RESEARCH ARTICLE
Neural Mechanisms Underlying the Processing
of Complex Sentences: An fMRI Study
Margreet Vogelzang1,2*
, Christiane M. Thiel2,3, Stephanie Rosemann2,3
Jochem W. Rieger2,4, and Esther Ruigendijk1,2
1Institute of Dutch Studies, University of Oldenburg, Oldenburg, Germany
2Cluster of Excellence “Hearing4all,” University of Oldenburg, Oldenburg, Germany
3Biological Psychology, Department of Psychology, Department for Medicine and Health Sciences,
University of Oldenburg, Oldenburg, Germany
4Applied Neurocognitive Psychology, Department of Psychology, University of Oldenburg, Oldenburg, Germany
*Now at the Department of Theoretical and Applied Linguistics, University of Cambridge.
Keywords: complex sentences, syntactic processing, word order, working memory, cognitive load,
fMRI
ABSTRACT
Previous research has shown effects of syntactic complexity on sentence processing. In
linguistics, syntactic complexity (caused by different word orders) is traditionally explained by
distinct linguistic operations. This study investigates whether different complex word orders
indeed result in distinct patterns of neural activity, as would be expected when distinct linguistic
operations are applied. Twenty-two older adults performed an auditory sentence processing
paradigm in German with and without increased cognitive load. The results show that without
increased cognitive load, complex sentences show distinct activation patterns compared with
less complex, canonical sentences: complex object-initial sentences show increased activity in
the left inferior frontal and temporal regions, whereas complex adjunct-initial sentences show
increased activity in occipital and right superior frontal regions. Increased cognitive load seems
to affect the processing of different sentence structures differently, increasing neural activity for
canonical sentences, but leaving complex sentences relatively unaffected. We discuss these
results in the context of the idea that linguistic operations required for processing sentence
structures with higher levels of complexity involve distinct brain operations.
INTRODUCTION
It is well known in psycholinguistic research that some sentences are more difficult to process
than others (per esempio., Bader & Meng, 1999; Bahlmann, Rodriguez-Fornells, Rotte, & Münte, 2007;
Tun, Benichov, & Wingfield, 2010). Less is known, Tuttavia, about the possibly differential
effects on processing of sentences with different syntactic complexities. To investigate this,
we present an fMRI experiment that assesses how listeners process sentences with two differ-
ent types of syntactic complexity at two levels of cognitive load and assess the brain mecha-
nisms underlying their processing.
The syntactic complexity of sentences can be varied through variations in word order. For
German and English, standard (cioè., canonical) word order in a main clause is Subject (S)-
Verb (V)-Object (O; Vedere, per esempio., Zwart, 1997 for German). Tuttavia, speakers frequently use struc-
tures that deviate from canonical word order for, among other things, pragmatic reasons. As an
a n o p e n a c c e s s
j o u r n a l
Citation: Vogelzang, M., Thiel, C. M.,
Rosemann, S., Rieger, J. W., &
Ruigendijk, E. (2020). Neural
mechanisms underlying the
processing of complex sentences: An
fMRI study. Neurobiology of Language,
1(2), 226–248. https://doi.org/10.1162/
nol_a_00011
DOI:
https://doi.org/10.1162/nol_a_00011
Supporting Information:
https://doi.org/10.1162/nol_a_00011
Received: 06 Dicembre 2019
Accepted: 01 April 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Margreet Vogelzang
mv498@cam.ac.uk
Handling Editor:
Michal Ben-Shachar
Copyright: © 2020 Massachusetts
Institute of Technology. Pubblicato
under a Creative Commons Attribution
4.0 Internazionale (CC BY 4.0) licenza.
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
esempio, compare sentences (1UN) E (1B). Sentence (1UN) shows the canonical word order of
SVO. In sentence (1B), Tuttavia, the object is placed before the subject, creating a more com-
plex syntactic structure.
(1) UN. The father is washing the boy.
B. It is the boy that the father is washing.
(2) UN. Der Igel berührt den Hasen.
TheNOM hedgehog touches theACC hare.
B. Den Hasen berührt der Igel.
TheACC hare touches theNOM hedgehog.
German has relatively rich morphology, which enables relatively free word order. For exam-
ple, it allows for object-initial sentences, in which the arguments switch places compared to
subject-initial sentences while keeping the lexical material constant (compare the SVO
sentence in [2UN] and the OVS sentence in [2B]). Such sentences have the same meaning in
the sense of who did what to whom, but differ pragmatically, for instance (2B) could occur
in a story with several animals, in which the hedgehog is touching some but not other
animals, and one would like to stress the importance of the hare as the one being touched,
and not some other animal. (2UN) would then be the version used where the animals are doing
several things to each other (per esempio., pushing, kissing, touching), and one wants to describe the
touching event: the hedgehog is touching the hare. Importantly, both word orders are used
in German.
In linguistic theory, it is assumed that different word orders are derived through distinct lin-
guistic operations (per esempio., Chomsky, 1981). This linguistic theory makes no explicit claims about
processing or the neural mechanisms underlying processing. Tuttavia, if we assume that dif-
ferent word orders are derived through distinct linguistic operations, differences in the process-
ing of different word orders would also be expected. More specifically, it has been argued that
different linguistic operations would be reflected in distinct (neural) processing of sentences
with different word orders (Shetreet & Friedmann, 2014). We will refer to this notion as the
linguistic operations account from now on.
When comparing structurally complex sentences with noncanonical word order to sen-
tences with canonical word order, it is known that complex sentences are generally more dif-
ficult to process (per esempio., Bader & Meng, 1999; Bahlmann et al., 2007 [embedded clauses];
Carroll & Ruigendijk, 2013 [OVS sentences]; Friederici, Fiebach, Schlesewsky, Bornkessel,
& von Cramon, 2006; Tun et al., 2010 [object relative clauses]). Per esempio, object-initial
sentences in German have been found to elicit longer reading times (Hemforth, 1993) E
more interpretation errors (mistakes in identifying who did what to whom; Vedere, per esempio., Carroll,
Uslar, Brand, & Ruigendijk, 2016) compared to subject-initial sentences. Such effects of com-
plex sentences compared to canonical sentences have been studied extensively in the litera-
ture, but less attention has been given to investigating the processing of different types of
noncanonical, complex sentences, which are of particular interest when evaluating the dis-
tinct linguistic analyses asserted by the linguistic operations account.
Auditory processing of complex sentences has consistently been found to increase neural
activity compared to processing of less complex sentences in the literature, which consists
mainly of studies on English, German, and Hebrew. Several brain regions seem to be involved
in the processing of multiple types of complexity, or even of complex sentences in general,
such as Broca’s area in the inferior frontal gyrus (per esempio., Ben-Shachar, Hendler, Kahn,
Neurobiology of Language
227
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
Ben-Bashat, & Grodzinsky, 2003; Ben-Shachar, Palti, & Grodzinsky, 2004; Caplan, Alpert, &
Waters, 1999; Röder, Stock, Neville, Bien, & Rösler, 2002; Santi & Grodzinsky, 2010;
Shetreet, Friedmann, & Hadar, 2009; Stromswold, Caplan, Alpert, & Rauch, 1996; even inde-
pendent of modality, Bahlmann et al., 2007; Bornkessel, Zysset, Friederici, von Cramon, &
Schlesewsky, 2005; Constable et al., 2004; Friederici et al., 2006; Grewe et al., 2005; Just,
Carpenter, Keller, Eddy, & Thulborn, 1996) and posterior superior temporal regions, including
Wernicke’s area (Ben-Shachar et al., 2003, 2004; Constable et al., 2004; Just et al., 1996; Santi
& Grodzinsky, 2010), which are traditionally considered as auditory and language processing
areas. Notably, Friederici et al. (2006) found, in a reading experiment with increasing numbers
of argument permutations, that activity in the left inferior frontal gyrus (IFG) increases as a
function of syntactic complexity, and similarly Makuuchi, Grodzinsky, Amunts, Santi, E
Friederici (2013) found that, in reading, increasing movement distance (as a manipulation of
a linguistic operation) increases activity in the same region (although not linearly). In a recent
meta-analysis of 54 fMRI studies, Rodd, Vitello, Woollams, and Adank (2015) confirmed the
critical role of the posterior left IFG and posterior temporal regions in different types of
syntactic processing. Investigating different noncanonical word orders, specifically two types
of linguistic operations, verb movement and wh- (who, what, Dove, Quando, why) movement,
Shetreet and Friedmann (2014) found that in Hebrew, OSV compared to canonical SVO
sentences (wh- movement) increased activity in the left IFG and bilateral posterior temporal
regions, whereas VSO (verb movement) compared to canonical sentences increased activity in
the left inferior occipital gyrus. When comparing OSV sentences directly to VSO sentences,
they found increased activity in left inferior frontal and temporal regions and in medial superior
frontal regions. Shetreet and Friedmann (2014) conclude that processing different word orders
activates different brain areas, reflecting distinct underlying linguistic operations (wh- movement vs.
verb movement). Così, some regions’ involvement may be more specific for the type of complexity,
such as inferior occipital regions for processing sentences in which the verb precedes its arguments
(Shetreet & Friedmann, 2014), the middle frontal gyrus for processing object-initial sentences
(Röder et al., 2002), or the superior frontal gyrus for producing verb-second clauses (in Dutch,
Den Ouden, Hoogduin, Stowe, & Bastiaanse, 2008). Inoltre, there is a large body of re-
search on (temporarily) ambiguous sentences (per esempio., Bahlmann et al., 2007; Bornkessel et al.,
2005; Friederici, Mecklinger, Spencer, Steinhauer, & Donchin, 2001; Friederici, Steinhauer,
Mecklinger, & Meyer, 1998), but since we are interested in the processing of complexity rather
than garden paths or ambiguity resolution, these will not be reviewed here.
To sum up, previous research has shown effects of complexity, questo è, effects of processing
a sentence with noncanonical word order. Inoltre, we know that in linguistics different
word orders are explained by different linguistic operations (per esempio., Chomsky, 1981). It is still
largely unknown, Tuttavia, whether these different linguistic operations indeed result in dis-
tinct patterns of neural activity. In the current study, we acoustically presented sentences with
two types of linguistic complexity, that all contain the same syntactic relation of who did what
to whom, to compare their neural processing. We used several cognitive measurements as
well as a dual-task paradigm to examine the role of cognitive capacities and cognitive load
in the processing of complex sentences. Specifically, a visual change-detection task was used
to increase demands of visual attention and executive control (which we will refer to with the
general term of cognitive load) without taxing working memory. Importantly, this visual task
should not interfere with the auditory processing, but may interact with the cognitive control
required to process the different word orders. This would be in line with previous findings
linking cognitive control explicitly to syntax in comprehension (in children, Engel de Abreu,
Gathercole, & Martin, 2011) and to the syntactic processing of sentences in general (Novick,
Neurobiology of Language
228
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
Trueswell, & Thompson-Schill, 2005). If the processing of different word orders can indeed be
explained by distinct linguistic operations, then increased cognitive load may influence the
processing of sentences with different word orders in different ways. If, on the other hand,
the processing of different word orders relies purely on differences in processing load rather
than linguistic operations and analyses, then increased cognitive load would be expected to
influence the processing of sentences with different word orders in similar ways. More specif-
ically, memory-based accounts argue that processing cost is the result of the distance between
a moved element and its original position, regardless of the type of movement, in the sense that
the larger the distance, the more processing costs, which could be reflected in stronger acti-
vation patterns (per esempio., Grodner & Gibson, 2005; Lewis, Vasishth, & Van Dyke, 2006). Although
admittedly this is a somewhat simplified description of these accounts, which additionally take
into account other influences such as feature overlap, and few claims have been made about
neural mechanisms underlying sentence processing based on these accounts, a memory-based
explanation may be expected to result in qualitatively similar activation patterns for different
linguistic operations, that could differ in strength. Experience-based accounts (per esempio., Hale,
2001; Levy, 2008), in turn, assume that processing is affected by the frequency of occurrence
of a certain pattern. Crucially, this would, in our interpretation, also not result in qualitatively
different activation patterns for different word orders, but in differences in the amount of ac-
tivation (see Shetreet & Friedmann, 2014, for a further discussion of these predictions).
Here we investigated these predictions with older adults, who are expected to show more
difficulties with complex sentence processing (cf. per esempio., Emery, 1985; Kemper, 1992; Wingfield,
McCoy, Peelle, Tun, & Cox, 2006) and a wider spread in their cognitive capacities than youn-
ger adults; this would create the opportunity to better investigate the correlations between cog-
nitive measurements and linguistic performance, as well as the influence of cognitive load. Noi
measured working memory capacity, as it has been argued that more information has to be
kept active in working memory in object-initial sentences compared to subject-initial sen-
tences (Schlesewsky, Fanselow, Kliegl, & Krems, 2000). Additionally, measurements of vocab-
ulary and cognitive flexibility were taken, as these have been found to influence older adults’
performance on sentence processing in adverse listening conditions (McAuliffe, Gibson, Kerr,
Anderson, & LaShell, 2013; Rosemann et al., 2017).
Our experiment used four word order conditions—canonical sentences, object-initial sen-
tences, adjunct-initial sentences, and adjunct-initial sentences in which the object precedes
the subject (Vedi la tabella 1)—to test the main hypothesis that different noncanonical sentences
are processed in distinct ways, which would be in line with our predictions based on the
linguistic operations account. These sentence conditions represent the operations of wh-
movement (movement of an argument, creating a sentence in which the object precedes
the subject) and adjunct movement (fronting of the adjunct, creating a sentence in which
the verb precedes its arguments). Note that in German main clauses, the surface order is
verb-second for finite verbs. In subordinate clauses, the surface order is verb-final, and there
is discussion on which of these is the actual underlying order (per esempio., Broekhuis, 2006). There is,
Tuttavia, general agreement in the literature that there is a difference between moving an ad-
junct and moving an argument over the verb to the first position of the sentence (Vedere, per esempio.,
Cinque, 1990, or Rizzi, 1997). An important difference between the two types of movement
is that argument movement changes the theta-role hierarchy (as in, per esempio., Jackendoff, 1990) In
which the agent should precede patient, whereas adjunct movement affects the position of the
verb in relation to its arguments in German. Both of these movements should arguably affect
sentence processing. Regarding the specific sentence conditions, we expect both object-initial
sentences (in line with Carroll et al., 2016) and adjunct-initial sentences to be more difficult to
Neurobiology of Language
229
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
Tavolo 1. Example of an experimental item in the four different sentence conditions, each containing
a subject (S), verb ( V), adjunct (UN), and object (O)
Condition
SVAO
OVAS
AVSO
AVOS
Example sentence
Der Igel berührt am Montag den Hasen.
TheNOM hedgehog touches on Monday theACC hare.
Den Hasen berührt am Montag der Igel.
TheACC hare touches on Monday theNOM hedgehog.
Am Montag berührt der Igel den Hasen.
On Monday touches theNOM hedgehog theACC hare.
Am Montag berührt den Hasen der Igel.
On Monday touches theACC hare theNOM hedgehog.
process and interpret than canonical sentences, and that changing the order of the subject and
object results in different processing than changing the position of the adjunct (in line with
Shetreet & Friedmann, 2014). Based on the findings of Bahlmann et al. (2007), Bornkessel
et al. (2005), Röder et al. (2002), and Shetreet and Friedmann (2014), we expect increased
activity in the left IFG and temporal regions for object-initial sentences compared to canonical
sentences. Based on the findings of Shetreet and Friedmann (2014), we expect increased
activity in occipital regions for adjunct-initial sentences compared to canonical sentences
and increased activity in inferior frontal, superior frontal, and temporal regions for object-initial
sentences compared to adjunct-initial sentences. The fourth word order condition, with the
adjunct at the beginning and the object before the subject (AVOS), is a new addition to the
literature, for which we expect combined effects of object-initial and adjunct-initial sentences.
Note that these predictions stem from previous literature on the neural processing of these
types of complex sentences rather than the linguistic operations account or memory-based
accounts directly, as these do not provide any specific predictions about the localization of
neural processing of complex sentences. Although the dual task manipulation is more explor-
ative and therefore the hypotheses less precise, we expect increased cognitive load to make all
sentences more difficult to interpret correctly and to affect the processing of the different sen-
tences distinctly, which again would be in line with the linguistic operations account. Nostro
reasoning is that if underlying linguistic operations are not the same, they may not be affected
to the same degree by a general cognitive load as induced by the dual task. Based on previous
research on the influence of cognitive capacities, we expect working memory (Payne et al.,
2014; Vos, Gunter, Schriefers, & Friederici, 2001), vocabulary, and cognitive flexibility
(McAuliffe et al., 2013; Rosemann et al., 2017) to correlate with the interpretation and pro-
cessing of complex sentences.
MATERIALS AND METHODS
Participants
Twenty-four volunteers participated in the study. One participant had to be excluded due to
technical issues and one other participant was excluded due to a lack of responses during the
Neurobiology of Language
230
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
dual task. Twenty-two participants remained for analysis (15 females, aged 51–70, mean age
61; 0, SD 6; 3). The participants had normal hearing as tested with an individual pure tone
audiogram (PTA-4 ≤ 15 dB, PTA-high ≤ 25 dB) and normal or corrected-to-normal vision.
Participants performed the Montreal Cognitive Assessment task (MoCA; Nasreddine et al.,
2005), a concise screening tool for mild cognitive impairment, which rendered a mean group
score of 27.6 out of 30, indicating normal cognitive functioning. The participants were all na-
tive speakers of German and reported no language impairment and no psychiatric or neuro-
logical disorders. They were all right-handed. The ethics committee of the University of
Oldenburg, Germany, approved the study (approval number Drs. 28/2017) and written in-
formed consent was obtained from all participants. Participants received monetary compen-
sation for their participation.
Materials and Design
The experiment used acoustically presented German sentences, each followed by two pictures
for a picture-selection task. Each sentence consisted of a subject, a transitive verb, a temporal
adjunct, and an object. Four different sentence conditions were used: SVAO sentences, Quale
have canonical word order, OVAS sentences in which the object is placed sentence-initially,
adjunct-initial AVSO sentences in which the verb is placed before its arguments, and adjunct-
initial AVOS sentences in which the subject-object order is additionally manipulated. Vedere
Tavolo 1 for an example of each of the four conditions. The subject and object of a sentence
were always animate masculine nouns, to allow for unambiguous nominative and accusative
markers on the determiners. The temporal adjunct always consisted of “am” (SU) followed by a
two-syllable day of the week.
Sentences were created based on the OLACS corpus (Uslar et al., 2013): Adjectives were
removed from the original sentences and temporal adjuncts were added. An online question-
naire was performed with 165 participants as a pretest to check for the reversibility of the ar-
gument roles. Only sentences that were equally plausible with and without role reversal were
selected for the main experiment; sentence pairs for which the difference in plausibility score
exceeded two standard deviations from the mean were excluded. Forty items remained for the
main experiment.
All sentences were recorded by a native German-speaking woman at 44,100 Hz sampling
rate. The mean sentence length was 2,653 ms. A high-pass filter of 50 Hz was applied post-
recording. The sentences were adjusted to 36.3 RMS and calibrated at 100 dB. Using the
Oldenburg (Matrix) Sentence Test (Wagener, Brand, & Kollmeier, 1999UN, 1999B; Wagener,
Kühnel, & Kollmeier, 1999), the loudness of the stimuli was adjusted for each participant in-
dividually to 80% intelligibility during the MRI measurements to ensure that all participants
could hear the stimuli equally well. The average adjusted loudness of stimuli presentation
era 72 dB (SD = 7.0). These were played over MR compatible headphones (Opto Active,
Optoacoustics Ltd, Israel) after applying noise cancellation to eliminate part of the scanner
noise.
After each sentence, two pictures were displayed. These presented both characters men-
tioned in the sentence performing the mentioned action. Note that the experiment thus did
not test the interpretation of the adjunct. Piuttosto, it tested the influence of the adjunct manip-
ulation, as well as the subject-object order manipulation, on the sentence structure, and there-
fore on sentence processing, as a whole. Participants could indicate the picture that best fit the
sentence with a response box: the left button for the left picture and the right button for the right
picture (with the right index and right middle finger, rispettivamente). The location of the target
Neurobiology of Language
231
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
picture on the screen (left or right) was counterbalanced across trials. Visual stimulation was
accomplished by a projector (DATAPixx2, VPixx Technologies Inc.) and a screen that was po-
sitioned behind the MRI at a distance of 50 cm from eye to screen.
The experiment tested the effect of cognitive load on sentence processing by means of a
secondary task. As the secondary task, a fixation cross change detection task was used, Quale
was chosen because it is a visual task that taxes cognitive control mechanisms without inter-
fering with sensory processing in the primary task. In the dual task condition, either the hor-
izontal or vertical line of the fixation cross, which was always displayed on the screen during
sentence presentation, could become slightly larger. Importantly, the fixation cross change ap-
peared during sentence presentation. Participants had to pay attention to the fixation cross
change and press a button when they detected it; the cross then turned grey until the pictures
appeared on the screen. No action was required when no change was detected.
Finalmente, a baseline condition without sound, but with pictures, was added to check the data
recording and analysis. In this condition, participants were instructed to select one of the pic-
tures randomly. The four sentence conditions and one silent baseline, presented either as a
single task or in combination with a secondary task (dual task condition), constituted a 5 X 2
within-subjects design. The experiment consisted of 240 trials in total (24 critical trials per con-
dizione), distributed over two sessions with a short break in between. In each session, 120 trials
were presented, divided into six blocks of 20 trials each. Each block presented either the single
or the dual task; these two types of blocks were alternated. Two pseudo-randomized lists were
created to prevent potential order effects.
Each trial lasted 8 S. Within a single trial, the critical sentence (or silent baseline) was pre-
sented acoustically accompanied by a fixation cross on the screen, followed by a jitter of 300
A 700 ms, before two pictures appeared on the screen for the response phase. The response
phase lasted 3,500 ms. No feedback was given throughout the practice trials and the main
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Overview of the presentation of the stimuli within one trial (pictures adjusted from
Wendt, Brand, & Kollmeier, 2014; Wendt, Kollmeier, & Brand, 2015).
Neurobiology of Language
232
Processing complex sentences: An fMRI study
experiment. After the response phase, a fixation cross appeared again with a jitter of 300 A
700 ms, before the next trial started automatically. A schematic overview of the presentation of
a trial is depicted in Figure 1. Instructions on whether the single or dual task should be per-
formed were displayed for 10 s prior to a new block of trials.
Cognitive Tests
Digit span
A standard backwards digit span task was applied as a measurement of working memory
( WM) capacity. Participants heard a sequence of digits (1–9) and were asked to repeat this
sequence in reverse order. Increasingly longer sequences were tested, each sequence length
twice, starting at length two and ending when the participant was not able to correctly repeat
any instances of a sequence length. For each correctly reported sequence, participants re-
ceived one point. The longest possible sequence was eight digits; Perciò, the maximal num-
ber of points was 14. The mean score of our participants was 7.0 (SD = 2.4).
Vocabulary
A German vocabulary test called “Wortschatztest” (Schmidt & Metzler, 1992) was used as a
measure of verbal intelligence. Participants were presented with rows of six words in which
five nonwords and one existing word were included and were asked to detect the existing
German word without guessing. Increasingly difficult words were tested. For each correctly
identified word, participants received one point. The task consists of 42 rows of words;
Perciò, the maximal number of points was 42. The mean score of our participants was
33.6 (SD = 2.7).
CTMT
The Comprehensive Trail-Making Test (CTMT; Reynolds, 2002) was used as a measure of cog-
nitive flexibility and task switching abilities. It uses a visual search task in which participants
have to connect encircled numbers and letters in ascending order on paper. Two CTMT sub-
tests were conducted: trail 1 with numbers from 1 A 25 and trail 5 with numbers and letters
that have to be connected in alternation (cioè., 1-A-2-B-3-C…). The time from start until com-
pletion of a trail was measured. Any errors made by the participant were corrected immedi-
ately. Participants’ final score on the CTMT was calculated as the difference in completion
time between trail 1 and trail 5; a lower score thus reflects better cognitive flexibility. IL
mean score of our participants was 26.4 S (SD = 17.8).
Procedure
Participants were tested individually in a quiet room at the University of Oldenburg. Primo, au-
diogram measurements were taken in a soundproof booth. Then, the CTMT, MoCA, and digit
span task were conducted, followed by a first practice for the main experiment, before starting
the part of the study that took place in the MRI scanner.
The tests in the scanner were split into two parts, with a break in between. The Oldenburg
Sentence Test, a second practice for the main experiment, and the first session of the main
experiment were performed in the first part. After this first part, participants would have a break
outside the scanner, in which they completed the vocabulary task. After a 10- to 15-min break,
the second session of the main experiment, a structural scan (T1, longitudinal or spin-lattice
relaxation time), resting state measurements, and DTI measurements were done in the scanner.
Neurobiology of Language
233
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
The resting state and DTI data will not be reported in this article. Each session of the main
experiment took about 18 min. The complete procedure took around 3 hr.
Behavioral Data Analysis
All responses to the picture-selection task were coded as either correct or incorrect.
Subsequently, the rates of correct responses were calculated for each of the four sentence con-
ditions in both the single and the dual task condition. We examined the differences between
the sentence conditions, the effect of the dual task, and the relationship between responses on
the linguistic task and scores on the cognitive tests (digit span, vocabulary, and CTMT). Post
hoc Bonferroni-corrected pairwise comparisons identified which sentence conditions differed
from each other.
The responses were analyzed using binomial generalized linear mixed-effect-models in R.
Sentence condition was coded in terms of subject-object order (two levels) and adjunct posi-
zione (two levels) using effect coding (cioè., −0.5, 0.5). We examined the effects of subject-object
order, adjunct position, task condition (cioè., single vs. dual task) and any interactions (fixed
effects) on the rate of correct responses (dependent variable). Note that for the offline compre-
hension, we only expected an effect of subject-object order and not necessarily of adjunct
position, since the task only directly addressed this aspect. To put it differently, for finding
the correct picture, the position of the adjunct was irrelevant. Adjunct position was hence only
included here for the sake of completeness. A maximal (converging) random effects structure
was used, with random intercepts for participants and items and random slopes for subject-
object order for subjects and for subject-object order and adjunct position for items. Additional
factors such as list, session, and trial order were tested for warranted inclusion in the model;
only session improved the model and was therefore included. The influence of digit span,
vocabulary, and CTMT was assessed subsequently by testing their warranted inclusion in
the model as well as possible interactions with subject-object order and adjunct position; digit
span, vocabulary, and CTMT were all included in the model as well as an interaction between
subject-object order and digit span.
MRI Data Acquisition
The functional and anatomical MRI measurements were conducted with a 3T Siemens
Magnetom Prisma MRI scanner located at the University of Oldenburg. A 20-channel head
coil was used. Echo-planar imaging was used to measure a sequence with blood oxygen level
dependent, or BOLD, contrasto (repetition time [TR] = 1,800 ms, echo time [TE] = 30 ms, flip
angle = 75 degrees, df = 20, slice thickness = 3 mm, field of view = 192 cm, 33 slices). For the
first part of the main experiment, 600 whole-brain volumes were acquired and for the second
part (which included four extra warm-up trials) this number was 617. Additionally, T1-weighted
anatomical images were recorded (TR = 2,000 ms, TE = 2.07 ms, flip angle = 9°, slice thickness =
0.75 mm, 224 slices).
fMRI Data Analysis
Image processing and analysis was performed using SPM12 (Statistical Parametric Mapping,
Wellcome Department of Imaging Neuroscience, University College London, http://www.fil.
ion.ucl.ac.uk/spm). Preprocessing for each session of the imaging data included motion cor-
rection and realignment to the first image for the functional data, coregistration, segmentation,
and normalization to the Montreal Neurological Institute, or MNI, space for the functional and
anatomical data, and smoothing with a Gaussian filter (8 mm kernel) for the functional data.
Neurobiology of Language
234
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
Figura 2. Percentages of correct responses in the linguistic task as a function of sentence type and
load. Error bars are 95% CI derived from logistic analysis. S = subject, V = verb, A = adjunct, O = object.
For the analysis of the fMRI data, we focused on the time window from the onset of the
sentence until the onset of the pictures. Note that this time window is 3,500 ms + jitter, so
the time window varied between 3,800 ms and 4,200 ms for every trial. At the request of
one of the reviewers, the data were additionally analyzed using a shorter duration (including
only the audio, without the jitter or pause following the audio). The results of this additional
analysis were qualitatively similar to the analysis presented in this article, so the results from
the longer time will be reported in the Results section. One case in which the results from the
different analyses differed, namely, activation in the left IFG for AVSO sentences, will be dis-
cussed in the Discussion section. First-level analyses were done per participant using a general
linear model. We applied a high-pass filter of 128 s and accounted for serial correlations with
an Auto-Regressive model of order 1—AR(1). Head movement parameters were added as re-
gressors for each session separately. No participants were excluded due to excessive head
movements (>3 mm). The models were used to estimate the regression coefficients for the sen-
tences, with which the contrasts 1) OVAS > SVAO (object-initial compared to canonical word
order), 2) AVSO > SVAO (adjunct-initial compared to canonical word order), 3) AVOS >
SVAO (adjunct-initial, object-before-subject compared to canonical word order), 4) OVAS >
AVSO (object-initial compared to adjunct-initial word order), 5) AVOS > OVAS (adjunct-
initial, object-before-subject compared to object-initial word order), E 6) AVOS >
AVSO (adjunct-initial, object-before-subject compared to adjunct-initial word order) were
calculated for both the single and the dual task. For the group (second-level) analyses, these
estimates were taken to calculate the effects of syntactic complexity in the single task and in
the dual task with simple t tests. Paired t tests were used to examine the interaction between
Tavolo 2. Pairwise comparisons between the responses to the different sentence conditions in the
behavioral task
OVAS − SVAO = 0
AVSO − SVAO = 0
AVOS − SVAO = 0
AVSO − OVAS = 0
AVOS − OVAS = 0
AVOS − AVSO = 0
β
−1.16
−0.13
−1.54
1.03
−0.38
−1.41
z-value
−9.41
−0.95
−12.70
8.56
−3.89
−11.92
Note. S = subject, V = verb, A = adjunct, O = object.
p value
< 0.001
0.77
< 0.001
< 0.001
< 0.001
< 0.001
235
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
n
o
_
a
_
0
0
0
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Processing complex sentences: An fMRI study
syntactic complexity and cognitive load. Effects are reported as significant when exceeding a
cluster-level (Family-Wise Error) corrected threshold of p < 0.05 (with a p < 0.001 cluster-forming
threshold). To localize the brain regions and Brodmann areas (BA) we used SPM12, xjView
(http://www.alivelearn.net/xjview), and BioImage Suite’s MNI2TAL converter (http://
sprout022.sprout.yale.edu/mni2tal/mni2tal.html). Subsequently, conjunction analyses were
performed to investigate which regions showed common activation in different contrasts
by creating a flexible factorial design with each of the three main contrasts of complex con-
dition > SVAO. Finalmente, the influence of different cognitive capacities was investigated with
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
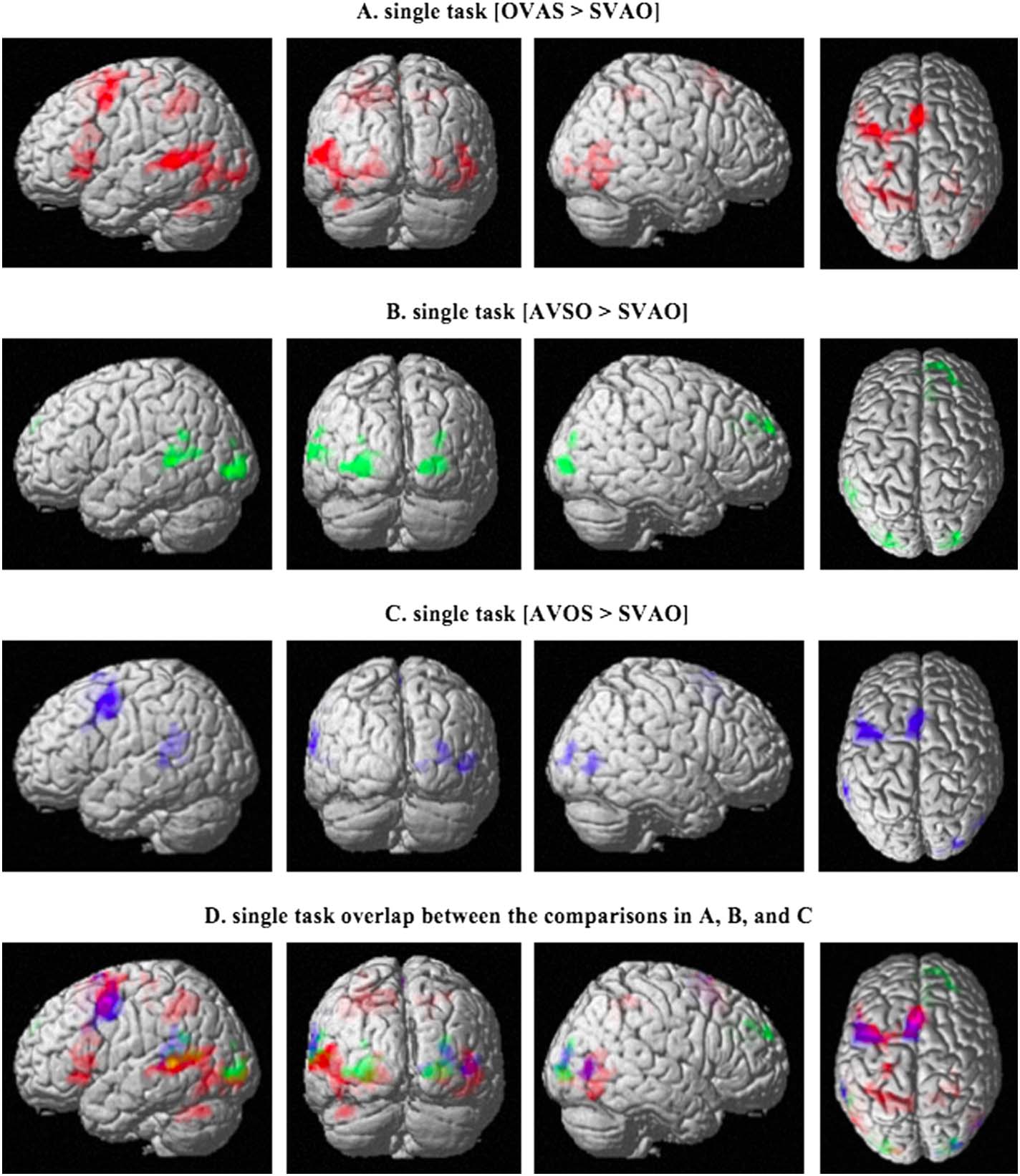
Figura 3. Activation patterns for the comparisons between sentence types in the single task. (UN)
shows the comparison between object-initial word order and canonical word order (OVAS >
SVAO), (B) shows the comparison between adjunct-initial word order and canonical word order
(AVSO > SVAO), E (C) shows the comparison between AVOS and the canonical SVAO.
Activated areas include the left auditory cortex and Broca’s area in (UN), left auditory cortex and
superior frontal regions in (B), and left auditory cortex and middle frontal regions in (C), P <
0.05; family-wise error corrected on the cluster level. The overlap between these comparisons is
shown in (D). S = subject, V = verb, A = adjunct, O = object.
Neurobiology of Language
236
Processing complex sentences: An fMRI study
multiple regression analyses, specifying the three factors of interest (digit span, vocabulary,
CTMT) as covariates.
RESULTS
Behavioral Results
The behavioral data are shown in Figure 2. The results showed main effects of subject-object
order (β = −3.34; z = −6.66; p < 0.001) and adjunct position (β = −0.42; z = −3.69; p < 0.001),
indicating that subject-before-object sentences were responded to correctly more often than
object-before-subject sentences and that adjunct-third sentences were responded to correctly
more often than adjunct-initial sentences. Post hoc pairwise comparisons showed that all con-
ditions significantly differed from each other except for the SVAO and AVSO conditions (see
Table 2). No effect of task type (single or dual task) was found (β = −0.06; z = −0.66; p = 0.51).
No significant interactions were found (all p’s > 0.05).
The model furthermore showed main effects of vocabulary and CTMT (rispettivamente, β =
0.18; z = 3.96; P < 0.001 and β = −0.02; z = −3.05; p < 0.01), indicating that participants
who performed better on the vocabulary task and CTMT task also performed better on the
linguistic task. Finally, an interaction between digit span score and subject-object order (β =
0.24; z = 3.57; p < 0.001) indicates that participants with a better digit span score also per-
formed better on the object-before-subject conditions in the linguistic task. Please see
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
n
o
_
a
_
0
0
0
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 3. MNI coordinates for the different comparisons under investigation in the single task
Comparison
single [OVAS > SVAO]
Peak coordinates
(X, sì, z)
(−40, −68, −28)
Z-value
5.09
Cluster size
2,765
Brain region
Left cerebellum exterior
single [AVSO > SVAO]
single [AVOS > SVAO]
(−48, 24, −4)
(−32, −50, 50)
(52, −70, −8)
(30, −42, 54)
(−26, −30, 64)
(−24, −92, 2)
(−54, −42, 4)
(18, −94, −2)
(12, 30, 28)
(8, 60, 30)
(−46, 4, 54)
(−6, 14, 56)
(−52, −38, 4)
(32, −92, 14)
4.64
4.59
4.10
4.09
3.99
5.62
4.28
4.25
4.10
3.93
5.20
4.62
4.13
3.94
2,311
1,445
843
549
253
569
471
469
241
212
817
991
391
244
Left frontal operculum
Left superior parietal lobule
Right inferior occipital gyrus
Right superior parietal lobule
Left postcentral gyrus
Left inferior occipital gyrus
Left middle temporal gyrus
Right inferior occipital gyrus
Right superior frontal gyrus (medial)
Right superior frontal gyrus
Left middle frontal gyrus
Left supplementary motor cortex
Left superior temporal gyrus
Right superior occipital gyrus
Note. S = subject, V = verb, A = adjunct, O = object.
Neurobiology of Language
237
Processing complex sentences: An fMRI study
Vogelzang, Thiel, Rosemann, Rieger, and Ruigendijk (2019) for a more detailed description
and plots of the effects of cognitive capacities on the offline sentence comprehension.
Imaging Results
Single task
A first analysis investigated the effects of sentence complexity in the single task. The first com-
parison, testing the effect of object- vs. subject-initial word order (OVAS > SVAO; Figure 3A,
Tavolo 3), showed activity in left cerebellar and temporal areas (BA 22), left inferior frontal areas
(including Broca’s area, BA 45 and BA 47), and superior parietal areas. Additional activity was
found in the inferior occipital gyrus and the left postcentral gyrus. The second comparison,
testing the effect of adjunct-initialization (AVSO > SVAO; Figure 3B, Tavolo 3), showed activity
in occipital areas and the left middle temporal gyrus (MTG; BA 21/22). Additional activity was
found in right superior frontal areas. The third comparison, testing the effect of adjunct-initial
word order with the object before the subject vs. canonical word order (AVOS > SVAO;
Figure 3C, Tavolo 3), showed activity in the left middle frontal gyrus (MFG; BA6) and left su-
perior temporal gyrus (STG), with additional activity in the left supplementary motor cortex
and right superior occipital gyrus. The overlap between these three comparisons is shown
in Figure 3D. Finalmente, we compared the noncanonical, complex conditions to each other
(OVAS > AVSO, AVOS > OVAS, and AVOS > AVSO). Although the comparisons of these
complex sentences to the canonical SVAO word order showed different activation patterns,
the differences in activity between the complex sentences did not reach significance. A
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 4. Average beta values extracted from the inferior frontal gyrus (IFG), middle temporal gy-
rus (MTG), superior temporal gyrus (STG), and inferior occipital gyrus (IOG) in all four sentence
conditions in the single task. Beta values were extracted from the entire mentioned regions using
WFU PickAtlas (Maldjian, Laurienti, & Burdette, 2004; Maldjian, Laurienti, Kraft, Burdette, & Kraft,
2003). Error bars indicate standard error of the mean. S = subject, V = verb, A = adjunct, O = object.
Neurobiology of Language
238
Processing complex sentences: An fMRI study
further examine the activity in each sentence condition, we extracted the beta values for these
conditions in four key regions for which hypotheses were formulated (the left IFG, left middle
and superior temporal gyri, and left inferior occipital gyrus) for visual inspection. These plots,
shown in Figure 4, confirm the overall differences in activity in these regions between complex
sentence conditions on the one hand and the canonical SVAO condition on the other. The key
regions show little differences in activity between the noncanonical sentences, which is in line
with the whole-brain–level analyses, in which differences in activity between the complex
sentences did not reach significance. One explanation for this may lie in the large variation
between participants, which is exemplified by their individual activation patterns in the con-
trast of complex sentences (all three types) vs. canonical SVAO sentences in the Supporting
Information file 1.
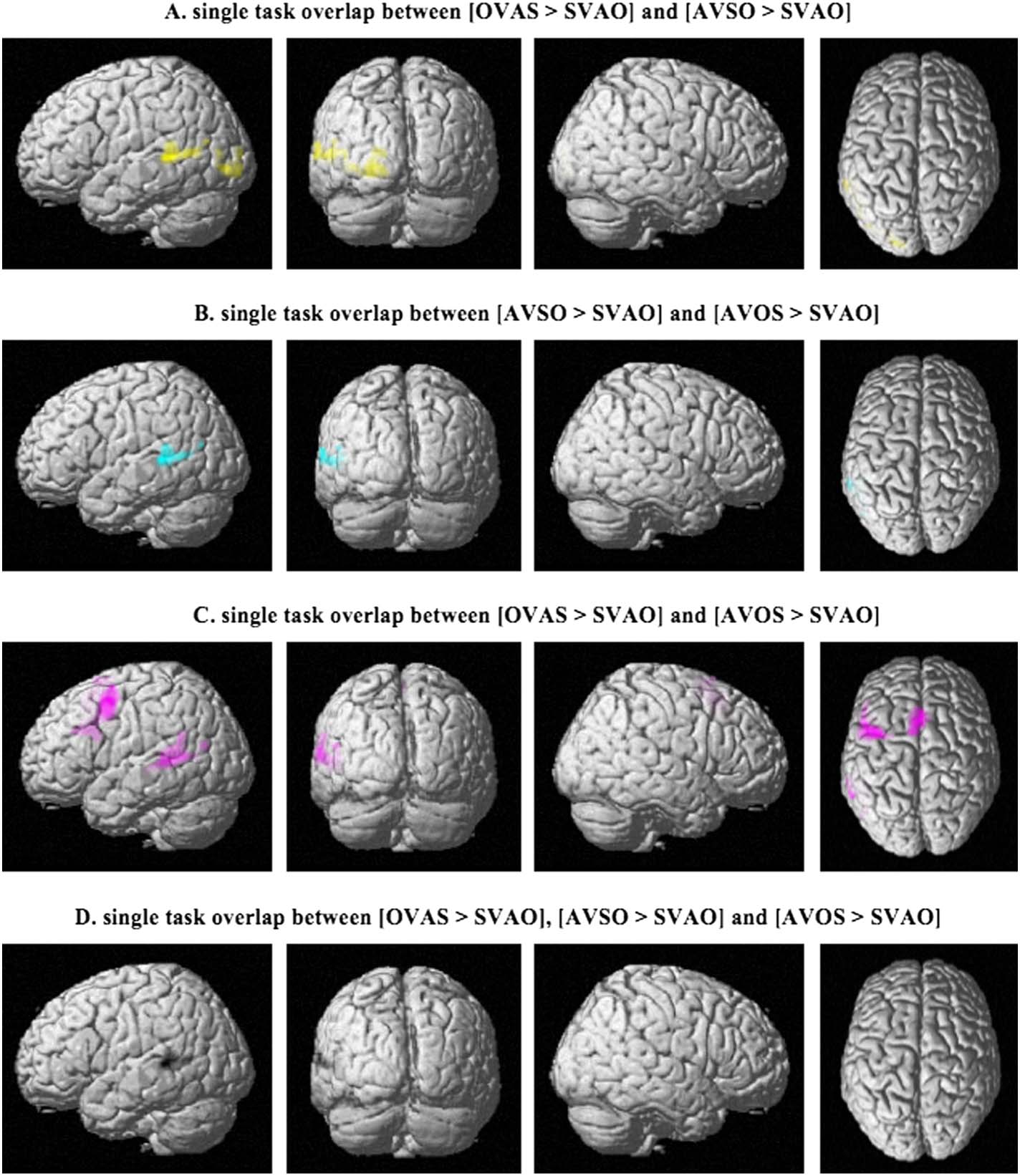
Conjunction analyses were used to quantify the overlap between the different comparisons
as shown in Figure 3D. These were performed for each set of contrasts separately in the single
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 5. Pairwise overlap in activity between the different sentence conditions in the single task
based on conjunction analyses ( P < 0.05; family-wise error corrected on the cluster level). The overlap
between all three comparisons is shown in (D). S = subject, V = verb, A = adjunct, O = object.
Neurobiology of Language
239
Processing complex sentences: An fMRI study
Table 4. Peak MNI coordinates for the overlap between the different sentence comparisons in the single task (based on conjunction analyses)
Conjunction
[OVAS > SVAO] E [AVSO > SVAO]
[AVSO > SVAO] E [AVOS > SVAO]
[OVAS > SVAO] E [AVOS > SVAO]
[OVAS > SVAO], [AVSO > SVAO]
E [AVOS > SVAO]
Peak coordinates
(X, sì, z)
(−50, −40, 4)
(−22, 94, −4)
(−50, −40, 4)
(−46, 4, 54)
(−52, −38, 4)
(−4, 18, 52)
(−50, −40, 4)
Z-value
4.11
Cluster size
422
Brain region
Left superior temporal gyrus
4.11
4.43
5.65
4.89
4.73
4.11
439
349
837
713
874
348
Left inferior occipital gyrus
Left superior temporal gyrus
Left middle frontal gyrus
Left superior temporal gyrus
Left supplementary motor cortex
Left superior temporal gyrus
Note. S = subject, V = verb, A = adjunct, O = object.
task (cioè., the overlap between OVAS > SVAO and AVSO > SVAO, the overlap between
AVSO > SVAO and AVOS > SVAO, and the overlap between OVAS > SVAO and AVOS >
SVAO), as well as the overlap between all three different contrasts. The results, presented in
Figura 5 and Table 4, show that although the complex sentence conditions show overlap in
their activation patterns, the only region that is activated in all three contrasts is the left
superior temporal gyrus (Figure 5D).
Dual task
The dual task analyses investigated the same contrasts as the single task analyses, as well as
the difference between the single and the dual task in these contrasts. Within the dual task, NO
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
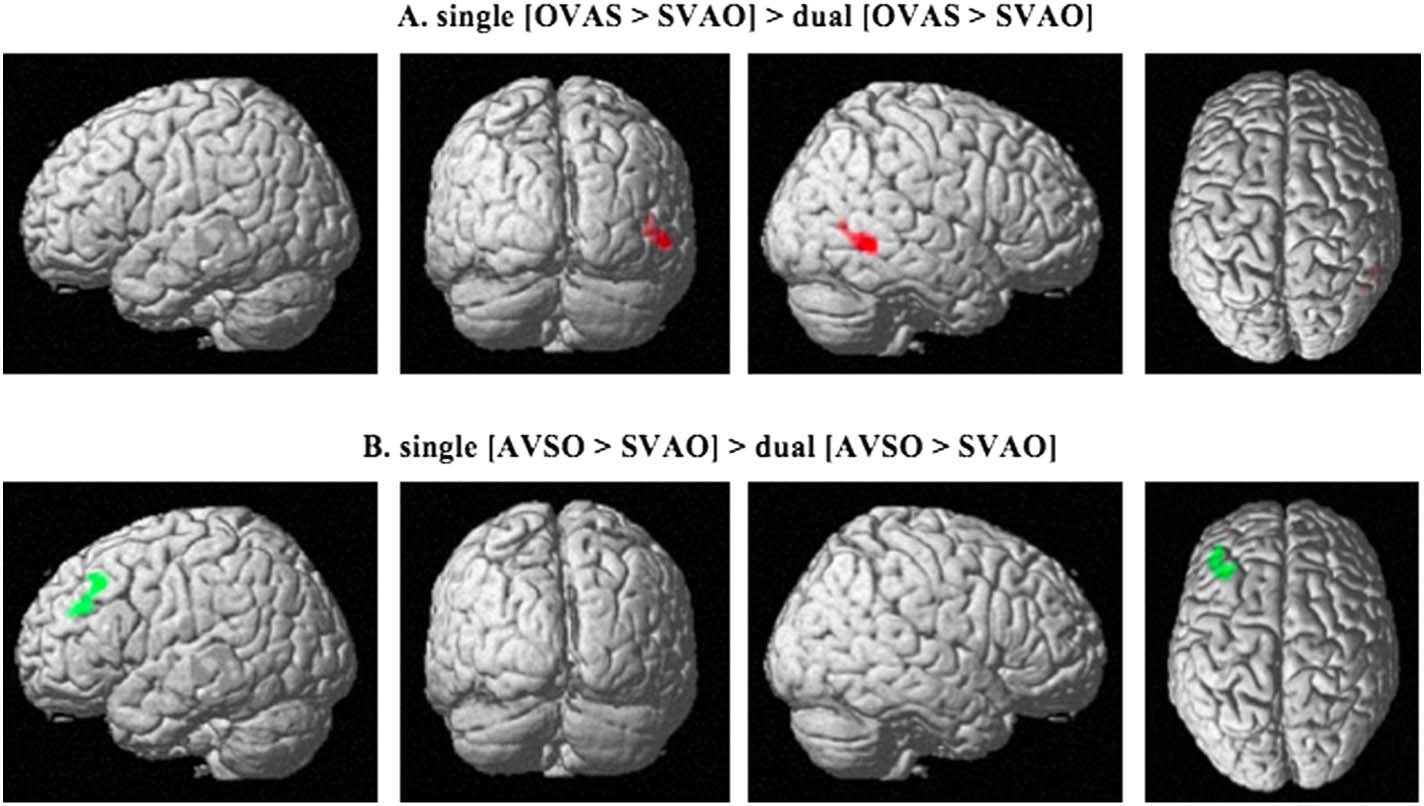
Figura 6. Activation patterns for the comparisons between sentence types in the single vs. dual task. The top row shows the comparison
between object-initial word order and canonical word order in the single compared to the dual task (single [OVAS > SVAO] > dual [OVAS >
SVAO]). The bottom row shows the comparison between adjunct-initial word order and canonical word order in the single compared to the
dual task (single [AVSO > SVAO] > dual [AVSO > SVAO]). Activated areas include the right middle temporal gyrus in (UN), and left middle
frontal gyrus in (B), P < 0.05; family-wise error corrected on the cluster level. S = subject, V = verb, A = adjunct, O = object.
Neurobiology of Language
240
Processing complex sentences: An fMRI study
Table 5. Peak MNI coordinates for the different comparisons between the single and the dual task
Comparison
single [OVAS > SVAO] >
dual [OVAS > SVAO]
single [AVSO > SVAO] >
dual [AVSO > SVAO]
Peak coordinates
(X, sì, z)
(54, −50, −4)
Z-value
4.56
Cluster size
238
Brain region
Right middle temporal gyrus
(−42, 42, 26)
3.88
214
Left middle frontal gyrus
Note. S = subject, V = verb, A = adjunct, O = object.
effects of object-initial word order compared to canonical word order (OVAS > SVAO), Di
adjunct-initial word order compared to canonical word order (AVSO > SVAO), or of adjunct-
initial object-before-subject word order compared to canonical word order (AVOS > SVAO)
were found. So, the dual task seems to reduce or even eliminate the effects of syntactic complex-
ity that were found in the single task. When comparing the object-initial to the adjunct-initial
condition (OVAS > AVSO), activity was found in the brain stem [peak coordinate (−6, −40, −24),
Z = 4.14, cluster size = 194].
Examining the difference between the single and the dual task, the results show effects for
both [OVAS > SVAO] E [AVSO > SVAO] in the [single-dual] comparison, but not in the
[dual-single] comparison. Specifically, an increased difference in activity was found in the sin-
gle task compared to the dual task in the [OVAS > SVAO] contrast in the right MTG (Figure 6A,
Tavolo 5) and in the [AVSO > SVAO] contrast in the left MFG (Figure 6B, Tavolo 5). No effects of
[AVOS > SVAO], [OVAS > AVSO], [AVOS > OVAS], O [AVOS > AVSO] were found in the
[single-dual] comparison or in the [dual-single] comparison. Così, as in the single task, IL
dual task comparisons show no differences in the activity between noncanonical conditions.
To investigate these effects in more detail, we extracted the beta values from the peak co-
ordinates listed in Table 5 and plotted these for visual inspection. In the right MTG, the OVAS
condition is largely unaffected by the task factor, whereas the activity associated with the pro-
cessing of canonical SVAO sentences is increased in the dual task compared to the single task
(Figure 7A). A similar pattern can be seen for the AVSO (unaffected by task) compared to
SVAO (affected by task) sentences in left MFG (Figure 7B). Così, it seems that the processing
of canonical SVAO sentences is affected by the dual task more than the processing of complex
OVAS or AVSO sentences.
Figura 7. Average beta values extracted from the right middle temporal gyrus (MTG) for the OVAS and
SVAO conditions in the single and the dual task (left graph) and from the left middle frontal gyrus (MFG)
for the AVSO and SVAO conditions in the single and the dual task (right graph), at the peak values listed
in Table 5. Error bars indicate standard error of the mean. S = subject, V = verb, A = adjunct, O = object.
Neurobiology of Language
241
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
Figura 8. Average beta values extracted from the inferior frontal gyrus (IFG), middle temporal
gyrus (MTG), superior temporal gyrus (STG), and inferior occipital gyrus (IOG) in all four sentence
conditions in the dual task. Beta values were extracted from the entire mentioned regions using
WFU PickAtlas (Maldjian et al., 2003, 2004). Error bars indicate standard error of the mean. S =
subject, V = verb, A = adjunct, O = object.
Again, the activity in each sentence condition was further examined by extracting the beta
values in the four key regions for visual inspection. The results for the single as well as the dual
task conditions are plotted in Figure 4 (single task) and Figure 8 (dual task). The graphs show, in line
with previous whole-brain analyses, that whereas in the single task condition SVAO sentences
show lower beta values than the complex sentences, in the dual task conditions these differences
have decreased, showing little difference in activity between canonical and complex sentence
conditions. We statistically investigated these differences with region of interest (ROI) analyses
and found a significant effect for the comparison between object-initial word order and canonical
word order in the single compared to the dual task (single [OVAS > SVAO] > dual [OVAS >
SVAO]) in the left MTG [peak coordinate (−52, −66, 16), Z = 3.56, cluster size = 83].
Correlations with cognitive capacities
Finalmente, we investigated the influence of different cognitive capacities with multiple regression
analyses for those comparisons that showed significant effects of word order in the previous
analyses. Therefore, we examined the relation between the processing of syntactically com-
plex sentences (OVAS > SVAO, AVSO > SVAO, and AVOS > SVAO) in the single task on the
one hand and digit span score, vocabulary score, and CTMT score on the other. No correla-
tions between these cognitive capacities and processing of complex sentences were found.
DISCUSSION
In this study, we investigated the mechanisms underlying the processing of different nonca-
nonical, complex sentences by older adults. Our main hypothesis was that distinct linguistic
Neurobiology of Language
242
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
operations underlie the processing of different noncanonical sentences, which were predicted
to be reflected in distinct patterns of neural activity. The behavioral results showed that object-
initial sentences and adjunct-initial sentences with the object before the subject lead to differ-
ent performance levels compared to canonical sentences and compared to each other in the
picture-selection task. These results support the idea that these noncanonical word orders af-
fect processing and therefore offline interpretation, even if the interpretation (of the adjunct) È
not directly assessed in the task. Adjunct-initial sentences with the subject before the object
were not found to be interpreted differently from canonical sentences.
In the imaging data, used to examine the online processing and neural patterns underlying
this processing, the results are less clear. Despite the finding that the differences in activity
between object-initial and adjunct-initial sentences were not significant, our noncanonical
sentence conditions did show clearly different neural processing when compared to canonical
sentences. Specifically, object-initial sentences showed activity in the left IFG and temporal
regions, which are more frequently associated with object-initialization (Bahlmann et al.,
2007; Bornkessel et al., 2005; Röder et al., 2002; Shetreet & Friedmann, 2014) and complex
sentence processing in general (Rodd et al., 2015). Adjunct-initial sentences with the subject
before the object showed activity in occipital regions, in line with Shetreet and Friedmann
(2014), and additionally showed activity in right superior frontal areas. Although typically as-
sociated with visual processing, occipital regions have been associated with the processing of
complex sentences before (Peelle, McMillan, Moore, Grossman, & Wingfield, 2004; Shetreet
& Friedmann, 2014) and are connected to the inferior frontal cortex through a ventral pathway
(Friederici & Gierhan, 2013). In congenitally blind adults, occipital regions have been found to
be sensitive to syntactic complexity, but not to complexity in mathematical equations (Lane,
Kanjlia, Omaki, & Bedny, 2015), and show behavior similar to classic language regions (left
frontal and temporal regions) during sentence processing (Bedny, Pascual-Leone, Dodell-
Feder, Fedorenko, & Saxe, 2011), although their sighted peers did not display such occipital
activity. This indicates that the role of occipital activation in sentence processing requires
further research. Finalmente, adjunct-initial sentences with the object before the subject com-
pared to canonical sentences showed activity that partially overlapped with activity found for
the object-initial sentences (left MFG, left STG, and left supplementary motor cortex) E
adjunct-initial sentences with the subject before the object (left STG). È interessante notare, neither
our study nor the study of Shetreet and Friedmann (2014) found increased activity in the IFG
for adjunct-initial sentences in which the verb precedes its arguments, whereas many previ-
ous studies, which used various sentence structures, systematically found activity in this re-
gion (see Rodd et al., 2015, for an overview). It has been shown, Tuttavia, that effects and
activity can vary between different types of sentence manipulations, such as embedding
(Santi & Grodzinsky, 2010) and scrambling (Makuuchi et al., 2013) compared to movement
or even between different types of movement (Shetreet & Friedmann, 2014), which may
explain the absence of an effect in the IFG for our adjunct-initial manipulations. Inoltre,
in the additional analyses including only the audio (without the jitter or pause following the
audio) described in the Materials and Methods section, left IFG activation did show for
adjunct-initial AVSO sentences, indicating that the length of the time window affects the
detection of this activity. Conjunction analyses showed that all noncanonical sentence
conditions shared activation in the left posterior STG, which is traditionally considered to
be involved in speech and language comprehension, and more specifically in the processing
of noncanonical sentences (Friederici, Kotz, Scott, & Obleser, 2010). Così, we found effects
of object-initialization and adjunct-initialization (and thus verb-argument order) that are in
line with the literature, and novel results for adjunct-initial sentences with the subject before
Neurobiology of Language
243
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
the object. Tuttavia, we could not replicate Shetreet and Friedmann’s (2014) findings on the
differences in neural activity between noncanonical object-initial and adjunct-initial sentences
in our sample of older, German-speaking adults. This may be explained by differences in the
experimental design (cioè., our pseudo-randomized design vs. the blocked design that
Shetreet and Friedmann used). Inoltre, the difference in findings may be explained
by differences between the two tested languages, such as differences in the case marking
system (only the accusative is morphologically marked in Hebrew, through an extra accusa-
tive marker “et,” whereas German marks all cases, in our study on the article of the noun
phrase). Inoltre, the position of the verb is different in Hebrew and German, Quale
may be an underlying factor. For Hebrew, it has been argued to always be in second position
(SVO); whereas for German, it has been argued to be in final position in the underlying word
order (see Introduction). Hence, one could argue that in our German sentences, verb move-
ment took place in all four conditions, whereas in Hebrew, verb movement was the actual
manipulation in one of the conditions. The finding that there are no significant differences in
the neural processing of different complex sentences seems more in line with a memory-based
account of sentence processing than a linguistic operations account. Tuttavia, since absence
of evidence is not evidence of absence, these results are to be interpreted with caution.
To investigate the processing of different noncanonical sentences further, a dual task par-
adigm was used. Contrary to our expectations, the behavioral results showed no differences
between the single and the dual task. As the dual task occurred during sentence processing, Esso
is possible that the effects of the increased load were not present anymore during the response
phase. In contrast to the behavioral results, the imaging results showed that neural processing
in the dual task was different compared to the single task. Specifically, whereas the single task
showed differences in activity between canonical sentences on the one hand and noncanon-
ical sentences on the other, these differences disappeared in the dual task. So, increased cog-
nitive load seems to be able to affect the processing of different sentence structures differently,
increasing neural activity for canonical sentences, but leaving the noncanonical sentences rel-
atively unaffected. Differences in differential activity for canonical vs. noncanonical sentences
between the single and the dual task were found in the right MTG and left MFG. Although
these were not the regions that we hypothesized finding differences in, thus making the effects
more difficult to interpret, additional examinations of the data suggest similar effects in the key
ROIs. Infatti, ROI analyses confirmed a similar effect in the left MTG. It is difficult to find direct
support for either the linguistic operations account or memory-based accounts in these find-
ing. The findings could be interpreted within the linguistic operations account, which as-
sumes that different word orders are derived from distinct linguistic analyses, which could
thus be affected differently by increased cognitive load. Tuttavia, based on this account
we would have expected to find differences between noncanonical sentences as well, Quale
were not found in the data. It is also difficult to reconcile with experience- or memory-based
accounts (Hale, 2001; Levy, 2008; Lewis et al., 2006), as in our understanding these accounts
would predict all sentences to be affected in a similar way, with the differences between sen-
tences still being present. Inoltre, the accounts themselves do not make claims about the
neural processing of different sentences, so more precise predictions on the basis of these
accounts should be formulated in future work. The finding that different sentence structures
are differentially affected by cognitive load may point more toward the linguistic operations
account, but certainly there is a role of memory and other cognitive resources in (complex)
sentence processing as well; the existence of examples in which different sentences are affected
by increased cognitive load in different ways merely suggests that a complete account cannot
be solely memory-based. Overall, we tentatively interpret the results as providing support for
Neurobiology of Language
244
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
the idea that distinct mechanisms, that could be explained by distinct underlying linguistic
operations, underlie the processing of different sentences. Nevertheless, the nature of these
processing differences and the relation between the identified brain regions and underlying
processing mechanisms for different sentences remains to be investigated further.
Finalmente, we examined the influence of several cognitive capacities on sentence processing.
In the behavioral data, effects of WM capacity, verbal intelligence, and cognitive flexibility
were found, indicating that these cognitive capacities interact with the interpretation of (com-
plex) sentences. In the imaging data, no effects of cognitive capacities were found. The differ-
ences between the behavioral and the imaging results could be due to (1) statistical corrections
(cioè., at the whole-brain level) for the imaging data, (2) the imaging data testing the correlation
of cognitive capacities with differential activity in the different sentence conditions rather than
with performance on one condition (as was done for the behavioral data), O (3) individual
variation in the brain areas that are influenced by these cognitive capacities. Further research
is needed to investigate these different explanations.
Some notes can be made with regards to the experimental design. We chose to test older
participants, as these were expected to show more difficulties with the processing of complex
sentences (cf. Emery, 1985; Kemper, 1992; Wingfield et al., 2006) and a wider spread in their
cognitive capacities than younger adults, which would aid the planned correlational analyses.
Infatti, the behavioral results show that our participants had quite some difficulties correctly
interpreting the two object-before-subject conditions. This may in part be due to their age and
in part be due to the MRI noise (essentially creating a speech-in-noise task), which can influ-
ence the processing of syntactically complex sentences (Carroll & Ruigendijk, 2013).
Tuttavia, the neuroimaging analyses did not show the expected effects between different non-
canonical sentences and no correlations with cognitive capacities. These comparisons may
have been constrained by the limited number of participants (although we had the same num-
ber of participants as Shetreet & Friedmann, 2014, namely, 22) and/or the strict corrections
that were applied at the whole-brain level (as recommended for SPM12). Future research
should therefore try to further clarify the differences in the processing of complex sentences
and the cognitive capacities that play a role in this processing, potentially focusing on different
populations (per esempio., older vs. younger) and different cognitive skill levels (per esempio., higher vs. inferiore
WM skills). Inoltre, sentences with different word orders inevitably also differ pragmati-
cally. Although we acknowledge this confound, previous studies on word order variations
have found very similar results for the syntactic processing of canonical compared to nonca-
nonical sentences, and the observed activated areas are therefore assumed to be due to syn-
tactic differences. Finalmente, as different sentence structures are affected differently and canonical
sentences behave more like complex sentences under increased cognitive load, our findings
indicate that investigations of language processing under challenging conditions will be par-
ticularly relevant, for instance, in hearing-impaired populations or during speech processing in
background noise (cf. Carroll & Ruigendijk, 2013).
In conclusion, we presented behavioral and imaging data on the processing of two types of
sentence complexity in German. The results are mixed, with the imaging results not replicating
differences between the processing of different sentences that were found in previous research
(Shetreet & Friedmann, 2014), but at the same time showing differential effects of increased
cognitive load for different sentence types. We tentatively interpret the results on complex sen-
tence processing under cognitive load as supporting the notion of distinct mechanisms under-
lying the processing of different sentence structures, in line with the idea of distinct linguistic
operations being performed. Tuttavia, follow-up research is needed to investigate the asserted
Neurobiology of Language
245
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
linguistic operations and their (neural) processing signatures in more detail, possibly with a
large-scale study.
ACKNOWLEDGMENTS
The authors would like to thank all participants for their participation. Inoltre, the authors
would like to thank Jan Michalsky for his help with recording the stimuli, Rebecca Carroll for
her help with recording and preparing the stimuli, our students and research assistants (Regina
Hert, Laura Peters, Anne Lina Voß, and Charlotte Sielaff ) for their help with the data collection,
and Gülsen Yanç and Katharina Grote for their support during MRI data acquisition.
FUNDING INFORMATION
This work was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research
Foundation) under Germany’s Excellence Strategy – EXC 2177/1 – Project ID 390895286.
This work was supported by the Neuroimaging Unit of the Carl von Ossietzky Universität
Oldenburg funded by grants from the German Research Foundation (3T MRI INST 184/152-1
FUGG and MEG INST 184/148-1 FUGG).
AUTHOR CONTRIBUTIONS
Margreet Vogelzang: Project administration, Conceptualization, Methodology, Formal analy-
sis, Writing. Christiane M. Thiel: Funding acquisition, Conceptualization, Writing. Stephanie
Rosemann: Methodology, Data curation, Writing. Jochem W. Rieger: Funding acquisition,
Conceptualization, Writing. Esther Ruigendijk: Funding acquisition, Conceptualization,
Methodology, Writing.
REFERENCES
Bader, M., & Meng, M. (1999). Subject-object ambiguities in
German embedded clauses: An across-the-board comparison.
Journal of Psycholinguistic Research, 28(2), 121–143. https://
doi.org/10.1023/A:1023206208142
Bahlmann, J., Rodriguez-Fornells, A., Rotte, M., & Münte, T. F.
(2007). An fMRI study of canonical and noncanonical word or-
der in German. Human Brain Mapping, 28(10), 940–949. https://
doi.org/10.1002/hbm.20318
Bedny, M., Pascual-Leone, A., Dodell-Feder, D., Fedorenko, E., &
Saxe, R. (2011). Language processing in the occipital cortex of
congenitally blind adults. Proceedings of the National
Academy of Sciences of the United States of America, 108(11),
4429–4434. https://doi.org/10.1073/pnas.1014818108
Ben-Shachar, M., Hendler, T., Kahn, I., Ben-Bashat, D., &
Grodzinsky, Y. (2003). The neural reality of syntactic transforma-
zioni. Psychological Science, 14(5), 433–440. https://doi.org/
10.1111/1467-9280.01459
Ben-Shachar, M., Palti, D., & Grodzinsky, Y. (2004). Neural corre-
lates of syntactic movement: Converging evidence from two
fMRI experiments. NeuroImage, 21(4), 1320–1336. https://doi.
org/10.1016/j.neuroimage.2003.11.027
Bornkessel, I., Zysset, S., Friederici, UN. D., von Cramon, D. Y., &
Schlesewsky, M. (2005). Who did what to whom? The neural ba-
sis of argument hierarchies during language comprehension.
NeuroImage, 26(1), 221–233. https://doi.org/10.1016/j.
neuroimage.2005.01.032
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Broekhuis, H. (2006). The universal base hypothesis: VO or OV? In
J. Van de Weijer & B. Los (Eds.), Linguistics in the Netherlands
(Vol. 23, pag. 28–39). Amsterdam: John Benjamins. https://doi.
org/10.1075/avt.23.06bro
Caplan, D., Alpert, N., & Waters, G. (1999). PET studies of syntactic
processing with auditory sentence presentation. NeuroImage, 9(3),
343–351. https://doi.org/10.1006/NIMG.1998.0412
Carroll, R., & Ruigendijk, E. (2013). The effects of syntactic com-
Jo ur na l o f
p l e x i t y o n p r o c e s s i n g s e n t e n c e s i n n o i s e .
Psycholinguistic Research, 42(2), 139–159. https://doi.org/
10.1007/s10936-012-9213-7
Carroll, R., Uslar, V., Brand, T., & Ruigendijk, E. (2016). in lavorazione
mechanisms in hearing-impaired listeners: Evidence from reac-
tion times and sentence interpretation. Ear and Hearing, 37(6),
e391–e401. https://doi.org/10.1097/AUD.0000000000000339
Chomsky, N. (1981). Lectures on government and binding: IL
Pisa lectures. Dordrecht, Netherlands: Foris Publications.
Cinque, G. (1990). Types of A’-dependencies. Cambridge, MA:
CON Premere.
Constable, R. T., Pugh, K. R., Berroya, E., Mencl, W. E., Westerveld,
M., Ni, W., & Shankweiler, D. (2004). Sentence complexity and
input modality effects in sentence comprehension: An fMRI
study. NeuroImage, 22(1), 11–21. https://doi.org/10.1016/ J.
NEUROIMAGE.2004.01.001
Den Ouden, D.-B., Hoogduin, H., Stowe, l. A., & Bastiaanse, R.
(2008). Neural correlates of Dutch Verb Second in speech
Neurobiology of Language
246
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
Processing complex sentences: An fMRI study
production. Brain and Language, 104(2), 122–131. https://doi.
org/10.1016/J.BANDL.2007.05.001
Emery, O. B. (1985). Language and aging. Experimental Aging
R e s e a r c h , 1 1 ( 1 ) , 3 – 6 0 . h t t p s : / / d o i . o r g / 1 0 . 1 0 8 0 /
03610738508259280
Engel de Abreu, P. M. J., Gathercole, S. E., & Martin, R. (2011).
Disentangling the relationship between working memory and
lingua: The roles of short-term storage and cognitive control.
Learning and Individual Differences, 21(5), 569–574. https://doi.
org/10.1016/j.lindif.2011.06.002
Friederici, UN. D., Fiebach, C. J., Schlesewsky, M., Bornkessel, IO. D.,
& von Cramon, D. Y. (2006). Processing linguistic complexity
and grammaticality in the left frontal cortex. Cerebral Cortex,
16(12), 1709–1717. https://doi.org/10.1093/cercor/bhj106
Friederici, UN. D., & Gierhan, S. M. (2013). The language network.
Current Opinion in Neurobiology, 23(2), 250–254. https://doi.
org/10.1016/J.CONB.2012.10.002
Friederici, UN. D., Kotz, S. A., Scott, S. K., & Obleser, J. (2010).
Disentangling syntax and intelligibility in auditory language
comprehension. Human Brain Mapping, 31(3), 448–457.
https://doi.org/10.1002/hbm.20878
Friederici, UN. D., Mecklinger, A., Spencer, K. M., Steinhauer, K., &
Donchin, E. (2001). Syntactic parsing preferences and their on-
line revisions: A spatio-temporal analysis of event-related brain
potentials. Cognitive Brain Research, 11, 305–323.
Friederici, UN. D., Steinhauer, K., Mecklinger, A., & Meyer, M.
(1998). Working memory constraints on syntactic ambiguity res-
olution as revealed by electrical brain responses. Biological
Psychology, 47(3), 193–221. https://doi.org/10.1016/S0301-
0511(97)00033-1
Grewe, T., Bornkessel, I., Zysset, S., Wiese, R., Von Cramon, D. Y.,
& Schlesewsky, M. (2005). The emergence of the unmarked:
A new perspective on the language-specific function of Broca’s
area. Human Brain Mapping, 26(3), 178–190. https://doi.org/
10.1002/ hbm.20154
Grodner, D., & Gibson, E. (2005). Consequences of the serial na-
ture of linguistic input for sentenial complexity. Cognitive
Scienza, 29(2), 261–290.
Hale, J. (2001). A probabilistic earley parser as a psycholinguistic
modello. In Proceedings of NAACL (pag. 159–166). https://doi.org/
10.3115/1073336.1073357
Hemforth, B. (1993). Kognitives Parsing: Repräsentation und
Verarbeitung sprachlichen Wissens [Cognitive parsing:
Representation and processing of linguistic knowledge]. Sankt
Augustin, Germany: Infix Verlag.
Jackendoff, R. (1990). Semantic structures. Cambridge, MA: MIT
Press.
Just, M. A., Carpenter, P. A., Keller, T. A., Eddy, W. F., & Thulborn,
K. R. (1996). Brain activation modulated by sentence compre-
hension. Scienza, 274(5284), 114–116. Retrieved from http://
www.ncbi.nlm.nih.gov/pubmed/8810246
Kemper, S. (1992). Language and aging. In F. IO. M. Craik &
T. UN. Salthouse (Eds.), The handbook of aging and cognition
(pag. 213–270). Hillsdale, NJ: Erlbaum.
Lane, C., Kanjlia, S., Omaki, A., & Bedny, M. (2015). “Visual” cor-
tex of congenitally blind adults responds to syntactic movement.
Journal of Neuroscience, 35(37), 12859–12868. https://doi.org/
10.1523/JNEUROSCI.1256-15.2015
Levy, R. (2008). Expectation-based syntactic comprehension.
Cognition, 106(3), 1126–1177. https://doi.org/10.1016/j.
cognition.2007.05.006
Lewis, R. L., Vasishth, S., & Van Dyke, J. UN. (2006). Computational
principles of working memory in sentence comprehension.
Trends in Cognitive Sciences, 10(10), 447–454. https://doi.org/
10.1016/j.tics.2006.08.007
Makuuchi, M., Grodzinsky, Y., Amunts, K., Santi, A., & Friederici,
UN. D. (2013). Processing noncanonical sentences in Broca’s re-
gion: Reflections of movement distance and type. Cerebral
Cortex, 23(3), 694–702. https://doi.org/10.1093/cercor/bhs058
Maldjian, J. A., Laurienti, P. J., & Burdette, J. H. (2004). Precentral
gyrus discrepancy in electronic versions of the Talairach Atlas.
NeuroImage, 21(1), 450–455. https://doi.org/10.1016/j.
neuroimage.2003.09.032
Maldjian, J. A., Laurienti, P. J., Kraft, R. A., Burdette, J. H., & Kraft,
R. UN. (2003). An automated method for neuroanatomic and cy-
toarchitectonic atlas-based interrogation of fMRI data sets.
NeuroImage, 19(3), 1233–1239. https://doi.org/10.1016/S1053-
8119(03)00169-1
McAuliffe, M. J., Gibson, E. M. R., Kerr, S. E., Anderson, T., &
LaShell, P. J. (2013). Vocabulary influences older and younger
listeners’ processing of dysarthric speech. The Journal of the
Acoustical Society of America, 134(2), 1358–1368. https://doi.
org/10.1121/1.4812764
Nasreddine, Z. S., Phillips, N. A., Bédirian, V., Charbonneau, S.,
Whitehead, V., Collin, I., … Chertkow, H. (2005). IL
Montreal Cognitive Assessment, MoCA: A brief screening tool
for mild cognitive impairment. Journal of the American
Geriatrics Society, 53(4), 695–699. https://doi.org/10.1111/
j.1532-5415.2005.53221.x
Novick, J. M., Trueswell, J. C., & Thompson-Schill, S. l. (2005).
Cognitive control and parsing: Reexamining the role of Broca’s
area in sentence comprehension. Cognitive, Affective and
Behavioral Neuroscience, 5(3), 263–281. https://doi.org/
10.3758/CABN.5.3.263
Payne, B. R., Grison, S., Gao, X., Christianson, K., Morrow, D. G.,
& Stine-Morrow, E. UN. l. (2014). Aging and individual differ-
ences in binding during sentence understanding: Evidence from
temporary and global syntactic attachment ambiguities.
Cognition, 130(2), 157–173. https://doi.org/10.1016/j.
cognition.2013.10.005
Peelle, J. E., McMillan, C., Moore, P., Grossman, M., & Wingfield,
UN. (2004). Dissociable patterns of brain activity during compre-
hension of rapid and syntactically complex speech: Evidence
from fMRI. Brain and Language, 91(3), 315–325. https://doi.org/
10.1016/j.bandl.2004.05.007
Reynolds, C. R. (2002). Comprehensive Trail-Making Test. Austin,
TX: Pro-Ed.
Rizzi, l. (1997). The fine structure of the left periphery. In L.
Haegeman (Ed.), Elements of grammar: A handbook of genera-
tive syntax (pag. 281–337). Dordrecht, Netherlands: Kluwer.
https://doi.org/10.1007/978-94-011-5420-8_7
Rodd, J. M., Vitello, S., Woollams, UN. M., & Adank, P. (2015).
Localising semantic and syntactic processing in spoken and writ-
ten language comprehension: An activation likelihood estima-
tion meta-analysis. Brain and Language, 141, 89–102. https://
doi.org/10.1016/j.bandl.2014.11.012
Röder, B., Stock, O., Neville, H., Bien, S., & Rösler, F. (2002). Brain
activation modulated by the comprehension of normal and pseudo-
word sentences of different processing demands: A functional mag-
netic resonance imaging study. NeuroImage, 15(4), 1003–1014.
https://doi.org/10.1006/nimg.2001.1026
Rosemann, S., Gießing, C., Özyurt, J., Carroll, R., Puschmann, S., &
Thiel, C. M. (2017). The contribution of cognitive factors to indi-
vidual differences in understanding noise-vocoded speech in young
and older adults. Frontiers in Human Neuroscience, 11(294), 1–13.
https://doi.org/10.3389/fnhum.2017.00294
Neurobiology of Language
247
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Processing complex sentences: An fMRI study
Santi, A., & Grodzinsky, Y. (2010). fMRI adaptation dissociates syn-
tactic complexity dimensions. NeuroImage, 51(4), 1285–1293.
https://doi.org/10.1016/j.neuroimage.2010.03.034
Schlesewsky, M., Fanselow, G., Kliegl, R., & Krems, J. (2000). IL
subject preference in the processing of locally ambiguous wh-
questions in German. In B. Hemforth & l. Konieczny (Eds.),
German sentence processing (pag. 65–93). Dordrecht,
Netherlands: Kluwer Academic. https://doi.org/10.1007/978-94-
015-9618-3_3
Schmidt, K.-H., & Metzler, P. (1992). Wortschatztest [Multiple
Choice Word Test]. Weinheim: Beltz Test GmbH.
Shetreet, E., & Friedmann, N. (2014). The processing of different
syntactic structures: fMRI investigation of the linguistic distinc-
tion between wh-movement and verb movement. Journal of
Neurolinguistics, 27(1), 1–17. https://doi.org/10.1016/j.
jneuroling.2013.06.003
Shetreet, E., Friedmann, N., & Hadar, U. (2009). An fMRI study of
syntactic layers: Sentential and lexical aspects of embedding.
NeuroImage, 48(4), 707–716. https://doi.org/10.1016/j.
neuroimage.2009.07.001
Stromswold, K., Caplan, D., Alpert, N., & Rauch, S. (1996).
Localization of syntactic comprehension by positron emission to-
mography. Brain and Language, 52(3), 452–473. https://doi.org/
10.1006/brln.1996.0024
Tun, P. A., Benichov, J., & Wingfield, UN. (2010). Response latencies
in auditory sentence comprehension: Effects of linguistic versus
perceptual challenge. Psychology and Aging, 25(3), 730–735.
https://doi.org/10.1037/a0019300
Uslar, V. N., Carroll, R., Hanke, M., Hamann, C., Ruigendijk, E.,
Brand, T., & Kollmeier, B. (2013). Development and evalua-
tion of a linguistically and audiologically controlled sentence
intelligibility test. The Journal of
the Acoustical Society
of America, 134(4), 3039–3056. https://doi.org/10.1121/
1.4818760
Vogelzang, M., Thiel, C. M., Rosemann, S., Rieger, J. W., &
Ruigendijk, E. (2019). Cognitive abilities to explain individual
variation in the interpretation of complex sentences by older
adults. In Proceedings of the 41th Annual Conference of the
Cognitive Science Society (pag. 3036–3043). Montreal, Canada:
Cognitive Science Society.
Vos, S. H., Gunter, T. C., Schriefers, H., & Friederici, UN. D. (2001).
Syntactic parsing and working memory: The effects of syntactic
complexity, reading span, and concurrent load. Language and
Cognitive Processes, 16(1), 65–103. https://doi.org/10.1080/
01690960042000085
Wagener, K. C., Brand, T., & Kollmeier, B. (1999UN). Entwicklung und
evaluation eines satztests in deutscher sprache II: Optimierung
des oldenburger satztests [Development and evaluation of a
german sentence test part II: Optimization of the oldenburg
sentence test]. Zeitschrift Für Audiologie, 38, 44–56.
Wagener, K. C., Brand, T., & Kollmeier, B. (1999B). Entwicklung
und evaluation eines satztests in deutscher sprache III: evaluation
des oldenburger satztests [Development and evaluation of a
German sentence test Part III: Evaluation of the Oldenburg sen-
tence test]. Zeitschrift Für Audiologie, 38, 86–95.
Wagener, K. C., Kühnel, V., & Kollmeier, B. (1999). Entwicklung
und evaluation eines satztests in deutscher sprache I: Design
des oldenburger satztests [Development and evaluation of a
German sentence test I: Design of the oldenburg oldenburg sen-
tence test]. Zeitschrift Für Audiologie, 38, 4–15.
Wendt, D., Brand, T., & Kollmeier, B. (2014). An eye-tracking par-
adigm for analyzing the processing time of sentences with differ-
ent linguistic complexities. PLOS ONE, 9(6). https://doi.org/
10.1371/journal.pone.0100186
Wendt, D., Kollmeier, B., & Brand, T. (2015). How hearing impair-
ment affects sentence comprehension: Using eye fixations to in-
vestigate the duration of speech processing. Trends in Hearing,
19, 1–18. https://doi.org/10.1177/2331216515584149
Wingfield, A., McCoy, S. L., Peelle, J. E., Tun, P. A., & Cox, C. l.
(2006). Effects of adult aging and hearing loss on comprehension
of rapid speech varying in syntactic complexity. Journal of the
American Academy of Audiology, 17(7), 487–497. https://doi.
org/10.3766/jaaa.17.7.4
Zwart, C. J. W. (1997). The Germanic SOV languages and the uni-
versal base hypothesis. In L. Haegeman (Ed.), The new compar-
ative syntax (pag. 246–267). London: Longman.
Neurobiology of Language
248
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
/
/
/
1
2
2
2
6
1
8
6
7
7
1
2
N
o
_
UN
_
0
0
0
1
1
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3