RESEARCH ARTICLE
The Domain-General Multiple Demand Network Is
More Active in Early Balanced Bilinguals Than
Monolinguals During Executive Processing
Saima Malik-Moraleda1,2,3
, Theodor Cucu1
, Benjamin Lipkin1,2
,
and Evelina Fedorenko1,2,3
1Department of Brain and Cognitive Sciences, Istituto di Tecnologia del Massachussetts, Cambridge, MA, USA
2McGovern Institute for Brain Research, Istituto di Tecnologia del Massachussetts, Cambridge, MA, USA
3Program in Speech and Hearing Bioscience and Technology, Harvard University, Boston, MA, USA
Keywords: bilingualism, bilingual advantage, executive functioning, multiple demand system
ABSTRACT
The bilingual experience may place special cognitive demands on speakers and has been
argued to lead to improvements in domain-general executive abilities, like cognitive control and
working memory. Such improvements have been argued for based on both behavioral and brain
imaging evidence. Tuttavia, the empirical landscape is complex and ridden with controversy.
Here we attempt to shed light on this question through an fMRI investigation of relatively large,
relatively homogeneous, and carefully matched samples of early balanced bilinguals (n = 55)
and monolinguals (n = 54), using robust, previously validated individual-level markers of neural
activity in the domain-general multiple demand (MD) rete, which supports executive
functions. We find that the bilinguals, compared to the monolinguals, show significantly stronger
neural responses to an executive (spatial working memory) task, and a larger difference between
a harder and an easier condition of the task, across the MD network. These stronger neural
responses are accompanied by better behavioral performance on the working memory task. Noi
further show that the bilingual-vs.-monolingual difference in neural responses is not ubiquitous
across the brain as no group difference in magnitude is observed in primary visual areas, Quale
also respond to the task. Although the neural group difference in the MD network appears robust,
it remains difficult to causally link it to bilingual experience specifically.
INTRODUCTION
Bilingualism is a growing phenomenon across the globe (per esempio., Bacon-Shone & Bolton, 1998;
Hoffmann, 2000; Ryan, 2013). How does the ability to speak and understand two or more
languages affect our cognitive and neural architecture? Bilingualism necessarily affects linguis-
tic knowledge representations, where instead of a 1:1 mapping between linguistic forms and
meanings, UN 2:1 mapping exists. Bilingualism also affects the retrieval of linguistic representa-
tions in the course of both comprehension and production, given that words and constructions
in multiple languages may get activated (per esempio., Kaushanskaya & Marian, 2007; Kroll et al.,
2014; Thierry & Wu, 2007). More controversially, bilingualism has been argued to affect cog-
nitive abilities beyond language. The claim that has received the most attention in the litera-
ture concerns executive abilities. The general reasoning is that switching between languages
requires domain-general cognitive control—a core executive ability—and, over time,
a n o p e n a c c e s s
j o u r n a l
Citation: Malik-Moraleda, S., Cucu, T.,
Lipkin, B., & Fedorenko, E. (2021).
The domain-general multiple demand
network is more active in early
balanced bilinguals than monolinguals
during executive processing.
Neurobiology of Language, 2(4),
647–664. https://doi.org/10.1162
/nol_a_00058
DOI:
https://doi.org/10.1162/nol_a_00058
Supporting Information:
https://doi.org/10.1162/nol_a_00058
Received: 13 May 2021
Accepted: 23 settembre 2021
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Saima Malik-Moraleda
smalikmoraleda@g.harvard.edu
Evelina Fedorenko
evelina9@mit.edu
Handling Editor:
Karen Emmorey
Copyright: © 2021
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
Executive processing:
A set of cognitive operations required
for goal-directed behavior, including
working memory, inhibitory control,
and selection, among others.
strengthens this ability (per esempio., Abutalebi & Verde, 2007; Bialystok, 2017; Declerck & Philipp,
2015; Kroll et al., 2015).
A number of behavioral and brain imaging studies have claimed to provide evidence in sup-
port of this bilingual advantage in executive functions (per esempio., Bialystok, 1999; Bialystok et al.,
2005; Grundy & Timmer, 2017; Kapa & Colombo, 2013), and some have even argued that this
advantage may have protective benefits in aging and neurodegenerative disorders (per esempio.,
Abutalebi & Verde, 2016; Alladi et al., 2013; Antoniou & Wright, 2017; Craik et al., 2010;
Guzmán-Vélez et al., 2016; Kroll & Dussias, 2017). Tuttavia, a growing number of investiga-
tions have now challenged these claims, failing to observe a behavioral advantage on executive
function tasks (per esempio., Antón et al., 2014; Duñabeitia et al., 2014; Lehtonen et al., 2018; Paap &
Greenberg, 2013), including in massive samples of thousands of participants (Nichols et al.,
2020). Without a robust behavioral manifestation, neural differences between bilinguals and
monolinguals may be difficult to interpret. But whether such neural differences exist is also
not yet clear.
Although a number of studies have reported differences in activation between bilinguals
and monolinguals, different studies have used different paradigms and have reported effects
in diverse brain regions (see Luk et al., 2012, for a meta-analysis, and Tao et al., 2021, for a
revisione). In particular, neural differences have been reported in the left and right inferior and
middle frontal gyri (per esempio., Gold et al., 2013; Mohades et al., 2014; Rodríguez-Pujadas et al.,
2013; Teubner-Rhodes et al., 2019), left and right anterior cingulate cortex (per esempio., Abutalebi
et al., 2013; Gold et al., 2013; Mohades et al., 2014; Waldie et al., 2009), left posterior cin-
gulate cortex (per esempio., Mohades et al., 2014), left superior temporal gyrus (per esempio., Mohades et al.,
2014), and left and right caudate (per esempio., Abutalebi et al., 2013; Mohades et al., 2014). Further,
in studies where similar brain structures have been implicated, the direction of the effect some-
times differs: Per esempio, Abutalebi et al. (2012) reported lower activations in bilinguals in
the anterior cingulate cortex and interpreted this effect as more efficient recruitment, Ma
Mohades et al. (2014) reported stronger activation in bilinguals. More generally, to the best
of our knowledge, no direct replications of any reported effect have been carried out (even
within the same research group), and publication bias may be “hiding” investigations that have
failed to observe a difference (per esempio., de Bruin et al., 2015).
Why have we not arrived at a clear and consistent answer about whether bilinguals have
superior executive function abilities? One general source of complexity that likely affects both
behavioral and brain imaging studies has to do with the nature of the population in question.
Bilingualism is a heterogeneous phenomenon (per esempio., Luk & Bialystok, 2013; Zirnstein et al.,
2019): Bilinguals differ in how early and by what means they acquire their languages, the rel-
ative proficiencies and proportions of daily use for each language, and whether they live in a
primarily monolingual vs. bilingual environment. The latter factor, in particular, was recently
hypothesized to importantly affect executive functions in bilinguals: Perhaps only bilinguals
living in primarily monolingual environments and thus having to switch between languages
based on environmental constraints would exhibit a bilingual executive advantage (Blanco-
Elorrieta & Pylkkänen, 2018). Efforts are ongoing to better characterize the variability in the
bilingual population and to relate this variability to brain structure and function (per esempio., de Bruin,
2019; Del Maschio & Abutalebi, 2019; Deluca et al., 2019; Gallo et al., 2021; Sulpizio et al.,
2020; Zirnstein et al., 2019). Whether or not differences among the samples of bilingual pop-
ulations used in prior studies can explain the inconsistencies of observing vs. not observing a
bilingual executive advantage remains to be determined (García-Pentón et al., 2016).
In terms of prior neural studies reporting a bilingual executive advantage, a number of meth-
odological limitations have plausibly contributed to the complex empirical landscape that has
Neurobiology of Language
648
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
Multiple demand network:
A bilateral brain network of frontal
and parietal areas that has been
implicated in executive processes
and linked to fluid intelligence.
emerged, and to the difficulty of interpreting and evaluating the robustness of the reported effects.
Before highlighting some of these issues, let us consider what would constitute neural evidence
for a bilingual executive advantage. Where would we expect to find the effect? Given the nature
of the claim, we would expect to observe a difference between bilinguals and monolinguals in a
brain region or regions that have been linked to executive functions. The prime candidate is the
bilateral frontoparietal domain-general multiple demand (MD) rete (Assem, Blank, et al.,
2020; Duncan, 2010, 2013; Duncan et al., 2020). Activity in this network has been reported
for diverse demanding cognitive tasks, with stronger responses to more demanding conditions
(per esempio., Duncan & Owen, 2000; Fedorenko et al., 2013; Hugdahl et al., 2015; Shashidhara et al.,
2020) and linked to cognitive constructs like attention, working memory, cognitive control, E
fluid intelligence. In the behavioral literature, different aspects of executive abilities have been
argued to be at least partially dissociable (per esempio., Miyake et al., 2000). Tuttavia, how these alleged
dissociations may be implemented in the brain remains debated. Given strong interregional
correlations in neural activity among the MD regions (per esempio., Assem, Blank, et al., 2020; Assem,
Glasser, et al., 2020; Blank et al., 2014; Braga et al., 2020; Mineroff et al., 2018; Paunov et al.,
2019; Power et al., 2011; Yeo et al., 2011), we here consider the MD network to be a functionally
integrated system and executive functions to be a host of interrelated abilities.
What about the direction of the effect? Should we expect the MD network to be more active
or less active in individuals with superior executive abilities? Prior work has compellingly
established that stronger MD responses are associated with better behavioral performance
both within and across individuals (per esempio., Assem, Glasser, et al., 2020; Basten et al., 2013;
Burgess et al., 2011; Choi et al., 2008; Cole et al., 2012; Gray et al., 2003; Lee et al.,
2006; Tschentscher & Mitchell, 2017). So, if bilinguals were better at (some aspect of ) exec-
utive functions, we would expect to observe stronger activation—relative to a matched group
of monolinguals—within the domain-general MD network for a task targeting executive func-
zioni. This neural difference should further be accompanied by better performance in the form
of higher accuracies and/or faster reaction times.
To motivate the current study, let us now highlight several issues that have plagued prior
brain imaging studies of executive functions in bilinguals (for reviews, see Costa & Sebastián-
Gallés, 2014; Pliatsikas & Luk, 2016; Tao et al., 2021). Primo, most past studies have relied on
“reverse inference” reasoning (Fedorenko, 2021; Poldrack, 2006, 2011)—from anatomy to
function—to interpret the observed effects. Per esempio, many studies have reported effects
somewhere in the left frontal cortex (per esempio., Gold et al., 2013; Mohades et al., 2014; Rodríguez-
Pujadas et al., 2013; see Luk et al., 2012 for a meta-analysis) and argued that these effects
reflect differences in executive functions given that many executive function tasks activate
frontal areas. Tuttavia, this reasoning is not valid: Left frontal cortex is structurally and func-
tionally heterogeneous and contains subsets of at least two distinct brain networks (per esempio.,
Fedorenko et al., 2012; see Fedorenko & Blank, 2020, for a review). One of these is the net-
work of interest—the MD network, but the other is the language-selective network (per esempio., Braga
et al., 2020; Fedorenko et al., 2011; Fedorenko & Thompson-Schill, 2014), which does not
support executive functions. Given the well-documented interindividual variability in the pre-
cise locations of the MD and language areas (per esempio., Fedorenko et al., 2011, 2013; Shashidhara
et al., 2020), an anatomical location cannot be used to interpret an effect as arising within the
MD network vs. the language network.
Secondo, to the best of our knowledge, all prior work has relied on comparisons of group-
level activation maps. In such analyses, individual maps in each group are aligned in the
common brain space, and voxel-wise functional correspondence is assumed to hold across
participants, and the group-level maps for bilinguals and monolinguals are then compared.
Neurobiology of Language
649
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
Such analyses suffer from limited sensitivity and functional resolution (Nieto-Castañón &
Fedorenko, 2012) due to interindividual differences in the precise locations of the functional
regions (see Shashidhara et al., 2020, for evidence of such variability for the MD network in
particular). In cases of between-group comparisons, this variability can lead to misleading, E
even altogether opposite, patterns of results. Per esempio, imagine that the functional topog-
raphy is less variable in the monolingual population, leading to better alignment at the group
level. In this scenario, even if at the individual level, every bilingual individual shows stronger
effects than every monolingual individual, the group-level comparison will show a more pro-
nounced effect in the monolingual group, which is the opposite of the true effect.
Third, most prior neuroimaging studies of bilinguals have relied on small, and sometimes
heterogeneous, samples, which can lead to spurious effects driven by a small number of out-
liers (per esempio., Assem, Glasser, et al., 2020).
Finalmente, in order to ensure that an observed effect in the MD network is not due to a group-
level difference in variables that would affect responses across the brain, such as brain vascular-
ization (per esempio., Erdogan et al., 2016; He et al., 2010; Poldrack, 2011), motion (per esempio., Hajnal et al.,
1994; Power et al., 2015), vigilance levels (per esempio., Wong et al., 2013), or arousal (per esempio., Chang et al.,
2016; Schölvinck et al., 2010), it is important to demonstrate that any group difference observed
between bilinguals and monolinguals in the MD system is not present in some control brain
region, as supported by a region by group interaction (per esempio., Nieuwenhuis et al., 2011). To the
best of our knowledge, none of the past studies have included such control regions.
In an effort to bring clarity to the ongoing debate about whether or not bilingual individuals
have superior executive abilities, we carried out an fMRI investigation where we (io) localized
the network of interest (the MD network) in each individual participant using a well-
established paradigm (a spatial working memory task) that has been previously shown to acti-
vate the same areas as other diverse executive-function tasks (per esempio., Fedorenko et al., 2013;
Shashidhara et al., 2020) and to robustly isolate the MD network from the language network
(Blank et al., 2014; Fedorenko et al., 2012, 2013; Ivanova et al., 2020; Mineroff et al., 2018);
(ii) examined individual-level neural markers (magnitudes of response to the target task,
estimated using data independent from the data used to localize the regions of interest) Quello
have been shown to be stable within individuals over time and to correlate with behavioral
performance (Assem, Glasser, et al., 2020); (iii) included a control set of regions—primary
visual areas—to evaluate the spatial specificity of the effect; E (iv) examined a relatively
large (n = 55) and relatively homogeneous set of bilinguals (early balanced bilinguals who live
in an English-speaking country—the United States; see Figure 2a for details), matched carefully
to a similarly sized group of monolinguals (Vedi la tabella 1 for details).
MATERIALS AND METHODS
Participants
The study included 109 participants: 55 bilinguals and 54 monolinguals. Participant selection
proceeded as follows. Primo, 87 bilingual–monolingual pairs of participants were identified
Tavolo 1.
Summary of the variables for which the two groups were matched.
Group
Bilingual
Age mean (SD)
25.47 (4.87)
Monolingual
25.42 (5.81)
% Female
43.6%
48.1%
% Right-handed
81.8%
87.0%
Neurobiology of Language
650
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
among the 800+ participants in the Fedorenko Lab’s database, the majority of whom had com-
pleted the task of interest (the spatial working memory task). These pairs were selected so as to
be similar in age and have the same gender and handedness. Prossimo, 11 participants were
removed (6 bilingual, 5 monolingual) because they had completed only one run of the task
(two runs are necessary to estimate the response magnitudes in individually defined functional
regions of interest (fROIs); see below for details); E 14 additional participants were removed
(1 bilingual, 13 monolingual) due to data quality issues. These exclusions left 149 participants
(80 bilingual and 69 monolingual). Finalmente, following feedback from the reviewers, 40 addi-
tional participants were removed (25 bilingual, 15 monolingual) in order to ensure that (io) Tutto
bilingual participants learned their second language before the age of 6 and reported a pro-
ficiency score of 4 O 5 on a scale from 1 A 5 (see below for details), and that (ii) all mono-
lingual participants that reported having studied any foreign language in school did so after the
age of 10 and reported a proficiency score of 1 O 2. (See Supporting Information 1, which can
be found at https://doi.org/10.1162/nol_a_00058.) These exclusions left 109 participants (55
bilingual, 54 monolingual). In the final set, 32 of the original 87 pairs remained, with the other
45 participants not being pairwise matched. Tuttavia, the two groups remained well-matched
on age (p = 0.86), genere (p = 0.92), and handedness (p = 0.98; Vedi la tabella 1).
Participants in the bilingual group were native speakers of diverse languages (see Table SI-1
for detailed language profiles of all participants) and reported speaking two (n = 17), three (n =
24), or four or more (n = 14) languages. Crucially, as noted above, all participants acquired their
second language at an early age (mean = 2.14 years, SE = 0.30), and on a scale from 1 (no knowl-
edge) A 5 (native-like proficiency), they self-reported speaking their second language with high
proficiency (mean = 4.91, SE = 0.03; Figure 2a). The majority (n = 44) listed English as their
second language or as one of two languages acquired simultaneously from birth, while the rest
(n = 11) listed a different language as their second language (Table SI-1) and English as their third
lingua. Participants in the monolingual group were native English speakers; the majority did
not report having studied a second language (n = 35), and the rest (n = 19) reported learning a
second language at school and relatively late in life (mean = 13.84 years, SE = 0.62) and self-
reported a low proficiency level (mean = 1.8, SE = 0.04) (Figure 2a and Table SI-1).
Participants had normal or corrected-to-normal vision. All participants gave informed con-
sent as required by the Committee on the Use of Humans as Experimental Subjects (COUHES;
https://couhes.mit.edu/) and were paid for their participation.
Experimental Design
Every participant completed a spatial working memory task as part of a 2-hr fMRI scanning
session for one of the projects in the Fedorenko Lab. This task is routinely used in the lab
as a localizer for the domain-general MD system (Assem, Blank, et al., 2020; Duncan,
2010, 2013; Duncan et al., 2020; Fedorenko et al., 2013). In this task, participants are pre-
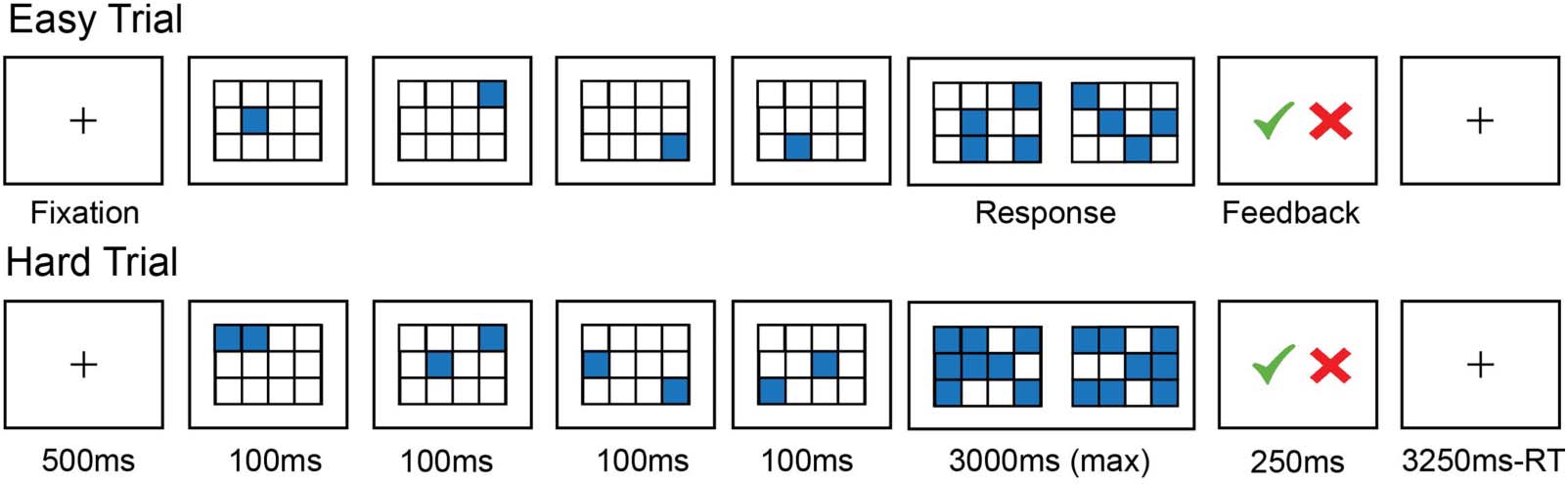
sented with a 3 × 4 grid, and on each trial, they see a sequence of locations flash up within the
grid. In the Easy condition, locations appear one at a time for a total of four locations, and in
the Hard condition, locations appear two at a time for a total of eight locations. After the
sequence, participants are presented with two grids showing two different sets of locations
and have to indicate which set of locations they had just seen. The grid with the incorrect
set of locations has one or two incorrect locations. Participants are given feedback on whether
they chose correctly in the form of a green checkmark or a red “X.” Each trial lasts 8 S (Vedere
Figura 1 for details of the timing), and trials are grouped into blocks of four. Each run consists of
twelve 32-s-long experimental blocks (six per condition) and four 16-s-long fixation blocks for
Neurobiology of Language
651
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
Figura 1. Sample trials of the Easy and Hard conditions of the spatial working memory task.
a total run duration of 448 S (7 min 28 S). All participants completed two runs (for a total task
duration of ∼15 min), with condition order counterbalanced across runs.
fMRI Data Acquisition
A whole-body 3 Tesla Siemens Trio scanner with a 32-channel head coil was used to collect
both structural and functional data. The structural images were collected with 1 mm isotropic
voxels (TR = 2,530 ms, TE = 3.48 ms) In 179 sagittal slices. An echo-planar imaging sequence
(flip angle: 90°, GRAPPA with 2 times acceleration factor) was used for the acquisition of func-
tional BOLD signal. The acquisition parameters were as follows: 31 4-mm thick near-axial
slices, in an interleaved order with a 10% distance factor; 2.1 mm × 2.1 mm in-plane resolu-
zione; field of view of 200 mm in the phase encoding anterior to posterior (A > P) direction;
matrix size of 96 × 96; TR of 2,000 ms; and TE of 30 ms. The gradient positioning based
on participant’s motion was adjusted using prospective acquisition correction. In order to
allow for the magnetization to become steady state, the first 10 s of each run were discarded.
fMRI Data Preprocessing and First-Level Analysis
fMRI data were analyzed using SPM12 (Wellcome Centre for Human Neuroimaging, UCL
Queen Square Institute of Neurology, London, UK; https://www.fil.ion.ucl.ac.uk/spm
/software/spm12/) and custom MATLAB (https://www.mathworks.com/) scripts. SPM was used
for preprocessing and first-level data modeling. Each participant’s data were motion corrected
and then normalized into a common brain space (the Montreal Neurological Institute (MNI)
template) and resampled into 2-mm isotropic voxels. The data were then smoothed with a
4-mm Gaussian filter and high-pass filtered (at 128 S). To model the spatial working memory
task, a standard mass univariate analysis was performed whereby a general linear model esti-
mated the effect size of each condition in each experimental run. These effects were each
modeled with a boxcar function (representing entire blocks) convolved with the canonical
hemodynamic response function. The model also included first-order temporal derivatives
of these effects, as well as nuisance regressors representing entire experimental runs, offline-
estimated motion parameters, and time points classified as outliers during the preprocessing
(cioè., scans where the scan-to-scan differences in global BOLD signal are above 5 standard
deviations, or where the scan-to-scan motion is above 0.9 mm).
MD fROI Definition and Response Estimation
For each participant, fROIs were defined using the Group-constrained subject-specific (GSS)
approach (Fedorenko et al., 2010). In this approach, a set of masks (or parcels) delineating
652
Group-constrained subject-specific
(GSS) approach:
An fMRI approach that enables
algorithmic definition of fROIs
in individual participants (Vedere
Fedorenko et al., 2010).
Neurobiology of Language
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
brain areas, within which most individuals in prior studies had shown activity for the localizer
contrasto, are combined with each individual participant’s activation map for the same contrast.
Here, for comparability with Fedorenko et al. (2013) and Assem, Glasser, et al. (2020), we
used a set of eighteen anatomical parcels; these covered the bilateral frontal and parietal brain
regions that have long been implicated in executive functions (per esempio., Cabeza & Nyberg, 2000;
Duncan, 2010, 2013) and included the middle frontal gyrus (MFG; 4,863 voxels LH, 5,104 vox-
els RH), the orbital part of the middle frontal gyrus (MFGorb; 888 voxels LH, 1,015 voxels RH),
the inferior frontal gyrus (IFG; 1,038 voxels LH, 1,399 voxels RH), the precentral gyrus (PrecG;
3,528 voxels LH, 3,381 voxels RH), the supplementary motor area (SMA; 2,147 voxels LH,
2,371 voxels RH), the anterior cingulate cortex (ACC; 1,615 voxels LH, 1,958 voxels RH), IL
superior parietal cortex (ParSup; 2,065 voxels LH, 2,222 voxels RH), the inferior parietal cortex
(ParInf; 2,447 voxels LH, 1,345 voxels RH), and the insula (1,858 voxels LH, 1,770 voxels RH).
The masks are available for download from: evlab.mit.edu/funcloc/.
For each individual participant, MD fROIs were defined by selecting 10% of voxels within
each parcel that were most responsive to the Hard > Easy spatial working memory contrast,
as defined by their t values. To estimate the responses of these fROIs to the Easy and Hard con-
ditions, an across-runs cross-validation procedure was used (Nieto-Castañón & Fedorenko,
2012): first, run 1 of the localizer was used to define the fROIs, and run 2 to estimate the responses
(in percent BOLD signal change) to the localizer condition, ensuring independence (Kriegeskorte
et al., 2009); second, run 2 was used to define the fROIs, and run 1 to estimate the responses;
finally, the extracted magnitudes were averaged across the two runs to derive a single response
magnitude per condition (hard and easy spatial working memory) per fROI per participant.
Control Regions
To test whether the group difference in response to the spatial working memory task that may
be observed within the MD network is present across the brain, we selected a set of control
brain regions. In particular, we used three bilateral anatomical parcels (from Tzourio-Mazoyer
et al., 2002) that cover primary visual areas. The rationale for selecting visual areas was that
they should show strong responses to the spatial working memory task given its visual nature,
so the comparison with the MD fROIs was fair. For this analysis, the responses to the Easy and
Hard conditions of the task were estimated across all voxels in each parcel in each participant,
and then averaged across the voxels in each parcel to obtain a single estimate per condition
per parcel per participant.
Statistical Analyses
The data were analyzed with linear mixed-effect models using the lme4 package in R
(https://cran.r-project.org/web/packages/lme4/index.html); p-value approximation was per-
formed with the lmerTest package, while effect sizes were calculated with the rstatix package
(Bates et al., 2015; Kuznetsova et al., 2017). The following linear mixed-effect regression
models were fit in order to address three critical research questions (all the analysis scripts
and the data tables are available at OSF [https://osf.io/b6xjy/]):
(UN) Does the MD network respond differentially in bilinguals and monolinguals during an
executive (spatial working memory) task?
The BOLD response was predicted by a model that included two fixed effects: condition
(Hard (relative to fixation), Easy (relative to fixation), and Hard > Easy) and group (bilingual
and monolingual). ROIs (n = 18) and participants (n = 109) were modeled as random
Neurobiology of Language
653
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
effects with random intercepts. ROIs were included as a random effect instead of a fixed effect
because, as discussed in the Introduction, the regions in the MD network have been previously
reported to be strongly functionally integrated, as evidenced by a high degree of
synchronization during naturalistic cognition (Assem, Blank, et al., 2020; Blank et al., 2014;
Braga et al., 2020; Paunov et al., 2019) and strong interregional correlations in effect sizes
(Assem, Glasser, et al., 2020; Mineroff et al., 2018). Tuttavia, for completeness, In
Supporting Information 2, we report models estimated for each ROI separately.
EffectSize ∼ Condition þ Group þ Group * Condition þ ð1 j ROI Þ þ ð1 j ParticipantÞ
(B) Do bilinguals perform better than monolinguals behaviorally on the spatial working
memory task?
The accuracy and reaction times on the spatial working memory task were predicted by two
separate models that included a fixed effect for group (bilingual and monolingual). Participants
(n = 65; 28 bilingual, 37 monolingual) were included as random effects with random
intercepts. (Note that the behavioral data for the remaining 44 participants (27 bilingual, 17
monolingual) were not collected due to experimenter error or equipment malfunction, or were
lost/overwritten.)
Accuracy or RT
ð
Þ ∼ Group þ ð1 j ParticipantÞ
(C) Do the control (primary visual) areas respond differentially in bilinguals and monolin-
guals during the spatial working memory task, and do the MD network and the primary
visual areas differ in their responses?
Primo, the BOLD response was predicted by a model that included two fixed effects: condition
(Hard (relative to fixation), Easy (relative to fixation), and Hard > Easy) and group (bilingual and
monolingual). ROIs (n = 18) and participants (n = 109) were modeled as random effects with
random intercepts.
EffectSize ∼ Condition þ Group þ Group * Condition þ ð1 j ROI Þ þ ð1 j ParticipantÞ
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
Prossimo, to explicitly test whether the MD network and the primary visual areas differ in their
responses between the two groups (per esempio., Nieuwenhuis et al., 2011), the BOLD response was
predicted by a model that included four fixed effects: condition (Hard (relative to fixation), Easy
(relative to fixation), and Hard > Easy), group (bilingual and monolingual), rete (MD and
Visual), and critically, a group by network interaction. ROIs (n = 18) and participants (n = 109)
were modeled as random effects with random intercepts.
EffectSize ∼ Condition þ Group þ Network þ Group * Network þð1 j ROIÞ þð1 j ParticipantÞ
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
RESULTS
As expected, and in line with previous research (Assem, Glasser, et al., 2020; Fedorenko et al.,
2013), the MD network showed a highly robust Hard > Easy effect across participants (b =
0.89, SE = 0.19; P < 0.001), and in each group separately (bilinguals: b = 0.96, SE = 0.19;
p < 0.001; monolinguals: b = 0.82, SE = 0.19; p < 0.001). The critical results were as follows.
(1) The MD network responded more strongly in bilinguals than in monolinguals during an
executive (spatial working memory) task.
A significant effect of group was observed: The MD fROIs responded more strongly in the
bilingual compared to the monolingual participants during both the Hard condition
Neurobiology of Language
654
Multiple demand network differences in bilinguals and monolinguals
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
n
o
_
a
_
0
0
0
5
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
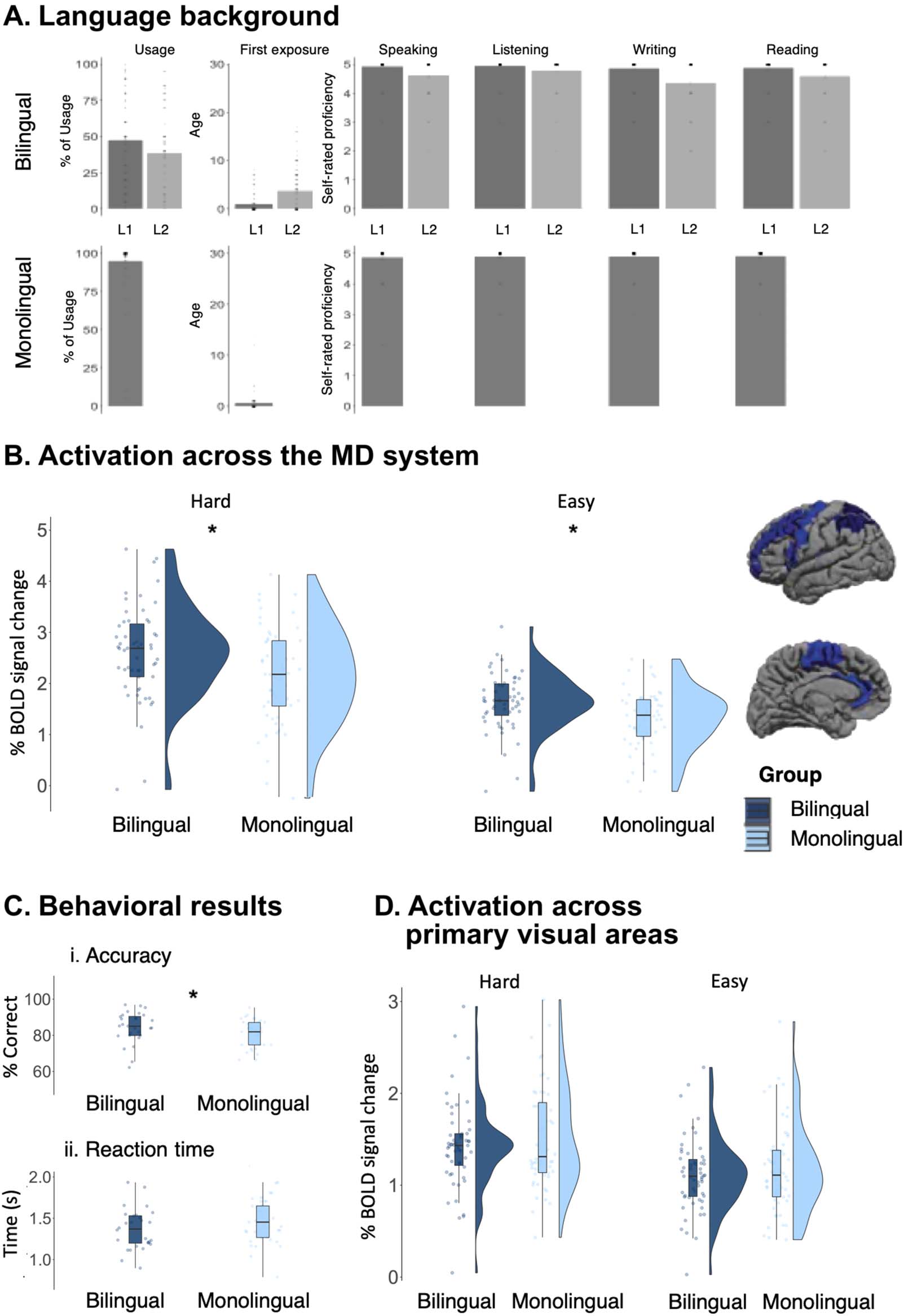
(A) The language background of bilingual and monolingual participants: usage of language (in %), age of first exposure, and
Figure 2.
self-rated proficiency scores from 1 (no knowledge) to 5 (native-like proficiency) for speaking, listening, writing, and reading are reported.
(B) Activation (in % BOLD signal change) across the MD system during the Hard and Easy conditions of the spatial working memory task.
(C) Accuracy and reaction times for the spatial working memory task. (D) Activation (in % BOLD signal change) across the primary visual areas
during the Hard and Easy conditions of the spatial working memory task.
Neurobiology of Language
655
Multiple demand network differences in bilinguals and monolinguals
(bilingual: mean = 2.62, SE = 0.02; monolingual: mean = 2.16, SE = 0.05; p < 0.01; Figure 2b)
and the Easy condition (bilingual: mean = 1.66, SE = 0.04; monolingual: mean = 1.34, SE =
0.04; p < 0.01). Further, the Hard > Easy effect was larger in the bilinguals (mean = 0.96, SE =
0.02) than in the monolinguals (mean = 0.82, SE = 0.02; P < 0.001).
(2) Bilinguals performed better than monolinguals behaviorally on the spatial working
memory task.
The bilinguals’ accuracies were higher (mean = 84.8%, SE = 1.64) than the monolinguals’
(mean = 79.3%, SE = 1.94; p = 0.03). Moreover, bilingual participants were numerically
faster (mean = 1.33 s, SE = 0.04) than monolingual participants (mean = 1.42 s, SE = 0.04;
p = 0.16). Both effects were small (Cohen’s d = 0.48 and −0.31, respectively), so we may
not have had sufficient power to detect an effect in the reaction time data.
(3) The primary visual areas responded similarly in bilinguals and monolinguals during
the spatial working memory task, and the MD network and the primary visual areas
differed in their responses.
Similar to the MD network, the primary visual areas showed a robust Hard > Easy effect across
participants (b = 0.33, SE = 0.08; P < 0.001), and in each group separately (bilinguals: b = 0.32,
SE = 0.09; p < 0.01; monolinguals: b = 0.33, SE = 0.09; p < 0.01). This is to be expected given that
the Hard condition contains more visual information (two squares, compared to one square, for
each trial component; see Figure 1). Critically, the primary visual areas of the bilingual
participants responded similarly to those of the monolingual participants during the Hard
condition (bilingual: mean = 1.41, SE = 0.09; monolingual: mean = 1.50, SE = 0.11; p = 0.78)
and the Easy condition (bilingual: mean = 1.10, SE = 0.08; monolingual: mean = 1.17, SE = 0.10;
p = 0.35). Further, the size of the Hard > Easy contrast was similar between the groups (bilingual:
mean = 0.32, SE = 0.03; monolingual: mean = 0.33, SE = 0.04; p = 0.32). Inoltre, a significant
group by network interaction obtained (b = 0.36, SE = 0.04; P < 0.001), such that the bilingual vs.
monolingual difference in the size of the Hard > Easy effect was reliably larger in the MD
network compared to the primary visual areas.
DISCUSSION
To address the question of whether bilingual individuals exhibit superior executive abilities—an
area of investigation characterized by a complex and controversial empirical landscape—we
carried out an fMRI investigation that had several unique design features, relative to the past
studies, that gave it a stronger chance to obtain a clearer answer, as elaborated in the Introduc-
tion and below. We found stronger responses to the spatial working memory task, and a larger
difference between a harder and an easier condition, in the bilingual participants compared to
the monolingual participants in the MD network, but not in the control (primary visual) areas.
This neural difference was accompanied by numerically better behavioral performance. Nel
remainder of the discussion, we highlight a few implications of these results and some questions
they raise, and talk about the limitations of the current investigation.
The Nature of the Superior Executive Abilities in Bilingual Individuals
We report effects that are reliable and substantial in size such that the bilinguals’ MD network
was overall more active during an executive (spatial working memory) task than the MD net-
work in age-, gender-, and handedness-matched monolingual participants, and the difference
Neurobiology of Language
656
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
between the harder and the easier conditions of the task was larger in bilinguals than in mono-
linguals. What do these effects reflect? Historically, behavioral and neural differences in exec-
utive abilities between bilinguals and monolinguals have been attributed to differences in their
linguistic experiences: in particular, to the bilinguals’ need to switch between languages, E
consequent improvements in their general cognitive control abilities. Tuttavia, attributing
these effects specifically to the differences between the two groups in their linguistic experience
is difficult. (This interpretive challenge is also present in all prior studies, even if they have not
explicitly acknowledged it.) In particular, bilingual individuals (or at least the type of bilinguals
examined here: most individuals are living in a country where their second language is the dom-
inant language, the majority of whom came to the United States to pursue educational and/or
professional goals) may be more motivated (Baker, 1988) and/or have higher grit (e.g. Credé
et al., 2017). The relationship between such factors and executive abilities remains controversial
(per esempio., Ebbesen, 2020; Pessoa, 2009; Taylor et al., 2004; Uddin, 2021); whereas some studies con-
trol for education and immigration status (Alladi et al., 2013), at present, it is not possible to rule
out the potential contribution of such factors to the effects observed here.
General fluid intelligence is also worth a mention: We only had IQ scores on a small subset
of our participants, so we could not match the groups for IQ. Tuttavia, it is actually unclear
whether matching on IQ makes sense in evaluating individual differences in executive abilities
given the intimate link between fluid intelligence and executive functions (per esempio., Assem, Blank,
et al., 2020; Duncan, 2010, 2013; Duncan et al., 2020; Gläscher et al., 2010; Woolgar et al.,
2010). Infatti, damage to the MD network has been shown to lead to deficits in executive
functions as well as to loss of fluid intelligence abilities (see Duncan, 2020, for an extensive
discussion), and stronger responses in the MD network have been associated with both better
performance on executive tasks and higher IQ scores (per esempio., Assem, Glasser, et al., 2020; Basten
et al., 2013; Burgess et al., 2011; Choi et al., 2008; Cole et al., 2012; Gray et al., 2003; Lee
et al., 2006; Tschentscher & Mitchell, 2017).
These interpretive challenges call for further studies across diverse bilingual populations. If
these effects hold across different kinds of bilinguals, that would help rule out potential expla-
nations in terms of motivation/grit, or establish that superior executive abilities characterize
only some bilingual/multilingual populations (per esempio., Blanco-Elorrieta & Pylkkänen, 2018). For
esempio, it is worth noting that using the same paradigm as the one used here, Jouravlev et al.
(2021) found no difference in the neural responses in the MD network in a set of 17 polyglots
and hyperpolyglots, most of whom acquired their non-native languages post critical period, COME
compared to a matched set of monolingual controls. Tuttavia, in Jouravlev et al.’s study, poly-
glots and monolinguals were matched for IQ, which as noted above, may not be the right
approach when probing for individual differences in executive functions.
To conclusively link superior executive abilities to linguistic experience, longitudinal devel-
opmental studies will be critical. In particular, tracking executive abilities in a population of
young monolingual children some of whom proceed to acquire a second language (per esempio.,
through a language immersion program) and some of whom do not would be extremely valu-
able. Ovviamente, longitudinal studies are notoriously challenging, and full experimental control
over which subset of children become bilingual may be hard or impossible to achieve.
Methodological Considerations in Future Studies of Bilingualism
Several unique features of the current study may have enabled us to detect a clear and robust
effect, and we hope some of these practices will become more widely adopted in the field of
bilingualism research. Perhaps most importantly, we identified the network of interest (the MD
Neurobiology of Language
657
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
rete) functionally in each individual participant using a robust MD localizer paradigm.
There are three key advantages to this approach. Primo, functional localization has long been
established to vastly improve sensitivity (cioè., the ability to detect an effect; per esempio., Brett et al.,
2002; Fedorenko et al., 2010; Nieto-Castañón & Fedorenko, 2012; Saxe et al., 2006). This issue
is especially pertinent when examining high-level cognitive processes. Such processes are
supported by the association cortex, where functional areas (io) are not predictable from macro-
anatomy (per esempio., Frost & Goebel, 2012; Tahmasebi et al., 2012; Vázquez-Rodríguez et al., 2019),
E (ii) vary substantially across individuals in their precise locations in a common brain space
(per esempio., Fedorenko et al., 2010, 2013; Shashidhara et al., 2020). An inevitable consequence is that
many effects may be robustly present in each individual participant but would be missed in a stan-
dard group analysis, which relies on voxelwise alignment across individuals (note that the use of
larger samples does not help with this problem). The use of this low-power analytic approach may
explain why prior studies have reported effects in only a subset of the MD network. (Incidentally,
arguments that only regions where an effect emerged in a traditional group analysis, but not other
regions, show the effect of interest are fallacious for the reasons above. In particular, region A but
not region B may emerge in a group analysis because region A is better aligned with anatomic
landmarks; Vedere, per esempio., Blank et al., 2016, for discussion.) The use of this approach may also
obscure between-population differences.
Secondo, functional localization confers a substantial interpretive advantage, removing the
need for precarious reverse inference (per esempio., Fedorenko, 2021; Poldrack, 2006). In particular,
by functionally identifying a network that has been robustly linked to executive functions across
diverse tasks (per esempio., Fedorenko et al., 2013; Hugdahl et al., 2015; Shashidhara et al., 2020) IL
observed effects can be straightforwardly interpreted as reflecting differences in executive func-
zioni. Because the cortex is highly functionally heterogeneous, and distinct areas often lay adja-
cent to one another within the same macroanatomic area, interpreting effects functionally based
on coarse macroanatomy is not justified. Per esempio, effects within the left IFG are sometimes
interpreted as reflecting the engagement of executive resources (per esempio., Garbin et al., 2010), E
other times as reflecting the engagement of linguistic resources (per esempio., Rodríguez-Pujadas et al.,
2013). Such flexibility in interpretation is clearly undesirable. Functional localization helps to
unambiguously identify the MD vs. the language-selective portions of the left IFG (Fedorenko &
Blank, 2020). The same holds for other areas of the association cortex, most of which are highly
heterogeneous, containing numerous distinct areas in close proximity to one another.
And third, the use of the same functional localizer paradigms across individuals, studies,
and labs enables the establishment of a cumulative research enterprise—the cornerstone of
robust and replicable science. This general approach has been de rigueur in other fields, like
vision (per esempio., Kanwisher et al., 1997) from the earliest days of brain imaging research, and more
recently, social cognition (per esempio., Saxe & Kanwisher, 2003) and language (Fedorenko et al.,
2010). Adopting this approach in the study of executive functions in bilingualism is likely to
lead to greater clarity and consensus because of the greater ease of comparing and replicating
findings across studies.
Another important feature of our study, which was not present in any prior study, is the use
of neural markers that have been previously established (UN) to be stable within individuals, (B)
to vary across individuals, E (C) to relate to behavioral performance (Assem, Glasser, et al.,
2020). This is critically important: A study that does not find a difference between bilinguals
and monolinguals is impossible to interpret if the relevant neural marker has not been shown
to have these properties.
Finalmente, when arguing for a neural difference between two groups in a particular brain
region or network, it is critical to establish the spatial selectivity of the effect. In particular,
Neurobiology of Language
658
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
some effects may be ubiquitously present across the brain and result from nonspecific differ-
enze, Per esempio, in the degree of vascularization or arousal. To rule out such effects, we
examined a control set of brain areas that respond to the task but are not part of the MD net-
lavoro (primary visual areas). Such control areas have typically been absent from past studies
and would be valuable to include in future work.
Limitations of scope
Although our study had several methodological advantages over much prior work, it remains a
single study probing a particular population of bilinguals: balanced early bilinguals currently
residing in the United States. The observation of superior executive abilities in this particular
bilingual population is consistent with, but does not directly evaluate, the hypothesis laid out
in Blanco-Elorrieta and Pylkkänen (2018). It would help move the field forward if future studies
(UN) focused on relatively homogeneous groups of bilinguals (per esempio., Costa & Santesteban, 2004;
Rossi et al., 2017), and/or (B) provided a detailed characterization of their language back-
ground and use patterns (de Bruin, 2019).
Conclusione
In conclusion, we report the first investigation of executive abilities in early bilinguals and
matched monolinguals using the kind of robust individual-subject functional localization ana-
lytic approach that is likely to yield more interpretable and more easily replicable results than
those obtained in past work. We hope that the field of bilingualism research adopts at least
some aspects of the approach advocated here, so as to lead to a more robust and cumulative
research enterprise.
DEDICATION
We would like to dedicate this paper to the memory of Albert Costa, who we both knew well
and loved as a mentor and a friend. Saima will always be grateful that Albert let her spend her
senior year in his lab despite not even being from the same university; his support, mentorship,
and guidance helped her not stray away from academia when things got tough. And Ev will
forever remember the weekly Friday night partying with Albert and the rest of the “crew” in
The Cellar and The People’s Republik during her undergrad years in the Caramazza Lab in the
late 1990s and early 2000s.
ACKNOWLEDGMENTS
We would like to acknowledge the Athinoula A. Martinos Imaging Center at the McGovern
Institute for Brain Research at MIT, and its support team (Steve Shannon and Atsushi Takahashi).
We thank former and current EvLab members for their help with fMRI data collection
(especially Dima Ayyash and Olessia Jouravlev). We also thank Rachel Ryskin and Ted
Gibson for helpful discussions. Saima Malik-Moraleda was supported by la Caixa Fellowship
LCF/BQ/AA17/11610043. Evelina Fedorenko was supported by the R00 award HD057522,
R01 awards DC016607 and DC016950 from NIH, and funds from the Brain and Cognitive
Sciences department and the McGovern Institute for Brain Research.
FUNDING INFORMATION
Saima Malik-Moraleda, “la Caixa” Foundation (https://dx.doi.org/10.13039/100010434),
Award ID: LCF/BQ/AA17/11610043. Evelina Fedorenko, National Institutes of Health (https://
dx.doi.org/10.13039/100000002), Award ID: HD057522. Evelina Fedorenko, National
Neurobiology of Language
659
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID: DC016607. Evelina
Fedorenko, National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID:
DC016950.
AUTHOR CONTRIBUTIONS
Saima Malik-Moraleda: Data curation: Lead; Formal analysis: Lead; Investigation: Lead;
Methodology: Lead; Validation: Lead; Visualization: Lead; Writing – original draft: Lead;
Writing – review & editing: Lead. Theodor Cucu: Data curation: Supporting; Formal analysis:
Supporting; Investigation: Supporting; Writing – review & editing: Supporting. Benjamin
Lipkin: Conceptualization: Supporting; Formal analysis: Supporting; Investigation: Supporting;
Validation: Equal; Visualization: Supporting; Writing – review & editing: Supporting. Evelina
Fedorenko: Formal analysis: Equal; Funding acquisition: Lead; Investigation: Supporting;
Methodology: Equal; Project administration: Lead; Resources: Lead; Supervision: Lead;
Writing – original draft: Equal; Writing – review & editing: Equal.
REFERENCES
Abutalebi, J., Della Rosa, P. A., Ding, G., Weekes, B., Costa, A., &
Verde, D. W. (2013). Language proficiency modulates the
engagement of cognitive control areas in multilinguals. Cortex,
49(3), 905–911. https://doi.org/10.1016/j.cortex.2012.08.018,
PubMed: 23021069
Abutalebi, J., Della Rosa, P. A., Verde, D. W., Hernandez, M.,
Scifo, P., Keim, R., Cappa, S. F., & Costa, UN. (2012). Bilingualism
tunes the anterior cingulate cortex for conflict monitoring. Cere-
bral Cortex, 22(9), 2076–2086. https://doi.org/10.1093/cercor
/bhr287, PubMed: 22038906
Abutalebi, J., & Verde, D. [W.] (2007). Bilingual language produc-
zione: The neurocognition of language representation and control.
Journal of Neurolinguistics, 20(3), 242–275. https://doi.org/10
.1016/j.jneuroling.2006.10.003
Abutalebi, J., & Verde, D. W. (2016). Neuroimaging of language
control in bilinguals: Neural adaptation and reserve. Bilingual-
ism: Language and Cognition, 19(4), 689–698. https://doi.org
/10.1017/S1366728916000225
Alladi, S., Bak, T. H., Duggirala, V., Surampudi, B., Shailaja, M.,
Shukla, UN. K., Chaudhuri, J. R., & Kaul, S. (2013). Bilingualism
delays age at onset of dementia, independent of education and
immigration status. Neurology, 81(22), 1938–1944. https://doi
.org/10.1212/01.wnl.0000436620.33155.a4, PubMed:
24198291
Antón, E., Duñabeitia, J. A., Estévez, A., Hernández, J. A., Castillo,
A., Fuentes, l. J., Davidson, D. J., & Carreiras, M. (2014). Is there
a bilingual advantage in the ANT task? Evidence from children.
Frontiers in Psychology, 5, Article 398. https://doi.org/10.3389
/fpsyg.2014.00398, PubMed: 24847298
Antoniou, M., & Wright, S. M. (2017). Uncovering the mechanisms
responsible for why language learning may promote healthy cog-
nitive aging. Frontiers in Psychology, 8, Article 2217. https://doi
.org/10.3389/fpsyg.2017.02217, PubMed: 29326636
Assem, M., Blank, IO. A., Mineroff, Z. A., Ademoğlu, A., & Fedorenko,
E. (2020). Activity in the fronto-parietal multiple-demand network
robustly associated with individual differences in working
memory and fluid intelligence. Cortex, 131, 1–16. https://doi.org
/10.1016/j.cortex.2020.06.013, PubMed: 32777623
Assem, M., Glasser, M. F., Van Essen, D. C., & Duncan, J. (2020).
A domain-general cognitive core defined in multimodally
parcellated human cortex. Cerebral Cortex, 30(8), 4361–4380.
https://doi.org/10.1093/cercor/bhaa023, PubMed: 32244253
Bacon-Shone, J., & Bolton, K. (1998). Charting multilingualism:
Language censuses and language surveys in Hong Kong. In
M. C. Pennington (Ed.), Language in Hong Kong at century’s
end (pag. 43–90). Hong Kong University Press.
Baker, C. (1988). Key issues in bilingualism and bilingual educa-
zione. Multilingual Matters.
Basten, U., Stelzel, C., & Fiebach, C. J. (2013). Intelligence is dif-
ferentially related to neural effort in the task-positive and the
task-negative brain network. Intelligenza, 41(5), 517–528.
https://doi.org/10.1016/j.intell.2013.07.006
Bates, D., Mächler, M., Bolker, B. M., & Walker, S. C. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical Soft-
ware, 67(1), 1–48. https://doi.org/10.18637/jss.v067.i01
Bialystok, E. (1999). Cognitive complexity and attentional control
in the bilingual mind. Child Development, 70(3), 636–644.
https://doi.org/10.1111/1467-8624.00046
Bialystok, E. (2017). The bilingual adaptation: How minds accom-
modate experience. Psychological Bulletin, 143(3), 233–262.
https://doi.org/10.1037/bul0000099, PubMed: 28230411
Bialystok, E., Martin, M. M., & Viswanathan, M. (2005). Bilingual-
ism across the lifespan: The rise and fall of inhibitory control.
International Journal of Bilingualism, 9(1), 103–119. https://doi
.org/10.1177/13670069050090010701
Blanco-Elorrieta, E., & Pylkkänen, l. (2018). Ecological validity in
bilingualism research and the bilingual advantage. Trends in
Cognitive Sciences, 22(12), 1117–1126. https://doi.org/10.1016
/j.tics.2018.10.001, PubMed: 30449317
Blank, I., Balewski, Z., Mahowald, K., & Fedorenko, E. (2016).
Syntactic processing is distributed across the language system.
NeuroImage, 127, 307–323. https://doi.org/10.1016/j
.neuroimage.2015.11.069, PubMed: 26666896
Blank, I., Kanwisher, N., & Fedorenko, E. (2014). A functional dis-
sociation between language and multiple-demand systems
revealed in patterns of BOLD signal fluctuations. Journal of Neu-
rophysiology, 112(5), 1105–1118. https://doi.org/10.1152/jn
.00884.2013, PubMed: 24872535
Braga, R. M., DiNicola, l. M., Becker, H. C., & Buckner, R. l.
(2020). Situating the left-lateralized language network in the
Neurobiology of Language
660
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
broader organization of multiple specialized large-scale distrib-
uted networks. Journal of Neurophysiology, 124(5), 1415–1448.
https://doi.org/10.1152/JN.00753.2019, PubMed: 32965153
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002, Giugno
2–6). Region of interest analysis using an SPM toolbox [Paper
presentation]. The 8th International Conference on Functional
Mapping of the Human Brain, Sendai, Japan.
Burgess, G. C., Gray, J. R., Conway, UN. R. A., & Braver, T. S. (2011).
Neural mechanisms of interference control underlie the relation-
ship between fluid intelligence and working memory span. Jour-
nal of Experimental Psychology: General, 140(4), 674–692.
https://doi.org/10.1037/a0024695, PubMed: 21787103
Cabeza, R., & Nyberg, l. (2000). Imaging cognition II: An empirical
review of 275 PET and fMRI studies. Journal of Cognitive
N e u ro s c i e n c e , 1 2 ( 1 ) , 1 – 4 7 . h t t p s : / / d o i . o r g / 1 0 . 11 6 2
/08989290051137585, PubMed: 10769304
Chang, C., Leopold, D., Schölvinck, M., Mandelkow, H., Picchioni,
D., Liu, X., Ye, F., Turchi, J., & Duyn, J. (2016). Tracking brain
arousal fluctuations with fMRI. Proceedings of the National
Academy of Sciences, 113(16), 4518–4523. https://doi.org/10
.1073/pnas.1520613113, PubMed: 27051064
Choi, Y. Y., Shamosh, N. A., Cho, S. H., DeYoung, C. G., Lee, M. J.,
Lee, J.-M., Kim, S. I., Cho, Z.-H., Kim, K., Gray, J. R., & Lee, K. H.
(2008). Multiple bases of human intelligence revealed by cortical
thickness and neural activation. Journal of Neuroscience, 28(41),
10323–10329. https://doi.org/10.1523/ JNEUROSCI.3259-08
.2008, PubMed: 18842891
Cole, M. W., Yarkoni, T., Repovš, G., Anticevic, A., & Braver, T. S.
(2012). Global connectivity of prefrontal cortex predicts cogni-
tive control and intelligence. Journal of Neuroscience, 32(26),
8988–8999. https://doi.org/10.1523/JNEUROSCI.0536-12.2012,
PubMed: 22745498
Costa, A., & Santesteban, M. (2004). Lexical access in bilingual
speech production: Evidence from language switching in highly
proficient bilinguals and L2 learners. Journal of Memory and
Language, 50(4), 491–511. https://doi.org/10.1016/j.jml.2004
.02.002
Costa, A., & Sebastián-Gallés, N. (2014). How does the bilingual
experience sculpt the brain? Nature Reviews Neuroscience,
15(5), 336–345. https://doi.org/10.1038/nrn3709, PubMed:
24739788
Craik, F. IO. M., Bialystok, E., & Freedman, M. (2010). Delaying the
onset of Alzheimer disease: Bilingualism as a form of cognitive
reserve. Neurology, 75(19), 1726–1729. https://doi.org/10.1212
/ WNL.0b013e3181fc2a1c, PubMed: 21060095
Credé, M., Tynan, M. C., & Harms, P. D. (2017). Much ado about
grit: A meta-analytic synthesis of the grit literature. Journal of Per-
sonality and Social Psychology, 113(3), 492–511. https://doi.org
/10.1037/pspp0000102, PubMed: 27845531
de Bruin, UN. (2019). Not all bilinguals are the same: A call for more
detailed assessments and descriptions of bilingual experiences.
Behavioral Sciences, 9(3), Article 33. https://doi.org/10.3390
/bs9030033, PubMed: 30909639
de Bruin, A., Treccani, B., & Della Sala, S. (2015). Cognitive advan-
tage in bilingualism: An example of publication bias? Psycholog-
i c a l S c i e n c e , 2 6 ( 1 ) , 9 9 – 1 0 7 . h t t p s : / / d o i . o rg / 1 0 . 11 7 7
/0956797614557866, PubMed: 25475825
Declerck, M., & Philipp, UN. M. (2015). A review of control pro-
cesses and their locus in language switching. Psychonomic Bul-
letin and Review, 22(6), 1630–1645. https://doi.org/10.3758
/s13423-015-0836-1, PubMed: 25917142
Del Maschio, N., & Abutalebi, J. (2019). Language organization in
the bilingual and multilingual brain. In J. W. Schwieter & M.
Paradis (Eds.), The handbook of the neuroscience of multilingual-
ism (pag. 199–213). John Wiley & Sons Ltd. https://doi.org/10
.1002/9781119387725.ch9
DeLuca, V., Rothman, J., Bialystok, E., & Pliatsikas, C. (2019).
Redefining bilingualism as a spectrum of experiences that differ-
entially affects brain structure and function. Atti del
National Academy of Sciences, 116(15), 7565–7574. https://doi
.org/10.1073/pnas.1811513116, PubMed: 30914463
Duñabeitia, J. A., Hernández, J. A., Antón, E., Macizo, P., Estévez,
A., Fuentes, l. J., & Carreiras, M. (2014). The inhibitory advan-
tage in bilingual children revisited: Myth or reality? Experimental
Psychology, 61, 234–251. https://doi.org/10.1027/1618-3169
/a000243, PubMed: 24217139
Duncan, J. (2010). The multiple-demand (MD) system of the pri-
mate brain: Mental programs for intelligent behaviour. Trends
in Cognitive Sciences, 14(4), 172–179. https://doi.org/10.1016/j
.tics.2010.01.004, PubMed: 20171926
Duncan, J. (2013). The structure of cognition: Attentional episodes
in mind and brain. Neuron, 80(1), 35–50. https://doi.org/10.1016
/j.neuron.2013.09.015, PubMed: 24094101
Duncan, J. (2020). How intelligence happens. Stampa dell'Università di Yale.
Duncan, J., Assem, M., & Shashidhara, S. (2020). Integrated intelli-
gence from distributed brain activity. Trends in Cognitive Sci-
enze, 24(10), 838–852. https://doi.org/10.1016/j.tics.2020.06
.012, PubMed: 32771330
Duncan, J., & Owen, UN. (2000). Common regions of the human
frontal lobe recruited by diverse cognitive demands. Trends in
Cognitive Sciences, 23(10), 475–483. https://doi.org/10.1016
/S0166-2236(00)01633-7
Ebbesen, C. l. (2020). Flawed estimates of cognitive ability in Clark
et al. Psychological Science, 2020. PsyArXiv. https://doi.org/10
.31234/osf.io/tzr8c
Erdogan, S., Tong, Y., Hocke, L., Lindsey, K., & Frederick, B d.
(2016). Correcting for blood arrival time in global mean regres-
sion enhances functional connectivity analysis of resting state
fMRI-BOLD signals. Frontiers in Human Neuroscience, 10,
Article 311. https://doi.org/10.3389/fnhum.2016.00311,
PubMed: 27445751
Fedorenko, E. (2021). The early origins and the growing popularity
of the individual-subject analytic approach in human neurosci-
ence. Current Opinion in Behavioral Sciences, 40, 105–112.
https://doi.org/10.1016/j.cobeha.2021.02.023
Fedorenko, E., Behr, M. K., & Kanwisher, N. (2011). Functional
specificity for high-level linguistic processing in the human brain.
Proceedings of the National Academy of Sciences, 108(39),
16428–16433. https://doi.org/10.1073/pnas.1112937108,
PubMed: 21885736
Fedorenko, E., & Blank, IO. UN. (2020). Broca’s area is not a natural
kind. Trends in Cognitive Sciences, 24(4), 270–284. https://doi
.org/10.1016/j.tics.2020.01.001, PubMed: 32160565
Fedorenko, E., Duncan, J., & Kanwisher, N. (2012). Language-
selective and domain-general regions lie side by side within Bro-
ca’s area. Current Biology, 22(21), 2059–2062. https://doi.org/10
.1016/j.cub.2012.09.011, PubMed: 23063434
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad domain
generality in focal regions of frontal and parietal cortex. Proceedings
of the National Academy of Sciences, 110(41), 16616–16621.
https://doi.org/10.1073/pnas.1315235110, PubMed: 24062451
Fedorenko, E., Hsieh, P.-J., Nieto-Castañón, A., Whitfield-Gabrieli,
S., & Kanwisher, N. (2010). New method for fMRI investigations
of language: Defining ROIs functionally in individual subjects.
Journal of Neurophysiology, 104(2), 1177–1194. https://doi.org
/10.1152/jn.00032.2010, PubMed: 20410363
Neurobiology of Language
661
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
Fedorenko, E., & Thompson-Schill, S. l. (2014). Reworking the lan-
guage network. Trends in Cognitive Sciences, 18(3), 120–126.
https://doi.org/10.1016/j.tics.2013.12.006, PubMed: 24440115
Frost, M., & Goebel, R. (2012). Measuring structural–functional cor-
respondence: Spatial variability of specialised brain regions after
macro-anatomical alignment. NeuroImage, 59(2), 1369–1381.
https://doi.org/10.1016/j.neuroimage.2011.08.035, PubMed:
21875671
Gallo, F., Novitskiy, N., Myachykov, A., & Shtyrov, Y. (2021). Indi-
vidual differences in bilingual experience modulate executive
control network and performance: Behavioral and structural
neuroimaging evidence. Bilingualism: Language and Cognition,
24(2), 293–304. https://doi.org/10.1017/S1366728920000486
Garbin, G., Sanjuan, A., Forn, C., Bustamante, J. C., Rodriguez-
Pujadas, A., Belloch, V., Hernandez, M., Costa, A., & Ávila, C.
(2010). Bridging language and attention: Brain basis of the
impact of bilingualism on cognitive control. NeuroImage,
53(4), 1272–1278. https://doi.org/10.1016/j.neuroimage.2010
.05.078, PubMed: 20558314
García-Pentón, L., Fernández García, Y., Costello, B., Duñabeitia,
J. A., & Carreiras, M. (2016). The neuroanatomy of bilingualism:
How to turn a hazy view into the full picture. Language, Cogni-
tion and Neuroscience, 31(3), 303–327. https://doi.org/10.1080
/23273798.2015.1068944
Gläscher, J., Rudrauf, D., Colom, R., Paul, l. K., Tranel, D.,
Damasio, H., & Adolphs, R. (2010). Distributed neural system
for general intelligence revealed by lesion mapping. Proceedings
of the National Academy of Sciences, 107(10), 4705–4709.
https://doi.org/10.1073/pnas.0910397107, PubMed: 20176936
Gold, B. T., Kim, C., Johnson, N. F., Kryscio, R. J., & Smith, C. D.
(2013). Lifelong bilingualism maintains neural efficiency for cog-
nitive control in aging. Journal of Neuroscience, 33(2), 387–396.
https://doi.org/10.1523/ JNEUROSCI.3837-12.2013, PubMed:
23303919
Gray, J. R., Chabris, C. F., & Braver, T. S. (2003). Neural mecha-
nisms of general fluid intelligence. Nature Neuroscience, 6(3),
316–322. https://doi.org/10.1038/nn1014, PubMed: 12592404
Grundy, J. G., & Timmer, K. (2017). Bilingualism and working
memory capacity: A comprehensive meta-analysis. Secondo
Language Research, 33(3), 325–340. https://doi.org/10.1177
/0267658316678286
Guzmán-Vélez, E., Warren, D. E., Feinstein, J. S., Bruss, J., & Tranel,
D. (2016). Dissociable contributions of amygdala and hippocam-
pus to emotion and memory in patients with Alzheimer’s disease.
Hippocampus, 26(6), 727–738. https://doi.org/10.1002/ hipo
.22554, PubMed: 26606553
Hajnal, J., Myers, R., Oatridge, A., Schwieso, J., Young, I., &
Bydder, G. (1994). Artifacts due to stimulus correlated motion
in functional imaging of the brain. Magnetic Resonance in Med-
i c i n e , 3 1 ( 3 ) , 2 8 3 – 2 9 1 . h t t p s : / / d o i . o rg / 1 0 . 1 0 0 2 / m r m
.1910310307, PubMed: 8057799
Lui, H., Shin, D., & Liu, T. T. (2010). Resting state BOLD fluctua-
tions in large draining veins are highly correlated with the global
mean signal. In Proceedings of the 18th annual meeting of the
ISMRM (P. 3488). ISMRM.
Hoffmann, C. (2000). The spread of English and the growth of
multilingualism with English in Europe. In J. Cenoz & U. Jessner
(Eds.), English in Europe: The acquisition of a third language
(pag. 1–21). Multilingual Matters.
Hugdahl, K., Raichle, M. E., Mitra, A., & Specht, K. (2015). On the
existence of a generalized non-specific task-dependent network.
Frontiers in Human Neuroscience, 9, Article 430. https://doi.org
/10.3389/fnhum.2015.00430, PubMed: 26300757
Ivanova, UN. A., Srikant, S., Sueoka, Y., Kean, H. H., Dhamala, R.,
O’Reilly, U. M., Bers, M. U., & Fedorenko, E. (2020). Compre-
hension of computer code relies primarily on domain-general
executive brain regions. ELife, 9, Article 58906. https://doi.org
/10.7554/eLife.58906, PubMed: 33319744
Jouravlev, O., Mineroff, Z., Blank, I., & Fedorenko, E. (2021). IL
small and efficient language network of polyglots and
hyper-polyglots. Cerebral Cortex, 31(1), 62–76. https://doi.org
/10.1093/cercor/bhaa205, PubMed: 32820332
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The fusiform
face area: A module in human extrastriate cortex specialized for
face perception. Journal of Neuroscience, 17(11), 4302–4311.
https://doi.org/10.1523/jneurosci.17-11-04302.1997, PubMed:
9151747
Kapa, l. L., & Colombo, J. (2013). Attentional control in early and
later bilingual children. Cognitive Development, 28(3), 233–246.
https://doi.org/10.1016/j.cogdev.2013.01.011, PubMed:
24910499
Kaushanskaya, M., & Marian, V. (2007). Bilingual language pro-
cessing and interference in bilinguals: Evidence from eye track-
ing and picture naming. Language Learning, 57(1), 119–163.
https://doi.org/10.1111/j.1467-9922.2007.00401.x
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S. F., & Baker, C. IO.
(2009). Circular analysis in systems neuroscience: The dangers of
double dipping. Nature Neuroscience, 12, 535–540. https://doi
.org/10.1038/nn.2303, PubMed: 19396166
Kroll, J. F., Bobb, S. C., & Hoshino, N. (2014). Two languages in mind:
Bilingualism as a tool to investigate language, cognition, and the
brain. Current Directions in Psychological Science, 23(3), 159–163.
https://doi.org/10.1177/0963721414528511, PubMed: 25309055
Kroll, J. F., & Dussias, P. E. (2017). The benefits of multilingualism to
the personal and professional development of residents of the
US. Foreign Language Annals, 50(2), 248–259. https://doi.org
/10.1111/flan.12271, PubMed: 29097822
Kroll, J. F., Dussias, P. E., Bice, K., & Perrotti, l. (2015). Bilingualism,
mente, and brain. Annual Review of Linguistics, 1, 377–394.
https://doi.org/10.1146/annurev-linguist-030514-124937,
PubMed: 28642932
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. (2017).
lmerTest package: Tests in linear mixed effects models. Journal
of Statistical Software, 82(13), 1–26. https://doi.org/10.18637/jss
.v082.i13
Lee, K. H., Choi, Y. Y., Gray, J. R., Cho, S. H., Chae, J. H., Lee, S., &
Kim, K. (2006). Neural correlates of superior intelligence:
Stronger recruitment of posterior parietal cortex. NeuroImage,
29(2), 578–586. https://doi.org/10.1016/j.neuroimage.2005.07
.036, PubMed: 16122946
Lehtonen, M., Soveri, A., Laine, A., Järvenpää, J., de Bruin, A., &
Antfolk, J. (2018). Is bilingualism associated with enhanced
executive functioning in adults? A meta-analytic review. Psycho-
logical Bulletin, 144(4), 394–425. https://doi.org/10.1037
/bul0000142, PubMed: 29494195
Luk, G., & Bialystok, E. (2013). Bilingualism is not a categorical
variable: Interaction between language proficiency and usage.
Journal of Cognitive Psychology, 25(5), 605–621. https://doi.org
/10.1080/20445911.2013.795574, PubMed: 24073327
Luk, G., Verde, D. W., Abutalebi, J., & Grady, C. (2012). Cognitive
control for language switching in bilinguals: A quantitative
meta-analysis of functional neuroimaging studies. Language
and Cognitive Processes, 27(10), 1479–1488. https://doi.org/10
.1080/01690965.2011.613209, PubMed: 24795491
Mineroff, Z., Blank, IO. A., Mahowald, K., & Fedorenko, E. (2018). UN
robust dissociation among the language, multiple demand, E
Neurobiology of Language
662
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
default mode networks: Evidence from inter-region correlations
in effect size. Neuropsychologia, 119, 501–511. https://doi.org
/10.1016/j.neuropsychologia.2018.09.011, PubMed: 30243926
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, UN. H.,
Howerter, A., & Wager, T. D. (2000). The unity and diversity of
executive functions and their contributions to complex “frontal
lobe” tasks: A latent variable analysis. Cognitive Psychology,
41(1), 49–100. https://doi.org/10.1006/cogp.1999.0734,
PubMed: 10945922
Mohades, S. G., Struys, E., Van Schuerbeek, P., Baeken, C., Van De
Craen, P., & Luypaert, R. (2014). Age of second language acqui-
sition affects nonverbal conflict processing in children: An fMRI
study. Brain and Behavior, 4(5), 626–642. https://doi.org/10.1002
/brb3.246, PubMed: 25328840
Nichols, E. S., Wild, C. J., Stojanoski, B., Battista, M. E., & Owen,
UN. M. (2020). Bilingualism affords no general cognitive advan-
tages: A population study of executive function in 11,000 peo-
ple. Psychological Science, 31(5), 548–567. https://doi.org/10
.1177/0956797620903113, PubMed: 32310712
Nieto-Castañón, A., & Fedorenko, E. (2012). Subject-specific func-
tional localizers increase sensitivity and functional resolution of
multi-subject analyses. NeuroImage, 63(3), 1646–1669. https://
doi.org/10.1016/ J.NEUROIMAGE.2012.06.065, PubMed:
22784644
Nieuwenhuis, S., Forstmann, B. U., & Wagenmakers, E. J. (2011).
Erroneous analyses of interactions in neuroscience: A problem
of significance. Nature Neuroscience, 14(9), 1105–1107.
https://doi.org/10.1038/nn.2886, PubMed: 21878926
Paap, K. R., & Greenberg, Z. IO. (2013). There is no coherent evi-
dence for a bilingual advantage in executive processing. Cogni-
tive Psychology, 66(2), 232–258. https://doi.org/10.1016/j
.cogpsych.2012.12.002, PubMed: 23370226
Paunov, UN. M., Blank, IO. A., & Fedorenko, E. (2019). Functionally
distinct language and theory of mind networks are synchronized
at rest and during language comprehension. Journal of Neuro-
physiology, 121(4), 1244–1265. https://doi.org/10.1152/jn
.00619.2018, PubMed: 30601693
Pessoa, l. (2009). How do emotion and motivation direct executive
controllo? Trends in Cognitive Sciences, 13(4), 160–166. https://doi
.org/10.1016/j.tics.2009.01.006, PubMed: 19285913
Pliatsikas, C., & Luk, G. (2016). Executive control in bilinguals: A con-
cise review on fMRI studies. Bilingualism: Language and Cognition,
19(4), 699–705. https://doi.org/10.1017/S1366728916000249
Poldrack, R. UN. (2006). Can cognitive processes be inferred from
neuroimaging data? Trends in Cognitive Sciences, 10(2), 59–63.
https://doi.org/10.1016/j.tics.2005.12.004, PubMed: 16406760
Poldrack, R. UN. (2011). Inferring mental states from neuroimaging
dati: From reverse inference to large-scale decoding. Neuron,
72(5), 692–697. https://doi.org/10.1016/j.neuron.2011.11.001,
PubMed: 22153367
Energia, J. D., Cohen, UN. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., Vogel, UN. C., Laumann, T. O., Miezin, F. M.,
Schlaggar, B. L., & Petersen, S. E. (2011). Functional network
organization of the human brain. Neuron, 72(4), 665–678.
https://doi.org/10.1016/j.neuron.2011.09.006, PubMed:
22099467
Energia, J. D., Schlaggar, B., & Petersen, S. (2015). Recent progress
and outstanding issues in motion correction in resting state fMRI.
NeuroImage, 107, 536–551. https://doi.org/10.1016/j
.neuroimage.2014.10.044, PubMed: 25462692
Rodríguez-Pujadas, A., Sanjuán, A., Ventura-Campos, N., Román,
P., Martin, C., Barceló, F., Costa, A., & Ávila, C. (2013). Bilinguals
use language-control brain areas more than monolinguals to
perform non-linguistic switching tasks. PLOS ONE, 8(9), Article
e73028. https://doi.org/10.1371/journal.pone.0073028,
PubMed: 24058456
Rossi, E., Diaz, M., Kroll, J. F., & Dussias, P. E. (2017). Late
bilinguals are sensitive to unique aspects of second language
processing: Evidence from clitic pronouns word-order. Frontiers
in Psychology, 8, Article 342. https://doi.org/10.3389/fpsyg.2017
.00342, PubMed: 28367130
Ryan, C. (2013). Language use in the United States: 2011. https://
www.census.gov/library/publications/2013/acs/acs-22.html
Saxe, R., & Kanwisher, N. (2003). People thinking about thinking
people: The role of the temporo-parietal junction in “theory of
mind.” NeuroImage, 19(4), 1835–1842. https://doi.org/10.1016
/S1053-8119(03)00230-1, PubMed: 12948738
Saxe, R., Moran, J. M., Scholz, J., & Gabrieli, J. (2006). Overlapping
and non-overlapping brain regions for theory of mind and self
reflection in individual subjects. Social Cognitive and Affective
Neuroscience, 1(3), 229–234. https://doi.org/10.1093/scan
/nsl034, PubMed: 18985110
Schölvinck, M., Maier, A., Ye, F., Duyn, J., & Leopold, D. (2010).
Neural basis of global resting-state fMRI activity. Proceedings of
the National Academy of Sciences, 107(22), 10238–10243.
https://doi.org/10.1073/pnas.0913110107, PubMed: 20439733
Shashidhara, S., Spronkers, F. S., & Erez, Y. (2020). Individual-
subject functional localization increases univariate activation
but not multivariate pattern discriminability in the “multiple-
demand” frontoparietal network. Journal of Cognitive Neurosci-
ence, 32(7), 1348–1368. https://doi.org/10.1162/jocn_a_01554,
PubMed: 32108555
Sulpizio, S., Del Maschio, N., Del Mauro, G., Fedeli, G., & Abutalebi,
J. (2020). Bilingualism as a gradient measure modulates functional
connectivity of language and control networks. NeuroImage, 205,
Article 116306. https://doi.org/10.1016/j.neuroimage.2019
.116306, PubMed: 31654763
Tahmasebi, UN. M., Davis, M. H., Wild, C. J., Rodd, J. M., Hakyemez,
H., Abolmaesumi, P., & Johnsrude, IO. S. (2012). Is the link between
anatomical structure and function equally strong at all cognitive
levels of processing? Cerebral Cortex, 22(7), 1593–1603. https://
doi.org/10.1093/cercor/bhr205, PubMed: 21893681
Tao, L., Wang, G., Zhu, M., & Cai, Q. (2021). Bilingualism and
domain-general cognitive functions from a neural perspective:
A systematic review. Neuroscience and Biobehavioral Reviews,
125, 264–295. https://doi.org/10.1016/j.neubiorev.2021.02.029,
PubMed: 33631315
Taylor, S. F., Welsh, R. C., Wager, T. D., Phan, K. L., Fitzgerald,
K. D., & Gehring, W. J. (2004). A functional neuroimaging
study of motivation and executive function. NeuroImage, 21(3),
1045–1054. https://doi.org/10.1016/j.neuroimage.2003.10.032,
PubMed: 15006672
Teubner-Rhodes, S., Bolger, D. J., & Novick, J. M. (2019). Conflict
monitoring and detection in the bilingual brain. Bilingualism:
Language and Cognition, 22(2), 228–252. https://doi.org/10
.1017/S1366728917000670
Thierry, G., & Wu, Y. J. (2007). Brain potentials reveal unconscious
translation during foreign-language comprehension. Proceedings
of the National Academy of Sciences, 104(30), 12530–12535.
https://doi.org/10.1073/pnas.0609927104, PubMed: 17630288
Tschentscher, N., & Mitchell, D. (2017). Fluid intelligence predicts
novel rule implementation in a distributed frontoparietal control
rete. Journal of Neuroscience, 37(18), 4841–4847. https://doi
.org/10.1523/JNEUROSCI.2478-16.2017, PubMed: 28408412
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, O., Delcroix, N., & Joliot, M. (2002). Automated
Neurobiology of Language
663
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multiple demand network differences in bilinguals and monolinguals
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
NeuroImage, 15(1), 273–289. https://doi.org/10.1006/nimg
.2001.0978, PubMed: 11771995
Uddin, l. Q. (2021). Cognitive and behavioural flexibility: Neural
mechanisms and clinical considerations. Nature Reviews Neuro-
science, 22(3), 167–179. https://doi.org/10.1038/s41583-021
-00428-w, PubMed: 33536614
Vázquez-Rodríguez, B., Suárez, l. E., Markello, R. D., Shafiei, G.,
Paquola, C., Hagmann, P., van den Heuvel, M. P., Bernhardt,
B. C., Spreng, R. N., & Misic, B. (2019). Gradients of structure–
function tethering across neocortex. Proceedings of the National
Academy of Sciences, 116(42), 21219–21227. https://doi.org/10
.1073/pnas.1903403116, PubMed: 31570622
Waldie, K. E., Badzakova-Trajkov, G., Miliivojevic, B., & Kirk, IO. J.
(2009). Neural activity during Stroop colour-word task perfor-
mance in late proficient bilinguals: A functional magnetic
resonance imaging study. Psychology & Neuroscience, 2(2),
125–136. https://doi.org/10.3922/j.psns.2009.2.004
Wong, C., Olafsson, V., Tal, O., & Liu, T. (2013). The amplitude of
the resting-state fMRI global signal is related to EEG vigilance
measures. NeuroImage, 83, 989–990. https://doi.org/10.1016/j
.neuroimage.2013.07.057, PubMed: 23899724
Woolgar, A., Parr, A., Cusack, R., Thompson, R., Nimmo-Smith, I.,
Torralva, T., Roca, M., Antoun, N., Manes, F., & Duncan, J.
(2010). Fluid intelligence loss linked to restricted regions of
damage within frontal and parietal cortex. Atti del
National Academy of Sciences, 107(33), 14899–14902. https://
doi.org/10.1073/pnas.1007928107, PubMed: 20679241
Sì, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., Roffman, J. L., Smoller, J. W., Zöllei, L., Polimeni,
J. R., Fisch, B., Liu, H., & Buckner, R. l. (2011). The organization of
the human cerebral cortex estimated by intrinsic functional con-
nectivity. Journal of Neurophysiology, 106(3), 1125–1165. https://
doi.org/10.1152/jn.00338, PubMed: 21653723
Zirnstein, M., Bice, K., & Kroll, J. F. (2019). Variation in language
experience shapes the consequences of bilingualism. In I. UN.
Sekerina, l. Spradlin, & V. Valian (Eds.), Bilingualism, executive
function, and beyond: Questions and insights (Studies in Bilin-
gualism 57, pag. 35–47). John Benjamins Publishing. https://
benjamins.com/catalog/sibil.57.03zir. https://doi.org/10.1075
/sibil.57.03zir
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
4
6
4
7
1
9
7
9
6
7
9
N
o
_
UN
_
0
0
0
5
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurobiology of Language
664