measured in terms of word report accuracy)
is causally linked to the degree of damage to the MD network but not to the language-selective
network (although the comparison of correlations was not statistically significant; see Table 3).
Conversely, the challenge of post-comprehension adaptation to semantically ambiguous
words in sentences (measured in terms of word meaning priming) causally depends on the
integrity of the language-selective but not the MD network; moreover, in this case there is a
reliable difference between the significant (language) and null (MD) correlations.
Here we tested a limited set of challenges to speech comprehension, thus we cannot make
general statements concerning dissociable contributions made by each of these cortical net-
works to all forms of perceptual or semantic challenge. However, our data provide initial evi-
dence for the task specificity of causal contributions. Focusing first on the effect of language
network damage, the correlation between language lesion volume and word meaning priming
was significantly different from the null correlation between lesion volume and coherence
judgement response times for ambiguous sentences, indicating a greater sensitivity to the
integrity of the language network for adaptation compared to initial comprehension of lexico-
semantic ambiguity. It is important to note that our results are based on data from individuals
without aphasia in whom lesions extended to a maximum of only ~11% of the language cortex
(see Supporting Information). It is likely that more extensive lesions, or indeed more sensitive
tests, would detect a contribution of the language network to comprehension. Furthermore,
the reliable correlation between lesion volume and word-meaning priming could also be dis-
sociated from the (null) effect of lesions on degraded speech perception, suggesting that the
integrity of the language network is more important for lexicosemantic than for acoustic or
perceptual challenges. We note, however, that our definition of the language network was
derived from studies using both written and spoken language and hence likely excluded early
auditory processing stages. It would be of interest to explore lesion definitions based on loca-
lising the speech perception system, which might reveal other systems that causally support
abilities at degraded speech perception.
Equivalent across-task comparisons of the effect of MD network damage did not reach sta-
tistical significance. Despite a reliable correlation between MD lesion volume and impaired
perception of acoustically degraded speech, this effect could not be clearly dissociated from
the null effects of lesion volume on tasks involving semantic processing, or perceptual and
semantic adaptation. We therefore cannot draw strong conclusions about the specificity of
the contribution of the MD network to degraded speech perception based on the current data.
For instance, it remains possible that the MD contribution is a secondary consequence of an
increase in demands on domain-general cognition (e.g., working memory) that would affect all
Neurobiology of Language
690
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
aspects of language functioning, but are emphasised by the word report task for degraded
speech. While we had expected that additional demands on domain-general operations would
be equivalent across each of our listening challenges and tasks, we do not have evidence that
this was the case. It might be that more careful titration of task difficulty will be required if we
are to demonstrate that network-specific lesions impair specific aspect(s) of language function
(perception vs. comprehension vs. adaptation to challenge types) rather than apparently spe-
cific effects being mediated by domain-general processes.
Relatedly, a reviewer raised the possibility that unmeasured auditory and cognitive impair-
ments may have had consequences for participants’ task performances. Such impairments
could affect performance even when participants achieved a high level of accuracy such as
reporting words for clear speech or distinguishing coherent and anomalous sentences. Further,
they could potentially account for the poorer task performance when challenge was intro-
duced. This concern is based on previous research that demonstrates a variety of higher-level
cognitive consequences of hearing impairment on tasks requiring speech perception and com-
prehension (for a review, see Humes et al., 2012). For example, a participant with a hearing
impairment may experience increased demands on domain-general functions depending on
the task, and the extent to which they can manage these demands may therefore depend on
cognitive abilities. As above, we had no reason to suppose differential effects of any auditory
or cognitive impairments on our different linguistic tasks, but we acknowledge the limitation
and suggest that future studies should measure auditory and cognitive abilities more broadly
alongside linguistic measures of interest.
Further studies exploring a wider range of challenges to speech comprehension and with
larger samples of participants might specify the causal contributions identified here in more
detail. For example, we might assess lesion correlates of perception and adaptation for other
forms of perceptual challenges to speech comprehension such as those arising when speech is
in background noise, or speech sounds are perceptually ambiguous (see Mattys et al., 2012,
for a review of these listening challenges). Future studies might also consider whether other
forms of semantic, syntactic, or lexical challenge to comprehension are also causally associ-
ated with the integrity of the MD or language networks. In this way, building on the current
methods and findings, one could map the hierarchy of cognitive processes involved in speech
perception and comprehension onto specific brain regions that support them. However, as
mentioned above, larger samples of patients will be needed if we are to conduct more ana-
tomically specific analyses at the level of individual voxels (e.g., using voxel-based lesion-
symptom mapping) or functional subregions within the larger networks studied here.
Conclusions
Speech comprehension in naturalistic situations requires listeners to accommodate and learn
in response to a range of perceptual and semantic challenges that make spoken sentences
more difficult to recognise and understand. Behavioural data from individuals with lesions
to language-selective and domain-general MD networks demonstrate different functional con-
tributions of these two networks depending on the source of the listening challenge. In partic-
ular, the MD network appears to be necessary for the perception of acoustically degraded
speech, whereas using recent experience to update meaning preferences for ambiguous words
appears to depend on anatomically distinct, frontotemporal regions argued to form a special-
ised language network.
In this work we considered two specific challenges, but future work should consider
whether differences in the ways in which acoustic degradation and lexical-semantic ambiguity
Neurobiology of Language
691
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
engage and depend on the domain-general MD network and domain-selective language net-
work translate to other perceptual and semantic challenges and to more naturalistic speech
processing. For example, speech perception must be resilient in the face of unfamiliar accents,
mispronunciations, and competing sounds. Comprehension processes must accommodate
multiple forms of syntactically or semantically complex and ambiguous speech. Many of these
are situations in which activation of inferior frontal regions has been observed (Blanco-
Elorrieta et al., 2021; Boudewyn et al., 2015; January et al., 2009; Kuperberg et al., 2003;
Novais-Santos et al., 2007) and an attribution to domain-general MD processing has some-
times been made. However, given the evidence for functionally distinct language-selective
and domain-general subregions lying in close proximity within the IFG, and the individual
variability in their precise locations (Fedorenko & Blank, 2020), such conclusions may be pre-
mature. Further studies of individuals with focal lesions can be used to determine whether
accommodating these other perceptual and semantic challenges to speech processing simi-
larly depends on the integrity of domain-general or language-selective brain regions. These
perceptual and semantic challenges are common for the noisy and ambiguous spoken lan-
guage that listeners perceive and comprehend every day.
ACKNOWLEDGMENTS
We would like to thank Rahel Schumacher and Vitor Zimmerer for discussion, and Peter
Watson for statistical advice. We also thank two anonymous reviewers for helpful comments
on earlier versions of the manuscript.
FUNDING INFORMATION
Matthew H. Davis, Medical Research Council UK, Award ID: MC_UU_00005/5. John
Duncan, Medical Research Council UK, Award ID: MC_UU_00005/6. Evelina Fedorenko,
National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID: R01-
DC016950. Evelina Fedorenko, National Institutes of Health (https://dx.doi.org/10.13039
/100000002), Award ID: R01-DC016607. Evelina Fedorenko, McGovern Institute for Brain
Research. Jennifer M. Rodd, Economic and Social Research Council UK, Award ID:
ES/S009752/1.
AUTHOR CONTRIBUTIONS
Lucy J. MacGregor: Data curation: Equal; Formal analysis: Lead; Methodology: Equal; Project
administration: Equal; Software: Lead; Visualization: Lead; Writing—original draft: Lead;
Writing—review & editing: Equal. Rebecca A. Gilbert: Data curation: Equal; Formal analysis:
Supporting; Methodology: Equal; Software: Supporting; Visualization: Supporting; Writing—
review & editing: Supporting. Zuzanna Balewski: Investigation: Lead. Daniel J. Mitchell: Data
curation: Equal; Methodology: Supporting; Software: Supporting; Writing—review & editing:
Supporting. Sharon W. Erzinçlioğlu: Data curation: Equal; Investigation: Supporting. Jennifer
M. Rodd: Conceptualization: Equal; Funding acquisition: Supporting; Resources: Equal;
Writing—review & editing: Supporting. John Duncan: Conceptualization: Equal; Funding
acquisition: Equal; Project administration: Equal; Resources: Equal; Supervision: Supporting;
Writing—review & editing: Supporting. Evelina Fedorenko: Conceptualization: Equal; Funding
acquisition: Equal; Project administration: Equal; Resources: Equal; Supervision: Supporting;
Writing—review & editing: Supporting. Matthew H. Davis: Conceptualization: Equal; Funding
acquisition: Equal; Methodology: Equal; Project administration: Equal; Resources: Equal;
Supervision: Lead; Writing—original draft: Supporting; Writing—review & editing: Equal.
Neurobiology of Language
692
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
DATA AVAILABILITY STATEMENT
Stimuli, data and analysis code are available at https://osf.io/fm67z/.
REFERENCES
Adank, P. (2012). The neural bases of difficult speech comprehen-
sion and speech production: Two activation likelihood estima-
tion (ALE) meta-analyses. Brain and Language, 122(1), 42–54.
https://doi.org/10.1016/j.bandl.2012.04.014, PubMed:
22633697
Altmann, G. T., & Kamide, Y. (1999). Incremental interpretation at
verbs: Restricting the domain of subsequent reference. Cognition,
73(3), 247–264. https://doi.org/10.1016/S0010-0277(99)00059-1,
PubMed: 10585516
Apperly, I. A., Samson, D., Carroll, N., Hussain, S., & Humphreys,
G. (2006). Intact first- and second-order false belief reasoning in
a patient with severely impaired grammar. Social Neuroscience,
1(3–4), 334–348. https://doi.org/10.1080/17470910601038693,
PubMed: 18633798
Assem, M., Blank, I. A., Mineroff, Z., Ademoğlu, A., & Fedorenko,
E. (2020). Activity in the fronto-parietal multiple-demand net-
work is robustly associated with individual differences in work-
ing memory and fluid intelligence. Cortex, 131, 1–16. https://doi
.org/10.1016/j.cortex.2020.06.013, PubMed: 32777623
Assem, M., Glasser, M. F., Van Essen, D. C., & Duncan, J. (2020). A
domain-general cognitive core defined in multimodally parcel-
lated human cortex. Cerebal Cortex, 30(8), 4361–4380. https://
doi.org/10.1093/cercor/bhaa023, PubMed: 32244253
Baayen, R. H., & Milin, P. (2010). Analyzing reaction times. Inter-
national Journal of Psychological Research, 3(2), 12–28. https://
doi.org/10.21500/20112084.807
Bates, D., Mächler, M., Bolker, B. M., & Walker, S. C. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical Soft-
ware, 67(1), 1–48. https://doi.org/10.18637/jss.v067.i01
Bates, E., Wilson, S. M., Saygin, A. P., Dick, F., Sereno, M. I.,
Knight, R. T., & Dronkers, N. F. (2003). Voxel-based lesion–
symptom mapping. Nature Neuroscience, 6(5), 448–450. https://
doi.org/10.1038/nn1050, PubMed: 12704393
Betts, H. N., Gilbert, R. A., Cai, Z. G., Okedara, Z. B., & Rodd, J. M.
(2018). Retuning of lexical-semantic representations: Repetition
and spacing effects in word-meaning priming. Journal of Experi-
mental Psychology: Learning, Memory and Cognition, 44(7),
1130–1150. https://doi.org/10.1037/xlm0000507, PubMed:
29283607
Bilenko, N. Y., Grindrod, C. M., Myers, E. B., & Blumstein, S. E.
(2009). Neural correlates of semantic competition during pro-
cessing of ambiguous words. Journal of Cognitive Neuroscience,
21(5), 960–975. https://doi.org/10.1162/jocn.2009.21073,
PubMed: 18702579
Binder, J. R., Frost, J. A., Hammeke, T. A., Cox, R. W., Rao, S. M., &
Prieto, T. (1997). Human brain language areas identified by
functional magnetic resonance imaging. Journal of Neurosci-
ence, 17(1), 353–362. https://doi.org/10.1523/ JNEUROSCI.17
-01-00353.1997, PubMed: 8987760
Bishop, D. (2003). Test for reception of grammar (TROG-2).
Pearson.
Blanco-Elorrieta, E., Gwilliams, L., Marantz, A., & Pylkkanen, L.
(2021). Adaptation to mis-pronounced speech: Evidence for a
prefrontal-cortex repair mechanism. Scientific Reports, 11(1), 97.
https://doi.org/10.1038/s41598-020-79640-0, PubMed:
33420193
Blank, I. A., & Fedorenko, E. (2017). Domain-general brain regions
do not track linguistic input as closely as language-selective
regions. Journal of Neuroscience, 37(41), 9999–10011. https://
doi.org/10.1523/ JNEUROSCI.3642-16.2017, PubMed:
28871034
Blank, I. A., Kanwisher, N., & Fedorenko, E. (2014). A functional
dissociation between language and multiple-demand systems
revealed in patterns of BOLD signal fluctuations. Journal of Neu-
rophysiology, 112(5), 1105–1118. https://doi.org/10.1152/jn
.00884.2013, PubMed: 24872535
Blott, L. M., Rodd, J. M., Ferreira, F., & Warren, J. E. (2021). Recov-
ery from misinterpretations during online sentence processing.
Journal of Experimental Psychology: Learning, Memory, and Cog-
nition, 47(6), 968–997. https://doi.org/10.1037/xlm0000936,
PubMed: 33252925
Boudewyn, M. A., Long, D. L., Traxler, M. J., Lesh, T. A., Dave, S.,
Mangun, G. R., Carter, C. S., & Swaab, T. Y. (2015). Sensitivity to
referential ambiguity in discourse: The role of attention, working
memory, and verbal ability. Journal of Cognitive Neuroscience,
27(12), 2309–2323. https://doi.org/10.1162/jocn_a_00837,
PubMed: 26401815
Braga, R. M., DiNicola, L. M., Becker, H. C., & Buckner, R. L.
(2020). Situating the left-lateralized language network in the
broader organization of multiple specialized large-scale distrib-
uted networks. Journal of Neurophysiology, 124(5), 1415–1448.
https://doi.org/10.1152/jn.00753.2019, PubMed: 32965153
Campbell, K. L., & Tyler, L. K. (2018). Language-related domain-
specific and domain-general systems in the human brain. Current
Opinions in Behavioral Sciences, 21, 132–137. https://doi.org/10
.1016/j.cobeha.2018.04.008, PubMed: 30057936
Cole, M., & Schneider, W. (2007). The cognitive control network:
Integrated cortical regions with dissociable functions. Neuro-
Image, 37(1), 343–360. https://doi.org/10.1016/j.neuroimage
.2007.03.071, PubMed: 17553704
Cutler, A., Dahan, D., & van Donselaar, W. (1997). Prosody in the
comprehension of spoken language: A literature review. Lan-
guage and Speech, 40(Part 2), 141–201. https://doi.org/10.1177
/002383099704000203, PubMed: 9509577
D’Ausilio, A., Bufalari, I., Salmas, P., & Fadiga, L. (2012). The role
of the motor system in discriminating normal and degraded
speech sounds. Cortex, 48(7), 882–887. https://doi.org/10.1016
/j.cortex.2011.05.017, PubMed: 21676385
D’Ausilio, A., Pulvermüller, F., Salmas, P., Bufalari, I., Begliomini,
C., & Fadiga, L. (2009). The motor somatotopy of speech percep-
tion. Current Biology, 19(5), 381–385. https://doi.org/10.1016/j
.cub.2009.01.017, PubMed: 19217297
Davis, M. H., Ford, M. A., Kherif, F., & Johnsrude, I. S. (2011). Does
semantic context benefit speech understanding through
“top-down” processes? Evidence from time-resolved sparse fMRI.
Journal of Cognitive Neuroscience, 23(12), 3914–3932. https://
doi.org/10.1162/jocn_a_00084, PubMed: 21745006
Davis, M. H., & Gaskell, M. G. (2009). A complementary systems
account of word learning: Neural and behavioural evidence.
Philosophical Transactions of the Royal Society B: Biological
Sciences, 364(1536), 3773–3800. https://doi.org/10.1098/rstb
.2009.0111, PubMed: 19933145
Neurobiology of Language
693
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
Davis, M. H., & Johnsrude, I. S. (2003). Hierarchical processing
in spoken language comprehension. Journal of Neuroscience,
23(8), 3423–3431. https://doi.org/10.1523/ JNEUROSCI.23-08
-03423.2003, PubMed: 12716950
Davis, M. H., Johnsrude, I. S., Hervais-Adelman, A., Taylor, K., &
McGettigan, C. (2005). Lexical information drives perceptual
learning of distorted speech: Evidence from the comprehension
of noise-vocoded sentences. Journal of Experimental Psychology:
General, 134(2), 222–241. https://doi.org/10.1037/0096-3445
.134.2.222, PubMed: 15869347
Diachek, E., Blank, I. A., Siegelman, M., Affourtit, J., & Fedorenko,
E. (2020). The domain-general multiple demand (MD) network
does not support core aspects of language comprehension: A
large-scale fMRI investigation. Journal of Neuroscience, 40(23),
4536–4550. https://doi.org/10.1523/JNEUROSCI.2036-19.2020,
PubMed: 32317387
Diedenhofen, B., & Musch, J. (2015). cocor: A comprehensive
solution for the statistical comparison of correlations. PLOS
ONE, 10(3), e0121945. https://doi.org/10.1371/journal.pone
.0121945, PubMed: 25835001
Dosenbach, N. U., Fair, D. A., Cohen, A. L., Schlaggar, B. L., &
Petersen, S. E. (2008). A dual-networks architecture of
top-down control. Trends in Cognitive Sciences, 12(3), 99–105.
https://doi.org/10.1016/j.tics.2008.01.001, PubMed: 18262825
Dosenbach, N. U., Fair, D. A., Miezin, F. M., Cohen, A. L., Wenger,
K. K., Dosenbach, R. A., Fox, M. D., Snyder, A. Z., Vincent, J. L.,
Raichle, M. E., Schlaggar, B. L., & Petersen, S. E. (2007). Distinct
brain networks for adaptive and stable task control in humans.
Proceedings of the National Academy of Sciences, 104(26),
11073–11078. https://doi.org/10.1073/pnas.0704320104,
PubMed: 17576922
Du, Y., Buchsbaum, B. R., Grady, C. L., & Alain, C. (2014). Noise
differentially impacts phoneme representations in the auditory
and speech motor systems. Proceedings of the National Academy
of Sciences, 111(19), 7126–7131. https://doi.org/10.1073/pnas
.1318738111, PubMed: 24778251
Du, Y., Buchsbaum, B. R., Grady, C. L., & Alain, C. (2016).
Increased activity in frontal motor cortex compensates impaired
speech perception in older adults. Nature Communications, 7,
12241. https://doi.org/10.1038/ncomms12241, PubMed:
27483187
Duffy, S. A., Morris, R. K., & Rayner, K. (1988). Lexical ambiguity
and fixation times in reading. Journal of Memory and Language,
27(4), 429–446. https://doi.org/10.1016/0749-596X(88)90066-6
Dumay, N., & Gaskell, M. G. (2007). Sleep-associated changes in
the mental representation of spoken words. Psychological
Science, 18(1), 35–39. https://doi.org/10.1111/j.1467-9280
.2007.01845.x, PubMed: 17362375
Duncan, J. (2010a). How intelligence happens. Yale University
Press.
Duncan, J. (2010b). The multiple-demand (MD) system of the pri-
mate brain: Mental programs for intelligent behaviour. Trends in
Cognitive Sciences, 14(4), 172–179. https://doi.org/10.1016/j.tics
.2010.01.004, PubMed: 20171926
Duncan, J. (2013). The structure of cognition: Attentional episodes
in mind and brain. Neuron, 80(1), 35–50. https://doi.org/10.1016
/j.neuron.2013.09.015, PubMed: 24094101
Duncan, J., & Owen, A. M. (2000). Common regions of the human
frontal lobe recruited by diverse cognitive demands. Trends in
Neuroscience, 23(10), 475–483. https://doi.org/10.1016/S0166
-2236(00)01633-7, PubMed: 11006464
Elliott, T. M., & Theunissen, F. E. (2009). The modulation transfer
function for speech intelligibility. PLOS Computational Biology,
5(3), e1000302. https://doi.org/10.1371/journal.pcbi.1000302,
PubMed: 19266016
Erb, J., Henry, M. J., Eisner, F., & Obleser, J. (2013). The brain
dynamics of rapid perceptual adaptation to adverse listening
conditions. Journal of Neuroscience, 33(26), 10688–10697.
https://doi.org/10.1523/ JNEUROSCI.4596-12.2013, PubMed:
23804092
Evans, S., & Davis, M. H. (2015). Hierarchical organization of audi-
tory and motor representations in speech perception: Evidence
from searchlight similarity analysis. Cerebral Cortex, 25(12),
4772–4788. https://doi.org/10.1093/cercor/ bhv136, PubMed:
26157026
Fedorenko, E. (2014). The role of domain-general cognitive control
in language comprehension. Frontiers in Psychology, 5, 335.
https://doi.org/10.3389/fpsyg.2014.00335, PubMed: 24803909
Fedorenko, E. (2021). The early origins and the growing popularity
of the individual-subject analytic approach in human neurosci-
ence. Current Opinion in Behavioral Sciences, 40, 105–112.
https://doi.org/10.1016/j.cobeha.2021.02.023
Fedorenko, E., Behr, M. K., & Kanwisher, N. (2011). Functional
specificity for high-level linguistic processing in the human brain.
Proceedings of the National Academy of Sciences, 108(39),
16428–16433. https://doi.org/10.1073/pnas.1112937108,
PubMed: 21885736
Fedorenko, E., & Blank, I. A. (2020). Broca’s area is not a natural
kind. Trends in Cognitive Sciences, 24(4), 270–284. https://doi
.org/10.1016/j.tics.2020.01.001, PubMed: 32160565
Fedorenko, E., Duncan, J., & Kanwisher, N. (2012). Language-
selective and domain-general regions lie side by side within Bro-
ca’s area. Current Biology, 22(21), 2059–2062. https://doi.org/10
.1016/j.cub.2012.09.011, PubMed: 23063434
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad domain
generality in focal regions of frontal and parietal cortex. Proceed-
ings of the National Academy of Sciences, 110(41), 16616–
16621. https://doi.org/10.1073/pnas.1315235110, PubMed:
24062451
Fedorenko, E., Hsieh, P.-J., Nieto-Castañón, A., Whitfield-Gabrieli,
S., & Kanwisher, N. (2010). New method for fMRI investigations
of language: Defining ROIs functionally in individual subjects.
Journal of Neurophysiology, 104(2), 1177–1194. https://doi.org
/10.1152/jn.00032.2010, PubMed: 20410363
Fedorenko, E., & Shain, C. (2021). Similarity of computations across
domains does not imply shared implementation: The case of lan-
guage comprehension. Current Directions in Psychological Science,
30(6), 526–534. https://doi.org/10.1177/09637214211046955,
PubMed: 35295820
Fedorenko, E., & Varley, R. A. (2016). Language and thought are not
the same thing: Evidence from neuroimaging and neurological
patients. Annals of the New York Academy of Sciences, 1369(1),
132–153. https://doi.org/10.1111/nyas.13046, PubMed:
27096882
Garrod, S., & Pickering, M. J. (2004). Why is conversation so easy?
Trends in Cognitive Sciences, 8(1), 8–11. https://doi.org/10.1016/j
.tics.2003.10.016, PubMed: 14697397
Gaskell, M. G., Cairney, S. A., & Rodd, J. M. (2019). Contextual
priming of word meanings is stabilized over sleep. Cognition,
182, 109–126. https://doi.org/10.1016/j.cognition.2018.09.007,
PubMed: 30227332
Gaskell, M. G., & Dumay, N. (2003). Lexical competition and the
acquisition of novel words. Cognition, 89(2), 105–132. https://
doi.org/10.1016/S0010-0277(03)00070-2, PubMed: 12915296
Gernsbacher, M. A., & Faust, M. E. (1991). The mechanism of sup-
pression: A component of general comprehension skill. Journal
Neurobiology of Language
694
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
of Experimental Psychology: Learning, Memory, and Cognition,
17(2), 245–262. https://doi.org/10.1037/0278-7393.17.2.245,
PubMed: 1827830
Gernsbacher, M. A., Varner, K. R., & Faust, M. E. (1990). Investigat-
ing differences in general comprehension skill. Journal of Exper-
imental Psychology: Learning, Memory, and Cognition, 16(3),
430–445. https://doi.org/10.1037/0278-7393.16.3.430,
PubMed: 2140402
Gilbert, R. A., Davis, M. H., Gaskell, M. G., & Rodd, J. M. (2018).
Listeners and readers generalize their experience with word
meanings across modalities. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 44(10), 1533–1561. https://
doi.org/10.1037/xlm0000532, PubMed: 29389181
Gilbert, R. A., & Rodd, J. M. (2022). Dominance norms and data for
spoken ambiguous words in British English. Journal of Cognition,
5(1), 4. https://doi.org/10.5334/joc.194, PubMed: 36072113
Guediche, S., Blumstein, S. E., Fiez, J. A., & Holt, L. L. (2014).
Speech perception under adverse conditions: Insights from
behavioral, computational, and neuroscience research. Frontiers
in Systems Neuroscience, 7, 126. https://doi.org/10.3389/fnsys
.2013.00126, PubMed: 24427119
Hagoort, P. (1993). Impairments of lexical semantic processing in
aphasia: Evidence from the processing of lexical ambiguities.
Brain and Language, 45(2), 189–232. https://doi.org/10.1006
/brln.1993.1043, PubMed: 8358597
Hagoort, P., Hald, L., Bastiaansen, M., & Petersson, K. M. (2004).
Integration of word meaning and world knowledge in language
comprehension. Science, 304(5669), 438–441. https://doi.org/10
.1126/science.1095455, PubMed: 15031438
Halai, A. D., Woollams, A. M., & Lambon Ralph, M. A. (2017).
Using principal component analysis to capture individual differ-
ences within a unified neuropsychological model of chronic
post-stroke aphasia: Revealing the unique neural correlates of
speech fluency, phonology and semantics. Cortex, 86, 275–289.
https://doi.org/10.1016/j.cortex.2016.04.016, PubMed:
27216359
Hardy, C. J. D., Marshall, C. R., Bond, R. L., Russell, L. L., Dick, K.,
Ariti, C., Thomas, D. L., Ross, S. J., Agustus, J. L., Crutch, S. J.,
Rohrer, J. D., Bamiou, D.-E., & Warren, J. D. (2018). Retained
capacity for perceptual learning of degraded speech in primary
progressive aphasia and Alzheimer’s disease. Alzheimer’s
Research & Therapy, 10(1), 70. https://doi.org/10.1186/s13195
-018-0399-2, PubMed: 30045755
Hervais-Adelman, A., Carlyon, R. P., Johnsrude, I. S., & Davis,
M. H. (2012). Brain regions recruited for the effortful comprehen-
sion of noise-vocoded words. Language and Cognitive Processes,
27(7–8), 1145–1166. https://doi.org/10.1080/01690965.2012
.662280
Hervais-Adelman, A., Davis, M. H., Johnsrude, I. S., & Carlyon,
R. P. (2008). Perceptual learning of noise vocoded words: Effects
of feedback and lexicality. Journal of Experimental Psychology:
Human Perception and Performance, 34(2), 460–474. https://
doi.org/10.1037/0096-1523.34.2.460, PubMed: 18377182
Hugdahl, K., Raichle, M. E., Mitra, A., & Specht, K. (2015). On the
existence of a generalized non-specific task-dependent network.
Frontiers in Human Neuroscience, 9, 430. https://doi.org/10
.3389/fnhum.2015.00430, PubMed: 26300757
Humes, L. E., Dubno, J. R., Gordon-Salant, S., Lister, J. J., Cacace,
A. T., Cruickshanks, K. J., Gates, G. A., Wilson, R. H., &
Wingfield, A. (2012). Central presbycusis: A review and evalua-
tion of the evidence. Journal of the American Academy of
Audiology, 23(8), 635–666. https://doi.org/10.3766/jaaa.23.8.5,
PubMed: 22967738
Huyck, J. J., & Johnsrude, I. S. (2012). Rapid perceptual learning of
noise-vocoded speech requires attention. Journal of the Acous-
tical Society of America, 131(3), EL236–EL242. https://doi.org
/10.1121/1.3685511, PubMed: 22423814
Huyck, J. J., Smith, R. H., Hawkins, S., & Johnsrude, I. S. (2017).
Generalization of perceptual learning of degraded speech across
talkers. Journal of Speech, Language, and Hearing Research,
60(11), 3334–3341. https://doi.org/10.1044/2017_JSLHR-H-16
-0300, PubMed: 28979990
Ivanova, A. A., Mineroff, Z., Zimmerer, V., Kanwisher, N., Varley, R.,
& Fedorenko, E. (2021). The language network is recruited but
not required for nonverbal event semantics. Neurobiology of
Language, 2(2), 176–201. https://doi.org/10.1162/nol_a_00030
Ivanova, A. A., Srikant, S., Sueoka, Y., Kean, H. H., Dhamala, R.,
O’Reilly, U.-M., Bers, M. U., & Fedorenko, E. (2020). Compre-
hension of computer code relies primarily on domain-general
executive brain regions. Elife, 9, e58906. https://doi.org/10
.7554/eLife.58906, PubMed: 33319744
January, D., Trueswell, J. C., & Thompson-Schill, S. L. (2009). Co-
localization of stroop and syntactic ambiguity resolution in Bro-
ca’s area: Implications for the neural basis of sentence processing.
Journal of Cognitive Neuroscience, 21(12), 2434–2444. https://doi
.org/10.1162/jocn.2008.21179, PubMed: 19199402
Johnsrude, I. S., & Rodd, J. R. (2015). Factors that increase process-
ing demands when listening to speech. In G. Hickok & S. Small
(Eds.), Neurobiology of language (pp. 491–502). Academic Press.
https://doi.org/10.1121/1.4920048
Kelly, H., & Armstrong, L. (2009) New word learning in people with
aphasia. Aphasiology, 23(12), 1398–1417. https://doi.org/10
.1080/02687030802289200
Khanna, M. M., & Boland, J. E. (2010). Children’s use of language
context in lexical ambiguity resolution. Quarterly Journal of
Experimental Psychology, 63(1), 160–193. https://doi.org/10
.1080/17470210902866664, PubMed: 19424907
Kuperberg, G. R., Holcomb, P. J., Sitnikova, T., Greve, D., Dale,
A. M., & Caplan, D. (2003). Distinct patterns of neural modula-
tion during the processing of conceptual and syntactic anoma-
lies. Journal of Cognitive Neuroscience, 15(2), 272–293. https://
doi.org/10.1162/089892903321208204, PubMed: 12676064
Linares, D., & López-Moliner, J. (2006). quickpsy: An R package to
fit psychometric functions for multiple groups. The R Journal,
8(1), 122–131. https://doi.org/10.32614/RJ-2016-008
Loebach, J. L., & Pisoni, D. B. (2008). Perceptual learning of spec-
trally degraded speech and environmental sounds. Journal of the
Acoustical Society of America, 123(2), 1126–1139. https://doi.org
/10.1121/1.2823453, PubMed: 18247913
Macmillan, N. A., & Kaplan, H. L. (1985). Detection theory analysis
of group data: Estimating sensitivity from average hit and
false-alarm rates. Psychological Bulletin, 98(1), 185–199.
https://doi.org/10.1037/0033-2909.98.1.185, PubMed: 4034817
MacSweeney, M., Woll, B., Campbell, R., McGuire, P. K., David, A. S.,
Williams, S. C. R., Suckling, J., Calvert, G. A., & Brammer, M. J.
(2002). Neural systems underlying British Sign Language and
audio-visual English processing in native users. Brain, 125(Part 7),
1583–1593. https://doi.org/10.1093/ brain/awf153, PubMed:
12077007
Mahowald, K., & Fedorenko, E. (2016). Reliable individual-level
neural markers of high-level language processing: A necessary
precursor for relating neural variability to behavioral and genetic
variability. NeuroImage, 139, 74–93. https://doi.org/10.1016/j
.neuroimage.2016.05.073, PubMed: 27261158
Mattys, S. L., Davis, M. H., Bradlow, A. R., & Scott, S. K. (2012).
Speech recognition in adverse conditions: A review. Language
Neurobiology of Language
695
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
and Cognitive Processes, 27(7–8), 953–978. https://doi.org/10
.1080/01690965.2012.705006
McClelland, J. L. (2013). Incorporating rapid neocortical learning of
new schema-consistent information into complementary learn-
ing systems theory. Journal of Experimental Psychology: General,
142(4), 1190–1210. https://doi.org/10.1037/a0033812, PubMed:
23978185
McGettigan, C., Rosen, S., & Scott, S. K. (2014). Lexico-semantic
and acoustic-phonetic processes in the perception of
noise-vocoded speech: Implications for cochlear implantation.
Frontiers in Systems Neuroscience, 8, 18. https://doi.org/10
.3389/fnsys.2014.00018, PubMed: 24616669
Meister, I. G., Wilson, S. M., Deblieck, C., Wu, A. D., & Iacoboni,
M. (2007). The essential role of premotor cortex in speech per-
ception. Current Biology, 17(19), 1692–1696. https://doi.org/10
.1016/j.cub.2007.08.064, PubMed: 17900904
Meng, X.-L., Rosenthal, R., & Rubin, D. B. (1992). Comparing cor-
related correlation coefficients. Psychological Bulletin, 111(1),
172–175. https://doi.org/10.1037/0033-2909.111.1.172
Mesulam, M. M., Rogalski, E. J., Wieneke, C., Hurley, R. S., Geula, C.,
Bigio, E. H., Thompson, C. K., & Weintraub, S. (2014). Primary
progressive aphasia and the evolving neurology of the language
network. Nature Reviews Neurology, 10(10), 554–569. https://doi
.org/10.1038/nrneurol.2014.159, PubMed: 25179257
Miller, G. A., Heise, G. A., & Lichten, W. (1951). The intelligibility
of speech as a function of the context of the test materials. Journal
of Experimental Psychology, 41(5), 329–335. https://doi.org/10
.1037/h0062491, PubMed: 14861384
Miller, G. A., & Isard, S. (1963). Some perceptual consequences of
linguistic rules. Journal of Verbal Learning and Verbal Behavior,
2(3), 217–228. https://doi.org/10.1016/S0022-5371(63)80087-0
Mineroff, Z., Blank, I. A., Mahowald, K., & Fedorenko, E. (2018). A
robust dissociation among the language, multiple demand, and
default mode networks: Evidence from inter-region correlations
in effect size. Neuropsychologia, 119, 501–511. https://doi.org
/10.1016/j.neuropsychologia.2018.09.011, PubMed: 30243926
Mirman, D., Chen, Q., Zhang, Y. S., Wang, Z., Faseyitan, O. K.,
Coslett, H. B., & Schwartz, M. F. (2015). Neural organization
of spoken language revealed by lesion-symptom mapping.
Nature Communications, 6, 6762. https://doi.org/10.1038
/ncomms7762, PubMed: 25879574
Mirman, D., & Thye, M. (2018). Uncovering the neuroanatomy of
core language systems using lesion-symptom mapping. Current
Directions in Psychological Science, 27(6), 455–461. https://doi
.org/10.1177/0963721418787486
Moineau, S., Dronkers, N. F., & Bates, E. (2005). Exploring the pro-
cessing continuum of single-word comprehension in aphasia.
Journal of Speech, Language, and Hearing Research, 48(4),
884–896. https://doi.org/10.1044/1092-4388(2005/061),
PubMed: 16378480
Monti, M. M., Parsons, L. M., & Osherson, D. N. (2012). Thought
beyond language: Neural dissociation of algebra and natural
language. Psychological Science, 23(8), 914–922. https://doi
.org/10.1177/0956797612437427, PubMed: 22760883
Morey, R. D. (2008). Confidence intervals from normalized data:
A correction to Cousineau (2005). Tutorial in Quantitative
Methods for Psychology, 4(2), 61–64. https://doi.org/10.20982
/tqmp.04.2.p061
Münster, K., & Knoeferle, P. (2018). Extending situated language
comprehension (accounts) with speaker and comprehender char-
acteristics: Toward socially situated interpretation. Frontiers in
Psychology, 8, 2267. https://doi.org/10.3389/fpsyg.2017.02267,
PubMed: 29416517
Musz, E., & Thompson-Schill, S. L. (2017). Tracking competition and
cognitive control during language comprehension with multi-voxel
pattern analysis. Brain and Language, 165, 21–32. https://doi.org/10
.1016/j.bandl.2016.11.002, PubMed: 27898341
Nelson, H. E. (1982). National Adult Reading Test (NART): For the
assessment of premorbid intelligence in patients with dementia:
Test manual. NFER-Nelson.
Nomura, E. M., Gratton, C., Visser, R. M., Kayser, A., Perez, F., &
D’Esposito, M. (2010). Double dissociation of two cognitive con-
trol networks in patients with focal brain lesions. Proceedings of
the National Academy of Sciences, 107(26), 12017–12022.
https://doi.org/10.1073/pnas.1002431107, PubMed: 20547857
Novais-Santos, S., Gee, J., Shah, M., Troiani, V., Work, M., &
Grossman, M. (2007). Resolving sentence ambiguity with
planning and working memory resources: Evidence from fMRI.
NeuroImage, 37(1), 361–378. https://doi.org/10.1016/j
.neuroimage.2007.03.077, PubMed: 17574445
Novick, J. M., Trueswell, J. C., & Thompson-Schill, S. L. (2005).
Cognitive control and parsing: Reexamining the role of Broca’s
area in sentence comprehension. Cognitive Affective and Behav-
ioral Neuroscience, 5(3), 263–281. https://doi.org/10.3758
/CABN.5.3.263, PubMed: 16396089
Obleser, J., Wise, R. J., Dresner, M. A., & Scott, S. K. (2007). Func-
tional integration across brain regions improves speech percep-
tion under adverse listening conditions. Journal of Neuroscience,
27(9), 2283–2289. https://doi.org/10.1523/JNEUROSCI.4663-06
.2007, PubMed: 17329425
Özyürek, A. (2014). Hearing and seeing meaning in speech and
gesture: Insights from brain and behaviour. Philosophical Trans-
actions of the Royal Society B: Biological Sciences, 369(1651),
20130296. https://doi.org/10.1098/rstb.2013.0296, PubMed:
25092664
Paunov, A. M., Blank, I. A., & Fedorenko, E. (2019). Functionally
distinct language and theory of mind networks are synchronized
at rest and during language comprehension. Journal of Neuro-
physiology, 121(4), 1244–1265. https://doi.org/10.1152/jn
.00619.2018, PubMed: 30601693
Peelle, J. E. (2018). Listening effort: How the cognitive conse-
quences of acoustic challenge are reflected in brain and behav-
ior. Ear and Hearing, 39(2), 204–214. https://doi.org/10.1097
/AUD.0000000000000494, PubMed: 28938250
Peelle, J. E., Gross, J., & Davis, M. H. (2013). Phase-locked
responses to speech in human auditory cortex are enhanced dur-
ing comprehension. Cerebral Cortex, 23(6), 1378–1387. https://
doi.org/10.1093/cercor/bhs118, PubMed: 22610394
Peelle, J. E., & Wingfield, A. (2005). Dissociations in perceptual
learning revealed by adult age differences in adaptation to
time-compressed speech. Journal of Experimental Psychology:
Human Perception and Performance, 31(6), 1315–1330. https://
doi.org/10.1037/0096-1523.31.6.1315, PubMed: 16366792
Pichora-Fuller, M. K., Kramer, S. E., Eckert, M. A., Edwards, B.,
Hornsby, B. W., Humes, L. E., Lemke, U., Lunner, T., Matthen,
M., Mackersie, C. L., Naylor, G., Phillips, N. A., Richter, M.,
Rudner, M., Sommers, M. S., Tremblay, K. L., & Wingfield, A.
(2016). Hearing impairment and cognitive energy: The framework
for understanding effortful listening (FUEL). Ear and Hearing, 37(S1),
5S–27S. https://doi.org/10.1097/AUD.0000000000000312,
PubMed: 27355771
Polk, M., & Kertesz, A. (1993). Music and language in degenerative
disease of the brain. Brain and Cognition, 22(1), 98–117. https://
doi.org/10.1006/brcg.1993.1027, PubMed: 7684592
Pulvermüller, F., & Fadiga, L. (2010). Active perception: Sensorimo-
tor circuits as a cortical basis for language. Nature Reviews
Neurobiology of Language
696
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
Neuroscience, 11(5), 351–360. https://doi.org/10.1038/nrn2811,
PubMed: 20383203
Quillen, I. A., Yen, M., & Wilson, S. M. (2021). Distinct neural cor-
relates of linguistic and non-linguistic demand. Neurobiology of
Language, 2(2), 202–225. https://doi.org/10.1162/nol_a_00031,
PubMed: 34585141
R Core Team. (2019). R: A language and environment for statistical
computing ( Version 3.6.1). Vienna, Austria.
Rayner, K., & Duffy, S. A. (1986). Lexical complexity and fixation
times in reading: Effects of word frequency, verb complexity, and
lexical ambiguity. Memory and Cognition, 14(3), 191–201.
https://doi.org/10.3758/BF03197692, PubMed: 3736392
Rodd, J. M. (2018). Lexical ambiguity. In S.-A. Rueschemeyer &
M. G. Gaskell (Eds.), The Oxford handbook of psycholinguistics
(2nd ed., pp. 96–117). Oxford University Press. https://doi.org/10
.1093/oxfordhb/9780198786825.013.5
Rodd, J. M. (2020). Settling into semantic space: An ambiguity-
focused account of word-meaning access. Perspectives in Psy-
chological Science, 15(2), 411–427. https://doi.org/10.1177
/1745691619885860, PubMed: 31961780
Rodd, J. M., Cai, Z. G. G., Betts, H. N., Hanby, B., Hutchinson, C.,
& Adler, A. (2016). The impact of recent and long-term experi-
ence on access to word meanings: Evidence from large-scale
internet-based experiments. Journal of Memory and Language,
87, 16–37. https://doi.org/10.1016/j.jml.2015.10.006
Rodd, J. M., Cutrin, B. L., Kirsch, H., Millar, A., & Davis, M. H.
(2013). Long-term priming of the meanings of ambiguous words.
Journal of Memory and Language, 68(2), 180–198. https://doi.org
/10.1016/j.jml.2012.08.002
Rodd, J. M., Davis, M. H., & Johnsrude, I. S. (2005). The neural
mechanisms of speech comprehension: fMRI studies of semantic
ambiguity. Cerebral Cortex, 15(8), 1261–1269. https://doi.org/10
.1093/cercor/bhi009, PubMed: 15635062
Rodd, J. M., Gaskell, G., & Marslen-Wilson, W. D. (2002). Making
sense of semantic ambiguity: Semantic competition in lexical
access. Journal of Memory and Language, 46(2), 245–266.
https://doi.org/10.1006/jmla.2001.2810
Rodd, J. M., Johnsrude, I. S., & Davis, M. H. (2010). The role of
domain-general frontal systems in language comprehension: Evi-
dence from dual-task interference and semantic ambiguity. Brain
and Language, 115(3), 182–188. https://doi.org/10.1016/j.bandl
.2010.07.005, PubMed: 20709385
Rodd, J. M., Johnsrude, I. S., & Davis, M. H. (2012). Dissociating
frontotemporal contributions to semantic ambiguity resolution
in spoken sentences. Cerebral Cortex, 22(8), 1761–1773.
https://doi.org/10.1093/cercor/bhr252, PubMed: 21968566
Rysop, A. U., Schmitt, L.-M., Obleser, J., & Hartwigsen, G. (2021).
Neural modelling of the semantic predictability gain under
challenging listening conditions. Human Brain Mapping, 42(1),
110–127. https://doi.org/10.1002/ hbm.25208, PubMed:
32959939
Schimke, E. A. E., Angwin, A. J., Cheng, B. B. Y., & Copland, D. A.
(2021). The effect of sleep on novel word learning in healthy
adults: A systematic review and meta-analysis. Psychonomic Bul-
letin and Review, 28(6), 1811–1838. https://doi.org/10.3758
/s13423-021-01980-3, PubMed: 34549375
Scott, T. L., Gallée, J., & Fedorenko, E. (2017). A new fun and robust
version of an fMRI localizer for the frontotemporal language sys-
tem. Cognitive Neuroscience, 8(3), 167–176. https://doi.org/10
.1080/17588928.2016.1201466, PubMed: 27386919
Seidenberg, M. S., Tanenhaus, M. K., Leiman, J. M., & Bienkowski,
M. (1982). Automatic access of the meanings of ambiguous
words in context: Some limitations of knowledge-based
processing. Cognitive Psychology, 14(4), 489–537. https://doi
.org/10.1016/0010-0285(82)90017-2
Shain, C., Blank, I. A., van Schijndel, M., Schuler, W., & Fedorenko,
E. (2020). fMRI reveals language-specific predictive coding
during naturalistic sentence comprehension. Neuropsychologia,
138, 107307. https://doi.org/10.1016/j.neuropsychologia.2019
.107307, PubMed: 31874149
Shannon, R. V., Zeng, F. G., Kamath, V., Wygonski, J., & Ekelid, M.
(1995). Speech recognition with primarily temporal cues. Science,
270(5234), 303–304. https://doi.org/10.1126/science.270.5234
.303, PubMed: 7569981
Shashidhara, S., Mitchell, D. J., Erez, Y., & Duncan, J. (2019).
Progressive recruitment of the frontoparietal multiple-demand
system with increased task complexity, time pressure, and reward.
Journal of Cognitive Neuroscience, 31(11), 1617–1630. https://doi
.org/10.1162/jocn_a_01440, PubMed: 31274390
Sohoglu, E., & Davis, M. H. (2016). Perceptual learning of
degraded speech by minimizing prediction error. Proceedings
of the National Academy of Sciences, 113(12), E1747–1756.
https://doi.org/10.1073/pnas.1523266113, PubMed: 26957596
Sohoglu, E., Peelle, J. E., Carlyon, R. P., & Davis, M. H. (2014). Top-
down influences of written text on perceived clarity of degraded
speech. Journal of Experimental Psychology: Human Perception
and Performance, 40(1), 186–199. https://doi.org/10.1037
/a0033206, PubMed: 23750966
Stacey, P. C., & Summerfield, A. Q. (2008). Comparison of word-,
sentence-, and phoneme-based training strategies in improving
the perception of spectrally distorted speech. Journal of Speech,
Language, and Hearing Research, 51(2), 526–538. https://doi.org
/10.1044/1092-4388(2008/038), PubMed: 18367694
Sumby, W. H., & Pollack, I. (1954). Visual contribution to speech
intelligibility in noise. Journal of the Acoustical Society of America,
26(2), 212–215. https://doi.org/10.1121/1.1907309
Swaab, T. Y., Brown, C., & Hagoort, P. (1998). Understanding
ambiguous words in sentence contexts: Electrophysiological
evidence for delayed contextual selection in Broca’s aphasia.
Neuropsychologia, 36(8), 737–761. https://doi.org/10.1016
/S0028-3932(97)00174-7, PubMed: 9751439
Swinney, D. A. (1979). Lexical access during sentence comprehen-
sion: (Re)consideration of context effects. Journal of Verbal
Learning and Verbal Behavior, 18, 645–659. https://doi.org/10
.1016/S0022-5371(79)90355-4
Swinney, D. A., Zurif, E., & Nicol, J. (1989). The effects of focal
brain damage on sentence processing: An examination of the
neurological organization of a mental module. Journal of Cogni-
tive Neuroscience, 1(1), 25–37. https://doi.org/10.1162/jocn
.1989.1.1.25, PubMed: 23968408
Tagarelli, K. M., Shattuck, K. F., Turkeltaub, P. E., & Ullman, M. T.
(2019). Language learning in the adult brain: A neuroanatomical
meta-analysis of lexical and grammatical learning. NeuroImage,
193, 178–200. https://doi.org/10.1016/j.neuroimage.2019.02
.061, PubMed: 30826361
Tahmasebi, A. M., Davis, M. H., Wild, C. J., Rodd, J. M.,
Hakyemez, H., Abolmaesumi, P., & Johnsrude, I. S. (2012). Is
the link between anatomical structure and function equally
strong at all cognitive levels of processing? Cerebral Cortex,
22(7), 1593–1603. https://doi.org/10.1093/cercor/ bhr205,
PubMed: 21893681
Tamminen, J., Payne, J. D., Stickgold, R., Wamsley, E. J., & Gaskell,
M. G. (2010). Sleep spindle activity is associated with the inte-
gration of new memories and existing knowledge. Journal of
Neuroscience, 30(43), 14356–14360. https://doi.org/10.1523
/JNEUROSCI.3028-10.2010, PubMed: 20980591
Neurobiology of Language
697
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks for challenges to speech comprehension
Thompson-Schill, S. L., D’Esposito, M., Aguirre, G. K., & Farah,
M. J. (1997). Role of left inferior prefrontal cortex in retrieval of
semantic knowledge: A reevaluation. Proceedings of the
National Academy of Sciences, 94(26), 14792–14797. https://
doi.org/10.1073/pnas.94.26.14792, PubMed: 9405692
Tuomiranta, L., Grönholm-Nyman, P., Kohen, F., Rautakoski, P.,
Laine, M., & Martin, N. (2011). Learning and maintaining new
vocabulary in persons with aphasia: Two controlled case studies.
Aphasiology, 25(9), 1030–1052. https://doi.org/10.1080
/02687038.2011.571384
Tuomiranta, L., Grönroos, A.-M., Martin, N., & Laine, M. (2014).
Vocabulary acquisition in aphasia: Modality can matter. Journal
of Neurolinguistics, 32, 42–58. https://doi.org/10.1016/j
.jneuroling.2014.08.006, PubMed: 25419049
Turken, A. U., & Dronkers, N. F. (2011). The neural architecture of
the language comprehension network: Converging evidence
from lesion and connectivity analyses. Frontiers in Systems Neu-
roscience, 5, 1. https://doi.org/10.3389/fnsys.2011.00001,
PubMed: 21347218
Vaden, K. I., Kuchinsky, S. E., Ahlstrom, J. B., Dubno, J. R., & Eckert,
M. A. (2015). Cortical activity predicts which older adults recog-
nize speech in noise and when. Journal of Neuroscience, 35(9),
3929–3937. https://doi.org/10.1523/JNEUROSCI.2908-14.2015,
PubMed: 25740521
Vaden, K. I., Kuchinsky, S. E., Cute, S. L., Ahlstrom, J. B., Dubno,
J. R., & Eckert, M. A. (2013). The cingulo-opercular network pro-
vides word-recognition benefit. Journal of Neuroscience, 33(48),
18979–18986. https://doi.org/10.1523/ JNEUROSCI.1417-13
.2013, PubMed: 24285902
Van Berkum, J. J. (2009). The neuropragmatics of ‘simple’ utterance
comprehension: An ERP review. In U. Sauerland & K. Yatsushiro
(Eds.), Semantics and pragmatics: From experiment to theory
(pp. 276–316). Palgrave Macmillan.
Varley, R. A., Klessinger, N. J., Romanowski, C. A., & Siegal, M.
(2005). Agrammatic but numerate. Proceedings of the National
Academy of Science, 102(9), 3519–3524. https://doi.org/10
.1073/pnas.0407470102, PubMed: 15713804
Varley, R. A., & Siegal, M. (2000). Evidence for cognition without
grammar from causal reasoning and ‘theory of mind’ in an
agrammatic aphasic patient. Current Biology, 10(12), 723–726.
https://doi.org/10.1016/S0960-9822(00)00538-8, PubMed:
10873809
Varley, R. A., Siegal, M., & Want, S. C. (2001). Severe impairment in
grammar does not preclude theory of mind. Neurocase, 7(6),
489–493. https://doi.org/10.1093/neucas/7.6.489, PubMed:
11788740
Vincent, J. L., Kahn, I., Snyder, A. Z., Raichle, M. E., & Buckner,
R. L. (2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Journal of Neuro-
physiology, 100(6), 3328–3342. https://doi.org/10.1152/jn
.90355.2008, PubMed: 18799601
Vitello, S., Warren, J. E., Devlin, J. T., & Rodd, J. M. (2014). Roles of
frontal and temporal regions in reinterpreting semantically
ambiguous sentences. Frontiers in Human Neuroscience, 8, 530.
https://doi.org/10.3389/fnhum.2014.00530, PubMed: 25120445
Wehbe, L., Blank, I. A., Shain, C., Futrell, R., Levy, R., von der
Malsburg, T., Smith, N., Gibson, E., & Fedorenko, E. (2021).
Incremental langauge comprehension difficulty predicts activity
in the language network but not the multiple demand network.
Cerebral Cortex, 31(9), 4006–4023. https://doi.org/10.1093
/cercor/bhab065, PubMed: 33895807
Wild, C. J., Yusuf, A., Wilson, D. E., Peelle, J. E., Davis, M. H., &
Johnsrude, I. S. (2012). Effortful listening: The processing of
degraded speech depends critically on attention. Journal of Neu-
roscience, 32(40), 14010–14021. https://doi.org/10.1523
/JNEUROSCI.1528-12.2012, PubMed: 23035108
Woolgar, A., Duncan, J., Manes, F., & Fedorenko, E. (2018). The
multiple-demand system but not the language system supports
fluid intelligence. Nature Human Behavior, 2(3), 200–204.
https://doi.org/10.1038/s41562-017-0282-3, PubMed:
31620646
Woolgar, A., Parr, A., Cusack, R., Thompson, R., Nimmo-Smith, I.,
Torralva, T., Roca, M, Antoun, N., Manes, F., & Duncan, J.
(2010). Fluid intelligence loss linked to restricted regions of dam-
age within frontal and parietal cortex. Proceedings of the
National Academy of Sciences, 107(33), 14899–14902. https://
doi.org/10.1073/pnas.1007928107, PubMed: 20679241
Zempleni, M.-Z., Renken, R., Hoeks, J. C. J., Hoogduin, J. M., &
Stowe, L. A. (2007). Semantic ambiguity processing in sentence
context: Evidence from event-related fMRI. NeuroImage, 34(3),
1270–1279. https://doi.org/10.1016/j.neuroimage.2006.09.048,
PubMed: 17142061
Zhang, Y., Frassinelli, D., Tuomainen, J., Skipper, J. I., & Vigliocco,
G. (2021). More than words: Word predictability, prosody,
gesture and mouth movements in natural language comprehen-
sion. Proceedings of the Royal Society B: Biological Sciences,
288(1955), 20210500. https://doi.org/10.1098/rspb.2021.0500,
PubMed: 34284631
Neurobiology of Language
698
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
6
6
5
2
0
7
2
3
4
2
n
o
_
a
_
0
0
0
8
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3