RESEARCH ARTICLE
Individual Differences in Indirect Speech Act
Processing Found Outside the Language Network
Katarina Bendtz1
, Sarah Ericsson1
Jana Bašnáková2,3, and Julia Uddén1,4
, Josephine Schneider1
, Julia Borg1
,
1Department of Psychology, Stockholm University, Sweden
2Donders Centre for Cognitive Neuroimaging, Nijmegen, The Netherlands
3Institute of Experimental Psychology, Centre of Social and Psychological Sciences SAS, Slovakia
4Department of Linguistics, Stockholm University, Sweden
Keywords: pragmatics, communication, fMRI, Indirect speech acts, lingua, theory of mind
ABSTRACT
Face-to-face communication requires skills that go beyond core language abilities. In
dialogue, we routinely make inferences beyond the literal meaning of utterances and
distinguish between different speech acts based on, per esempio., contextual cues. È, Tuttavia, non
known whether such communicative skills potentially overlap with core language skills or
other capacities, such as theory of mind (ToM). In this functional magnetic resonance imaging
(fMRI) study we investigate these questions by capitalizing on individual variation in pragmatic
skills in the general population. Based on behavioral data from 199 participants, we selected
participants with higher vs. lower pragmatic skills for the fMRI study (N = 57). In the scanner,
participants listened to dialogues including a direct or an indirect target utterance. IL
paradigm allowed participants at the whole group level to (passively) distinguish indirect from
direct speech acts, as evidenced by a robust activity difference between these speech acts in
an extended language network including ToM areas. Individual differences in pragmatic skills
modulated activation in two additional regions outside the core language regions (one cluster
in the left lateral parietal cortex and intraparietal sulcus and one in the precuneus). IL
behavioral results indicate segregation of pragmatic skill from core language and ToM. In
conclusion, contextualized and multimodal communication requires a set of interrelated
pragmatic processes that are neurocognitively segregated: (1) from core language and
(2) partly from ToM.
INTRODUCTION
Dialogue constitutes a fundamental form of human communication (Levinson, 2006). In
accordance with this suggestion, being able to communicate face-to-face with another person
may require skills that go beyond “core” language skills, such as lexical retrieval or syntactic
composition. Communication in real-world settings places specific requirements on the inter-
locutors, Per esempio, the ability to go beyond the literal meaning of utterances based on con-
textual cues to understand the speech acts forming the basis of conversational turns, or to tailor
each utterance to the perspective or knowledge of the addressee. There is an ongoing debate on
the nature of these so-called pragmatic skills and the extent to which they overlap with different
core language skills or other capacities, such as executive functions or theory of mind (ToM)
skills (Vedere, per esempio., Matthews et al., 2018, for a developmental approach). Behavioral investigations
a n o p e n a c c e s s
j o u r n a l
Citation: Bendtz, K., Ericsson, S.,
Schneider, J., Borg, J., Bašnáková, J.,
& Udde(cid:1)N, J. (2022). Individual
differences in indirect speech act
processing found outside the language
rete. Neurobiology of Language,
3(2), 287–317. https://doi.org/10.1162
/nol_a_00066
DOI:
https://doi.org/10.1162/nol_a_00066
Supporting Information:
https://doi.org/10.1162/nol_a_00066
Received: 22 Giugno 2021
Accepted: 5 Gennaio 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Julia Uddén
Julia.udden@psychology.su.se
Handling Editor:
Sonja A. Kotz
Copyright: © 2022
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Theory of speech acts:
A perspective on utterances as
conveying not only literal
information but also an action,
such as a request.
Pragmatics:
Communication beyond what is
given in the literal message.
Indirect speech act:
A speech act conveying information
that is not explicitly given literally,
but is nonetheless expressed. “It
would be nice with some coffee” is
an example of an indirect request.
of conversational implicature comprehension suggest that the pragmatic skills necessary for
interpreting at least some aspects of dialogue are relatively independent of structural language
skills, per esempio., vocabulary and syntax (Wilson & Bishop, 2019).
Neurocognitive Dissociation of Pragmatics from Core Language, ToM, and Cognitive Control Functions
In questo articolo, we adopt a neuroscience approach to contribute an additional perspective on
the complex knowledge domain of pragmatics and its representation in the human brain. Questo
approach enables us to investigate whether there are specific neural signatures of pragmatic
skills, or whether these skills draw on general cognitive skills, as would be indicated by mod-
ulation of activity in three other networks: the left-lateralized perisylvian language network,
the ToM network, and the multiple demand (MD) network subserving executive functions.
As our primary test bed, we chose indirect speech acts (ISAs), since interpreting their commu-
nicative meaning requires fully fledged context-based pragmatic inferencing.
Our approach capitalizes on the fact that even within the neurotypical population, there is
substantial variability in pragmatic skills. By combining neuroimaging techniques with behav-
ioral measurements of this variability, we investigate several novel questions. Primo, we ask
whether, Dove, and how individual pragmatic variability will manifest itself in neural activity
during pragmatic inferencing. More specifically, we wonder where this variability will be man-
ifested in relation to the three networks specified above. Secondo, we ask to what extent indi-
vidual behavioral and neural pragmatic variability can be explained away by variability in
core language skills, ToM skills, and executive functions. If it cannot, this would be a sign
of neurocognitive segregation of pragmatics from the other aspects of cognition. In other
parole, such a segregation would mean that neither core language processes, nor core lan-
guage processes together with ToM, nor these two domains together with cognitive control
processes suffice to fully characterize pragmatic processing. A theoretical reason that we study
these three cognitive skills in relation to pragmatics is that they can be easily argued to influ-
ence or be influenced by pragmatics during the development of language and communication.
Infatti, these three skills have received most attention in relation to pragmatic skill in devel-
opmental research (Matthews et al., 2018).
The Indirect Speech Acts fMRI Experiment – Dissociations from Core Language
Interpreting the communicative meaning of ISAs requires pragmatic inferences about what
the speakers truly meant. Così, in order to measure individual differences in pragmatic pro-
cessing in the functional magnetic resonance imaging (fMRI) scanner, we used an established
neurocognitive paradigm on ISA processing. In this paradigm, called the indirect speech acts
experiment, participants listened to a dialogue including an introductory context and a target
utterance with literal and prosodic cues. In several previous studies, the paradigm has
allowed most participants to (passively) discern indirect from direct speech acts, as evidenced
by robust activity differences between these two speech acts in the core language network,
but crucially also in an extended language network including ToM/mentalizing areas in young
adults (Bašnáková et al., 2014; van Ackeren et al., 2016) as well as adolescents (Asaridou et al.,
2019). The effect has been stable across written and auditory versions of the experiment and is
also present when gestures provide the target information expressing the direct or indirect
speech act (Bašnáková, 2019). Two studies have in addition found areas beyond core language
areas, including mentalizing areas, in related pragmatic contrasts: Egorova et al. (2016) con-
trasted two different direct speech acts, and Hellbernd and Sammler (2016, 2018), where par-
ticipants categorized speech acts based on prosody.These existing ISA results thus already
Neurobiology of Language
288
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
provide neural evidence that pragmatic processes cannot be reduced to core language pro-
cesses. Inoltre, if we compare areas reported in these studies to the neurosynth mask (cioè.,
a meta-analytical statistical activation map associated with a keyword) based on the term
language comprehension, we see overlap with the ISA literature in both medial frontal and
the bilateral temporoparietal junction (TPJ) regions, which are also not considered part of the
core language network. Inoltre, the typical left-hemisphere dominance of the perisylvian
language network is absent, in favor of a more bilaterally balanced (Asaridou et al., 2019;
Egorova et al., 2016; Hellbrend & Sammler, 2018) or a right dominant activation pattern
(Bašnáková, 2019). We expected not only that we would replicate the previously found areas
but also that individual variability across groups would be manifested in the vicinity
(localization-wise) of these areas, although the exact location was more difficult to predict, COME
the study is the first investigating individual variability in higher-level pragmatic inferencing.
We use the term language network to refer to the cortical areas recruited for several inter-
related processes that at the same time are partly separable from a neurocognitive point of
view. Depending on the perspective taken, core language processing can, for instance, be
considered as separable lexical, combinatorial, phonological, syntactic, and semantic pro-
cesses, as well as comprehension and production processes (Hagoort, 2017). The included
areas (inferior frontal, anterior temporal, and posterior temporal cortices, in particular from
the middle temporal gyrus (MTG) and up, as well as the angular gyrus) are highly character-
istic, often observed bilaterally but with left dominance (although lateralization depends on
the process studied). The network is to some extent modular and distinguishable as a resting
state network (Paunov et al., 2019), yet at the same time it is possible to divide it into, for
esempio, dorsal vs. ventral pathways (Friederici, 2012; Hickok & Poeppel, 2004).
It should also be noted that some areas in the language network (per esempio., the inferior frontal
gyrus (IFG) and the posterior superior temporal cortices) are commonly observed in other
compiti, possibly also sometimes in ToM related tasks (Schurz et al., 2014, 2015). Using factor
analysis Wilson and Bishop (2019) showed that a two-factor model, using pragmatic skill and
core language skill as two separate factors, captures variation in young healthy adults better
than a one-factor model, although the two factors are robustly related. Tuttavia, in a large-
scale meta-analysis across lifespan, figurative language use has been shown to be highly
related to core language skills in autism spectrum disorders (ASD; Kalandadze et al., 2018).
We wanted to test the extent to which the pragmatic skills and processes we measured
depended on individual differences in, Per esempio, reading speed, vocabulary, word recog-
nition, speed of lexical access, or cultural literacy (see a description of used tasks in the sec-
zione: Behavioral tests of language ability). Accordingly, participants were given a series of tests
measuring their formal language abilities: nonpragmatic or core linguistic skills as measured
by tests for print exposure/reading skill (the author recognition test (AR); Moore & Gordon,
2015), lexical access (the lexical decision task (LDT); Holmer et al., 2016), and vocabulary
(from the Vocabulary, Swedish Scholastic Aptitude Test (vSweSAT); Cliffordson, 2004); Vedere
the section: Behavioral tests of language ability. Our expectation was that individual variance
in core language processes would neither suffice alone to account for variance in pragmatics,
nor suffice when combined with variation in ToM processes (see a description of the used ToM
task in the section: Low/high pragmatic ability groups). We now turn to discuss the latter.
No Unitary Theory of Mind Skill
Based on theoretical predictions (Grice, 1975), understanding what speakers really mean is
primarily achieved through arriving at the underlying intentions behind specific utterances.
Neurobiology of Language
289
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Under this view, widely accepted in the field of pragmatics, communicative meaning interpre-
tation should fall under the scope of ToM processing, as intentions are unobservable mental
stati (similar to beliefs and desires). Tuttavia, firstly, it is now clear that ToM is not well-
defined, in particular not as one domain-general function. Recent meta-analyses indicate that
different categories of tasks result in different activation patterns (Schurz et al., 2014, 2015),
even though there are core hubs that span across all types of tasks, Per esempio, the hallmark
task of false-belief reasoning (bilateral TPJ and medial prefrontal cortex (mPFC); Schurz et al.,
2014). Along these lines, Enrici et al. (2019) suggested that communicative intention process-
ing should be considered a fundamental cognitive component of a reformulated ToM system
(for further details see Schaafsma et al., 2015). Secondly, many pragmatic phenomena may not
require access to the mental state of the speaker in the first place—see, Per esempio, I’ve eaten
breakfast meaning “I’ve eaten breakfast today” (Garrett & Harnish, 2007) and It’s cold in here
as an indirect request (Kissine, 2016)—suggesting the existence of pragmatic processes distin-
guished from mentalizing. Thirdly, we have reasons to question evidence from developmental
studies and clinical populations showing covariation between ToM and pragmatic skills
(Domanesci & Bambini, 2020). As Bosco and colleagues (2018) argue, the extent of the cor-
relations between pragmatic phenomena and ToM is probably inflated because tests used to
assess ToM are often, Infatti, pragmatic tasks; Per esempio, the strange stories test used to
assess advanced ToM reasoning instead probes the implicit and explicit understanding of
figurative language and irony (Bosco et al., 2018). Così, it is generally not helpful to equate
pragmatic processing skills with ToM a priori. We need empirical studies where a reduction
of pragmatic processes to ToM is not assumed. Designs should instead allow for a distinction
to be made between the processes. The extent to which segregation is possible should then
be left as an empirical question. Including measures of core language skills in the same
experiment is also crucial.
If pragmatics is indeed not simply reducible to ToM plus core language skills, this also
makes clear predictions for patterns of individual variation in the general population. Behav-
iorally, these predictions include small or absent correlations of pragmatic skills vs. core lan-
guage and ToM skills. This constitutes the basic rationale of the empirical tests in the current
study. We measured 199 participants from the young adult general population using a novel
behavioral battery. Based on a subset of these tests measuring two different pragmatic skills
(see descriptions in the section: Behavioral tests of pragmatic ability), we selected two groups
of participants with good (top 50% performers, a high scoring (HS) group) vs. poor (lowest
50% performers, a low scoring (LS) group) communicative skill for the fMRI study (N = 57,
Dove 29 participants had poor communicative skills). The pragmatic skills tested for this pre-
selection included both production and comprehension. The participants’ mentalizing (O
ToM) ability was also assessed using a nonverbal test, the reading the mind in the eyes test
(RMET; Baron-Cohen et al., 2001; see the section: Non-verbal behavioral ToM).
Cognitive Control Functions and the MD Network
Cognitive control functions (CCFs) allow us to plan, controllo, and regulate complex higher-
order tasks in a flexible manner. For instance, the transient representation and manipulation
of task-relevant information in working memory is needed for proper cognitive control. (Note
that for our purposes here, CCFs will be used interchangeably with executive functions.) Nel
current study, we ask whether proficiency in pragmatic processes is dissociable from CCFs. Noi
start tackling this question mainly by studying individual variation in behavioral pragmatic
skills in relation to CCFs, more specifically variance in a complex operation span task (OSpan;
Foster et al., 2015). Inhibitory control and working memory have been shown to be associated
Neurobiology of Language
290
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
with performance on some pragmatic tasks (Matthews et al., 2018), where for instance Ryskin
et al. (2014) have used the Ospan task specifically. The OSpan task is further described in the
section: Cognitive control – Operation span (OSpan).
As mentioned above, there is also a neural perspective on this matter. One of the questions
in the current study is where, in terms of large-scale brain networks, individual variation in the
ISA paradigm presented above will manifest itself. The three most relevant large-scale brain
networks we consider are the core language network, the ToM network, and the MD net-
lavoro (Duncan, 2010). These networks were described in detail in a series of studies from the
Fedorenko lab (Blank et al., 2014; Paunov et al., 2019), including internal replication
esperimenti. The MD network specifically has been emerging from fMRI data as a relevant
network across varied tasks (per esempio., cognitive control tasks including working memory, atten-
tional control, or general intelligence) but how to best describe its function is less clear
(Duncan, 2010). A domain-general function is, Tuttavia, often assumed, and hence the net-
work is also sometimes referred to as the task-positive network, or the frontoparietal control
rete (FPCN; Chein et al., 2011; Vincent et al., 2008). In previous research it has been
shown that the OSpan task we use indeed activates the intended MD network (Faraco
et al., 2011). There have also been indications that individuals with higher OSpan perfor-
mance activate this network more during the task (Faraco et al., 2011; Osaka et al., 2003).
Anatomically, three reviews or meta-analyses converge on the MD network as consisting of
areas in bilateral dorsolateral prefrontral cortex (per esempio., BA 9/46), bilateral parietal areas (per esempio.,
BA 7 ), and dorsal medial prefrontal/anterior cingulate (per esempio., BA 32), and/or medial
presupplementary/supplementary motor cortex (Chein et al., 2011; Duncan, 2010; Vincent
et al., 2008). These studies also agree on using the label cognitive control (or alternatively
executive functions, which is largely used interchangeably in the literature) when discussing
the cognitive function of this network. As we thought there was a substantial probability that
CCFs would influence the ISA task and since this work focuses on individual variation in
communicative skills, we decided to match the LS and HS group on their CCF performance.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Approach Summary
Our approach can be summarized with six points. (1) Two sets of experiments were con-
ducted, an fMRI experiment and a preceding behavioral experiment. (2) We used the estab-
lished ISA paradigm in the fMRI experiment to compare two conditions: indirect vs. direct
speech acts. (3) We compared activity between the HS and LS groups that were formed
based on the behavioral experiment of a larger sample (N = 199). We used two behavioral
pragmatic tasks in this behavioral experiment, while at the same time controlling for what
we perceived as the general cognitive skill most likely to actually influence the ISA variabil-
ità, which was CCF. (4) By design, the regions showing a Group × Condition interaction are
assumed to be involved in pragmatic inference. (5) To test for neurocognitive segregation,
regions in (4) are compared with neurosynth networks for core language, ToM, and CCF. (6)
In a further test of segregation, individual activity in regions in (4) that were further tested in
(5) and do not show overlap with neurosynth networks is correlated with the following skills
measured in the behavioral battery: nonpragmatic (“core”) language – vocabulary (vSweSat)
and lexical access (LDT); ToM – nonverbal test of mentalizing ability (RMET); and CCF–
OSpan. Inoltre, all behavioral tasks used are correlated with each other, testing whether
pragmatic skills segregate from core language, ToM, and CCF at the cognitive level. Noi
consider absences of overlap in (5) and nonsignificant correlations in (6) as evidence of
neurocognitive segregation.
Neurobiology of Language
291
Pragmatic individual differences using fMRI
Audience design:
The adjustment of utterances to suit
the needs of interlocutors.
MATERIALS AND METHODS
Participants
Sixty participants (age 18–36, 28 males) were recorded in the fMRI experiment. No participant
had any history of neurological impairment, brain surgery, or ASD/Asperger diagnosis. Partic-
ipants with a history of language impairment were generally excluded prior to participation.
They all gave informed consent and received 350 in SEK [Swedish krona] for their participa-
zione. The study was approved by the Swedish Regional Ethical Review Authority in Stockholm.
The participants of the fMRI experiment were selected from a larger group who participated
in a preceding behavioral experiment (N = 199; 99 males; average age 28.7 (males) E
29.3 (females)). Participants were invited into the fMRI experiment based on their results
in two tests in the behavioral experiment battery: audience design (AD) and prosodic com-
prehension of request for response (PC-RR; see the section: Behavioral Tests of ToM and
Cognitive Control. For more information on how we formed these groups, see the section:
fMRI Task: The indirect speech acts experiment.) Further details on the participants are given
in the supplementary material and Table S1 (Supporting Information can be found at https://
doi.org/10.1162/nol_a_00066).
Behavioral Tests of Pragmatic Ability
Prosodic comprehension of requests for response
We wanted to cover a wide range of pragmatic skills, crossing the division of production and
comprehension without neglecting the multimodal aspects of communicative signals. For the
comprehension test, we settled on testing participants’ ability to identify a speaker’s request
for feedback in a communicative situation based on prosody. Speech prosody is known to be
an important ingredient in spoken language, not only expressing emotions but also potentially
modifying, or even completely altering, the communicative meaning of an utterance (per esempio.,
irony). Hellbernd and Sammler (2016) found that prosodic patterns of different speech acts, come
as criticism, suggestion, or wish, were dissociable in terms of prosodic cues to a high degree and
also consistent across speakers and utterances. We chose to focus on the prosodic modulation of
utterances in the process of establishing meaning (see meaning establishment in Clark, 1994).
For this purpose, we developed the prosodic comprehension of request for response (PC-RR)
test. In our test, participants judged whether an auditory utterance (per esempio., I don’t think your com-
puter has the right port for this charger) is meant as a simple statement (actors used few pauses,
steadily falling intonation) or as soliciting feedback from the listener (actors paused between
has the right and port, variable intonation shifting between falling and rising). Sentence mate-
rials were constructed so that the same literal material could be sensibly used in both condi-
zioni. We recorded 12 sentences where speakers (two male, two female) had been asked to
produce prosody indicating a request for response from the listener, a request for response
trial, or a statement trial. The sentences were presented to the participants whose task was
to determine by button-press whether the sentence was a request for response or a statement
trial. Each participant listened to all 12 sentences, half in the request for response condition,
and half as statement sentences (order and mapping between condition and sentence were
randomized). The test was implemented using PsychoPy (Peirce et al., 2019).
Production: Advanced audience design
While there are many psycho/neurolinguistic studies of production processes, they have
always been outnumbered by comprehension studies, largely for methodological reasons
Neurobiology of Language
292
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
(per esempio., the issue of combining elicitation in production with experimental manipulation and
controllo, as well as the issue of movement artifacts in neuroimaging). Perhaps the most com-
mon task that can be described as a pragmatic production task is the director’s task, Quale
builds on an important insight from conversation analysis (Sacks & Schegloff, 1979)—inter-
locutors have to tailor each contribution to the specific needs of the listener by means of
so-called audience design (Campana, 1984; Clark et al., 1983), alternatively called recipient
progetto (Sacks & Schegloff, 1979).
In the director’s task, the participant produces labels for visually presented objects based on
whether the objects are uniquely in their own field of view or seen mutually with the listener
(Krauss & Glucksberg, 1977; Ryskin et al., 2015; Wardlow, 2013). The standard director’s task
ha, Tuttavia, been criticized for possibly testing selective attention rather than pragmatic
inferencing, as the participant may make use of the strategy to pay attention only to objects
that are not occluded. This attentional strategy might for instance be implemented using
shallow visual attentional mechanisms, as occluded/non-occluded objects have particular
background colors (Rubio-Fernández, 2017). Another study showed similar performance
when participants take the perspective of a camera instead of another person, which questions
task validity when studying pragmatics (Santiesteban et al., 2015). To address these concerns,
we designed a version of the director’s task where the speaker instead needs to consider the
age, genere, and cultural background of the listener. In other words, the speaker needs to tailor
their utterance to the specific addressee, based on their inferences about what the addressee
knows. Validity is improved since it is the addressee’s knowledge that must be taken into
account, just as in naturalistic communicative situations. The more participants take their
knowledge of the listener into account, the higher their performance on the task. We thus
consider this task a state-of-the-art task for measuring valid individual differences in AD, Quale
was our goal in the context of the fMRI study. More generally, this new task probes how inter-
locutor characteristics (per esempio., age) influence conversational processes.
Participants were instructed to describe an object that could not be assumed to be known
in the unknown condition (test condition), or that could be assumed to be known in the
known condition (control condition) to a fictitious addressee. An example is given in
Tavolo 1. A successful way of describing an object to the addressee in the unknown condition
is to describe it in other, simpler words. Successfully paraphrasing the object in the test trial
resulted in 1 point, while not paraphrasing (using a label that could not have been known to
the addressee) gave 0 points.
Each trial consisted of an image of a bookshelf where five objects were placed along with a
picture of the addressee specific to that trial. The target object was the object the participant had
to describe. The competitor object (Vedi la tabella 1) belonged to the same general category as the
Tavolo 1.
Examples of four trials in the audience design task for two different addressees
Addressee
6-year-old child
91-year-old person
Target
Corset
Drone
Test trial: Unknown condition
Example of paraphrasing
The beige thing that looks like a dress
Competitor
Bikini
Control trial: Known condition
Target
Banana
The machine with four propellers
Helicopter
Recorder
Note. For the test trials, the competitor object and an example of a paraphrase indicating that the participant has succeeded in taking the addressee’s perspective
are given. The two objects are shown in Figure S2. In this example, the object to be named was a corset (the competitor was a bikini), and when the addressee
was a small child, an appropriate label would be the beige thing that looks like a dress, whereas a pragmatically inappropriate label would be undergarment
since a child probably does not know what a corset’s function is, and bikini also fits this label.
Neurobiology of Language
293
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
target object to make the participant avoid a more general description, which could be taken as a
paraphrase. Three filler objects were also on the bookshelf. Each participant did 10 test trials and
10 control trials. The test was implemented using PsychoPy (Peirce et al., 2019).
Behavioral Tests of ToM and Cognitive Control
Nonverbal behavioral ToM
An overall goal of the study was to delineate pragmatic processes that are potentially partly
independent of more basic nonverbal ToM. We measured ToM from a standard nonverbal test
of mentalizing ability: RMET (Baron-Cohen et al., 2001). Based solely on pictures of the eye
region of human faces, participants had to attribute complex emotions to the pair of eyes by
choosing from four alternatives. The test is independent of language abilities but related to the
severity of ASD symptoms (Senju et al., 2002). We wanted to minimize the risk of a more trivial
association between ToM and pragmatic ability, which might have resulted from some RMET
items requiring the participants to resort to verbal strategies. We thus eliminated items where
any communicative intention (cioè., sarcasm or joking) was cued. Out of the remaining trials we
chose 12 in a way so as to maximize the variation of emotions and the difficulty of the test
(using the correctness rates reported in Baron-Cohen et al., 2001). The test was implemented
using PsychoPy (Peirce et al., 2019). Even though the RMET was originally intended as a test of
ToM and has been extensively used as such, we note that there is now research rather linking it
to the ability to recognize complex emotions (Oakley et al., 2016), somewhat independent of
intention recognition. While emotions can be considered to be mental states and thus be part
of ToM, Oakley et al. (2016) use Alexithymia and ASD patients to show that they are poten-
tially segregated from domain-general ToM. Therefore, we refer to this task as a domain-
general ToM/complex emotion recognition task in the discussion. It could thus potentially
be viewed as reflecting “affective” rather than “cognitive” ToM (Shamay-Tsoory & Aharon-
Peretz, 2007). Note, Tuttavia, that there are known issues in measuring individual differences
in ToM (Conway et al., 2019). This suggests that an alternative formulation of this part of the
design is that we are testing the uniqueness of pragmatic processes relative to complex emo-
tion recognition (or again, affective ToM). Tuttavia, similar restrictive generalizability applies
to other ToM tests.
Cognitive control – Operation span
The operation span (OSpan) test (Foster et al., 2015) has been designed to probe CCF, anche
as working memory, as a part of CCF. IL 199 participants in the behavioral experiment mem-
orized a sequence of letters while resisting frequent distractors, as a test of their CCF. CCF,
especially inhibitory control and working memory, are associated with performance on some
pragmatic tasks, at least during development (Matthews et al., 2018).
The mixed adult literature includes Roßnagel (2000, 2004), who found that AD was nega-
tively affected by higher cognitive load as induced by a dual task situation. Initial correlations
between pragmatic processes and tasks measuring individual differences in general executive
function and nonverbal IQ, such as the Stroop task, Raven’s matrices, and simple working
memory span tasks, have been observed, while replication attempts have failed (Brown-
Schmidt, 2009; Brown-Schmidt & Fraundorf, 2015; Ryskin et al., 2014). Ryskin et al. (2014)
found a significant relation between a perspective-taking task and the OSpan task, providing at
least one correlational result yet to be replicated or not.
Così, Ancora, as a measure of cognitive control, participants completed the OSpan test
(Foster et al., 2015) as implemented in E-prime (Engle lab; https://englelab.gatech.edu
Neurobiology of Language
294
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
/taskdownloads.html). In this test, participants are presented with a sequence of letters, one by
one, and then asked to recall the sequence. Between each letter presentation, the participants
must complete a distractor task in the form of a simple calculus problem (per esempio., 3 * 4 + 1). For
more details, see the supplementary material.
Behavioral Tests of Language Ability
As indicated in the introduction, previous research has segregated pragmatic comprehension
processes from core language ability, in the form of vocabulary and grammar. The partici-
pants in the behavioral experiment were assessed on core language tasks, and the fMRI partic-
ipants on two additional tasks. These structural language measures were the AR test, Quale
tests print exposure and predicts reading skill (Moore & Gordon, 2015) and cultural literacy
(Stanovich & West, 1989; West et al., 1993); a vocabulary test taken from the Vocabulary,
Swedish Scholastic Aptitude Test (vSweSAT of 2016; Cliffordson, 2004) an LDT (Holmer et al.,
2016) measuring low level reading and lexical access processes. The vSweSAT and LDT were
administered for the fMRI participants only. The vSweSAT consists of 10 words where the correct
synonym is to be selected from five alternatives. There was no time limit to complete the test.
What we refer to here as core language skills or structural language skills are actually a set of often
interrelated subskills, commonly assessed and evaluated together in the clinical setting (Vedere
further section S1.4.1 in the Supporting Information).
Lexical decision task
In the LDT (Holmer et al., 2016), the participants were presented with 46 three-letter com-
binations (two consonants and one vowel) within a 5 s period, and were instructed to determine
as fast as possible whether the combination was a real word (half of the trials) or not; order was
randomized. Half of the words that were not real words were pseudowords, questo è, they were
orthographically legal but non-lexicalized items. The other half were non-words, questo è, ortho-
graphically illegal items. For each participant, the reaction times for the correctly answered tri-
als were averaged and multiplied by −1 to yield the final score where performance increases
with score value.
Author recognition task
In the AR task, the participants were presented with a list of names in alphabetical order, half of
which were real authors and half of which were not. The task was to identify which names were
authors. A correctly selected author yielded 1 point, an incorrectly selected author yielded −1,
and all other cases yielded zero. There was no time limit to complete the test. We adapted the AR
task as given in Moore and Gordon (2015) to a Swedish reading audience and slightly revised the
task by increasing the ratio of better known to lesser known authors in order to increase sensi-
tivity to individual variation among participants with poor performance. This was advised by the
authors in Moore and Gordon (2015) based on an item response analysis of their data. For more
details, see the Supporting Information.
Low/High Pragmatic Ability Groups
Our aim was to study individual variation in pragmatic ability in general, rather than perfor-
mance on specific tests. We thus combined the different pragmatic tests of relevance to the
Neurobiology of Language
295
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Figura 1. PC-RR and AD test scores for all participants. The participants in the high (low) score
group are shown as red squares (blue triangles) and participated in the fMRI study. Participants who
scored in the lower 50% on both the AD and PC-RR tests were placed in the LS group and those
who scored in the higher 50% on both tests were placed in the HS group. Both of these groups
include a wide range of scores within these limits, not only very low/high ones.
fMRI paradigm, PC-RR and AD, and constructed two pools of participants, one with low and
one with high pragmatic ability. The low (high) ability pool was formed out of those with
scores lower (higher) than the mean on both the PC-RR and the AD tasks; Guarda la figura 1. Par-
ticipants in the fMRI experiment were then invited from these two pools, forming one low and
one high ability participant group, the LS and HS groups. There were 41 participants in each
pool, and participants were invited randomly until 30 in each group had been measured. In
order to match the groups on cognitive control or complex working memory ability, as indi-
cated by performance on the OSpan test, some participants from the pools were omitted.
Due to some further exclusion after measurements, the final LS (N = 29, 13 males) E
HS (N = 28, 15 males) groups were somewhat smaller. One participant was excluded from
the experiment after measurements due to excessive head movements (>4 mm) at several
volumes. Another participant was excluded due to <80% correctness rate on the compliance
questions (see below). Finally, one participant was excluded due to technical failure. The
average scores and standard deviations of the PC-RR and AD tests across groups along with
two-sample t tests of AD and PC-RR across groups are given in Table 2. While there were no
significant differences between the two participant groups in terms of age, RMET, OSpan, or
LDT, the HS group had significantly higher AR and vSweSAT scores (see Table 2).
fMRI Task: The Indirect Speech Acts Experiment
Stimulus material
The stimulus material in the ISA task consisted of 78 experimental trials. These were short
recorded dialogues: a question and a reply, preceded by a short context depicting the setting
of the dialogue. There were two experimental conditions, indirect and direct; see Table 3. In
Neurobiology of Language
296
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
n
o
_
a
_
0
0
0
6
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pragmatic individual differences using fMRI
Table 2.
Average behavioral test scores, age, and gender for the two communicative skill groups
PC-RR*
AD*
RMET
AR*
LDT
vSweSAT*
OSpan
Age
Gender
High score group
0.97 ± 0.04
Low score group
0.74 ± 0.12
0.70 ± 0.13
0.83 ± 0.12
0.14 ± 0.07
1,019 ± 190
0.86 ± 0.13
0.72 ± 0.17
29.97 ± 4.04
15 males
0.25 ± 0.18
0.82 ± 0.15
0.09 ± 0.05
1,026 ± 287
0.76 ± 0.20
0.70 ± 0.16
29.10 ± 4.97
13 males
Statistical tests
t(34) = 10.11, p < 0.001
t(50) = 10.62, p < 0.001
t(52) = 0.35, p = 0.73
t(53) = 2.58, p = 0.01
t(45) = −0.10, p = 0.92
t(45) = 2.22, p = 0.03
t(55) = 0.55, p = 0.59
t(52) = 0.71, p = 0.48
X(1) = 0.42, p = 0.52
Note. Average test scores and age (± standard deviation) for the two groups in the prosodic comprehension of requests for response (PC-RR) task, audience
design (AD) task, reading the mind in the eyes test (RMET), author recognition (AR) task, lexical decision task (LDT), vocabulary scholastic aptitude test (vSwe-
SAT), and operation span (OSpan) test along with statistical tests ( Welch’s two-sample t test) of difference of these quantities between the groups. In the final row
we report a statistical test (Pearson’s chi-square test with Yates’s continuity correction) of the difference in gender between the groups. An asterisk marks a
significant difference in test scores between the groups.
Condition
Direct
Indirect
Compliance
question
Filler trial
Table 3.
Example of an ISA test trial in the direct and indirect version and a following compliance question
Context
Magnus and Emilia are old friends. They are discussing how hard
it is to find restaurants which both you and your partner fancy.
Emilia asks Magnus:
Question
Why doesn’t your
girlfriend like
Japanese food?
Magnus and Emilia are old friends. They are talking about the last
time Magnus visited Emilia in her student dormitory. Emilia asks
Magnus:
Did your girlfriend like
my vegan noodles?
Answer
She is not so
used to Asian
seasoning.
Was there someone who was not so used to Asian seasoning?
Benny and Ellinor are doing their laundry. Benny went out walking
the dog but is now back. He asks Ellinor, who was the last to
visit the laundry room.
Is the washing
machine done?
Yes, just put
everything in
the dryer.
the direct condition, the reply sentence was simply an informative answer to the question. In
other words, the speaker meaning (see Introduction) coincided with the literal meaning. In the
indirect condition, the literal reading of the reply is often or partly irrelevant (Grice, 1975) to
the question. In these trials, the speaker meaning in the reply was different from, or included
more meaning than, the literal meaning. While the literal content of the reply sentence was the
same in both conditions, the context and question sentences varied, making the reply sentence
direct or indirect (see Table 3). This was to ensure that any differences in neural activation
between indirect and direct conditions observed during the reply would be a consequence
of inferring the speaker meaning, rather than decoding the literal reply. To interpret the indirect
replies, the participants thus had to make a pragmatic inference. In addition, these replies were
affective in the sense that they “saved the face” of the addressee. According to face theory
Neurobiology of Language
297
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
n
o
_
a
_
0
0
0
6
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pragmatic individual differences using fMRI
(Goffman, 1967), the face of a speaker can be defined as the “positive social value” that a
speaker holds in relation to others in a communicative situation. In the indirect condition,
the speakers saved the face by paraphrasing a message that would be face threatening if given
in its explicit form (Brown & Levinson, 1987).
We focused on face saving speech acts as opposed to purely informative indirect replies
based on previous research on ISAs in different social contexts (Holtgraves, 1998). Holtgraves
found that for replies that seem to diverge from relevance (Grice, 1975), i.e., replies that do not
explicitly (literally) answer the question, a face saving context made them more likely to be
interpreted as ISAs. Half of the trials (39) delivered a critical opinion from the speaker, referred
to as Criticism trials. The other half delivered a bad message or critical opinion from another
person not present in the dialogue setting, referred to as Bad message trials. These two kinds of
sentences were introduced to provide varied, yet controlled trial types. We did not have any
specific hypothesis on how trial type would affect processing, and analysis across types was
beyond the scope of the study.
We created 14 filler trials formatted like the other trials: a context, a question, and a reply.
The replies were always direct, but in contrast to the replies in the indirect and direct trials, the
filler replies were initiated by a yes or no. The filler trials were presented to make the pattern of
indirect and direct answers less apparent. In order to make sure participants paid attention to
the stimuli, they answered 12 compliance yes/no questions about the content of the previous
trial; see Table 3. The questions regarded practical information that could be given in any part
of the trial (context, question, or answer).
All participants answered with an accuracy of >83% on the compliance questions and had
at most one error per run. There was no significant difference (T(59) = −0.76, p = .45) in the
accuracy between the groups. No participants passed the far outlier threshold in terms of reac-
tion times defined as the Q3 + 3 × IQR, where Q3 is the 75% percentile and IQR is the inter-
quartile range.
Fifty-eight of the dialogues were revised and translated from Asaridou et al. (2019). We cre-
ated 20 additional dialogues, as well as short contexts for all 78 dialogues. The number of
words in the contexts and the questions was balanced across the two conditions (two-sample
t tests, p > 0.73). Care was taken to make the contexts as similar as possible between the two
conditions of each trial, as well as to use what we perceived as frequent words throughout.
The average durations of the trial components are presented in Table 4. The intertrial intervals
(ITIs) were sampled from a flat distribution between 2 E 4 s with bin size 0.1 S.
fMRI procedure
Participants received scripted oral instructions about the ISA experiment. They were instructed
to pay attention to what the protagonists “really intended to say.” The experiment was
Tavolo 4.
Average durations of trial components [S]
Condition
Indirect
Direct
Context
11.6 ± 2.5
11.6 ± 2.5
Question
2.9 ± 1.0
2.6 ± 1.0
Answer
4.2 ± 1.0
3.8 ± 1.2
Note. Average durations ± standard deviations of context, question, and answer for the two conditions sepa-
rately. No significant differences between the indirect and the direct conditions were found for the context,
question, or answer.
Neurobiology of Language
298
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
performed double-blind, such that neither the experimenters nor the participants knew which
group (low- or high-scoring pragmatic ability) the participant belonged to.
The ISA task, with a total duration of approximately 38 min, was divided into 3 runs. Each
run consisted of 26 unique experimental trials, 1–3 filler trials, and 1–5 compliance questions.
Every participant was presented with 78 experimental trials (half indirect, half direct), 14 filler,
E 12 compliance questions in total. Every experimental trial was presented in its direct vs. its
indirect version to an equal number of participants (but a specific participant only heard any
specific trial in either its direct or indirect version), to control for the exact literal meaning of
each reply, across participants. For more details, see the Supporting Information.
fMRI and T1 data acquisition
The participants were scanned in an interleaved fashion with a Siemens 3T Magnetom Prisma
MRI-scanner, using a 20-channel surface coil. Functional scans were acquired with a repeti-
tion time (TR) Di 2.1 s and echo time (TE) Di 30 ms; each volume consisted of 70 slices, 2.0 mm
thick, con un 1 mm slice gap. The voxel size was 2.2 × 2.2 × 2.0 mm3 and the field of view was
210 mm. The flip angle was 70°. A whole-brain high-resolution structural T1-weighted
magnetization-prepared rapid acquisition with gradient echo (MPRAGE) sequence (using
generalized autocalibrating partially parallel acquisitions (GRAPPA) to accelerate acquisition)
was performed to characterize a participant’s anatomy (TR = 2,300 ms, TE = 2.85 ms, 192 slices
con 1.3 mm isotropic resolution, and field of view = 256 mm).
Analysis of the Behavioral Data
The statistical analyses were performed using R (version 3.6.1; https://cran.r-project.org/bin
/windows/base/old/). We tested for correlations between the AD and the PC-RR tests using a
one-tailed Pearson correlation test. A one-tailed test was used due to the strong expectation
that the two pragmatic tests would be positively rather than negatively correlated. Come il
scores of the behavioral tests were found not to be normally distributed as tested by the
Shapiro-Wilk normality test, a rank based inverse normal (RIN) transformation (Bishara &
Hittner, 2012) was performed on the data before testing the correlations with a Pearson
product-moment correlation test (see further supplemental section S1.5).
fMRI Data Analysis
The data were preprocessed and statistically analyzed with the statistical parametric mapping
software SPM12 (https://www.fil.ion.ucl.ac.uk/spm/; Friston, 2007).
Preprocessing
We first performed motion correction (realignment), then coregistration of the functional
images to the anatomical image, normalization to a standard Montreal Neurological Institute
(MNI) spazio, and spatial smoothing. After realignment, the head movements in x, sì, and z
were checked independently. All participants had a head movement <3 mm, considering
all directions. The normalization was carried out using affine regularization, and a resampling
of the voxels to 2 × mm 4th degree B-spline interpolation. During
the normalization, white grey matter segmentation bias correction were also per-
formed. spatial smoothing 3D isotropic Gaussian kernel of full-width
at half-maximum (FWHM) =8 applied functional data. We included temporal
high-pass filter (cycle cut-off at 128 s), account for various low-frequency effects.
Neurobiology Language
299
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
> 0.05). All tests are two-tailed except for the AD and
PC-RR correlation. No correction for multiple comparisons was applied. We refrain from drawing any conclusions from the Pearson correlation coefficients
themselves, which is why we are not showing them here (see further discussion in the section: Analysis of the behavioral data).
Neurobiology of Language
301
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Tavolo 6.
Activations for contrast indirect vs. direct, interaction between groups, and for both groups
Anatomical region
(UN) Indirect vs. direct interaction between groups (F test)
Left superior parietal lobe/left SMG
Bilateral precuneus
MNI local
maxima
sì
X
−40
8
−48
−66
(B) Indirect vs. direct between groups HS > LS (follow up t test for direction)
Left superior parietal lobule/left SMG
Bilateral precuneus
−40
8
−48
−66
(C) Indirect vs. direct between groups LS > HS (follow up t test for direction)
No significant clusters or voxels
(D) Indirect vs. direct for both groups
Cluster
Voxel
z
Size
pFWE
T / F value
pFWE
46
54
46
54
368
<0.001
399
<0.001
n.s.
n.s.
732
<0.001
t(55) = 5.37
0.040
525
<0.001
n.s.
Right MTG, angular gyrus, temporal pole, and IFG/frontal orbital
50
28
−4
5642
<0.001
t(56) = 9.38
<0.001
cortex
Bilateral superior frontal gyrus
8
50
40
4070
<0.001
t(56) = 8.93
<0.001
Left cerebellum
Left MTG, angular gyrus and IFG/frontal orbital cortex
Bilateral precuneus
(e) Indirect group comparison
No significant clusters or voxels
(f ) Direct group comparison
No significant clusters or voxels
−26
−60
6
−82
−50
−54
−36
26
36
418
0.001
t(56) = 6.90
<0.001
4534
0.001
t(56) = 6.88
<0.001
438
<0.001
n.s.
Note. See the section First level statistical analysis for detailed explanations of the contrasts. The cluster-forming threshold was p = 0.001. Coordinates are given
in MNI space. n.s. stands for nonsignificant.
significantly with the LDT; see Table 5. Finally, the two language tests AR and vSweSAT cor-
related significantly. AR was dropped from further analysis for this reason and because we
were more interested in a test of vocabulary, as it is a central, well-researched core language
ability (it is less clear what exactly AR tests).
fMRI Results
Indirect vs. direct contrast
We report the indirect vs. direct contrast for both HS and LS groups in Table 6 and for each
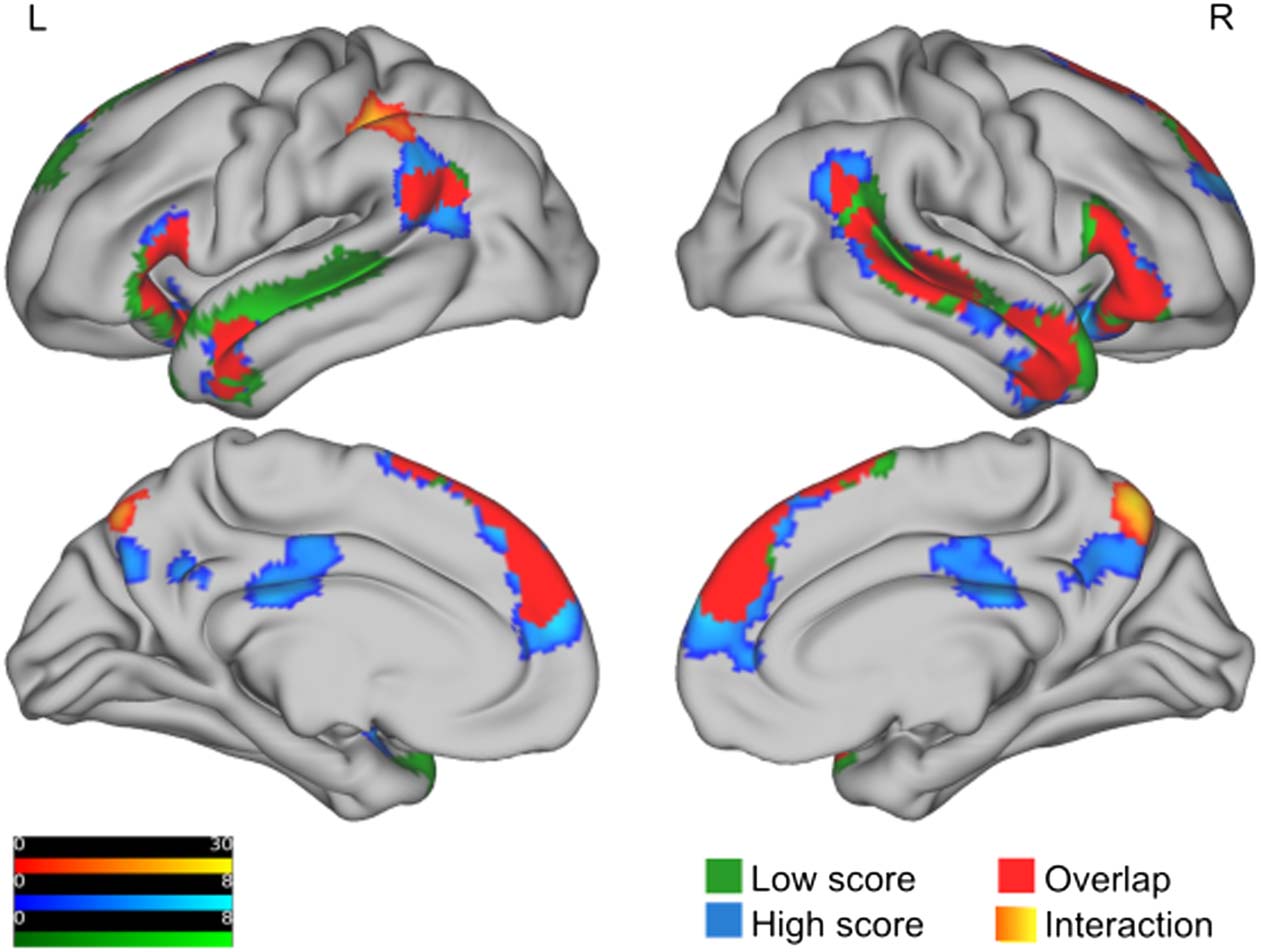
group separately in Table 7. We observed overlapping activity for the HS and LS groups in
bilateral IFG, TPJ, anterior temporal lobe, medial superior frontal gyrus (SFG)/dorsomedial pre-
frontal cortex (dmPFC), and right mid and posterior MTG/superior temporal sulcus (STS; see
Figure 2). In addition, the LS group showed left posterior MTG/STS activity. The HS group acti-
vated a larger portion of the same cortices, ventrally and dorsally in the left TPJ, and
Neurobiology of Language
302
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
n
o
_
a
_
0
0
0
6
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pragmatic individual differences using fMRI
Table 7.
Activations for contrast indirect vs. direct for both groups separately
Anatomical region
(a) Indirect vs. direct for HS group
Right M/STG
Right SFG/anterior paracingulate gyrus
Left angular gyrus/occipital cortex/pSMG
Right frontal orbital cortex/insula
Right IFG/frontal pole
Right temporal pole
Left frontal orbital cortex/insula
Bilateral posterior cingulate
Left cerebellum
Bilateral precuneus
Left temporal pole
(b) Indirect vs. direct for LS group
Right frontal orbital cortex/IFG/temporal pole/STG/MTG
Bilateral SFG
Left frontal orbital cortex/IFG
Left angular gyrus/SMG
MNI local maxima
z
y
x
Cluster
Voxel
Size
pFWE
t / F value
pFWE
54
6
−62
30
48
48
−28
0
−32
−4
−54
48
−6
−50
−60
−24
62
−54
18
32
10
14
−26
−86
−68
6
26
54
26
−56
−12
24
28
−14
−2
−32
−18
24
−34
34
−28
−8
24
−6
16
0.001
0.002
0.009
0.010
0.015
0.044
0.041
1,465
<0.001
t(28) = 7.94
2,969
<0.001
t(28) = 7.65
937
205
669
583
774
375
330
387
218
<0.001
t(28) = 6.85
0.026
t(28) = 6.81
<0.001
t(28) = 6.62
<0.001
t(28) = 6.13
<0.001
t(28) = 6.07
0.001
0.003
0.001
0.020
n.s.
n.s.
n.s.
n.s.
3,339
<0.001
t(27) = 8.69
<0.001
1,920
<0.001
t(27) = 7.37
1,736
<0.001
t(27) = 7.00
0.004
0.009
272
0.003
n.s.
Note. See the section First level statistical analysis for detailed explanations of the contrasts. The cluster-forming threshold was p = 0.001. n.s. stands for
nonsignificant.
ventrally/rostrally in the medial SFG/dmPFC. This group also showed significant activity
(absent in the LS group) in the left cerebellum, bilateral posterior cingulate, and precuneus,
a pattern which resulted in a significant group interaction for only the precuneus cluster. Note
that differences across groups that are evident in this latter kind of analysis are not straightfor-
ward to interpret as they are not statistically tested interactions with groups (for this see instead
the section: Indirect vs. Direct contrast – Interaction between the HS and LS groups). In
Figure 2, we also report some of these analyses for illustrative purposes.
Indirect vs. direct contrast – Interaction between the HS and LS groups
Our main analysis testing for interaction between groups for the indirect vs. direct contrast
with an F test revealed two significant clusters; see Figure 2 and Table 6. One significant clus-
ter was found in the parietal lobe, with parts in the superior part of the posterior as well as the
anterior division of the supramarginal gyrus (SMG). Slightly more medial parts extended into
the superior parietal lobe, where anterior parts connected to the postcentral gyrus and poste-
rior parts connected to the angular gyrus (and anterior and mid-parts of intraparietal sulcus
(IPS); Juelich Histological Atlas (https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/Atlases/Juelich)). This clus-
ter was adjacent to significant activity in the lateral left inferior parietal cluster that was only
Neurobiology of Language
303
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
n
o
_
a
_
0
0
0
6
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pragmatic individual differences using fMRI
Figure 2. Brain activation for the indirect vs. direct contrast, for each group separately as well as
for the interaction between the groups. The figure shows F values for clusters with a cluster-forming
threshold of puncorrected = 0.001 (no extent-level threshold, k = 0). As a multiple comparison correc-
tion method, we used family wise error (FWE) correction at the cluster and peak level. We only
report clusters and voxels with a pFWE < 0.05. All clusters are projected onto a pial surface. The
activation for the interaction is shown in orange to yellow. The activation for the high (low) score
group is shown in blue (green). The regions where the activations in both groups overlap are shown
in solid red and no activation information is given. Note that the range of the scales do not reflect
the exact max (min) value of the unit of activation (which is customary) but are adjusted slightly so
as to include values from all hemispheres.
present for the HS group for the indirect vs. direct contrast, but it was located at the superior
bank of this cluster (see Figure 2). Another significant cluster was found in the lateral parts of
the right precuneus, extending into the left precuneus. A follow-up t test revealed that these
two activations were stronger for the HS group compared to the LS group (see Table 6). Note
that, as we selected the F test for analysis, this analysis was performed to find out the direction
of the effect in the cluster, referred to in the section: Indirect vs. direct contrast – Interaction
between the HS and LS groups, that was found in the F test only (i.e., it is not a parallel but a
subordinate test). A lateralization test of the parietal cluster showed the lateralization not to be
significant: A paired t test of the blood oxygen level-dependent (BOLD) responses from the
indirect vs. direct contrast, between the left parietal cluster and corresponding right regions,
was nonsignificant across all participants (p = 0.34) and in the HS (p = 0.48) and LS (p = 0.35)
groups separately.
Context vs. ITI contrast
Using an F test identical to the one reported above (in the section: Indirect vs. direct contrast –
Interaction between the HS and LS groups), we found no significant group differences (p >
0.88) during listening to dialogue contexts. This p value refers to the smallest FWE-corrected
p value from a pool of p values at both the cluster and peak levels. Così, our groups did not
differ in semantic or structural processing at the level of sentences or mini-discourses (maximal
number of sentences in a context is 4).
Neurobiology of Language
304
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Tavolo 8. Overlap between indirect vs. direct between-group interactions and networks
Number of overlapping voxels per neurosynth mask
Cluster (total number
of voxels)
Parietal (368 voxels)
Language comprehension
(association)
0/368 = 0%
ToM
(association)
0/368 = 0%
Cognitive control
(association)
7/368 = 2%
Cognitive control
(uniformity)
301/368 = 82%
Precuneus (399 voxels)
0/399 = 0%
3/399 = 1%
0/399 = 0%
36/399 = 9%
Note. The number (and percentage) of voxels from each cluster in the indirect vs. direct interaction analyses (rows) that lie in each neurosynth mask (column).
Note that cognitive control was used as a term to create a network corresponding to the MD network.
Overlap between indirect vs. direct interaction between groups result, and the language, ToM, E
MD/cognitive control networks
Tavolo 8 presents the results of the analysis described in the section Second level statistical
analysis, where neurosynth masks were used to estimate the overlap between the core lan-
guage, ToM, and MD/cognitive control networks on one hand and the parietal and precuneus
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 3. Parietal cluster. Single subject signal average (across voxels in the cluster) of indirect vs. direct first level contrast values for the
parietal cluster vs. scores for four behavioral tests: RMET, OSpan, vSweSAT, and LDT. Data for participants from the high (low) group are shown
as filled (not filled) dots. The p values indicate the outcome of two-tailed Pearson correlation tests with RIN-transformed data.
Neurobiology of Language
305
Pragmatic individual differences using fMRI
cluster found in the indirect vs. direct interaction analysis, on the other hand. The overlap was
generally low or non-existing, except for the uniformity cognitive control mask, which showed
an 82% overlap with the parietal cluster.
Correlation analyses for the indirect vs. direct contrast vs. behavior
As previously described in the section Second level statistical analyses, indirect vs. direct con-
trast values, extracted from the interaction analysis clusters (the parietal cluster and the pre-
cuneus cluster), were plotted against and tested for correlation with, the behavioral scores of
RMET, OSpan, vSweSAT, and LDT (see Figures 3 E 4, as well as Figure S4). This revealed no
significant correlations for RMET, OSpan, vSweSAT, or LDT; see Figures 3 E 4.
The results for RMET, OSpan, and LDT were expected as no significant difference between
the groups in terms of RMET, OSpan, or LDT score existed. In contrasto, there was a significant
difference between the groups in vSweSAT performance, and thus, the absence of correlation
between activation data and the vSweSAT score is perhaps more unexpected and more
informative.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 4. Precuneus cluster. Single subject signal average (across voxels in the cluster) of indirect vs. direct first level contrast values for the
precuneus cluster vs. scores for four behavioral tests: RMET, OSpan, vSweSAT, and LDT. Data for participants from the high (low) group are
shown as filled (not filled) dots. The p values indicate the outcome of two-tailed Pearson correlation tests with RIN-transformed data.
Neurobiology of Language
306
Pragmatic individual differences using fMRI
DISCUSSION
In an established paradigm comparing indirect vs direct speech acts across all participants, we
report our first results as expected activations in core language regions and beyond. Interesse-
ingly, individuals belonging to a group with high communicative skills (the HS group) showed
significantly more activity than those in a group with a low communicative skills (the LS
group) in two cortical clusters: (1) left superior and inferior lateral parietal cortex, E (2) bilat-
eral dorsal precuneus (Guarda la figura 2 and Table 6). These regions are located outside core lan-
guage and ToM regions (neurosynth language comprehension and ToM association maps; Vedere
Tavolo 8). Their activity did not depend on either of the core language skills (speed of lexical
access and vocabulary) or on ToM skills (see Figures 3 E 4). The absence of correlations
with the vocabulary test are noteworthy since behaviorally, our two pragmatic tests did cor-
relate with this test, which is not the case for the ToM test (Tavolo 5). There was no effect of
high vs. low communicative skill when participants initially listened to the informative con-
testo, which strengthens the specificity of the results to making communicative inferences. Noi
suggest that complex aspects of intention processing are subserved by a network including
these two regions (and potentially additional regions as discussed below). Based on the ToM
related results (behavioral correlations, neuroimaging segregation, and fMRI vs. behavioral
correlation), these complex aspects of intention processing may be specific to communicative
situations. Importantly, our results show a neurocognitive separation between these commu-
nicative inferences and comprehension processes that are rather involved with, Per esempio,
lexical and sentence level semantic and syntactic processing of the literal material, which are
subserved by different regions in the core language network.
The Indirect vs. Direct Effect, in Both Groups, as Potential Low Level Pragmatics
We verified that the activation pattern for the indirect vs. direct contrast for both groups rep-
licated the results in previous studies with similar paradigms (Asaridou et al., 2019; Bašnáková
et al., 2014); Guarda la figura 2, Tavolo 6, and Table 7. The same basic pattern was also present for the
HS group and LS group, although there were visible differences between HS and LS, per esempio., In
the lateral left inferior parietal region and the medial prefrontal cortex. The interpretation of
our reported individual differences rests on this replicability of the basic contrast of passive
observation of indirect vs. direct speech acts across different versions of the ISA experiment
that has been observed in the literature, Per esempio, across modalities (Asaridou et al., 2019;
Bašnáková et al., 2014). In this basic contrast, we observed overlapping activity for the HS
and LS groups in bilateral IFG, TPJ, anterior temporal lobe, medial SFG/dmPFC, and right mid
and posterior MTG/STS (Guarda la figura 2). The HS group activated a larger portion of cortices,
ventrally and dorsally, of the left TPJ. This group also showed significant activity (absent in the
LS group) in the precuneus, a pattern which partly resulted in a significant group interaction.
Essentially, all areas reported in the HS and LS groups, separately or in the overlap between
groups, were thus part of either the neurosynth language comprehension or ToM association
maps, in sharp contrast to the areas in the interaction that showed no overlap with these
maps, as reported in Table 8.
There are many possibilities of what these frontotemporo and frontoparietal regions
revealed by the indirect > direct contrast, might reflect. One line of explanation is to simply
see them as reflecting core language processes and ToM processes that are recruited more for
the indirect vs. the direct condition. The frontotemporal regions overlapping with the neuro-
synth language comprehension map could then correspond to top-down (downstream) effects
of pragmatic processing on core language processing. For instance, there is evidence that
Neurobiology of Language
307
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
listeners recognize certain speech acts similar to ISAs early on in utterance (Gisladottir et al.,
2015), which would allow for enhanced literal (or ultimately sentence level) processing, as a
downstream consequence of an ISA. It should be noted that by design, these activations can-
non, Tuttavia, correspond to bottom-up literal processing (bottom-up here relative to pragmatic
processes). The frontoparietal regions overlapping with the ToM association maps, on the other
hand, could correspond to intention processing, perspective-taking, and other ToM processes. UN
second line of explanation is that both these frontotemporal and frontoparietal regions reflect
processes such as the construction of a situation model, as the indirectness introduces a relative
complication in the situation model (Heidlmayr et al., 2020). More generally, these areas could
reflect low-level pragmatic processing, whereas the interaction would reflect more high-level
pragmatic processing.
Interpreting the Group Interaction: Pragmatics vis-à-vis ToM and Cognitive Control Functions
Recall that the HS group showed higher indirect vs. direct activity than the LS group in the left
lateral parietal cortex and dorsal precuneus, outside core language and ToM regions (Vedere
Figura 2 and Tables 6 E 8). Note that this group difference is present despite an absence
in other parameters (Vedi la tabella 2): There were no group differences in age, genere, behavioral
ToM skill (RMET), and CCF (OSpan). Inoltre, there was an absence of group differences
in processing the informative context, and no differences in performance of comprehension
questions.
As reported in Table 8, one of the neurosynth masks created by the cognitive control search
term did have a large overlap with the superior/inferior lateral parietal cluster. Additionally,
based on other reports in the literature on the location of the relevant networks, it would
be possible to consider the superior/inferior lateral parietal cluster to be roughly located in
the overlap of the ToM network and the MD network (Vincent et al., 2008). Likewise, IL
dorsal precuneus cluster could be considered a ToM area (see further discussion). Tuttavia,
it is notable that the complete ToM or MD networks were not activated. Così, considering the
locational information alone, activity in the two reported clusters could reflect MD/cognitive
control processes. There was, Tuttavia, no overlap with the neurosynth ToM masks and the
two clusters. There is additional evidence of segregation from the behavioral results (Vedere
Tavolo 5) as the performance on the ToM task (RMET) did not correlate with any of the prag-
matic tasks (PC-RR or AD). For CCF, the behavioral data shows that even though there seems
to be an interrelation between the OSpan test and the prosodic comprehension test (PC-RR),
there was a nonsignificant correlation with the AD production task. When we used the indi-
vidual variation in the RMET behavioral task as a proxy for variation in ToM as a neurocog-
nitive system, we found that neither of the two regions reported were modulated by individual
differences in ToM/complex emotion recognition (see Figures 3 E 4). The behavioral results
follow the same pattern, and thus the absence of modulation was expected due to the absence
of difference between the two groups (similar reasoning applies to the OSpan task). Taken
together, these results suggest a potential neurocognitive segregation of domain-specific com-
municative inference from domain-general ToM/complex emotion recognition.
Interpreting the Group Interaction as Potential Higher Order Pragmatics
As for sketching the nature of the potentially higher order pragmatic processing in the precu-
neus and parietal areas in the interaction between groups, we suggest that complex aspects of
intention processing (see further the sections: Interpreting the group interaction: Pragmatics
vis-à-vis ToM and cognitive control functions; Precuneus findings and large-scale functional
Neurobiology of Language
308
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
networks; Inferior and superior parietal cortex) and related internal and external attentional
processes (see further sections: Inferior and superior parietal cortex; Intraparietal sulcus, atten-
tion and intention) are subserved by these two regions (and potentially additional regions).
Again, the ToM part of the results suggests some specificity of these intentional and attentional
processes to the communicative situation. While communicative intention is fairly well
explored in the literature (Brunetti et al., 2014; Enrici et al., 2019; Rasgado-Toledo et al.,
2021; Tettamanti et al., 2017), there is less written about attentional processes specific to
the communicative situation, but we suggest that they might entail contextually driven atten-
tion to verbal and prosodic cues that would reveal an indirectness in an utterance, even before
these cues occur. Importantly, the absence of an interaction with groups when listening to the
contextual information, as well as the absence of an interaction with group performance on
the comprehension questions argues against group differences in terms of paying bare atten-
zione (as opposed to attention to, per esempio., specific cues) to the stimuli or comprehending the literal
content. Because of the overlap of the superior parietal cluster in the interaction between
groups and the cognitive control uniformity neurosynth map, we cannot exclude the possibil-
ity that these areas represent more general cognitive control processes (attentional processes
can again be mentioned as a possibility). (See further sections: Interpreting the group interac-
zione: Pragmatics vis-à-vis ToM and cognitive control functions; Inferior and superior parietal
cortex; Intraparietal sulcus, attention and intention.)
Precuneus Finding and Large-Scale Functional Networks
Unlike the parietal cluster we observed, the precuneus is a highly connected hub area (Nijhuis
et al., 2013). As such, it should not come as a surprise that it has been implicated in several
large-scale functional networks. First of all, it is often considered part of a general ToM network
(Paunov et al., 2019), but not always (Schurz et al., 2014), as is clear from its absence from the
ToM neurosynth mask used in Table 8. It has also been proposed as the very center of a sug-
gested intention processing network (Enrici et al., 2019). Our finding of increased precuneus
activity for participants with high pragmatic skills indicates that this area is indeed an impor-
tant cortical node subserving communicative intention processing and more specifically, as in
our task, the discernment of different speech acts. Tuttavia, second, while the precuneus is
only sometimes included in the ToM network, it is a core node in the default mode network
(DMN), and thus is a possible interface (and sometimes overlaps) between the ToM and DMN
(Amft et al., 2015; Mars et al., 2012). Third, precuneus has been implicated in the (right later-
alized) FPCN. We will come back to these observations when we discuss the parietal findings
in the following two paragraphs, from two different perspectives.
Inferior and Superior Parietal Cortex
The finding of increased activity for high skill participants indicates that this area is indeed an
important cortical node subserving communicative intention processing.
This suggests that previous views on intention processing in lateral parietal and temporal
areas might have to be modified. For instance, in an overview of the intention processing net-
lavoro, Enrici et al. (2019) suggest that the precuneus, right TPJ, and the mPFC—all three acti-
vated in our group averages (Figura 2)—encode “prospective social intentions,” while these
regions plus the left TPJ encode communicative intentions. The left TPJ is thus the region they
suggest to be specifically involved in communicative intentions; note, Tuttavia, that the sug-
gestion of specific intentional roles (per esempio., prospective social intentions) is only empirically sup-
ported by one study with a small sample size of N = 12 (Ciaramidaro et al., 2007). Our results
Neurobiology of Language
309
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
suggest that the literature should not ignore the intention processing role played by lateral pari-
etal areas dorsal of the TPJ (in addition to the precuneus, TPJ, and mPFC), as activity in these
areas may speculatively indicate routine recruitment linked to successful pragmatic perfor-
mance. As a comment on lateralization, with respect to this region, it is important to first note
that in a direct test of asymmetry, the right hemisphere area corresponding to the left latera-
lized cluster we report does not show a significantly different pattern (we discuss this further in
the supplemental discussion).
Turning to the inferior parietal cortex, specifically the angular and SMG, these areas were
activated by both groups (Guarda la figura 2), while the group interaction was in the superior parts of
these regions (extending into the IPS; see the next paragraph). The most likely interpretation is
that activity in the angular gyrus reflects conceptual processing of (indirect) speech acts, given
its presumed role as a main hub for conceptual/semantic processing (Binder et al., 2009).
Intraparietal Sulcus, Attention, and Intention
Given our results, in particular the findings in the mid and anterior IPS, it is relevant to also
discuss two additional large-scale brain networks that we did not choose to focus on a priori.
These networks are (1) the dorsal attention network (DAN), including the bilateral IPS, superior
parietal cortex, precuneus, and the frontal as well as supplementary eye fields; E (2) IL
mirror-neuron system (MNS), including the bilateral anterior IPS, premotor cortex, and some-
times inferior frontal cortex. The IPS is included in the top-down DAN. A specific suggestion
has been made for the domain-general attentional function of IPS. According to Bisley and
Goldberg (2010), the IPS consists of a set of priority maps selecting relevant features (hence
a top–down attentional process) across different tasks. This kind of explanation has also been
suggested in the context of a language task (Kristensen et al., 2013). From this point of view,
the IPS activity of the high score communicators might be providing more relevant top–down
attentional guidance, in the form of more precise and goal-effective priority maps, per esempio., to turn
attention to the potential verbal and prosodic cues that reveal the indirectness in the target
utterance, before they occurred in the stimuli. According to the view of Vincent et al.
(2008), our findings in the parietal cortex (including lateral inferior and superior parietal cor-
tices and the IPS) lie at the intersection of the DAN and the FPCN (or MD) rete, while our
precuneus findings lie at the intersection of their FPCN and the DMN network. A possible
interpretation is that these findings reflect the HS group’s better external and internal atten-
tional control, rispettivamente. We cannot exclude that the HS group found the task easier, Quale
might have enabled a larger amount of mind-wandering, potentially activating the DMN.
Tuttavia, it is then unclear why only this part of the DMN (rather than the whole network)
would be activated.
For the action system, Hamilton and Grafton (2006) suggest a hierarchical model of motor
regions with the anterior IPS at the top. This area is thought to represent action goals and inten-
zioni. While this suggestion is based on a large literature of hand-actions (Hamilton & Grafton,
2006; Iacoboni et al., 2005), per esempio., grasping actions and tool use, the question is whether this
analysis has any relevance for speech actions that we make during conversation. A reasonable
amount of skepticism is needed as the study of the MNS has suffered from severe hype
(Hickok, 2009), making the literature hard to navigate. Some authors interested in language,
communication, and other forms of joint action have indeed asked the question (Brennan
et al., 2010) or more directly suggested that the MNS is also involved in these tasks (Bekkering
et al., 2009). Does increased IPS activity for good communicators correspond to increased
intentional processing? The explanation is simple enough as stated, but there are also several
Neurobiology of Language
310
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
comments that need to be made alongside this suggestion. First of all, if there is any truth to this
statement, we would like to decouple this interpretation from the MNS, as we do not observe
any activity in other regions interpretable as mirror-neuron regions. If the anterior IPS is indeed
at the top of the action hierarchy, encoding intentions and goals, then this model is worth also
considering when studying conversational processes, but we do not see any reasons for further
reference here to mirror-neurons. Secondo, a limitation for this interpretation is that the IPS is not
routinely observed in communicative paradigms. For instance, it is not observed in many of the
studies using versions of the ISA experiment (Asaridou et al., 2019; Bašnáková et al., 2014),
although the left anterior IPS was observed in Egorova et al. (2016), who contrasted request
vs. naming, only using direct speech acts (see also van Ackeren et al., 2016). It is possible that
the individual differences approach improved sensitivity for the very top of the hierarchy, Quale
might not be as activated in all participants and hence not detected under conditions of group
averaging. In summary, we here provide possible attentional and intentional interpretations of
the unexpected IPS activity.
Theoretical Considerations
Our results can be discussed in the light of the theoretical suggestion of a submodule for com-
municative inferences (Sperber & Wilson, 2002). In Sperber and Wilson’s original suggestion
from relevance theory, inferential processes in comprehension are subserved by dedicated
domain-specific mechanisms, constituting a module (Fodor, 1983). Our results suggest that
pragmatic processes are likely mainly instantiated with mechanisms distinct from core lan-
guage mechanisms, as well as distinct from domain-general ToM mechanisms, even though
we investigate a broader set of pragmatic processes than originally suggested by Sperber and
Wilson (2002). Instead of referring to mentalizing as a unitary process, it is an interesting sug-
gestion that mentalizing is a large and multidimensional space of mental state representations,
a “mind space,” which would dynamically inform any situation where there is a need to con-
sult somebody’s mental state (Conway et al., 2019). This is a promising future direction for
ToM research.
Note that our findings do not allow us to take a stance on whether pragmatics constitutes a
(weak) module or a submodule of ToM, as suggested by relevance theory. Our position is that
pragmatics is neurocognitively separable from core language as well as (at least partly) from
aspects of ToM, and that the concept of ToM will need differentiation. In other words, prag-
matic inferencing does not equal mentalizing. We endorse the approach to study pragmatics in
its own right, which should continue.
Pragmatic analyses of different types of utterances even suggest that not all interpretations of
pragmatic phenomena require access to the mental state of the speaker in the first place, as is
the case of some generalized conversational implicatures pointed out in the introduction.
Some authors argue that even certain ISAs, such as indirect requests (per esempio., It’s cold in here)
can be interpreted without necessarily considering the mental state of the speaker (Kissine,
2016). Piuttosto, the interpretive process can sometimes be based purely on the accessibility
of certain salient information in the context; Per esempio, the listener knows that closing an
open window will relieve the speaker’s unpleasant feeling of being cold (see further the
effort/effect trade-off used in these inferences, below).
Another perspective from relevance theory (Sperber & Wilson, 1996) is that optimal rele-
vance is a trade-off between cognitive effort and effect on the listener. In cases of ambiguity
“hearers tend to choose the most salient or accessible meaning, the one that costs the least pro-
cessing effort to construct” (Sperber & Wilson, 2002, pag. 6–7; see also Gernsbacher, 1995). For
Neurobiology of Language
311
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
them, this shows that communicative meaning is not coded, but inferred outside the explicit and
conventionalized linguistic code system. Cognitive effort is thus suggested to be at the very heart
of processes establishing meaning. This effort might of course partly, or even mainly, be related to
pragmatic processes, but there is no particular reason why limited domain-general capacities,
per esempio., limited CCF, would not also produce similar effects on inferential processes. We provide
some evidence in this direction, as our behavioral comprehension measure PC-RR correlated to
a robust significant degree with OSpan in the behavioral study (N = 198), while the production
AD task did not. This finding has clinical relevance (Cummings, 2015).
Neurobiological and Clinical Perspectives
From a neurobiological perspective we provide evidence of two clusters constituting part of a
network with the function of interpreting communicative meaning: left superior and inferior
lateral parietal cortex (including IPS) and the bilateral dorsal precuneus. Individual differences
in our ISA experiment are related to other pragmatic processes, which is also informative for
future research in neuropragmatics. More specifically, a correlation between the two prag-
matic measures of prosodic speech act comprehension and AD in production was found
behaviorally (note, Tuttavia, the one-tailed threshold and absence of multiple comparison cor-
rections for this test; Vedi la tabella 5). A group interaction analysis revealed significant activation
during the ISA task. Since the groups were formed based on the pragmatic tests, this result
suggests (albeit somewhat indirectly) a relation between three different pragmatic processes,
one in production (AD) and two in comprehension (PC-RR and ISA), together suggesting that
contextualized (and multimodal) communication requires interrelated skill sets. These results,
including the neural results, are relevant for clinical research, for instance, in relation to the
more recent and specific diagnoses pragmatic language impairment (4th ed.; DSM-IV;
American Psychiatric Association, 2000) and social (pragmatic) communication disorder
(5th ed.; DSM-5; American Psychiatric Association, 2013). Activity in these two areas could
tentatively be seen as a biomarker for pragmatic competence, although our results at the same
time show that there is no one such thing as pragmatic competence (per esempio., as correlations
between pragmatic tasks are not very high; see further discussion on the relation between
different pragmatic measures in the supplemental discussion). If high skill participants stan-
dardly recruit these two areas to a larger degree when establishing communicative meaning,
the consequence might be increased connectivity with other areas involved in pragmatic pro-
cessing for these participants or enhanced (per esempio., more relevant or more detailed) representation
of speech acts for the HS participants in these areas.
Limitations
Regarding limitations of the current study, it should be noted that we can really only make
conclusions regarding the exact core language, ToM, and cognitive control aspects we mea-
sure through the selected tasks. A different selection of tasks may have manifested itself differ-
ently. While we partly base our statements about segregation on nonsignificant behavioral and
brain-behavioral correlations, we want to emphasize that we do not base our conclusions on
these alone. We also use converging evidence from locational information on the overlap
between the clusters in the interaction analysis and the functional networks we study. IL
neurocognitive separation from the language network is clearer than the segregation from
domain-general ToM, although we think we have also demonstrated some level of segregation
with aspects of ToM (per esempio., complex emotion recognition). From a skeptical point of view, if the
individual differences in the ISA experiment were interpreted as reflecting domain-general
processes rather than pragmatic processes, our data (per esempio., the left lateral parietal and IPS
Neurobiology of Language
312
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
activation) might be considered to point to attentional (per esempio., feature selection) rather than cog-
nitive control processes (per esempio., including maintenance, manipulation, and inhibition). An addi-
tional limitation of the study is that we do not have the behavioral data to test whether the HS
group would perform better or faster, per esempio., as a consequence of using higher order areas when
processing the dialogues. Finalmente, we would like to clarify that while we suggest that the left
lateral parietal and precuneus findings here should be understood as indexes of making com-
municative inferences, we do not suggest that these are the only areas involved in communi-
cative inferences (see supplemental discussion S3.4).
Conclusione
We interpret our data as suggesting that pragmatics should be reduced to neither (1) an exten-
sion of core language processes in special circumstances nor (2) domain-general
ToM/complex emotion processing or its combination with core language processes, even
though we cannot rule out there being partly shared processing. We suggest that the highly
replicable contrast of passive observation of indirect vs. direct speech could reflect low-level
pragmatic processing, whereas areas in the lateral parietal lobe and dorsal precuneus, that are
more active in this contrast for high communicative skill participants, could reflect more
high-level pragmatic processing. Inoltre, we suggest that individual differences in the
ISA experiment should be interpreted as reflecting the establishment of communicative
inference—a pragmatic process that our evidence indicates is related to other pragmatic pro-
cesses that we measured. While the data show some interrelations between different pragmatic
compiti, additional research would be needed to exclude the possibility that the individual dif-
ferences in pragmatic processing in the ISA experiment are partially related to differences in
some domain-general aspect of cognition that we did not measure. In conclusion, we suggest
that contextualized (and multimodal) communication requires interrelated and partly uniquely
communicative skill sets that cannot simply be reduced to other seemingly related cognitive
skills, such as core language skills, ToM skills, or cognitive control.
ACKNOWLEDGMENTS
This research was funded by the Promobilia Foundation, Riksbanken Jubileumsfond, IL
Swedish Collegium of Advances Studies, and the Magnus Bergvall Foundation. Data acquisi-
tion was supported by a grant via the Stockholm University Brain Imaging Centre (SU FV-5.1.2-
1035-15). We would also like to thank Daniel Pedersen for contributing to the translation of
the AR task to a Swedish audience and Salomi Asaridou for important feedback on the design
and manuscript.
FUNDING INFORMATION
Katarina Bendtz, Stiftelsen Promobilia. Julia Udde(cid:1)N, Riksbankens Jubileumsfond (http://dx.doi
.org/10.13039/501100004472). Julia Udde(cid:1)N, Swedish Collegium of Advanced Studies. Julia
Udde(cid:1)N, Magnus Bergvalls Stiftelse (http://dx.doi.org/10.13039/501100006285). Julia Udde(cid:1)N,
data acquisition supported by a grant to the Stockholm University Brain Imaging Centre,
Award ID: SU FV-5.1.2-1035-15.
AUTHOR CONTRIBUTIONS
Katarina Bendtz: Data curation; Formal analysis: Lead; Funding acquisition: Equal; Investigation:
Lead; Methodology: Equal; Project administration: Equal; Software; Visualization; Writing –
original draft: Equal; Writing – review and editing: Equal. Sarah Ericsson: Methodology:
Neurobiology of Language
313
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Supporting; Writing – review and editing: Supporting. Josephine Schneider: Investigation:
Supporting; Writing – review and editing: Supporting. Julia Borg: Investigation: Supporting.
Jana Bašnáková: Writing – review and editing: Supporting. Julia Uddén: Conceptualization;
Formal analysis; Funding acquisition: Equal; Methodology: Equal; Project administration:
Equal; Resources; Supervision; Writing – original draft: Equal; Writing – review and editing:
Equal.
REFERENCES
American Psychiatric Association. (2000). Diagnostic and statistical
manual of mental disorders (4th ed., text rev.).
American Psychiatric Association. (2013). Diagnostic and statistical
manual of mental disorders (5th ed.). https://doi.org/10.1176
/appi.books.9780890425596
Amft, M., Bzdok, D., Laird, UN. R., Fox, P. T., Schilbach, L., & Eickhoff,
S. B. (2015). Definition and characterization of an extended
social-affective default network. Brain Structure & Function,
220(2), 1031–1049. https://doi.org/10.1007/s00429-013-0698-0,
PubMed: 24399179
Asaridou, S. S., Demir-Lira, Ö. E., Uddén, J., Goldin-Meadow, S., &
Small, S. l. (2019). Pragmatic language processing in the adoles-
cent brain. BioRxiv, Article 871343. https://doi.org/10.1101
/871343
Baron-Cohen, S., Wheelwright, S., Hill, J., Raste, Y., & Plumb, IO.
(2001). The “Reading the mind in the eyes” test revised version:
A study with normal adults, and adults with Asperger syndrome
or high-functioning autism. Journal of Child Psychology and Psy-
chiatry, 42(2), 241–251. https://doi.org/10.1111/1469-7610
.00715, PubMed: 11280420
Bašnáková, J. (2019). Beyond the language given: The neurobiolog-
ical infrastructure for pragmatic inferencing [Doctoral thesis,
Radboud University Nijmegen]. Radboud Repository. https://
repository.ubn.ru.nl/handle/2066/200653
Bašnáková, J., Weber, K., Petersson, K. M., van Berkum, J., &
Hagoort, P. (2014). Beyond the language given: The neural cor-
relates of inferring speaker meaning. Cerebral Cortex, 24(10),
2572–2578. https://doi.org/10.1093/cercor/ bht112, PubMed:
23645715
Bekkering, H., De Bruijn, E. R. A., Cuijpers, R. H., Newman-
Norlund, R., Van Schie, H. T., & Meulenbroek, R. (2009). Joint
action: Neurocognitive mechanisms supporting human interac-
zione. Topics in Cognitive Science, 1(2), 340–352. https://doi.org
/10.1111/j.1756-8765.2009.01023.x, PubMed: 25164937
Campana, UN. (1984). Language style as audience design. Language in Soci-
ety, 13(2), 145–204. https://doi.org/10.1017/S004740450001037X
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, l. l. (2009).
Where is the semantic system? A critical review and
meta-analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19(12), 2767–2796. https://doi.org/10.1093/cercor
/bhp055, PubMed: 19329570
Bishara, UN. J., & Hittner, J. B. (2012). Testing the significance of a
correlation with nonnormal data: Comparison of Pearson, Spear-
Uomo, transformation, and resampling approaches. Psicologico
Methods, 17(3), 399–417. https://doi.org/10.1037/a0028087,
PubMed: 22563845
Bisley, J. W., & Goldberg, M. E. (2010). Attention, intention, E
priority in the parietal lobe. Annual Review of Neuroscience,
33, 1–21. https://doi.org/10.1146/annurev-neuro-060909
-152823, PubMed: 20192813
Blank, I., Kanwisher, N., & Fedorenko, E. (2014). A functional dis-
sociation between language and multiple-demand systems
revealed in patterns of BOLD signal fluctuations. Journal of Neu-
rophysiology, 112(5), 1105–1118. https://doi.org/10.1152/jn
.00884.2013, PubMed: 24872535
Bosco, F. M., Tirassa, M., & Gabbatore, IO. (2018). Why pragmatics
and theory of mind do not (completely) sovrapposizione. Frontiers in Psy-
chology, 9, Article 1453. https://doi.org/10.3389/fpsyg.2018
.01453, PubMed: 30150960
Brennan, S. E., Galati, A., & Kuhlen, UN. K. (2010). Two minds,
In
one dialog: Coordinating speaking and understanding.
B. H. Ross (Ed.), The psychology of learning and motivation:
Advances in research and theory (pag. 301–344). Elsevier
Academic Press. https://doi.org/10.1016/S0079-7421(10)53008
-1
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002, Giugno
2–6). Region of interest analysis using an SPM toolbox [Paper
presentation]. 8th International Conference on Functional Map-
ping of the Human Brain, Sendai, Japan. Available at https://
www.scienceopen.com/document?vid=f9a14c27-7c57-4539
-933d-d73833ff5232
Brown, P., & Levinson, C. (1987). Politeness: Some universals in
language usage. Cambridge University Press. https://doi.org/10
.1017/CBO9780511813085
Brown-Schmidt, S. (2009). The role of executive function in per-
spective taking during online language comprehension. Psycho-
nomic Bulletin & Review, 16(5), 893–900. https://doi.org/10
.3758/PBR.16.5.893, PubMed: 19815795
Brown-Schmidt, S., & Fraundorf, S. H. (2015). Interpretation of
informational questions modulated by joint knowledge and into-
national contours. Journal of Memory and Language, 84, 49–74.
https://doi.org/10.1016/j.jml.2015.05.002
Brunetti, M., Zappasodi, F., Marzetti, L., Perrucci, M. G., Cirillo, S.,
Romani, G. L., Pizzella, V., & Aureli, T. (2014). Do you know
what I mean? Brain oscillations and the understanding of
communicative intentions. Frontiers in Human Neuroscience,
8, Article 36. https://doi.org/10.3389/fnhum.2014.00036,
PubMed: 24550813
Chein, J. M., Moore, UN. B., & Conway, UN. R. UN. (2011). Domain-
general mechanisms of complex working memory span. Neuro-
Image, 54(1), 550–559. https://doi.org/10.1016/j.neuroimage
.2010.07.067, PubMed: 20691275
Ciaramidaro, A., Adenzato, M., Enrici, I., Erk, S., Pia, L., Bara, B. G.,
& Walter, H. (2007). The intentional network: How the brain
reads varieties of intentions. Neuropsychologia, 45(13), 3105–3113.
https://doi.org/10.1016/j.neuropsychologia.2007.05.011, PubMed:
17669444
Clark, H. H. (1994). Managing problems in speaking. Speech Com-
munication, 15(3–4), 243–250. https://doi.org/10.1016/0167
-6393(94)90075-2
Neurobiology of Language
314
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Clark, H. H., Schreuder, R., & Buttrick, S. (1983). Common ground
and the understanding of demonstrative reference. Journal of Ver-
bal Learning & Comportamento verbale, 22(2), 245–258. https://doi.org
/10.1016/S0022-5371(83)90189-5
Cliffordson, C. (2004). Effects of practice and intellectual growth on
performance on the Swedish Scholastic Aptitude Test (SweSAT).
European Journal of Psychological Assessment, 20(3), 192–204.
https://doi.org/10.1027/1015-5759.20.3.192
Conway, J. R., Catmur, C., & Bird, G. (2019). Understanding
individual differences in theory of mind via representation of
minds, not mental states. Psychonomic Bulletin & Review, 26(3),
798–812. https://doi.org/10.3758/s13423-018-1559-x, PubMed:
30652239
Cummings, l. (2015). Theory of mind in utterance interpretation:
The case from clinical pragmatics. Frontiers in Psychology, 6,
Article 1286. https://doi.org/10.3389/fpsyg.2015.01286,
PubMed: 26379602
Domanesci, F., & Bambini, V. (2020). Pragmatic competence. In E.
Fridland & C. Pavese (Eds.), Routledge handbook of skill and
expertise. Springer. PsyArXiv. https://doi.org/10.31234/osf.io
/q8msa
Duncan, J. (2010). The multiple-demand (MD) system of the pri-
mate brain: Mental programs for intelligent behaviour. Trends
in Cognitive Sciences, 14(4), 172–179. https://doi.org/10.1016/j
.tics.2010.01.004, PubMed: 20171926
Egorova, N., Shtyrov, Y., & Pulvermüller, F. (2016). Brain basis of
communicative actions in language. NeuroImage, 125, 857–867.
https://doi.org/10.1016/j.neuroimage.2015.10.055, PubMed:
26505303
Enrici, I., Bara, B. G., & Adenzato, M. (2019). Theory of Mind,
pragmatics and the brain: Converging evidence for the role of
intention processing as a core feature of human communication.
Pragmatics & Cognition, 26(1), 5–38. https://doi.org/10.1075/pc
.19010.enr
Faraco, C. C., Unsworth, N., Langley, J., Terry, D., Li, K., Zhang, D.,
Liu, T., & Mugnaio, l. S. (2011). Complex span tasks and hippocam-
pal recruitment during working memory. NeuroImage, 55(2),
773–787. https://doi.org/10.1016/j.neuroimage.2010.12.033,
PubMed: 21182968
Fodor, J. (1983). The modularity of mind. CON Premere. https://doi.org
/10.7551/mitpress/4737.001.0001
Foster, J. L., Shipstead, Z., Harrison, T. L., Hicks, K. L., Redick, T. S.,
& Engle, R. W. (2015). Shortened complex span tasks can reliably
measure working memory capacity. Memory & Cognition, 43(2),
226–236. https://doi.org/10.3758/s13421-014-0461-7, PubMed:
25217113
Friederici, UN. D. (2012). The cortical language circuit: From audi-
tory perception to sentence comprehension. Trends in Cognitive
Scienza, 16(5), 262–268. https://doi.org/10.1016/j.tics.2012.04
.001, PubMed: 22516238
Friston, K. J. (2007). Statistical parametric mapping. In W. Penny,
K. J. Friston, J. Ashburner, S. Kiebel, & T. Nichols (Eds.), Statistical
parametric mapping: The analysis of functional brain images
(pag. 10–31). Elsevier. https://doi.org/10.1016/B978-012372560
-8/50002-4
Garrett, M., & Harnish, R. M. (2007). Experimental pragmatics:
Testing for implicitures. Pragmatics & Cognition, 15(1), 65–90.
https://doi.org/10.1075/pc.15.1.07gar
Gernsbacher, M. (1995). Handbook of psycholinguistics. Academic
Press.
Gisladottir, R. S., Chwilla, D. J., & Levinson, S. C. (2015). Conver-
sation electrified: ERP correlates of speech act recognition in
underspecified utterances. PLOS ONE, 10(3), Article e0120068.
https://doi.org/10.1371/journal.pone.0120068, PubMed:
25793289
Goffman, E. (1967). Interaction ritual: Essays on face-to-face inter-
action. Aldine.
Grice, H. P. (1975). Logic and conversation. In P. Cole & J. l.
Morgan (Eds.), Syntax and Semantics, Vol. 3, Speech Acts
(pag. 41–58). Elsevier Academic Press. https://doi.org/10.1163
/9789004368811_003
Hagoort, P. (2017). The core and beyond in the language-ready
brain. Neuroscience & Biobehavioral Reviews, 81(Pt B),
194–204. https://doi.org/10.1016/j.neubiorev.2017.01.048,
PubMed: 28193452
Hamilton, UN. F., & Grafton, S. T. (2006). Goal representation in
human anterior intraparietal sulcus. Journal of Neuroscience,
26(4), 1133–1137. https://doi.org/10.1523/JNEUROSCI.4551-05
.2006, PubMed: 16436599
Heidlmayr, K., Weber, K., Takashima, UN. & Hagoort, P. (2020). No
titolo, no theme: The joined neural space between speakers and
listeners during production and comprehension of multi-sentence
discourse. Cortex, 130, 111–126. https://doi.org/10.1016/j.cortex
.2020.04.035, PubMed: 32652339
Hellbernd, N., & Sammler, D. (2016). Prosody conveys speaker’s
intentions: Acoustic cues for speech act perception. Journal of
Memory and Language, 88, 70–86. https://doi.org/10.1016/j.jml
.2016.01.001
Hellbernd, N., & Sammler, D. (2018). Neural bases of social com-
municative intentions in speech. Social Cognitive and Affective
Neuroscience, 13(6), 604–615. https://doi.org/10.1093/scan
/nsy034, PubMed: 29771359
Hickok, G. (2009). Eight problems for the mirror neuron theory of
action understanding in monkeys and humans. Journal of Cogni-
tive Neuroscience, 21(7), 1229–1243. https://doi.org/10.1162
/jocn.2009.21189, PubMed: 19199415
Hickok, G., & Poeppel, D. (2004). Dorsal and ventral streams: UN
framework for understanding aspects of the functional anatomy
of language. Cognition, 92(1–2), 67–99. https://doi.org/10.1016/j
.cognition.2003.10.011, PubMed: 15037127
Holmer, E., Heimann, M., & Rudner, M. (2016). Evidence of an
association between sign language phonological awareness
and word reading in deaf and hard-of-hearing children. Research
in Developmental Disabilities, 48, 145–159. https://doi.org/10
.1016/j.ridd.2015.10.008, PubMed: 26561215
Holtgraves, T. (1998). Interpreting indirect replies. Cognitive Psy-
chology, 37(1), 1–27. https://doi.org/10.1006/cogp.1998.0689,
PubMed: 9787060
Iacoboni, M., Molnar-Szakacs, I., Gallese, V., Buccino, G.,
Mazziotta, J. C., & Rizzolatti, G. (2005). Grasping the intentions
of others with one’s own mirror neuron system. PLOS Biology,
3(3), Article e79. https://doi.org/10.1371/journal.pbio.0030079,
PubMed: 15736981
Kalandadze, T., Norbury, C., Nærland, T., & Næss, K. B. (2018).
Figurative language comprehension in individuals with autism
spectrum disorder: A meta-analytic review. Autism, 22(2),
99–117. https://doi.org/10.1177/1362361316668652, PubMed:
27899711
Kissine, M. (2016). Pragmatics as metacognitive control. Frontiers
in Psychology, 6, Article 2057. https://doi.org/10.3389/fpsyg
.2015.02057, PubMed: 26834671
Krauss, R. M., & Glucksberg, S. (1977). Social and nonsocial
speech. Scientific American, 236(2), 100–105. https://doi.org/10
.1038/scientificamerican0277-100
Kristensen, l. B., Wang, L., Petersson, K. M., & Hagoort, P.
(2013). The interface between language and attention:
Neurobiology of Language
315
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
Prosodic focus marking recruits a general attention network in
spoken language comprehension. Cerebral Cortex, 23(8),
1836–1848. https://doi.org/10.1093/cercor/ bhs164, PubMed:
22763170
Levinson, S. C. (2006). On the human “interaction engine”. In N. J.
Enfield & S. C. Levinson (Eds.), Roots of human sociality: Culture,
cognition and interaction (pag. 39–69). Berg. https://doi.org/10
.4324/9781003135517-3
Mars, R., Neubert, F.-X., Noonan, M., Sallet, J., Toni, I., &
Rushworth, M. (2012). On the relationship between the “default
mode network” and the “social brain”. Frontiers in Human
Neuroscience, 6, Article 189. https://doi.org/10.3389/fnhum
.2012.00189, PubMed: 22737119
Matthews, D., Biney, H., & Abbot-Smith, K. (2018). Individual dif-
ferences in children’s pragmatic ability: A review of associations
with formal language, social cognition, and executive functions.
Language Learning and Development, 14(3), 186–223. https://doi
.org/10.1080/15475441.2018.1455584
Moore, M., & Gordon, P. C. (2015). Reading ability and print expo-
sure: Item response theory analysis of the author recognition test.
Behavior Research Methods, 47(4), 1095–1109. https://doi.org
/10.3758/s13428-014-0534-3, PubMed: 25410405
Nijhuis, E. H., van Cappellen van Walsum, A.-M., & Norris, D. G.
(2013). Topographic hub maps of the human structural neocorti-
cal network. PLOS ONE, 8(6), Article e65511. https://doi.org/10
.1371/journal.pone.0065511, PubMed: 23935801
Oakley, B. F. M., Brewer, R., Bird, G., & Catmur, C. (2016). Theory
of mind is not theory of emotion: A cautionary note on the Read-
ing the Mind in the Eyes Test. Journal of Abnormal Psychology,
125(6), 818–823. https://doi.org/10.1037/abn0000182, PubMed:
27505409
Osaka, M., Osaka, N., Kondo, H., Morishita, M., Fukuyama, H.,
Aso, T., & Shibasaki, H. (2003). The neural basis of individual
differences in working memory capacity: An fMRI study. Neuro-
Image, 18(3), 789–797. https://doi.org/10.1016/S1053-8119(02)
00032-0, PubMed: 12667855
Paunov, UN. M., Blank, IO. A., & Fedorenko, E. (2019). Functionally
distinct language and theory of mind networks are synchronized
at rest and during language comprehension. Journal of Neuro-
physiology, 121(4), 1244–1265. https://doi.org/10.1152/jn
.00619.2018, PubMed: 30601693
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M., Höchenberger, R.,
Sogo, H., Kastman, E., & Lindeløv, J. K. (2019). PsychoPy2:
Experiments in behavior made easy. Behavior Research Methods,
51(1), 195–203. https://doi.org/10.3758/s13428-018-01193-y,
PubMed: 30734206
Rasgado-Toledo, J., Valles-Capetillo, E., Giudicessi, A., &
Giordano, M. (2021). Neural processing of facial expressions
as modulators of communicative intention. PsyArXiv. https://doi
.org/10.31234/osf.io/tsem8
Roßnagel, C. (2000). Cognitive load and perspective-taking: Applying
the automatic-controlled distinction to verbal communication.
European Journal of Social Psychology, 30(3), 429–445. https://doi
.org/10.1002/(SICI )1099-0992(200005/06)30:3<429::AID
-EJSP3>3.0.CO;2-V
Roßnagel, C. S. (2004). Lost in thought: Cognitive load and the pro-
cessing of addressees’ feedback in verbal communication. Exper-
imental Psychology, 51(3), 191–200. https://doi.org/10.1027
/1618-3169.51.3.191, PubMed: 15267127
Rubio-Fernández, P. (2017). The director task: A test of theory-of-
mind use or selective attention? Psychonomic Bulletin & Review,
24(4), 1121–1128. https://doi.org/10.3758/s13423-016-1190-7,
PubMed: 27822775
Ryskin, R. A., Benjamin, UN. S., Tullis, J., & Brown-Schmidt, S.
(2015). Perspective-taking in comprehension, production, E
memory: An individual differences approach. Journal of Experi-
mental Psychology: General, 144(5), 898–915. https://doi.org/10
.1037/xge0000093, PubMed: 26214163
Ryskin, R. A., Brown-Schmidt, S., Canseco-Gonzalez, E., Yiu, l. K., &
Nguyen, E. T. (2014). Visuospatial perspective-taking in conversation
and the role of bilingual experience. Journal of Memory and Lan-
guage, 74, 46–76. https://doi.org/10.1016/j.jml.2014.04.003
Sacks, H., & Schegloff, E. UN. (1979). Two preferences in the
organization of reference to persons in conversation and their
interaction. In G. Psathas (Ed.), Everyday language: Studies in
ethnomethodology (pag. 15–21). Irvington Publishers.
Santiesteban, I., Shah, P., White, S., Bird, G., & Heyes, C. (2015).
Mentalizing or submentalizing in a communication task? Evi-
dence from autism and a camera control. Psychonomic Bulletin
& Review, 22(3), 844–849. https://doi.org/10.3758/s13423-014
-0716-0, PubMed: 25149442
Schaafsma, S. M., Pfaff, D. W., Spunt, R. P., & Adolphs, R. (2015).
Deconstructing and reconstructing theory of mind. Trends in
Cognitive Science, 19(2), 65–72. https://doi.org/10.1016/j.tics
.2014.11.007, PubMed: 25496670
Schurz, M., Kronbichler, M., Weissengruber, S., Surtees, A.,
Samson, D., & Perner, J. (2015). Clarifying the role of theory
of mind areas during visual perspective taking: Issues of spon-
taneity and domain-specificity. NeuroImage, 117, 386–396.
https://doi.org/10.1016/j.neuroimage.2015.04.031, PubMed:
25907759
Schurz, M., Radua, J., Aichhorn, M., Richlan, F., & Perner, J. (2014).
Fractionating theory of mind: A meta-analysis of functional brain
imaging studies. Neuroscience & Biobehavioral Reviews, 42,
9–34. https://doi.org/10.1016/j.neubiorev.2014.01.009,
PubMed: 24486722
Senju, A., Tojo, Y., Konno, M., Dairoku, H., & Hasegawa, T. (2002).
[Reading mind from pictures of eyes: Theory of mind, lingua
ability, general intellectual ability, and autism]. Shinrigaku Ken-
kyu: The Japanese Journal of Psychology, 73(1), 64–70. https://doi
.org/10.4992/jjpsy.73.64, PubMed: 12101592
Shamay-Tsoory, S. G., & Aharon-Peretz, J. (2007). Dissociable pre-
frontal networks for cognitive and affective theory of mind: UN
lesion study. Neuropsychologia, 45(13), 3054–3067. https://doi
.org/10.1016/j.neuropsychologia.2007.05.021, PubMed:
17640690
Sperber, D., & Wilson, D. (1996). Relevance: Communication and
cognition (2nd ed.). Wiley.
Sperber, D., & Wilson, D. (2002). Pragmatics, modularity and
mind-reading. Mind & Language, 17(1–2), 3–23. https://doi.org
/10.1111/1468-0017.00186
Stanovich, K. E., & West, R. F. (1989). Exposure to print and ortho-
graphic processing. Reading Research Quarterly, 24(4), 402–433.
https://doi.org/10.2307/747605
Tettamanti, M., Vaghi, M. M., Bara, B. G., Cappa, S. F., Enrici, I., &
Adenzato, M. (2017). Effective connectivity gateways to the
theory of mind network in processing communicative intention.
NeuroImage, 155, 169–176. https://doi.org/10.1016/j
.neuroimage.2017.04.050, PubMed: 28438665
van Ackeren, M. J., Smaragdi, A., & Rueschemeyer, S.-A. (2016).
Neuronal interactions between mentalising and action systems
during indirect request processing. Social Cognitive and Affective
Neuroscience, 11(9), 1402–1410. https://doi.org/10.1093/scan
/nsw062, PubMed: 27131039
Vincent, J. L., Kahn, I., Snyder, UN. Z., Raichle, M. E., & Buckner,
R. l. (2008). Evidence for a frontoparietal control system
Neurobiology of Language
316
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pragmatic individual differences using fMRI
revealed by intrinsic functional connectivity. Journal of Neuro-
physiology, 100(6), 3328–3342. https://doi.org/10.1152/jn
.90355.2008, PubMed: 18799601
Wardlow, l. (2013). Individual differences in speakers’ perspective
taking: The roles of executive control and working memory. Psy-
chonomic Bulletin & Review, 20(4), 766–772. https://doi.org/10
.3758/s13423-013-0396-1, PubMed: 23408369
West, R. F., Stanovich, K. E., & Mitchell, H. R. (1993). Reading in
the real world and its correlates. Reading Research Quarterly,
28(1), 34–50. https://doi.org/10.2307/747815
Wilson, UN. C., & Bishop, D. V. M. (2019). “If you catch my drift…":
Ability to infer implied meaning is distinct from vocabulary and
grammar skills. Wellcome Open Research, 4, 68. https://doi.org
/10.12688/wellcomeopenres.15210.2, PubMed: 31245632
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
3
2
2
8
7
2
0
0
8
4
7
5
N
o
_
UN
_
0
0
0
6
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurobiology of Language
317