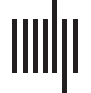Research
Core language brain network for fMRI language
task used in clinical applications
Qiongge Li
1,2,∗
, Gino Del Ferraro1,3,∗
, Luca Pasquini3,4, Kyung K. Peck3,5,
Hernán A. Makse1, and Andrei I. Holodny3,6,7
1Levich Institute and Physics Department, City College of New York, New York, USA
2Physics Department, The Graduate Center of City University of New York, New York, USA
3Department of Radiology, Memorial Sloan Kettering Cancer Center, New York, USA
4Neuroradiology Unit, NESMOS Department Sant’Andrea Hospital La Sapienza University, Rome, Italy
5Department of Medical Physics, Memorial Sloan Kettering Cancer Center, New York, USA
6New York University School of Medicine, New York, USA
7Neuroscience, Weill Medical College of Cornell University, New York, USA
* Equal contributions as first authors.
Keywords: Task-based fMRI, Functional
Presurgical langugage mapping, k-core
language networks, Healthy controls, Graph theory,
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
ABSTRACT
Functional magnetic resonance imaging (fMRI) is widely used in clinical applications to
highlight brain areas involved in specific cognitive processes. Brain impairments, ad esempio
tumors, suppress the fMRI activation of the anatomical areas they invade and, così,
brain-damaged functional networks present missing links/areas of activation. IL
identification of the missing circuitry components is of crucial importance to estimate the
damage extent. The study of functional networks associated with clinical tasks but performed
by healthy individuals becomes, Perciò, of paramount concern. These “healthy” networks
can, Infatti, be used as control networks for clinical studies. In this work we investigate the
functional architecture of 20 healthy individuals performing a language task designed for
clinical purposes. We unveil a common architecture persistent across all subjects under
study, that we call “core” network, which involves Broca’s area, Wernicke’s area, IL
premotor area, and the pre-supplementary motor area. We study the connectivity of this
circuitry by using the k-core centrality measure, and we find that three of these areas belong
to the most robust structure of the functional language network for the specific task under
study. Our results provide useful insights on primarily important functional connections.
AUTHOR SUMMARY
Neurosurgeons employ language fMRI to localize important language areas for patients
with brain impairment. Yet, brain pathologies (per esempio., brain tumors, strokes, epilepsy) affect
functional connectivity by disrupting functional links and suppressing the activation of
brain areas. Così, although clinical tasks are designed to guarantee robust activation, IL
functional connectivity of patients with brain pathologies is ultimately damaged by brain
impairments. To better quantify the damage produced by the brain pathology on the
functional connectivity, it is paramount to have, as a benchmark, functional networks of
healthy individuals who perform a task for clinical cases. Our findings identify a group of
functional regions of interest linked together in a functional circuitry that have a decisive role
for the language task used in clinical applications.
a n o p e n a c c e s s
j o u r n a l
Citation: Li, Q., Del Ferraro, G.,
Pasquini, L., Peck, K. K., Makse, H. A.,
& Holodny, UN. IO. (2020). Core language
brain network for fMRI language task
used in clinical applications. Network
Neuroscience, 4(1), 134–154. https://
doi.org/10.1162/netn_a_00112
DOI:
https://doi.org/10.1162/netn_a_00112
Supporting Information:
https://doi.org/10.1162/netn_a_00112
http://www-levich.engr.ccny.cuny.edu/
webpage/hmakse/brain/
Received: 12 Giugno 2019
Accepted: 28 ottobre 2019
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Andrei I. Holodny
holodnya@mskcc.org
Handling Editor:
Alex Arenas
Copyright: © 2019
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
Core language brain network
Aphasia:
Language impairment affecting the
production or comprehension of
speech and the ability to read or
write. Aphasia is caused by brain
injuries such as strokes, head
traumas, brain tumors, or brain
infections.
INTRODUCTION
Broca’s area (BA) and Wernicke’s area (WA) have long been recognized as essential language
centers. Studies of aphasic patients have shown that damage to BA and WA causes loss of
ability to produce speech (expressive aphasia) and difficulty understanding language (recep-
tive aphasia), rispettivamente (Dronkers, Plaisant, Iba-Zizen, & Cabanis, 2007; Wernicke, 1970).
Further evidence has shown that other secondary and tertiary anatomical brain areas are also
involved in language Friederici (2011), including the pre-supplementary motor area (pre-SMA;
Hertrich, Dietrich, & Ackermann, 2016), the premotor area (preMA; Duffau et al., 2003), E
the basal ganglia (Booth, Legna, Lu, Houk, & Bitan, 2007). Despite this evidence, a full char-
acterization of the language network is still debated (Fedorenko & Kanwisher, 2009; Friederici,
Chomsky, Berwick, Moro, & Bolhuis, 2017).
Functional MRI (fMRI) has been largely used to investigate the blood-oxygen-level depen-
dent (BOLD) activation of the human brain, for both clinical and research purposes. Although
it cannot fully resolve the issue of “functional specialization” of brain regions by itself, it sheds
light on which regions are engaged in certain cognitive processes. Therefore fMRI allows us to
constrain hypotheses on the structure of the language network.
Language task fMRI:
Functional MRI scan where
participants perform specific
language tasks while the fMRI brain
activity is measured. It is used as a
radiology clinical routine to
determine the localization of
language areas in patients with brain
impairments.
Language has been investigated using both resting-state fMRI (rs-fMRI) and tasked-based
fMRI (tb-fMRI). The former studies brain activation of subjects at rest (Lee, Smyser, & Shimony,
2013), whereas tb-fMRI delineates brain areas functionally involved in the performance of a
specific task (Bookheimer, 2002). Task-based fMRI is task-dependent, questo è, different language
tasks may activate different areas involved in language function (Xiong et al., 2000). Conse-
quently, clinical studies employ a specific class of language tasks that have been shown to
produce robust activation in individual participants and thus facilitate the localization of the
language-sensitive cortex (Brennan et al., 2007; Ramsey, Sommer, Rutten, & Kahn, 2001).
In this paper we analyze fMRI scans of 20 healthy individuals who perform the same lan-
guage task designed for clinical purposes. From the correlation of the BOLD signal we construct
the functional connectivity network for each subject, which is standardly employed to inves-
tigate statistical interdependencies among brain regions (Bullmore & Sporns, 2009; Gallos,
Makse, & Sigman, 2012; Hermundstad et al., 2013). We then employ graph theory to study
the networks’ properties as successfully done in Del Ferraro et al. (2018) to investigate mem-
ory formation. The motivation for this study is to use the resulting functional connectivity of
these healthy individuals as a benchmark for clinical study, as we explain next. We employ
a language clinical task because we are interested in studying the fMRI activation associated
with this specific type of task. We employ healthy subjects and not patients with brain impair-
ments because we want to study the fMRI activation without any interference that might arise
because of the brain impairment. Brain pathologies (per esempio., brain tumors, Wang et al., 2013;
strokes, Tombari et al., 2004; epilepsy, Rosenberger et al., 2009) indeed affect functional con-
nectivity by disrupting functional links and reducing the fMRI activation of brain areas (per esempio.,
the neurovascular decoupling effect due to brain tumor; Aubert, Costalat, Duffau, & Benali,
2002). The reconstruction of the functional connectivity in clinical cases, Perciò, is influ-
enced by the presence of brain pathology (Wang et al., 2013). In other words, the functional
connectivity of a patient with a pathology such as a brain tumor, for instance, presents miss-
ing links and missing fMRI active areas compared with the healthy case, for the same specific
task. To better understand what functional damage was produced by the brain impairment, Esso
is important to have, as a benchmark, functional networks of healthy individuals performing
the same language task normally used for clinical cases. In this way, using clinical language
tasks performed by healthy subjects, we can study functional networks associated with clinical
Network Neuroscience
135
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
fROI:
Functional region of interest. Group
of fMRI active voxels located in the
same anatomical brain region that
are identified as part of the same
active brain area.
tasks without perturbations that arise from brain impairments. These functional networks can
be used as benchmarks for other studies that use the same type of task but employ patients
with brain damage. The comparison between a healthy control and a patient’s functional net-
work relative to the same task could in principle establish what is the damage produced by the
brain impairment on the functional network and might, among others, guide tumor resection
to preserve functional links.
Motivated by these considerations, we investigated which is the language functional archi-
tecture shared among healthy subjects, questo è, the functional subnetwork that persists in each
analyzed individual beyond the intersubject variability. This architecture is indicative of a core
structure for the language task under study shared across individuals. Core architectures have
been identified in other contexts (Bassett et al., 2013), but very little is known about the core
for language tasks (Chai, Mattar, Blank, Fedorenko, & Bassett, 2016) and its investigation is
one of the main goals of our study.
Inoltre, we aim to uncover the functional connectivity of the subdivisions of the Broca’s
area (pars-opercularis, op-BA, and pars-triangularis, tri-BA, cioè., Brodmann area 44 E 45 Rif-
spectively), which plays a pivotal role in language function (Dronkers et al., 2007; Friederici,
2011). Previous studies based on fMRI showed that BA’s subdivisions perform different func-
tions in language processing. Newman, Just, Keller, Roth, and Carpenter (2003) showed that
tri-BA is more implicated in thematic processing whereas op-BA is more involved in syntac-
tic processing. Studies based on transcranial magnetic stimulation have shown that op-BA is
more specialized in phonological tasks and tri-BA more in semantic tasks (Devlin, Matthews,
& Rushworth, 2003; Gough, Nobre, & Devlin, 2005; Nixon, Lazarova, Hodinott-Hill, Gough,
& Passingham, 2004). Patients who show speech impairment often have direct damage to the
Broca’s area. Così, understanding how BA subdivisions are functionally wired to other brain
regions in healthy controls may be used for comparison in some clinical cases and could poten-
tially be of help to better clarify the effect of brain pathologies on this decisive language area.
From our analysis we find that the functional architecture shared by most of the subjects un-
der study wires together Broca’s area (op-BA and tri-BA), Wernicke’s area, the pre-supplementary
motor area, and the premotor area. By investigating network properties at the subject level
we find that, in each individual functional network, these areas belong to an innermost core,
more specifically the maximum k-core of the functional connectivity, which is a robust and
highly connected substructure of the functional architecture. The k-core measure has received
vast attention in network analysis since it provides a topological notion of the structural skele-
ton of a circuitry (Dorogovtsev, Goltsev, & Mendes, 2006; Kitsak et al., 2010; Pittel, Spencer,
& Wormald, 1996; Rubinov & Sporns, 2010). More recently, the maximum k-core has been
related to the stability of complex biological systems (Morone, Del Ferraro, & Makse, 2019)
and of resilient functional structures in the brain (Lucini, Del Ferraro, Sigman, & Makse, 2019).
Our results demonstrate that the functional architecture that persists beyond intersubject vari-
ability is part of the maximum k-core structure, an innermost highly connected subnetwork,
associated with a system’s resilience and stability (Morone et al., 2019).
Overall, our findings identify a group of functional regions of interest (fROIs) linked to-
gether in a functional circuitry that play a decisive role for the language task used in clinical
applications.
MATERIALS AND METHODS
The study was approved by the Institutional Review Board and an informed consent was ob-
tained from each subject. The study was carried out according to the declaration of Helsinki.
Network Neuroscience
136
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
Twenty healthy right-handed adult subjects (13 males and 7 females; age range 36 years,
mean = 36.6; SD = 11.56) without any neurological history were included.
Functional Task
For the fMRI task, all subjects performed a verbal fluency task using verb generation in response
to auditory nouns. During the verb generation task, subjects were presented with a noun (for
esempio, baby) by oral instruction and then asked to generate action words (Per esempio,
cry, crawl) associated with the noun. Four nouns were displayed over six stimulation epochs,
with each epoch lasting 20 S, which allowed for a total of 24 distinct nouns to be read over
the entire duration. Each epoch consisted of a resting period and a task period (see BOLD
activation in Figures 1A and 1B). In order to avoid artifacts from jaw movements, subjects
were asked to silently generate the words. Brain activity and head motion were monitored
using Brainwave software (GE, Brainwave RT, Medical Numerics, Germantown, MD), allowing
real-time observation.
Data Acquisition
A GE 3T scanner (General Electric, Milwaukee, Wisconsin, USA) and a standard quadrature
head coil was employed to acquire the MR images. Functional images covering the whole
brain were acquired using a T2*-weighted gradient echo planar imaging sequence (repeti-
tion time, TR/echo time, TE = 4, 000/40 ms; slice thickness = 4.5 mm; matrix = 128 × 128;
FOV = 240 mm). Functional matching axial T1-weighted images (TR/TE = 600/8 ms; slice
thickness = 4.5 mm) were acquired for anatomical coregistration purposes. Additionally,
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
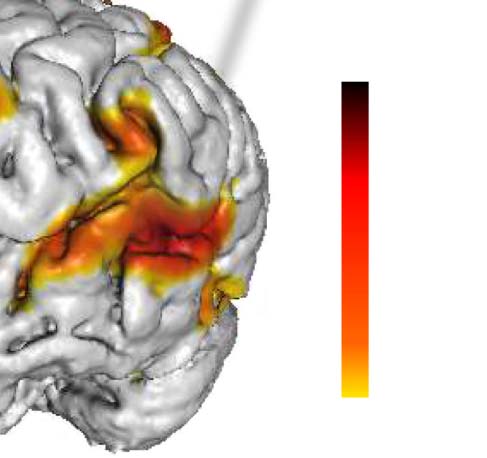
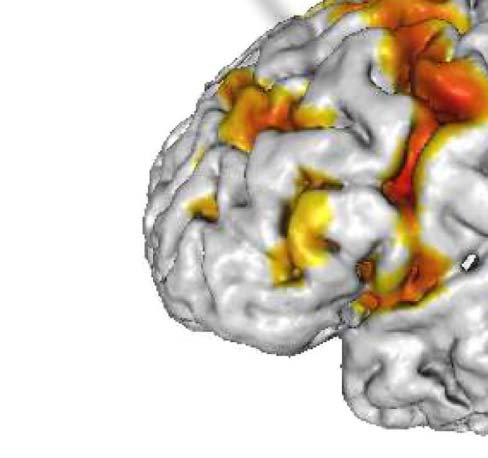
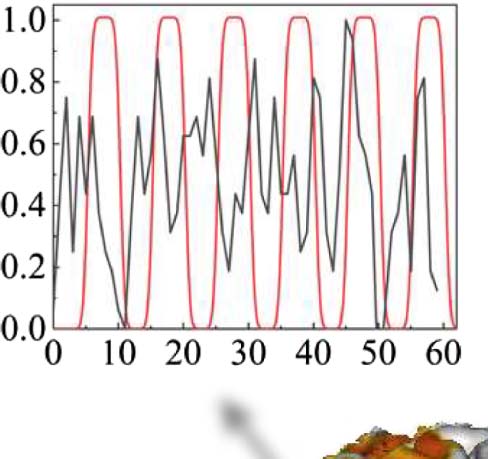
Figura 1. Activation map for a representative subject. BOLD signal for a nonactive and active
voxel are shown respectively in panels A and B together with the smoothed boxcar language model,
which depicts the auditory stimulus. The black curve represents BOLD signal, while the red curve
represents the smoothed boxcar task model design. The red curve’s peaks represent the continuous
stimuli (verb task), presented for a certain period of time (10 S), the “task” or “on” period. The red
curve’s troughs represent the rest period (no task) for the participant, which lasted 10 S, (C) 3D
visualization of the brain with fMRI active areas and corresponding p values.
Network Neuroscience
137
Core language brain network
3D T1-weighted SPGR (spoiled gradient recalled) sequences (TR/TE = 22/4 ms; slice thick-
ness = 1.5 mm; matrix = 256 × 256) covering the entire brain were acquired.
Data Processing
Functional MRI data were processed and analyzed using the software program Analysis of
Functional NeuroImages (AFNI; Cox, 1996). Head motion correction was performed using 3D
rigid-body registration. The first volume was selected to register all other volumes. The first
volume was chosen because it was acquired right next to the anatomical scan. During the
registration, the motion profile was saved and during the statistical analysis any voxels highly
correlated with the profile were regressed out. Spatial smoothing was applied to improve the
signal-to-noise ratio using a Gaussian filter with 4 mm full width of half maximum. Corrections
for linear trend and high-frequency noise were also applied. To obtain the activation map, IL
rectangular train representing the single-task (verb generation) block design is convolved with a
canonical hemodynamic response function (HRF; see the red curve in Figures 1A and 1B). Questo
time series is one regressor in the general linear model where the preprocessed BOLD response
is to fit to this as well as a baseline (intercept). The BOLD signal is represented by the black
curve in Figures 1A and 1B). The test statistics for the activation maps are determined by cross-
correlation analysis within AFNI software. They were generated in the individual native space
at a minimum threshold of p < 0.0001 to identify activated voxels set by a neuroradiologist
(see Figure 1C, for a representative subject).
The individual voxel threshold level (uncorrected p < 0.0001) was set to minimize the con-
tribution of false positives that could be caused by stimulus-correlated head motion and/or
random noise fluctuation. The level was set for all subjects. We looked at FDR (q value cal-
culated in AFNI software) to control false positives among the detected voxels, and the FDR
corrected is q < 0.001. We verified that the activated areas identified and considered in our
graph theory analysis are significant.
Network Construction
The following sections describe the functional network construction. In the first subsection, we
first describe how to create, from the fMRI signal of the active voxels, a brain network for each
individual separately. The second subsection discusses the group analysis or how we obtain,
from the individual brain networks, a common architecture that unveils a persistent circuitry
across all the single-subject brain networks.
For each subject we construct a functional network. This
Individual brain network construction.
network can be seen at two different scales or levels: (a) at the voxel level and (b) at the fROI
level, as we explain in more detail below.
At the voxel level, active voxels in the individual activation map (p < 0.0001) define the
nodes of our functional network, where a voxel is the lowest resolution measured by fMRI.
Functional links are inferred by thresholding pairwise Pearson correlations (see Equation 1)
between a pair of voxels, as standard in the literature (Bullmore & Sporns, 2009; Gallos et al.,
2012; Hermundstad et al., 2013).
The pairwise correlation is defined as follows:
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cij =
(cid:2)
Network Neuroscience
(cid:3)xixj(cid:4) − (cid:3)xi(cid:4)(cid:3)xj(cid:4)
i (cid:4) − (cid:3)xi(cid:4))2((cid:3)x2
((cid:3)x2
j (cid:4) − (cid:3)xj(cid:4))2
,
(1)
138
Core language brain network
Voxel-level functional network:
Functional connectivity network
where each node represents a voxel
and a link depicts the functional
interdependency between a pair of
voxels.
where xi is a vector encoding the fMRI time response of voxel i and (cid:3)·(cid:4) indicates a temporal
average.
Accordingly, pairs of voxels with correlation above a fixed threshold are connected by a link
(Bullmore & Sporns, 2009; Del Ferraro et al., 2018; Lucini et al., 2019). The threshold is an
absolute threshold, which means we pick a reference value of the correlation as the smallest
and assign a zero to any value below. The link weight is given by the correlation strength
as defined above in Equation 1. Nearby active voxels are grouped together based on each
subject’s individual anatomy and are considered part of the same fROI. Figure 2, upper panel,
shows a realization of the voxel-level functional network for a representative subject, where
voxels that are part of the same fROI are colored equally.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
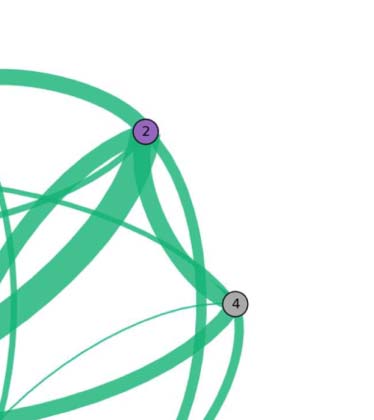
Figure 2. Two visualizations of the individual functional network. The figure shows functional
networks for a representative subject relative to the fMRI active brain areas during the language task.
Upper panel: voxel-level network. Each node in the network represents a voxel, each link connects
a pair of voxels in different brain modules, and it is indicative of functional interdependency. Links
connecting voxels within the same brain module are not visible but exist. Lower panel: fROI-level
network for the same voxel-level architecture shown in the upper panel. Voxels belonging to the
same anatomical region are grouped into an fROI, represented as a node in the network. Node’s
size is proportional to the number of voxels in the fROI. Colored borders have no meaning and are
used only for illustrative purpose. Each link’s thickness connecting two fROIs is proportional to the
sum of link’s weight connecting all the voxels in the two fROI (exact definition given in Equation 2).
Network Neuroscience
139
Core language brain network
We define fROIs within each subject individually, based on the activation and anatomy
of the specific subject (Del Ferraro et al., 2018; Fedorenko, Hsieh, Nieto-Castañón, Whitfield-
Gabrieli, & Kanwisher, 2010; Lucini et al., 2019). For instance, all the active voxels in
Brodmann area 22 of the superior temporal lobe define the Wernicke’s area fROI. The rea-
son for choosing individual-based fROIs is that group-based ROI-level analysis suffers from
intersubject variability in the location of activation. In contrast, individual-subject-based fROI
analysis can reveal greater functional specificity (Fedorenko et al., 2010). Furthermore, working
in individual native space prevents the propagation of errors due to coregistration to universal
ATLAS.
fROI-level functional network:
Course-grained representation of a
voxel-level functional network.
Each node represents an fROI and
a link depicts the functional
interdependency between a pair of
fROIs.
At the fROI level or module level a node represents an entire fROI, that is, a group (col-
lection) of nearby active voxels in the spatially (anatomically) proximate area. Hereafter, we
might use the word “module” or brain “region” as substitute of the word fROI. At this level, a
functional link connects two fROIs if and only if there exists at least one link, at the voxel level,
between a pair of voxels in the two fROIs. The functional link’s weight between two fROIs i
and j (Wij) is defined as the sum of the number of links connecting pairs of voxels between
the two fROIs, normalized by the sum of the two fROIs’ size:
Wij = #links connecting i ↔ j
size(fROIi) + size(fROIj) .
(2)
For each individual, we then normalize each Wij by the value of the largest W for that individual
(Wmax):
˜Wij =
Wij
Wmax ,
for all i and j fROIs.
(3)
In this way the link’s weight scale is the same across subjects (see Supplementary Table S1)
and the maximum weight is ˜W = 1 in each individual. Figure 2, lower panel, illustrates the
functional network at the fROI level for a representative subject.
For each individual, we are interested in uncovering the functional architecture of the
subdivision of the Broca’s area, that is, tri-BA and op-BA, which correspond to all active
voxels in Brodmann area 44 and 45, respectively. Each of these subareas has been associ-
ated with different language processes in previous studies (Devlin et al., 2003; Gough et al.,
2005; Newman et al., 2003; Nixon et al., 2004). Through our analysis we aim to find out
the specificity of their functional connectivity, to unveil whether their different engagement in
language processing may be associated with a different functional wiring with the rest of the
brain. Thus, when building the individual functional network, we group the active voxels of
the BA into two different and separate fROIs: op-BA and tri-BA (see Figure 2).
We named the fROIs according to their main anatomical boundaries as follows. We re-
tained the classical designations of BA (Brodmann area 44–45, inferior frontal gyrus) and WA
(Brodmann area 22, superior temporal gyrus), as these designations still predominate in neuro-
surgery, which dominates clinical practice (Friederici, 2011). We defined the ventral premotor
area (v-preMA) as the ventral portion of the premotor cortex, which includes the inferior part
of Brodmann area 6, centered on the posteriormost portion of the middle frontal gyrus (MFG;
Friederici, 2011). The superior portion of Brodmann area 6 was considered dorsal premotor
area (d-preMA). The anteriormost part of the middle frontal gyrus was identified as anterior
middle frontal gyrus (aMFG). The pre-SMA was defined within the medial frontal cortex, at the
level of Brodmann area 6 (Nachev, Kennard, & Husain, 2008). The precentral gyrus was iden-
tified with Brodmann area 4, the supramarginal gyrus was identified with Brodmann area 40
Network Neuroscience
140
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core language brain network
and angular gyrus with Brodmann area 39 (Friederici, 2011). The deep opercular cortex (DOC)
included the innermost portion of the frontal operculum (Friederici, 2011).
The visual and the auditory cortex, which are active areas that support nonlinguistic pro-
cessing, were excluded from the analysis (Fedorenko et al., 2010; Fedorenko & Kanwisher,
2009). These areas are indeed activated because the subject is presented with auditory stimuli
and may keep the eyes open.
The same functional network construction as described above is carried over for all 20
subjects individually, both at the voxel and the fROI level. Next, we carry out a group analysis
to identify the common functional network shared across individuals, beyond the intersubject
variability, as described in following section.
Common network construction across subjects. Our interest in studying functional networks for
single individuals performing language tasks is aimed at uncovering functional architectures
that are persistent across healthy subjects and could be useful and informative when dealing
with clinical cases. Individual functional networks have innate subject variability (e.g., one
subject activates in one specific area or has a functional link while another does not). Therefore,
after we reconstructed the individual functional networks, we performed a group analysis at
the fROI level by investigating which set of links and brain areas is persistent across subjects
or, in other words, which functional subarchitecture is common among all the individuals.
This functional architecture is informative of which areas and functional links persist beyond
the intersubject variability, and therefore it represents a language core structure for the specific
language task under study. Accordingly, surgical intervention, as for instance tumor resection,
should operate by preserving such core structure existing across healthy controls. In addition,
functional damage to this structure due to brain pathologies—and observed from the functional
connectivity of the patient—may be informative of the damage extent (e.g., a missing functional
link in the core may signify a larger harm than a missing connection between more peripheral
areas not in the core). We name this most persistent functional architecture across subjects,
at the fROI level, common network. This common architecture is defined retaining a pair of
fROIs and a functional link connecting them only if these areas and link are present across
subjects.
The weight of the functional link connecting two fROIs i and j in the common network (WC
ij )
is defined as the average of the ˜Wij connecting those fROIs across subjects:
WC
ij = 1
N
N
∑
l=1
˜W(l)
ij
,
(4)
where N is the total number of individuals.
We report and discuss the results of this quantitative analysis in the following sections.
RESULTS
Individual Networks
For each individual we observe fMRI activation in both hemispheres, however, left dominance
is clearly observed, as expected since all the subjects are right-handed (Isaacs, Barr, Nelson,
& Devinsky, 2006; Knecht et al., 2000). The number of left hemisphere areas of activation is
greater and in most cases their frequency of activation is greater as well.
Network Neuroscience
141
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core language brain network
Active fROIs across subjects include the following, in alphabetic order: angular gyrus (L),
Broca’s area (L; op-BA and tri-BA), Broca’s area (R), caudate (L and R), deep opercular cortex
(L and R; Friederici, 2011), aMFG (L and R), precentral gyrus (L and R), ventral and dorsal
preMA (L), ventral preMA (R), pre-SMA, supramarginal gyrus (L and R), and Wernicke’s area
(L and R). Detailed information on the frequency of activation of each area across subjects is
summarized in Supporting Information Table S2. In the following, for brevity, we will refer to
left hemisphere brain areas simply with the name of the areas, omitting the specification (L).
The functional network for a representative subject at both the voxel and the fROI level is
shown in Figure 2. All the single subjects’ functional connectivity at the fROI level for each
of the 20 healthy individuals considered in our study are shown in Supporting Information
Figure S1, and all the connectivity values between pairs of fROIs are reported in Supporting
Information Table S1.
We observe that, overall, the preMA is the most connected area across subjects, in terms of
connectivity weight. In 8 over 20 individuals the strongest functional connection is between
preMA–op-BA and in 7 over 20 cases it is between preMA–pre-SMA. In total, the preMA turns
out to have the strongest connection with one of the other areas in 17 out of 20 subjects; in
3 cases the strongest functional connection is between op-BA and tri-BA. For further details
on the connectivity of each area, see Supporting Information Table S1.
Wernicke’s area is known to structurally connect to BA through the arcuate fasciculus, a
bundle of axons linking the inferior frontal gyrus with the superior temporal gyrus. We inves-
tigated the functional connections of the BA subdivisions with the rest of the brain and, with
focus on WA, at the fROI level, we compared how frequent op-BA connects to WA versus how
frequent tri-BA connects to WA. We find that op-BA connects to WA in 18 out of 20 subjects
(90% of the cases), while tri-BA connects to WA in 15 out of 20 individuals (75% of the cases).
In terms of connectivity weight, in 10 out of 20 subjects (50%) WA connects more strongly to
op-BA than to tri-BA, whereas in 7 subjects (35%) we have the opposite finding, tri-BA con-
nects more to WA than the opercular counterpart. In 2 individuals the functional connectivity
of op-BA and tri-BA to WA is, instead, approximately the same. One subject does not show
WA activation at all.
Regarding other relevant areas such as preMA and pre-SMA, we find that the connectivity
frequency of these areas with op-BA and tri-BA is about the same. Indeed the preMA connects
to op-BA in 18 subjects and to tri-BA in 17 out of 20. The pre-SMA connects to op-BA in
19 subjects and to tri-BA in 18 individuals. So, overall, the connectivity frequency of the BA
subdivisions with preMA and pre-SMA is similar. In terms of connectivity weight, op-BA con-
nects more strongly to both preMA and pre-SMA compared with tri-BA. Thus, although the BA
subdivisions connect to preMA and pre-SMA with about the same frequency across subjects,
op-BA has, overall, a larger connectivity weight.
To investigate the overall functional connectivity that each fROI has with the rest of the
common network shown in Figure 3 we computed, for each individual, the sum of the func-
tional links of each fROI with the rest of the network. In each subject, each fROI that appears
in the common network can then be ranked according to this connectivity strength (most con-
nected at the top and least connected at the bottom). We present these results in Supporting
Information Figure S2. The results show that preMA is either the most connected or the second
most connected area in 95% of the cases. On the contrary, WA is either the least connected
or the second least connected area in 95% of the cases.
Network Neuroscience
142
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core language brain network
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
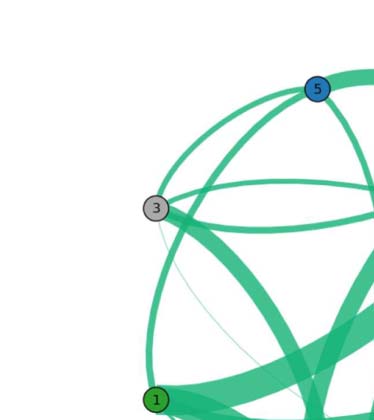
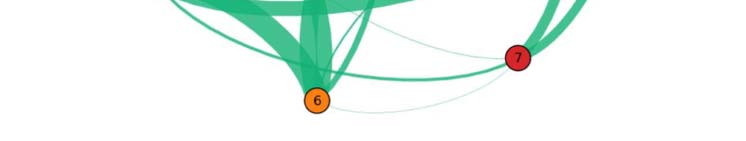
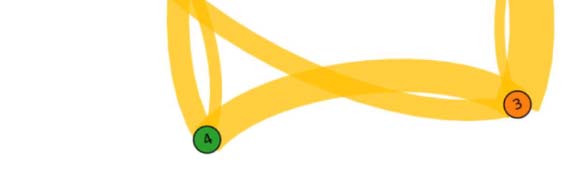
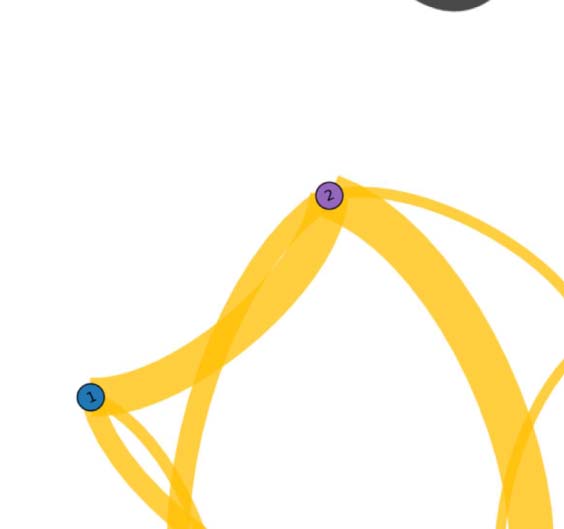
Figure 3. Common network across subjects for the language task under study. The figure illustrates
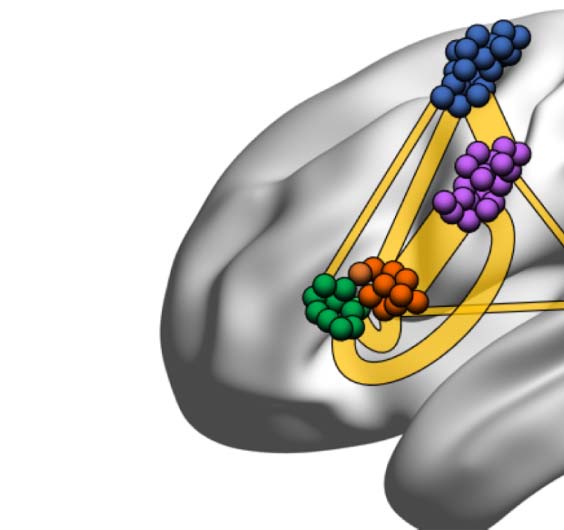
the functional network, beyond intersubject variability, shared across individuals (17 out of 20).
The weight of a link connecting two fROIs is proportional to the average of the functional links
connecting those fROIs across subjects. Upper panel: fROIs are located on their anatomical loca-
tion on the brain. Lower panel: pictorial illustration of the network in the upper panel, with the
fROIs equally spaced on a plane.
Common Network Across Subjects and Functional Subdivisions of Broca’s Area
The common network at the fROI level, as described in the Common Network Construction
section, is made by those fROIs and links present (persistent) across the majority of subjects. As
a result of the left dominance at the individual level, no consistent overlap of right-hemisphere
activation has been found across subjects.
We find that the persistent structure across individuals (17 over 20), beyond intersubject
variability, is made by op-BA, tri-BA, WA, preMA, and pre-SMA connected together in a func-
tional architecture (see Figure 3). This circuitry represents the core structure for the specific
clinical language task under investigation since it is the functional architecture that prevails
in nearly all subjects. We find this network in 17 over 20 subjects and not in all of them be-
cause three subjects show lack of activation for either the op-BA (1 case), the tri-BA (1 case),
or neither WA nor tri-BA (1 case). The common network shown in Figure 3 is therefore the one
prevailing in closely all the subjects and, thus, the functional structure that is persistent be-
yond intersubject variability. We tested the robustness of these results by varying the threshold
in each individual network (discussed in the Individual Brain Network Construction section)
by 5−10% and by recomputing the common network analysis. We find that within this range
of variation the common network is made by the same brain modules (fROIs) and links shown
in Figure 3.
Network Neuroscience
143
Core language brain network
Furthermore, this conclusion is additionally supported by additional findings we obtained
on a study conducted on bilingual healthy subjects when they speak their native language (Li,
Pasquini, et al., 2019). In Li, Pasquini, et al. (2019) we study the functional network of English
monolingual subjects and of bilinguals (native in Spanish, bilingual in English). We find that
for both groups, when subjects speak their native language the persistent structure is made by
BA, WA, preMA, and pre-SMA. Additionally, data in Li, Pasquini, et al. (2019) were acquired
for a letter-generation language task, that is, a different task that we use in the present paper
(verb generation). Thus, results of Li, Pasquini, et al. (2019) together with the present findings
support evidence that the core language network is the most consistent functional architecture
beyond intersubject variability and it is not limited to one specific language task.
max
max) is between op-BA and preMA (WC
In terms of functional connectivity, the strongest connectivity weight in the common net-
work (WC
= 0.74 ± 0.31, where the average is made
across all the subjects that have such link). The triangular BA also connects with the preMA but
with about half of the magnitude (WC = 0.37 ± 0.29). Detailed information on the functional
connection of the other areas in the common network is reported in Supporting Information
Table S3. Broca’s area has been long recognized as a central language area; its strong connec-
tivity with the preMA(L) is of particular interest since the preMA(L) has been more recently iden-
tifiedas an area with dominant role for language (Duffau et al., 2003). We discuss this result further
in the Discussion subsection: Functional and Structural Connectivity of the Common Network.
When we look at the connectivity of the BA subdivisions with Wernicke’s areas, a primary
area for language comprehension, we observe that, in the common network, WA only connects
to op-BA. This reveals the existence of larger coactivation of the BOLD signal between these
two areas that might also be driven by their spatial vicinity (WA is anatomically closer to op-BA
than to tri-BA). In more detail, as discussed in Results section, at the individual level we find that
WA connects to tri-BA in 15 out of 20 cases whereas it connects to op-BA in 18 out of 20 cases.
Therefore, overall, BA subdivisions both connect to WA in several different individuals with
a slightly larger presence of WA–op-BA connectivity across subjects. In terms of connectivity
weight, when we count only subjects where both op-BA and tri-BA connect to WA, we find
that op-BA connects slightly more strongly to WA compared with tri-BA (WC = 0.17 ± 0.23
versus WC = 0.15 ± 0.20, respectively).
Furthermore, we observe that op-BA has a larger connectivity than tri-BA both on the
number of connections with the rest of the areas in this network (4 versus 3 respectively, the
extra one being WA–op-BA) and in terms of functional connectivity weight. Indeed, the av-
erage connectivity of the op-BA, across subjects and across areas, in the common network is
WC = 0.45 ± 0.25 (WC = 0.55 ± 0.20 without the link WA–op-BA), whereas the comprehen-
sive connectivity weight of the tri-BA is WC = 0.32 ± 0.18.
Finally, we observe that the average values for the common-network functional weights
reported in Supporting Information Table S3 have large standard deviations (magnitude com-
parable with the mean). This result signals a large intersubject variability for the weight of the
single functional link across subjects. To investigate this further, we plot the empirical distri-
bution of all the functional links’ weights across subjects and observe that it displays a long
tail shape (see Supporting Information Figure S3), which is explicative of the large standard
deviation values.
The Common Network is Part of the Maximum k-core: The Most Resilient Architecture
The notion of k-core in theoretical physics has been used as a fundamental measure of cen-
trality and robustness within a network (Morone et al., 2019). Since it was first introduced in
Network Neuroscience
144
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core language brain network
social sciences (Seidman, 1983) it has been used in several contexts (Kitsak et al., 2010), as
in random network theory (Pittel et al., 1996) or to describe large-scale structure in the brain
(Hagmann et al., 2008).
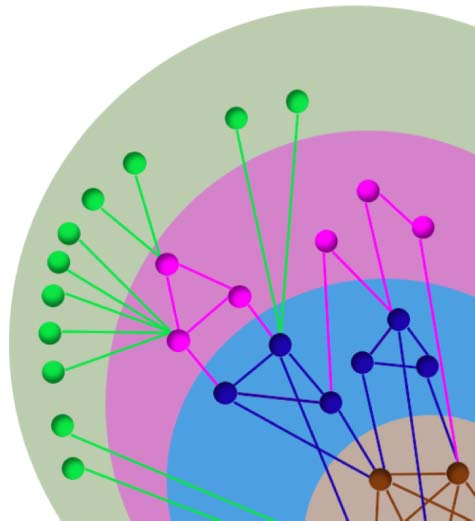
The k-core of a given architecture is defined as the maximal subgraph, not necessarily glob-
ally connected, made of all nodes having degree (number of connections) at least k. In practice,
the k-core subgraph can be derived by removing from the network all nodes with degree less
than k. The removal of these nodes reduces the degree of their neighbors, and if the degree
of the latter drops below k then also these nodes should in turn be removed. The procedure
iterates until there are no further nodes that can be pulled out from the network. The remain-
ing graph is the k-core of the network. A k-core structure includes subnetworks with higher ks:
k + 1, k + 2, and so forth. For instance, the 1-core includes the 2-core which, in turn, includes
the 3-core and so forth (see Figure 4). In each k-core, nodes in the periphery (not included in
the k + 1-core) are called k-shell (ks). Thus, in each network, k-core (and k-shell) structures are
nested within each other with increasing k. The innermost structure of the network corresponds
to the graph with the maximum k-core (Dorogovtsev et al., 2006). Thus, by definition, the max
k-core is not nested into any other structure with higher k. As a consequence, by definition,
the max k-shell always coincides with the max k-core. Figure 4A illustrates k-core and k-shell
structures in a simple explanatory network.
Recently, the maximum k-core (kmax
core) has been linked to the most resilient structure of bio-
logical systems with positive interactions (Morone et al., 2019) and, in an fMRI study of human
brains, the kmax
core of the functional connectivity for a visual-task-based experiment has been
found to be the most robust structure, which remains active even during subliminal conscious
states (subject not aware of seeing images; Lucini et al., 2019).
Motivated by these recent findings, we pruned each voxel-level individual functional net-
work until the maximum k-core structure, and we investigated to which k-core each node
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
k-core and k-shell of a network. Panel A illustrates pictorially a network. Nodes in
Figure 4.
the same disk have the same k-core. A k-core structure includes subnetworks with higher ks, so the
1-core includes the 2-core which, in turn, includes the 3-core, and so forth. Nodes that are in the
k-core but not in the k + 1-core are called k-shell and are colored differently. The maximum k-core
= 4 and depicted with brown nodes.
coincides with the maximum k-shell, in this network is kmax
core
Panel B illustrates pictorially the construction of the k-core histogram shown in Figure 5. Note that
here nodes in each k-shell are colored differently, whereas in Figure 5 different colors indicate nodes
in different fROIs, piled up according to their k-shell as in this panel.
Network Neuroscience
145
Core language brain network
(voxel) belongs. We focused on the areas part of the common network (BA, WA, pre-SMA,
and preMA) because these are the fROIs that form a persistent language structure across in-
dividuals, as shown in Figure 3. We aim to explore whether these regions are part of some
significative k-core structure that might shed light on the architecture of the network. Our goal
is to investigate, across subjects, which fROIs characterize the occupancy of each k-shell and,
thus, we proceed as follows. For each individual network we compute the k-core and k-shell
of all the nodes (voxels) as described above. Each subject has, in general, a different kmax
core
(k-shell), which is a integer number that can go up to the maximum degree of the subject’s
network. Thus, in order to compare results across subjects, we normalize the k-core range-
of-values to 1, in each subject. We do this by dividing all the k-core (k-shell) values in each
individual network by the individual kmax
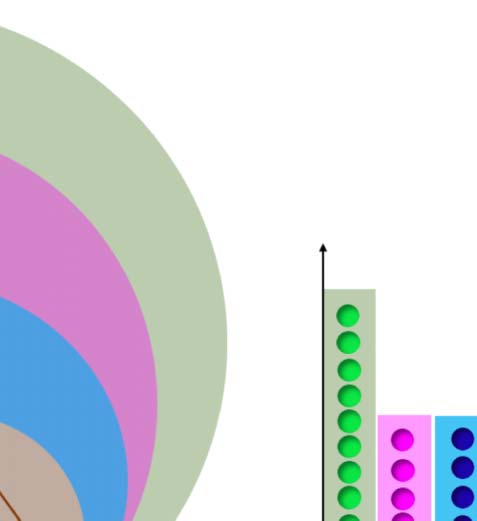
core for that network. In Figure 5 we then plot the total
k-shell occupancy for all the individuals, and we color differently the contribution of each fROI
in order to visualize to which k-shell they belong.
Results in Figure 5 show that the maximum k-shell (which is in turn the maximum k-core)
is the most populated of all the k-shells of the common network. More importantly, if we look
at each area individually, we observe that the largest concentration of pre-SMA, op-BA, tri-BA,
and v-preMA is in the maximum k-shell. Among the areas of the common network, WA is the
only area that does not have the largest portion in the kmax
core but, rather, populates smaller k-shell
values. These results are robust to 5−10% variation of the threshold used to build individual
functional networks (discussed in the Individual Brain Network Construction section). In order
to asses how much of our results are given by construction we compared them with randomized
voxel assignation to fROI and recomputed the k-shell occupancy as a null model. Results are
shown in Supporting Information Figure S4 and show that the relative occupancy of each fROI
in the maximum k-shell is different from the random case. Furthermore, WA in the real model
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
n
e
n
_
a
_
0
0
1
1
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
k-shell occupancy. The histogram shows the k-shell occupancy for nodes in the four
Figure 5.
fROIs of the common network of Figure 3. Overall, the majority of the nodes of this structure are
located in the maximum k-shell, which coincides with kmax
core, a quantity linked to the robustness of
a complex network (Morone et al., 2019). Of the four fROIs of the common network, the pre-SMA,
op-BA, tri-BA, and ventral preMA are mostly part of the kmax
core. Wernicke’s area (WA) is more an
outlier; it is mostly located in lower k-shells and minimally located in the kmax
core.
Network Neuroscience
146
Core language brain network
populates more the smaller shells compared with the null model case. We conclude that our
results are not due to a random effect.
In Morone et al. (2019) the authors have shown that, for complex networks with positive
couplings, the kmax
core of the network is the most resilient structure under decreasing of the cou-
pling weight. In our functional networks, all the links are obtained through thresholding of
pairwise correlations that, from our findings, turn out to be all positive. This is because the
BOLD signal is extracted from a task-based fMRI experiment, stimulated by an external input.
In this way, active voxels are those mostly correlated with the task model and, when comput-
ing pairwise correlations among voxels correlated with the same external stimulus, most likely
one finds positive correlations, as we observe from our data analysis. This result allows us to
interpret the functional networks wired by positive interactions and, therefore, the theory of
Morone et al. (2019) applies. Accordingly, we can interpret the maximum k-core structure of
our network as the most resilient one under decreasing of the correlation weight.
In other words, the circuitry made by the pre-SMA, BA, and preMA represents the most
robust structure of the functional network. Wernicke’s area, although it is part of the common
network, for the most part does not lay in the kmax
core of the network, probably because of its more
peripheral anatomical location compared with the other fROIs of this common architecture.
Therefore, although it is one of the most important areas for language, it is not part of the most
resilient core.
Rich-Club Response to Changes in Threshold
The rich-club coefficient is a measure of the connectivity as a function of degree (Zhou &
Mondragón, 2004). If subnetworks of nodes have a tendency towards a higher percentage of
connectivity for larger degrees, then the network is said to be a “rich club.” It has been shown
that the brain structural network typically conforms to this paradigm (van den Heuvel & Sporns,
2011). Since the number of connections will naturally increase with degree, it is standard to
normalize the rich-club coefficient by the coefficient obtained for a random network with the
same number of nodes and degree distribution. The rich-club coefficient is defined as
φ(k) =
2E>k
N>k(N>k − 1) ,
(5)
where E>k and N>k is the number of edges and nodes in the network where all nodes with
φ(k)
degree less than k have been removed. The normalized coefficient is then defined by
φrand(k)
where φrand(k) refers to the rich-club coefficient calculated in the same network that has been
randomly rewired while preserving the degree distribution.
In the Individual Brain Network Construction section we defined an absolute threshold for
the voxel-voxel correlation in order to define what represents a minimum edge weight that
defines links in the functional language networks. Tuttavia, the choice of threshold is some-
what arbitrary and may have an effect on the results. In order to probe how the connectivity
responds to changes in the choice of threshold, we show normalized rich-club coefficients for
a representative subject in Figure 6.
The threshold was lowered by 5% and raised by 5% in each case, and we can see that
the rich-club structure is evident as shown by the rising curve above 1 as a function of k up
to the high-degree sector of
the network where there is high volatility. We also ob-
serve that a 5% change in threshold value does not change qualitatively the behavior of the
rich-club coefficient. These results show that, in addition to the rich-club behavior found in
Network Neuroscience
147
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
Figura 6. Rich clubs. Normalized rich-club coefficients for a representative subject with 5%
(lowered/raised) variation of the current threshold. All other subjects have qualitatively the same
normalized rich-club coefficient. The red curve corresponds to the original threshold (rete).
The blue curve corresponds to a 5% lower variation of the threshold, which leads to a denser net-
lavoro. The green curve corresponds to a 5% higher variation of the threshold, which leads to a
sparser network. The figure shows rich-club behavior of the functional brain network, showing an
increasing normalized rich-club coefficient.
(van den Heuvel & Sporns, 2011), functional brain networks, such as those we studied in this
lavoro, also show the same feature.
DISCUSSION
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
T
.
In this study, we reconstructed the functional language network of 20 healthy subjects from
tb-fMRI data providing information about the functional connectivity between active areas on
fMRI maps, with both a voxel- and an fROI-level resolution. The language task designed for the
experiment is customarily used in clinical cases and has shown to produce robust activation in
previous studies (Brennan et al., 2007; Li, Dong, et al., 2019; Ramsey et al., 2001; Xiong et al.,
2000). Functional activation is generally sensitive to the fMRI task employed; our interest in
reconstructing functional networks for this specific task aimed to create benchmark results for
healthy individuals that can be used as reference for functional networks affected by brain
pathologies. Infatti, brain impairments are known to create damage to the functional con-
nectivity. It is therefore paramount to have healthy functional architectures relative to clinical
language tasks in order to make the comparison between healthy individuals, and patients’
functional networks possible.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Our main finding is the existence of a common persistent functional network across subjects
that wires together BA, WA, ventral preMA, and pre-SMA in the left dominant hemisphere
for 17 out of 20 right-handed healthy subjects (Guarda la figura 3). We interpret this circuitry as a
core structure for the language task under study since this network persists across nearly all
individuals.
Inoltre, we compute the k-core of each node (voxel) in the common network—the
maximum value of which has been recently linked to network resilience in ecosystems and
Network Neuroscience
148
Core language brain network
Direct cortical stimulation:
Electrical stimulation of the cerebral
cortex with the aim of identifying
brain areas that take part in a specific
system function (per esempio., lingua,
motor). It remains the gold standard
for presurgical mapping of the motor
cortex and language areas to prevent
unnecessary functional damage.
fMRI studies (Lucini et al., 2019; Morone et al., 2019). We find that three out of four areas
of the common architecture (specifically pre-SMA, BA, and preMA) are mostly concentrated
in the maximum k-core of the network (Guarda la figura 5). This led us to conclude, following the
findings of Lucini et al. (2019) and Morone et al. (2019), that these areas are the most robust
of the language network in terms of fMRI-correlated signal.
Wernicke’s area is a crucial language area and indeed appears as part of the common net-
work across individuals, yet its type of connectivity with the rest of the fROIs in this architecture
is slightly different from the connectivity of the other areas. Infatti, overall, WA shares two con-
nections with other areas in this network, one with BA and one with the preMA, whereas each
of the other fROIs has at least three total functional connections. This might be a by-product
of the more peripheral location of the WA compared with the other fROIs, which being spa-
tially closer to each other are facilitated to coactivate because of white fibers wiring them
together. Wernicke’s area is also the only area among the four in the common network that
is not largely part of the maximum k-core (Guarda la figura 5). This result is in agreement with the
discussion above about this area and, Ancora, might be due to the more perimetric location of
the WA in the common network.
Finalmente, we investigated the functional architecture of the BA anatomical subareas, revealing
a different connectivity between tri-BA, op-BA, and the other areas of the common network
for this specific language task. In the first subsection, we discuss our findings regarding the
functional connectivity of the BA subdivisions, contextualizing them with known white matter
connections that these areas share with the rest of the brain, found in other studies.
Functional and Structural Connectivity of the Common Network
We observe that the left ventral preMA is the most connected area of the common network,
with four total connections and the strongest connectivity with op-BA (WC = 0.74 ± 31) E
with pre-SMA (WC = 0.64 ± 0.31). Come mostrato in figura 2 for a representative subject, the ven-
tral preMA is functionally connected to all the main cortical language areas of the dominant
hemisphere, suggesting that this area may play an important role in speech production (other
subjects show qualitatively the same feature; see Supporting Information Figure S1).
Tate, Herbet, Moritz-Gasser, Tate, and Duffau (2014) investigated the crucial cortical epi-
centers of human language function by means of intraoperative direct cortical stimulation in
165 consecutive patients affected by low-grade glioma. The study shows that speech arrest is
localized to the ventral preMA instead of the classical BA. Inoltre, the presence of gliomas
growing in the left ventral preMA has been related to a higher percentage of speech deficits
than gliomas infiltrating the classical BA, providing a possible clinical correlate of the results
of Tate et al. (Bizzi et al., 2012; Tate et al., 2014). Inoltre, Duffau et al. (2003), through
intraoperative functional mapping in awake patients, have concluded that the left dominant
preMA seems to play a major role in language since its electrical stimulation causes speech
disturbances. All these results, together with the findings of our study, pinpoint a central role
of the preMA in language production.
Tuttavia, one must be careful not to overinterpret these results, as the highest connectivity
does not necessarily imply a central or essential role of that particular fROI in the network.
Using advanced graph theoretical analysis, Morone and Makse (2015) demonstrated that the
most connected nodes in a network often do not correspond to the most essential nodes,
the elimination of which would lead to collapse of that particular network. This idea has been
Network Neuroscience
149
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
recently tested on functional networks obtained from fMRI of rodent brains and verified through
in vivo pharmaco-genetic intervention (Del Ferraro et al., 2018).
Although the correspondence between structural and functional connectivity is not fully
understood yet (Honey et al., 2009), the arrangement displayed by our study is supported by
structural evidence. The existence of a physical connection between ventral preMA and BA
seems realistic, given their spatial contiguity. Besides representing a shared origin for the main
bundles of the dorsal pathway (Chang, Raygor, & Berger, 2015; Dick, Bernal, & Tremblay,
2014), the two areas may be directly connected by a specific opercular-premotor fascicle
(described in the next section; Lemaire et al., 2013).
The pre-SMA shows connectivity with both ventral preMA and BA (Guarda la figura 3, Supporting
Information Figure S1, and Supporting Information Table S3). These functional connections
are consistent with the organization of the structural language connectome to some extent:
the frontal aslant tract (FAT), an association motor pathway that underlies verbal fluency and
connects pre-SMA and BA (Catani et al., 2013; Ford, McGregor, Case, Crosson, & White,
2010; Jenabi, Peck, Young, Brennan, & Holodny, 2014), likely includes projection to posterior
regions of the MFG, corresponding to the ventral preMA (Chang et al., 2015).
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
The low connectivity weight between ventral preMA and WA (see Supporting Information
Table S3) may be explained by the increased distance between the two structures. Of note, we
find that the functional connectivity weight between op-BA and WA is similar to that of the
ventral preMA and WA (Supporting Information Table S3), which is consistent with their struc-
tural connection through the same white matter tract, corresponding to the arcuate component
of the AF/SLF system (Chang et al., 2015; Dick et al., 2014).
Broca’s Area Subdivisions
Our findings show that the subdivisions of Broca’s area present different patterns of connectivity
within the language network, with the opercular portion appearing more connected to all the
significant nodes of the common network compared with the triangular part. This evidence
appears in line with the structural architecture of the network.
The prominent interaction between ventral preMA and op-BA found in this study (WC =
0.74 ± 0.31, see Supporting Information Table S3) supports the evidence of a structural link
between op-BA and preMA, as suggested by Lemaire et al. (2013) using DTI analysis. IL
authors investigated the structural connectome of the extended BA, identifying the U-shaped
opercular-premotor fasciculus that connects the op-BA to the ipsilateral preMA (Lemaire
et al., 2013). On the contrary, tri-BA and ventral preMA showed lower functional connectivity
(WC = 0.37 ± 0.29), possibly suggesting indirect communication through the op-BA.
The second strongest functional connection between BA’s subareas and other fROIs of the
common network that we find is the link between op-BA–pre-SMA (WC = 0.35 ± 0.23). These
two areas are connected by the FAT (Ford et al., 2010), which originates in the SMA/pre-SMA
and terminates into the posteriormost aspect of the inferior frontal gyrus (IFG) (Catani et al.,
2013). Triangular BA and pre-SMA share a lower functional connectivity weight (WC = 0.20 ±
0.21) compared with op-BA and pre-SMA, reflecting the anatomic boundaries of the FAT.
Finalmente, the functional link between op-BA and WA is in line with the evidence of a dorsal
pathway of language between op-BA and STG through the AF/SLF system (dorsal pathway II;
Friederici, 2011).
Network Neuroscience
150
Core language brain network
Limitations of the Study
Functional networks are task dependent and brain impairments, such as brain tumors, affect
their functional connectivity (per esempio., by destroying functional links or preventing the fMRI acti-
vation of entire brain regions). In our study, we focused on a clinical language task since we
wanted to enhance a functional network that emerges in clinical studies. We employed healthy
subjects because we wanted to find out the network associated with the clinical language task
without possible functional damage induced by brain impairments. A comparison with func-
tional networks of patients with brain impairments is of great interest but goes beyond the
purpose of the present paper and it is, instead, presented in a follow-up (Del Ferraro, Pasquini,
Peck, Holodny, & Makse, 2019). In Del Ferraro et al. (2019) we compare the functional net-
work as well as the structural network (obtained through diffusion tractography imaging) Di
patients who present speech impairment under awake cortical electrostimulation and patients
who do not. As a control, we also compare these networks with the same architecture for
healthy subjects, and the results of the present paper are further used as a benchmark.
Our study is limited to a specific clinical language task (cioè., verb generation), and since
functional networks are task-dependent, in principle, our results could be limited to this par-
ticular language task. In a follow-up work (Li, Pasquini, et al., 2019) we investigate the func-
tional connectivity differences between healthy subjects who are monolinguals, native English
speakers, and subjects who are bilingual, native Spanish speakers who speak English as a sec-
ond language. We employ a different type of language task (cioè., letter generation), and in both
datasets we find that the core language network is the same as the one we find in the present
paper by using a verb-generation task and discussed in the Results section. This strengthens
our findings showing that the core common network is not limited to a specific language task
but that it might be a robust structure shared across several tasks. Further studies that make
use of other language tasks are needed to elucidate this point and the generality of the core
common network.
As a final limitation of our study we remark that the sample size of our data is limited to 20
individuals, and it would be important, in following works, to test the core network analysis
on larger datasets.
ACKNOWLEDGMENTS
We thank Mehrnaz Jenabi for help with AFNI and FSL software and Medeleine Gene for help
with mining the data.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00112.
Data that support the findings of this study are publicly available and have been deposited in
http://www-levich.engr.ccny.cuny.edu/webpage/hmakse/brain/ (Li, Del Ferraro, et al., 2019).
AUTHOR CONTRIBUTIONS
Qiongge Li: Formal analysis; Methodology; Software; Visualization; Writing – Original Draft;
Writing – Review & Editing. Gino Del Ferraro: Methodology; Software; Supervision; Writing –
Original Draft; Writing – Review & Editing. Luca Pasquini: Methodology; Validation; Writing –
Review & Editing. Kyung K. Peck: Data curation; Writing – Review & Editing. Hernán A. Makse:
Network Neuroscience
151
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
Funding acquisition; Investigation; Supervision; Writing – Review & Editing. Andrei I. Holodny:
Funding acquisition; Investigation; Supervision; Writing – Review & Editing.
FUNDING INFORMATION
Hernán A. Makse, National Institutes of Health (http://dx.doi.org/10.13039/100000002), Award
ID: 1R01EB022720. Craig Thompson, National Institutes of Health (http://dx.doi.org/10.13039/
100000002), Award ID: P30 CA008748. Hernán A. Makse, National Science Foundation
(http://dx.doi.org/10.13039/100000001), Award ID: 1515022. Tim Ahles, National Institutes
of Health (http://dx.doi.org/10.13039/100000002), Award ID: U54CA137788. Tim Ahles, Na-
tional Institutes of Health (http://dx.doi.org/10.13039/100000002), Award ID: U54CA132378.
Luca Pasquini, Italian Scientists and Scholars in North America Foundation (http://dx.doi.org/
10.13039/100009799), Award ID: Imaging chapter award 2018. Luca Pasquini, European
Society of Radiology, Award ID: Bracco clinical fellowship 2018. Qiongge Li, City University
of New York (http://dx.doi.org/10.13039/100006462), Award ID: Doctoral student research
grant.
REFERENCES
Aubert, A., Costalat, R., Duffau, H., & Benali, H. (2002). Modeling
of pathophysiological coupling between brain electrical activa-
zione, energy metabolism and hemodynamics: Insights for the in-
terpretation of intracerebral tumor imaging. Acta Biotheoretica,
50, 281–295.
Bassett, D. S., Wymbs, N. F., Rombach, M. P., Porter, M. A., Mucha,
P. J., & Grafton, S. T. (2013). Task-based core-periphery organ-
ization of human brain dynamics. PLoS Computational Biology,
9, e1003171.
Bizzi, A., Nava, S., Ferrè, F., Castelli, G., Aquino, D., Ciaraffa, F., . . .
Piacentini, S. (2012). Aphasia induced by gliomas growing in the
ventrolateral frontal region: Assessment with diffusion MR trac-
tography, functional MR imaging and neuropsychology. Cortex,
48, 255–272.
Bookheimer, S.
(2002). Functional MRI of language: New ap-
proaches to understanding the cortical organization of semantic
processing. Annual Review of Neuroscience, 25, 151–188.
Booth, J. R., Legna, L., Lu, D., Houk, J. C., & Bitan, T. (2007). IL
role of the basal ganglia and cerebellum in language processing.
Brain Research, 1133, 136–144.
Brennan, N. M. P., Whalen, S., de Morales Branco, D., O’Shea, J. P.,
Norton, IO. H., & Golby, UN. J. (2007). Object naming is a more
sensitive measure of speech localization than number counting:
Converging evidence from direct cortical stimulation and fMRI.
NeuroImage, 37, S100–S108.
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10, 312.
Catani, M., Mesulam, M. M., Jakobsen, E., Malik, F., Martersteck,
A., Wieneke, C., . . . Rogalski, E. (2013). A novel frontal pathway
underlies verbal fluency in primary progressive aphasia. Brain,
136, 2619–2628.
Chai, l. R., Mattar, M. G., Blank, IO. A., Fedorenko, E., & Bassett,
D. S. (2016). Functional network dynamics of the language sys-
tem. Cerebral Cortex, 26, 4148–4159.
Chang, E. F., Raygor, K. P., & Berger, M. S. (2015). Contemporary
model of language organization: An overview for neurosurgeons.
Journal of Neurosurgery, 122, 250–261.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29, 162–173.
Del Ferraro, G., Moreno, A., Min, B., Morone, F., Pérez-Ramírez,
Ú., Pérez-Cervera, L., . . . Makse, H. UN. (2018). Finding influential
nodes for integration in brain networks using optimal percolation
theory. Nature Communications, 9, 2274.
Del Ferraro, G., Pasquini, L., Peck, K. K., Holodny, A., & Makse,
(2019). Structural and functional connectivity differ-
H. UN.
ences between brain tumor patients who exhibit intra-operative
speech impairments vs patients who show no speech deficit.
(In preparation)
Devlin, J. T., Matthews, P. M., & Rushworth, M. F. (2003). Seman-
tic processing in the left inferior prefrontal cortex: A combined
functional magnetic resonance imaging and transcranial mag-
netic stimulation study. Journal of Cognitive Neuroscience, 15,
71–84.
Dick, UN. S., Bernal, B., & Tremblay, P. (2014).The language connectome:
New pathways, new concepts. Neuroscientist, 20, 453–467.
Dorogovtsev, S. N., Goltsev, UN. V., & Mendes, J. F. F. (2006). K-core
organization of complex networks. Physical Review Letters, 96,
040601.
Dronkers, N. F., Plaisant, O., Iba-Zizen, M. T., & Cabanis, E. UN.
(2007). Paul Broca’s historic cases: High resolution MR imaging
of the brains of Leborgne and Lelong. Brain, 130, 1432–1441.
Duffau, H., Capelle, L., Denvil, D., Gatignol, P., Sichez, N., Lopes,
M., . . . Van Effenterre, R. (2003). The role of dominant premotor
cortex in language: A study using intraoperative functional map-
ping in awake patients. NeuroImage, 20, 1903–1914.
Fedorenko, E., Hsieh, P.-J., Nieto-Castañón, A., Whitfield-Gabrieli,
S., & Kanwisher, N. (2010). New method for fMRI investigations
of language: Defining ROIs functionally in individual subjects.
Journal of Neurophysiology, 104, 1177–1194.
Network Neuroscience
152
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
Fedorenko, E., & Kanwisher, N. (2009). Neuroimaging of language:
Why hasn’t a clearer picture emerged? Language and Linguistics
Compass, 3, 839–865.
Ford, A., McGregor, K. M., Case, K., Crosson, B., & White, K. D.
(2010). Structural connectivity of Broca’s area and medial frontal
cortex. NeuroImage, 52, 1230–1237.
Friederici, UN. D. (2011). The brain basis of language processing: From
structure to function. Physiological Reviews, 91, 1357–1392.
Friederici, UN. D., Chomsky, N., Berwick, R. C., Moro, A., & Bolhuis,
J. J. (2017). Language, mind and brain. Nature Human Behavior,
1, 713–722.
Gallos, l. K., Makse, H. A., & Sigman, M. (2012). A small world of
weak ties provides optimal global integration of self-similar mod-
ules in functional brain networks. Proceedings of the National
Academy of Sciences, 109, 2825–2830.
Gough, P. M., Nobre, UN. C., & Devlin, J. T. (2005). Dissociat-
ing linguistic processes in the left inferior frontal cortex with
transcranial magnetic stimulation. Journal of Neuroscience, 25,
8010–8016.
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Wedeen, V. J., & Sporns, O. (2008). Mapping the structural core
of human cerebral cortex. PLoS Biology, 6, e159.
Hermundstad, UN. M., Bassett, D. S., Brown, K. S., Aminoff, E. M.,
Clewett, D., Freeman, S., . . . Carlson, J. M. (2013). Structural
foundations of resting-state and task-based functional connectiv-
ity in the human brain. Proceedings of the National Academy of
Scienze, 110, 6169–6174.
Hertrich, I., Dietrich, S., & Ackermann, H. (2016). The role of the
supplementary motor area for speech and language processing.
Neuroscience and Biobehavioral Reviews, 68, 602–610.
Honey, C., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J.-P.,
Meuli, R., & Hagmann, P. (2009). Predicting human resting-state
functional connectivity from structural connectivity. Proceedings
of the Natinal Academy of Sciences, 106, 2035–2040.
Isaacs, K. L., Barr, W. B., Nelson, P. K., & Devinsky, O. (2006).
Degree of handedness and cerebral dominance. Neurology, 66,
1855–1858.
Jenabi, M., Peck, K. K., Young, R. J., Brennan, N., & Holodny,
UN. IO. (2014). Probabilistic fiber tracking of the language and
motor white matter pathways of the supplementary motor area
(SMA) in patients with brain tumors. Journal of Neuroradiology,
41, 342–349.
Kitsak, M., Gallos, l. K., Havlin, S., Liljeros, F., Muchnik, L.,
Stanley, H. E., & Makse, H. UN. (2010). Identification of in-
fluential spreaders in complex networks. Nature Physics, 6,
888–893.
Knecht, S., Dräger, B., Deppe, M., Bobe, L., Lohmann, H., Flöel, A.,
. . . Henningsen, H. (2000). Handedness and hemispheric lan-
guage dominance in healthy humans. Brain, 123, 2512–2518.
Lee, M. H., Smyser, C. D., & Shimony, J. S. (2013). Resting-state
fMRI: A review of methods and clinical applications. AJNR, 34,
1866–1872.
Lemaire, J.-J., Golby, A., Wells III, W. M., Pujol, S., Tie, Y., Rigolo,
L., . . . Kikinis, R. (2013). Extended Broca’s area in the connec-
tome of language in adults: Subcortical single-subject analysis
using DTI tractography. Brain Topography, 26, 428–441.
Li, Q., Del Ferraro, G., Pasquini, L., Peck, K., Makse, H., & Holodny,
UN. (2019). Task-based fMRI of 20 healthy individual performing a
language task used for clinical studies. http://www-levich.engr.
ccny.cuny.edu/webpage/hmakse/brain/
Li, Q., Dong, J. W., Del Ferraro, G., Petrovich Brennan, N., Peck,
K. K., Tabar, V., . . . Holodny, UN. IO. (2019). Functional translo-
cation of Broca’s area in a low-grade left frontal glioma: Graph
theory reveals the novel, adaptive network connectivity. Frontiers
in Neurology, 10, 702.
Li, Q., Pasquini, L., Del Ferraro, G., Gene, M., Peck, K. K., Makse,
H. A., & Holodny, UN. (2019). Functional connectivity core dif-
ferences between monolinguals and bilinguals healthy brains.
arXiv:1909.03109
Lucini, F. A., Del Ferraro, G., Sigman, M., & Makse, H. UN. (2019).
How the brain transitions from conscious to subliminal percep-
zione. Neuroscience, 411, 280–290.
Morone, F., Del Ferraro, G., & Makse, H. UN. (2019). The k-core as a
predictor of structural collapse in mutualistic ecosystems. Nature
Physics, 15, 95–102.
Morone, F., & Makse, H. UN. (2015). Influence maximization in com-
plex networks through optimal percolation. Nature, 524, 65–68.
Nachev, P., Kennard, C., & Husain, M. (2008). Functional role of
the supplementary and pre-supplementary motor areas. Nature
Reviews Neuroscience, 9, 856–869.
Newman, S. D., Just, M. A., Keller, T. A., Roth, J., & Carpenter, P. UN.
(2003). Differential effects of syntactic and semantic processing
on the subregions of Broca’s area. Cognitive Brain Research, 16,
297–307.
Nixon, P., Lazarova, J., Hodinott-Hill, I., Gough, P., & Passingham,
R. (2004). The inferior frontal gyrus and phonological processing:
An investigation using rTMS. Journal of Cognitive Neuroscience,
16, 289–300.
Pittel, B., Spencer, J. H., & Wormald, N. C. (1996). Sudden emer-
gence of a giant k-core in a random graph. Journal of Combina-
torial Theory, Series B, 67, 111–151.
Ramsey, N., Sommer, I., Rutten, G., & Kahn, R. (2001). Combined
analysis of language tasks in fMRI improves assessment of hemi-
spheric dominance for language functions in individual subjects.
NeuroImage, 13, 719–733.
Rosenberger, L., Zeck, J., Berl, M., Moore, E., Ritzl, E., Shamim,
S., . . . Gaillard, W. D. (2009). Interhemispheric and intrahemi-
spheric language reorganization in complex partial epilepsy.
Neurology, 72, 1830–1836.
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52,
1059–1069.
Seidman, S. B. (1983). Network structure and minimum degree.
Social Networks, 5, 269–287.
Tate, M. C., Herbet, G., Moritz-Gasser, S., Tate, J. E., & Duffau, H.
(2014). Probabilistic map of critical functional regions of the hu-
man cerebral cortex: Broca’s area revisited. Brain, 137, 2773–2782.
Tombari, D., Loubinoux, I., Pariente, J., Gerdelat, A., Albucher, J.-F.,
Tardy, J., . . . Chollet, F. (2004). A longitudinal fMRI study: In re-
covering and then in clinically stable sub-cortical stroke patients.
NeuroImage, 23, 827–839.
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organi-
zation of the human connectome. Journal of Neuroscience, 31,
15775–15786.
Network Neuroscience
153
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Core language brain network
Wang, L., Chen, D., Yang, X., Olson, J. J., Gopinath, K., Fan, T.,
& Mao, H. (2013). Group independent component analysis and
functional MRI examination of changes in language areas as-
sociated with brain tumors at different locations. PLoS ONE, 8,
e59657.
Wernicke, C. (1970). The aphasic symptom-complex: A psycholog-
ical study on an anatomical basis. Archives of Neurology, 22,
280–282.
Xiong,
J., Rao, S.,
Jerabek, P., Zamarripa, F., Woldorff, M.,
Lancaster, J., & Fox, P. T. (2000). Intersubject variability in corti-
cal activations dung a complex language task. NeuroImage, 12,
326–339.
Zhou, S., & Mondragón, R.
(2004). The rich-club phenom-
enon in the internet topology. IEEE Communications Letters, 8(3),
180–182.
J.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
4
1
1
3
4
1
8
6
6
6
9
2
N
e
N
_
UN
_
0
0
1
1
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network Neuroscience
154