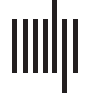RESEARCH ARTICLE
Distinct Neural Correlates of Linguistic and
Non-Linguistic Demand
Ian A. Quillen
, Melodie Yen
, and Stephen M. Wilson
Department of Hearing and Speech Sciences, Vanderbilt University Medical Center, Nashville, TN, USA
a n o p e n a c c e s s
j o u r n a l
Keywords: linguistic demand, task difficulty, accuracy, reaction time, multiple demand network, aphasia
ABSTRACT
In this study, we investigated how the brain responds to task difficulty in linguistic and non-
linguistic contexts. This is important for the interpretation of functional imaging studies of
neuroplasticity in post-stroke aphasia, because of the inherent difficulty of matching or controlling
task difficulty in studies with neurological populations. Twenty neurologically normal individuals
were scanned with fMRI as they performed a linguistic task and a non-linguistic task, each of which
had two levels of difficulty. Critically, the tasks were matched across domains (linguistico, non-
linguistico) for accuracy and reaction time, such that the differences between the easy and difficult
conditions were equivalent across domains. We found that non-linguistic demand modulated the
same set of multiple demand (MD) regions that have been identified in many prior studies. In
contrasto, linguistic demand modulated MD regions to a much lesser extent, especially nodes
belonging to the dorsal attention network. Linguistic demand modulated a subset of language
regions, with the left inferior frontal gyrus most strongly modulated. The right hemisphere region
homotopic to Broca’s area was also modulated by linguistic but not non-linguistic demand. When
linguistic demand was mapped relative to non-linguistic demand, we also observed domain by
difficulty interactions in temporal language regions as well as a widespread bilateral semantic
rete. In sum, linguistic and non-linguistic demand have strikingly different neural correlates.
These findings can be used to better interpret studies of patients recovering from aphasia. Some
reported activations in these studies may reflect task performance differences, while others can be
more confidently attributed to neuroplasticity.
Citation: Quillen, IO. A., Yen, M., &
Wilson, S. M. (2021). Distinct
neural correlates of linguistic and
non-linguistic demand. Neurobiology
of Language, 2(2), 202–225. https://
doi.org/10.1162/nol_a_00031
DOI:
https://doi.org/10.1162/nol_a_00031
Supporting Information:
https://doi.org/10.1162/nol_a_00031
Received: 5 Marzo 2020
Accepted: 13 Gennaio 2021
Competing Interests: The authors have
declared that no competing interests
exist.
INTRODUCTION
Corresponding Author:
Stephen M. Wilson
stephen.m.wilson@vanderbilt.edu
Handling Editor:
Yanchao Bi
Copyright: © 2021 Massachusetts
Institute of Technology. Pubblicato
under a Creative Commons Attribution
4.0 Internazionale (CC BY 4.0) licenza.
The MIT Press
How does the brain respond to task difficulty in linguistic and non-linguistic contexts? Our mo-
tivation for addressing this question is that it bears on the interpretation of functional imaging
studies of neuroplasticity in post-stroke aphasia. Individuals with aphasia, by their nature, find
language tasks more difficult than do neurologically normal control participants. Inoltre, COME
patients recover over time, language tasks generally become easier. These facts imply that com-
parisons between patients and controls, and longitudinal analyses as patients recover, are con-
founded by task difficulty. Therefore, when activation differences are observed, it is difficult to
determine whether they reflect functional reorganization or effects of task difficulty (Binder et al.,
2005; Fridriksson & Morrow, 2005; Geranmayeh et al., 2014; Price et al., 2006). Attempts have
been made to manipulate and match task difficulty between patients and controls (Brownsett
et al., 2014; Raboyeau et al., 2008; Sharp et al., 2004; Sharp et al., 2010; Wilson et al., 2018;
Wilson et al., 2019), but precise matching has proven difficult to achieve.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Linguistic and non-linguistic demand
The MD network:
A bilateral network including the
inferior frontal junction, anterior
insula, pre-supplementary motor
area, anterior-mid cingulate, E
intraparietal sulcus that supports
cognitive flexibility in many
diverse contexts.
Linguistic demand:
Refers to the additional processing
requirements of more difficult
linguistic tasks relative to easier
linguistic tasks.
Non-linguistic demand:
Refers to the additional processing
requirements of more difficult non-
linguistic cognitive tasks relative to
easier tasks.
The brain regions that are modulated by task difficulty are generally remarkably consistent
across tasks. A bilateral network including the inferior frontal junction, anterior insula, pre-
supplementary motor area (SMA), anterior-mid cingulate, and intraparietal sulcus has been
described as constituting a “multiple-demand” (MD) rete, supporting cognitive flexibility
in many contexts (Duncan & Owen, 2000; Fox et al., 2005). A compelling demonstration of
the generality of this network came from a study in which comparisons between difficult and
easy conditions of seven diverse cognitive tasks yielded similar patterns of fronto-parietal acti-
vation (Fedorenko et al., 2013).
Based on these findings, it could be speculated that the additional task difficulty that individ-
uals with aphasia experience when performing language tasks would yield increased activity in
MD regions (Geranmayeh et al., 2014). Tuttavia, it is not clear that linguistic demand is anal-
ogous to other types of cognitive demand. While two of the seven tasks investigated by
Fedorenko et al. (2013) involved linguistic stimuli, both of these were verbal working memory
compiti. Many previous studies have manipulated linguistic factors such as syntactic complexity,
ambiguity, word frequency, or difficulty of semantic decisions. Some studies have reported that
these manipulations modulated likely MD regions (Binder et al., 2004; Binder et al., 2005;
Blumstein et al., 2005; Eckert et al., 2009; Erb et al., 2013; Ihnen et al., 2015; Mollica et al.,
2020; Noppeney & Price, 2004; Piai et al., 2013; Sabsevitz et al., 2005; Vaden et al., 2013;
Wilson et al., 2016); while others have reported modulation of likely left hemisphere language
regions (Binder et al., 2005; Bornkessel et al., 2005; Graves et al., 2007; Graves et al., 2010; Just
et al., 1996; Makuuchi et al., 2009; Mason et al., 2003; Obleser et al., 2011; Rodd et al., 2005;
Roskies et al., 2001; Sabsevitz et al., 2005; Stromswold et al., 1996; Thompson et al., 2007;
Wilson et al., 2016). Often it is difficult to determine whether language or MD regions are in-
volved, given the close proximity of some of these regions (Fedorenko et al., 2012), and the fact
that most studies have not explicitly assessed both possibilities. Only a few studies have manip-
ulated both linguistic and non-linguistic demand, with modulation in common most often ob-
served in the anterior insula and/or anterior cingulate (Eckert et al., 2009; Erb et al., 2013; Piai
et al., 2013). No studies, to our knowledge, have matched task structure and behavioral factors
across linguistic and non-linguistic tasks so as to permit direct statistical comparisons.
The goal of the present study is to directly compare the brain regions that are modulated by
linguistic demand and non-linguistic demand. This entails matching task structure across
domains and ensuring that the difference in difficulty between easy and difficult conditions is
precisely matched across domains in terms of accuracy and reaction time. A clear picture
of similarities and differences between the neural correlates of linguistic demand and non-
linguistic demand may facilitate the interpretation of studies of neuroplasticity in aphasia, In
which task difficulty is so notoriously hard to control.
METHODS
Participants
Twenty neurologically normal individuals (age 26.6 ± 6.1 (SD) years, range 18–40 years; 3 male,
17 female; 17 right-handed, 3 left-handed; all native speakers of English; formazione scolastica 15.7 ± 1.5
years, range 12–18 years) were successfully scanned with fMRI. Participants were recruited by
word of mouth from Vanderbilt University Medical Center community in Nashville, Tennessee.
All participants, including the three left-handed participants, were left-lateralized for language,
as revealed by contrasts of semantic and perceptual conditions.
Prior to running the imaging study, a separate group of seven participants (age 26.7 ± 3.4 years,
range 23–30 years; 2 male, 5 female; all right-handed; all native speakers of English; formazione scolastica
Neurobiology of Language
203
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Linguistic and non-linguistic demand
16.9 ± 0.9 years, range 16–18 years) took part in behavioral studies for optimizing the experi-
mental design. These participants were recruited similarly; none of them were scanned.
All participants gave written informed consent and were compensated for their time. IL
study was approved by the institutional review board at Vanderbilt University Medical Center.
Experimental Design
In the fMRI study, five conditions were presented in a block design: (1) Semantic Easy; (2)
Semantic Difficult; (3) Perceptual Easy; (4) Perceptual Difficult; (5) Rest (Figura 1). All blocks
were 16 s in duration, and each block (except for the rest condition) consisted of eight stimuli,
which were presented every 2 S. Each run consisted of six blocks per condition in pseudorandom
order, for a total of 30 blocks (cioè., exactly 8 min).
Each participant was first trained on the task with untimed presentation of example items from
each condition, and specific instructions as described in detail below. Then, they performed one
complete practice run prior to entering the scanner, so that they would be familiarized with the
four active conditions and would settle on strategies for each condition. Finalmente, they performed
two runs in the scanner while echo-planar images were acquired.
In the four active conditions, each trial consisted of a pair of words or a pair of symbol strings
that were visually presented one above the other in the center of the screen. Participants were
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Experimental design and behavioral data. (UN) Example items from the four conditions.
Matching semantic items are shown, as indicated by the white box that appeared when participants
pressed the “match” button. Mismatching perceptual items are shown. Note that items were pre-
sented in green or red to cue participants as to the difficulty of the current condition. (B) Precisione
by condition. (C) Reaction time by condition.
Neurobiology of Language
204
Linguistic and non-linguistic demand
instructed to press a button with a finger of their left hand if the words “go together” or if the
symbol strings “are identical,” and to do nothing otherwise. If they pressed the button, a box
appeared around the words or symbols to acknowledge the button press, but no feedback
was provided as to whether the response was correct. The number of matching items varied
pseudorandomly by block and ranged from two out of eight (25%) to six out of eight (75%).
Responses were recorded between 300 ms and 1,900 ms, but the words or symbol strings dis-
appeared after 1,300 ms. Participants were instructed to respond as quickly as possible, but were
informed that responses after the stimuli had disappeared would still count.
In the Semantic Easy condition, half of the word pairs were semantically related, and half
were not. The words were relatively high frequency, concrete, and acquired early (Tavolo 1),
and the semantic relationships between the matching word pairs were chosen to be relatively
transparent. The words were presented in green text so that participants knew when they were
performing the easy condition. They were instructed: “Green words are easy. Matches will be
obvious: the words will clearly go together. You should be able to respond quickly.” The 96 In-
scanner word pairs and 48 practice word pairs for this condition were selected from the easier
items of the larger stimulus set described by Wilson et al. (2018).
The Semantic Difficult condition was the same as the Semantic Easy condition except that the
words were relatively low frequency, abstract, and acquired later (Tavolo 1), and the semantic
relationships between the matching word pairs were chosen to be relatively opaque. In this con-
dizione, the words were presented in red text so that participants knew when they were perform-
ing the difficult condition. They were instructed: “Red words are difficult. Matches may be more
subtle: the relationship between the words may be less obvious. Respond as quickly as you can,
Condition
Semantic Easy
Length
11.0 ± 0.8
Frequency
7.51 ± 0.89
Age of
acquisition
4.62 ± 0.88
Concreteness
558 ± 60
Tavolo 1.
Characteristics of the stimuli
Semantic Difficult Sub-level 1
11.0 ± 0.8
5.09 ± 0.94
8.61 ± 0.98
456 ± 87
Semantic Difficult Sub-level 2
11.0 ± 0.8
4.50 ± 0.89
9.63 ± 1.00
438 ± 87
Match
esempio
RABBIT
Mismatch
esempio
TOMATO
CARROT
BEACH
SENSE
LOGIC
SOAR
ERRAND
KERNEL
WHIFF
FLUTTER
OUTCOME
Semantic Difficult Sub-level 3
11.0 ± 0.8
3.54 ± 0.88
11.49 ± 1.37
400 ± 84
DECREE
KALE
Perceptual Easy
11.0 ± 0.7
Perceptual Difficult
11.0 ± 1.0
—
—
—
—
—
—
EDICT
ƟƕΨΓƜ
INCLINE
ϞΦΓƟƜδ
ƟƕΨΓƜ
ƜΓΘŒƧ
ϞƩЖЖΦ
ΞδΦƕƏƕ
ϞƩЖЖΦ
ΞδΦŒƏƕ
Note. Length is the total number of letters summed across both items in each pair; Frequency is the average log lemma frequency across each pair, based on the
American National Corpus (Reppen et al., 2005); Age of acquisition is the average across each pair in years from Kuperman et al. (2012); Concreteness is the
average across each pair obtained from the MRC database (Coltheart, 1981).
Neurobiology of Language
205
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Linguistic and non-linguistic demand
but take the time you need. If you need to respond after the words have disappeared from the
screen, that is fine.” Although 96 in-scanner word pairs and 48 practice word pairs were needed
for this condition, three times that many were selected from the more difficult items of the larger
stimulus set described by Wilson et al. (2018), such that there were three sub-levels of difficulty
among the difficult items. This was done to facilitate matching of accuracy between conditions.
During the experiment, items were selected from the three sub-levels depending on the partic-
ipant’s current relative accuracies on the Semantic Difficult and Perceptual Difficult conditions.
If semantic accuracy exceeded perceptual accuracy by more than 5%, then items from the most
difficult sub-level were presented. If semantic accuracy was more than 5% below perceptual
accuracy, then items from the least difficult sub-level were presented. Otherwise, items from
the middle sub-level were presented.
In the Perceptual Easy condition, the differences between matching and mismatching symbol
strings were readily apparent (Tavolo 1). Mismatching strings differed in every symbol and one
string was always five symbols long while the other was six symbols long. Matching strings were
identical and were five or six symbols long. The symbols were presented in green so that partic-
ipants knew when they were performing the easy condition. They were instructed: “Green sym-
bols are easy. If they mismatch, they will be very different. You should be able to respond
quickly.”
The Perceptual Difficult condition was the same as the Perceptual Easy condition, except that
pairs of mismatching strings always had the same number of symbols as each other (either 5 O 6)
and only one symbol differed between the two (Tavolo 1). In this condition, the symbols were
presented in red so that participants knew when they were performing the difficult condition.
They were instructed: “Red symbols are difficult. If they mismatch, only one of the symbols will
be different. Respond as quickly as you can, but take the time you need. If you need to respond
after the symbols have disappeared from the screen, that is fine.”
Note that the specificity of the instructions for each of the four conditions and the complete
8-min practice run prior to scanning were designed to maximize homogeneity of processing
strategies across participants. Also, it was important that participants knew when they were per-
forming easy or hard conditions. Without explicit cues to indicate easy conditions, participants
might seek subtle semantic relationships between words in Semantic Easy mismatching items,
and would have to scour Perceptual Easy matching items for potentially mismatching symbols,
which would result in both easy conditions being more demanding than intended.
Norming Experiment
To compare the neural correlates of linguistic and non-linguistic demand, we needed differ-
ences in accuracy and reaction time to be matched across the linguistic and non-linguistic do-
mains. To achieve this, we ran seven participants on preliminary versions of the experiment
without scanning them. Based on these participants’ behavioral data, we iteratively adjusted
several aspects of the experimental design until we arrived at the design described above, Quale
seemed likely to yield performance that would be balanced as required in the subsequent im-
aging study. Specifically, (1) we adjusted the degree of mismatch of the Perceptual Easy and
Perceptual Difficult mismatch trials; (2) we adjusted the length of time that the stimuli were
shown before being removed from the screen; (3) we introduced color cues to condition diffi-
culty; (4) we tweaked the instructions to encourage quick responses on easy conditions; (5) we
made the Semantic Difficult condition adaptive to participant performance; E (6) we decided
to have participants perform a complete run before scanning so that they would become familiar
with the different strategies entailed by the four conditions.
Neurobiology of Language
206
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Linguistic and non-linguistic demand
Neuroimaging
Participants were scanned on a Philips Achieva 3 Tesla scanner with a 32-channel head coil at
the Vanderbilt University Institute of Imaging Science. Visual stimuli were projected onto a
screen at the end of the bore, which participants viewed through a mirror mounted to the head
coil. T2*-weighted blood oxygen level-dependent (BOLD) echo planar images were collected
with the following parameters: 240 volumes + 4 initial volumes discarded; 35 axial slices in
interleaved order; slice thickness = 3.0 mm with 0.5 mm gap; field of view = 220 × 220 mm;
matrix = 96 × 96; repetition time (TR) = 2,000 ms; echo time (TE) = 30 ms; flip angle = 75°; SENSE
factor = 2; voxel size = 2.3 × 2.3 × 3.5 mm. T1-weighted structural images and coplanar T2-
weighted images were also acquired.
Behavioral Data Analysis
The behavioral data were analyzed with repeated measures ANOVAs in JMP version 12.0.1
(SAS Institute). Reaction times from all trials with button presses (cioè., hits and false alarms) were
included in the analyses. Within each condition for each participant, reaction times for each trial
were clipped at 2.5 standard deviations from the mean for that condition and that participant.
Neuroimaging Data Analysis
The functional imaging data were first preprocessed with tools from AFNI (Cox, 1996). Head
motion was corrected, with six translation and rotation parameters saved for use as covariates.
Prossimo, the data were detrended with a Legendre polynomial of degree 2, and smoothed with a
Gaussian kernel (FWHM = 6 mm). Then, independent component analysis was performed using
the FSL tool melodic (Beckmann & Smith, 2004). Noise components were manually identified
with reference to the criteria of Kelly et al. (2010) and removed using fsl_regfilt.
First level models were fit for each of the two functional runs using boxcar models of each active
condition convolved with a hemodynamic response function based on the difference of two
gamma density functions (time to first peak = 5.4 S; FWHM = 5.2 S; time to second peak = 15 S;
FWHM = 10 S; coefficient of second gamma density = 0.09) with the program fmrilm from the
FMRISTAT package (Worsley et al., 2002) in MATLAB R2019a (Mathworks). The six motion
parameters were included as covariates, as were time series from white matter and cerebrospinal
fluid regions to account for nonspecific global fluctuations, and three cubic spline temporal trends.
The T1-weighted anatomical images were warped to MNI space using unified segmentation
in SPM5 (Ashburner & Friston, 2005). Functional images were coregistered with structural
images via coplanar T2-weighted structural images using SPM, and warped to MNI space.
Contrasts were created to compare each of the four conditions to the implicit Rest baseline.
Linguistic demand was modeled with the contrast Semantic Difficult − Semantic Easy. Although
this contrast captures linguistic demand only in the specific context of a lexical-semantic task, it is
noteworthy that this contrast has previously been demonstrated to robustly activate core language
regions in general, not just lexical-semantic regions (Wilson et al., 2018), supporting its use as a
proxy for linguistic demand in general. Non-linguistic demand was modeled with the contrast
Perceptual Difficult − Perceptual Easy. Again this is a specific instantiation of non-linguistic
demand, but the striking similarity between regions modulated by non-linguistic demand across
a range of diverse cognitive tasks (Fedorenko et al., 2013) suggests that any kind of non-linguistic
demand can effectively serve as a proxy for non-linguistic demand in general. Language regions
were identified with the contrast (Semantic Easy + Semantic Difficult) − (Perceptual Easy +
Perceptual Difficult). Finalmente, the interaction of domain by difficulty was modeled by the contrast
Neurobiology of Language
207
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Linguistic and non-linguistic demand
(Semantic Difficult − Semantic Easy) − (Perceptual Difficult − Perceptual Easy). Second level ran-
dom effects analyses were performed and a cluster-defining threshold of p < 0.005 was applied.
Correction for multiple comparisons (p < 0.05) was carried out based on permutation testing of
the maximum cluster extent with the FSL tool randomise (Winkler et al., 2014). Specifically, null
distributions were created by randomly inverting the signs of individual contrast images.
A region of interest (ROI) analysis was carried out to examine responses to each of the four
active conditions in the MD network and the language network, using ROIs that were function-
ally defined in individual participants (Fedorenko et al., 2010). MD ROIs were defined based on
the symmetrical image of the MD network described by Fedorenko et al. (2013) and available for
download at http://imaging.mrc-cbu.cam.ac.uk/imaging/MDsystem. We used this image rather
than our own data in order to avoid circularity, since we intended to test whether ROIs were
modulated by perceptual demand, which is the same contrast we would use to define the
MD network. We defined spheres of radius 8 mm around prominent peaks in each hemisphere.
Twelve spheres were defined in the bilateral inferior frontal junction, anterior insula, pre-SMA/
anterior-mid cingulate, dorsal premotor cortex, intraparietal sulcus, and occipito-temporal cor-
tex. For the pre-SMA/anterior-mid cingulate ROI, we used peaks from sagittal slices x = ±6, be-
cause this region had only a single peak centered on the midline. Then, for each of the two runs,
we defined individual ROIs as the top 10% of voxels within each sphere that had the highest
t statistics for modulation by non-linguistic demand in the other run, and that were not modu-
lated by language (uncorrected p > 0.1).
Language ROIs were based on the language regions identified in our own data. This did not
introduce circularity, since the contrast to identify language regions was orthogonal to the con-
trasts under investigation in the ROI analysis. We further smoothed the language contrast image
using a Gaussian filter with FWHM = 8 mm in order to identify maximally general peaks, Poi
defined spheres of radius 8 mm around seven prominent peaks in the left hemisphere: the dorsal
pars opercularis of the inferior frontal gyrus (IFG), pars triangularis of the IFG, SMA, fusiform
gyrus, posterior superior temporal sulcus (STS), anterior STS, and hippocampus. Three additional
spheres were also defined around prominent right hemisphere peaks that did not reach statistical
significance but were clearly homotopic to left hemisphere language areas: the pars triangularis
of the IFG, posterior STS, and anterior STS. Then, for each of the two runs, we defined individual
ROIs as the top 10% of voxels within each sphere that had the highest t statistics for the language
contrast in the other run, and that were not modulated by non-linguistic demand (uncorrected
p > 0.1).
We used omnibus repeated measures ANOVAs in JMP and a series of t tests to determine
which regions were modulated by linguistic demand, which regions were modulated by percep-
tual demand, and which regions showed interactions of domain by difficulty, and to compare
the magnitude of these effects between the regions comprising each network. To account
for multiple comparisons, p values for all of these tests were obtained from permutation testing
using a custom procedure implemented in MATLAB. Specifically, null distributions for the
effects of linguistic and non-linguistic demand and the interaction of domain by difficulty were
created by randomly inverting the signs of individual contrast images, while null distributions for
differential effects between regions were created by randomly permuting contrast estimates
between regions within participants.
RESULTS
Our study is concerned with cortical networks, so only cortical activations will be described in
the text. Subcortical and cerebellar findings can be observed in the figures and tables.
Neurobiology of Language
208
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
N
o
_
UN
_
0
0
0
3
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Linguistic and non-linguistic demand
Behavioral Data
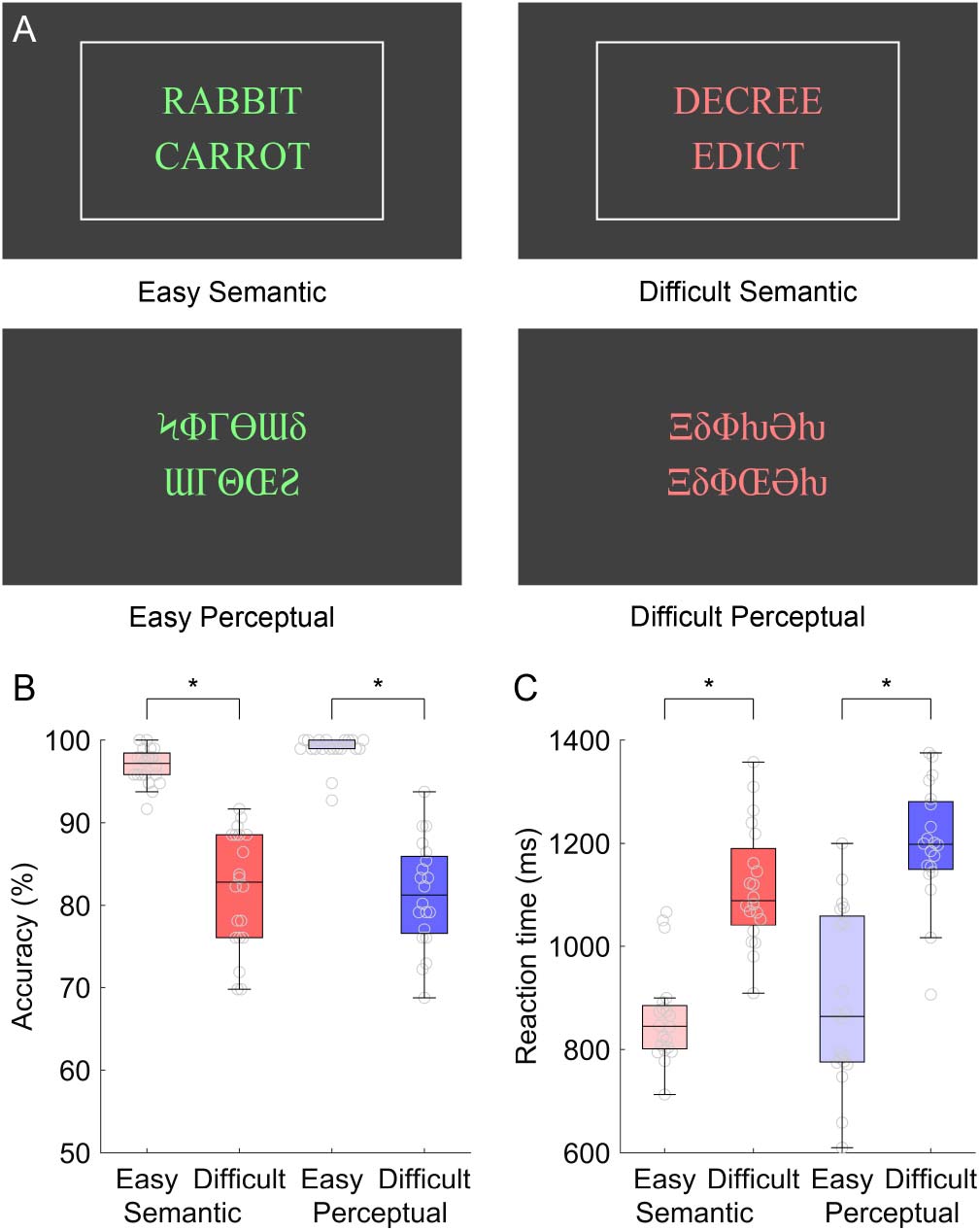
For accuracy, a repeated measures ANOVA with two within-subjects factors (domain, difficulty)
revealed a main effect of difficulty with difficult conditions less accurate than easy conditions,
F(1, 19) = 173.01, P < 0.0001, but no main effect of domain, F(1, 19) = 0.53, p = 0.48, and no
interaction of domain by difficulty, F(1, 19) = 2.22, p = 0.15 (Figure 1B).
For reaction time, a repeated measures ANOVA revealed a main effect of difficulty with
slower responses to difficult conditions compared to easy conditions, F(1, 19) = 151.13, p <
0.0001, but no main effect of domain, F(1, 19) = 4.21, p = 0.054, and no interaction of domain
by difficulty, F(1, 19) = 1.88, p = 0.19 (Figure 1C).
The absence of significant domain by difficulty interactions for accuracy or reaction time is
important, because it means that we can compare the neural correlates of linguistic demand and
non-linguistic demand without this comparison being confounded by accuracy or reaction time.
The lack of main effects of domain was welcome, though less important, since only the contrast
used to identify language regions could be impacted by main effects of domain; for that reason, we
were not concerned that the main effect of domain approached significance for reaction time.
One respect in which the domains did differ was in the nature of the errors made on the dif-
ficult conditions. On the Semantic Difficult condition, the total error rate of 18.0 ± 7.1% (SD) was
made up of 14.4 ± 6.6% misses and 3.6 ± 3.2% false alarms, while on the Perceptual Difficult
condition, the total error rate of 18.7 ± 6.5% was made up of 3.5 ± 1.9% misses and 15.2 ± 6.7%
false alarms. To ensure that our imaging findings were not impacted by the differing ratios of “go”
and “no-go” responses across conditions, all analyses described in the following sections were
repeated with the inclusion of an additional explanatory variable modeling “go” versus “no-go”
responses within blocks. This variable picked up left-hand button presses as expected, but its
inclusion did not meaningfully change the results for any contrasts of interest.
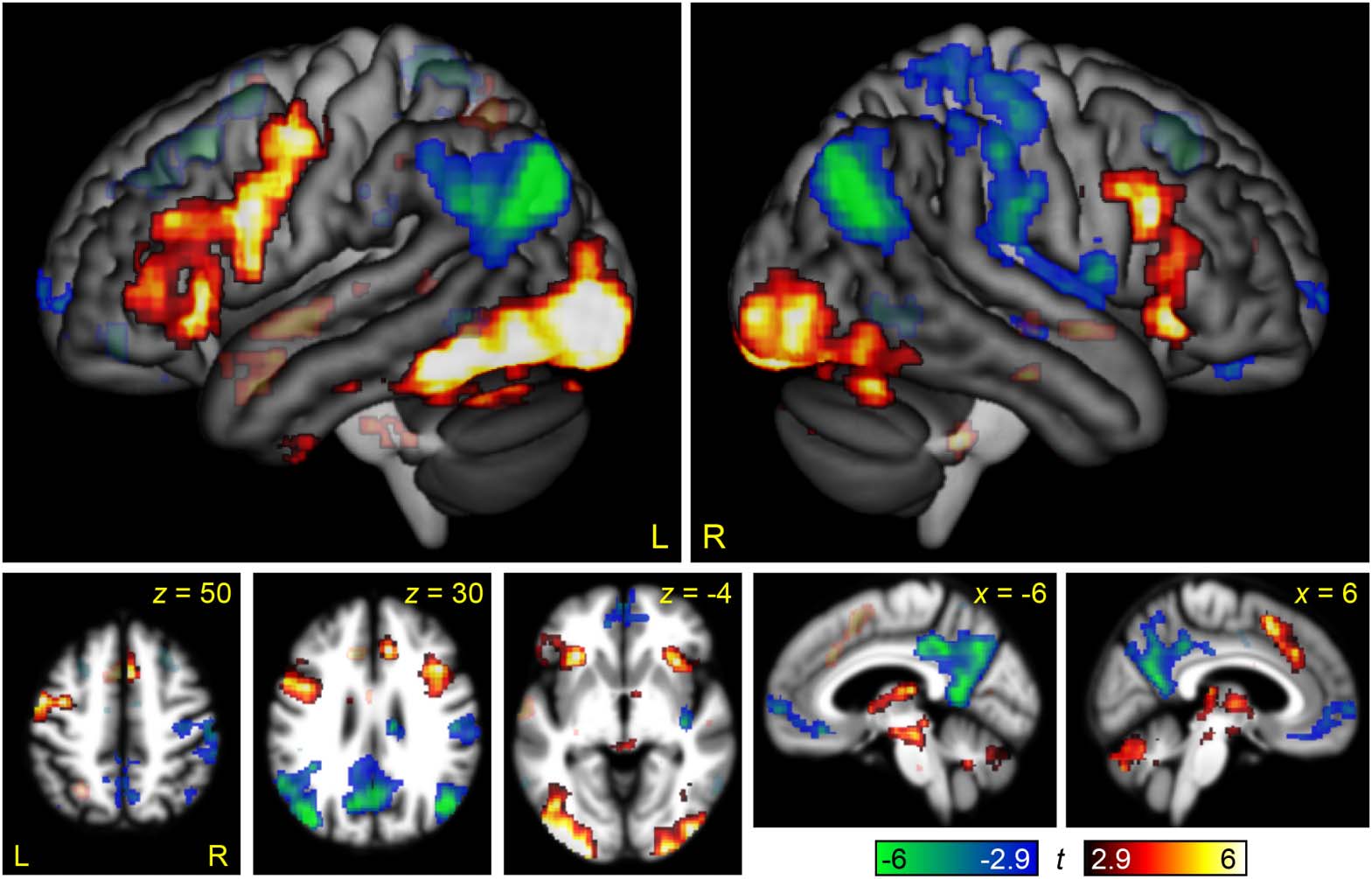
Brain Regions Modulated by Linguistic Demand
The contrast between the Semantic Difficult and Semantic Easy conditions was used to identify
brain regions modulated by linguistic demand (Figure 2, Table 2). In the left hemisphere, the
regions that were differentially active for the more difficult condition included the left IFG (pars
opercularis and triangularis) and sulcus, the inferior frontal junction, the precentral gyrus, the
anterior insula, and an extensive occipito-temporal region extending anteriorly along the fusi-
form gyrus almost to the temporal pole; also notable was activation of the left pre-SMA/anterior-
mid cingulate that did not meet the cluster extent threshold ( p = 0.082). Activations in the right
hemisphere were similar but less extensive: The inferior frontal activation was largely restricted
to the ascending ramus of the Sylvian fissure, the inferior frontal junction, and the anterior insula,
while the occipito-temporal region did not extend nearly as anteriorly; the right pre-SMA/
anterior-mid cingulate did, however, reach significance. The regions that were deactivated
by this contrast were bilateral and reflected the default mode network: the angular gyrus, pre-
cuneus, posterior cingulate, and ventromedial prefrontal cortex.
Voxelwise Comparisons Between Regions Modulated by Linguistic Demand, the MD Network, and the
Language Network
We carried out several different analyses to investigate how the brain regions modulated by lin-
guistic demand relate to the MD network and the language network.
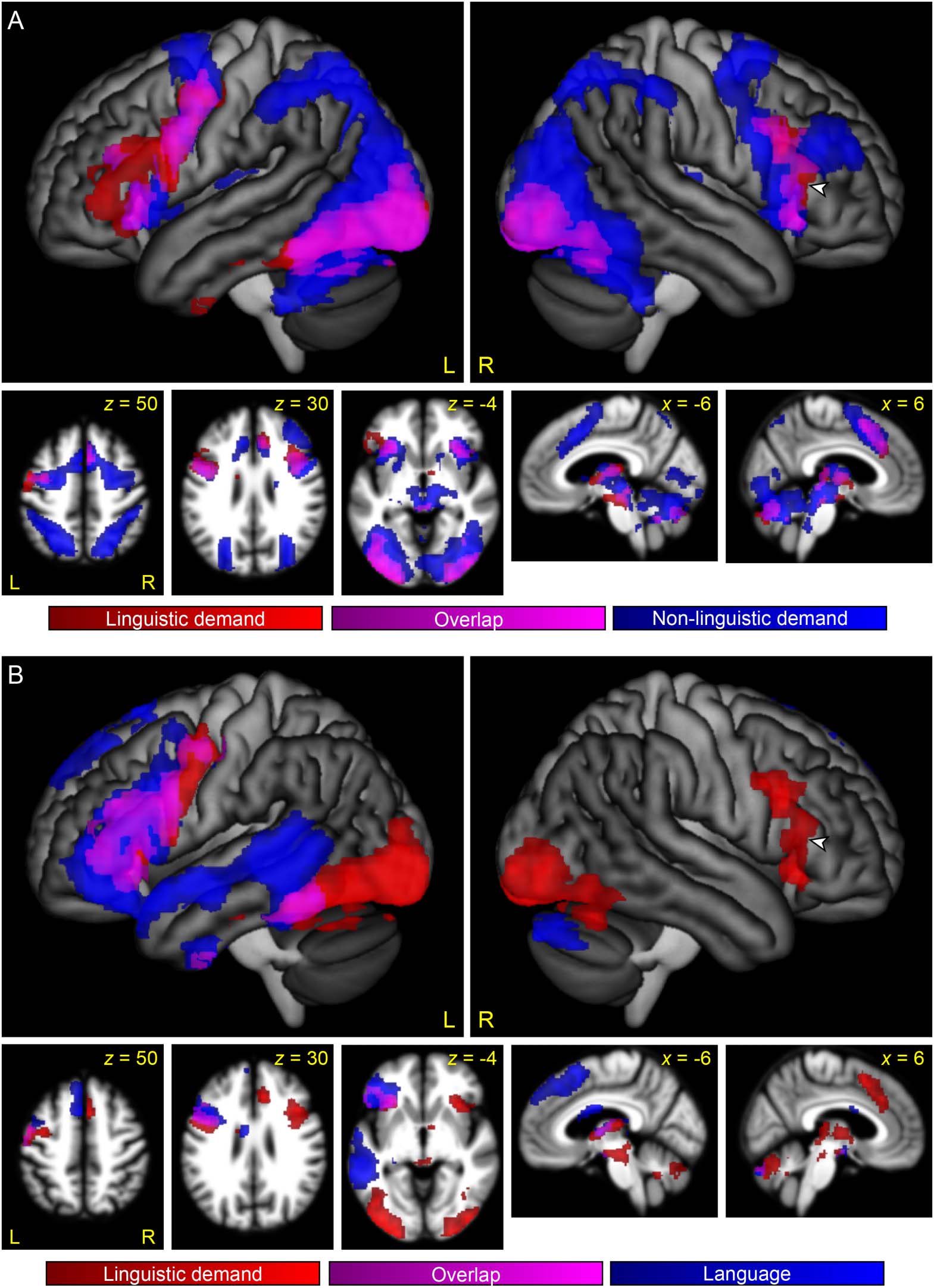
First, we plotted the overlap between linguistic demand regions and MD regions (Figure 3A).
MD regions were identified by the contrast between the Perceptual Difficult and Perceptual Easy
Neurobiology of Language
209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
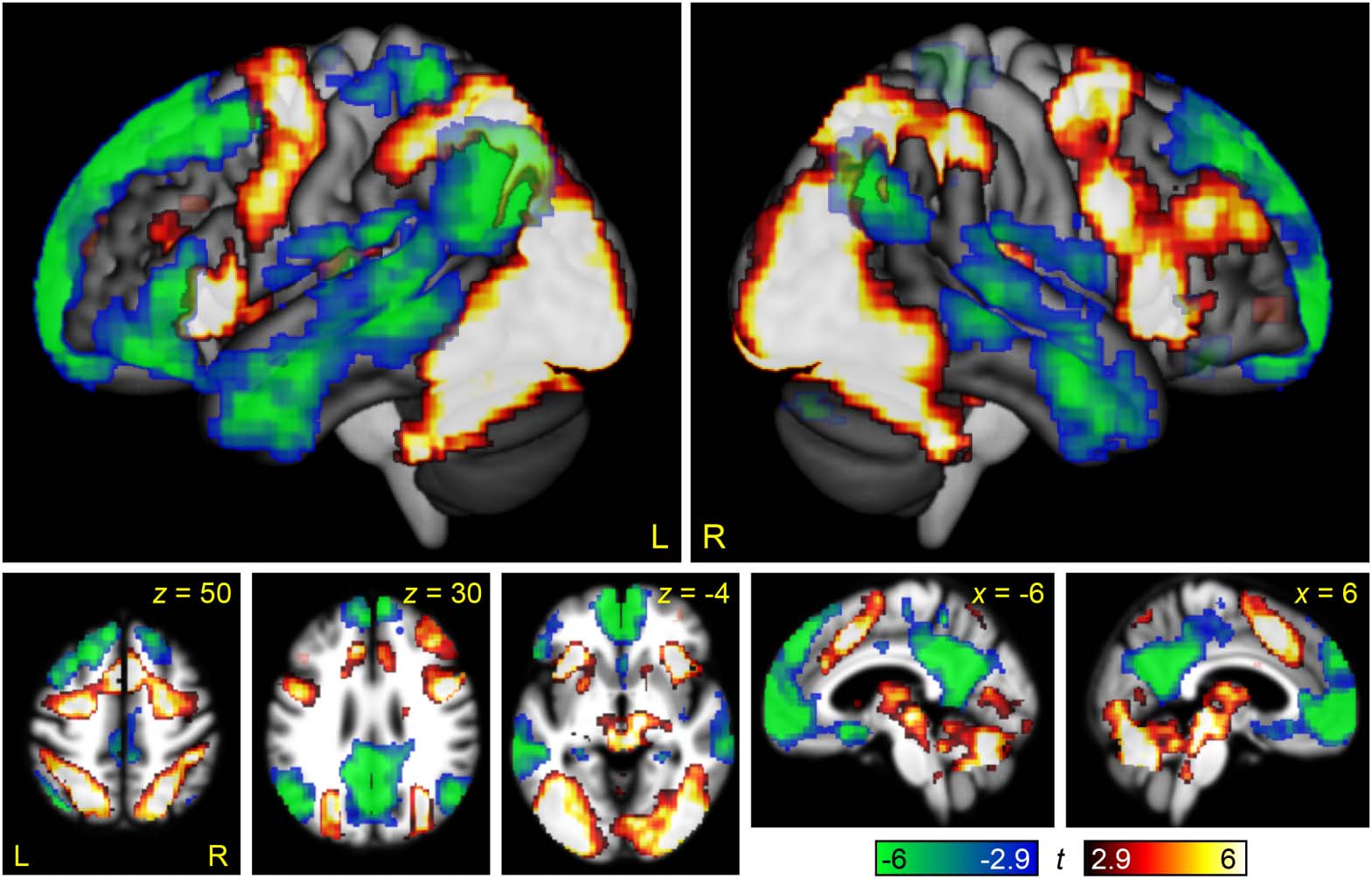
Figure 2. Brain regions modulated by linguistic demand. The contrast between the Semantic
Difficult and Semantic Easy conditions is shown in hot colors, while the reverse contrast is shown
in cool colors. Opaque = statistically significant, corrected for multiple comparisons; transparent =
voxelwise p < 0.005, but did not meet cluster extent threshold.
conditions, which activated a widespread bilateral network (Figure 4), consistent with prior
research.
We found that many of the regions that were modulated by linguistic demand belonged to the
MD network, specifically, the bilateral inferior frontal junction extending especially in the left
hemisphere onto the adjacent precentral gyrus, the bilateral anterior insula, the ventrolateral
component of occipito-temporal regions bilaterally, and the right pre-SMA/anterior-mid cingu-
late. (Note that the left pre-SMA/anterior-mid cingulate narrowly missed significance for modu-
lation by linguistic demand, as seen in Figure 2).
Next, we plotted the overlap between linguistic demand regions and language regions
(Figure 3B). Language regions were identified by the contrast of both semantic matching condi-
tions to both perceptual matching conditions, which activated strongly left-lateralized frontal
and temporal language regions (Figure 5), consistent with prior research.
This analysis showed that several of the regions that were modulated by linguistic demand
were language regions, specifically, left frontal regions (the pars opercularis and pars triangularis
of the IFG, inferior frontal sulcus, dorsal precentral gyrus, and anterior insula) and a region ex-
tending anteriorly from the visual word form area along the length of the fusiform gyrus.
The largest cluster that was modulated by linguistic demand but was neither an MD region
nor a left hemisphere language region was located in the right IFG, centered on the ascending
ramus of the Sylvian fissure, which separates the pars opercularis from the pars triangularis (clus-
ter extent = 1,456 mm3, maximum t = 4.67; center of mass MNI coordinates = 52, 26, 10, see
arrowheads in Figure 3).
Finally, we computed a whole brain interaction contrast of domain by difficulty, in order to
directly compare modulation by linguistic and non-linguistic demand (Figure 6, Table 2). This
Neurobiology of Language
210
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
Brain region(s)
Linguistic demand
Table 2.
Coordinates of activated regions
Extent (mm3) Max t
MNI coordinates
y
x
z
p
Left fusiform gyrus, inferior temporal gyrus, lingual gyrus, inferior
33,432
13.61
−37
−71
−12
0.0017
and middle occipital lobe
Left IFG pars opercularis and triangularis, inferior frontal sulcus,
30,224
7.20
−43
15
20
0.0028
inferior frontal junction, precentral gyrus, anterior insula
Right inferior occipital lobe
Bilateral thalamus, basal ganglia, midbrain
Right IFG ascending ramus of the Sylvian fissure, inferior frontal
junction, anterior insula
Bilateral cerebellum
Right pre-SMA/anterior-mid cingulate
Negative linguistic demand
Bilateral precuneus, posterior cingulate
Left angular gyrus
Right supramarginal gyrus, postcentral gyrus
Right angular gyrus
Bilateral ventromedial prefrontal cortex
Right posterior insula
Interaction of domain by difficulty
17,824
13,928
12,008
6,792
5,680
6.51
7.44
8.10
6.44
6.20
37,016
9.86
16,616
10.46
15,408
8,672
7,280
5,712
5.21
8.24
5.64
5.84
36
−3
41
3
9
−1
−45
48
47
2
41
Left IFG pars triangularis and orbitalis, superior temporal gyrus,
45,112
7.22
−47
STS, middle temporal gyrus, hippocampus
Bilateral ventromedial prefrontal cortex, superior frontal gyrus
Bilateral precuneus, posterior cingulate
Right superior temporal gyrus, STS, middle temporal gyrus
Negative interaction of domain by difficulty
37,008
26,840
15,584
8.61
7.70
6.84
Much of the bilateral multiple demand network, cerebellum
232,624
10.66
Left dorsal precentral gyrus and sulcus, superior frontal sulcus,
14,776
7.47
pre-SMA, anterior-mid cingulate
Left anterior insula, basal ganglia
Right thalamus
Note. Coordinates are centers of mass.
Neurobiology of Language
7,672
7,584
6.84
7.21
−1
−1
60
7
−20
−28
16
−78
−12
19
−71
24
−53
−65
−27
−66
49
−6
−4
50
−51
−14
−53
−1
18
−15
−11
1
17
−28
38
29
30
44
31
−6
5
−9
18
28
−12
16
54
8
11
0.0085
0.013
0.017
0.036
0.046
0.0010
0.0091
0.011
0.027
0.036
0.049
0.0013
0.0020
0.0038
0.0099
< 0.0001
0.012
0.029
0.029
211
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. The relationship between regions modulated by linguistic demand, the multiple demand
network, and the language network. (A) Regions modulated by linguistic demand are shown in red
or violet. Regions modulated by non-linguistic demand, as revealed by the contrast between the
Perceptual Difficult and the Perceptual Easy conditions, are shown in blue or violet. Violet indicates
overlap. (B) Regions modulated by linguistic demand are shown in red or violet. Language regions,
as revealed by the contrast of both semantic conditions to both perceptual conditions, are shown in
blue or violet. Violet indicates overlap. Arrowheads indicate right IFG region that was modulated by
linguistic demand, but was neither an MD region nor a language region.
Neurobiology of Language
212
Linguistic and non-linguistic demand
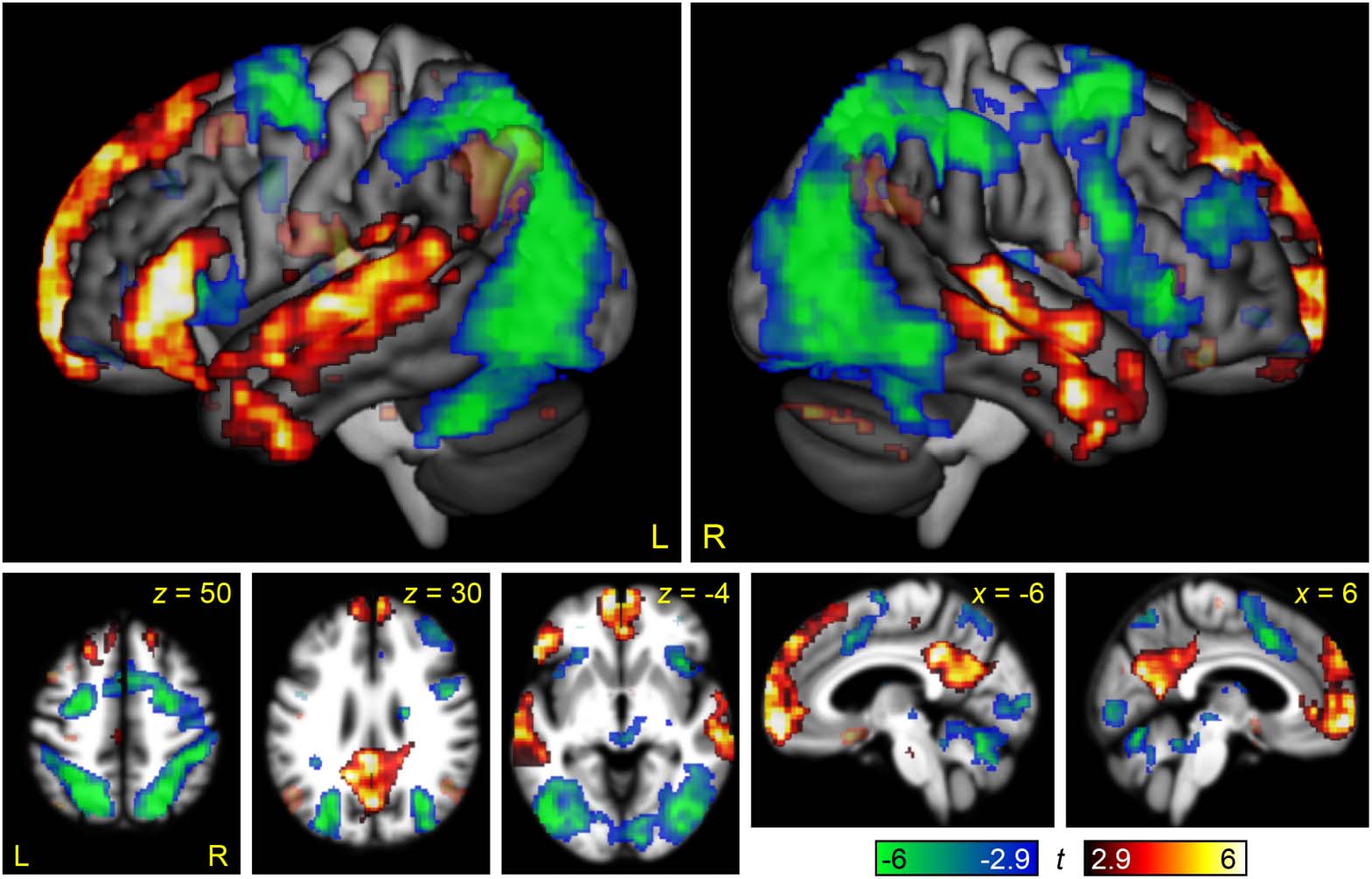
Figure 4. Brain regions modulated by non-linguistic demand. The contrast between the Perceptual
Difficult and Perceptual Easy conditions is shown in hot colors, while the reverse contrast is shown
in cool colors. Opaque = statistically significant, corrected for multiple comparisons; transparent =
voxelwise p < 0.005, but did not meet cluster extent threshold.
interaction map revealed large scale differences between linguistic demand and non-linguistic
demand. Core language regions in the left IFG (pars triangularis and orbitalis) and left superior
temporal gyrus, STS, and middle temporal gyrus were differentially modulated by linguistic
demand relative to non-linguistic demand, as were right hemisphere homotopic regions in
the temporal lobe, and midline semantic/default mode regions in the anterior cingulate, medial
prefrontal cortex, and the precuneus. Conversely, almost the entire MD network was differen-
tially modulated by non-linguistic demand. It is important to note that many of these signifi-
cant interactions were the consequence not of positive modulation by linguistic demand, but
of negative modulation by non-linguistic demand, especially throughout the default mode
network.
Functionally Defined Regions of Interest
A limitation of voxelwise group analyses is that participants are aligned anatomically but not
functionally, potentially resulting in the conflation of adjacent but functionally distinct brain re-
gions (I. A. Blank et al., 2017; Fedorenko et al., 2010). Therefore, we next carried out a ROI
analysis in which we plotted signal change as a function of domain (linguistic, non-linguistic)
and difficulty (easy, difficult) in MD network and language network nodes that were functionally
defined in individual participants (Figure 7, Table 3).
All twelve MD regions examined showed similar functional profiles. All twelve were modu-
lated by linguistic demand (except for one region with p = 0.07), reflecting greater sensitivity of
functional ROI-based analysis compared to the voxelwise analysis. All twelve regions were
modulated by non-linguistic demand, and all twelve showed interactions whereby modulation
was greater by non-linguistic demand than linguistic demand; these findings were consistent
Neurobiology of Language
213
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
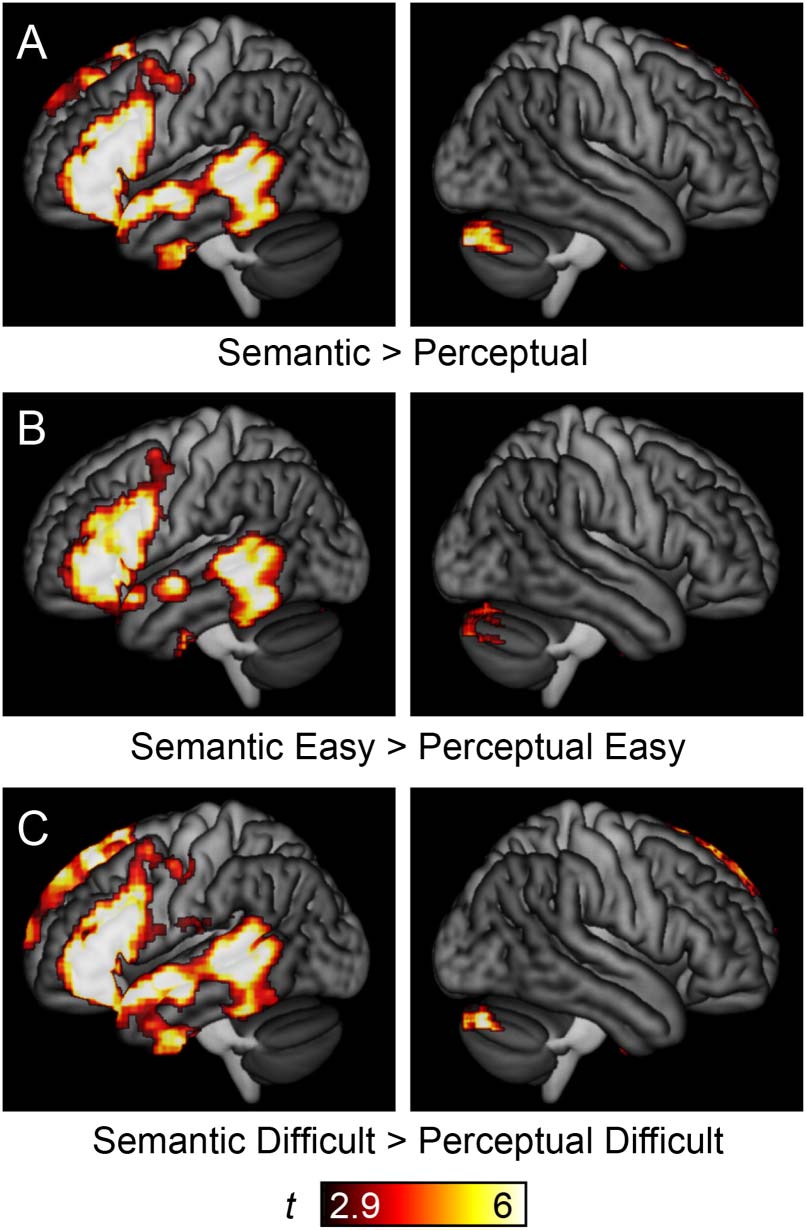
Figure 5.
Language regions of the brain. (A) Language areas, as revealed by the contrast of
(Semantic Easy + Semantic Difficult) − (Perceptual Easy + Perceptual Difficult). Note that contrasts
between (B) the two easy conditions or (C) the two difficult conditions yielded similar results.
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. The interaction between linguistic demand and domain-general demand. Regions where
modulation by linguistic demand was greater than modulation by domain-general demand are shown
in hot colors, while the reverse contrast is shown in cool colors. Opaque = statistically significant, corrected
for multiple comparisons; transparent = voxelwise p < 0.005, but did not meet cluster extent threshold.
Neurobiology of Language
214
Linguistic and non-linguistic demand
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
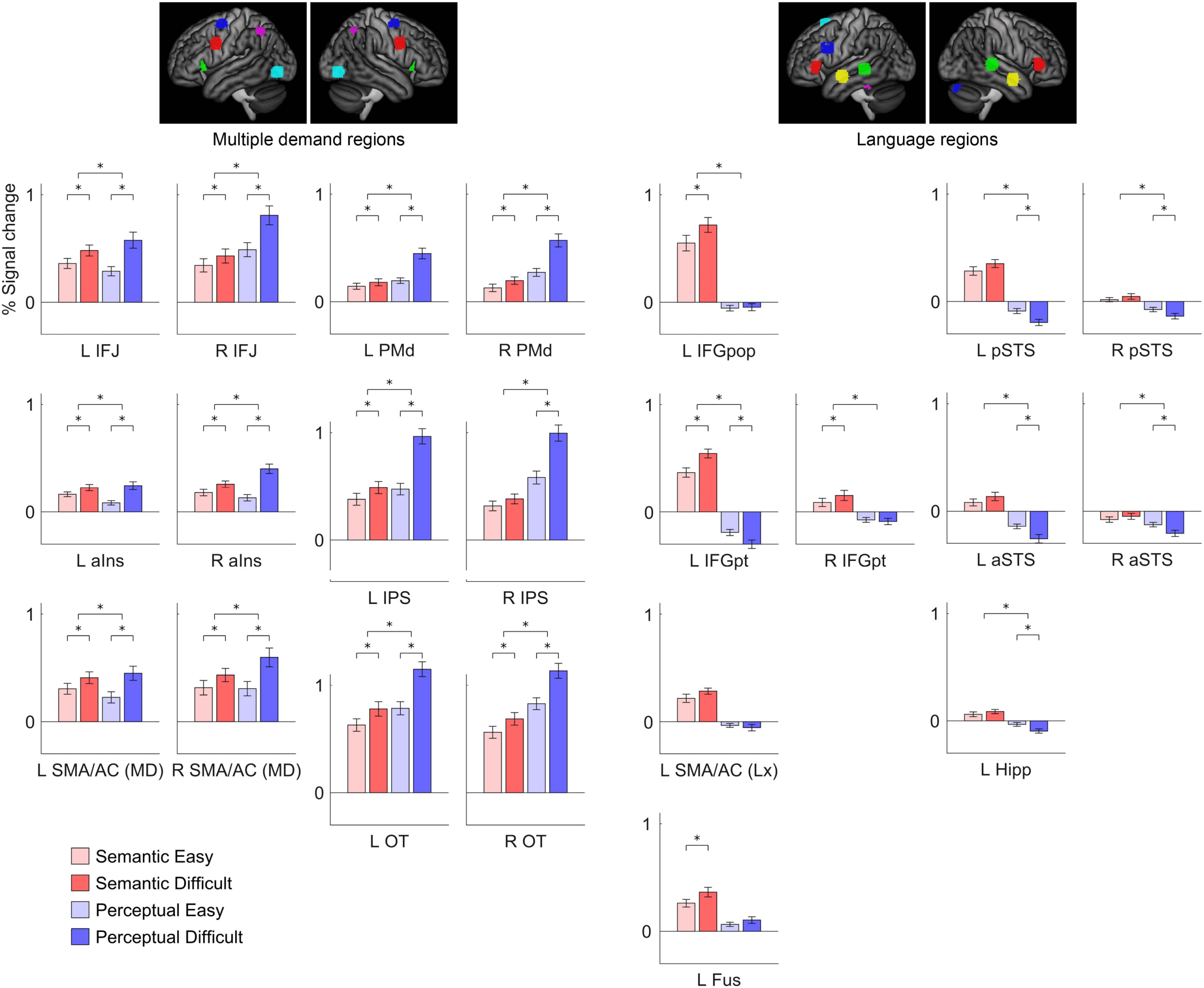
Figure 7.
Functional region of interest analysis. Signal change for the four conditions in multiple demand regions, language regions, and right
hemisphere regions homotopic to language regions. Error bars show standard error of the mean. The upper significance markers indicate the
significance of the interaction of domain by difficulty. L = left; R = right; IFJ = inferior frontal junction; aIns = anterior insula; SMA/AC = pre-
supplementary motor area/anterior-mid cingulate; PMd = dorsal premotor cortex; IPS = intraparietal sulcus; OT = occipitotemporal cortex;
IFGpop = inferior frontal gyrus, pars opercularis; IFGpt = inferior frontal gyrus, pars triangularis; Fus = fusiform gyrus; pSTS = posterior superior
temporal sulcus; aSTS = anterior superior temporal suclus; Hipp = hippocampus.
with the voxelwise analyses. Although all twelve MD regions were modulated by linguistic
demand, they differed in the extent to which they were modulated, F(4.85, 92.23) = 5.37,
p = 0.0003 (Supplementary Table 1; supporting information can be found online at https://
doi.org/10.1162/nol_a_00031). The left inferior frontal junction and left occipito-temporal re-
gion were most strongly modulated, while left dorsal premotor cortex was least strongly modu-
lated. The twelve regions were also not equivalent in their relative modulation by linguistic and
non-linguistic demand, F(5.06, 96.18) = 14.49, p < 0.0001 (Supplementary Table 2). The left
Neurobiology of Language
215
Linguistic and non-linguistic demand
Modulation by linguistic demand
dz
punc
p
Table 3.
Region of interest analysis
Modulation by non-linguistic demand
dz
punc
p
Interaction of domain by difficulty
dz
punc
p
MD network
L IFJ
R IFJ
L aIns
R aIns
1.37
< 0.0001
0.0002 *
1.08
0.77
0.95
0.0001
0.0012 *
0.0027
0.016
*
0.0004
0.0033 *
L SMA/AC
0.94
0.0005
0.0034 *
R SMA/AC 1.18
< 0.0001
0.0007 *
L PMd
R PMd
L IPS
R IPS
L OT
R OT
0.64
0.71
1.02
0.60
0.0098
0.048
0.005
0.026
*
*
0.0002
0.0019 *
0.015
0.071
1.33
< 0.0001
0.0002 *
1.07
0.0001
0.0014 *
Language network
L IFGpop
1.06
0.0001
0.001
*
L IFGpt
1.20
< 0.0001
0.0002 *
L SMA/AC
0.51
0.033
0.18
L Fus
L pSTS
L aSTS
L Hipp
R IFGpt
R pSTS
R aSTS
1.15
0.42
0.49
0.38
0.73
0.39
0.45
0.0001
0.0002 *
0.077
0.042
0.10
0.35
0.22
0.44
0.0042
0.032
*
0.097
0.057
0.42
0.27
1.25
1.86
1.27
1.89
1.83
1.81
1.65
1.78
2.29
2.31
2.29
1.83
0.09
−1.13
−0.22
0.49
−1.12
−0.98
−0.80
−0.19
−0.91
−1.39
< 0.0001
0.0002 *
< 0.0001
< 0.0001 *
< 0.0001
0.0002 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
< 0.0001
< 0.0001 *
−0.72
−1.43
−0.98
−1.39
−1.02
−0.94
−1.52
−1.59
−1.96
−1.82
−1.57
−1.31
0.0044
0.018 *<
< 0.0001
< 0.0001 *<
0.0003
0.0022 *<
< 0.0001
< 0.0001 *<
0.0002
0.0015 *<
0.0005
0.0029 *<
< 0.0001
< 0.0001 *<
< 0.0001
< 0.0001 *<
< 0.0001
< 0.0001 *<
< 0.0001
< 0.0001 *<
< 0.0001
< 0.0001 *<
< 0.0001
0.0002 *<
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
0.68
1.00
0.73
0.004
0.025 *
0.0001
0.0007 *<
1.63
< 0.0001
< 0.0001 *
0.34
0.041
0.96
0.27
0.0001
0.0009 *<
0.0003
0.0026 *<
0.0019
0.016
*<
0.40
0.98
0.0006
0.0051 *<
< 0.0001
< 0.0001 *<
0.57
0.57
1.07
0.84
0.75
0.69
0.93
1.16
0.020
0.019
0.11
0.10
0.0001
0.0017 *
0.0014
0.011 *
0.0034
0.022 *
0.0058
0.037 *
0.0005
0.005 *
0.0001
0.0007 *
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Note. punc = uncorrected p value; * = statistically significant after correction for multiple comparisons; < = negative modulation; see Figure 7 caption for
abbreviations.
anterior insula showed the most similar modulation across domains, while the bilateral intrapar-
ietal sulci and dorsal premotor cortex showed the greatest discrepancy between domains. Of
note, no homotopic pairs of regions differed from one another in terms of modulation by linguis-
tic demand or the interaction of domain by difficulty, though the anterior insula showed a trend
toward an interaction, with less of a discrepancy between domains in the left anterior insula
compared to the right ( p = 0.058).
Neurobiology of Language
216
Linguistic and non-linguistic demand
The ten language regions (including homotopic regions) examined showed a more diverse
array of functional profiles (Figure 7, Table 3). All language regions were numerically modulated
by linguistic demand, though this modulation was statistically significant only for the left IFG
pars opercularis, left IFG pars triangularis, left fusiform gyrus, and right IFG pars triangularis,
consistent with the voxelwise analysis described in the previous section. The differences
between regions in extent of modulation were significant, F(3.68, 69.90) = 8.62, p < 0.0001
(Supplementary Table 3), and were driven mainly by the left IFG pars opercularis and left IFG
pars triangularis, which were more strongly modulated than almost every other region. None of
the language regions were modulated by non-linguistic demand, and several of them were
negatively modulated: the left IFG pars triangularis, left posterior STS, left anterior STS, left
hippocampus, and right posterior STS. Eight of the ten language regions showed significant
interactions of domain by difficulty: the left IFG pars opercularis, left IFG pars triangularis, left
posterior STS, left anterior STS, left hippocampus, right IFG pars triangularis, right posterior STS,
and right anterior STS, and there were differences between regions in their relative modulation
by linguistic and non-linguistic demand, F(4.25, 80.74) = 7.05, p < 0.0001 (Supplementary
Table 4). Specifically, the left IFG pars triangularis showed a greater discrepancy between
domains than almost every other region. Among the homotopic pairs of nodes examined, the
left IFG pars triangularis showed greater modulation by linguistic demand than the right IFG pars
triangularis ( p = 0.034) as well as a greater discrepancy of modulation relative to non-linguistic
demand ( p = 0.0001). There were no interhemispheric differences in the temporal lobes.
DISCUSSION
In many diverse cognitive paradigms, task difficulty modulates the MD network in a highly ste-
reotyped manner (Duncan & Owen, 2000; Fedorenko et al., 2013). Our results indicate that the
neural correlates of linguistic demand are strikingly different. We found that linguistic demand
modulated the MD network to a lesser extent than non-linguistic demand, and that this discrep-
ancy was more pronounced for some nodes than others. Outside of the MD network, linguistic
demand modulated a subset of language regions, with the most robust modulation in the left IFG;
a homotopic region in the right IFG was also modulated. Finally, if linguistic demand is concep-
tualized relative to non-linguistic demand, linguistic demand could be seen to modulate left
temporal language regions as well as a wider bilateral semantic network.
How MD Regions Respond to Linguistic Demand
All twelve MD regions examined were modulated by linguistic demand, but all twelve were less
modulated by linguistic demand than they were by non-linguistic demand, suggesting that task
difficulty in a linguistic context depends less on the MD network than task difficulty in non-
linguistic contexts. This may reflect the fact that linguistic demand draws also on brain regions
outside the MD network, as discussed below. The discrepancy between linguistic and non-
linguistic demand was particularly pronounced in the dorsal premotor and intraparietal sulcus
nodes, which belong to the dorsal attention network and are involved in spatial attention and eye
movements (Corbetta & Shulman, 2002, 2011).
A number of previous studies have shown that manipulations of various kinds of linguistic
difficulty modulate likely MD regions (Binder et al., 2004; Binder et al., 2005; Blumstein
et al., 2005; Eckert et al., 2009; Erb et al., 2013; Ihnen et al., 2015; Mollica et al., 2020;
Noppeney & Price, 2004; Piai et al., 2013; Sabsevitz et al., 2005; Vaden et al., 2013; Wilson
et al., 2016). What is novel about our study is that it is the first, to our knowledge, to match task
Neurobiology of Language
217
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
structure, accuracy, and reaction time across linguistic and non-linguistic tasks so as to permit
direct statistical comparisons between domains.
There was no indication that recruitment of the MD network for increasing linguistic demand
was left-lateralized. This contrasts with recruitment of the MD network for language tasks rela-
tive to resting baselines, which is modestly left-lateralized (Diachek et al., 2020). Taken together,
these two findings suggest that the basic demands of performing a language task draw somewhat
more on left hemisphere MD regions, but that additional demands as the language task becomes
more difficult recruit MD regions in both hemispheres similarly.
It is unclear to what extent MD regions support language processing in real world language
use, as opposed to only in metalinguistic tasks. I. A. Blank and Fedorenko (2017) examined in-
tersubject correlations between participants listening to the same narratives in MD regions and
language regions. They found stronger intersubject correlations in language regions than MD
regions, and focused their interpretation on this difference, but it is noteworthy that intersubject
correlations throughout the MD network were still highly significant. We interpret this as evi-
dence for modulation of the MD network by the systematic time-varying demands of (relatively)
ecologically valid language comprehension. However other studies from the same group have
provided evidence against MD involvement in real world language processing, including lack of
MD recruitment in the absence of overt tasks (Diachek et al., 2020), lack of MD modulation by
surprisal (Shain et al., 2020), and lack of MD modulation by online measures of incremental
processing load (Wehbe et al., 2020). This question is not central to our study, because we
are primarily concerned with informing the interpretation of studies of language processing in
aphasia, which have usually involved metalinguistic tasks. But going forward, the resolution of
this question will be important, because we are interested in understanding the neural mecha-
nisms of real world language processing, not just performance of language tasks.
How Language Regions Respond to Linguistic Demand
Left frontal language regions—the pars triangularis and pars opercularis of the IFG—were strongly
modulated by linguistic demand, consistent with many previous findings (Binder et al., 2005;
Bornkessel et al., 2005; Graves et al., 2007; Graves et al., 2010; Just et al., 1996; Makuuchi
et al., 2009; Mason et al., 2003; Obleser et al., 2011; Rodd et al., 2005; Roskies et al., 2001;
Sabsevitz et al., 2005; Stromswold et al., 1996; Wilson et al., 2016). Ventral temporal language
regions extending along the fusiform gyrus were also modulated by linguistic demand.
In contrast, the core language regions of the lateral temporal lobe presented a more complex
picture. These regions were not modulated by linguistic demand in either the whole brain anal-
ysis or the ROI analysis. Several previous studies employing semantic tasks have similarly shown
no modulation of temporal lobe regions by task difficulty (Binder et al., 2005; Noppeney & Price,
2004; Sabsevitz et al., 2005; but cf. Badre et al., 2005 for a positive finding). One possible in-
terpretation is that these regions are recruited for semantic processing in an “all or none” manner
(Sabsevitz et al., 2005). However, our preferred interpretation builds on the observation that
temporal lobe regions are involved in conceptual processing even in the resting state (Binder
et al., 1999). We propose that linguistic demand as manipulated in the present study may shift
the relative balance of introspective versus externally cued semantic processing, without greatly
affecting the overall extent of such processing (see also McKiernan et al., 2003).
Importantly, left lateral temporal language regions were deactivated by perceptual process-
ing relative to rest, and were deactivated further still in the Difficult Perceptual condition. These
patterns can be interpreted as reflecting the inhibition of conceptual processing by an attention-
demanding task (Binder et al., 1999; McKiernan et al., 2003; Shulman et al., 1997). The lack of
Neurobiology of Language
218
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
modulation by linguistic demand in combination with differential deactivation by perceptual
demand yielded significant domain by difficulty interactions. If linguistic demand is conceptu-
alized as a relative concept, with non-linguistic demand as a point of reference, then we can
argue that these left temporal lobe language regions are in fact modulated by linguistic demand,
but that this is masked in the simple linguistic demand contrast because of the simultaneous
decrease in introspective conceptual processing that occurs as a general function of increasing
task difficulty.
How Regions Outside the MD and Language Networks Respond to Linguistic Demand
Most regions modulated by linguistic demand belonged to either the MD network or the
language network. In the voxelwise analysis, the largest cluster meeting neither of these criteria
was localized to the right IFG, between the pars opercularis and pars triangularis. This region
was homotopic to a left hemisphere language region, and was surrounded by MD regions in the
anterior insula, inferior frontal junction, and middle frontal gyrus, but was not itself modulated by
non-linguistic demand. The modulation of the right IFG by linguistic demand but not by percep-
tual demand was confirmed in the ROI analysis.
This positive finding of right hemisphere recruitment as language processing becomes more
demanding stands in contrast to two previous studies that sought to identify right hemisphere
recruitment for greater linguistic demand using functional transcranial Doppler sonography
(Dräger & Knecht, 2002) and fMRI (Dräger et al., 2004); however, it is possible that the more
difficult condition in those studies did not result in a greater extent of language processing, since
a word completion task was used, and participants generated more words in the easier condi-
tion. Another recent study found that the IFG maintained a lateralized activation pattern as the
difficulty of a verbal working memory task increased, unlike other prefrontal regions in which
activations became more bilateral with increasing task difficulty (Höller-Wallscheid et al.,
2017). However, in that study, the maintained lateralization pattern reflected modulation of both
left and right IFG by task difficulty, consistent with our findings.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
Right hemisphere temporal lobe regions, as well as the wider bilateral semantic network,
which is closely related to the default mode network (Binder et al., 2009), showed a similar pat-
tern to left temporal language regions. These regions were not modulated by linguistic demand,
but they were negatively modulated by non-linguistic demand, and therefore significant domain
by difficulty interactions were observed in whole brain and ROI analyses. Similar to left temporal
regions, a relative conception of linguistic demand would imply that these regions actually are in
a sense modulated by linguistic demand, in that they do not deactivate as a function of difficulty,
as would be expected in a non-linguistic task.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Challenges in Operationalizing Linguistic Demand
The most significant limitations of our study relate to the operationalization of linguistic demand.
First and foremost, we investigated only one type of language task: a semantic decision task. Our
rationale for choosing this task as a proxy for language processing in general was that it has pre-
viously been demonstrated to activate core language regions in a reliable and valid manner
(Wilson et al., 2018). Some partial support for the choice of this task as a proxy for language
processing in general comes from our finding that the language regions most strongly modulated
by linguistic demand in this task were the pars triangularis and opercularis of the IFG, neither of
which are generally considered semantic regions (Binder et al., 2009). But the language system
has many subcomponents and there are many different kinds of linguistic demand (Deschamps
Neurobiology of Language
219
Linguistic and non-linguistic demand
& Tremblay, 2014; Graves et al., 2010; McGettigan et al., 2011; Wilson et al., 2009). Some
linguistic processes depend on regions outside the core language network identified by the
semantic decision task, such as phonological encoding, which depends on the left supramargi-
nal gyrus (Price et al., 1997; Yen et al., 2019). Moreover, even language domains that depend
predominantly on the core regions identified by our task may differ from our task in terms of
modulation by demand. For example, a number of studies that have shown that syntactic
complexity modulates not just inferior frontal cortex (consistent with our findings) but also the
posterior temporal language region (which was not significantly modulated in the present study)
(I. Blank et al., 2016; Wilson et al., 2010).
Second, the Semantic Easy and Semantic Difficult conditions differ not just in the demands
they make on the linguistic system, but also in the demands they make on the conceptual
semantic system. This is relevant to interpreting the domain by difficulty interaction effects we
observed throughout the bilateral semantic network, which are more likely to reflect modulation
by “semantic demand” rather than linguistic demand per se.
Third, the linguistic and non-linguistic conditions differ in ways other than the presence
versus absence of linguistic content. One is a task of conceptual matching, the other a task of
judging perceptual identity. Experientially, the semantic task involves retrieving meanings for
each word, and then searching for any possible connection between them. In contrast, the per-
ceptual identity task entails looking back and forth between the two stimuli, searching for dif-
ferences. In the semantic task, a successful search leads to a button press, while in the perceptual
task, a successful search (i.e., the identification of a difference between the symbol strings) leads
to the withholding of a button press. These structural differences between the tasks explain why
most of the errors in the Semantic Difficult condition were misses, while most of the errors in the
Perceptual Difficult condition were false alarms. While an additional explanatory variable
accounting for the difference between “go” and “no-go” trials did not substantially change
any of our findings, it is nevertheless a limitation that the tasks, while structured similarly,
entailed somewhat different strategies.
Finally, our study was focused on modulation by demand, and not on initial recruitment of
MD regions or other regions in the easy conditions. There were potentially informative differ-
ences between the linguistic and non-linguistic conditions even on the easy conditions. For
example, the left inferior frontal junction showed more activity for the Semantic Easy condition
than the Perceptual Easy condition, while the reverse was true of the right inferior frontal junc-
tion. It is possible that regions that were more strongly activated for the easy conditions may have
had less potential to demonstrate further increases in activation for the difficult conditions.
Implications for Studies of Neuroplasticity in Aphasia
Notwithstanding the limitations described in the previous section, our findings have clear impli-
cations for the interpretation of functional imaging studies of neuroplasticity in aphasia.
Variables associated with task difficulty such as accuracy and reaction time have dramatic
effects on functional signal (Binder et al., 2005; Fedorenko et al., 2013; Yarkoni et al., 2009),
and task difficulty has been shown to modulate extent of activation in individuals with aphasia
(Fridriksson & Morrow, 2005). Recently, Geranmayeh et al. (2014) suggested that many activa-
tions that have been attributed to reorganization of language processing may instead reflect
effects of task difficulty. They speculated that such effects might be observed in several domain-
general networks: the dorsal attention network, cingulo-opercular network, and fronto-parietal
control network. Our findings—that there are major differences between the neural correlates of
linguistic and non-linguistic demand—suggest some refinements to this argument. Our findings
Neurobiology of Language
220
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
provide a more specific set of expectations about which brain regions might be expected to show
activation due to task difficulty effects: Some MD regions are more strongly modulated by
linguistic demand than others, and not just MD regions are modulated by linguistic demand,
but also some language regions, especially left frontal regions, along with the homotopic region
in the right IFG.
Geranmayeh et al. (2014) addressed several specific reported findings. They argued that the
transient upregulation of the right anterior insula reported by Saur et al. (2006) might reflect
increased activity related to task difficulty in a node of the cingulo-opercular network. Our
findings confirm that the right insula is indeed robustly modulated by linguistic demand. In
another case study, Geranmayeh et al. (2014) suggested that an atypical functional response
profile in the right STS reported by Leff et al. (2002) might reflect engagement of a node of the
ventral attention network. We found no evidence for modulation of the right STS or any nearby
region by linguistic demand, suggesting that this finding is unlikely to reflect a task difficulty
confound.
The right IFG, specifically the pars opercularis, is the brain region that has most often been
reported to show increased activation in individuals with aphasia relative to matched control
participants (S. C. Blank et al., 2003; Wilson & Schneck, 2021). The present finding that this
region is modulated by linguistic demand suggests that this often-replicated finding may reflect
the increased task difficulty experienced by individuals with aphasia.
We concur with Geranmayeh et al. (2014) in advocating caution in the interpretation of find-
ings localized to MD regions, or in other regions that are modulated by linguistic demand, but it
is also important to emphasize that such activations are not necessarily artifactual. It is plausible
that the neural changes that support recovery from aphasia may include upregulation or even
repurposing of domain-general regions, and that this may represent genuine compensatory re-
organization rather than simply task performance confounds (Brownsett et al., 2014; DeMarco
et al., 2018; Geranmayeh et al., 2017). It may prove difficult to distinguish between these two
interpretations.
In general, in studies of neuroplasticity in aphasia, the interpretation of activation differences
between groups, or correlations with behavior or time, remains complex and challenging. In
many cases, performance on the control conditions will also need to be considered, which will
entail considering the nature of interactions of domain by difficulty, which as we have shown
yield a somewhat different picture than linguistic demand alone.
A promising direction for future research will be to investigate the neural correlates of linguis-
tic demand in individuals with aphasia (Fridriksson & Morrow, 2005). We studied only neuro-
logically normal individuals; moreover, our participants were younger and more educated
than the typical stroke population. The lesions that cause aphasia almost always impact the
MD network as well, so it is an important open question how damage to various nodes of both
networks impacts the recruitment of surviving brain regions as a function of linguistic demand.
ACKNOWLEDGMENTS
We thank Michael de Riesthal, Evgeniia Diachek, Melissa Duff, Jillian Entrup, Evelina
Fedorenko, Deborah Levy, Sarah Schneck, and Lucina Uddin for valuable discussions and
input, Marisa Bush, Josh Hageman, Clair Jones, Leslie McIntosh, and Chris Thompson for assis-
tance with acquisition of MRI data, two anonymous reviewers for constructive feedback, and the
individuals who participated in the study. This research was supported in part by the National
Institute on Deafness and Other Communication Disorders.
Neurobiology of Language
221
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
FUNDING INFORMATION
Stephen M. Wilson, National Institute on Deafness and Other Communication Disorders (http://
dx.doi.org/10.13039/100000055), Award ID: R01 DC013270. Stephen M. Wilson, National
Institute on Deafness and Other Communication Disorders (http://dx.doi.org/10.13039
/100000055), Award ID: R21 DC016080.
AUTHOR CONTRIBUTIONS
Ian A. Quillen: Methodology; Investigation; Formal analysis; Visualization; Writing – original
draft; Writing – review & editing. Melodie Yen: Methodology; Investigation; Writing – review &
editing. Stephen M. Wilson: Conceptualization; Methodology; Formal analysis; Visualization;
Software; Writing – original draft; Writing – review & editing; Funding acquisition.
REFERENCES
Ashburner, J., & Friston, K. J. (2005). Unified segmentation. NeuroImage,
26, 839–851. DOI: https://doi.org/10.1016/j.neuroimage.2005
.02.018, PMID: 15955494
Badre, D., Poldrack, R. A., Paré-Blagoev, E. J., Insler, R. Z., &
Wagner, A. D. (2005). Dissociable controlled retrieval and gen-
eralized selection mechanisms in ventrolateral prefrontal cortex.
Neuron, 47, 907–918. DOI: https://doi.org/10.1016/j.neuron
.2005.07.023, PMID: 16157284
Beckmann, C. F., & Smith, S. M. (2004). Probabilistic independent
component analysis for functional magnetic resonance imaging.
IEEE Transactions on Medical Imaging, 23, 137–152. DOI:
https://doi.org/10.1109/TMI.2003.822821, PMID: 14964560
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral Cortex,
19, 2767–2796. DOI: https://doi.org/10.1093/cercor/bhp055,
PMID: 19329570, PMCID: PMC2774390
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S. F., Rao,
S. M., & Cox, R. W. (1999). Conceptual processing during the
conscious resting state: A functional MRI study. Journal of
Cognitive Neuroscience, 11, 80–93. DOI: https://doi.org/10
.1162/089892999563265, PMID: 9950716
Binder, J. R., Liebenthal, E., Possing, E. T., Medler, D. A., & Ward,
B. D. (2004). Neural correlates of sensory and decision pro-
cesses in auditory object identification. Nature Neuroscience,
7, 295–301. DOI: https://doi.org/10.1038/nn1198, PMID:
14966525
Binder, J. R., Medler, D. A., Desai, R., Conant, L. L., & Liebenthal,
E. (2005). Some neurophysiological constraints on models of
word naming. NeuroImage, 27, 677–693, DOI: https://doi.org/10
.1016/j.neuroimage.2005.04.029, PMID: 15921937
Blank, I., Balewski, Z., Mahowald, K., & Fedorenko, E. (2016). Syntactic
processing is distributed across the language system. NeuroImage,
127, 307–323. DOI: https://doi.org/10.1016/j.neuroimage
.2015.11.069, PMID: 26666896, PMCID: PMC4755877
Blank, I. A., & Fedorenko, E. (2017). Domain-general brain regions
do not track linguistic input as closely as language-selective
regions. Journal of Neuroscience, 37, 9999–10011. DOI: https://
doi.org/10.1523/JNEUROSCI.3642-16.2017, PMID: 28871034,
PMCID: PMC5637122
Blank, I. A., Kiran, S., & Fedorenko, E. (2017). Can neuroimaging
help aphasia researchers? Addressing generalizability, variability,
and interpretability. Cognitive Neuropsychology, 34, 377–393.
DOI: https://doi.org/10.1080/02643294.2017.1402756, PMID:
29188746, PMCID: PMC6157596
Blank, S. C., Bird, H., Turkheimer, F., & Wise, R. J. S. (2003).
Speech production after stroke: The role of the right pars opercu-
laris. Annals of Neurology, 54, 310–320. DOI: https://doi.org/10
.1002/ana.10656, PMID: 12953263
Blumstein, S. E., Myers, E. B., & Rissman, J. (2005). The perception
of voice onset time: An fMRI investigation of phonetic category
structure. Journal of Cognitive Neuroscience, 17, 1353–1366.
DOI: https://doi.org/10.1162/0898929054985473, PMID:
16197689
Bornkessel, I., Zysset, S., Friederici, A. D., Von Cramon, D. Y., &
Schlesewsky, M. (2005). Who did what to whom? The neural
basis of argument hierarchies during language comprehension.
NeuroImage, 26, 221–233. DOI: https://doi.org/10.1016/j
.neuroimage.2005.01.032, PMID: 15862222
Brownsett, S. L. E., Warren, J. E., Geranmayeh, F., Woodhead, Z.,
Leech, R., & Wise, R. J. S. (2014). Cognitive control and its impact
on recovery from aphasic stroke. Brain, 137, 242–254. DOI:
https://doi.org/10.1093/brain/awt289, PMID: 24163248,
PMCID: PMC3891442
Coltheart, M. (1981). The MRC psycholinguistic database. The
Quarterly Journal of Experimental Psychology A: Human
Experimental Psychology, 33A, 497–505. DOI: https://doi.org
/10.1080/14640748108400805
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215. DOI: https://doi.org/10.1038/nrn755,
PMID: 11994752
Corbetta, M., & Shulman, G. L. (2011). Spatial neglect and atten-
tion networks. Annual Review of Neuroscience, 34, 569–599.
DOI: https://doi.org/10.1146/annurev-neuro-061010-113731,
PMID: 21692662, PMCID: PMC3790661
Cox, R. W. (1996). AFNI: Software for analysis and visualization of
functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29, 162–173. DOI: https://doi.org/10
.1006/cbmr.1996.0014, PMID: 8812068
DeMarco, A. T., Wilson, S. M., Rising, K., Rapcsak, S. Z., & Beeson,
P. M. (2018). The neural substrates of improved phonological
processing following successful treatment in a case of phonolog-
ical alexia and agraphia. Neurocase, 24, 31–40. DOI: https://doi
.org/10.1080/13554794.2018.1428352, PMID: 29350575,
PMCID: PMC5843561
Neurobiology of Language
222
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
Deschamps, I., & Tremblay, P. (2014). Sequencing at the syllabic
and supra-syllabic levels during speech perception: An fMRI
study. Frontiers in Human Neuroscience, 8, 492. DOI: https://
doi.org/10.3389/fnhum.2014.00492, PMID: 25071521, PMCID:
PMC4086203
Diachek, E., Blank, I., Siegelman, M., & Fedorenko, E. (2020). The
domain-general multiple demand (MD) network does not support
core aspects of language comprehension: A large-scale fMRI
investigation. Journal of Neuroscience, 40, 4536–4550. DOI:
https://doi.org/10.1523/JNEUROSCI.2036-19.2020, PMID:
32317387, PMCID: PMC7275862
Dräger, B., Jansen, A., Bruchmann, S., Forster, A., Pleger, B.,
Zwitserlood, P., & Knecht, S. (2004). How does the brain accom-
modate to increased task difficulty in word finding? A functional
MRI study. NeuroImage, 23, 1152–1160. DOI: https://doi.org/10
.1016/j.neuroimage.2004.07.005, PMID: 15528114
Dräger, B., & Knecht, S. (2002). When finding words becomes dif-
ficult: Is there activation of the subdominant hemisphere?
NeuroImage, 16, 794–800. DOI: https://doi.org/10.1006/nimg
.2002.1095, PMID: 12169263
Duncan, J., & Owen, A. M. (2000). Common regions of the human
frontal lobe recruited by diverse cognitive demands. Trends in
Neurosciences, 23, 475–483. DOI: https://doi.org/10.1016
/S0166-2236(00)01633-7, PMID: 11006464
Eckert, M. A., Menon, V., Walczak, A., Ahlstrom, J., Denslow, S.,
Horwitz, A., & Dubno, J. R. (2009). At the heart of the ventral
attention system: The right anterior insula. Human Brain Mapping,
30, 2530–2541. DOI: https://doi.org/10.1002/hbm.20688, PMID:
19072895, PMCID: PMC2712290
Erb, J., Henry, M. J., Eisner, F., & Obleser, J. (2013). The brain dy-
namics of rapid perceptual adaptation to adverse listening con-
ditions. Journal of Neuroscience, 33, 10688–10697. DOI: https://
doi.org/10.1523/JNEUROSCI.4596-12.2013, PMID: 23804092,
PMCID: PMC6618499
Fedorenko, E., Duncan, J., & Kanwisher, N. (2012). Language-selective
and domain-general regions lie side by side within Broca’s area.
Current Biology, 22, 2059–2062. DOI: https://doi.org/10.1016
/j.cub.2012.09.011, PMID: 23063434, PMCID: PMC3494832
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad domain
generality in focal regions of frontal and parietal cortex. Proceedings
of the National Academy of Sciences of the United States of
America, 110, 16616–16621. DOI: https://doi.org/10.1073/pnas
.1315235110, PMID: 24062451, PMCID: PMC3799302
Fedorenko, E., Hsieh, P.-J., Nieto-Castañón, A., Whitfield-Gabrieli,
S., & Kanwisher, N. (2010). New method for fMRI investigations
of language: Defining ROIs functionally in individual subjects.
Journal of Neurophysiology, 104, 1177–1194. DOI: https://doi
.org/10.1152/jn.00032.2010, PMID: 20410363, PMCID:
PMC2934923
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen, D. C.,
& Raichle, M. E. (2005). The human brain is intrinsically organized
into dynamic, anticorrelated functional networks. Proceedings of
the National Academy of Sciences of the United States of
America, 102, 9673–9678. DOI: https://doi.org/10.1073
/pnas.0504136102, PMID: 15976020, PMCID: PMC1157105
Fridriksson, J., & Morrow, L. (2005). Cortical activation and language
task difficulty in aphasia. Aphasiology, 19, 239–250. DOI: https://
doi.org/10.1080/02687030444000714, PMID: 16823468,
PMCID: PMC1486765
Geranmayeh, F., Brownsett, S. L. E., & Wise, R. J. S. (2014). Task-
induced brain activity in aphasic stroke patients: What is driving
recovery? Brain, 137, 2632–2648. DOI: https://doi.org/10.1093
/brain/awu163, PMID: 24974382, PMCID: PMC4163030
Geranmayeh, F., Chau, T. W., Wise, R. J. S., Leech, R., &
Hampshire, A. (2017). Domain-general subregions of the medial
prefrontal cortex contribute to recovery of language after stroke.
Brain, 140, 1947–1958. DOI: https://doi.org/10.1093/brain
/awx134, PMID: 29177494, PMCID: PMC5903407
Graves, W. W., Desai, R., Humphries, C., Seidenberg, M. S., & Binder,
J. R. (2010). Neural systems for reading aloud: A multiparametric
approach. Cerebral Cortex, 20, 1799–1815. DOI: https://doi.org
/10.1093/cercor/bhp245, PMID: 19920057, PMCID: PMC2901017
Graves, W. W., Grabowski, T. J., Mehta, S., & Gordon, J. K. (2007).
A neural signature of phonological access: Distinguishing the
effects of word frequency from familiarity and length in overt
picture naming. Journal of Cognitive Neuroscience, 19, 617–631.
DOI: https://doi.org/10.1162/jocn.2007.19.4.617, PMID:
17381253
Höller-Wallscheid, M. S., Thier, P., Pomper, J. K., & Lindner, A.
(2017). Bilateral recruitment of prefrontal cortex in working
memory is associated with task demand but not with age.
Proceedings of the National Academy of Sciences of the
United States of America, 114, E830–E839. DOI: https://doi
.org/10.1073/pnas.1601983114, PMID: 28096364, PMCID:
PMC5293052
Ihnen, S. K. Z., Petersen, S. E., & Schlaggar, B. L. (2015). Separable
roles for attentional control sub-systems in reading tasks: A
combined behavioral and fMRI study. Cerebral Cortex, 25,
1198–1218. DOI: https://doi.org/10.1093/cercor/bht313, PMID:
24275830, PMCID: PMC4397571
Just, M. A., Carpenter, P. A., Keller, T. A., Eddy, W. F., & Thulborn,
K. R. (1996). Brain activation modulated by sentence compre-
hension. Science, 274(5284), 114–116. DOI: https://doi.org
/10.1126/science.274.5284.114, PMID: 8810246
Kelly, R. E., Jr., Alexopoulos, G. S., Wang, Z., Gunning, F. M.,
Murphy, C. F., Morimoto, S. S., Kanellopoulos, D., Jia, Z., Lim,
K. O., & Hoptman, M. J. (2010). Visual inspection of indepen-
dent components: Defining a procedure for artifact removal from
fMRI data. Journal of Neuroscience Methods, 189, 233–245.
DOI: https://doi.org/10.1016/j.jneumeth.2010.03.028, PMID:
20381530, PMCID: PMC3299198
Kuperman, V., Stadthagen-Gonzalez, H., & Brysbaert, M. (2012).
Age-of-acquisition ratings for 30,000 English words. Behavior
Research Methods, 44, 978–990. DOI: https://doi.org/10.3758
/s13428-012-0210-4, PMID: 22581493
Leff, A., Crinion, J., Scott, S., Turkheimer, F., Howard, D., & Wise, R.
(2002). A physiological change in the homotopic cortex following
left posterior temporal lobe infarction. Annals of Neurology, 51,
553–558. DOI: https://doi.org/10.1002/ana.10181, PMID: 12112100
Makuuchi, M., Bahlmann, J., Anwander, A., & Friederici, A. D.
(2009). Segregating the core computational faculty of human
language from working memory. Proceedings of the National
Academy of Sciences of the United States of America, 106,
8362–8367. DOI: https://doi.org/10.1073/pnas.0810928106,
PMID: 19416819, PMCID: PMC2688876
Mason, R. A., Just, M. A., Keller, T. A., & Carpenter, P. A. (2003).
Ambiguity in the brain: What brain imaging reveals about the
processing of syntactically ambiguous sentences. Journal of
Experimental Psychology: Learning, Memory, and Cognition,
29, 1319–1338. DOI: https://doi.org/10.1037/0278-7393
.29.6.1319, PMID: 14622064
McGettigan, C., Warren, J. E., Eisner, F., Marshall, C. R.,
Shanmugalingam, P., & Scott, S. K. (2011). Neural correlates of
sublexical processing in phonological working memory. Journal of
Cognitive Neuroscience, 23, 961–977. DOI: https://doi.org/10
.1162/jocn.2010.21491, PMID: 20350182, PMCID: PMC3376447
Neurobiology of Language
223
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
McKiernan, K. A., Kaufman, J. N., Kucera-Thompson, J., & Binder,
J. R. (2003). A parametric manipulation of factors affecting task-
induced deactivation in functional neuroimaging. Journal of
Cognitive Neuroscience, 15, 394–408. DOI: https://doi.org
/10.1162/089892903321593117, PMID: 12729491
Mollica, F., Siegelman, M., Diachek, E., Piantadosi, S. T., Mineroff,
Z., Futrell, R., Kean, H., Qian, P., & Fedorenko, E. (2020).
Composition is the core driver of the language-selective network.
Neurobiology of Language, 1, 104–134. DOI: https://doi.org/10
.1162/nol_a_00005
Noppeney, U., & Price, C. J. (2004). Retrieval of abstract semantics.
NeuroImage, 22, 164–170. DOI: https://doi.org/10.1016/j
.neuroimage.2003.12.010, PMID: 15110006
Obleser, J., Meyer, L., & Friederici, A. D. (2011). Dynamic assign-
ment of neural resources in auditory comprehension of complex
sentences. NeuroImage, 56, 2310–2320. DOI: https://doi.org
/10.1016/j.neuroimage.2011.03.035, PMID: 21421059
Piai, V., Roelofs, A., Acheson, D. J., & Takashima, A. (2013).
Attention for speaking: Domain-general control from the anterior
cingulate cortex in spoken word production. Frontiers in Human
Neuroscience, 7, 832. DOI: https://doi.org/10.3389/fnhum
.2013.00832, PMID: 24368899, PMCID: PMC3856851
Price, C. J., Crinion, J., & Friston, K. J. (2006). Design and analysis of
fMRI studies with neurologically impaired patients. Journal of
Magnetic Resonance Imaging, 23, 816–826. DOI: https://doi
.org/10.1002/jmri.20580, PMID: 16649208
Price, C. J., Moore, C. J., Humphreys, G. W., & Wise, R. J. S. (1997).
Segregating semantic from phonological processes during reading.
Journal of Cognitive Neuroscience, 9, 727–733. DOI: https://doi
.org/10.1162/jocn.1997.9.6.727, PMID: 23964595
Raboyeau, G., De Boissezon, X., Marie, N., Balduyck, S., Puel, M.,
Bézy, C., Démonet, J. F., & Cardebat, D. (2008). Right hemi-
sphere activation in recovery from aphasia: Lesion effect or
function recruitment? Neurology, 70, 290–298. DOI: https://
doi.org/10.1212/01.wnl.0000287115.85956.87, PMID:
18209203
Reppen, R., Ide, N., & Suderman, K. (2005). American National
Corpus (ANC) Second Release. Philadelphia, PA: Linguistic Data
Consortium.
Rodd, J. M., Davis, M. H., & Johnsrude, I. S. (2005). The neural
mechanisms of speech comprehension: fMRI studies of semantic
ambiguity. Cerebral Cortex, 15, 1261–1269. DOI: https://doi.org
/10.1093/cercor/bhi009, PMID: 15635062
Roskies, A. L., Fiez, J. A., Balota, D. A., Raichle, M. E., & Petersen,
S. E. (2001). Task-dependent modulation of regions in the left
inferior frontal cortex during semantic processing. Journal of
Cognitive Neuroscience, 13, 829–843. DOI: https://doi.org/10
.1162/08989290152541485, PMID: 11564326
Sabsevitz, D. S., Medler, D. A., Seidenberg, M., & Binder, J. R.
(2005). Modulation of the semantic system by word imageability.
NeuroImage, 27, 188–200. DOI: https://doi.org/10.1016/j
.neuroimage.2005.04.012, PMID: 15893940
Saur, D., Lange, R., Baumgaertner, A., Schraknepper, V., Willmes,
K., Rijntjes, M., & Weiller, C. (2006). Dynamics of language re-
organization after stroke. Brain, 129, 1371–1384. DOI: https://
doi.org/10.1093/brain/awl090, PMID: 16638796
Shain, C., Blank, I. A., van Schijndel, M., Schuler, W., Fedorenko, E.
(2020). fMRI reveals language-specific predictive coding during
naturalistic sentence comprehension. Neuropsychologia, 138,
107307. DOI: https://doi.org/10.1016/j.neuropsychologia
.2019.107307, PMID: 31874149, PMCID: PMC7140726
area. Annals of Neurology, 56, 836–846. DOI: https://doi.org
/10.1002/ana.20294, PMID: 15514975
Sharp, D. J., Turkheimer, F. E., Bose, S. K., Scott, S. K., & Wise, R. J. S.
(2010). Increased frontoparietal integration after stroke and cogni-
tive recovery. Annals of Neurology, 68, 753–756. DOI: https://doi
.org/10.1002/ana.21866, PMID: 20687116
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L., Miezin, F. M.,
Raichle, M. E., & Petersen, S. E. (1997). Common blood flow
changes across visual tasks: II. Decreases in cerebral cortex.
Journal of Cognitive Neuroscience, 9, 648–663. DOI: https://doi
.org/10.1162/jocn.1997.9.5.648, PMID: 23965122
Stromswold, K., Caplan, D., Alpert, N., & Rauch, S. (1996).
Localization of syntactic comprehension by positron emission
tomography. Brain and Language, 52, 452–473. DOI: https://
doi.org/10.1006/brln.1996.0024, PMID: 8653390
Thompson, C. K., Bonakdarpour, B., Fix, S. C., Blumenfeld, H. K.,
Parrish, T. B., Gitelman, D. R., & Mesulam, M.-M. (2007). Neural cor-
relates of verb argument structure processing. Journal of Cognitive
Neuroscience, 19, 1753–1767. DOI: https://doi.org/10.1162
/jocn.2007.19.11.1753, PMID: 17958479, PMCID: PMC2253656
Vaden, K. I., Kuchinsky, S. E., Cute, S. L., Ahlstrom, J. B., Dubno, J. R., &
Eckert, M. A. (2013). The cingulo-opercular network provides word-
recognition benefit. Journal of Neuroscience, 33, 18979–18986.
DOI: https://doi.org/10.1523/JNEUROSCI.1417-13.2013, PMID:
24285902, PMCID: PMC3841458
Wehbe, L., Blank, I. A., Shain, C., Futrell, R., Levy, R., von der
Malsburg, T., Smith, N., Gibson, E., & Fedorenko, E. (2020).
Incremental language comprehension difficulty predicts activity
in the language network but not the multiple demand network.
bioRxiv. DOI: https://doi.org/10.1101/2020.04.15.043844
Wilson, S. M., DeMarco, A. T., Henry, M. L., Gesierich, B., Babiak,
M., Miller, B. L., & Gorno-Tempini, M. L. (2016). Variable dis-
ruption of a syntactic processing network in primary progressive
aphasia. Brain, 139, 2994–3006. DOI: https://doi.org/10.1093
/brain/aww218, PMID: 27554388, PMCID: PMC5091045
Wilson, S. M., Dronkers, N. F., Ogar, J. M., Jang, J., Growdon, M. E.,
Agosta, F., Henry, M. L., Miller, B. L., & Gorno-Tempini, M. L.
(2010). Neural correlates of syntactic processing in the nonfluent
variant of primary progressive aphasia. Journal of Neuroscience,
30, 16845–16854. DOI: https://doi.org/10.1523/JNEUROSCI
.2547-10.2010, PMID: 21159955, PMCID: PMC3024013
Wilson, S. M., Eriksson, D. K., Yen, M., Demarco, A. T., Schneck, S. M.,
& Lucanie, J. M. (2019). Language mapping in aphasia. Journal of
Speech, Language, and Hearing Research, 62, 3937–3946. DOI:
https://doi.org/10.1044/2019_JSLHR-L-RSNP-19-0031, PMID:
31756153, PMCID: PMC7203526
Wilson, S. M., Isenberg, A. L., & Hickok, G. (2009). Neural corre-
lates of word production stages delineated by parametric modu-
lation of psycholinguistic variables. Human Brain Mapping, 30,
3596–3608. DOI: https://doi.org/10.1002/hbm.20782, PMID:
19365800, PMCID: PMC2767422
Wilson, S. M., & Schneck, S. M. (2021). Neuroplasticity in post-
stroke aphasia: A systematic review and meta-analysis of func-
tional imaging studies of reorganization of language processing.
Neurobiology of Language, 2, 22–82. DOI: https://doi.org/10
.1162/nol_a_00025
Wilson, S. M., Yen, M., & Eriksson, D. K. (2018). An adaptive
semantic matching paradigm for reliable and valid language
mapping in individuals with aphasia. Human Brain Mapping,
39, 3285–3307. DOI: https://doi.org/10.1002/hbm.24077,
PMID: 29665223, PMCID: PMC6045968
Sharp, D. J., Scott, S. K., & Wise, R. J. S. (2004). Retrieving meaning
after temporal lobe infarction: The role of the basal language
Winkler, A. M., Ridgway, G. R., Webster, M. A., Smith, S. M., &
Nichols, T. E. (2014). Permutation inference for the general linear
Neurobiology of Language
224
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Linguistic and non-linguistic demand
model. NeuroImage, 92, 381–397. DOI: https://doi.org/10.1016/j
.neuroimage.2014.01.060, PMID: 24530839, PMCID: PMC4010955
Worsley, K. J., Liao, C. H., Aston, J., Petre, V., Duncan, G. H.,
Morales, F., & Evans, A. C. (2002). A general statistical analysis
for fMRI data. NeuroImage, 15, 1–15. DOI: https://doi.org/10
.1006/nimg.2001.0933, PMID: 11771969
Yarkoni, T., Barch, D. M., Gray, J. R., Conturo, T. E., & Braver, T. S.
(2009). BOLD correlates of trial-by-trial reaction time variability
in gray and white matter: A multi-study fMRI analysis. PLoS One,
4, e4257. DOI: https://doi.org/10.1371/journal.pone.0004257,
PMID: 19165335, PMCID: PMC2622763
Yen, M., DeMarco, A. T., & Wilson S. M. (2019). Adaptive para-
digms for mapping phonological regions in individual partici-
pants. NeuroImage, 189, 368–379. DOI: https://doi.org/10
.1016/j.neuroimage.2019.01.040, PMID: 30665008, PMCID:
PMC6424113
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
0
2
1
9
1
1
8
6
2
n
o
_
a
_
0
0
0
3
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
225