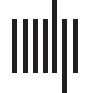RESEARCH
Distance-dependent consensus thresholds
for generating group-representative
structural brain networks
Richard F. Betzel1,2,3,4, Alessandra Griffa5, Patric Hagmann6, and Bratislav Miši´c7
1Department of Psychological and Brain Sciences, Indiana University, Bloomington, IN, USA
2Cognitive Science Program, Indiana University, Bloomington, IN, USA
3Program in Neuroscience, Indiana University, Bloomington, IN, USA
4Network Science Institute, Indiana University, Bloomington, IN, USA
5Dutch Connectome Lab, Department of Complex Trait Genetics, Center for Neurogenomics and Cognitive
Research, Amsterdam Neuroscience, VU University, Amsterdam, The Netherlands
6Lausanne University Hospital and University of Lausanne (CHUV-UNIL), Lausanne, Svizzera
7Montréal Neurological Institute, McGill University, Montréal, Quebec, Canada
Keywords: Complex networks, Wiring cost, Connectome, Group-representative, Consensus
ABSTRACT
Large-scale structural brain networks encode white matter connectivity patterns among
distributed brain areas. These connection patterns are believed to support cognitive processes
E, when compromised, can lead to neurocognitive deficits and maladaptive behavior.
A powerful approach for studying the organizing principles of brain networks is to construct
group-representative networks from multisubject cohorts. Doing so amplifies signal to noise
ratios and provides a clearer picture of brain network organization. Here, we show that
current approaches for generating sparse group-representative networks overestimate the
proportion of short-range connections present in a network and, as a result, fail to match
subject-level networks along a wide range of network statistics. We present an alternative
approach that preserves the connection-length distribution of individual subjects. We have
used this method in previous papers to generate group-representative networks, though to
date its performance has not been appropriately benchmarked and compared against other
metodi. As a result of this simple modification, the networks generated using this approach
successfully recapitulate subject-level properties, outperforming similar approaches by better
preserving features that promote integrative brain function rather than segregative. IL
method developed here holds promise for future studies investigating basic organizational
principles and features of large-scale structural brain networks.
AUTHOR SUMMARY
Sparse structural connectivity data from many subjects can be succinctly represented using
appropriate averaging procedures. We show, Tuttavia, that several popular procedures for
doing so generate group-averaged networks with statistics that are dissimilar from the
subject-level networks they are intended to represent. These dissimilarities, we argue, arise
from the over- and underexpression of short-range and long-distance connections,
rispettivamente, in the group-averaged matrix. We present a distance-dependent thresholding
procedure that preserves connection length distributions and consequently better matches
subject-level networks and their statistics. These findings inform data-driven exploratory
analyses of connectomes.
a n o p e n a c c e s s
j o u r n a l
Citation: Betzel, R. F., Griffa, A.,
Hagmann, P., & Miši ´c, B. (2019).
Distance-dependent consensus
thresholds for generating
group-representative structural brain
networks. Network Neuroscience, 3(2),
475–496. https://doi.org/10.1162/
netn_a_00075
DOI:
https://doi.org/10.1162/netn_a_00075
Supporting Information:
https://doi.org/10.1162/netn_a_00075
https://www.richardfbetzel.com/code/
Received: 15 settembre 2018
Accepted: 3 Dicembre 2018
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Richard Betzel
rbetzel@indiana.edu
Bratislav Miši ´c
bratislav.misic@mcgill.ca
Handling Editor:
Alex Fornito
Copyright: © 2018
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
INTRODUCTION
The human brain is a network composed of neural elements—neurons, populations, E
areas—interconnected to one another via synapses, axonal projections, and myelinated fiber
tracts, depending on the scale considered (Sporns, Tononi, & Kötter, 2005). These connections
shape neural elements’ patterns of input and output and play an important role in determining
any given element’s functional properties (Passingham, Stephan, & Kötter, 2002). By modeling
neural elements and their connections as the nodes and edges of a graph, we can quantify
with summary statistcs the network organization of brains and shed light on their function in
health, disease, and development (Sporns, Tononi, & Edelman, 2000).
Though considerable effort has been expended to better understand how different aspects of
brain network architecture vary across individuals (Yeh et al., 2016) and covary with behavioral
and clinical traits (Gollo et al., 2018; Miši´c & Sporns, 2016), studying group-representative
brain networks has also proven profitable for understanding the network organization and
properties of a typical or average brain (Hagmann et al., 2008; van den Heuvel & Sporns,
2011). It is often the case that group-representative networks are generated by aggregating
network data from many subjects while preserving those properties that are consistently ex-
pressed at the subject level (de Reus & van den Heuvel, 2013; Roberts, Perry, Roberts, Mitchell,
& Breakspear, 2017; Zalesky et al., 2016). This approach, when performed carefully, can the-
oretically enhance signal while suppressing noise and artifacts, affording a clearer view of the
brain’s network organization.
Most methods for constructing group-representative networks are variants of “consensus-
based thresholding.” That is, a sparse group network is generated by specifying a threshold
whose value ranges between 0 E 1, and retaining connections that are observed in at least
some fraction of subjects, τ. The retained connections are usually associated with a weight
while all others are set to 0 (Roberts et al., 2017) (Figures 1A, 1B). In almost every application,
the same consensus threshold is applied uniformly over all possible connections. This so-
called uniform consensus-based thresholding is common and group-representative networks
generated using this approach appear frequently in the network neuroscience literature. È
critical to note, Tuttavia, that the “correct” threshold is generally unknown and is often selected
according to heuristics. Inoltre, the very act of thresholding can introduce biases and cloud
subsequent interpretations of network organization and statistics (Garrison, Scheinost, Finn,
Shen, & Constable, 2015).
Group-representative networks are intended to serve as exemplars by preserving features
consistently expressed at the level of individual subjects while reducing the level of noise and
uncertainty. Among the most salient features of subject-level structural brain networks is the
dependence of their topological features on their spatial embedding (Stiso & Bassett, 2018).
Both the probability that two brain areas are connected and the weight of that connection,
should it exist, decay monotonically with interareal distance. This effect has been reported
in human structural networks reconstructed from diffusion MRI with tractography algorithms
(Betzel & Bassett, 2018; Roberts et al., 2016; Samu, Seth, & Nowotny, 2014), as well as net-
works reconstructed using invasive methods, such as tract-tracing (Ercsey-Ravasz et al., 2013;
Horvát et al., 2016), suggesting that these dependencies are not simply artifacts of any specific
network construction approach, but an evolutionarily conserved feature of large-scale brain
networks (van den Heuvel, Bullmore, & Sporns, 2016).
The preference for strong, short-range connections can be explained parsimoniously by
cost-reduction mechanisms (Bullmore & Sporns, 2012; Vértes et al., 2012). Intuitively, longer
Group-representative network:
A single network intended to
represent the connectivity data
of many subjects.
Consensus thresholding:
A procedure for generating
group-representative networks by
retaining only those connections
present in a given fraction
(the threshold) of subjects.
Network Neuroscience
476
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
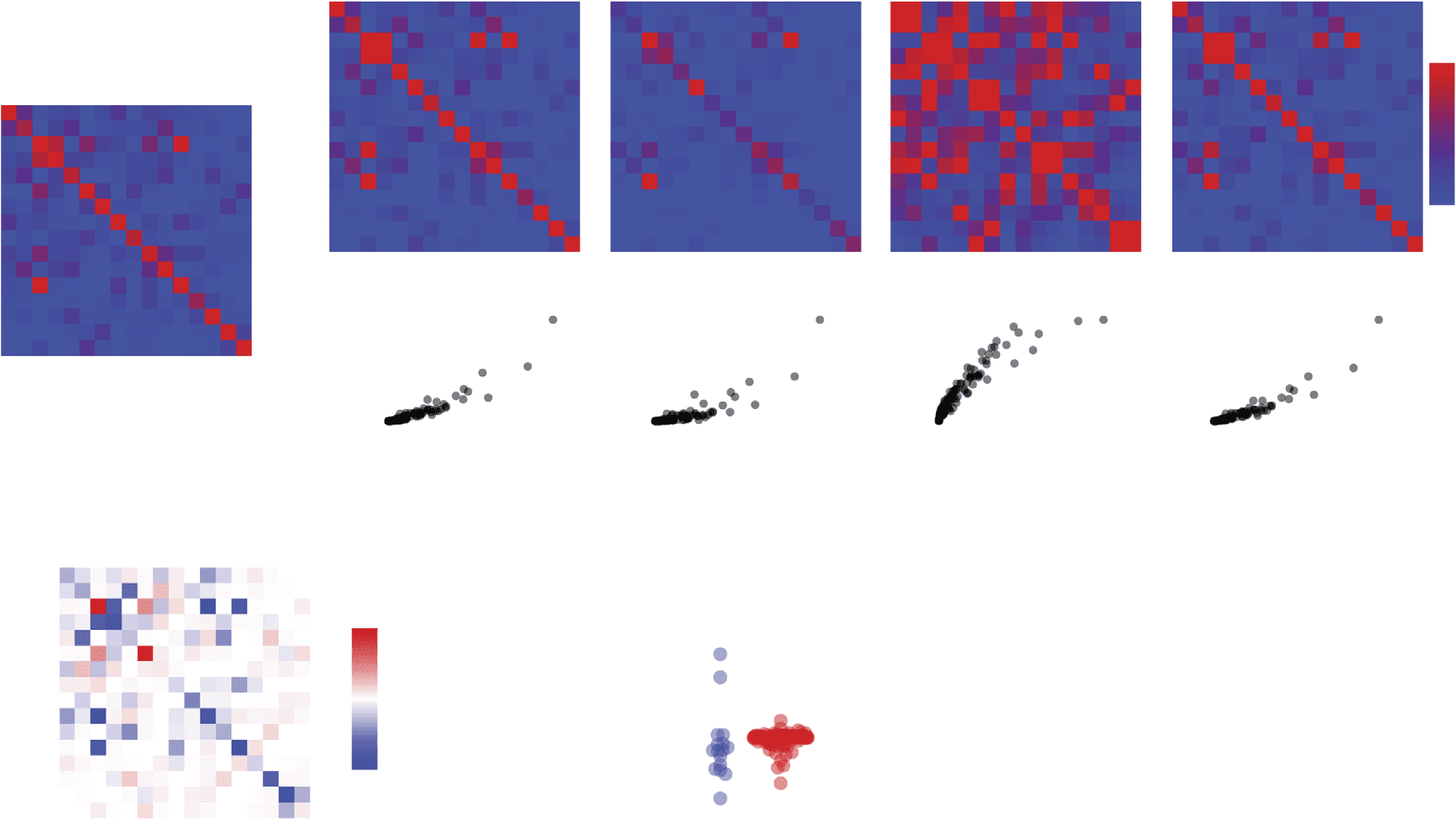
Figura 1. Construction and superficial comparison of group-representative matrices. Group-representative connectivity matrices summarize
subject-level network data (UN) by retaining features that are consistently expressed across subjects. In most applications the features of interest
are the edges between brain areas and their weights. The most straightforward approach for generating a group-representative matrix involves
first constructing a consensus matrix (B), whose elements denote the fraction of all subjects in which edges are expressed. Group-representative
matrices can be estimated by retaining all connections expressed in at least τ subjects and populating those connections with weights. Though
this approach is common, it suffers from a number of shortcomings. Generalmente, because the probability of observing any given short-range
connection is greater than the probability of observing a long-range connections, short-range connections also appear more consistently
across subjects. Di conseguenza, imposing a uniform consensus-based threshold across all elements of the consensus matrix will result in a group-
representative matrix in which short-range connections are expressed with much greater frequency than any single subject (C). To circumvent
this issue, we present a simple alternative approach. Briefly, this involves dividing all connections into m bins according to their length
E, within each bin, retaining the connection that is most frequently expressed. This distance-dependent consensus-based thresholding
approach results in networks with almost the exact same edge length distribution as the typical subject. We also show differences in the
group-representative matrices generated using the distance-dependent and uniform consensus-based threshold (here, we choose τ for the
uniform method such the resulting matrix has a number of connections equal to that of the average subject). (D) Connections present only in
the uniform method are depicted in blue and those present only in the distance-depdendent method are shown in red. (E) We plot these same
method-specific connections on the brain, and color them according to their length (in millimeters). (F) Generalmente, the connections unique to
the uniform method are short range (blue curve) while those unique to the distance-dependent method include long-distance connections.
connections are more costly; they require additional material to form and extra energy for
sustained use compared with short-range connections. As a consequence, nervous systems
have evolved to favor shorter, low-cost connections. Despite this, brain networks still exhibit
some long-distance connections (Betzel, Medaglia, & Bassett, 2018). It is generally understood
that longer connections play critical functional roles in order to offset their cost, though their
precise function is still not fully understood.
Network Neuroscience
477
Distance-dependent structural brain networks
Whatever their precise functional role, long-distance connections are arguably one of the
most important subject-level features to preserve in any group-representative network. They
play an important role in increasing shortest-path efficiency (Kaiser & Hilgetag, 2006) and en-
gender diverse network dynamics and information processing (Betzel & Bassett, 2018). How-
ever, uniform consensus-based thresholding can produce networks that vastly underestimate
the number of observed long-distance connections. This bias emerges because the consen-
sus of connections across subjects is, itself, distance-dependent, with short-range connections
appearing more consistent than longer-range connections. As a consequence, for a given con-
sensus threshold the distribution of suprathreshold connections will always favor short-range
connections at
È, group-representative
networks generated using a uniform consensus-based thresholding procedure will exhibit
more short-range connections and fewer long-distance connections than the typical subject
(Figures 1C–F). Because long-distance connections are responsible for driving certain network
statistics, these group-representative networks will also fail to match subject-level networks in
terms of those metrics.
long-distance connections. Quello
the expense of
Here, we present an alternative method for constructing group-representative networks.
Our approach, which we haved used in previous papers (Betzel & Bassett, 2018; Betzel, Gu,
Medaglia, Pasqualetti, & Bassett, 2016; Betzel et al., 2017; Miši´c et al., 2015) but never ap-
propriately benchmarked, builds upon the consensus-based thresholding framework; Piuttosto
than imposing a threshold uniformly over all connections, we allow our threshold to vary as a
function of distance, retaining the most consistent connections conditional upon their length.
In contrast to existing approaches, we derive the distance-dependent threshold nonparamet-
rically so as to match the pooled edge length distribution of subject-level data. We compare
networks generated using this distance-dependent thresholding procedure with those gener-
ated using more traditional methods and show that, across a wide range of network statistics
and comparative measures, networks generated using the distance-dependent approach out-
perform others. The distance-dependent procedure successfully recapitulates many of the im-
portant organizational features of subject-level networks and demonstrates promise for future
exploratory studies of structural brain networks.
RESULTS
In this section we compare four different approaches for generating group-representative struc-
tural connectivity networks.
Connections are retained if they appear in at least one subject. We refer to this as the
“Simple” method.
Connections are retained if they appear in at least 50% of subjects. We refer to this as
the “τ = 0.5” method.
Connections are retained if they appear in at least τ
Avg is the con-
sensus threshold that results in a binary density equal to that of the typical subject. Noi
refer to this as the “τ = Avg” method. Note: We calculate this threshold separately for
inter-/intra-hemispheric connections.
Connections are retained using a distance-dependent consensus threshold . The resulting
network preserves, approximately, the edge length distribution of the typical subject. Noi
refer to this as the “Dist.” method. As with the “τ = Avg” method, the distance-dependent
threshold is introduced separately for inter-/intra-hemispheric connections.
Avg subjects, where τ
478
Distance-dependent consensus
thresholding:
Same as consensus thresholding, Ma
where the threshold for connection
retention varies with distance.
Network Neuroscience
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
Structural connectivity:
A map of physical connections
among neural elements
(per esempio., axonal projections among
neurons/populations, white matter
fiber pathways among brain
areas/parcels).
This section is further divided into four subsections. In the first two subsections, we compare
statistics of group-representative networks with those of individual subjects. In the next subsec-
zione, we characterize connectivity patterns of the group-representative matrices with respect
to cognitive systems and discuss implications for our understanding of brain function. Nel
final subsection, we characterize how hubs are redistributed depending upon the approach
used for generating group-representative brain networks.
Throughout this section, we report results of analyses using the high-resolution parcella-
tion of the Lausanne dataset (N = 1, 000 nodes), where white matter fiber tracts are recon-
structed from diffusion spectrum imaging data using deterministic streamline tractography (Vedere
the Materials and Methods section for processing details). These results are representative of
our findings using coarser parcellations. Those additional results are included in the Support-
ing Information, Figure S1 and Figure S2 (Betzel, Griffa, Hagmann, & Miši´c, 2019). Finalmente, we
also compare the results of the distance-dependent consensus method with a weight-based
thresholding approach in which we generate a new group-representative network by imposing
a weight threshold on the “Simple” group-representative network. These results of this analysis
are included show in the supplement (Figure S3; Betzel, Griffa, et al., 2019).
Uniform and Distance-Dependent Consensus-Based Thresholding Generate Systematically
Different Networks
The presence and weights of edges in structural connectivity networks exhibit spatial depen-
dencies because of cost-reduction principles and reconstruction artifacts that cause short-range
connections to be more consistently expressed across subjects. As a consequence, procedures
for generating group-representative networks that retain connections using uniform consensus
thresholds will necessarily overestimate the number of short-range connections in a network.
In the following subsections we explore the implications of these biases in greater detail.
Here, we simply show that uniform consensus thresholds generate group-representative net-
works with different spatial statistics than those generated using distance-dependent consensus
thresholds, wherein the threshold for edge retention varies as a function of Euclidean distance.
In Figure 1D, we show an adjacency matrix containing connections that are present in either
the uniform or distance-dependent group-representative matrix but not both. Alongside this
panel and in Figure 1E, we plot these same connections in anatomical space and color connec-
tions according to their lengths, with long/short connections appearing bright red/dark blue.
In the left and right subpanel we show connections that are present in distance-dependent
method but not the uniform method and vice versa. Note that the connections shown in
the left subpanel tend to be long (red), indicating that the distance-dependent method retains
long-distance connections that are not preserved in the uniform method. Conversely, the con-
nections in the right subpanel tend to be short (blue), indicating that the uniform method
retains short-range connections not observed in distance-dependent method. These trends can
be summarized by examining the distribution of connections present in one method but not the
other. In Figure 1F we show that, as expected, connections retained exclusively by the uniform
method are sharply peaked around 25 mm, whereas the connections retained by the distance-
dependent method are more broadly distributed and include many long-distance connections.
We note that the τ = 0.5 and distance-dependent methods have equal binary density, making
a direct comparison of those matrices appropriate.
These observations about differences in the connection length distributions of group-
representative brain networks, though superficial, have important practical consequences for
the structural properties of those networks. An overexpression of short-range connections could
Network Neuroscience
479
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
result in excessively cohesive brain network modules (Roberts et al., 2016; Samu et al., 2014),
missing out on potentially rich intermodular connectivity patterns (Betzel, Medaglia, et al.,
2018). Conversely, group-representative networks that overexpress long-distance connections
may lack segregative network properties such as local clustering (Sporns & Zwi, 2004). These
observations are in line with the fact that, in general, network properties are not independent
of one another, and variation in one property has implications for others (Rubinov, 2016).
Here, and throughout this paper, we argue that mischaracterizations of edge length distribu-
tions have profound implications for the spectrum of network properties that are exhibited by
group-representative networks and whether those properties are in line with those of individual
subjects’ networks.
Consistency-Based Thresholding Does Not Preserve Subject-Level Network Statistics
There are many criteria by which group-representative connectivity matrices could be evalu-
ated and judged. Arguably among the most important is their ability to recover and recapitulate
the topological properties of the subject-level data that they supposedly represent. In this sec-
tion we compare four approaches for generating group-representative networks according to
how well each matches individual subjects in terms of an ensemble of network statistics. Noi
focus specifically on local statistics such as degree, strength, clustering coefficient, between-
ness centrality, and edge length distribution and global statistics such as number of binary
connections, total weight, mean clustering, topological efficiency, mean path length, modu-
larity, diameter, and degree assortativity.
We divided network meausures into two categories based on whether they were defined lo-
cally or globally. We compared subject-level and group-representative local measures, questo è,
those defined at the level of individual nodes, using Kolmogorov-Smirnov (KS) tests. The KS test
statistic measures the maximum distance between two cumulative distributions, and therefore
smaller values indicate closer correspondence. In Figures 2A–E, we overlay cumulative dis-
tributions of degree, strength, clustering coefficient, betweenness centrality, and edge length
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
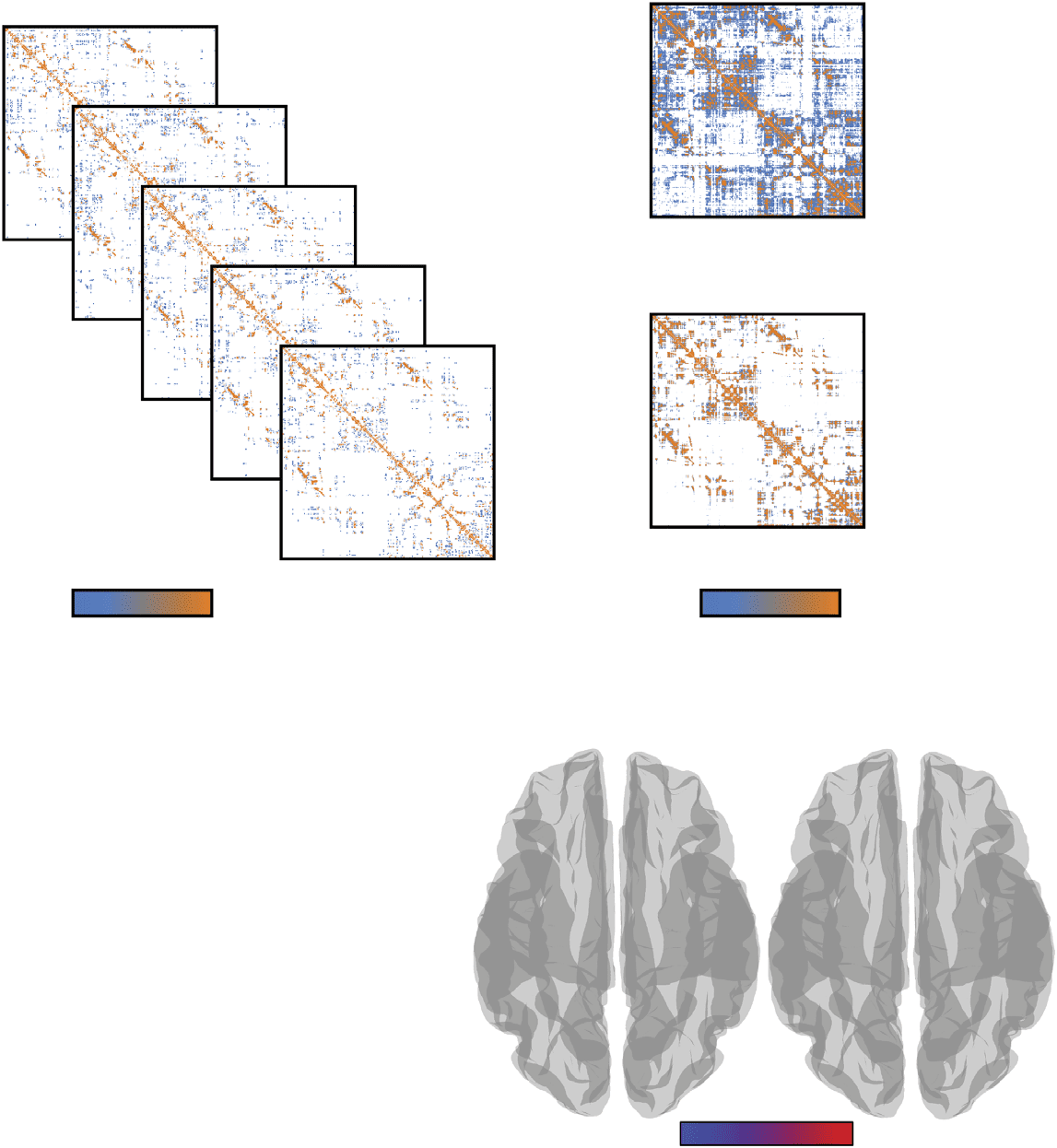
Figura 2. Comparing distributions across methods. We show cumulative distributions for (UN)
degree, (B) strength, (C) clustering coefficient, (D) betweenness centrality, E (E) connection length.
Subject-level data are shown in gray. Superimposed on those distributions are curves associated
with the four methods that we tested (in color). In panels (F)–(J) we show Kolmogorov-Smirnov (KS)
statistics for each network measure, which compare cumulative distribution curves of methods with
individual subjects.
Network Neuroscience
480
Distance-dependent structural brain networks
for each of the four group-representative networks on top of the cumulative distributions for
individual subjects. The KS statistics comparing these distributions are plotted in Figures 2F–J.
For all five measures, we found that the distance-dependent consensus-based thresholding
approach outperformed the other three methods, questo è, smaller KS statistics (P < 0.05; Bonfer-
roni corrected). These findings indicate that the distance-dependent method better preserved
multiple node- and edge-level measures than the other methods, suggesting that network statis-
tics computed on the other group-representative networks may be misleading, in that they are
not necessarily representative of the typical subject.
We performed similar comparisons of the global network measures. Here, rather than com-
paring distributions using the KS test, we z-scored the measures computed on the group-
representative networks against the corresponding subject-level distributions. A z-score close
to 0 implied that the group-representative network was close to the mean subject-level value
for a given network measure. In Figures 3A–H, we show binary and weighted analogs of the
total number of connections in the network, mean clustering, efficiency, and modularity. We
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
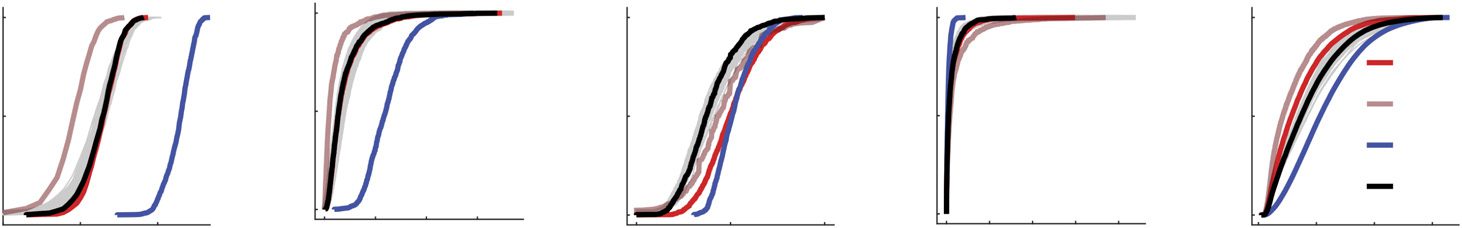
Figure 3. Comparing scalar network statistics. Here, we compare the performances of four differ-
ent methods of group-representative brain networks to those of individual subjects. (A) Each bar rep-
resents the total number of binary connections for single subjects (gray), a uniform method with
approximately the same number of connections as the average subject (bright red), a uniform method
with a consensus threshold of τ = 0.5 (dark red), a “Simple” method that retains a connection if it
is observed in even one subject (blue), and the distance-dependent method (black). Panels (B)–(D)
show similar plots but for mean clustering, efficiency, and modularity. Panels (E)–(H) depict those
same measures, but computed over weighted analogs of the binary networks. (I) For each measure
shown (along with several others), we identified the method that was closest to that of the average
across all subjects. In general, we find that the distance-dependent method consistently outperforms
or performs comparably to the other tested methods, achieving rank 1 or 2 across all metrics.
Network Neuroscience
481
Distance-dependent structural brain networks
found similar results when comparing network diameter, assortativity, and mean path length
(Figures 3I, 3J).
As with the local network measures, these findings suggest that decisions about how to
generate a group-representative connectivity matrix have implications for its topological or-
ganization. Importantly, the most popular approach—uniform consensus-based thresholding—
preserves a greater number of short-range connections compared with the typical subject and,
as a result, exhibits topological properties that are inconsistent with those exhibited by that
subject.
Implications for Structure-Function Relationships
In the previous section, we compared group-representative networks in terms of how well they
recapitulated topological properties of subject-level networks. Another important dimension
along which group-representative networks can be compared is in terms of how their struc-
tural connections map onto the brain’s functional and cognitive systems. Here, we explore this
structure-function relationship by averaging structural connectivity weights among previously
described cognitive systems—resting-state networks (RSNs; Schaefer et al., 2017). Resting-state
networks are composed of brain areas with similar functional connectivity profiles and reca-
pitulate the collections of brain areas that coactivate during cognitively demanding tasks. We
compute inter- and intra-RSN connectivity density for each of the four group-representative
methods, in each case generating a system-by-system connection density matrix (Figure 4B).
We also do the same for individual subjects, averaging over these subject-level matrices to ob-
tain a single matrix whose elements represent the mean intersystem connection density across
all subjects (Figure 4A). To compare group and subject matrices, we compute the Pearson
correlation of their upper triangle + diagonal elements.
In general, we find that each group-representative matrix is positively correlated with the
subject-level matrix, indicating that, overall, system-to-system connectivity at the subject level
is preserved at the group level by all methods. Nonetheless, there is considerable variability
across group-representative methods in terms of correlation magnitude and deviation from
the identity line (Figure 4C). For instance, the “Simple” method exhibited a correlation of
r ≈ 0.92 but massively overestimated the amplitude of connection densities. Similarly, the
uniform method with a threshold of τ = 0.5 exhibited a much weaker correlation of r = 0.85.
The remaining two methods, on the other hand, exhibited much stronger correlations with
magnitudes in excess of r ≈ 0.93 and approximately the same spread of data points around
the identity line.
The two best-performing methods were the uniform method for which we selected a thresh-
old resulting in the same density as the average subject and the distance-dependent method.
We compared these two methods in greater detail to better understand the implications of using
one method versus the other. First, we computed the difference in inter-RSN connectivity den-
sity (Figure 4D). We found that there were subtle yet systematic differences. In particular, we
found that the distance-dependent method exhibited much weaker within-RSN density com-
pared with that of the uniform method while also exhibiting stronger between-RSN connection
density (p < 0.05, permutation test; Figures 4E, 4F).
These findings have important implications for the analysis and interpretation of brain net-
work data. This is especially true for studies that aim to link features of structural and functional
brain networks to one another. Past studies using group-representative data constructed us-
ing a uniform consensus threshold may fail to match the specificity of subject-level networks,
Network Neuroscience
482
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Distance-dependent structural brain networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
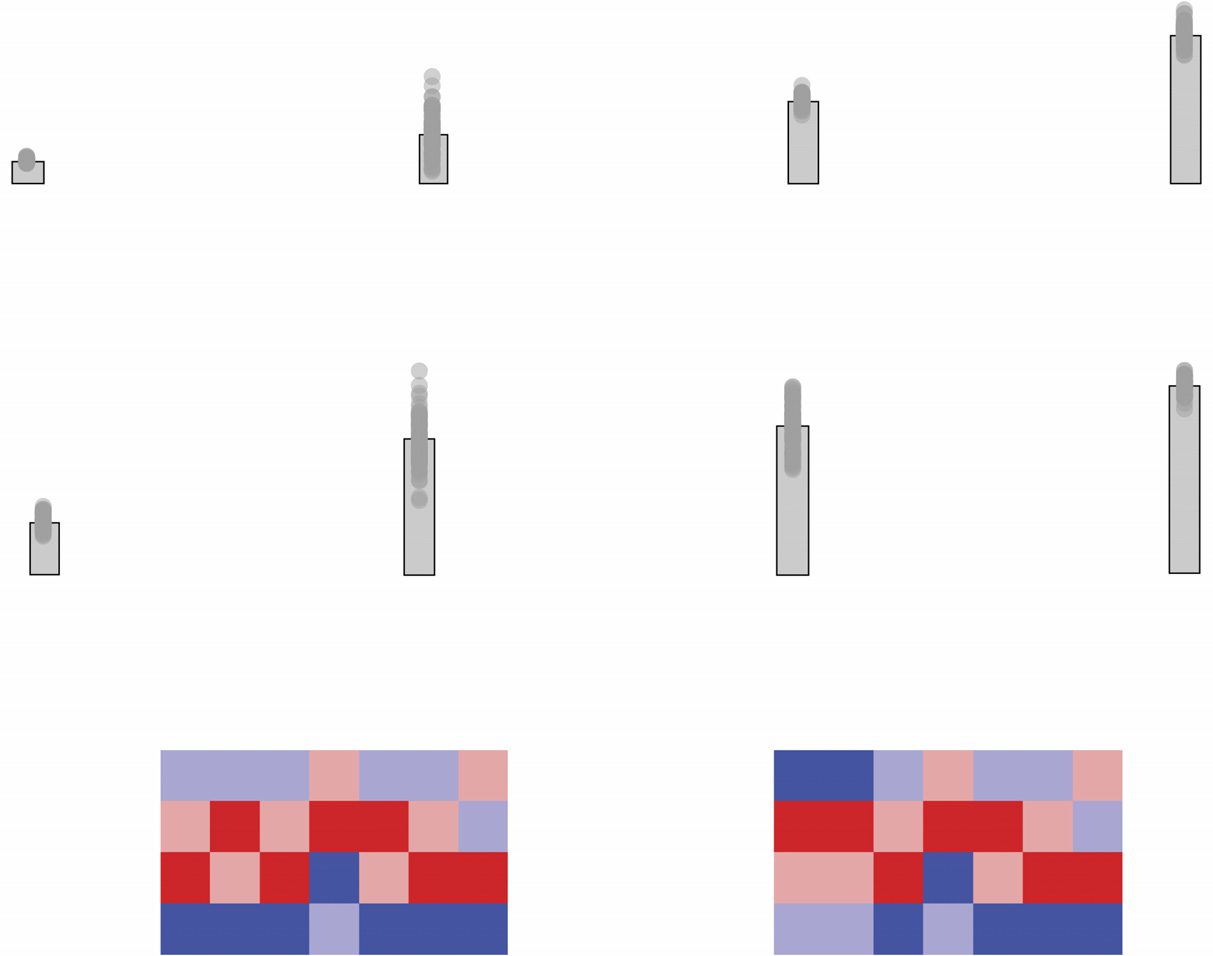
Figure 4. Comparing within-/between-RSN connectivity patterns. We compared different group-representative networks in terms of connec-
tion densities within and between canonical brain systems taken from Schaefer et al. (2017). (A) Inter-RSN connection density of the typical
subject. (B) Inter-RSN connection densities for four different group-representative networks: (from left to right) uniform consensus method with
same density as subjects, uniform consensus method with threshold set at τ = 0.5, “Simple” method, and distance-dependent method. (C) We
show the correlation patterns of inter-RSN densities for each method (y-axis) with that of the subject average (x-axis). Of the methods com-
pared here, the distance-dependent and the uniform method with same density as the typical subject performed the best. We compare these
methods so as to better understand their differences. (D) Difference in inter-RSN connection density between distance-dependent and τ = 0.5
threshold methods. Blue colors indicate that connection density is greater in uniform method while red density indicates that connection
density is greater in distance-dependent method. (E) We find that, on average, the uniform method results in weaker within-RSN density than
the distance-dependent method, while the distance-dependent method has greater between-RSN density. (F) We show the observed difference
in within- and between-RSN density and compare it against a null method. Here, we show the null distribution (blue) and the observed value
(red). The null distribution was constructed by independently and randomly permuting rows/columns of each original connectivity matrix and
reaggregating according to the RSN system labels. Then we compute the mean difference of within-/between-RSN densities.
while the simple averaging procedure may overestimate the weights of short-range connections
(van den Heuvel & Sporns, 2013). These failings could, in principle, lead to mischaracteriza-
tions of structure-function associations.
More importantly, these findings suggest that differences in the construction method for
group-representative networks can result
in networks that emphasize either segregative
features—that is, stronger within-RSN connection densities, as expressed by the uniform
method—or integrative features—that is, stronger between-RSN connection densities as ex-
pressed by the distance-dependent method. The balance between information segregation
and integration is thought to be an important organizational principle responsible for shap-
ing brain network topology (Cohen & D’Esposito, 2016; Deco, Tononi, Boly, & Kringelbach,
2015; Sporns, 2013). Our findings indicate that different group-representative methods differ-
entially emphasize these characteristics, indicating that a user’s seemingly arbitrary choice in
method can have implications for measures made on a network.
Network Neuroscience
483
Distance-dependent structural brain networks
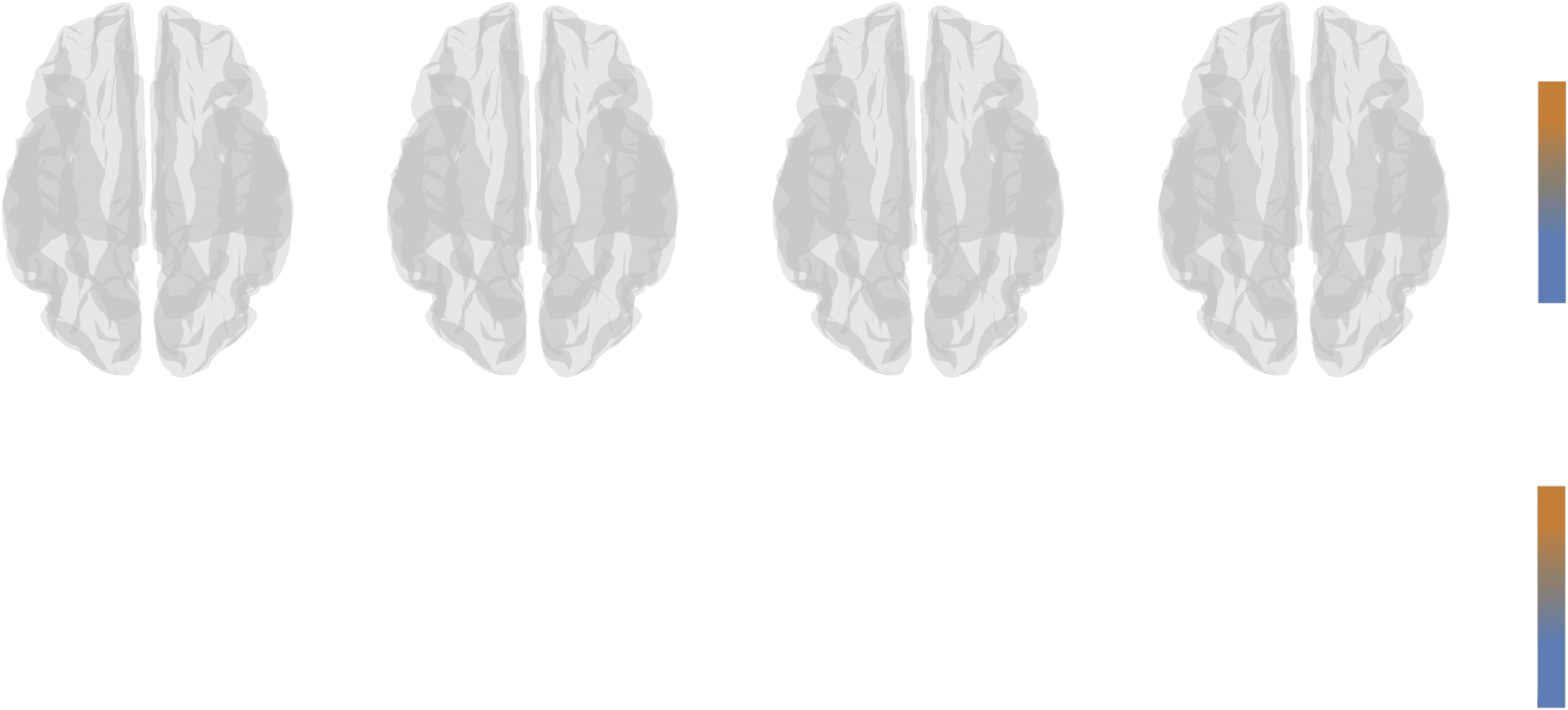
Hub (Re)distribution
A third means of comparing group-representative networks against one another is to measure
the redistribution of hub areas, that is, assessing changes in the locations of “central” brain
areas as a result of choosing one group method versus another. Here, we compare the spatial
distribution of betweenness centrality, node degree, clustering coefficient, and participation
coefficient under uniform and distance-dependent methods. To ensure that comparisons are
as fair as possible, we rank-transformed all measures prior to comparison.
In general, we found widespread and hemispherically symmetric redistribution of hub
regions. In the case of betweenness centrality (Figure 5A), we found that under the distance-
dependent method, areas associated with cognitive control are increasingly central, while
areas in the somatomotor system become less central (p < 0.05; corrected for multiple com-
parisons by controlling false discovery rate at 5%; Figure 5E). In terms of degree (Figure 5B), we
find that control and limbic systems make a greater number of connections, while dorsal at-
tention, salience/ventral attention, somatomotor, and visual systems exhibit fewer connections
(Figure 5F). For clustering coefficient (Figure 5C), we find that components of default mode and
motor systems are more clustered while multiple components of the control network are less
clustered (Figure 5G). Finally, in terms of participation coefficient (Figure 5D), we find that the
dorsal attention and visual systems exhibit greater participation, whereas somatomotor and
other visual systems exhibit decreased participation (Figure 5H).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Comparing spatial distribution of hubs. We compare four measures of hubness: (A) betweenness centrality, (B) degree, (C) clustering
coefficient, and (D) participation coefficient. Rather than compare raw values, which can fluctuate because of small differences in global
network properties like total number of connections or weight, we compare ranked values of each measure and observe whether a node’s
rank is smaller/greater under the distance-dependent or uniform method. Orange-colored nodes indicate that a node’s value is greater under
the distance-dependent method than it is under the uniform method. Blue-colored nodes indicate the opposite. We then aggregated node-level
differences in ranked measures by cognitive systems and compared the mean system-level values with those obtained under a null method.
In panels (E)–(H) we show the z-scored system means. In general, large-magnitude z-scores indicate bigger greater system-level differences
between the two methods.
Network Neuroscience
484
Distance-dependent structural brain networks
These findings have important implications for our understanding of brain function. Hubs
and central brain areas are believed to be important for controlling interareal communciation
and regulating the flow of information within and between brain network modules. Indeed,
the designation of an area as a “hub” has been important for hypothesis generation and has
also played an important confirmatory role in other studies. Our findings suggest that these
definitions are, at least to some extent, dependent upon the method used to generate a group-
representative network. Moreover, some of the most salient differences between methods ap-
pear localized to specific cognitive systems, which has additional implications for how we
interpret findings related to hubs and brain function.
DISCUSSION
In this paper we flesh out the details of a new method for generating group-representative
brain networks that outperforms more conventional methods in terms of preserving subject-
level statistics. Specifically, we show that this method better preserves local and global network
statistics, that its structure-function relationships are more consistent with those of individual
subjects, and that it gives rise to a different intuition of where highly central hub regions are
located in the brain.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural Networks Need Long-Distance Connections
Here, we found that compared with uniform consensus-based thresholding, a distance-
dependent threshold preserves to a greater degree the connection length distributions observed
in individual subjects. We argue that this difference in connection length distributions has both
practical (i.e., measurable) and theoretical consequences. Practically, we show that expressing
fewer long-distance connections results in networks that are more clustered and, as a result of
increased rate of triadic closure, more modular than that of the typical subject (Henderson &
Robinson, 2013). Similarly, lacking long-distance shortcuts results in networks that are less effi-
cient and that possess longer characteristic path length than the average subject (Henderson
& Robinson, 2013; Samu et al., 2014). Overall, the uniform method results in networks that
emphasize segregative traits at the expense of those that support integration of information
(Sporns, 2013). This is confirmed further when we compared the intersystem connection den-
sities of the distance and uniform methods, observing that within-community density was less,
on average, using the distance method compared with the uniform method. Additionally, the
differences in the features preserved by each method contribute to shaping the spatial distri-
bution of hubs across the brain. Overall, these findings suggest that the principal advantage
of the distance-dependent method is that it better preserves network features that emphasize
information integration.
The presence of long-distance connections, though useful in theory for brain network func-
tion, also presents a methodological challenge and their inclusion in network models demands
careful attention. For instance, recent studies have shown that in highly modular networks, the
addition of spurious long-distance intermodular connections induce larger changes in network
statistics like clustering, modularity, and efficiency compared with the addition of short-range,
intramodular connections (Zalesky et al., 2016). The results of our study, on the other hand,
suggest that by including long-distance connections, we better recapitulate the network proper-
ties of individual subjects. This discrepancy between these findings is a direct result of two dis-
tinct sets of assumptions: in Zalesky et al. (2016), the authors treat the “true” network to be
one generated using uniform consensus-based thresholding with τ = 0.5, resulting in a highly
modular network, whereas we treat the “true” network to be one with statistics similar to that
Network Neuroscience
485
Distance-dependent structural brain networks
Connectome:
A comprehensive network map of
connections among neural elements.
of the typical subject, which is less modular, less clustered, and more efficient. In general, the
organization of the ground-truth connectome remains unknown, and it is unclear which set of
assumptions is more appropriate. With advances in cellular-level reconstruction, it may soon
be possible to obtain a clearer picture of true structural connections, which could be used to in-
form out macroscale models of connectivity and resolve this debate (Briggman, Helmstaedter,
& Denk, 2011; Helmstaedter et al., 2013).
The Role of Group-Representative Network Analysis
In this study we focus on group-representative networks. Analysis of these group networks has
been and remains an important component of network neuroscience. In the case of nonhuman
datasets, group network analysis is almost always performed out of necessity. Invasive methods
like tract-tracing limit the number of experiments that can be performed on any one animal
brain. As a result, whole-brain networks are necessarily composites of many animals (Markov
et al., 2012; Noori et al., 2017; Oh et al., 2014). Human structural networks constructed from
diffusion MRI data using tractography algorithms are sensitive to scan parameters and prone
to false positives and negatives (Maier-Hein et al., 2016; Reveley et al., 2015). Analyses of
human networks, therefore, benefit from aggregation of multisubject cohorts into a group-
representative network, which serves to enhance signal while reducing the level of noise and
uncertainty. The resulting networks can be treated as exemplars and used to uncover key struc-
tural traits and organizing principles (Hagmann et al., 2008; van den Heuvel & Sporns, 2011),
as the basis for dynamic methods (Miši´c et al., 2015), and serve as a sort of “prior” for other
machine and statistical learning approaches (Rosenthal et al., 2018).
However, analysis and interpretation of group-level networks presume that those networks
are, in fact, representative of the typical subject. Group networks that violate this assumption
can contribute misleading or inaccurate insight into brain network organization and function.
We show here that group-representative networks constructed using a uniform consensus-
based threshold, which fail to preserve important spatial properties of subject-level brain net-
works, may be especially susceptible to such inaccuracies. Our work suggests that the uniform
method generates networks that tend to overestimate the cohesiveness of communities. In ad-
dition, the uniform method also presents a conflicting account of hub distributions throughout
the brain when compared with the distance-dependent method. Because analysis of group-
representative networks remains a powerful approach, understanding and accounting for their
limitations and biases should be investigated in future research.
Limitations
Here, we present a method for constructing group-representative networks, demonstrating that
this approach results in group networks that better preserve subject-level properties than ex-
isting approaches. Nonetheless, our study suffers from some limitations.
First, we make the overarching assumption that the long-distance connections observed in
single-subject networks (which we preserve in our group-representative network) are “real” and
not strictly artifacts of the tractography algorithm. This assumption is supported, first, by the fact
that long-distance connections are, in general, more challenging to reconstruct using common
tractography parameters. Completing long streamlines requires strong spatial coherence of the
diffusion field over distances greater than 150 mm, which is unlikely to occur in the presence of
high background noise (Jones, Knösche, & Turner, 2013). Second, long-distance connections,
typically, do not appear randomly distributed, but are clustered (Betzel & Bassett, 2018). That is,
if regions i and j are connected by a long-distance tract, it is likely that other regions in i’s spatial
Network Neuroscience
486
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Distance-dependent structural brain networks
neighborhood are connected to j and j’s spatially proximal neighbors (and vice versa). These
observations suggest that long-distance connections cannot easily be explained as errant “one-
off” reconstructions. Nonetheless, tractography has known shortcomings (Maier-Hein et al.,
2016; Reveley et al., 2015), and the verisimilitude surrounding long connections remains un-
clear. Advances in hardware, fiber reconstruction software (Pestilli, Yeatman, Rokem, Kay, &
Wandell, 2014), and detailed comparisons of tractography with tract-tracing data (Calabrese,
Badea, Cofer, Qi, & Johnson, 2015) will help future studies overcome these issues.
A second limitation concerns the network measures and metrics used to compare group-
representative networks to one another and to individual subjects. These measures were
selected because they emphasized network topology as well as its relationship to neuroscientif-
ically relevant metadata (i.e., cognitive functional systems). However, these measures are, first,
not necessarily an exhaustive list and it is unclear whether the distance-dependent method’s
performance would be better than other methods were we to select a different set of measures.
Second, network measures tend to be correlated with one another—for example, a network
with high efficiency will tend to have short path length. Therefore, the comparisons we made
were not necessarily independent of one another. Though we intentionally selected a wide
range of measures to help address these issues, our analyses could be extended in future work
to include an even broader range of measures and comparative metrics.
A third limitation concerns the applicability of the distance-dependent thresholding pro-
cedure (or any consensus-based thresholding procedure for that matter) to fully weighted
connectivity matrices, like those generated from probabilistic tractography or functional con-
nectivity data. One strategy to extend our approach is to first impose a threshold on subject-
level fully weighted matrices, which is fairly common in the analysis of the aforementioned
data types (Fallani, Latora, & Chavez, 2017; Gollo et al., 2018). With sparse subject-level data,
the consensus-based thresholding procedures can be carried out as reported here. Generating
consensus matrices that, themselves, are fully weighted remains an outstanding challenge.
In that case, consensus-based thresholding fails (all connections are present across all sub-
jects). Moreover, in the case of functional connectivity, which is usually estimated as a corre-
lation matrix, special care must be taken to preserve statistically defined transitive relationships
(Zalesky, Fornito, & Bullmore, 2012). Future work should be directed to more explicitly inves-
tigate consensus methods for full matrices.
Yet another limitation concerns the ability of modern tractography methods for reconstruct-
ing long-distance tracts. We show that traditional consensus-based thresholding methods prune
away long-distance streamlines in single subjects because the locations of those tracts are less
consistent across subjects than short-range tracts. Here, we interpret the reduction in long-
distance tracts (which are expressed in every subject) as evidence of a failing in consensus-
based thresholding procedures. Another interpretation, however, is that long-distance tracts
appear less consistent because they are spurious. We argue that this is likely not the case, as
long-distance connections (though inconsistent) nonetheless exhibit high levels of clustering
and structure (Betzel & Bassett, 2018), suggesting that they are distributed in a far from random
manner. This issue should be investigated in more detail in future work.
In this study, we generated group-representative networks by retaining connections with
the greatest consensus across subjects, that is, those that were expressed in a large fraction
of individuals. This ensures that group matrices preserve those connections most consistently
expressed across subjects rather than the connections with the strongest weights (another com-
mon procedure for denoising network data; Fallani et al., 2017; van Wijk, Stam, & Daffertshofer,
Network Neuroscience
487
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Distance-dependent structural brain networks
2010). Moreover, it is expected that connection weights vary over several orders of magnitude
(Buzsáki & Mizuseki, 2014). This provides some rationale for retaining connections with high
consensus across subject irrespective of their weight. In contrast, imposing a weight-based
threshold across connections effectively truncates a heavy-tailed distribution and restricts con-
nection weights to a narrower regime. Nonetheless, there may be scenarios in which weight-
based thresholding is more appropriate for constructing group-representative networks, for
example, when the noise level is exceptionally high and there is poor consensus across sub-
jects. Much additional work is needed to identify the situations where one or the other ap-
proach should be preferred.
A final limitation is the necessity that the subject-level matrices used to estimate the group-
representative network be sparse. Both the uniform and the distance-dependent methods rely
on the intuition that some connections are more common across individuals than others. For
some diffusion MRI and tractography algorithms—such as probabilistic tractography—this is
not always the case (Descoteaux, Deriche, Knosche, & Anwander, 2009). Nonetheless, it may
be possible to adapt the approaches used here with sparse deterministic tractography to the
probabilistic case by substituting connection probability measures for the consensus. Care
would have to be taken to deal with the potentially confounding geometric and spatial biases
(Roberts et al., 2016). Future work should investigate this in greater detail.
Conclusion
Overall, our findings suggest that care must be taken when studying and analyzing group-
representative networks. We presented an approach for limiting discrepancies between subject-
and group-level networks by adding a distance-dependence to the consensus threshold. This
approach will aid in future studies that seek to investigate general properties of structural brain
networks.
MATERIALS AND METHODS
Connectome Dataset
In this study we compared methods for constructing group-representative brain networks from
structural connectivity data. We carried out these comparisons using diffusion spectrum MRI
data parcellated into networks at three different organizational scales. Here, we describe those
processing steps in greater detail.
A total of 70 healthy participants (age 28.8 ± 9.1 years old, 43 males) were
MRI acquistion.
scanned on a 3T scanner with a 32-channel head coil (Magnetom TrioTim, Magnetom Prisma,
Siemens Medical, Germany). The session included (a) a magnetization-prepared rapid acqui-
sition gradient echo (MPRAGE) sequence (1 × 1 × 1.2 mm resolution, 240 × 257 × 160 voxels;
TR = 2,300 ms, TE = 2.98 ms, TI = 900 ms); (b) a diffusion spectrum imaging (DSI) sequence
(2.2 × 2.2 × 3 mm resolution; 96 × 96 × 34 voxels; TR = 6,100 ms, TE = 144 ms; q4half ac-
quisition with maximum b-value 8,000 s/mm2, one b0 volume). Informed written consent was
in accordance with institutional guidelines and the protocol was approved by the Ethics Com-
mittee of Clinical Research of the Faculty of Biology and Medicine, University of Lausanne,
Switzerland.
The individual connection matrices were computed using the open ag-
MRI preprocessing.
gregation software Connectome Mapper (http://www.connectomics.org; Daducci et al., 2012),
Network Neuroscience
488
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
n
e
n
_
a
_
0
0
0
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Distance-dependent structural brain networks
which calls different tools at different processing steps using the parameters described in the
sequelae.
MPRAGE volumes were segmented into white matter, gray matter, and cerebrospinal fluid
using FreeSurfer software version 5.0.0 (Dale, Fischl, & Sereno, 1999). Cortical volumes were
segmented into five progressively finer parcellations, with 68, 114, 219, 448, and 1,000 ap-
proximately equally sized parcels (Cammoun et al., 2012). Here, we analyze the 68-, 219-,
and 1,000-parcel divisions. DSI data were reconstructed following the protocol described by
Wedeen and colleagues (V. J. Wedeen, Hagmann, Tseng, Reese, & Weisskoff, 2005), thus
estimating an orientation distribution function (ODF) in each voxel. Up to three main stream-
line orientations were idenntified in each voxel as the maxima of the ODF (Diffusion Toolkit
software, http://www.trackvis.org/dtk).
Structural connectivity matrices were estimated for individual participants using deter-
ministic streamline tractography on reconstructed DSI data, initiating 32 streamline propa-
gations per diffusion direction per white matter voxel (V. Wedeen et al., 2008). The MPRAGE
and the brain parcellation were linearly registered to the subject diffusion space (b0) using a
boundary-based cost function (FreeSurfer software; Greve & Fischl, 2009). For each starting
point, streamlines were grown in two opposite directions with a fixed step size equal to 1 mm.
As the streamline entered new voxels, growth contributed along the ODF maximum direction
that produced the least curvature. Streamlines were terminated if changes in direction were
greater than 60 deg/mm. Tractography completed when both ends of the streamline left the
white matter mask. Structural connectivity between pairs of parcels was estimated in terms of
streamline density, defined as the number of streamlines between two parcels normalized by
the mean length of the streamlines and the mean surface area of the parcels.
Single-Subject Networks and Connection Consensus
Let As ∈ RN×N
be the weighted and symmetric structural connectivity matrix for subject
s = 1, . . . , T, whose element Aijs indicates the normalized streamline count between brain
areas i and j. Given the set of matrices A = {As} we can calculate the consensus matrix,
C ∈ RN×N
[Aijs > 0]. Intuitively, Poi, Cij ∈ [0, 1] indicates the
fraction of T subjects for whom the connection {io, j} is expressed.
, whose element Cij = 1
T ∑T
s=1
Group-Representative Network Construction
In this paper we compare several strategies for constructing group-representative networks
for structural connectivity estimated from dMRI and reconstructed using tractography. In this
section, we introduce several approaches for doing so.
The most naïve approach for generating a group-representative connectivity
Simple average.
matrix is to let each connection’s weight be its mean value over all subjects, ignoring those for
whom a connection is not expressed, questo è, Aijs = 0. We refer to this approach as the simple
average and denote the estimated group-representative connectivity matrix as Asimp
, whose
elements are defined as Asimp
Aijs. Here, T>0 is the set of subjects satisfying
| ∑s∈T>0
Aijs > 0.
ij = 1
|T>0
A more common approach for generating group-representative
Consistency-based thresholding.
matrices is to impose a threshold, τ, over the elements of a consensus matrix, C, so that the only
elements retained are those that satisfy Cij ≥ τ. The intuition is that a good group-representative
Network Neuroscience
489
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
matrix should preserve the features, in this case connections, that are consistently expressed
across individual subjects. Within the broader consensus-based thresholding framework there
are many strategies for implementation. In this section we hightlight those that are explored in
this paper.
The most common variant of consensus-based thresholding is the imposition of a uniform
threshold over all connections. Questo è, all possible elements of the consensus matrix are con-
sidered simultaneously and those that survive Cij ≥ τ are retained. In contrasto, a restricted
threshold is one in which connections are grouped into K classes according to some criteria
and a class-dependent threshold, τ(k), where k ∈ {1, . . . , K}, is imposed separately over each
class. Per esempio, connections could be classified according to whether their starting and
termination points fall within the same or different hemispheres. The restricted threshold is
not limited to ordinal data, but can also be used with continuous variables through discretiza-
zione. Interareal distance, for instance, is a continuous variable that measures the Euclidean
distance between areal centroids. One could impose a distance-dependent threshold, τ(D),
by discretizing the interval of possible Euclidean distances into nonoverlapping bins. Each bin
would include connections that span a particular range of distances, and distinct thresholds
could be imposed within each bin.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Note, for all group-representative networks generated using any of the three consensus-
based thresholding procedures, connection weights were determined using the following two-
step process. Primo, each retained connection was assigned its corresponding weight from the
“simple average” connectivity matrix. Once all connections had an assigned weight, we re-
assigned weights from the pooled subject-level connection weights via linear interpolation.
Così, the resulting matrices had connection weight distributions approximately identical to
that of the typical subject.
Models tested in this submission. Here, we tested four different methods for generating group-
representative networks. The first was the “Simple” method, which retained a connection and
its average weight if it was observed in at least one subject. The second and third methods
were variants of the uniform consensus-based threshold method. The first of these imposed a
threshold of τ = 0.5 over all connections, so that the group network included only those con-
nections expressed in at least half of the subject cohort. The second method involved choosing
a consensus threshold such that resulting network had a binary density as close as possible to
that of the average subject. We refer to this method as the τ = Avg method. The fourth and
final method was a distance-dependent threshold. In this method, subjects’ edge lengths are
combined. Given this list of edges and their lengths, we define M bins based on edge length
percentiles where M is the total number of edges in the consensus network. Prossimo, and for
each bin, we identify all possible edges who, based on their lengths, fall into that bin. Of those
possible edges, we choose the one with the greatest consensus across subjects. If there exists
a tie, we choose the edge with the greatest weight on average. This procedure generates a
network with approximately the same edge length distribution of the original network while
still selectively preserving edges with high consensus across subjects.
We note that, in general, there are many thresholding procedures with different aims
and different criteria for retaining or pruning a connection. Here, we focused on some of
the most common thresholding procedures, all of which operated upon the consensus matrix,
C. Specifically, the τ = 0.5 method can be regarded as a realization of the common “ab-
solute thresholding” procedure, whereas the τ = Avg realizes a “proportionality threshold.”
While these procedures are common in the network neuroscience literature, they are generally
Network Neuroscience
490
Distance-dependent structural brain networks
applied to matrices whose elements represent connection weight, such as fiber density, correla-
tion magnitude, fractional anisotropy, and so on. Here, because our goal is dually to threshold
a matrix but also to retain features that are common across subjects, we apply these thresh-
olding methods to a consensus matrix rather than a weight matrix. It remains unclear whether
weight-based thresholding is an appropriate method for generating group-representative ma-
trices. This should be investigated more explicitly in future work.
We note that of the methods tested here, τ = 0.5 and the distance-dependent method have
the same density (equal to that of the average subject’s density). The other methods, τ = 0.5
and the “Simple” method, both have different densities. This has important implications, COME
fluctuations in network density is known to bias many network measures. Therefore, only the
τ = 0.5 and the distance-dependent methods are directly comparable.
Network Measures
We compared subject-level and group-representative networks using a set of measures that
quantify different aspects of network topology (all measures were computed using func-
tions provided as part of the Brain Connectivity Toolbox (https://sites.google.com/site/bctnet/;
Rubinov & Sporns, 2010). These measures included binary and weighted total connection
weight, degree, clustering coefficient (nodal and global), betweenness centrality, efficiency,
path length, diameter, multiscale modularity, participation coefficient, assortativity, and con-
nection length. In this section, we describe those measures in greater detail.
Among the simplest structural measures one can calculate given a
Degree and total weight.
connectivity matrix, A = {Aij}, is the degree of node i, which summarizes the total number
(or weight) of its connections:
ki = ∑
j
Aij.
(1)
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
Given the vector of nodes’ degrees, k = {ki}, we can then calculate the total weight of the
network as 2m = ∑i ki (the factor of two is necessary in this case because of the undirectedness
of the networks considered here).
Another simple measure is clustering coefficient of each node, io. IL
Clustering coefficient.
clustering coefficient measures the density of connections among all of i’s neighbors and is
calculated as
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
ci =
2ti
ki(ki − 1) ,
(2)
where ti = ∑j,h Aij Aih Ajh is the number of triangles surrounding node i. The node-level
clustering coefficient can be averaged to summarize the mean clustering of a network, C =
N ∑i ci.
1
Path-based measures: Characteristic path length, diameter, betweenness centrality, and efficiency. Given
the connectivity matrix A = {Aij}, we define the matrix D = {Dij} to be the shortest paths
Network Neuroscience
491
Distance-dependent structural brain networks
matrix, whose element Dij is equal to the length of the shortest topological path between
nodes i and j. For binary networks, shortest paths are calculated in terms of geodesic distance,
questo è, number of steps. For weighted networks, Tuttavia, shortest paths are calculated based
on a transformation of edge weights to length. Here, we use a reciprocol weight-to-length
transformation, questo è, Lij = 1
Wij . Once the shortest paths matrix has been calcualted, we
can define characteristic path length as
L =
2
N(N − 1) ∑
io,j>i
Dij.
We can also define network diameter as
Δ = arg max Dij.
Finalmente, we define efficiency as
E =
2
N(N − 1) ∑
io,j>i
1
Dij
.
(3)
(4)
(5)
A measure related to the concept of shortest paths is betweenness centrality. Let ρhj be the
number of shortest paths between nodes h and j and ρhj(io) be the number of shortest paths
between h and j that pass through node i. Then the betweenness centrality of node i, Quale
measures the fraction of all shortest paths that pass through node i, is calculated as
bi =
1
(N − 1)(N − 2)
∑
H,j,H(cid:5)=i,H(cid:5)=j,io(cid:5)=j
ρhj(io)
ρhj
.
(6)
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
Modularity maximization, and the participation coefficient. Many real-world networks exhibit
modular architecture, meaning that their nodes can be meaningfully decomposed subnetworks
(also called “communities” or “modules”) that are internally cohesive but segregated from
one another. Though there are many approaches for detecting modules in networks, one of
the most popular is modularity maximization, which partitions each node i into one of K
communities σi ∈ {1, . . . K} by maximizing a modularity quality function, designed by the
variable Q (Newman & Girvan, 2004). Modularity functions have the following form:
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Q = 1
2m ∑
C
[Aij − Pij].
∑
io,j∈c
(7)
Here, Aij and Pij are the observed and expected weights of the connection between nodes
{io, j}. The double summation ensures that, effettivamente, the sum counts only pairs of nodes
that fall within communities, questo è, σi = σj. Accordingly, Q is optimized when the observed
density of connections within communities is maximally greater than what would be expected
by chance. Here, we let Pij = kik j
2M , which corresponds to a chance method in which each
node’s degree is preserved but where its connections are otherwise made at random.
The standard formulation of modularity, Q, suffers from what is known as a “resolution limit”
rendering it incapable of detecting communities below some characteristic scale determined
Network Neuroscience
492
Distance-dependent structural brain networks
by a network’s overall density (Fortunato & Barthelemy, 2007). To circumvent this issue, UN
parameterized version of the modularity function exists:
Q(γ) = 1
2m ∑
C
[Aij − γPij].
∑
io,j∈c
(8)
Here, γ is the structural resolution parameter, which scales the relative contribution of the
expected weight. Effectively, the value of γ determines the scale of detected communities,
with small and large values of γ returning correspondingly larger or smaller communities. Questo
parameterization does not directly address the issue of the resolution limit; it simply shifts the
scale below which communities are undetectable.
Optimizing Q(γ) is computationally intractable. Tuttavia, there exist many methods and
heuristics for approximating the optimal solution. One of the most common is the so-called
Louvain algorithm, which has proven both fast and accurate in benchmarking tests (Blondel,
Guillaume, Lambiotte, & Lefebvre, 2008). The Louvain algorithm is stochastic, Tuttavia, and its
estimate of the optimal partitions depends upon initial conditions. Accordingly, it is common
to repeat the algorithm many times to generate a sample of near-optimal solutions.
Here, we vary γ over a range of 0.7 A 2.1 in increments of 0.1. At each discrete value,
we optimize Q(γ) using a generalization of the Louvain algorithm 100 times (Jutla, Jeub, &
Mucha, 2011).
The partitions generated by modularity maximization and related community detection al-
gorithms can be used to further characterize different aspects of network organization and
function. One such metric is the participation coefficient, which measures the extent to which
a node’s connections are distributed across modules or concentrated within its own module
(Guimera & Amaral, 2005). Let κiσ denote the total weight of connections node i makes to
module σ. Participation coefficient is calculated as
pi = 1 − ∑
P
(cid:2)
(cid:3)
2
.
κiσ
ki
(9)
Intuitively, the closer pi is to 1, the greater the extent that i’s connections are distributed across
many different modules. Values of pi close to 0 indicate that i’s connections are conncentrated
within a few modules.
Degree assortativity. We also computed degree assortativity, a measure that quantifies the
extent to which nodes of a given degree tend to connect with other nodes of similar degree
(Newman, 2002). Intuitively, degree assortativity is a Pearson correlation of the degrees at
different endpoints of each edge in the network and is calculated as
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
r =
M−1 ∑i jiki − [M−1 ∑i
io + k2
io ) − [M−1 ∑i
(j2
1
2
M−1 ∑i
1
2
(ji + ki)2]
(ji + ki)]2
1
2
.
(10)
Here, i = 1, . . . , M indexes the edges in the network, and ji and ki are the degrees of nodes
connected by the ith edge.
Finalmente, we also computed connection length distributions. We define the
Connection length.
length of a connection between nodes {io, j} as the Euclidean distance separating the centroids
Network Neuroscience
493
Distance-dependent structural brain networks
of those regions. Generalmente, connections are curvilinear and do not adhere to straight-line dis-
tances. Tuttavia, in studies that measured both Euclidean distance and the curvilinear fiber
lengths of connections, these measures were found to be highly correlated (Betzel, Avena-
Koenigsberger et al., 2016). This implies that, while Euclidean distance is not a perfect substi-
tute for a connection’s length, it is a very good first-order approximation.
SUPPORTING INFORMATION
Code for generating the uniform and distance-dependent group-representative networks is
available at https://www.richardfbetzel.com/code/.
AUTHOR CONTRIBUTIONS
Richard Betzel: Conceptualization; Formal analysis; Software; Writing – original draft;
Writing – review & editing. Alessandra Griffa: Data curation; Resources; Writing – review &
editing. Patric Hagmann: Data curation; Resources; Writing – review & editing. Bratislav Miši´c:
Conceptualization; Formal analysis; Supervision; Writing – original draft; Writing – review &
editing.
FUNDING INFORMATION
McGill University, Canada First Research Excellence Fund: McGill University for the Healthy
Brains for Healthy Lives initiative, Award ID: 1a-DF-1. Bratislav Miši´c, Natural Sciences and
Engineering Research Council of Canada, Award ID: RGPIN #017-04265. Bratislav Misic,
Fonds de recherche Quebec – Sante (ChercheurBoursier) and the Canadian Institutes of Health
Research, Award ID: #391300. Richard Betzel, Indiana University Office of the Vice President
for Research Emerging Area of Research Initiative, Apprendimento: Brains, Machines and Children.
REFERENCES
Betzel, R. F., Avena-Koenigsberger, A., Goñi, J., Lui, Y., De Reus,
M. A., Griffa, A., . . . Sporns, O. (2016). Generative models of
the human connectome. NeuroImage, 124, 1054–1064.
long-distance connections in weighted,
Betzel, R. F., & Bassett, D. S. (2018). Specificity and robustness
Di
interareal con-
nectomes. Proceedings of the National Academy of Sciences,
201720186.
Betzel, R. F., Griffa, A., Hagmann, P., & Miši´c, B. (2019). Supporto-
ing information for “Distance-dependent consensus thresholds
for generating group-representative structural brain networks.”
Network Neuroscience, 3(2), 475–496. https://doi.org/10.1162/
netn_a_00075
Betzel, R. F., Gu, S., Medaglia, J. D., Pasqualetti, F., & Bassett, D. S.
(2016). Optimally controlling the human connectome: The role
of network topology. Scientific Reports, 6, 30770.
Betzel, R. F., Medaglia, J. D., & Bassett, D. S. (2018). Diversity of
meso-scale architecture in human and non-human connectomes.
Nature Communications, 9(1), 346.
Betzel, R. F., Medaglia, J. D., Papadopoulos, L., Baum, G. L., Gur,
R., Gur, R., . . . Bassett, D. S. (2017). The modular organization
of human anatomical brain networks: Accounting for the cost of
wiring. Network Neuroscience, 1(1), 42–68.
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks. Jour-
nal of Statistical Mechanics: Theory and Experiment, 2008(10),
P10008.
Briggman, K. L., Helmstaedter, M., & Denk, W. (2011). Wiring
the retina.
specificity in the direction-selectivity circuit of
Nature, 471(7337), 183.
Bullmore, E., & Sporns, O. (2012). The economy of brain network
organization. Nature Reviews Neuroscience, 13(5), 336.
Buzsáki, G., & Mizuseki, K. (2014). The log-dynamic brain: How
skewed distributions affect network operations. Nature Reviews
Neuroscience, 15(4), 264.
Calabrese, E., Badea, A., Cofer, G., Qi, Y., & Johnson, G. UN. (2015).
A diffusion MRI tractography connectome of the mouse brain and
comparison with neuronal tracer data. Cerebral Cortex, 25(11),
4628–4637.
Cammoun, L., Gigandet, X., Meskaldji, D., Thiran, J., Sporns, O.,
Fare, K., . . . Hagmann, P. (2012). Mapping the human connec-
tome at multiple scales with diffusion spectrum MRI. Journal of
Neuroscience Methods, 203(2), 386–397.
Cohen, J. R., & D’Esposito, M. (2016). The segregation and integra-
tion of distinct brain networks and their relationship to cognition.
Journal of Neuroscience, 36(48), 12083–12094.
Daducci, A., Gerhard, S., Griffa, A., Lemkaddem, A., Cammoun, L.,
Gigandet, X., . . . Thiran, J.-P. (2012). The connectome mapper:
An open-source processing pipeline to map connectomes with
MRI. PloS ONE, 7(12), e48121.
Dale, UN. M., Fischl, B., & Sereno, M. IO. (1999). Cortical surface-
IO. Segmentation and surface reconstruction.
based analysis:
NeuroImage, 9(2), 179–194.
Network Neuroscience
494
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
Deco, G., Tononi, G., Boly, M., & Kringelbach, M. l. (2015). Re-
thinking segregation and integration: Contributions of whole-
brain modelling. Nature Reviews Neuroscience, 16(7), 430.
de Reus, M. A., & van den Heuvel, M. P.
(2013). Estimating
false positives and negatives in brain networks. NeuroImage, 70,
402–409.
Descoteaux, M., Deriche, R., Knosche, T. R., & Anwander, UN.
(2009). Deterministic and probabilistic tractography based on
complex fibre orientation distributions. IEEE Transactions on
Medical Imaging, 28(2), 269–286.
Ercsey-Ravasz, M., Markov, N. T., Lamy, C., Van Essen, D. C.,
Knoblauch, K., Toroczkai, Z., & Kennedy, H. (2013). A predic-
tive network model of cerebral cortical connectivity based on a
distance rule. Neuron, 80(1), 184–197.
Fallani, F. D. V., Latora, V., & Chavez, M. (2017). A topological cri-
terion for filtering information in complex brain networks. PLoS
Computational Biology, 13(1), e1005305.
Fortunato, S., & Barthelemy, M. (2007). Resolution limit in commu-
nity detection. Proceedings of the National Academy of Sciences,
104(1), 36–41.
Garrison, K. A., Scheinost, D., Finn, E. S., Shen, X., & Constable,
R. T. (2015). IL (In) stability of functional brain network mea-
sures across thresholds. NeuroImage, 118, 651–661.
Gollo, l. L., Roberts, J. A., Cropley, V. L., Di Biase, M. A., Pantelis,
C., Zalesky, A., & Breakspear, M. (2018). Fragility and volatility of
structural hubs in the human connectome. Nature Neuroscience,
21(8), 1107.
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain im-
age alignment using boundary-based registration. NeuroImage,
48(1), 63–72.
Guimera, R., & Amaral, l. UN. N. (2005). Functional cartography of
complex metabolic networks. Nature, 433(7028), 895.
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Wedeen, V. J., & Sporns, O. (2008). Mapping the structural core
of human cerebral cortex. PLoS Biology, 6(7), e159.
Helmstaedter, M., Briggman, K. L., Turaga, S. C., Jain, V., Seung,
H. S., & Denk, W. (2013). Connectomic reconstruction of the in-
ner plexiform layer in the mouse retina. Nature, 500(7461), 168.
Henderson, J. A., & Robinson, P. UN. (2013). Using geometry to un-
cover relationships between isotropy, homogeneity, and modu-
larity in cortical connectivity. Brain Connectivity, 3(4), 423–437.
Horvát, S., G˘am˘anu, R., Ercsey-Ravasz, M., Magrou, L., G˘am˘anu,
B., Van Essen, D. C., . . . Kennedy, H. (2016). Spatial embedding
and wiring cost constrain the functional layout of the cortical net-
work of rodents and primates. PLoS Biology, 14(7), e1002512.
Jones, D. K., Knösche, T. R., & Turner, R. (2013). White matter in-
tegrity, fiber count, and other fallacies: The do’s and don’ts of
diffusion MRI. NeuroImage, 73, 239–254.
Jutla, IO. S., Jeub, l. G., & Mucha, P. J. (2011). A generalized Louvain
method for community detection implemented in Matlab. http://
netwiki.amath.unc.edu/GenLouvain
Kaiser, M., & Hilgetag, C. C. (2006). Nonoptimal component place-
ment, but short processing paths, due to long-distance projec-
tions in neural systems. PLoS Computational Biology, 2(7), e95.
Maier-Hein, K., Neher, P., Houde, J.-C., Cote, M.-A., Garyfallidis,
(2016). Tractography-
. Descoteaux, M.
E., Zhong,
J.,
.
.
based connectomes are dominated by false-positive connections.
bioRxiv:084137
Markov, N. T., Ercsey-Ravasz, M., Ribeiro Gomes, A., Lamy, C.,
Magrou, L., Vezoli, J., . . . Kennedy, H. (2012). A weighted
and directed interareal connectivity matrix for macaque cerebral
cortex. Cerebral Cortex, 24(1), 17–36.
Miši´c, B., Betzel, R. F., Nematzadeh, A., Goni,
J., Griffa, A.,
Hagmann, P., . . . Sporns, O. (2015). Cooperative and competitive
spreading dynamics on the human connectome. Neuron, 86(6),
1518–1529.
Miši´c, B., & Sporns, O. (2016). From regions to connections and
networks: New bridges between brain and behavior. Current
Opinion in Neurobiology, 40, 1–7.
Newman, M. E. (2002). Assortative mixing in networks. Physical
Review Letters, 89(20), 208701.
Newman, M. E., & Girvan, M. (2004). Finding and evaluating
community structure in networks. Physical Review E, 69(2),
026113.
Noori, H. R., Schöttler, J., Ercsey-Ravasz, M., Cosa-Linan, A., Varga,
M., Toroczkai, Z., & Spanagel, R. (2017). A multiscale cerebral
neurochemical connectome of the rat brain. PLoS Biology, 15(7),
e2002612.
Oh, S. W., Harris, J. A., Di, L., Winslow, B., Cain, N., Mihalas, S., . . .
Zeng, H. (2014). A mesoscale connectome of the mouse brain.
Nature, 508(7495), 207.
Passingham, R. E., Stephan, K. E., & Kötter, R. (2002). The anatomi-
cal basis of functional localization in the cortex. Nature Reviews
Neuroscience, 3(8), 606.
Pestilli, F., Yeatman, J. D., Rokem, A., Kay, K. N., & Wandell, B. UN.
(2014). Evaluation and statistical inference for human connec-
tomes. Nature Methods, 11(10), 1058.
Reveley, C., Seth, UN. K., Pierpaoli, C., Silva, UN. C., Yu, D.,
Saunders, R. C., . . . Frank, Q. Y. (2015). Superficial white matter
fiber systems impede detection of long-range cortical connec-
tions in diffusion MR tractography. Proceedings of the National
Academy of Sciences, 201418198.
Roberts,
J. A., Perry, A., Lord, UN. R., Roberts, G., Mitchell,
P. B., Smith, R. E., . . . Breakspear, M. (2016). The contribu-
tion of geometry to the human connectome. NeuroImage, 124,
379–393.
Roberts, J. A., Perry, A., Roberts, G., Mitchell, P. B., & Breakspear,
M. (2017). Consistency-based thresholding of the human con-
nectome. NeuroImage, 145, 118–129.
Rosenthal, G., Váša, F., Griffa, A., Hagmann, P., Amico, E., Goñi, J.,
. . . Sporns, O. (2018). Mapping higher-order relations between
brain structure and function with embedded vector representa-
tions of connectomes. Nature Communications, 9(1), 2178.
Rubinov, M. (2016). Constraints and spandrels of interareal connec-
tomes. Nature Communications, 7, 13812.
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069.
Samu, D., Seth, UN. K., & Nowotny, T. (2014). Influence of wiring
cost on the large-scale architecture of human cortical connectiv-
ità. PLoS Computational Biology, 10(4), e1003557.
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, UN. J., . . . Sì, B. T. (2017). Local-global parcellation of
Network Neuroscience
495
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Distance-dependent structural brain networks
the human cerebral cortex from intrinsic functional connectivity
MRI. Cerebral Cortex, 1–20.
Sporns, O. (2013). Network attributes for segregation and integra-
tion in the human brain. Current Opinion in Neurobiology, 23(2),
162–171.
Sporns, O., Tononi, G., & Edelman, G. M. (2000). Theoretical neu-
roanatomy: Relating anatomical and functional connectivity in
graphs and cortical connection matrices. Cerebral Cortex, 10(2),
127–141.
Sporns, O., Tononi, G., & Kötter, R. (2005). The human connectome:
A structural description of the human brain. PLoS Computational
Biology, 1(4), e42.
Sporns, O., & Zwi, J. D. (2004). The small world of the cerebral
cortex. Neuroinformatics, 2(2), 145–162.
Stiso, J., & Bassett, D. (2018). Spatial embedding imposes constraints
on the network architectures of neural systems. arXiv:1807.04691
van den Heuvel, M. P., Bullmore, E. T., & Sporns, O. (2016).
Comparative connectomics. Trends in cognitive sciences, 20(5),
345–361.
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organiza-
tion of the human connectome. Journal of Neuroscience, 31(44),
15775–15786.
van den Heuvel, M. P., & Sporns, O. (2013). An anatomical sub-
strate for integration among functional networks in human cor-
tex. Journal of Neuroscience, 33(36), 14489–14500.
van Wijk, B. C., Stam, C. J., & Daffertshofer, UN. (2010). Comparing
brain networks of different size and connectivity density using
graph theory. PloS ONE, 5(10), e13701.
Vértes, P. E., Alexander-Bloch, UN. F., Gogtay, N., Giedd, J. N.,
Rapoport, J. L., & Bullmore, E. T. (2012). Simple models of human
brain functional networks. Proceedings of the National Academy
of Sciences, 201111738.
Wedeen, V., Wang, R., Schmahmann, J., Benner, T., Tseng, W., Dai,
G., . . . de Crespigny, UN. (2008). Diffusion spectrum magnetic
resonance imaging (DSI) tractography of crossing fibers. Neuro-
Image, 41(4), 1267–1277.
Wedeen, V. J., Hagmann, P., Tseng, W.-Y. I., Reese, T. G., &
Weisskoff, R. M. (2005). Mapping complex tissue architecture
with diffusion spectrum magnetic resonance imaging. Magnetic
Resonance in Medicine, 54(6), 1377–1386.
Yeh, F.-C., Vettel, J. M., Singh, A., Poczos, B., Grafton, S. T.,
Erickson, K. I., . . . Verstynen, T. D. (2016). Quantifying differ-
ences and similarities in whole-brain white matter architecture
using local connectome fingerprints. PLoS Computational Biol-
ogy, 12(11), e1005203.
Zalesky, A., Fornito, A., & Bullmore, E. (2012). On the use of corre-
lation as a measure of network connectivity. NeuroImage, 60(4),
2096–2106.
Zalesky, A., Fornito, A., Cocchi, L., Gollo, l. L., van den Heuvel,
M. P., & Breakspear, M. (2016). Connectome sensitivity or speci-
ficity: Which is more important? NeuroImage, 142, 407–420.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
3
2
4
7
5
1
0
9
2
6
4
8
N
e
N
_
UN
_
0
0
0
7
5
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Network Neuroscience
496