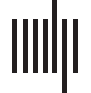Research
Modeling the influence of the hippocampal
memory system on the oculomotor system
Jennifer D. Ryan
1
John Griffiths
1,2∗
, Kelly Shen
3
, Gleb Bezgin
1∗
1
, Arber Kacollja
, Heather Tian
1
,
, and Anthony R. McIntosh
1,2
1Rotman Research Institute, Baycrest, Toronto, Ontario, Canada
2Department of Psychology, University of Toronto, Toronto, Ontario, Canada
Montreal Neurological Institute, McGill University, Montreal, Quebec, Canada
3
Keywords: Hippocampus, Eye movements, Functional Dynamics, Memory, Oculomotor guidance
a n o p e n a c c e s s
j o u r n a l
ABSTRACT
Visual exploration is related to activity in the hippocampus (HC) and/or extended medial
temporal lobe system (MTL), is influenced by stored memories, and is altered in amnesic
cases. An extensive set of polysynaptic connections exists both within and between the HC
and oculomotor systems such that investigating how HC responses ultimately influence
neural activity in the oculomotor system, and the timing by which such neural modulation
could occur, is not trivial. We leveraged TheVirtualBrain, a software platform for large-scale
network simulations, to model the functional dynamics that govern the interactions between
the two systems in the macaque cortex. Evoked responses following the stimulation of the
MTL and some, but not all, subfields of the HC resulted in observable responses in
oculomotor regions, including the frontal eye fields, within the time of a gaze fixation.
Modeled lesions to some MTL regions slowed the dissipation of HC signal to oculomotor
regions, whereas HC lesions generally did not affect the rapid MTL activity propagation to
oculomotor regions. These findings provide a framework for investigating how information
represented by the HC/MTL may influence the oculomotor system during a fixation and
predict how HC lesions may affect visual exploration.
AUTHOR SUMMARY
No major account of oculomotor (eye movement) guidance considers the influence of the
hippocampus (HC) and broader medial temporal lobe (MTL) system, yet it is clear that
information is exchanged between the two systems. Prior experience influences current
viewing, and cases of amnesia due to compromised HC/MTL function show specific
alterations in viewing behavior. By modeling large-scale network dynamics, we show that
stimulation of subregions of the HC, and of the MTL, rapidly results in observable responses
in oculomotor control regions, and that HC/MTL lesions alter signal propagation. These
findings suggest that information from memory may readily guide visual exploration, and call
for a reconsideration of the neural circuitry involved in oculomotor guidance.
INTRODUCTION
Memory influences ongoing active exploration of the visual environment (Hannula et al.,
2010). For instance, more viewing is directed to novel versus previously viewed items (Fagan,
1970; Fantz, 1964), and more viewing is directed to areas that have been altered from a
prior viewing (Ryan, Althoff, Whitlow, & Cohen, 2000; Smith, Hopkins, & Squire, 2006). UN
Citation: Ryan, J. D., Shen, K., Kacollja,
A., Tian, H., Griffiths, J., Bezgin, G., &
McIntosh, UN. R. (2020). Modeling the
influence of the hippocampal memory
system on the oculomotor system.
Network Neuroscience, 4(1), 217–233.
https://doi.org/10.1162/netn_a_00120
DOI:
https://doi.org/10.1162/netn_a_00120
Supporting Information:
https://doi.org/10.1162/netn_a_00120
Received: 31 Luglio 2019
Accepted: 4 Dicembre 2019
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Jennifer D. Ryan
jryan@research.baycrest.org
Kelly Shen
kshen@research.baycrest.org
Handling Editor:
Michael Cole
Copyright: © 2019
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Simulated dynamics of hippocampal responses in oculomotor regions
Hippocampus:
A region within the medial temporal
lobe of the brain that has a critical
role in the formation of memory
representations that consist of the
relations among distinct elements.
Medial temporal lobe:
A collection of neural regions that
includes the hippocampus, subicular
complex, and surrounding entorhinal
cortex, parahippocampal cortex, E
perirhinal cortex, which collectively
support cognitive functions such as
perception, navigation, and memory.
Visual exploration:
The manner by which the eyes move
to explore the visual world. Entails
the active process of alternating
ballistic movements of the eyes
(saccades)and pauses by the eyes in
order to inspect regions in space
(gaze fixations).
Oculomotor system:
A distributed network of neural
regions that provide cognitive and
motor control over the movement of
the eyes.
number of studies have implicated a network of subregions within the hippocampus (HC)
and/or broader medial temporal lobe (MTL) responsible for the influence of memory on viewing
behavior. Amnesic cases who have severe memory impairments due to compromised function
of the HC and/or MTL show changes in their viewing behavior compared with neurologically
intact cases (Chau, Murphy, Rosenbaum, Ryan, & Hoffman, 2011; Hannula, Ryan, Tranel, &
Cohen, 2007; Olsen et al., 2015; Ryan et al., 2000; Warren, Duff, Tranel, & Cohen, 2010).
Similar findings have been observed in older adults who have suspected HC/MTL compromise
(Ryan, Leung, Turk-Browne, & Hasher, 2007), and certain viewing patterns have been shown
to track with entorhinal cortex (ERC) volumes (Yeung et al., 2017). Visual exploration predicts
HC activity during encoding (Liu, Shen, Olsen, & Ryan, 2017), E, conversely, HC/MTL ac-
tivity predicts ongoing visual exploration that is indicative of memory retrieval (Hannula &
Ranganath, 2009; Ryals, Wang, Polnaszek, & Voss, 2015). The relationship between visual
sampling and HC activity is weakened in aging, presumably because of decline in HC struc-
ture or function (Liu, Shen, Olsen, & Ryan, 2018). Such evidence collectively demonstrates that
HC/MTL function is related to oculomotor behavior. The indirect implication of these studies
is that the HC must influence neural activity in the oculomotor system.
Studies in nonhuman primates have shown that HC/MTL activity is linked to oculomotor
behavior. The activity of grid cells in the ERC is tied to eye position (Killian, Jutras, & Buffalo,
2012), while HC/MTL activity is modulated by saccades (Sobotka, Nowicka, & Ringo, 1997)
and fixations (Hoffman et al., 2013; Leonard et al., 2015). How HC/MTL activity traverses the
brain to influence the oculomotor system has not been shown to date. The oculomotor system
is itself a highly recurrent and distributed network (Parr & Friston, 2017) composed of corti-
cal and subcortical regions responsible for the execution of a saccade (per esempio., frontal eye field,
FEF; superior colliculus, SC) as well as regions that exert cognitive control over where the eyes
should go (per esempio., dorsolateral prefrontal cortex, dlPFC; anterior cingulate cortex, ACC; lateral
intraparietal area, area LIP) (Bisley & Mirpour, 2019; Johnston & Everling, 2008). Prior work
has speculated as to which regions of the brain may be important for bridging the memory and
oculomotor systems (per esempio., Meister & Buffalo, 2016; Micic, Ehrlichman, & Chen, 2010), Ma
these discussions were limited to regions examined in isolation. There are no known direct
connections between hippocampal subfields and the oculomotor system. Yet, by examining
whole-cortex connectivity, we have shown that there is an extensive set of polysynaptic path-
ways spanning extrastriate, posterior parietal, and prefrontal regions that may mediate the ex-
change of information between the oculomotor and memory systems (Shen, Bezgin, Selvam,
McIntosh, & Ryan, 2016). Given the vast anatomical connectivity within and between the
memory and oculomotor systems, trying to discern the functional network involved in bridg-
ing them is not a trivial problem to tackle. Specifically, the large and complex contribution
of recurrent anatomical connections to the functional dynamics of large-scale brain networks
must be considered (Spiegler, Hansen, Bernard, McIntosh, & Jirsa, 2016). One crucial ques-
tion concerning such functional dynamics is whether HC/MTL activity is able to influence the
activity related to the preparation of a saccade. To impact ongoing visual exploration, HC/MTL
activity would likely need to resolve in the oculomotor system within the time of an average
duration of a gaze fixation (∼250–400 ms) (Henderson, Nuthmann, & Luke, 2013).
To examine the extent to which HC/MTL activity could influence the oculomotor system,
we leveraged a computational modeling and neuroinformatics platform, TheVirtualBrain, E
simulated the functional dynamics of a whole-cortex directed macaque network when stim-
ulation is applied to HC and MTL nodes of interest. Critically, we examined whether and
when evoked activity culminated in responses in key regions within the oculomotor system.
Finalmente, we observed the extent to which the propagation and timing of such activity was altered
Network Neuroscience
218
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Simulated dynamics of hippocampal responses in oculomotor regions
Amnesia:
A severe memory disorder that arises
because of lesion or dysfunction of
the hippocampus and/or surrounding
medial temporal lobe. Individuals
with amnesia may be unable to
retrieve previously established
ricordi (retrograde amnesia) E
form new memories (anterograde
amnesia).
following lesions to one or more HC/MTL regions in order to understand the neural dynam-
ics that may underlie altered visual exploration in cases of HC/MTL dysfunction, such as in
amnesia or aging.
RESULTS
We modeled the influence of HC/MTL activity on the oculomotor system using a connectome-
based approach using TheVirtualBrain (see Methods for details). Following Spiegler and col-
leagues (Spiegler et al., 2016), we assigned a neural mass model to each node and set each to
operate near criticality, which is considered to be the point at which information processing
capacity is maximal (Deco et al., 2014; Ghosh, Rho, McIntosh, Kötter, & Jirsa, 2008). Nodes
were then connected together as defined by a weighted and directed macaque structural con-
nectivity matrix and the distance between them defined by a tract lengths matrix. As we were
interested in examining signal propagation from the memory system to the oculomotor system
while taking into account the extensive recurrent connectivity between them, we chose to use
the macaque connectome because of the available information from tracer data on the direc-
tionality of fiber tracts. Without stimulation, this network exhibits no activity. Tuttavia, con
stimulation, activity dissipates throughout the network according to the spatiotemporal con-
straints imposed by the connectivity weights and distances. Despite having no spontaneous
activity, this model has been shown to exhibit the emergent properties of spontaneous activity
(Spiegler et al., 2016). Questo è, with stimulation, the model produces a diverse set of resting-state
networks that are typically detected from spontaneous activity in empirical studies. Cortical
network dynamics were set via additional parameter tuning such that stimulating V1 resulted
in biologically plausible timing of evoked responses in downstream visual cortical regions.
Finalmente, we systematically stimulated HC subfields and MTL regions of interest and detected
evoked responses across the rest of the network.
HC Subregion Stimulation
Stimulation of HC subfields and MTL regions of interest evoked widespread activation across
the network, similar to previous surface-based model simulations (Spiegler et al., 2016).
Figura 1 shows an example of activity dissipation following CA1 stimulation. Evoked responses
were first detected in other HC subfields and MTL regions but then spread to prefrontal and
extrastriate cortices, and later to posterior parietal cortex. The full list of activation times for
each of the 77 nodes can be found in Supporting Information Table S2. Tuttavia, in all sub-
sequent analyses, we present only the results pertaining to our nodes of interest, identified as
those along the shortest paths between HC/MTL and oculomotor regions (Shen et al., 2016) O
those that have been specifically suggested in the literature to be potentially relevant (Meister
& Buffalo, 2016).
Stimulation of HC subregions CA1, subiculum (S), presubiculum (PrS), and parasubiculum
(PaS) resulted in observable responses in almost all of the cortical nodes of interest, and within
regions 46, 24, and FEF, of the oculomotor system (for CA1 example; see Figure 2A). Within
our oculomotor regions of interest, activity was first observed in area 46, followed by 24, E
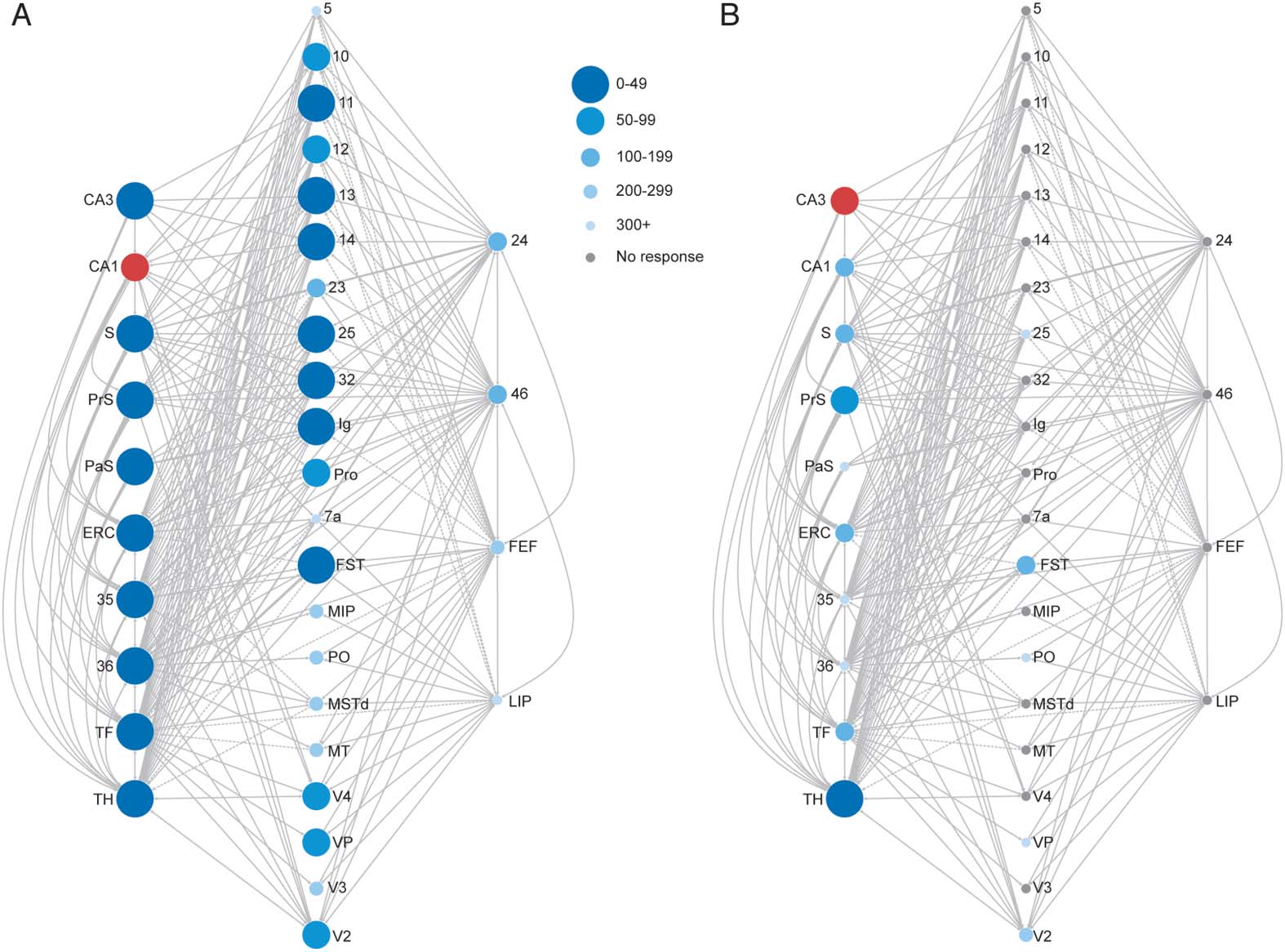
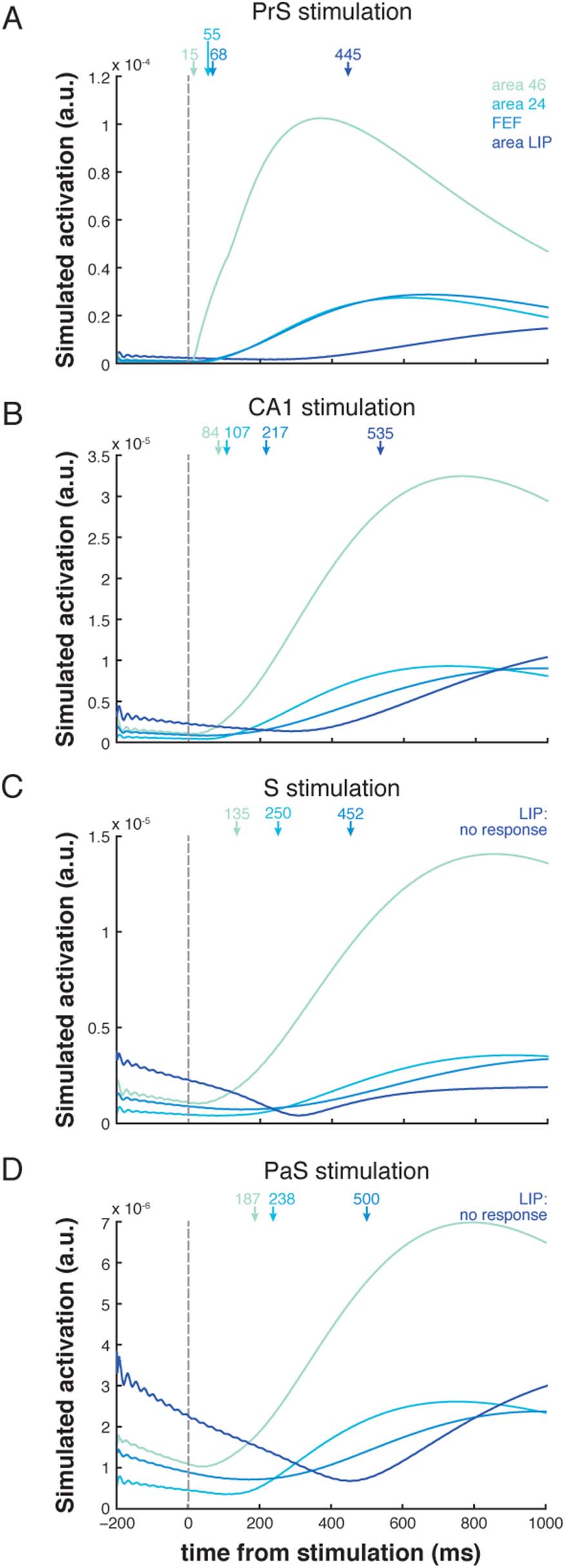
FEF, regardless of HC stimulation site. Stimulation of the PrS resulted in the fastest observable
responses in these oculomotor areas (under 70 ms; Figure 3A). Stimulation of CA1 resulted in
rapid activity that culminated in oculomotor regions in under 220 ms (Figure 3B). Stimulation
of either the S or the PaS resolved into area 46 activity by 200 ms, into area 24 by 250 ms, E
finally into FEF by 500 ms (Figure 3C and 3D). Responses in area LIP occurred substantially
later than the other oculomotor areas and even later than all other cortical nodes for CA1 and
Network Neuroscience
219
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Simulated dynamics of hippocampal responses in oculomotor regions
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Dissipation of activity over time across the cortex following simulated stimulation of
CA1. Average above-threshold simulated activation (arbitrary units) for each node for a 10-ms epoch
following each time point is plotted on the macaque cortical surface. Activations were log scaled
for the purposes of visualization. From left to right: ventral, dorsal, medial, and lateral views of the
macaque cortical surface.
PrS stimulations (>440 ms; Tavolo 1). No evoked response was detected in area LIP following
stimulation of S or PaS. Responses were not observed in the majority of the predefined cortical
hubs following CA3 stimulation, and activity did not culminate in observable responses in the
oculomotor areas (Figure 3B). See Table 1 for activation times for all nodes of interest.
MTL Stimulation
Stimulation of any of the broader regions within the MTL (entorhinal cortex, ERC; perirhinal
cortex, 35, 36; parahippocampal cortex, TF, TH) resulted in observable responses within ocu-
lomotor areas 46, 24, and FEF well under 100 ms, faster than the responses observed from
HC subfield stimulation. Of the MTL regions, stimulation of area 35/36 resulted in the earliest
responses in areas 46, 24, and FEF (within 25 ms). Although evoked responses in area LIP
occurred in under ∼250 ms for all but ERC stimulations, area LIP again exhibited the most
delayed response across all nodes of interest following MTL stimulations. See Table 1 for acti-
vation times for all nodes of interest.
Network Neuroscience
220
Simulated dynamics of hippocampal responses in oculomotor regions
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
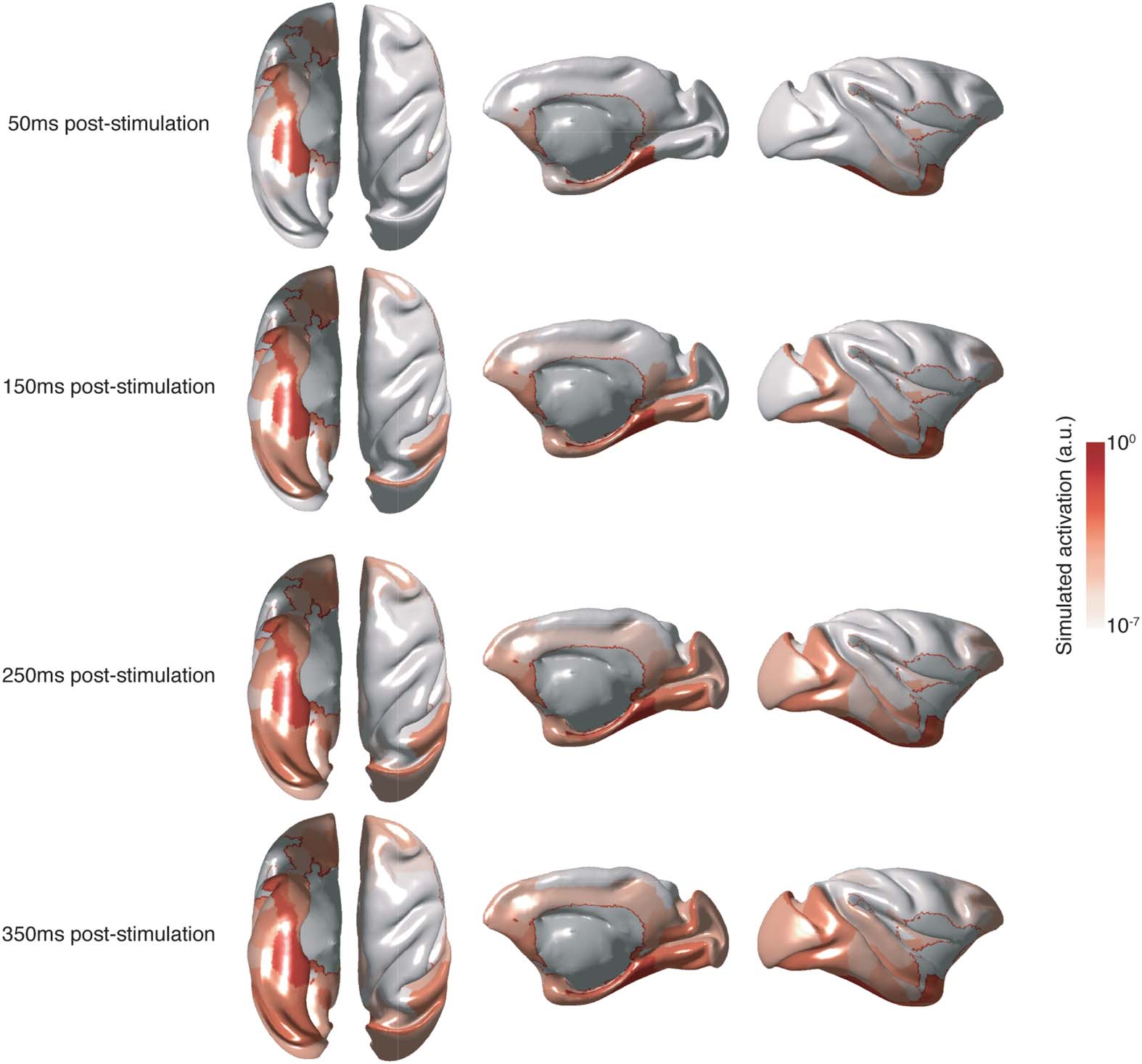
(UN) Simulated stimulation of the CA1 (red circle) resulted in observable responses (blue
Figura 2.
circles) in multiple hippocampus (HC)/medial temporal lobe (MTL) nodes, intermediary nodes, E
in regions governing oculomotor control, including the frontal eye fields (FEFs). (B) Simulated stim-
ulation of the CA3 (red circle) resulted in observable responses (blue circles) limited to HC/MTL
nodes. Very few responses were observed in cortical areas and none were observed in oculomotor
areas. Size and shade of the circles scale with elapsed time prior to an observed response. Gray
lines denote direct structural connections between nodes. For visualization purposes, only regions
that contribute to the shortest paths from HC/MTL to oculomotor nodes are shown. Connections
between intermediary nodes (middle column) are not shown. Connections that are unidirectional
and away from oculomotor areas (cioè., to HC/MTL) are indicated by dashed lines.
Cortical Responses
HC and MTL region stimulation (except for CA3) resulted in signal propagation across all of
our preidentified cortical regions of interest. When CA3 was stimulated, cortical responses
were only observed in areas V2 and 25; no other signal was observed. Notably, responses in
areas 5 and 7a were generally observed following activity from oculomotor regions, includ-
ing FEF, suggestive of a possible feedback response. The exception is S stimulation, in which
responses in area 5 preceded responses in oculomotor regions by ∼100 ms. Responses in V4
also followed oculomotor responses, except in cases of CA1, TF, and TH stimulation. Sim-
ilarly, responses in area 23 followed oculomotor responses, except in cases of PrS and TH
stimulation. See Table 1 for activation times for all nodes of interest.
Lesion Models: HC Subregions
Some models of HC and MTL lesions showed an appreciable effect on activation times while
others did not. Only the results for lesions that affected any activation time by at least ±10 ms
are shown. Lesion of CA3 changed neither the pattern nor the timing of observable responses
following stimulation of each of the other HC/MTL regions (data not shown). Lesion of CA1
Network Neuroscience
221
Simulated dynamics of hippocampal responses in oculomotor regions
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Figura 3. Simulated response profiles (envelope of region time series) of oculomotor areas fol-
lowing stimulation of PrS (UN), CA1 (B), S (C), and PaS (D). Activation is given in arbitrary units (a.u.).
The onsets of the responses for each oculomotor area indicated by arrows. Area LIP did not exhibit
a response that exceeded its baseline threshold following S and PaS stimulation.
Network Neuroscience
222
Simulated dynamics of hippocampal responses in oculomotor regions
Tavolo 1. Simulated activation times (ms) following stimulation of hippocampal subfields and medial
temporal lobe regions.
Stimulated node
e
D
o
N
N
o
io
T
UN
v
R
e
S
B
O
CA3
CA1
S
PrS
PaS
ERC
35
36
TF
TH
5
10
11
12
13
14
23
25
32
Ig
Pro
7UN
FST
MIP
PO
MSTd
MT
V4
VP
V3
V2
24
46
FEF
LIP
CA3
0
137
115
84
350
169
567
317
137
7
N/A
N/A
N/A
N/A
N/A
N/A
N/A
435
N/A
N/A
N/A
N/A
186
N/A
656
N/A
N/A
N/A
753
N/A
215
N/A
N/A
N/A
N/A
CA1
48
0
17
23
17
40
6
1
0
0
322
59
12
96
10
9
165
6
38
11
52
381
47
237
208
286
260
76
55
260
56
107
84
217
535
S
31
0
0
119
96
0
21
14
20
29
50
24
68
186
18
27
384
19
63
63
96
202
147
523
593
664
857
353
266
841
274
250
135
452
N/A
PrS
37
0
26
0
28
4
49
10
0
0
92
83
74
93
62
57
12
35
88
75
12
336
52
215
190
287
272
103
86
257
48
55
15
68
445
PaS
27
46
37
25
0
0
50
6
44
0
555
106
166
285
132
76
231
57
89
189
48
795
79
296
248
494
483
244
259
409
66
238
187
500
N/A
ERC
0
0
0
6
50
0
0
0
11
20
249
12
10
15
8
9
43
6
10
11
3
256
6
405
355
33
189
280
223
398
139
24
12
79
561
35
21
8
16
13
15
0
0
0
12
29
129
10
7
9
7
9
37
11
13
4
1
53
1
113
99
23
107
147
98
154
16
22
9
19
233
36
20
2
10
11
0
0
0
0
6
18
141
12
8
10
11
11
52
8
14
6
4
52
3
126
107
23
23
142
101
163
17
25
11
15
230
TF
48
0
0
0
0
7
9
6
0
0
69
81
38
24
19
13
24
9
14
21
29
103
0
70
60
94
36
5
2
68
6
22
17
34
176
TH
1
0
0
0
32
13
19
14
0
0
64
20
48
34
19
20
13
12
17
13
20
59
6
48
39
53
83
15
41
68
0
16
23
71
144
Note. Only nodes of interest (HC/MTL regions, oculomotor regions, and regions that are involved
in the shortest paths between HC/MTL and oculomotor nodes) are shown. S= subiculum; PrS =
presubiculum; PaS = parasubiculum; ERC = entorhinal cortex; 35/36 = perirhinal cortex; TF/TH =
parahippocampal cortex; 0 = stimulation onset; N/A = no response observed. For a comprehensive
set of activation times for all nodes in the network, see Supporting Information Table S2.
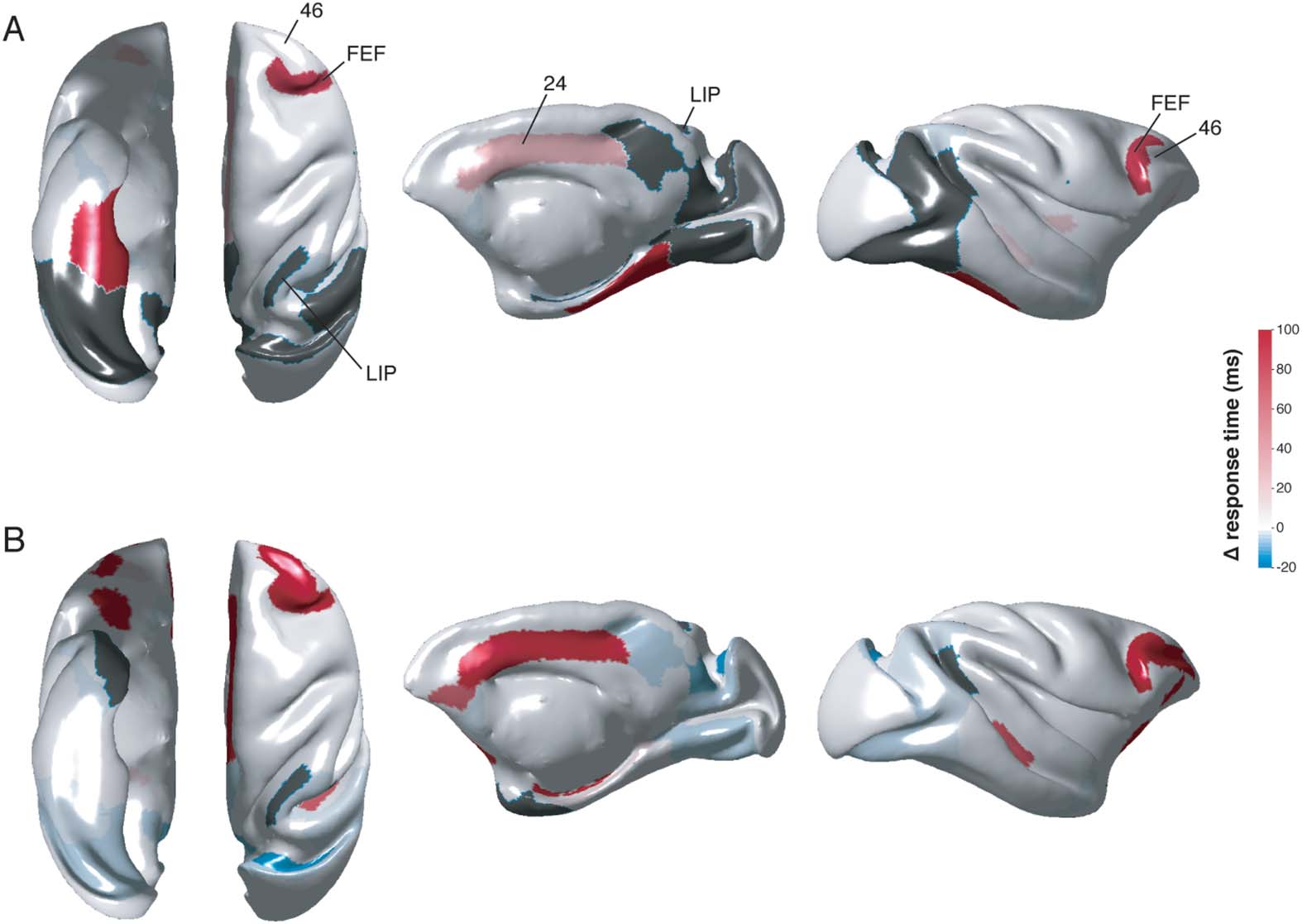
resulted in a lack of signal to V2, V4, area 23, and slowing of signal from the subicular com-
plex to various regions, including oculomotor regions FEF and area 24 (Figure 4A; Supporting
Information Table S3). Lesion of CA1 also led to small increases in the speed of signal follow-
ing CA3 stimulation to the subicular complex, and from MTL regions to TF/TH, and to other
regions within the subicular complex (all less than 10 ms).
Network Neuroscience
223
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Simulated dynamics of hippocampal responses in oculomotor regions
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Figura 4. Changes in simulated activation times following HC lesions. Subicular stimulation fol-
lowing CA1 (UN) and ERC (B) lesions. Only nodes of interest are presented on the brain surface plots.
Activation time differences were computed by subtracting the prelesion activation times from the
postlesion ones. Absence of response following a lesion indicated in gray. From left to right: ventral,
dorsal, medial, and lateral views of the macaque cortical surface.
Lesions to either the S or PaS produced little change to either the pattern or timing of re-
sponses following stimulation of the other HC/MTL regions (data not shown). Lesions to the
PrS produced moderate changes (<20 ms) in timing: there was some slowing of activity prop-
agation from PaS to cortical regions, including oculomotor and speeding
of signal propagation within the HC subfields TF>140 ms)
(Supporting Information Table S5).
Lesion Models: MTL Regions
Lesion of the ERC resulted in considerable slowing of observable signal in areas 24, 46, E
FEF (30–340 ms) following S (Figure 3B) or PaS stimulation (Supporting Information Table S6).
TF and/or TH lesions resulted in slowing (10–400 ms) of signal following CA1, S, and PaS
stimulation to one or more of areas 24, 46, and FEF, and a lack of response in FEF following
PaS stimulation (only the combined TF/TH is shown; Supporting Information Table S7). Area
35 and/or 36 lesions also resulted in slowing (10–90 ms) of signal following CA1, S, and PaS
stimulation to one or more of areas 24, 46, and FEF, although not as severe as the slowing
Network Neuroscience
224
Simulated dynamics of hippocampal responses in oculomotor regions
observed following TF/TH lesions (only the combined 35/36 lesion is shown; Supporting
Information Table S8).
Other Cortical Lesions
In our original stimulations, signals in regions 5, 7UN, 23, and V4 were predominantly observed
following observable responses in oculomotor areas 24, 46, and FEF, suggesting these cortical
areas are receiving feedback signals rather than primarily serving as hubs to transfer signal
from the HC/MTL to the oculomotor regions. To explore this in more depth, we simulated a
combined lesion of 5/7a/23/V4 and examined signal propagation. Following this combined
cortical lesion, stimulation of each of the HC/MTL regions (except for CA3) continued to re-
sult in observable signal in areas 24, 46, and FEF. È interessante notare, while area LIP exhibited the
slowest overall responses in the intact simulations, the combined cortical lesion led to sub-
stantial speeding of signal to LIP from HC (200–270 ms faster) and MTL (80–300 ms faster) Rif-
gions owing to speeded responses in intermediary visual cortical and parietal areas (Supporting
Information Table S9).
DISCUSSION
A preponderance of evidence has demonstrated a correlation between HC/MTL neural activity
and oculomotor behavior (Hannula et al., 2010; Killian, Potter, & Buffalo, 2015; Liu et al.,
2017), but research had not shown whether HC/MTL activity can reach the oculomotor system
in time to influence the preparation of a saccade. The HC is well connected anatomically to
the oculomotor system through a set of polysynaptic pathways that span MTL, frontal, parietal,
and visual cortices (Shen et al., 2016), but the existence of anatomical connections does not
provide conclusive evidence of the functional relevance of specific pathways. By considering
the functional dynamics and recurrent interactions of the large-scale network involved in the
HC/MTL guidance of eye movements, we show that propagation of evoked HC/MTL neural
activity results in neural activity observable in areas 24 (ACC), 46 (dlPFC), and FEF, Quale
are important for the cognitive and motoric control of eye movements, rispettivamente (Johnston
& Everling, 2008). Critically, the culmination of neural signal in these oculomotor regions
occurred within the time of a typical gaze fixation (∼250–400 ms; Buswell, 1935; Henderson
et al., 2013): within 200 ms following HC subfield stimulation (except for CA3), and within
100 ms following stimulation of each MTL region. Our findings suggest that the underlying
neural dynamics of the memory and oculomotor systems allow for representations mediated
by the HC/MTL to guide visual exploration—what is foveated and when—on a moment-to-
moment basis.
The lack of responses in the FEF following CA3 stimulation is not surprising, given that there
are no known direct connections, and fewer polysynaptic pathways, between the CA3 and the
oculomotor regions investigated here (Shen et al., 2016). These functional and anatomical dif-
ferences align well with the purported representational functions of CA3 versus CA1. Foveated
information may be bound into detailed memory representations via the autoassociative net-
work of the CA3 (pattern separation; Norman & O’Reilly, 2003; Yassa & Stark, 2011), whereas
CA1 would enable the comparison of stored information to the external visual world ( pattern
completion; Rolls, 2013; Yassa & Stark, 2011).
Stimulation of the subiculum and parasubiculum resulted in relatively slower responses
observed in each of the oculomotor regions, whereas stimulation of presubiculum resulted in
rapid responses observed in the oculomotor regions. The subiculum and parasubiculum may
largely provide information that supports the grid cell mapping of the ERC (Boccara et al.,
Network Neuroscience
225
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Simulated dynamics of hippocampal responses in oculomotor regions
2010; Peyrache, Schieferstein, & Buzsáki, 2017; Tang et al., 2016). These regions may then
function as a “pointer” by providing online information of an individual’s location in space
(Tang et al., 2016). This slowly changing spatial layout may not then require a rapid influence
on the oculomotor system, but instead, may allow for the presubiculum, which has cells that are
responsive to head direction (Robertson, Rolls, Georges-François, & Panzeri, 1999) to precisely
locate and foveate visual objects. These functional distinctions are speculative and remain to
be tested.
Stimulation of each of the MTL cortices resulted in observable responses in each of areas
24, 46, and FEF that were faster than any of the responses observed following HC subregion
stimulation. The MTL cortices are intermediary nodes that may permit the relatively rapid trans-
fer of information from HC to the oculomotor system. The unique representational content
supported by each region may influence ongoing visual exploration in a top-down manner.
The PRC provides lasting information regarding the features of objects (Erez, Cusack, Kendall,
& Barense, 2016; Graham, Barense, & Lee, 2010), the PHC provides information regarding
the broader spatial environment (Alvarado & Bachevalier, 2005; Eichenbaum, Yonelinas, &
Ranganath, 2007; Sato & Nakamura, 2006), and the ERC may provide information regarding
the relative spatial arrangements of features within (Yeung et al., 2017), and among, objects
within the environment (Buckmaster, 2004; Yeung et al., 2019). Signal from the MTL may be
used to accurately, and rapidly, prioritize gaze fixations to areas of interest.
HC subfield lesions only minimally altered the timing of activity from MTL to oculomotor
regions; the relatively rapid propagation of signal from MTL to FEF (<100 ms) was preserved.
Lesions to MTL regions resulted in slowing of signal from some HC subfields oculomotor
regions. This pattern results suggests that different patterns visual exploration (i.e., rate,
area) may occur cases>3.0.CO;2-H
Rolls, E. T. (2013). The mechanisms for pattern completion and pat-
tern separation in the hippocampus. Frontiers in Systems Neuro-
science. https://doi.org/10.3389/fnsys.2013.00074
Ryals, UN. J., Wang, J. X., Polnaszek, K. L., & Voss, J. l. (2015).
Hippocampal contribution to implicit configuration memory ex-
pressed via eye movements during scene exploration. Hippo-
campus, 25(9), 1028–1041. https://doi.org/10.1002/hipo.22425
Ryan, J. D., & Cohen, N. J. (2004). The nature of change detec-
tion and online representations of scenes. Journal of Experimental
Network Neuroscience
232
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Simulated dynamics of hippocampal responses in oculomotor regions
Psychology: Human Perception and Performance, 30(5), 988–1015.
https://doi.org/15462635
Ryan, J. D., Althoff, R. R., Whitlow, S., & Cohen, N. J. (2000). Amne-
sia is a deficit in relational memory. Psychological Science, 11(6),
454–461. https://doi.org/10.1111/1467-9280.00288
Ryan, J. D., & Cohen, N. J. (2003). Evaluating the neuropsycho-
logical dissociation evidence for multiple memory systems. Cog-
nitive, Affective and Behavioral Neuroscience, 3(3), 168–185.
https://doi.org/10.3758/CABN.3.3.168
Ryan, J. D., Leung, G., Turk-Browne, N. B., & Hasher, l. (2007).
Assessment of age-related changes in inhibition and binding us-
ing eye movement monitoring. Psychology and Aging, 22(2),
239–250. https://doi.org/10.1037/0882-7974.22.2.239
Sato, N., & Nakamura, K. (2006). Visual response properties of neu-
rons in the parahippocampal cortex of monkeys. https://doi.org/
10.1152/jn.01089.2002
Schmolesky, M. T., Wang, Y., Hanes, D. P., Thompson, K. G.,
Leutgeb, S., Schall, J. D., & Leventhal, UN. G. (2017). Signal timing
across the macaque visual system. Journal of Neurophysiology.
https://doi.org/10.1152/jn.1998.79.6.3272
Seelke, UN. M. H., Padberg,
J., Disbrow, E., Purnell, S. M.,
Recanzone, G., & Krubitzer, l. (2012). Topographic maps within
Brodmann’s area 5 of macaque monkeys. Cerebral Cortex.
https://doi.org/10.1093/cercor/bhr257
J.
Shen, K., Bezgin, G., Schirner, M., Ritter, P., Everling, S., &
McIntosh, UN. R. (2019). A macaque connectome for large-scale
network stimulations in TheVirtualBrain. Scientific Data, 6(1),
123. https://doi.org/10.1038/s41597-019-0129-z
Shen, K., Bezgin, G., Selvam, R., McIntosh, UN. R., & Ryan, J. D. (2016).
An anatomical interface between memory and oculomotor sys-
tems. Journal of Cognitive Neuroscience, 28(11), 1772–1783.
https://doi.org/10.1162/jocn_a_01007
Shen, K., Goulas, A., Grayson, D. S., Eusebio, J., Gati, J. S., Menon,
R. S., . . . Everling, S. (2019). Exploring the limits of network topol-
ogy estimation using diffusion-based tractography and tracer
studies in the macaque cortex. NeuroImage, 191, 81–92. https://
doi.org/10.1016/j.neuroimage.2019.02.018
Smith, C. N., Hopkins, R. O., & Squire, l. R. (2006). Experience-
dependent eye movements, awareness, and hippocampus-
dependent memory. The Journal of Neuroscience: The Official
Journal of the Society for Neuroscience, 26(44), 11304–11312.
https://doi.org/10.1523/JNEUROSCI.3071-06.2006
Smith, C. N., & Squire, l. R. (2017). When eye movements express
memory for old and new scenes in the absence of awareness and
independent of hippocampus. Learning and Memory. https://doi.
org/10.1101/lm.043851.116
Sobotka, S., Nowicka, A., & Ringo, J. l. (1997). Activity linked to ex-
ternally cued saccades in single units recorded from hippocam-
pal, parahippocampal, and inferotemporal areas of macaques.
J Neurophysiol, 78(4), 2156–2163.
Spiegler, A., Hansen, E. C. A., Bernard, C., McIntosh, UN. R., & Jirsa,
V. K. (2016). Selective activation of resting-state networks follow-
ing focal stimulation in a connectome-based network model of
the human brain. Eneuro, 3(5).
Stephan, K. E., Kamper, L., Bozkurt, A., Burns, G., Young, M., &
Kötter, R. (2001). Advanced database methodology for the Col-
lation of Connectivity data on the Macaque brain (CoCoMac).
Philosophical Transactions of the Royal Society of London. Series
B, Biological Sciences, 356, 1159–1186. https://doi.org/10.1098/
rstb.2001.0908
Tang, Q., Burgalossi, A., Ebbesen, C. L., Sanguinetti-Scheck, J. I.,
Schmidt, H., Tukker, J. J., . . . Brecht, M. (2016). Functional archi-
tecture of the rat parasubiculum. Journal of Neuroscience. https://
doi.org/10.1523/JNEUROSCI.3749-15.2016
Vincent, J. L., Kahn, I., Van Essen, D. C., & Buckner, R. l. (2010).
Functional connectivity of the macaque posterior parahippocam-
pal cortex. Journal of Neurophysiology. https://doi.org/10.1152/
jn.00546.2009
Vlachos, I., Aertsen, A., & Kumar, UN. (2012). Beyond statistical sig-
nificance: Implications of network structure on neuronal activ-
ità. PLoS Computational Biology, 8(1). https://doi.org/10.1371/
journal.pcbi.1002311
Warren, D. E., Duff, M. C., Tranel, D., & Cohen, N. J. (2010).
Medial temporal lobe damage impairs representation of simple
stimuli. Frontiers in Human Neuroscience, 4, 35. https://doi.org/
10.3389/fnhum.2010.00035
Wynn, J. S., Ryan, J. D., & Moscovitch, M. (2019). Effects of prior
knowledge on active vision and memory in younger and older
adults. Journal of Experimental Psychology: General. https://doi.
org/10.1037/xge0000657
Yassa, M. A., & Stark, C. E. l. (2011). Pattern separation in the
hippocampus. Trends in Neurosciences. https://doi.org/10.1016/
j.tins.2011.06.006
Yeung, L.-K., Olsen, R. K., Bild-Enkin, H. E. P., D’Angelo, M. C.,
Kacollja, A., McQuiggan, D. A., . . . Barense, M. D. (2017). An-
terolateral entorhinal cortex volume predicted by altered intra-
item configural processing. The Journal of Neuroscience. https://
doi.org/10.1523/JNEUROSCI.3664-16.2017
Yeung, L.-K., Olsen, R. K., Hong, B., Mihajlovic, V., D’Angelo,
M. C., Kacollja, A., . . . Barense, M. D. (2019). Object-in-
place memory predicted by anterolateral entorhinal cortex and
parahippocampal cortex volume in older adults. Journal of Cog-
nitive Neuroscience, 31(5), 711–729. https://doi.org/10.1162/
jocn_a_01385
Network Neuroscience
233
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
2
1
7
1
8
6
6
7
5
4
N
e
N
_
UN
_
0
0
1
2
0
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3