RESEARCH ARTICLE
Neural Representation of Articulable and
Inarticulable Novel Sound Contrasts:
The Role of the Dorsal Stream
a n o p e n a c c e s s
j o u r n a l
David I. Saltzman and Emily B. Myers
University of Connecticut, Storrs, CT
Keywords: speech perception, fMRI, multivoxel pattern analysis, speech articulation, dorsal stream,
phonetics
ABSTRACT
The extent that articulatory information embedded in incoming speech contributes to the
formation of new perceptual categories for speech sounds has been a matter of discourse for
decades. It has been theorized that the acquisition of new speech sound categories requires
a network of sensory and speech motor cortical areas (the “dorsal stream”) to successfully
integrate auditory and articulatory information. Tuttavia, it is possible that these brain regions
are not sensitive specifically to articulatory information, but instead are sensitive to the abstract
phonological categories being learned. We tested this hypothesis by training participants over
the course of several days on an articulable non-native speech contrast and acoustically
matched inarticulable nonspeech analogues. After reaching comparable levels of proficiency
with the two sets of stimuli, activation was measured in fMRI as participants passively listened
to both sound types. Decoding of category membership for the articulable speech contrast
alone revealed a series of left and right hemisphere regions outside of the dorsal stream that
have previously been implicated in the emergence of non-native speech sound categories,
while no regions could successfully decode the inarticulable nonspeech contrast. Although
activation patterns in the left inferior frontal gyrus, the middle temporal gyrus, and the
supplementary motor area provided better information for decoding articulable (speech)
sounds compared to the inarticulable (sine wave) sounds, the finding that dorsal stream
regions do not emerge as good decoders of the articulable contrast alone suggests that other
factors, including the strength and structure of the emerging speech categories are more likely
drivers of dorsal stream activation for novel sound learning.
INTRODUCTION
Whether an infant learning her first language, or an adult learning his fifth language, in lan-
guage acquisition the learner must learn to perceive as well as produce new speech sounds.
In typical language acquisition, these two processes (perceptual and articulatory learning)
develop in tandem, although not necessarily on the same timeline (Tsao, Liu, & Kuhl,
2004). Tuttavia, in laboratory conditions, we know that individuals can learn to distinguish
complex auditory speech and nonspeech categories without learning to produce these same
categorie. At issue is the degree to which articulatory information associated with speech is
recruited even in the absence of articulatory learning. Questo è, when one learns a new pho-
netic category without learning to produce the same category, are speech motor represen-
tations recruited?
Citation: Saltzman, D. I., & Myers, E. B.
(2020). Neural representation of
articulable and inarticulable novel
sound contrasts: The role of the dorsal
stream. Neurobiology of Language,
1(3), 339–364. https://doi.org/10.1162/
nol_a_00016
DOI:
https://doi.org/10.1162/nol_a_00016
Supporting Information:
https://doi.org/10.1162/nol_a_00016
Received: 21 ottobre 2019
Accepted: 23 May 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
David I. Saltzman
david.saltzman@uconn.edu
Handling Editor:
Patrick Wong
Copyright: © 2020 Massachusetts
Institute of Technology. Pubblicato
under a Creative Commons Attribution
4.0 Internazionale (CC BY 4.0) licenza.
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
Models of speech perception like the motor theory of speech (Liberman, Cooper,
Shankweiler, & Studdert-Kennedy, 1967) and subsequent direct realist approaches (Best,
1995; Fowler, 1986), make explicit predictions about the formative role of articulatory codes
(cioè., the gestures made by the articulators to create a speech sound) in speech perception. In
these models, motor or articulatory representations are the objects of perception, and must be
acquired and then accessed to achieve robust comprehension (see Galantucci, Fowler, &
Turvey, 2006 for a review). Così, learning to discriminate between a pair of speech sounds
like /b/ and /d/ (which are similar acoustically) requires that the listener access information the
articulatory gestures used to produce those sounds.
Though not explicitly linked, the dual stream model of speech processing (Hickok & Poeppel,
2007) makes biological predictions that also implicate motor systems not in speech sound
perception, but in speech sound learning. Specifically, the dorsal stream pathway, consisting of
such brain regions as the inferior frontal gyrus (IFG), the pre-motor cortex, and the temporal-
parietal junction, is claimed to play a critical role in auditory-motor integration and serves as a
“neural mechanism that both codes and maintains instances of speech sounds,” which can then
be later used to aid in production as well as in perception (Hickok & Poeppel, 2007, P. 399).
According to this view, during the learning of new speech sounds, listeners code the incoming
articulatory gestures in dorsal stream areas in preparation for future speech output (Hickok &
Buchsbaum, 2003). Notably, once speech sound representations have been learned, dorsal stream
activation is no longer thought to be required during perception-only tasks.
The role of the dorsal stream during speech perception (for established speech categories and
newly learned ones) has been a matter of significant debate, as has the role of articulatory codes
during speech perception in general (per esempio., Lotto, Hickok, & Holt, 2009; Schomers &
Pulvermüller, 2016; Wilson, 2009; Scott, McGettigan, & Eisner, 2009). If articulatory codes
are accessed as a matter of course during perception of speech categories, it should follow that
the encompassed brain regions are sensitive to differences in articulation between speech
sounds during speech perception, especially during speech sound learning (see Meister,
Wilson, Deblieck, Wu, & Iacoboni, 2007; Wilson & Iacoboni, 2006; Wilson, Saygin, Sereno,
& Iacoboni, 2004). While evidence for sensitivity to place/manner of articulation differences is
apparent in the left superior temporal gyrus (STG) using electrocorticography (Mesgarani,
Cheung, Johnson, & Chang, 2014), it is less clear that dorsal stream speech motor regions are
involved in the perception of established, mature speech categories. Using a univariate fMRI
approach, Pulvermüller et al. (2006) found differential activation in lip- and tongue-dedicated
regions of the precentral gyrus in response to the syllables /p/ and /t/, which differ in place of
articulation. These results were supported by findings from Correia, Jansma, and Bonte
(2015), who used a multivoxel pattern analysis (MVPA) to identify brain regions that could clas-
sify neural responses to stimuli based on specific articulatory features independent of phonemic
categoria (and subsequent acoustic variation). Activation patterns in pre-motor and motor areas
(among other regions) could distinguish place and manner of articulation contrasts, even though
the classifier was trained on responses to one phonological category (stop consonants) E
tested on responses to another (fricatives). Tuttavia, both Arsenault and Buchsbaum (2015)
and Arsenault and Buchsbaum (2016) failed to find any dorsal stream regions that were sensitive
to different articulatory dimensions of passively heard syllables in both univariate (attempting
to replicate Pulvermüller et al.’s results) and multivariate analyses, calling into question the
sensitivity of the dorsal stream to learned phonological contrasts.
As noted above, there is contradictory evidence that dorsal stream regions are sensitive to
articulatory contrasts during speech perception. Tuttavia, even in the studies that support the
Neurobiology of Language
340
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
activation of the dorsal stream during speech processing, an alternative explanation exists: È
possible that what is being observed is IFG sensitivity to abstract sound categories at large, E
not specifically the articulatory differences between sounds. Questo è, during speech perception,
inferior frontal structures may not be coding articulatory differences between sounds (per esempio., IL
difference in lip protrusion in the production of /i/ and /y/), but instead reflect the ultimate
product of those gestures, which are two distinct phonetic categories. IFG sensitivity to speech
categories has been established in a number of studies (Hutchison, Blumstein, Myers, 2008;
Lee, Turkeltaub, Granger, & Raizada, 2012; Myers, 2007; Myers, Blumstein, Walsh, &
Eliassen, 2009), but these studies do not disentangle the issue of articulatory codes versus ab-
stract categories as the two are fundamentally confounded in established speech categories. In
each of these previous studies, participants were presented with isolated speech sounds in
their native language. These well-established perceptual categories may activate articulatory
representations of the same sound by association rather than as a means to perceive the sound
itself. Questo è, upon hearing “da,” one may automatically activate gestural codes for producing
“da,” whether or not this articulatory information is necessary for retrieving the speech sound
categoria. Further, as argued in the dual stream model, the dorsal stream may only be recruited
during learning, when participants are learning the links between auditory and motor repre-
sentations of sounds. Così, to fully test the predictions of this model, it is necessary to present
listeners with a situation in which a new speech category is acquired alongside an acoustically
matched, inarticulable contrast in order to fully establish what dorsal stream activation may
actually reflect.
Liebenthal, Binder, Spitzer, Possing, and Medler (2005) and Liebenthal et al. (2010) ex-
plored this idea by contrasting brain activation from familiar speech sounds (a continuum from
/b/ to /d/) with that of an acoustically unfamiliar matched nonspeech continuum (created by
spectrally inverting the first formant of the speech continuum). Although when participants had
minimal exposure to the speech and nonspeech continua (Liebenthal et al., 2005), no dorsal
stream differences emerged, after participants were trained on the same stimuli for four days,
(Liebenthal et al., 2010), some dorsal stream recruitment was seen. That dorsal stream recruit-
ment arises only during the learning process is consistent with the dual stream model, but par-
adoxically, dorsal stream engagement is not preferentially seen for speech (articulable) stimuli.
Invece, several significant clusters in the left IFG, the bilateral postcentral gyrus, and the bi-
lateral supplementary motor area display greater activation at posttest for a nonspeech contin-
uum that could not be articulated by a human vocal tract. In a conjunction analysis, in which
the neural changes as a result of training were compared across the speech and nonspeech
continua, the left IFG was identified as sensitive specifically to learning in the nonspeech con-
tinuum. This provides further evidence that during learning, IFG activation more likely reflects
the emergent category status of the nonspeech stimuli as listeners begin to form perceptual
categories rather than a reflection of articulatory codes (Myers et al., 2009).
Desai, Liebenthal, Waldron, and Binder (2008), used a similar paradigm to Liebenthal et al.
(2005), but critically substituted sine-wave speech versions of both the speech (/b/-/d/) and non-
speech continua (spectrally rotated /b/-/d/). Adopting the labels from this study, we will refer to
them hereafter as “phonemic” and “nonphonemic,” respectively, to differentiate from the nat-
ural speech versions of these stimuli in previous studies. Sine-wave speech is created by taking
the spectral center and power of each of the formants in a speech sound, and then creating a
sine-wave tone at the same frequency and power (Remez, Rubin, Pisoni, & Carrell, 1981). IL
resulting sound contains approximately the same distinctive spectral information as the original
speech sound but is not perceived as speech or speechlike until the listener is informed of the
intended message, after which listeners often report being able to understand the message. These
Neurobiology of Language
341
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
stimuli allowed the authors to control for familiarity and to manipulate the availability of artic-
ulatory information by revealing to participants midway through the experiment that one of the
continua (the phonemic continuum) was made from familiar speech sounds. Dorsal stream
recruitment was observed at posttest, where greater activation in the left IFG and the left precen-
tral gyrus was seen for the phonemic continuum compared to the nonphonemic continuum. It
seems possible that after being informed that the phonemic continuum was supposed to reflect
two familiar speech sound categories, listeners began to recover some of the articulatory code
used to create those sounds, and as a result the dorsal stream was recruited during learning. Ma
Ancora, this possibility does not exclude that neural activation reflects the emergence of two
distinct categories, as perception of the phonemic continuum shifted from a continuous mode
to a categorical mode during posttest.
The Current Study
To investigate the role of the dorsal stream during the learning of new speech sounds, we at-
tempted to equate the degree of proficiency with the two stimulus sets being learned, anche
as to manipulate articulatory feasibility. While both variables have been manipulated indepen-
dently in previous studies, to our knowledge no study has controlled for both of these variables
simultaneously. We take a similar approach to Liebenthal et al. (2005, 2010) by introducing a set
of articulable (nonnative speech) versus inarticulable (impoverished sine-wave speech) sounds.
Like Desai et al. (2008), we used sounds that did not initially have established categories for our
listeners, therefore requiring participants to learn both sets of sounds (unlike Liebenthal et al.,
2005, 2010). Tuttavia, in Desai et al., articulatory information is only indirectly implied in their
phonemic continuum, as listeners must recover a phonological category from sine-wave
speech. There is no guarantee that participants are activating articulatory representations of
the source stimuli, and results could reflect listeners treating the sine-wave speech as inarticul-
able sounds but with a defined category structure. Therefore, we chose to use a non-native
speech sound contrast to ensure that articulatory information is theoretically available, while
the contrast is still as unfamiliar as the impoverished sine-wave speech contrast. Listeners are
then trained to the same accuracy on both contrasts, after which two competing hypotheses
can be investigated. Primo, following the logic of the dual stream model of speech perception,
dorsal stream regions should only be recruited for auditory-motor integration during the learning
of articulable sounds. Alternatively, dorsal stream regions (especially the IFG) may play a more
general role in categorical decisions and will be equally recruited during the learning of both
articulable and inarticulable sounds (so long as the two sounds are equally well learned).
The question at hand is especially suited to the use of MVPA (Haxby et al., 2001) because
these techniques can provide a clue as to the nature of the information stored in different areas
of the cortex. If the patterns of activation are distinctive enough to classify stimulus types, Esso
implies that there is systematic neural representation for some property of that stimulus.
Further, the differences in activation in response to a contrast like /b/ and /t/ may not be lo-
cated in separate areas of the cortex, but may rather be better characterized by a distributed
pattern of activation (including nonadjacent voxels), the kind of informational code that MVPA
is sensitive to. By using signal decoding techniques, we ask not only whether articulatory ver-
sus auditory regions are recruited, but also whether the information content of those regions
distinguishes between the two categories in each contrast.
We attempted to answer this question by using a multiday training study in which participants
were trained to equal degrees of familiarity on an unfamiliar (but articulable) non-native phonetic
contrasto (the /i/ vs. /y/ contrast found in languages like French), as well as acoustically matched
Neurobiology of Language
342
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
impoverished sine-wave analogues of the same contrast. After three days of training, participants
then engaged in a passive fMRI task in which they heard tokens from the two continua they were
trained on. A series of univariate and multivariate analyses were performed on the fMRI data to
determine the neural architecture involved in the learning of new sounds, and how the articu-
latory feasibility of that sound impacts that architecture.
MATERIALS AND METHOD
Participants
Thirty-four undergraduate students were recruited from the University of Connecticut. All par-
ticipants indicated that they were right-handed, monolingual native English speakers with nor-
mal hearing and no history of neurological or language disorders. Handedness and hearing
abilities were self-reported by the participant, but not assessed in any other way. Eight partic-
ipants were excluded (because of failure to complete all four sessions, n = 5, incomplete MRI
dati, n = 2, withdrawal, n = 1), leaving data from 26 participants (ages = 18–22 years, females =
16, males = 10) for analysis. Written informed consent was obtained from every participant in
accordance with the guidelines of the University of Connecticut ethics committee. Participants
were paid for their participation.
Stimuli
Speech stimuli
The speech stimuli consisted of tokens taken from a seven-step vowel continuum from /i/ (front
unrounded vowel, found in English in words like “leaf”) to /y/ (front rounded vowel found in
languages such as French). This contrast was chosen because /y/ is not found in American
English phonology and monolingual English speakers consistently struggle to discriminate these
vowels without training (Levy & Strange, 2008; Darcy et al., 2012).
Natural speech tokens of /y/ and /i/ were produced by a male native speaker of Parisian
French, and the /y/ token was then transformed into synthesized speech using the to
KlattGrid function in Praat (Boersma & Weenink, 2020) to allow for precise control over
acoustic properties. F1, F2, and F4 were fixed at the average values derived from the natural
/y/ production, and F3 was systematically lowered in 7 equal Bark scale steps until the F3
value was equivalent to the naturally produced /i/ token’s F3 value, creating a continuum from
/y/ to /i/. F5 was removed as it is uninformative to the contrast of interest. The synthesized /i/
and /y/ endpoints were submitted for pilot testing to five native speakers of French, who ac-
curately identified the intended vowels and deemed them natural sounding. All tokens had a
duration of 432 ms and were scaled to an intensity of 84 dB. Stimuli were delivered over head-
phones at a volume adjusted by the participant to a loud yet comfortable listening level.
Sine-wave stimuli
Nonspeech acoustic analogues of the speech stimuli were created by transforming the vowel
continuum described in the previous section into sine-wave speech. This was achieved using
a Praat script created by Chris Darwin (available at http://www.lifesci.sussex.ac.uk/ home/
Chris_Darwin/Praatscripts/SWS). The resulting seven tokens were then stripped of the sine-wave
equivalents of F1 and F4, leaving only the critical F2–F3 contrast in place to reduce the likeli-
hood that they would be perceived as speech-like (Guarda la figura 1). The resultant two-formant sine
sweeps are thus acoustically matched to the diagnostic information in the vowel continuum
(the frequency of F3), but critically during initial pilot testing (N = 11) they were never indicated
Neurobiology of Language
343
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
Figura 1. Spectrograms of the first step of the vowel (left) and sine-wave (right) continua.
to be perceived as speech (participants frequently described them as “chirps,” “bird sounds,” or
“robot sounds”). The same pilot testing also revealed that a single session was sufficient for par-
ticipants to reach near ceiling accuracy in categorizing sine-wave analogue tokens using the
explicit perceptual fading training task described in the following sections (M = 97.7%, SD =
15%). All tokens had a duration of 432 ms and were scaled to an intensity of 80 dB.
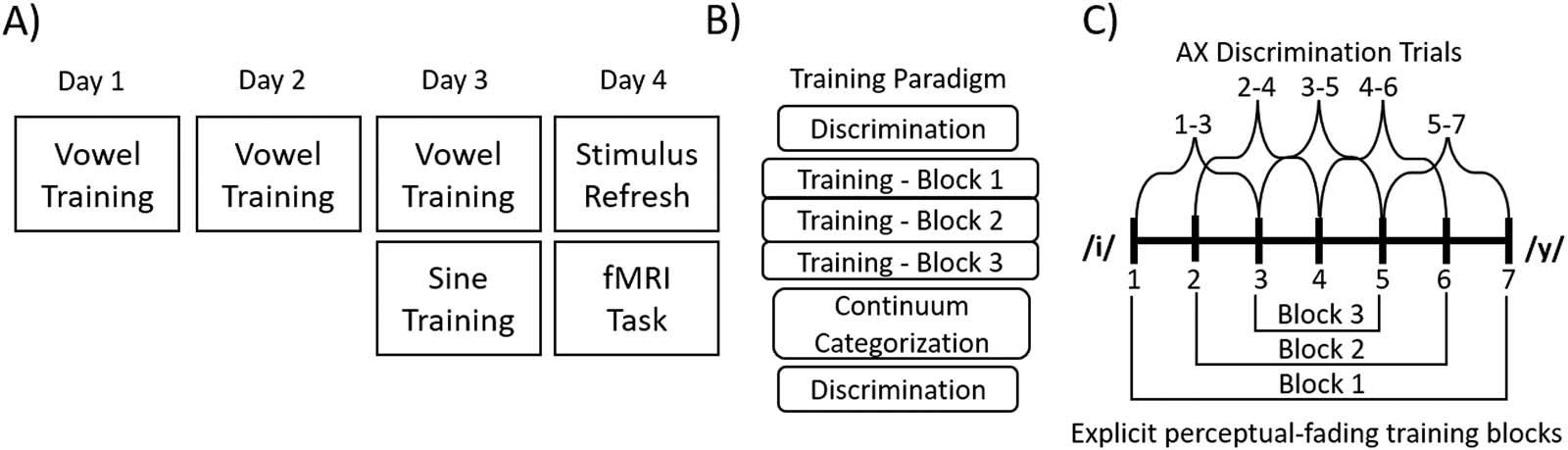
Experiment Schedule
The study took place over four consecutive days (Guarda la figura 2); on the first day, participants went
through the informed consent process and completed the training paradigm for the vowel to-
kens. On the second day, participants returned and repeated training for the vowel tokens. On
the third day, participants again repeated the vowel training followed by the sine-wave training
paradigm. Finalmente, on the fourth day participants completed a short refresher on both stimulus
sets, in which participants were refamiliarized with the correspondence between continuum
sounds and the category they map on to, and then completed the fMRI task. The discrepancy
in the number of training sessions between the vowel stimuli and the sine-wave stimuli was a
result of earlier pilot testing, which indicated that it took participants one session to reach a
threshold of at least 80% accuracy in the most difficult training block on the sine-wave stimuli,
but three days to reach the same criteria for the vowel stimuli. The fMRI task was completed on a
fourth day to allow for consolidation of the recently learned sine-wave stimuli, as sleep consol-
idation has been shown to impact non-native phonetic learning (Earle & Myers, 2015).
Behavioral Training Regimen
The behavioral training paradigm consisted of three subtasks that were identical in structure
across both Vowel and Sine training. OpenSesame 3.2.4 (Mathôt, Schreij, & Theeuwes, 2012)
Figura 2.
(UN) Schedule of tasks completed over the course of the four days of training. (B) Schedule of the subtasks that comprised both the
vowel and the sine-wave training sessions. (C) Overview of the stimulus pairs used in the discrimination (above the horizontal continuum line)
and the explicit perceptual fading training (below the horizontal continuum line).
Neurobiology of Language
344
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
was used for stimulus presentation, and all responses were made via a Cedrus RB-840 response
box. Participants wore Sony MDR-7506 headphones and were allowed to adjust volume to
their most comfortable listening level. All response options were counterbalanced across
participants.
Discrimination
At both the beginning and the end of the behavioral training paradigm, participants completed
an identical AX discrimination task in which they were presented with a pair of tokens from the
appropriate stimulus set that were either identical (per esempio., step 1 vs. step 1) or two steps apart on
the continuum (per esempio., steps 1 vs. step 3; steps 4 vs. step 6) and asked to indicate as quickly and
accurately as possible if the two sounds were the same or different. All pairs were presented
both “forwards” (per esempio., step 1 followed by step 3) or “backwards” (per esempio., step 3 followed by step 1),
with an intertrial interval of 500 ms. There were seven pair types where the two tokens pre-
sented were identical and ten pair types where the stimuli differed. Each pair was presented
11 times, for a total of 187 trials. Accuracy of response and reaction time were both recorded
0
as outcomes of interest, and d
, an unbiased measure of sensitivity, was calculated from the
participants’ responses. Trials in which participants failed to respond before the trial timed out
(6,000 ms) were considered incorrect responses.
Perceptual fading training
Explicit training on the sound categories used a perceptual fading paradigm where participants
were asked to map the two halves of the continuum to a different colored square. On each
successive block, the acoustic difference between the two tokens that participants were asked
to identify was reduced. To familiarize participants with the two categories they would be
hearing, they were first shown a colored square and played 10 repetitions of the continuum
endpoint token that would thereon be arbitrarily associated with that colored square (per esempio., UN
red square would appear on the screen and the /i/ endpoint token would play).
In the first training block (labeled as “easy”), participants were asked to identify as quickly
and accurately as possible which colored square corresponded to step 1 and step 7. On the
second training block (labeled as “medium”), participants were asked to identify which col-
ored square corresponded to continuum step 2 and continuum step 6. On the third training
block (labeled as “hard”), participants were asked to identify which colored square corre-
sponded to continuum step 3 and continuum step 5, the tokens that lay on opposing sides
of the acoustic boundary. Notice that step 4, the midpoint of the continuum, was not presented
during training.
Each block consisted of 60 trials (30 repetitions of each token), and participants were re-
quired to achieve an 80% accuracy threshold in order to ascend to the next block. If partic-
ipants did not reach threshold by the end of the block, they would repeat that same block up to
three times before being forced to move on to the subsequent block. Therefore, a given par-
ticipant could experience between one and three repetitions of each block, depending on how
quickly they advanced to more difficult blocks. Participants received explicit feedback during
the task. Accuracy of response and reaction time were both recorded as outcomes of interest.
Continuum categorization
Following training, participants engaged in a two-alternative forced choice task in which they
were asked to identify the colored square that corresponded to the sound being presented, con
Neurobiology of Language
345
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
no feedback. There were 10 randomly presented repetitions of each token from the full seven-
step continuum, for a total of 70 trials. The proportion of step 7 responses (which corresponded
to the /y/Vowel and /y/Sine endpoints in the Vowel and Sine conditions respectively) per contin-
uum step were then calculated. Trials in which participants failed to respond were omitted from
analysis.
Imaging Methods
fMRI design and procedure
Before entering the scanner, participants completed a brief task in which they were refamiliar-
ized with the stimuli that would be presented during the fMRI task. Participants were first pre-
sented with a colored square while five repetitions of each of two tokens from that category
were played. Following this, participants engaged in a two-alternative forced choice (2AFC)
task in which they were asked to identify which colored square corresponded to the sound
presented. Each of the four tokens were repeated 30 times, and this was repeated for both
the Vowel tokens and the Sine tokens, leading to a total of 240 trials for the 2AFC task.
Explicit feedback was provided during this task.
Participants were instructed to listen attentively and told that on some trials a stimulus
would be presented more quietly than normal. When this happened, they should press any
button on the MRI-safe response devices held in both hands. These catch stimuli consisted
of a randomly chosen token from the eight Vowel and Sine tokens presented at 20 dB lower
intensity. Participants completed a brief training during the structural scan acquisition to famil-
iarize them with this task. Imaging data from catch trials were modeled in participant-level
analyses but did not contribute to group-level analyses. Participants were on average quite
accurate at detecting catch trials (M = 90%, SD = 30%). Stimuli were delivered by
OpenSesame 3.2.4. The volume was set to a comfortable listening level as determined by each
participant.
The fMRI experimental paradigm consisted of 10 runs, with run order randomized between
participants. Trials within each run were presented in a fixed, pseudorandom order determined
using the OptSeq2 tool (https://surfer.nmr.mgh.harvard.edu/optseq/). Steps 1, 3, 5, E 7 from
both the Vowel and Sine continua were each repeated 15 times per run, along with 15 stimuli
used for the catch task described in the previous paragraph, resulting in 150 presentations of
each stimulus over the course of the experiment. An auditory stimulus appeared in all but 25
of the silent gaps during scanning, for an average intertrial interval of 2,390 ms. Each run con-
sisted of 161 volumes.
Acquisition parameters
Anatomical and functional MRI data were collected with a 3-T Siemens Prisma scanner
(Erlangen, Germany). High resolution three dimensional T1-weighted anatomical images were
acquired using a multiecho magnetization prepared rapid gradient echo sequence (MPRAGE;
repetition time [TR] = 2,400 ms, echo time = 2.22 ms, inversion time = 1,000 ms, 0.8 mm3
isotropic voxels, 320 × 320 matrix) and reconstructed into 208 slices. Functional echo planar
images were acquired using an accelerated multiband sequence in ascending, interleaved
order (52 slices, 2.5 mm thick, 2 mm2 axial in-plane resolution, 110 × 110 matrix, 220 mm3
field of view, flip angle = 62°) and followed a fast, sparse sampling design: Each functional
volume was acquired with a 1,000 ms acquisition time, followed by 1,000 ms of silence during
Neurobiology of Language
346
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
which auditory stimuli were presented (effective TR = 2,000 ms). Stimuli were always presented
during the silent gap.
Univariate fMRI analyses
Data were analyzed using Analysis of Functional NeuroImages (AFNI; Cox, 1996). Preprocessing
for the univariate analyses consisted of first transforming images from oblique to cardinal orienta-
zione, followed by motion correction using a six-parameter rigid body transform aligned with each
participant’s anatomical data set, normalization to Talairach space (Talairach & Tournoux, 1988),
and spatial smoothing with a 4 mm Gaussian kernel. Masks were created using each participant’s
anatomical data to eliminate voxels located outside the brain. Individual masks were used to gen-
erate a group mask. Motion and signal fluctuation outliers were removed following standard
procedures.
We generated time series vectors for each of the eight continua tokens ( Vowel step 1,
Vowel step 3, Vowel step 5, Vowel step 7, Sine step 1, Sine step 3, Sine step 5, Sine step 7) COME
well as the catch trials and any false alarm trials for each participant in each run. These vectors
contained the onset time of each stimulus and were convolved with a stereotypic gamma hemo-
dynamic function. IL 10 condition vectors along with six additional nuisance movement
parameters were submitted to a regression analysis. This analysis generated by-voxel fit coeffi-
cients for each condition for each participant.
For group level analysis, beta coefficients were submitted to a 2 × 3 analysis of variance
(ANOVA) in AFNI (using 3dANOVA3). In the ANOVA, main effects of sound type ( Vowel vs.
Sine), continuum step (1 vs. 3 vs. 5 vs. 7), and their interaction were estimated.
Multivariate fMRI analyses
Data were preprocessed again using AFNI with a modified pipeline compared to the univariate
analyses. Specifically, multivariate preprocessing consisted of first transforming images from
oblique to cardinal orientation, followed by motion correction using a six-parameter rigid
body transform aligned with each participant’s anatomical data set. No blurring or spatial nor-
malization were performed until after the participant-level analysis to preserve fine-grained
spatial information. The same 10 condition vectors from the univariate analyses along with
six additional nuisance movement parameters were submitted to an individually modulated
regression analysis (using the stim_times_IM flag in AFNI’s 3dDeconvolve), in which each trial
for a condition is given its own amplitude estimation, yielding trial-wise beta coefficients (COME
opposed to run-wise beta coefficients, see Rissman, Gazzaley, & D’Esposito, 2004). The ma-
trices from the output of the individually-modulated regression were then submitted to a least
squares sum estimation using AFNI’s 3dLSS, in accordance with best practices for performing
multivariate analyses on fast event-related fMRI designs (Mumford, Turner, Ashby, & Poldrack,
2012; Mumford, Davis, & Poldrack, 2014).
The beta coefficient output from 3dLSS was then used in an MVPA performed in MATLAB
using the Decoding Toolbox (TDT; Hebart, Görgen, & Haynes, 2015). A linear support vector
machine (SVM) classification was used with 3 voxel (6 mm) radius searchlight and a leave-
one-run-out cross validation scheme, in which the SVM was trained on data from nine runs
and tested on a tenth, held-out run. The regularization parameter for the SVM was set at C = 1.
In a searchlight analysis, a roving sphere moves across the whole brain, and in each search-
light sphere an SVM was trained and tested (Kriegeskorte, Goebel, & Bandettini, 2006). IL
classification accuracy score was assigned to the central voxel of each searchlight. Questo
yielded a participant-level classification accuracy map for each participant in their brain’s
Neurobiology of Language
347
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
N
o
_
UN
_
0
0
0
1
6
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Neural decoding of articulable and inarticulable sounds
native space. The following classifications were performed: /i/ versus /y/ for vowel tokens (defined
as step 1 and step 3 vs. step 5 and step 7, hereafter “/i/Vowel vs. /y/Vowel
"), and the analogous
sine version (which we will hereafter refer to as “/i/Sine vs. /y/Sine
").
In preparation for group-level analyses, participant-level classification maps were then
normalized to Talairach space and blurred with a 4 mm Gaussian kernel. Following this,
single-sample t tests against chance levels were performed for the /i/Vowel vs. /y/Vowel and
/i/Sine vs. /y/Sine (using 3dttest++ in AFNI): t test output was masked by a group mask which
only included voxels present in all 26 participants. Inoltre, a paired-samples t test was
performed comparing the /i/Vowel vs. /y/Vowel and /i/Sine vs. /y/Sine decodings.
Cluster level correction for multiple comparisons was determined by running 10,000 iter-
ations of Monte Carlo simulations on the group mask. Specifically, we used the -acf option in
3dFWHMx and 3dClustSim (AFNI) to estimate the spatial smoothness and generate voxel-wise
and cluster-wise inferences. Data were corrected at a cluster level correction of p < 0.05 (voxel
level threshold of p < 0.025, 149 contiguous voxels).
RESULTS
Behavioral Data Analysis and Results
Vowel training results
All the following behavioral analyses were conducted using the R packages afex and lme4
(Bates, Mächler, Bolker, & Walker, 2015; Singmann, Bolker, Westfall, Aust, & Ben-Shachar,
2020). Model selection was achieved by first creating and running all possible permutations
of random effects structures, after which a backwards stepping selection procedure was applied
to the models that converged (Matuschek, Kliegl, Vasishth, Baayen, & Bates, 2017). Output
from the ANOVA table generated by the afex command “mixed” (run on the appropriate model
selected from the earlier described procedure) is reported for all mixed-effects model analyses in
this study. Residuals were inspected for each selected model with no deviations from normal
encountered.
Perceptual fading training. On average, accuracy during training improved over the course of
the three days (see Table 1, Figure 3A). Given the adaptive nature of the paradigm, participants
completed varying repetitions of each level of difficulty (easy: M = 1.29 blocks, SD = 0.58;
medium: M = 1.28 blocks, SD = 0.58; hard: M = 2.12 blocks, SD = 0.90), and as expected
there was a strong negative correlation between the number of blocks completed and average
accuracy on the explicit perceptual fading training (r(24) = −0.92, p < 0.0001), reflecting the
fact that less proficient learners had to complete more blocks to progress through training.
Accuracy data during training was submitted to a generalized linear mixed-effects model
using the R packages afex and lme4 (Bates, Mächler, Bolker, & Walker, 2015; Singmann,
Bolker, Westfall, Aust, & Ben-Shachar, 2020). Model selection was achieved by first creating
and running all possible permutations of random effects structures, after which a backwards
stepping selection procedure was applied to the models that converged (Matuschek, Kliegl,
Vasishth, Baayen, & Bates, 2017). Output from the ANOVA table generated by the afex com-
mand “mixed” (run on the appropriate model selected from the earlier described procedure) is
reported for all mixed-effects model analyses in this study.
To examine the degree to which participants’ accuracy during vowel training increased
over time, a generalized linear mixed-effects model was fit The selected model included fixed
Neurobiology of Language
348
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
Table 1. Accuracy during explicit perceptual fading training
Training
Day 1: Vowel
Easy (1–7)
M = 87.7% (SD = 33%)
Medium (2–6)
M = 89% (SD = 31%)
Hard (3–5)
M = 73.9% (SD = 43.9%)
Day 2: Vowel
M = 96.2% (SD = 19%)
M = 93.1% (SD = 25.3%)
M = 76.9% (SD = 42.2%)
Day 3: Vowel
M = 95.7% (SD = 20.3%)
M = 93.3% (SD = 25%)
M = 79.4% (SD = 40.5%)
Day 3: Sine
M = 93.7% (SD = 24.3%)
M = 94.9% (SD = 22.1%)
M = 83.9% (SD = 36.7%)
Note: Means (M) and standard deviations (SD) displayed.
effects for training difficulty level, training day, and the interaction of training day and level, as
well as by-subject random slopes and intercepts for the interaction between training difficulty
level and training day. The model revealed a significant main effect of training difficulty ( χ2 =
89.27, p < 0.0001) and a main effect of training day ( χ2 = 7.45, p = 0.02), with higher accu-
racy on easier blocks and higher accuracy on successive days of training The interaction be-
tween training difficulty and training day did not reach significance ( p = 0.62).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Behavioral data from the vowel training paradigm. (A) Accuracy on the explicit perceptual fading training task for the vowel stimuli
as a function of training level difficulty. Each point indicates an individual participant’s performance, and the shape of the point indicates the
–/y/vowel con-
number of times that participant repeated that block. (B) Responses to the continuum categorization task as a function of /i/vowel
0
tinuum step and training day. Error bars indicate standard error. (C) d
scores from the AX discrimination task for the vowel stimuli across each
0
stimulus as a function of pretest and posttest as well as training day. Error bars indicate standard error. (D) d
scores from the AX discrimination
task for vowel stimuli with respect to the participant-specific determination of the between-category (BC) and within-category ( WC) tokens as a
function of pretest and posttest as well as training day. Error bars indicate standard error.
Neurobiology of Language
349
Neural decoding of articulable and inarticulable sounds
In general, participants were successful at identifying the end-
Vowel categorization posttest.
points of the continuum, with at least 90% accuracy on average across each of the three train-
ing days (see Figure 3B). Responses from the categorization posttest were transformed into a
binary outcome variable and then submitted to a generalized linear mixed-effects model with
a logit link function (also instantiated in the R package afex) to assess how categorization of the
full continuum changed over the course of the three training days. P values were estimated
using a likelihood-ratio test. The model selected included fixed effects for continuum step
(centered) and training day, with by-subject random slopes and intercepts for continuum step,
training day, and their interaction. Output from the model indicated a main effect of contin-
uum step ( χ2 = 44.15, p < 0.0001) and a main effect of training day ( χ2 = 6.08, p = 0.05),
indicating that participants’ /y/Vowel responses increased as a result of the F3 manipulation
across the continuum as expected, and that participants’ overall proportion of /y/Vowel re-
sponses decreased with more experience with the continuum. The interaction was not signif-
icant ( p = 0.82), indicating no significant change in the steepness of the categorization
function over the three days of training.
AX discrimination task. Discrimination data was first separated into pretest (the first discrimina-
tion task completed during that day’s training) and posttest (the final discrimination task follow-
ing that day’s continuum categorization task), and “different” trials were collapsed such that
discrimination token “step 1–step 3” was considered the same as “step 3–step 1.” Following
was calculated for each of the resulting five “different” tokens for each subject and
this, d
training day. For the purposes of analysis, the token “3–5” was considered to be “between-
category” (as step 4 of the continuum represents the most ambiguous token in terms of
acoustics), and all other tokens were considered “within-category” (Figure 3C).
0
0
A linear mixed-effects model was then fit for participants’ d
scores, with fixed effects of
token type (between-category vs. within-category), training day, block (pretest vs. posttest)
and their interactions, as well as by-subject random intercepts. A main effect of token type
0
was found ( χ2 = 12.30, p < 0.001), in that d
scores were higher for the between-category
token (step 3 – step 5) than the within-category tokens. In addition, a main effect of training
day ( χ2 = 3.09, p = 0.05) arose, indicating that d
scores increased over the course of the
experiment. All other main effects and interactions were nonsignificant.
0
Using the acoustic boundary to determine the between-category discrimination token ig-
nores the fact that participants’ perceptual boundary did not necessarily correspond to the
midpoint of the continuum. To remedy this concern, we estimated participant-relative percep-
tual boundaries and fit a linear mixed-effects model using the participant-relative determina-
tions of the between-category and within-category tokens. The model selected contained fixed
effects of token type (between-category vs. within-category), training day, block (pretest vs.
posttest) and their interactions, as well as by-subject random intercepts. This model was then
compared to the model previously fit using the acoustic boundary to determine whether a
token was between-category or within-category. The model that used participant-relative
boundaries had superior fit but yielded largely the same findings: a main effect of token type
( χ2 = 38.76, p < 0.0001), a main effect of training day ( χ2 = 5.19, p < 0.01), and a main effect of
block ( χ2 = 6.78, p < 0.01). All interactions were nonsignificant.
Sine training results
Explicit perceptual fading training. Participants again completed varying repetitions of each level
of difficulty (easy: M = 1.27 blocks, SD = 0.60; medium: M = 1.35 blocks, SD = 0.56; hard:
Neurobiology of Language
350
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
M = 1.69 blocks, SD = 0.84), and there was a similarly strong negative correlation between
the number of blocks completed and the average accuracy on the explicit perceptual fading
training, r(7) = −0.86, p = 0.003.
To examine the efficacy of the single training session on identification of the two Sine
tokens, a generalized linear mixed-effects model was performed. The model selected included
fixed effects for training difficulty level and by-subject random intercepts (the maximal model
for this analysis). The model revealed a significant main effect of training difficulty ( χ2 =
102.98, p < 0.0001), indicating higher accuracy on easier blocks consisting of larger intervals
on the continuum.
Sine AX discrimination task. Discrimination data for the sine-wave stimuli was analyzed using
the same procedure as the vowel stimuli. A linear mixed-effects model was then used for par-
ticipants’ d
scores, with fixed effects of token type (between-category vs. within-category),
block (pretest vs. posttest), and their interaction, as well as by-subject random slopes and
intercepts for all fixed effects and interactions.
0
The model revealed that there was no significant difference in d
between the between-
category and within-category tokens ( p = 0.26). However, there was a main effect of block,
0
scores overall increased from pretest to posttest (F = 9.43, p < 0.01). The interac-
in which d
tion between token type and block was significant (F = 5.60, p < 0.05), indicating a larger
increase in sensitivity at posttest for the between-category token than the within-category
token. Recoding the data to use participant-relative determinations of the between-category
token did not alter the results (Figure 4D).
0
Comparison of vowel and sine-wave behavioral performance
Explicit perceptual fading training. Accuracy on the training task was comparable in sine-wave
and vowel tokens, despite participants only having a single day of training (see Table 1,
Figure 4A), which was expected from previous pilot testing. Data from the third day of training
was analyzed with a generalized linear mixed-effects model with logit link function (the de-
pendent variable, accuracy, was coded as binary value for each trial). Fixed effects of training
difficulty level, stimulus type ( Vowel vs. Sine), and their interaction, along with by-subject
random slopes and intercepts for all fixed effects and their interactions were entered into
the model. P values were estimated using a likelihood ratio test. The model output yielded
a significant main effect of block ( χ2 = 29.33, p < 0.0001), indicating that participants’ accu-
racy decreased as training difficulty increased, regardless of stimulus type, as well as a main
effect of stimulus type ( χ2 = 4.39, p = 0.04), in that accuracy was higher for the Sine stimuli
compared to the Vowel stimuli. The interaction between block and stimulus type was nonsig-
nificant ( p = 0.07).
Categorization posttest. Participants’ accuracy at identifying the endpoints of the Sine con-
tinuum was not as robust as with the Vowel continuum, but still reached at least 75% ac-
curacy at each endpoint token (Figure 4B). Posttest categorization accuracy was analyzed
using a generalized linear mixed-effects model where categorization of the Vowel (on the
third training day) and Sine continua were compared. Fixed effects of continuum step, con-
tinuum type (Sine vs. Vowel), and their interactions, as well as random by-subject slopes
and intercepts for all fixed effects and interactions were entered into the model. P values
were estimated using a likelihood ratio test. There was a significant effect of continuum step
( χ2 = 42.19, p < 0.0001), but the effect of continuum type was nonsignificant ( p = 0.34).
However, there was a significant continuum step by continuum type interaction ( χ2 = 6.17,
Neurobiology of Language
351
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
Figure 4. Behavioral data from the sine-wave training paradigm. (A) Accuracy on the explicit perceptual fading training task for the sine-wave
stimuli as a function of training level difficulty. Each point indicates an individual participant’s performance, and the shape of the point indicates
–/y/Sine continuum step. Error bars
the number of times that participant repeated that block. (B) Categorization data as a function of the /i/Sine
indicate standard error. (C) d
scores from the AX discrimination task for the sine-wave stimuli across each stimulus as a function of pretest
0
and posttest. Error bars indicate standard error. (D) d
scores from the AX discrimination task for sine-wave stimuli with respect to the participant-
relative determination of the between-category (BC) and within-category ( WC) tokens as a function of pretest and posttest. Error bars indicate
standard error.
0
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
p = 0.01). Post hoc pairwise comparisons were performed using the R package lsmeans to
investigate the source of the continuum step by continuum type interaction. There were
significant differences (after Bonferroni correction) between the categorization of sine-wave
and vowel stimuli at step 1 ( p = 0.025), step 5 ( p = 0.012), step 6 ( p = 0.005), and step 7
( p = 0.004), which reflects the lesser degree of certainty at the endpoints for the sine-wave
continuum.
Another metric of differences in the mode of categorization between the two stimulus sets
(i.e., categorical perception vs. more continuous perception) is the slope of the categorization
function. To this end, psychometric functions were fit to participants’ continuum categoriza-
tion responses from the third day of training for both the sine-wave stimuli and the vowel stim-
uli using the package quickpsy (Linares & López-Moliner, 2016) in R. The slope of the
psychometric function was used as a tool to quantify how strongly categorical an individual’s
perception is, similar to the “categorical index” approach in Liebenthal et al. (2005, 2010),
with the assumption that a steeper slope is among the behavioral hallmarks of more rigidly
categorical perception. A linear mixed-effects model was then fit to the slopes of the psycho-
metric functions, with a fixed effect of stimulus type ( Vowel vs. Sine-wave) and random by-
subject intercepts. There was no significant difference in slope values between stimulus sets
(F = 0.55, p = 0.47).
Neurobiology of Language
352
Neural decoding of articulable and inarticulable sounds
Finally, to achieve converging evidence regarding the mode of categorization between the
vowel and sine-wave stimuli, we also analyzed participants’ reaction times in the phonetic
categorization task, as categorical perception is usually accompanied by slower responses
for more ambiguous tokens in the center of a continuum than the clearer endpoint tokens
(Pisoni & Tash, 1974). We again used responses only from the third day of training. To account
for overall slower responses to the sine-wave stimuli compared to the vowel stimuli (sine-
wave: M = 638.83 ms, SD = 577.35 ms, vowel: M = 473.64 ms, SD = 464.99 ms), we con-
verted reaction times to z-scores. A linear mixed-effects model was fit to the z-scored reaction
times, with main effects of stimulus type ( Vowel vs. Sine-wave), continuum step (squared, to
better reflect the quadratic shape of participants’ responses), and their interaction. The model
selected included by-subject random slopes and intercepts for stimulus type and continuum
step. There was no significant main effect of stimulus type ( p = 0.93) or continuum step ( p =
0.16), and critically no interaction between stimulus type and continuum step ( p = 0.85), in-
dicating that the pattern of responses for each step of the continuum, regardless of stimulus set,
was not significantly different (see Figure 5).
AX discrimination task. To evaluate the behavioral equivalence across stimulus sets, the AX
discrimination data from the third day of training for the vowel stimuli was combined with
the data from the sine-wave stimuli and analyzed using a linear mixed-effects model on the
0
d
scores. The model selected included main effects of token type (between-category vs. within-
category), block (pretest vs. posttest), stimulus set ( Vowel vs. Sine), and their interactions, as
well as by-subject random slopes and intercepts for each main effect.
A main effect of token type was found (F = 4.53, p < 0.05), indicating higher overall sen-
sitivity for between-category tokens than within-category tokens, as well as a main effect of
0
block (F = 6.58, p < 0.05), indicating higher d
scores at posttest than pretest, and a main effect
of stimulus set (F = 21.68, p < 0.0001), with participants showing better overall sensitivity to
sine-wave stimuli. In addition, a significant interaction of token type and stimulus set was
found (F = 12.06, p < 0.001), as well as a significant interaction between block and stimulus
set (F = 9.72, p < 0.01).
Post hoc pairwise comparisons on both interactions were again performed using the R
package lsmeans with Bonferroni correction. The interaction between token type and stimulus
set was driven by a significant difference between within-category and between-category
Figure 5. Reaction time (z-scored) in the continuum categorization task on the third day of training
as a function of continuum step and stimulus set. Error bars indicate standard error.
Neurobiology of Language
353
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
Figure 6. Regions that showed greater activation for sine-wave tokens than vowel tokens in the
ANOVA. Clusters corrected at the voxel level of p < 0.025, with 149 contiguous voxels and a cor-
rected threshold of p < 0.05.
tokens in the vowel data ( p = 0.001), but not the sine-wave data ( p = 0.34), while the inter-
action between block and stimulus set reflected an overall increase in d
from pretest to
posttest in the sine-wave condition ( p = 0.0004) but no difference in the vowel condition
( p = 0.51).
0
fMRI results
Imaging data from Sine and Vowel tokens were pooled in two analyses; a univariate approach
to look for differences in activation magnitude, and a multivariate approach to investigate dif-
ferences in activation patterns.
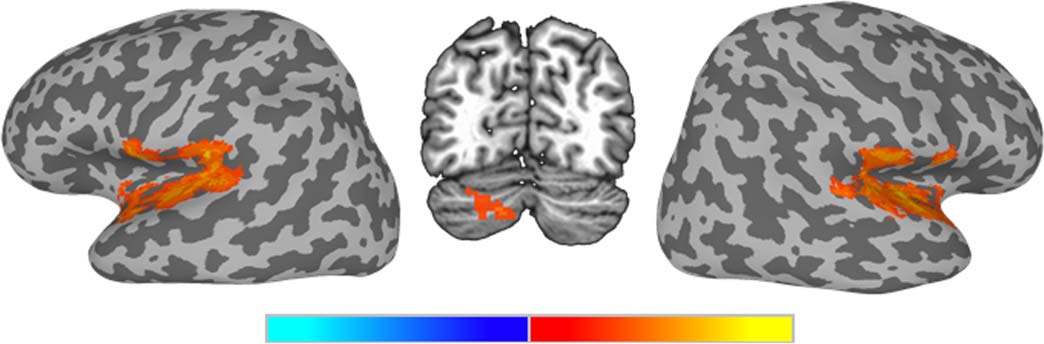
Univariate analysis. Contrasting functional activation for vowel tokens compared to sine-wave
tokens (Figure 6, Table 2) revealed three prominent clusters, all showing more activation for
sine-wave tokens compared to vowel tokens. Significant activation differences were found in a
large right hemisphere cluster in the STG extending into the Rolandic operculum, the insula,
the temporal pole, and Heschl’s gyrus; an analogous left hemisphere cluster in the STG ex-
tending into Heschl’s gyrus, the Rolandic operculum, and the temporal pole and a smaller
cluster in the left cerebellum.
To investigate how individual differences in proficiency with the two continua may have
impacted neural results, a linear mixed-effects model (with fixed effects of stimulus type and
continuum step, and by-subject random intercepts) was performed on the functional data
using AFNI’s 3dLME. Accuracy on the hardest level of difficulty for the explicit perceptual
fading training for both the vowel tokens and sine-wave tokens on the final day of training
was selected as a covariate. After controlling for participants’ proficiency with the two
Table 2. Results of univariate ANOVA comparing activation to Sine versus Vowel stimuli
Area
Sine > Vowel
L STG, L RO
R STG, R RO
L Cere (Crus 2), L Cere (Crus 1)
Cluster size ( Voxels)
1,807
1,777
150
X
−45
47
−15
sì
−11
−9
−69
z
Maximum t value
6
6
−34
7.85
9.55
3.90
Note. L = left; R = right; STG = superior temporal gyrus; RO = Rolandic operculum; Cere = cerebellum. Clusters corrected at the voxel level of p < 0.025, with
149 contiguous voxels and a corrected threshold of p < 0.05.
Neurobiology of Language
354
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
continua, similar bilateral STG clusters emerged in which sine-wave tokens elicited greater
activation than vowel tokens.
Multivariate analysis. Participant-level classification accuracy maps from the searchlight
MVPA analysis were submitted to single sample t tests against chance level (50%) to determine
cortical regions that were able to discriminate category differences (e.g., all “A” tokens vs. all
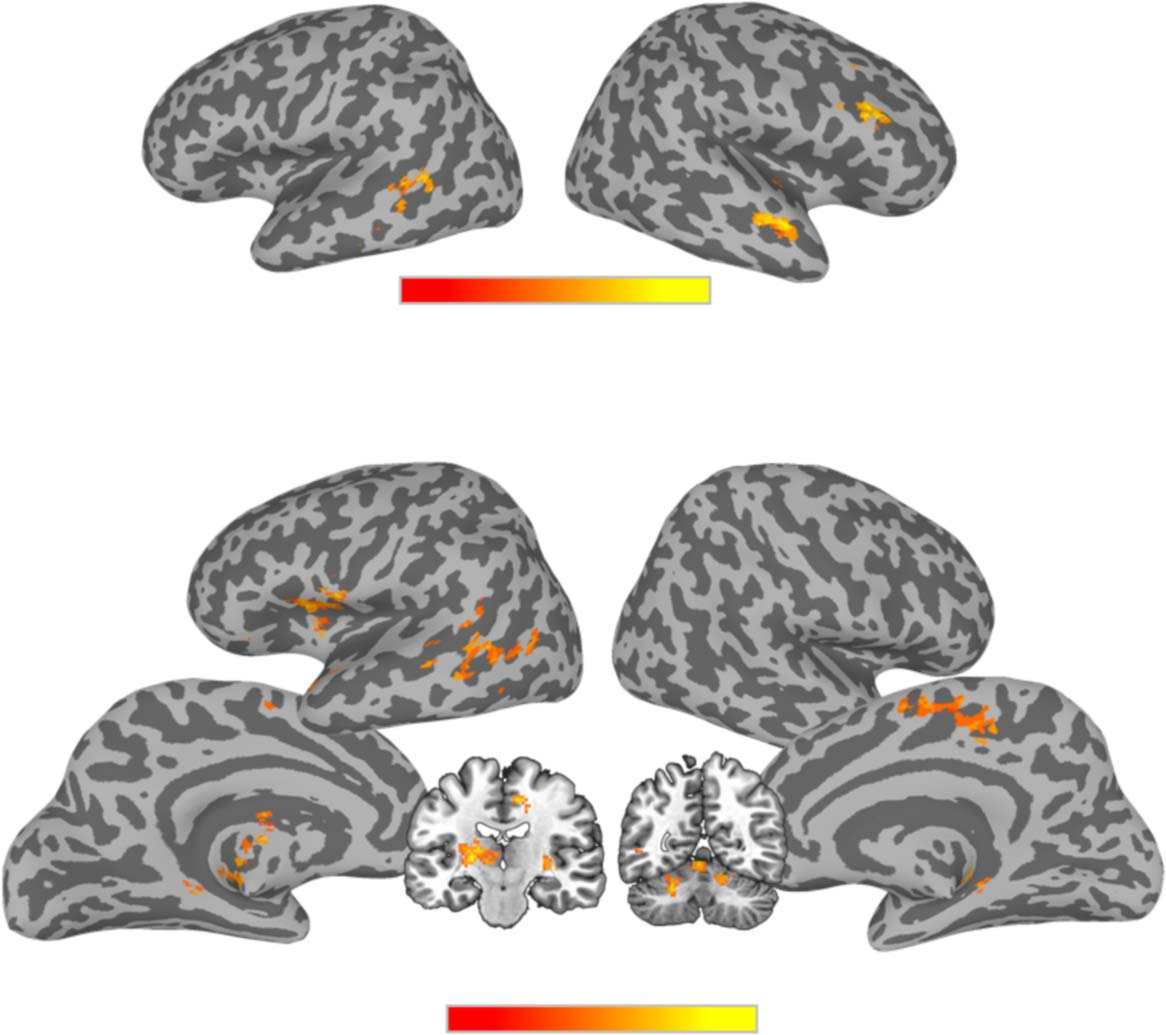
“B” tokens). For the decoding of the two vowel categories (/i/Vowel vs. /y/Vowel), three significant
clusters were found: (1) a cluster in the right middle temporal gyrus (MTG) extending into the
right STG, (2) a cluster in the right IFG (specifically in the pars triangularis and the pars oper-
cularis), and (3) a cluster in left MTG (see Table 3, Figure 7A). The same analysis for the sine-
wave speech tokens, decoding /i/Sine versus /y/Sine did not yield any regions that met the thresh-
old for significance, but voxel-level decoding results from the searchlight analysis contributed
to the Vowel decoding−Sine decoding comparison below.
Next, we contrasted the sine-wave and vowel decoding maps to identify regions that dif-
fered significantly in decoding performance for the two stimulus sets. Participant-level classi-
fication accuracy maps from the sine-wave and vowel decoding analyses were submitted to a
paired-samples t test to identify regions that showed significantly different decoding perfor-
mance for Vowel category differences than for Sine category differences. All such clusters
showed superior decoding of Vowel contrasts and included a cluster that extended from the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
Table 3. Results of t tests performed on MVPA classification data
Area
/i/vowel vs. /y/vowel decoding
Cluster size (voxels)
x
R MTG, R STG
R IFG
L MTG
/i/Sine vs. /y/Sine decoding
No significant clusters
Vowel decoding−Sine decoding
L Thalamus
L Insula, L TP, L RO, L IFG
L MTG
R SMA, R MCC, L SMA
R Cere ( VI), R Cere (IV-V)
L Cere (IV-V), Cere Verm (4/5)
R HC, R Put
L Cere ( VI), L Cere (Crus 1)
200
181
159
–
422
404
288
227
191
186
171
162
y
−7
19
−43
47
49
−53
–
–
−21
−33
−47
9
15
−3
27
−23
−17
−5
−59
−17
−53
−57
−29
−55
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
z
−12
30
2
–
6
20
6
44
−22
−8
0
−26
Peak accuracy
51.64%
51.66%
51.47%
–
Maximum t value
3.04
2.94
2.87
2.93
2.97
3.18
3.06
2.93
Note. L = left; R = right; MTG = middle temporal gyrus; STG = superior temporal gyrus; IFG = inferior frontal gyrus; TP = temporal pole; RO = Rolandic
operculum; SMA = supplementary motor area; MCC = middle cingulate cortex; Cere = cerebellum; HC = hippocampus; Put = putamen; Cere Verm = cerebral
vermis. Clusters corrected at the voxel level of p < 0.025, with 149 contiguous voxels yielding a corrected threshold of p < 0.05.
Neurobiology of Language
355
Neural decoding of articulable and inarticulable sounds
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Results from the multivoxel pattern analyses. (A) Regions that showed significant classi-
fication of the two Vowel categories. Clusters corrected at the voxel level of p < 0.025, with 149
contiguous voxels and a corrected threshold of p < 0.05. (B) Output from the paired t test showing
regions that classified Vowel categories significantly more accurately than Sine stimuli. The pop-out
image displays cerebellar clusters and a thalamus cluster not rendered in surface image.
left insula into the left temporal pole and the left IFG, as well as another cluster in the left MTG
(Table 3, Figure 7B).
DISCUSSION
Behavioral Results
The intent in this study was to train participants to equal proficiency on two novel contrasts, a
non-native vowel contrast and an acoustically matched sine-wave contrast. There are many
metrics that can be considered when evaluating the degree to which we were successful in
doing so; overall accuracy during training reflects the degree to which contrasts were learn-
able in the moment, the slope of the categorization curve reflects the precision with which
participants separated the categories, and differences in discrimination performance (in partic-
ular spikes in discriminability of tokens that cross the category boundary) reflect the emerging
“categorical” status of the sound category. It is notable that these metrics do not necessarily
pattern together, even in well-developed native language categories. For instance, several
studies have found no relationship between the categorization function and discrimination
performance (Gerrits & Schouten, 2004; Hary & Massaro, 1982; van Hessen & Schouten,
1999; Schouten, Gerrits, & van Hessen, 2003).
On balance, participants showed successful learning of both contrasts: Participants’ ability
to both identify and discriminate between the two categories in each continuum increased
Neurobiology of Language
356
Neural decoding of articulable and inarticulable sounds
with training, and ultimately, performance on the training task, the slope of the category
boundary, and the shape of the reaction time function were not significantly different between
the two continua. The continua differed, however, in the degree of “acquired distinctiveness”
in the discrimination data, with the vowel stimuli showing marked increases in sensitivity to
between-category tokens, while sine-wave stimuli showed much higher overall discriminabil-
ity, but no obvious peak for between-category tokens. This pattern may reflect qualitative dif-
ferences in the way that speech versus nonspeech categories emerge, with speech sound
categories more likely to have perceptual warping within the category, a classic hallmark of
native language speech categories. Below we consider our results in the context of other lit-
erature on speech and nonspeech category learning.
The vowel contrast in particular showed a behavioral pattern that qualitatively resembles
nativelike perception (i.e., an increased sensitivity across category boundaries, a decreased
sensitivity within category boundaries, a sharp categorization curve, and an increased reaction
time to near-boundary decisions (Liberman, Harris, Hoffman, & Griffith, 1957; Pisoni & Tash
0
1974). On the last day of training, d
values from the AX discrimination task for within-category
0
tokens, (e.g., 1–3 or 5–7) were much lower than on the first day of training, whereas d
scores
increased for the tokens most often containing participants’ perceptual boundary (2–4 and 3–5).
These results are congruent with previous studies of native English speakers’ perception of the
front-rounded vowel /y/, which shows that listeners will initially assimilate /y/ to the American
English vowel /i/. Nonetheless, this contrast is ultimately learnable to a high level of proficiency
(Levy & Strange, 2008; Levy, 2009).
Sine-wave versions of speech stimuli are not automatically mapped to speech analogues. In
our study, we removed F1 and F4 to prevent listeners from reconstructing the formants for the
/i/ and /y/ vowels and thus retrieving those speech categories. Studies of the perception of sine-
wave speech continua have found that participants’ responses in identification tasks were ei-
ther random or largely continuous (Grunke & Pisoni, 1982; Johnson & Ralston, 1994, exp. 1;
Desai et al., 2008), though a sine-wave speech version of a vowel continuum was perceived
categorically regardless of whether the listeners interpreted them as speech or nonspeech
(Johnson & Ralston, 1994, exp. 2). As discussed above, the sine-wave behavior is betrayed
by one finding that suggests that the sine-wave category was more poorly learned (or less fully
elaborated) than the vowel stimuli, namely a lack of an obvious peak in the discrimination
function around the category boundary. However, the significant interaction present in the
sine-wave AX discrimination analysis, wherein there was a greater increase in sensitivity for
the between-category token than the within-category tokens at posttest, does point to an
emerging pattern for acquired distinctiveness. Without additional exposure, this point will re-
main speculative as there is a lack of converging research on discrimination performance for
sine-wave speech after multiple days of training.
fMRI Results
In general, greater activation for the sine-wave tokens in the univariate analysis points to the
possibility that they were being processed less efficiently by a general auditory, and not a
speech-specific, mechanism. A broad scale contrast of areas where sine-wave tokens elicited
greater activation than vowel tokens revealed bilateral clusters in STG, similar to the findings
in Liebenthal et al. (2010), where nonspeech stimuli showed greater activation than speech
stimuli at posttest in STG. This could be due to the fact that the sine-wave tokens were still
more novel and participants had less of an opportunity to habituate to them. However, our
results replicate Liebenthal et al. (2010), where participants were trained over the course of
Neurobiology of Language
357
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
four sessions on their nonspeech stimuli, suggesting that mere lack of exposure is not the only
driving factor behind the sine-wave vs. vowel activation differences. Given the broad scale of
dimensions that vary between the two stimulus sets (e.g., articulatory information or acoustic
complexity), it is difficult to draw conclusions about the driving force of these neural differ-
ences using only univariate methods.
Regions identified in vowel decoding analysis are outside of dorsal stream
As discussed earlier, the dual stream model posits that the dorsal stream is recruited during the
learning of speech sounds, where it serves as an auditory-motor integration mechanism.
Evidence for this role has been difficult to ascertain given that many previous studies have used
stimuli where only the inarticulable stimuli were unfamiliar (e.g., Liebenthal et al. 2005, 2010).
To more conclusively answer this question, we trained participants on an unfamiliar vowel con-
trast, and used multivariate decoding analyses to investigate which brain regions could represent
the differences between these sounds. Any region that can decode these newly learned vowel
categories could do so by virtue of access to (1) articulatory differences (lip rounding differences
that listeners infer or extract from the auditory input), (2) abstract phonological categories (/i/ vs.
/y/), or (3) the acoustic differences between the two sounds (F3 is higher in /y/).
We found that the right MTG and the right STG were sensitive to vowel category information.
Crucially these areas are not included in the dorsal stream or implicated in speech-motor pro-
cessing but have been identified in previous imaging studies of non-native phonetic learning
(Callan et al., 2003; Wong, Perrachione, & Parrish, 2007). Additionally, another significant clus-
ter was found in the right IFG, which along with the right MTG, has been implicated as playing a
role in perceptual learning for speech, as Myers and Mesite (2014) found it to be sensitive to
changes in representation of an /s/-/∫/ contrast in response to a talker-specific artificial accent.
The right IFG has also been shown to be sensitive to nonlinguistic acoustic processing, like pitch
(Wang, Sereno, Jongman, & Hirsch, 2003), as well as to play a role in non-native phonetic learn-
ing (Myers & Swan, 2012). Thus, we take it as more likely that decoding sensitivity in these right
hemisphere regions is based upon differential representations of the abstract categories of /i/Vowel
and /y/Vowel and not the differences in articulation between them (or simply the acoustic differ-
ences). The only left hemisphere region that successfully decoded the two vowel categories was
the left MTG, and the cluster overlaps strongly with the phonological network described as part
of the ventral stream (Hickok & Poeppel, 2007). As such, our preferred interpretation of this pat-
tern is that the left MTG likely represents phonological category differences between /i/ and /y/.
The failure to find evidence for dorsal stream representation of different speech sounds during
phonetic learning, when it is predicted by the dual stream model, suggests that articulatory in-
formation is either coded in areas not involved in speech-motor gestures (e.g., the right MTG and
the STG) along with phonological representations, or represented at a level that our multivariate
analysis is not sensitive to, or else simply not recruited during this task.
Dorsal stream identified only when vowel decoding is contrasted against sine-wave decoding
It is possible the sensitivity to newly learned vowel categories reflects acquisition of any new
auditory categories, and is not speech-specific. In order to differentiate these possibilities, we
compared classification results from the /i/Vowel vs. /y/Vowel and the /i/Sine vs. /y/Sine decoding
maps. The resulting statistical map indicates regions that better decoded the two different
vowel categories than the two sine-wave categories, or put differently, regions that are sensitive
specifically to auditory contrasts that differ in articulation. The critical comparison of interest is
whether dorsal stream regions show discriminable patterns of activation only during the
Neurobiology of Language
358
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
learning of articulable sounds ( Vowel) but not nonarticulable sounds (Sine), as would be pre-
dicted by the conception of the dorsal stream as an auditory-motor integration mechanism.
Several dorsal stream regions emerged from this analysis; the left IFG and the left insula, the
bilateral supplementary motor areas, and a left MTG extending into posterior STG (see
Figure 7B) all showed superior decoding of vowel tokens compared to the sine-wave
analogues.
These results are consistent with several transcranial magnetic stimulation (TMS) studies of
the role of motor regions in speech perception, in which “artificial lesions” of speech motor
regions (i.e., primary and pre-motor cortices) lead to changes in discriminating or identifying
native language speech sounds, indicating a causal role of the articulatory regions during
speech perception (Rogers, Möttönen, Boyles, & Watkins, 2014; Möttönen & Watkins,
2009; Krieger-Redwood, Gaskell, Lindsay, & Jefferies, 2013; Sato, Tremblay, & Gracco,
2009; Meister et al. 2007; Smalle, Rogers, & Möttönen, 2015). However, these studies have
not necessarily addressed the dorsal stream as circuit but have instead focused on its constit-
uent regions one at a time. For instance, Krieger-Redwood et al. (2013) showed that stimula-
tion of dorsal stream regions affected phonetic decisions but not performance, on a more
naturalistic task where listeners had to match a word to a sample picture. Murakami, Kell,
Restle, Ugawa, and Ziemann (2015) applied TMS simultaneously to the left pre-motor cortex
and the left IFG (two of the three regions of the dorsal stream) and found decrements in pho-
nological processing only in noisy conditions, which suggests that the dorsal stream may only
be brought online during perception when stimulus identity is unclear. In our study, it is pos-
sible that the dorsal stream is recruited when the newly learned vowel /y/ must be distin-
guished from similar English vowels already in the repertoire. By contrast, the sine-wave
stimuli most likely do not assimilate to any existing auditory categories, and thus their identity
is more easily resolved.
The results from the Vowel decoding−Sine-wave decoding analysis do provide support for
the dual stream model’s conception of the dorsal stream as an auditory-motor integration
mechanism, but we hesitate to conclusively accept this interpretation. As we will discuss in
the next section, no brain regions significantly decoded sine-wave stimuli, which means that
the Vowel decoding−Sine-wave decoding analysis is displaying areas that have higher accuracy
in the vowel decoding, but nevertheless do not reach significance in the vowel decoding alone
(there was essentially no overlap between the map generated by the Vowel decoding−Sine-wave
decoding analysis and the map generated by the vowel decoding alone beyond the left MTG).
To that end, we believe the results from the vowel decoding, which do not involve any dorsal
stream regions, sufficiently address the original motivation of this study. Based upon these
results, future research should step back and more thoroughly examine if the dorsal stream is
truly involved in phonetic learning at all.
Another potential issue with the Vowel decoding−Sine-wave decoding lies in the present
study’s ability to completely control the equivalence of category structure across the vowel
and sine-wave stimuli. As our motivating goal was to investigate whether dorsal stream recruit-
ment in previous studies reflected auditory-motor integration or simply the formation of new
auditory categories, it was necessary to have two sets of stimuli that differed only on the
dimension of articulatory feasibility. As discussed above, although our stimuli were equated
in terms of participants’ success on the training task, that equivalent accuracy may have led to
qualitative differences in the shape of the emerging categories. Specifically, by one metric
(discrimination) our sine-wave stimuli differed from our vowel stimuli in their degree of
categoricity. Therefore, it is possible that the regions found in the Vowel decoding−Sine-wave
Neurobiology of Language
359
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
decoding could reflect differences in the emergence of categorical perception and not just
articulatory information. Future research will need to more strongly control for category struc-
ture to more precisely equivalate between speech and nonspeech sounds.
Gradient neural representations of sine-wave stimuli may have impeded decoding
The sine-wave decoding analysis did not uncover any brain regions that reliably represented
the two sine-wave categories. This is a bit of a puzzle given strong behavioral evidence that
participants were able to distinguish the categories. One explanation for the failure to find
neural sensitivity to sine-wave tokens appeals to the degree to which these stimuli showed
“acquired equivalence” within the sound category. In speech processing, acoustic cues to
speech sounds are initially encoded continuously at the neural level (Blumstein, Myers, &
Rissman, 2005; Frye et al., 2007; Toscano, McMurray, Dennhardt, & Luck, 2010), then cate-
gorical structure is imposed on them quickly and neural responses begin to reflect phonolog-
ical category membership early in processing (Toscano, Anderson, Fabiani, Gratton, &
Garnsey, 2018). We speculate that by contrast, nonspeech categories such as the sine-wave
speech tokens used in the current study may continue to be represented in a graded fashion at
the neural level. Because our analysis favored grouping of two more perceptually distinct sine-
wave stimuli to reflect a single category (e.g., steps 1 and 3 for /i/Sine), this could have created a
more difficult classification problem for sine-wave tokens. Submitting just the endpoints of the
sine-wave continuum (step 1 and step 7) to the same decoding analysis yielded a significant
cluster in the right MFG. Notably, this analysis suffers from less power than that reported in the
main body of the text, using only half of the tokens in the decoding analysis. This finding con-
trasts with a previous multivariate fMRI study of trained nonspeech novel auditory categories
that found that bilateral primary auditory cortex activation could successfully classify the two
categories (Ley et al., 2012).
On the topic of classifier accuracy, observed accuracy at the group levels was numerically
low, but statistically reliable. For the goals of this experiment, maximizing accuracy of the
classifier is not of primary importance; significantly above-chance findings indicate that there
are structured neural patterns in response to different stimuli, and using accuracy levels as a
measure of the size of this effect is a faulty assumption. For a detailed explanation of the dif-
ferences between MVPA for interpretation versus prediction, see Hebart and Baker (2018).
Subcortical regions implicated only in learning of speech sounds
Speech sound processing, especially during learning and adapting to speech sound variants,
may not be limited to cortical regions. In the present data, better decoding for the vowel con-
trast compared to the sine-wave contrast was found in the left thalamus. Previous work has
observed that the thalamus is sensitive to human speech sounds: Dehaene-Lambertz et al.
(2005) found that the thalamus was generally more active for speech rather than sine-wave
speech analogues in humans, while Kraus et al. (1994) found that the guinea pig thalamus
is sensitive to complex spectral differences between human speech sounds even when animals
were not exposed to any kind of training on these sounds. However, more relevant to this goal
of this study is the relationship between the thalamus and articulatory information; neuropsy-
chological investigations have found that damage to the thalamus often yields difficulties with
the articulation of speech sounds (Jonas, 1982; Wallesch et al., 1983), and our results suggest
that during the learning of new speech sounds, the thalamus may be representing the articu-
latory codes that will later be used for production, similar to the role of the dorsal stream in the
dual stream model.
Neurobiology of Language
360
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
More broadly, there has been increasing interest in the contribution of subcortical brain
structures in experience-dependent plasticity in the auditory system (Chandrasekaran, Skoe,
& Kraus, 2014), and it is suggested that the cerebellum is involved in adaptation to alterations
in speech through its functional connectivity to cortical language areas (Guediche, Holt,
Laurent, Lim, & Fiez, 2015). In the current study, the significant right cerebellar clusters
may reflect the engagement of this language plasticity network (similar to Guediche et al.,
2015) as a result of the nascent formation of a speech sound category. Language related cer-
ebellar function is heavily right lateralized, while the left cerebellum, especially lobule VI and
Crus 1, are implicated as having a role in executive functioning. Left cerebellar engagement in
this study could reflect a decision-making process about category membership for the vowel
contrast (for a review, see Stoodley & Schmahmann, 2009).
Conclusion
In sum, the MVPA results from the present experiment provide support for the interpretation
that dorsal stream regions are recruited during the learning of articulable sounds only, a ques-
tion that has been obscured in previous studies due to stimuli not being equated on profi-
ciency or articulatory feasibility. We did find evidence for dorsal stream recruitment when
looking for regions that could preferentially discriminate category membership for articulable
contrasts (our vowel contrast) rather than inarticulable contrasts (our sine-wave contrast);
however, differences in category structure between the articulable and inarticulable contrasts
(in that the former was perceived more categorically by some metrics than the latter) also
allow for the possibility that this recruitment reflects sensitivity to category structure and
not articulatory feasibility. However, these regions did not appear when examining decoding
of the two articulable vowel sounds alone. Instead, a series of regions outside of the dorsal
stream previously implicated in non-native phonetic learning could successfully classify the
two vowel categories. Future work will need to address the interplay between recruitment of
the dorsal stream in the initial stages of phonetic learning and the contribution of right hemi-
sphere cortical regions.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
FUNDING INFORMATION
Emily B. Myers, National Institute on Deafness and Other Communication Disorders (http://dx.
doi.org/10.13039/100000055), Award ID: R01DC013064. James Magnuson, National
Science Foundation (http://dx.doi.org/10.13039/100000001), Award ID: 1735225.
AUTHOR CONTRIBUTIONS
Emily Myers proposed the original motivation and design of the experiment. Emily Myers and
David Saltzman created the materials and final design. David Saltzman collected the data and
performed all analyses. David Saltzman wrote the initial draft of the manuscript, with additions
and revisions by Emily Myers. David Saltzman responded to reviewers’ comments with addi-
tions from Emily Myers.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Arsenault, J. S., & Buchsbaum, B. R. (2015). Distributed neural rep-
resentations of phonological features during speech perception.
Journal of Neuroscience, 35(2), 634–642. https://doi.org/
10.1523/JNEUROSCI.2454-14.2015
Arsenault, J. S., & Buchsbaum, B. R. (2016). No evidence of soma-
totopic place of articulation feature mapping in motor cortex
during passive speech perception. Psychonomic Bulletin &
Review, 23(4), 1231–1240. https://doi.org/10.3758/s13423-015-
0988-z
Bates, D, Mächler, M, Bolker, B, & Walker, S. (2015). Fitting linear
mixed-effects models using lme4. Journal of Statistical Software,
67(1), 1–48. https://doi.org/10.18637/jss.v067.i01
Neurobiology of Language
361
Neural decoding of articulable and inarticulable sounds
Best, C. T. (1995). A direct realist view of cross-language speech
perception. Speech Perception and Linguistic Experience,
171–206.
Blumstein, S. E., Myers, E. B., & Rissman, J. (2005). The perception
of voice onset time: An fMRI investigation of phonetic category
structure. Journal of Cognitive Neuroscience, 17(9), 1353–1366.
https://doi.org/10.1162/0898929054985473
Boersma, P., & Weenink, D. (2020). Praat: Doing phonetics by
computer [Computer program]. Version 6.1.16, retrieved 6
June 2020 from http://www.praat.org/
Callan, D. E., Tajima, K., Callan, A. M., Kubo, R., Masaki, S., &
Akahane-Yamada, R. (2003). Learning-induced neural plasticity
associated with improved identification performance after train-
ing of a difficult second-language phonetic contrast.
NeuroImage, 19(1), 113–124. https://doi.org/10.1016/S1053-
8119(03)00020-X
Chandrasekaran, B., Skoe, E., & Kraus, N. (2014). An integrative
model of subcortical auditory plasticity. Brain Topography, 27(4),
539–552. https://doi.org/10.1007/s10548-013-0323-9
Correia, J. M., Jansma, B. M. B., & Bonte, M. (2015). Decoding ar-
ticulatory features from fMRI responses in dorsal speech regions.
Journal of Neuroscience, 35(45), 15015–15025. https://doi.org/
10.1523/JNEUROSCI.0977-15.2015
Cox, R. W. (1996). AFNI: Software for analysis and visualization of
functional magnetic resonance neuroimages. Computers and
Biomedical Research, an International Journal, 29(3), 162–173.
https://doi.org/10.1006/cbmr.1996.0014
Darcy, I., Dekydtspotter, L., Sprouse, R. A., Glover, J., Kaden, C.,
McGuire, M., & Scott, J. H. (2012). Direct mapping of acoustics
to phonology: On the lexical encoding of front rounded vowels
in L1 English–L2 French acquisition. Second Language Research,
28(1), 5–40. https://doi.org/10.1177/0267658311423455
Dehaene-Lambertz, G., Pallier, C., Serniclaes, W., Sprenger-
Charolles, L., Jobert, A., & Dehaene, S. (2005). Neural correlates
of switching from auditory to speech perception. NeuroImage,
24(1), 21–33. https://doi.org/10.1016/j.neuroimage.2004.09.039
Desai, R., Liebenthal, E., Waldron, E., & Binder, J. R. (2008). Left
posterior temporal regions are sensitive to auditory categoriza-
tion. Journal of Cognitive Neuroscience, 20(7), 1174–1188.
https://doi.org/10.1162/jocn.2008.20081
Earle, F. S., & Myers, E. B. (2015). Sleep and native language inter-
ference affect non-native speech sound learning. Journal of
Experimental Psychology. Human Perception and Performance,
41(6), 1680–1695. https://doi.org/10.1037/xhp0000113
Fowler, C. A. (1986). An event approach to the study of speech per-
ception from a direct–realist perspective. Journal of Phonetics, 14(1),
3–28. https://doi.org/10.1016/S0095-4470(19)30607-2
Frye, R. E., Fisher, J. M., Coty, A., Zarella, M., Liederman, J., &
Halgren, E. (2007). Linear coding of voice onset time. Journal
of Cognitive Neuroscience, 19(9), 1476–1487. https://doi.org/
10.1162/jocn.2007.19.9.1476
Galantucci, B., Fowler, C. A., & Turvey, M. T. (2006). The motor
theory of speech perception reviewed. Psychonomic Bulletin &
Review, 13(3), 361–377. https://doi.org/10.3758/BF03193857
Gerrits, E., & Schouten, M. E. H. (2004). Categorical perception de-
pends on the discrimination task. Perception & Psychophysics,
66(3), 363–376. https://doi.org/10.3758/BF03194885
Grunke, M. E., & Pisoni, D. B. (1982). Some experiments on per-
ceptual learning of mirror-image acoustic patterns. Perception &
Psychophysics, 31(3), 210–218. https://doi.org/10.3758/
BF03202525
Guediche, S., Holt, L. L., Laurent, P., Lim, S.-J., & Fiez, J. A. (2015).
Evidence for cerebellar contributions to adaptive plasticity in
speech perception. Cerebral Cortex, 25(7), 1867–1877. https://
doi.org/10.1093/cercor/bht428
Hary, J. M., & Massaro, D. W. (1982). Categorical results do not
imply categorical perception. Perception & Psychophysics, 32(5),
409–418. https://doi.org/10.3758/BF03202770
Haxby, J. V., Gobbini, M. I., Furey, M. L., Ishai, A., Schouten, J. L., &
Pietrini, P. (2001). Distributed and overlapping representations of
faces and objects in ventral temporal cortex. Science, 293(5539),
2425–2430. https://doi.org/10.1126/science.1063736
Hebart, M. N., & Baker, C. I. (2018). Deconstructing multivariate
decoding for the study of brain function. NeuroImage, 180, 4–18.
https://doi.org/10.1016/j.neuroimage.2017.08.005
Hebart, M. N., Görgen, K., & Haynes, J.-D. (2015). The Decoding
Toolbox (TDT): A versatile software package for multivariate anal-
yses of functional imaging data. Frontiers in Neuroinformatics, 8.
https://doi.org/10.3389/fninf.2014.00088
Hickok, G., & Buchsbaum, B. (2003). Temporal lobe speech percep-
tion systems are part of the verbal working memory circuit:
Evidence from two recent fMRI studies. Behavioral and Brain
S c i e n c e s , 2 6 ( 6 ) , 7 4 0 – 7 4 1 . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 7 /
S0140525X03340166
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8(5), 393–402.
https://doi.org/10.1038/nrn2113
Hutchison, E. R., Blumstein, S. E., & Myers, E. B. (2008). An event-
related fMRI investigation of voice-onset time discrimination.
NeuroImage, 40(1), 342–352. https://doi.org/10.1016/j.
neuroimage.2007.10.064
Johnson, K., & Ralston, J. V. (1994). Automaticity in speech percep-
tion: Some speech/nonspeech comparisons. Phonetica, 51(4),
195–209. https://doi.org/10.1159/000261975
Jonas, S. (1982). The thalamus and aphasia, including transcortical
aphasia: A review. Journal of Communication Disorders, 15(1),
31–41. https://doi.org/10.1016/0021-9924(82)90042-9
Kraus, N., Mcgee, T., Carrell, T., King, C., Littman, T., & Nicol, T.
(1994). Discrimination of speech-like contrasts in the auditory
thalamus and cortex. The Journal of the Acoustical Society of
America, 96, 2758. https://doi.org/10.1121/1.411282
Krieger-Redwood, K., Gaskell, M. G., Lindsay, S., & Jefferies, E.
(2013). The selective role of premotor cortex in speech percep-
tion: A contribution to phoneme judgements but not speech
comprehension. Journal of Cognitive Neuroscience, 25(12),
2179–2188. https://doi.org/10.1162/jocn_a_00463
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006). Information-
based functional brain mapping. Proceedings of the National
Academy of Sciences, 103(10), 3863–3868. https://doi.org/
10.1073/pnas.0600244103
Lee, Y.-S., Turkeltaub, P., Granger, R., & Raizada, R. D. S. (2012).
Categorical speech processing in Broca’s area: An fMRI study
Journ al o f
using m ultiva riate pa ttern-based analysis.
Neuroscience, 32(11), 3942–3948. https://doi.org/10.1523/
JNEUROSCI.3814-11.2012
Levy, E. S. (2009). Language experience and consonantal context
effects on perceptual assimilation of French vowels by
American-English learners of French. The Journal of the
Acoustical Society of America, 125(2), 1138–1152. https://doi.
org/10.1121/1.3050256
Levy, E. S., & Strange, W. (2008). Perception of French vowels by
American English adults with and without French language expe-
rience. Journal of Phonetics, 36(1), 141–157. https://doi.org/
10.1016/J.WOCN.2007.03.001
Ley, A., Vroomen, J., Hausfeld, L., Valente, G., De Weerd, P., &
Formisano, E. (2012). Learning of new sound categories shapes
Neurobiology of Language
362
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
neural response patterns in human auditory cortex. Journal of
Neuroscience, 32(38), 13273–13280. https://doi.org/10.1523/
JNEUROSCI.0584-12.2012
Liberman, A. M., Cooper, F. S., Shankweiler, D. P., & Studdert-
K e n n e d y , M . ( 1 9 6 7 ) . P e r c e p t i o n o f t h e s p e e c h c o d e .
Psychological Review, 74(6), 431–461. https://doi.org/10.1037/
h0020279
Liberman, A. M., Harris, K. S., Hoffman, H. S., & Griffith, B. C.
(1957). The discrimination of speech sounds within and across
phoneme boundaries. Journal of Experimental Psychology, 54(5),
358–368. https://doi.org/10.1037/h0044417
Liebenthal, E., Binder, J. R., Spitzer, S. M., Possing, E. T., & Medler, D.
A. (2005). Neural substrates of phonemic perception. Cerebral
Cortex, 15(10), 1621–1631. https://doi.org/10.1093/cercor/
bhi040
Liebenthal, E., Desai, R., Ellingson, M. M., Ramachandran, B.,
Desai, A., & Binder, J. R. (2010). Specialization along the left su-
perior temporal sulcus for auditory categorization. Cerebral
Cortex, 20(12), 2958–2970. https://doi.org/10.1093/cercor/
bhq045
Linares, D., & López-Moliner, J. (2016). quickpsy: An R Package to
fit psychometric functions for multiple groups. http://diposit.ub.
edu/dspace/handle/2445/116040
Lotto, A. J., Hickok, G. S., & Holt, L. L. (2009). Reflections on mirror
neurons and speech perception. Trends in Cognitive Sciences, 13(3),
110–114. https://doi.org/10.1016/j.tics.2008.11.008
Mathôt, S., Schreij, D., & Theeuwes, J. (2012). OpenSesame: An
open-source, graphical experiment builder for the social sci-
ences. Behavior Research Methods, 44(2), 314–324. https://doi.
org/10.3758/s13428-011-0168-7
Matuschek, H., Kliegl, R., Vasishth, S., Baayen, H., & Bates, D.
(2017). Balancing type I error and power in linear mixed models.
Journal of Memory and Language, 94, 305–315. https://doi.org/
10.1016/j.jml.2017.01.001
Meister, I. G., Wilson, S. M., Deblieck, C., Wu, A. D., & Iacoboni,
M. (2007). The essential role of premotor cortex in speech per-
ception. Current Biology, 17(19), 1692–1696. https://doi.org/
10.1016/J.CUB.2007.08.064
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal gyrus.
Science, 343(6174), 1006–1010. https://doi.org/10.1126/
science.1245994
Möttönen, R., & Watkins, K. E. (2009). Motor representations of ar-
ticulators contribute to categorical perception of speech sounds.
Journal of Neuroscience, 29(31), 9819–9825. https://doi.org/
10.1523/JNEUROSCI.6018-08.2009
Mumford, J. A., Davis, T., & Poldrack, R. A. (2014). The impact of
study design on pattern estimation for single-trial multivariate
pattern analysis. NeuroImage, 103, 130–138. https://doi.org/
10.1016/j.neuroimage.2014.09.026
Mumford, J. A., Turner, B. O., Ashby, F. G., & Poldrack, R. A.
(2012). Deconvolving BOLD activation in event-related designs
for multivoxel pattern classification analyses. NeuroImage, 59(3),
2636–2643. https://doi.org/10.1016/j.neuroimage.2011.08.076
Murakami, T., Kell, C. A., Restle, J., Ugawa, Y., & Ziemann, X. U.
(2015). Left dorsal speech stream components and their con-
tribution to phonological processing. Journal of Neuroscience,
35(4), 1411–1422. https://doi.org/10.1523/ JNEUROSCI.0246-
14.2015
Myers, E. B. (2007). Dissociable effects of phonetic competition
and category typicality in a phonetic categorization task: An
fMRI investigation. Neuropsychologia, 45(7), 1463–1473.
https://doi.org/10.1016/j.neuropsychologia.2006.11.005
Myers, E. B., Blumstein, S. E., Walsh, E., & Eliassen, J. (2009).
Inferior frontal regions underlie the perception of phonetic cate-
gory invariance. Psychological Science, 20(7), 895–903. https://
doi.org/10.1111/j.1467-9280.2009.02380.x
Myers, E. B., & Mesite, L. M. (2014). Neural systems underlying
perceptual adjustment to non-standard speech tokens. Journal
of Memory and Language, 76, 80–93. https://doi.org/10.1016/j.
jml.2014.06.007
Myers, E. B., & Swan, K. (2012). Effects of category learning on neu-
ral sensitivity to non-native phonetic categories. Journal of
Cognitive Neuroscience, 24(8), 1695–1708. https://doi.org/
10.1162/jocn_a_00243
Pisoni, D. B., & Tash, J. (1974). Reaction times to comparisons
within and across phonetic categories. Percep tion &
Psychophysics, 15(2), 285–290. https://doi.org/10.3758/
BF03213946
Pulvermüller, F., Huss, M., Kherif, F., Martin, F. M. del P., Hauk,
O., & Shtyrov, Y. (2006). Motor cortex maps articulatory features
of speech sounds. Proceedings of the National Academy of
Sciences, 103(20), 7865–7870. https://doi.org/10.1073/
pnas.0509989103
Remez, R. E., Rubin, P. E., Pisoni, D. B., & Carrell, T. D. (1981).
Speech perception without traditional speech cues. Science,
212(4497), 947–949. https://doi.org/10.1126/science.7233191
Rissman, J., Gazzaley, A., & D’Esposito, M. (2004). Measuring
functional connectivity during distinct stages of a cognitive task.
NeuroImage, 23(2), 752–763. https://doi.org/10.1016/j.
neuroimage.2004.06.035
Rogers, J. C., Möttönen, R., Boyles, R., & Watkins, K. E. (2014).
Discrimination of speech and non-speech sounds following theta-
burst stimulation of the motor cortex. Frontiers in Psychology, 5.
https://doi.org/10.3389/fpsyg.2014.00754
Sato, M., Tremblay, P., & Gracco, V. L. (2009). A mediating role of
the premotor cortex in phoneme segmentation. Brain and
Language, 111(1), 1–7. https://doi.org/10.1016/j.bandl.
2009.03.002
Schomers, M. R., & Pulvermüller, F. (2016). Is the sensorimotor cor-
tex relevant for speech perception and understanding? An inte-
grative review. Frontiers in Human Neuroscience, 10, 435–435.
https://doi.org/10.3389/fnhum.2016.00435
Schouten, B., Gerrits, E., & van Hessen, A. (2003). The end of cat-
egorical perception as we know it. Speech Communication, 41(1),
71–80. https://doi.org/10.1016/S0167-6393(02)00094-8
Scott, S. K., McGettigan, C., & Eisner, F. (2009). A little more con-
versation, a little less action—Candidate roles for the motor cor-
tex in speech perception. Nature Reviews Neuroscience, 10(4),
295–302. https://doi.org/10.1038/nrn2603
Singmann, H., Bolker, B., Westfall, J., Aust, F., & Ben-Shachar, M. S.
(2020). afex: Analysis of factorial experiments. R package version
0.27-2. https://CRAN.R-project.org/package=afex
Smalle, E. H. M., Rogers, J., & Möttönen, R. (2015). Dissociating
contributions of the motor cortex to speech perception and re-
sponse bias by using transcranial magnetic stimulation.
Cerebral Cortex, 25(10), 3690–3698. https://doi.org/10.1093/cer-
cor/bhu218
Stoodley, C. J., & Schmahmann, J. D. (2009). Functional topogra-
phy in the human cerebellum: A meta-analysis of neuroimaging
studies. NeuroImage, 44(2), 489–501. https://doi.org/10.1016/j.
neuroimage.2008.08.039
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic atlas of
the human brain. Stuttgart: George Thieme.
Toscano, J. C., Anderson, N. D., Fabiani, M., Gratton, G., &
Garnsey, S. M. (2018). The time-course of cortical responses to
Neurobiology of Language
363
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural decoding of articulable and inarticulable sounds
speech revealed by fast optical imaging. Brain and Language,
184, 32–42. https://doi.org/10.1016/j.bandl.2018.06.006
Toscano, J. C., McMurray, B., Dennhardt, J., & Luck, S. J. (2010).
Continuous perception and graded categorization: Electrophys-
iological evidence for a linear relationship between the acoustic
signal and perceptual encoding of speech. Psychological Science,
21(10), 1532–1540. https://doi.org/10.1177/0956797610384142
Tsao, F.-M., Liu, H.-M., & Kuhl, P. K. (2004). Speech perception in
infancy predicts language development in the second year of life:
A longitudinal study. Child Development, 75(4), 1067–1084.
https://doi.org/10.1111/j.1467-8624.2004.00726.x
van Hessen, A. J., & Schouten, M. E. H. (1999). Categorical percep-
tion as a function of stimulus quality. Phonetica, 56(1–2), 56–72.
https://doi.org/10.1159/000028441
Wallesch, C.-W., Kornhuber, H. H., Brunner, R. J., Kunz, T.,
Hollerbach, B., & Suger, G. (1983). Lesions of the basal ganglia,
thalamus, and deep white matter: Differential effects on language
functions. Brain and Language, 20(2), 286–304. https://doi.org/
10.1016/0093-934X(83)90046-9
Wang, Y., Sereno, J. A., Jongman, A., & Hirsch, J. (2003). fMRI ev-
idence for cortical modification during learning of Mandarin lex-
ical tone. Journal of Cognitive Neuroscience, 15(7), 1019–1027.
https://doi.org/10.1162/089892903770007407
Wilson, S. M. (2009). Speech perception when the motor system is
compromised. Trends in Cognitive Sciences, 13(8), 329–331.
https://doi.org/10.1016/j.tics.2009.06.001
Wilson, S. M., & Iacoboni, M. (2006). Neural responses to non-
native phonemes varying in producibility: Evidence for the
sensorimotor nature of speech perception. NeuroImage, 33(1),
316–325. https://doi.org/10.1016/J.NEUROIMAGE.2006.05.032
Wilson, S. M., Saygin, A. P., Sereno, M. I., & Iacoboni, M. (2004).
Listening to speech activates motor areas involved in speech
production. Nature Neuroscience, 7, 701–702. https://doi.org/
10.1038/nn1263
Wong, P. C. M., Perrachione, T. K., & Parrish, T. B. (2007). Neural
characteristics of successful and less successful speech and word
learning in adults. Human Brain Mapping, 28(10), 995–1006.
https://doi.org/10.1002/hbm.20330
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
3
9
1
8
6
7
7
4
6
n
o
_
a
_
0
0
0
1
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
364