RESEARCH
Catecholaminergic manipulation alters dynamic
network topology across cognitive states
James M. Shine
1,2
Sander Nieuwenhuis
, Ruud L. van den Brink
3,4
1
, and Russell A. Poldrack
3,4
5
, Dennis Hernaus
,
1Department of Psychology, Stanford University, Stanford, CA, USA
2Brain and Mind Centre, The University of Sydney, Sydney, NSW, Australia
3Institute of Psychology, Leiden University, Leiden, The Netherlands
4Leiden Institute for Brain and Cognition, Leiden, The Netherlands
5Maryland Psychiatric Research Center, University of Maryland School of Medicine, MD, USA
a n o p e n a c c e s s
j o u r n a l
Keywords: fMRI; Noradrenaline; Integration; Flexibility; Network; Atomoxetine
ABSTRACT
The human brain is able to flexibly adapt its information processing capacity to meet a
variety of cognitive challenges. Recent evidence suggests that this flexibility is reflected in the
dynamic reorganization of the functional connectome. The ascending catecholaminergic
arousal systems of the brain are a plausible candidate mechanism for driving alterations
in network architecture, enabling efficient deployment of cognitive resources when the
environment demands them. We tested this hypothesis by analyzing both resting-state and
task-based fMRI data following the administration of atomoxetine, a noradrenaline reuptake
inhibitor, compared with placebo, in two separate human fMRI studies. Our results
demonstrate that the manipulation of central catecholamine levels leads to a reorganization
of the functional connectome in a manner that is sensitive to ongoing cognitive demands.
AUTHOR SUMMARY
There is emerging evidence that the flexible network structure of the brain is related to
activity within the ascending arousal systems of the brain, such as the noradrenergic locus
coeruleus. Here, we explored the role of catecholaminergic activity on network architecture
by analyzing the graph structure of the brain measured using functional MRI following the
administration of atomoxetine, a selective noradrenaline reuptake inhibitor. We estimated
functional network topology in two double-blind, placebo-controlled datasets: one from the
resting state and another from a parametric N-back task. Our results demonstrate that the
nature of catecholaminergic network reconfiguration is differentially related to cognitive state
and provide confirmatory evidence for the hypothesis that the functional network signature of
the brain is sensitive to the ascending catecholaminergic arousal system.
INTRODUCTION
A fundamental question facing modern neuroscience is how local computations are integrated
across the brain to support the vast repertoire of mammalian behavior and cognition. Conver-
gent results from multimodal neuroimaging studies (de Pasquale, Penna, Sporns, Romani, &
Corbetta, 2016; Kitzbichler, Henson, Smith, Nathan, & Bullmore, 2011; Shine et al., 2016;
Vatansever, Menon, Manktelow, Sahakian, & Stamatakis, 2015) have demonstrated that brain
Citation: Shine, J. M., van den Brink,
R. L., Hernaus, D., Nieuwenhuis,
S., & Poldrack, R. UN. (2018).
Catecholaminergic manipulation
alters dynamic network topology
across cognitive states. Network
Neuroscience, 2(3), 381–396.
https://doi.org/10.1162/netn_a_00042
DOI:
https://doi.org/10.1162/netn_a_00042
Supporting Information:
https://doi.org/10.1162/netn_a_00042
Received: 6 ottobre 2017
Accepted: 2 Gennaio 2018
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
James M. Shine
mac.shine@sydney.edu.au
Handling Editor:
Olaf Sporns
Copyright: © 2018
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
N
e
N
_
UN
_
0
0
0
4
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Catecholaminergic manipulation and cognitive states
Segregation:
A state in which there is a relatively
high modularity in a network, cioè.,
there are distinct modules with weak
interconnections.
Integration:
A state in which there is a relatively
low modularity in a network, cioè.,
there are less distinct modules with
strong interconnections.
Network:
A system comprising a set of distinct
nodes, interconnected by a set of
edges.
Noradrenaline:
A catecholaminergic
neurotransmitter that provides a
widespread, modulatory role over
neuronal processing in the central
nervous system.
Atomoxetine:
A chemical that inhibits the
transporter chemical that normally
facilitates the reuptake of
noradrenaline.
Module:
A group of nodes that is more tightly
interconnected than the connections
between modules.
Network Neuroscience
activity during cognitive tasks reflects a balance between regional segregation and network-
level integration (Shine & Poldrack, in press), in which communication across distributed
circuits enables fast and effective cognitive performance (Shine et al., 2016).
There is growing evidence that ascending catecholaminergic neuromodulatory projections
from the brainstem mediate this integration (Samuels & Szabadi, 2008; Shine et al., 2016). Pro-
jections from arousal-related nuclei, such as the noradrenergic locus coeruleus (Sara, 2009),
arborize widely in target regions and putatively alter network architecture by modulating the
impact of incoming neuronal input in an activity-dependent manner (Aston-Jones & Cohen,
2005). Previous neuroimaging studies in humans have highlighted a close relationship be-
tween noradrenaline, network topology, and cognitive performance (Eldar, Cohen, & Niv,
2013; Shine et al., 2016). Specifically, increased free noradrenaline has been shown to in-
crease the phasic-to-tonic ratio of neuronal firing in both the locus coeruleus and the cortex.
As such, neurons that are less tonically active during the unstimulated state may also si-
multaneously demonstrate a heightened responsivity to relevant stimuli (Bari & Aston-Jones,
2013; Devilbiss & Waterhouse, 2011). We have previously used a biophysical computational
model to demonstrate that fluctuations in neural gain, the potential computational role of cat-
echolamines (Aston-Jones & Cohen, 2005; Servan-Schreiber, Printz, & Cohen, 1990), controls
the balance between network-level segregation and integration (Shine, Aburn, Breakspear, &
Poldrack, in press). Tuttavia, it is not yet known whether directly manipulating noradrenaline
shapes network topology, or indeed whether the effects of noradrenergic function on network
topology differ across behavioral contexts.
To test the hypothesis that ascending catecholamines modulate global network topology as
a function of cognitive state, we analyzed two separate fMRI datasets in which individuals
were scanned following administration of either atomoxetine (ATX), a noradrenergic reuptake
In the first
inhibitor (Robbins & Arnsten, 2009), or a pharmacologically inactive placebo.
study, subjects were scanned in the “resting” state (van den Brink et al., 2016); in the sec-
ond, subjects were scanned while performing a cognitively challenging N-back task (Hernaus,
Casales Santa, Offermann, & Van Amelsvoort, 2017). Based on the opposing effects of ATX
on functional connectivity observed in these two studies (Hernaus et al., 2017; van den Brink
et al., 2016), animal studies that highlight differential effects of ATX on phasic versus tonic
locus coeruleus activity (Bari & Aston-Jones, 2013) and the hypothesized link between nor-
adrenaline and network topology (Eldar et al., 2013; Shine et al., 2016), we expected that ATX
administration would manifest distinct topological effects as a function of cognitive state.
RESULTS
Effect of Atomoxetine on the Topological Signature of the Resting State
In the double-blind, placebo-controlled crossover resting-state study (van den Brink et al.,
2016), 24 healthy subjects (age = 19–26) underwent fMRI scanning prior to (t = −20 minutes)
and following (t = +90 minutes) the administration of either 40 mg of ATX or placebo. A
estimate time-resolved network topology, we submitted preprocessed BOLD fMRI data from
each subject to a preregistered analysis pipeline that calculates sliding-window connectivity
between regional time series (Shine et al., 2015) (Supporting Information Figure S1a, Shine,
van den Brink, Hernaus, Nieuwenhuis, & Poldrack, 2018) and then estimates the resulting
topological signature of each windowed graph (Shine et al., 2016). Specifically, we used a
weighted- and signed-version of the Louvain algorithm (Rubinov & Sporns, 2010) to identify
tightly connected communities of regions within each temporal window. We then determined
how strongly each region was connected to other regions within its own module (quantified
382
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
N
e
N
_
UN
_
0
0
0
4
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Catecholaminergic manipulation and cognitive states
using the within-module degree Z-score: WT) as well as to regions outside of its own module
(quantified using the participation coefficient: BT) over time. The resultant topology can be
summarized at the regional level (per esempio., to determine which regions were the most integrated
during a particular cognitive state) or at the global level by using a joint histogram of WT
and BT values (known as a “cartographic profile”). Rightward fluctuations in the density of
the cartographic profile along the horizontal (cioè., BT) axis reflect a more highly integrated
functional connectome and have been shown to relate positively to individual differences
in effective cognitive performance (Mattar, Cole, Thompson-Schill, & Bassett, 2015; Shine
et al., 2016). Importantly, the participation coefficient measures the distribution of connections
rather than their magnitude, such that BT can be elevated in cases with relatively weak overall
connectivity between regions.
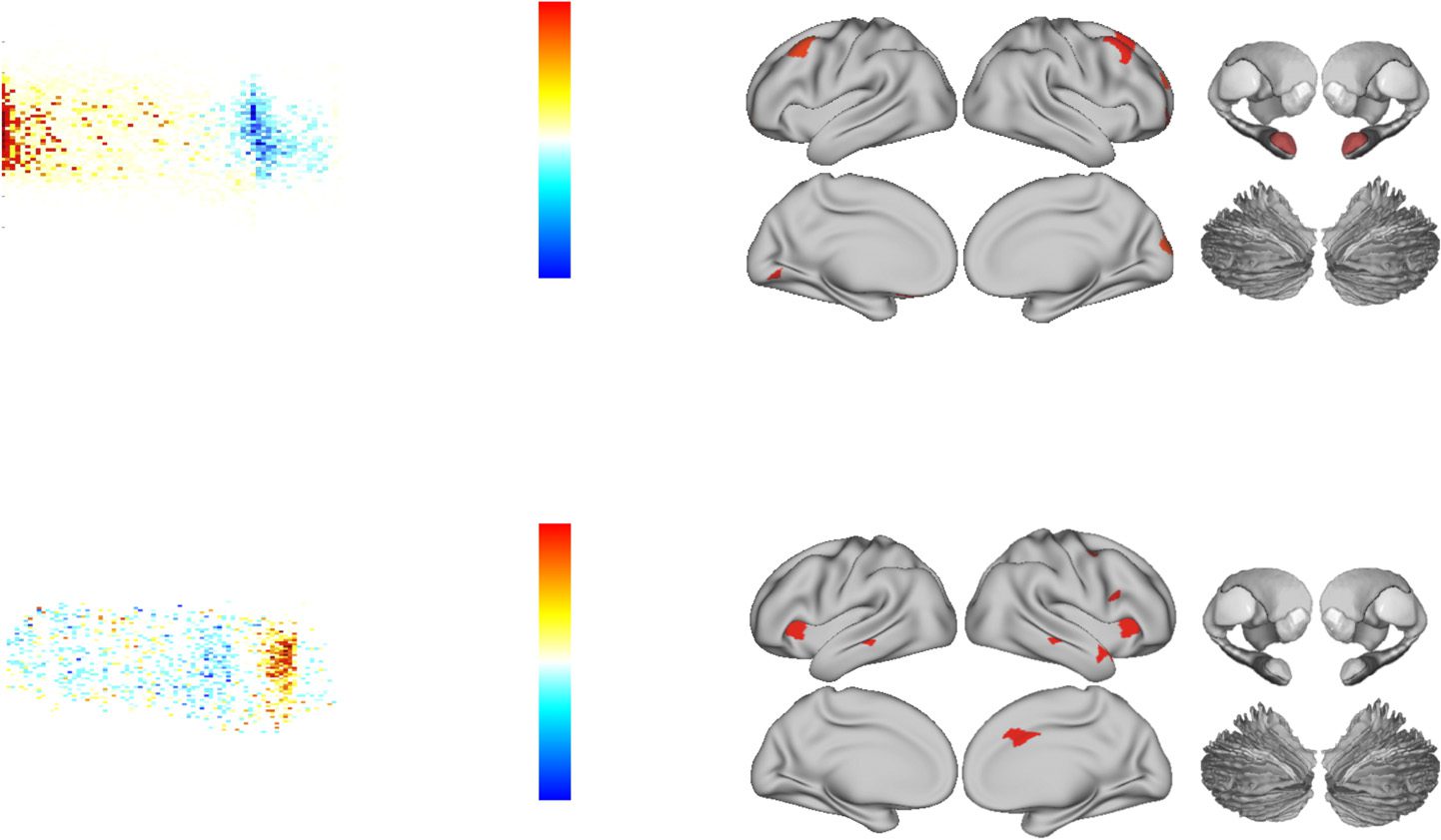
As predicted (https://osf.io/utqq2), the administration of ATX compared with placebo at rest
led to a significant reconfiguration of network-level topology (Figure 1A). Specifically, ATX
administration caused a global shift toward segregation that was maximal in lateral frontal,
frontopolar, and occipital cortices, along with the bilateral amygdala (Figure 1B). A parsimo-
nious explanation for this result is that increases in free synaptic noradrenaline are known to
downregulate tonic activity within the locus coeruleus, which has a dense expression of in-
hibitory α2-autoreceptors (Bari & Aston-Jones, 2013; Sara, 2009). As such, our results suggest
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
N
e
N
_
UN
_
0
0
0
4
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1.
(UN) Effect of atomoxetine versus placebo on the cartographic profile, which demonstrates
a shift toward segregation: red/yellow, increased frequency postatomoxetine; and blue, decreased
frequency postatomoxetine (FDR q ≤ 0.05).
(B) Parcels with decreases in their between-module
connectivity (cioè., participation coefficient) following atomoxetine (vs. placebo); see Supporting
Information Table S1 (Shine et al., 2018) for parcel MNI coordinates (FDR q ≤ 0.05). (C) Effect of
atomoxetine versus placebo on the relationship between the cartographic profile and pupil diam-
eter, which demonstrates a shift toward integration: red/yellow, increased frequency postatomoxe-
tine; and blue, decreased frequency postatomoxetine (FDR q ≤ 0.05). (D) Parcels with increased
time-varying connectivity between between-module connectivity (cioè., participation coefficient) E
pupil diameter following atomoxetine (vs. placebo); see Table S1 for parcel MNI coordinates (FDR
q ≤ 0.05). Key: ATX, atomoxetine; BT, between-module connectivity; WT, within-module connec-
attività; see Table S1 for parcel coordinates.
Network Neuroscience
383
Catecholaminergic manipulation and cognitive states
that network topology in the resting state became segregated because of a reduction in the
tonic firing rate of the locus coeruleus (Bari & Aston-Jones, 2013). This interpretation is consis-
tent with recent computational models that demonstrate a strong link between the tonic firing
rate of the locus coeruleus and functional signatures of brain network activity (Safaai, Neves,
Eschenko, Logothetis, & Panzeri, 2015).
Network Topology is Sensitive to Phasic and Tonic Catecholaminergic Levels
Although the topological signature observed in the resting state is consistent with a decrease in
tonic noradrenaline, in vivo experiments in rodents have demonstrated that ATX administration
also enhances phasic firing patterns in the locus coeruleus (Bari & Aston-Jones, 2013). This in
turn should be expected to potentiate phasic noradrenergic responses and hence integrate the
brain; Tuttavia, this will happen only under conditions necessary to elicit phasic noradrenergic
signaling, such as sensory salience (Nieuwenhuis, De Geus, & Aston-Jones, 2011) and acute
stress (Hermans, van Marle, & Ossewaarde, 2011). Così, in the context of an increase in free
noradrenaline, we might expect that the strength of the relationship between network topology
and phasic noradrenaline should increase following ATX administration. Questo è, the presence
of extra noradrenaline should facilitate additional network reconfiguration as a function of
behavioral requirements.
The lack of cognitive constraints during the resting state make it inherently difficult to directly
test whether the predicted alterations in phasic catecholaminergic activity were indeed related
to changes in network topology. Fortunately, we could interrogate this hypothesis by lever-
aging the relationship between the locus coeruleus and the descending sympathetic circuitry
that controls pupil dilation (Nieuwenhuis et al., 2011), which in turn has been linked to cog-
nitively relevant alterations in cortical arousal ( Joshi, Li, Kalwani, & Gold, 2016; McGinley,
David, & McCormick, 2015UN; Reimer et al., 2016). In a previous study, we demonstrated a
positive relationship between pupil diameter and fluctuations in network topology (Shine et al.,
2016), suggesting that ascending neuromodulatory signals may facilitate network-level inte-
gration. In the current study, we hypothesized that the increase in free catecholamines follow-
ing atomoxetine (Warren, van den Brink, Nieuwenhuis, & Bosch, 2017) should heighten this
relationship, and hence lead to a stronger relationship between pupil diameter and network-
and regional-level integration. Consistent with this hypothesis, we observed a stronger rela-
tionship between pupil diameter and network topology following ATX administration than
following placebo (Figure 1C and 1D). Together, these results provide evidence to suggest that
during quiescence, network topology is sensitive to both phasic and tonic patterns of ongoing
noradrenergic activity.
Effect of Atomoxetine on the Topological Signature of Cognitive Function
A potential benefit of increasing the concentration of free noradrenaline (Invernizzi & Garattini,
2004) is that the liberated catecholamines can be utilized in appropriate contexts to facilitate
activity within task-relevant neural circuits. In other words, ATX may downregulate tonic nor-
adrenergic release during rest, but when required, it may conversely facilitate an increase in
phasic noradrenergic release (Bari & Aston-Jones, 2013) and hence increase network-level
integration. To directly test this hypothesis, we analyzed data from a separate dataset of 19
subjects (age range 18–30) who underwent a cognitively challenging, parametric N-back task
after the administration of either ATX (60 mg) or placebo (Hernaus et al., 2017) (Supporting
Information Figure S1b, Shine et al., 2018). We hypothesized that because of a heightened
Network Neuroscience
384
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
N
e
N
_
UN
_
0
0
0
4
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Catecholaminergic manipulation and cognitive states
phasic noradrenergic response, the main effect of network-level integration should be more
pronounced during the task following ATX, when compared with the placebo condition.
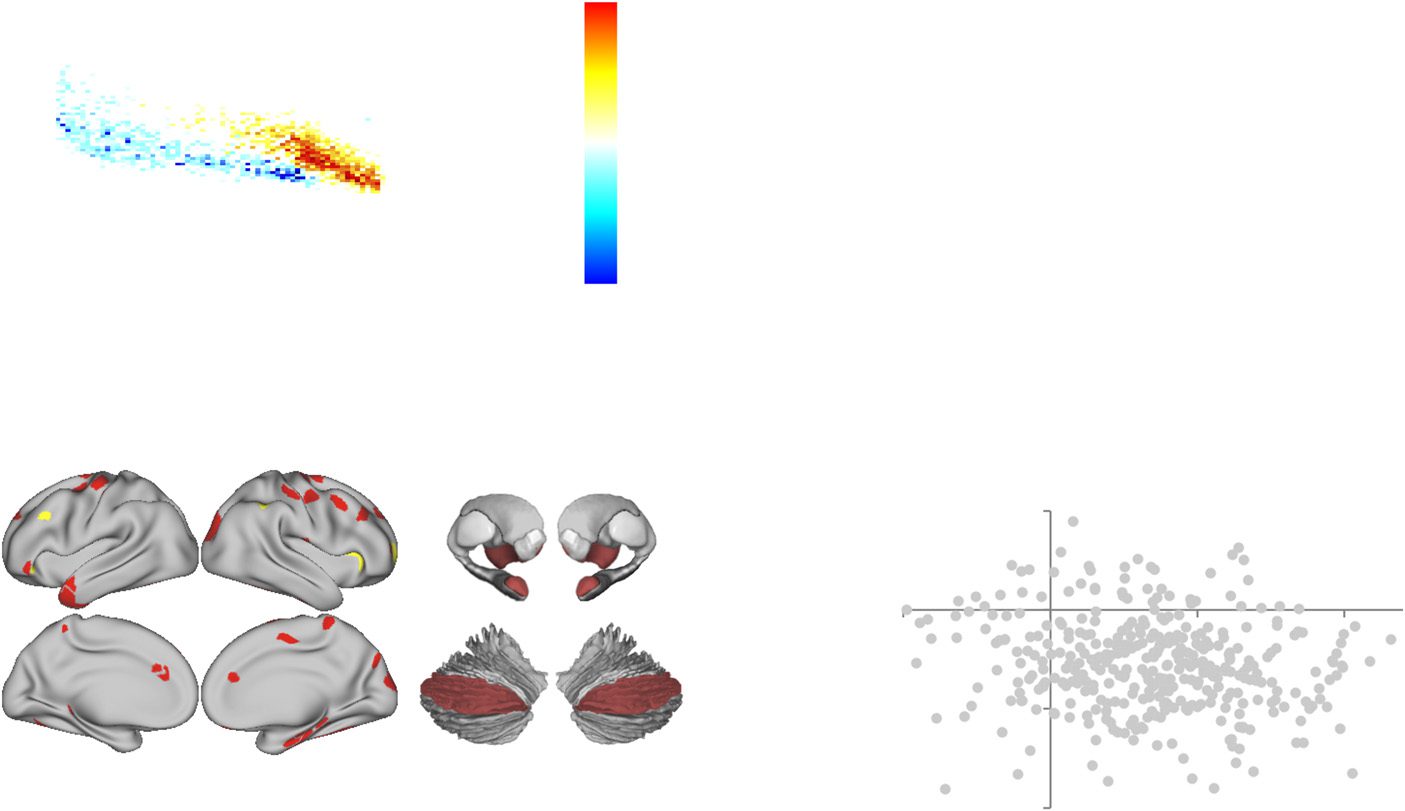
Consistent with our hypothesis, we observed a significant increase in network-level integra-
tion during task performance following ATX (Figure 2A). Specifically, there was an inverted
U-shaped relationship between cognitive load and network integration in both conditions that
was significantly elevated in the post-ATX session (t = 2.47; p = 0.009; Figure 2B). This main
effect of ATX was maximal across frontal, parietal, and temporal cortices, along with thalamus,
amygdala, and Crus II of the cerebellum (Figure 2C, red). Importantly, there is a long-standing
research literature linking catecholamines with cognitive function via an inverse U-shaped
relationship (Cools & D’Esposito, 2011; Robbins & Arnsten, 2009); Tuttavia, few studies have
provided a potential explanation for the algorithmic benefits that such a mechanism might
confer. Here, we demonstrate that network integration may mirror the inverted U-shaped
relationship between catecholamine levels and cognitive performance.
Regional Topological Signatures Change as a Function of Cognitive Load
Although the majority of regions across the brain demonstrated an inverse U-shaped relation-
ship with load, there was a subset of regions that demonstrated a linear increase with cogni-
tive load following atomoxetine administration (Figure 2C, yellow). Specifically, the bilateral
anterior insula, left dorsolateral prefrontal cortex, and right frontopolar cortex demonstrated a
higher extent of integration (BT) with increasing task complexity following ATX, suggesting that
the additional free catecholamines may have facilitated enhanced topological involvement of
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
N
e
N
_
UN
_
0
0
0
4
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
(UN) Mean cartographic profile across all four blocks of load comparing post-ATX to
Figura 2.
postplacebo; similar patterns were observed in each block (FDR q ≤ 0.05). (B) Mean parcelwise B
for each N-back load in both the placebo (PLC, blue) and atomoxetine (ATX, red) conditions (error
bars represent standard error across subjects). (C) Parcels with higher BT post-ATX as a function of
task performance (FDR q ≤ 0.05); main effect (red) and load effect (yellow). (D) Correlation between
the regions that showed highest BT during task performance (ATX > Placebo) and regions that
were shifted toward segregation in the rest study (ATX(Post>Pre) > Placebo(Post>Pre)); see Supporting
Information Table S1 (Shine et al., 2018) for parcel MNI coordinates (FDR q ≤ 0.05). Key: ATX,
atomoxetine; BT, between-module connectivity; WT, within-module connectivity.
Network Neuroscience
385
Catecholaminergic manipulation and cognitive states
these regions as a function of task performance. Together, the relationship between these re-
gions and ATX suggests that phasic noradrenaline may selectively enhance performance in
task-relevant regions, perhaps through arousal-mediated alterations in neural gain (Devilbiss
& Waterhouse, 2011; Reimer et al., 2014; Waterhouse, Moises, & Woodward, 1980).
Differential Effects of Catecholamines on Network Topology as a Function of Cognitive State
The topological dissociation across the two studies analyzed suggests that the effect of ATX on
network topology may be mediated by a set of similar regions across cognitive states. When
we directly compared the effect of ATX in the two datasets, we observed a spatial correspon-
dence between the effects of ATX on network topology during rest and task. Specifically, IL
regional topological signature observed during the N-back task was inversely correlated with
the regional signature observed during rest (r = −0.147 [bootstrapped 95% CI: −0.246 to
−0.037]; P < 0.002; Figure 2D), suggesting that ATX affected similar regions during rest and
task, albeit by shifting them in different topological directions.
Flexibility:
The proportion of time that an
individual node switches modules
over time
Differential Effects of Catecholamines on Network Flexibility as a Function of Cognitive State
Previous work has shown that topological flexibility is important for cognitive performance
(Cole et al., 2013; Medaglia, Lynall, & Bassett, 2015). That is, regions that are important for
defining network architecture should both play a crucial role in network topology (e.g., in-
terconnect otherwise disparate modules), while also maintaining temporal flexibility (e.g., so
that network resources can be deployed at the appropriate time). To test whether atomoxetine
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
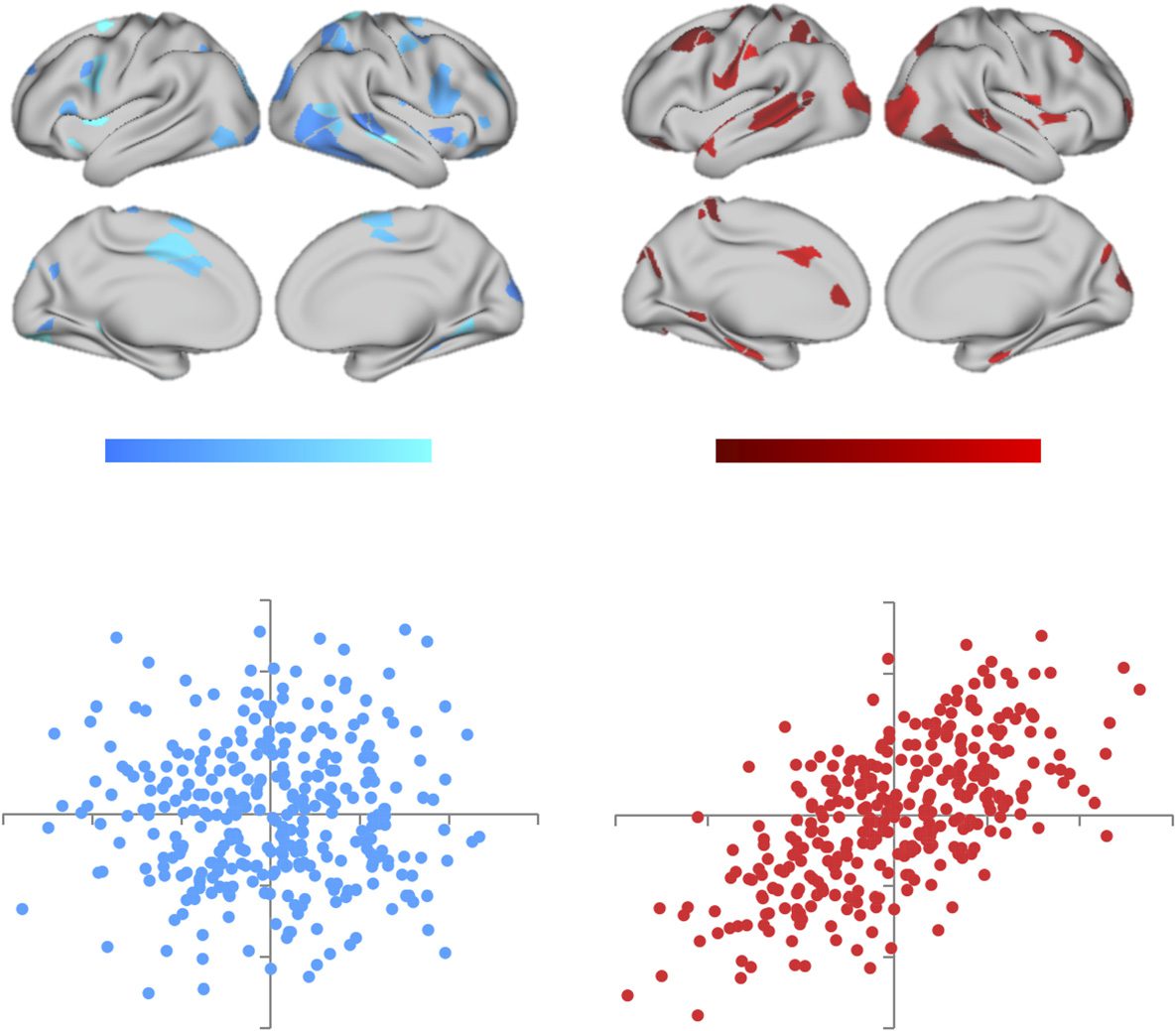
(A) Effect of atomoxetine (vs. placebo) on regional flexibility in the resting state (left, blue)
Figure 3.
and during the N-back task (right, red); regions depicted with increased flexibility (FDR q ≤ 0.05).
(B) Correlation between effect of atomoxetine (vs. placebo) on BT and regional flexibility during
rest (left, blue; r = 0.02) and during the N-back task (right, red; r = 0.61); the difference between
the two correlations was also significant (ZI* = 2.08; p = 0.037).
Network Neuroscience
386
Catecholaminergic manipulation and cognitive states
administration exerted a significant effect on time-resolved network topology as a function of
cognitive state, we calculated the topological flexibility of each region following atomoxetine
(vs. placebo) under both task conditions. Specifically, we estimated the flexibility of each
brain parcel by calculating the percentage of temporal windows in which an individual region
“switched” between modules over the course of each session. We found that atomoxetine
caused an increase in flexibility in a distributed network of regions across temporal, parietal,
and frontal cortex, with distinct signatures in rest and task (Figure 3A). Interestingly, the ad-
ministration of atomoxetine caused an increase in flexibility and integration in the same set of
regions during the N-back task, but not during rest (ZI* = 2.08; p = 0.037; Figure 3B), suggest-
ing that heightened free catecholamine levels may have facilitated network-level integration,
but only during situations with tight cognitive constraints.
DISCUSSION
Our results provide direct evidence that the manipulation of catecholamine levels in the hu-
man brain leads to substantial shifts in network topology. Furthermore, we were also able to
demonstrate that the alterations in network topology critically depend on cognitive state. In the
resting state, an overabundance of free catecholamine levels following ATX administration was
associated with a relatively segregated network topology (Figure 1A). The lack of effortful cog-
nitive engagement during the resting state may have facilitated a decrease in ascending arousal
via ATX-mediated autoinhibition of the locus coeruleus (Bari & Aston-Jones, 2013; Sara, 2009),
allowing the network to settle into a segregated architecture, potentially as a way to minimize
energy expenditure (Bullmore & Sporns, 2012). In contrast, when presented with a complex
cognitive challenge following ATX, an increase in the phasic-to-tonic ratio of noradrenergic
function (Bari & Aston-Jones, 2013) may have facilitated functional connectivity between
otherwise segregated circuits, integrating the functional connectome (Figure 2A) and putatively
increasing the temporal coordination between the brain circuitry required to successfully com-
plete the N-back task. Together, these results thus provide evidence that the ascending arousal
system mediates the balance between network-level integration and segregation as a function
of cognitive demands.
The administration of atomoxetine also differentially effected time-varying network topology
as a function of cognitive state. During the resting state, atomoxetine affected switching and
network integration relatively independently (Figure 3B). In contrast, the administration of
atomoxetine during N-back task performance led to an increase in topological flexibility in
the same regions that showed an increase in network-level integration (as measured by BT;
Figure 3B). These results suggest that fluctuating levels of catecholaminergic neurotransmitters
shape the spatiotemporal architecture of the brain in a manner that is sensitive to ongoing
cognitive demands (Shine & Poldrack, in press). However, it bears mention that some of the
observed differences between rest and task in our study may have been due to dose-related
differences, particularly given the known differences in the sensitivity of different classes of
noradrenergic receptors (Robbins & Arnsten, 2009). Irrespective of any potential dose-related
effects, our results do provide empirical support for the hypothesis that fluctuations in nor-
adrenaline are responsible for reconfiguring the network architecture of the brain. Indeed, the
shift toward higher integration is consistent with the existing hypothesis that heightened phasic
noradrenergic responses bias the network toward salient stimuli by modulating the sensitivity
of the network to incoming sensory input (Sara & Bouret, 2012).
The precise biological mechanism underlying these effects is currently a topic of active inves-
tigation, but there is emerging evidence that the network-level impact of catecholamines may
relate to their ability to modulate the “gain” of neurons across the brain (Aston-Jones & Cohen,
Network Neuroscience
387
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
2005; McGinley et al., 2015b). For instance, it has previously been shown that stimulation
of the locus coeruleus, both using electrical (Toussay, Basu, Lacoste, & Hamel, 2013) and
optogenetic approaches (Carter et al., 2010), leads to the widespread activation of the cortex,
producing a high-frequency, low-amplitude signature that is a known correlate of the awake
brain (Berridge & Waterhouse, 2003). Similar patterns have also been observed during spon-
taneous activity in awake mice, confirming that firing in the locus coeruleus directly facilitates
high-frequency cortical activity during natural behavior (Safaai et al., 2015). We recently used
a biophysical computational model to show that manipulating neural gain transitioned the
network from a segregated to an integrated architecture (Shine et al., 2018). The results of this
study are consistent with these findings, and further confirm the role of catecholaminergic tone
in simultaneously balancing the key topological properties of integration and segregation in a
state-dependent manner. They also provide a mechanistic explanation for the brain’s response
to periods of acute stress, which are also mediated by ascending noradrenergic systems
(Hermans et al., 2011). Future studies will play an important role in solidifying this mech-
anistic explanation and determining the contexts in which the balance between these factors
is most crucial for understanding complex behavior.
Although our experimental results suggest a crucial role for noradrenaline in the topological
reconfiguration of brain network architecture, it bears mention that biological systems rarely
demonstrate sharp boundaries between function systems. For instance, in addition to modu-
lating noradrenaline, ATX administration has also been shown to modulate the central con-
centrations of other arousal-related neurotransmitters, including serotonin (Ding et al., 2014),
histamine (Liu et al., 2008), and dopamine (Robbins & Arnsten, 2009), suggesting that the ef-
fects observed in our study may relate to the reconfiguration of the ascending arousal system
as a whole. This systemic interdependence is perhaps best exemplified when comparing the
relationship between noradrenaline and dopamine, the two major catecholaminergic neuro-
transmitters in the central nervous system. Whereas the majority of dopaminergic synapses
utilize their own specific transporter (Chen & Reith, 2000), a subgroup of dopaminergic ter-
minals in the cortex can also exploit noradrenergic transporters to reenter presynaptic axons
(Morón, Brockington, & Wise, 2002).
In addition, it has been shown that locus coeruleus
neurons can corelease noradrenaline and dopamine (Devoto, Flore, Pira, & Longu, 2004). As
such, our results may reflect the combined improvements in cortical signal-to-noise that relate
to some combination of dopaminergic and noradrenergic effects on neuronal projection tar-
gets (Robbins & Arnsten, 2009). The different concentrations of ATX used in the two studies
may also have affected these nonselective aspects of ATX. Fortunately, future studies that con-
trast the roles of the related neurotransmitter systems at different concentrations across a range
of cognitive states will help to clarify this issue.
Together, our results demonstrate a relationship between network topology and catecholamin-
ergic function that is sensitive to cognitive state. Future experiments should now be designed
to decipher the relative impact of other neurotransmitter systems, both in health and disease.
METHODS
Resting-State Study
Twenty-four right-handed individuals (age 19–26 years; 5 men) were included
Participants.
in this study. All participants were screened by a physician for physical health and drug
contraindications. Exclusion criteria included the following: standard contraindications for
MRI; current use of psychoactive or cardiovascular medication; a history of psychiatric ill-
ness or head trauma; cardiovascular disease; renal failure; hepatic insufficiency; glaucoma;
Network Neuroscience
388
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
hypertension; drug or alcohol abuse; learning disabilities; poor eyesight; smoking >5 cigarettes
a day; and current pregnancy. All participants gave written, informed consent before the
experiment.
Study design. We used a double-blind placebo-controlled crossover design (van den Brink
et al., 2016). In each of two sessions, scheduled 1 week apart at the same time of day, partici-
pants received either a single oral dose of atomoxetine (40 mg) or placebo (125 mg of lactose
monohydrate with 1% magnesium stearate, visually identical to the drug). In both sessions,
participants were scanned once before pill ingestion (t = −20 min) and once following inges-
zione (t = 90 min), when approximate peak plasma levels were reached. Each scan comprised
8 min of eyes-open resting-state fMRI. During scanning, the room was dark, and participants
fixated on a black fixation cross presented on a gray background. Drug uptake was confirmed
using cortisol and α-amylase levels in the saliva (van den Brink et al., 2016; Warren et al.,
2017).
All MRI data were collected with a Philips 3T MRI scanner. In each of the scan-
MRI data.
ning sessions, we collected T2*-weighted EPI resting-state images (echo time 30 ms, repetition
time 2.2 S, flip angle 80◦
, FOV 80 × 80 × 38 voxels of size 2.75 mm isotropic, E 216 vol-
umes). To allow magnetic equilibrium to be reached, the first five volumes were automati-
cally discarded. Inoltre, each time the participant entered the scanner, we collected a b0
field inhomogeneity scan (echo time 3.2 ms, repetition time 200 ms, flip angle 30◦
, and FOV
256 × 256 × 80 voxels with a reconstructed size of 0.86 × 0.86 mm with 3-mm-thick slices).
Finalmente, at the start of the first session, we collected a high-resolution anatomical T1 image
(echo time 4.6 ms, repetition time 9.77 ms, flip angle 8◦
, and FOV 256 × 256 × 140 voxels
with size 0.88 × 0.88 mm with 1.2-mm-thick slices) for image normalization and registration.
After realignment (using FSL’s MCFLIRT) and skull stripping (using BET),
Data preprocessing.
b0 unwarping was used to control for potential differences in head position across sessions.
The b0 scans were first reconstructed into an unwrapped phase angle and magnitude im-
age. The phase image was then converted to units of radians per second and median filtered,
and the magnitude image was skull-stripped. We then used FEAT to unwarp the EPI images in
the y-direction with a 10% signal loss threshold and an effective echo spacing of 0.333. IL
unwarped EPI images were then prewhitened, smoothed at 5 mm FWHM, and coregistered
with the anatomical T1 to 2-mm isotropic MNI space (degrees of freedom: EPI to T1, 3; T1/EPI
to MNI, 12). FMRIB’s ICA-based X-noiseifier (Salimi-Khorshidi, Douaud, & Beckmann, 2014)
was used with pretrained weights to de-noise the imaging data.
Temporal artifacts were identified in each dataset by calculating framewise displacement (FD)
from the derivatives of the six rigid-body realignment parameters estimated during standard
volume realignment (Power et al., 2014), as well as the root-mean-square change in BOLD
signal from volume to volume (DVARS). Frames associated with FD > 0.25 mm or DVARS >
2.5% were identified; Tuttavia, as no participants were identified with greater than 10% Di
the resting time points exceeding these values, no trials were excluded from further analysis.
There were no differences in head motion parameters between the four sessions (p > 0.500).
Following artifact detection, nuisance covariates associated with the six linear head move-
ment parameters (and their temporal derivatives), DVARS, physiological regressors (created
using the RETROICOR method), and anatomical masks from the CSF and deep cerebral WM
were regressed from the data by using the CompCor strategy (Behzadi, Restom, Liau, & Liu,
Network Neuroscience
389
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
N
e
N
_
UN
_
0
0
0
4
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Catecholaminergic manipulation and cognitive states
2007). Finalmente, in keeping with previous time-resolved connectivity experiments (Bassett, Yang,
Wymbs, & Grafton, 2015), a temporal band-pass filter (0.01 < f < 0.125 Hz) was applied to
the data. In addition, all results were replicable both with and without frame censoring at the
predefined levels, as well as with and without global signal regression.
Following preprocessing,
the mean time series was extracted from
Brain parcellation.
375 predefined regions-of-interest (ROIs). To ensure whole-brain coverage, we extracted the
following: 333 cortical parcels (161 and 162 regions from the left and right hemispheres,
respectively) by using the Gordon atlas (Gordon et al., 2014), 14 subcortical regions from
Harvard-Oxford subcortical atlas (bilateral thalamus, caudate, putamen, ventral striatum,
globus pallidus, amygdala, and hippocampus; http://fsl.fmrib.ox.ac.uk/), and 28 cerebellar re-
gions from the SUIT atlas (Diedrichsen et al., 2009) for each participant in the study.
To estimate time-resolved functional connectivity
Time-resolved functional connectivity.
(Hutchison et al., 2013; Sako˘glu et al., 2010) between the 375 ROIs, we used the multipli-
cation of temporal derivatives approach (MTD; http://github.com/macshine/coupling/; Shine
et al., 2015). The MTD is computed by calculating the pointwise product of temporal deriva-
tive of pairwise time series (Equation 1). To reduce the contamination of high-frequency noise
in the time-resolved connectivity data, the MTD is averaged by calculating the mean value
over a temporal window, w. Time-resolved functional connectivity was calculated between
all 375 brain regions by using the MTD within a sliding temporal window of 15 time points
(33 seconds at 2.2 s per window), which allowed for estimates of signals amplified at ap-
proximately 0.1 Hz. Individual functional connectivity matrices were then calculated within
each temporal window, thus generating an unthresholded (i.e., signed and weighted) three-
dimensional adjacency matrix (region × region × time) for each participant. The MTD for the
pairwise interaction between region i and j is defined according to Equation 1:
MTDijt =
1
w ∑t+w/2
t−w/2
(cid:2)
(cid:4)
dtit × dtjt
× σdtj
σdti
(cid:3)
(cid:5)
(1)
where dt is the first temporal derivative of the ith
time series at time t, σ is the standard
deviation of the temporal derivative time series for region i or j, and w is the window length
of the simple moving average. This equation can then be calculated over the course of a time
series to obtain an estimate of time-resolved connectivity between pairs of regions.
or jth
The Louvain modularity algorithm from the Brain Con-
Time-resolved community structure.
nectivity Toolbox (Rubinov & Sporns, 2010) was used to estimate both time-averaged and
time-resolved community structure. The Louvain algorithm iteratively maximizes the modu-
larity statistic, Q, for different community assignments until the maximum possible score of
Q has been obtained (Equation 2). The modularity estimate for a given network is therefore a
quantification of the extent to which the network may be subdivided into communities with
stronger within-module than between-module connections.
QT =
(cid:4)
1
v+ ∑ij
(cid:5)
ij − e+
w+
ij
δMi Mj
−
1
v+ + v− ∑ij
(cid:4)
ij − e−
w−
ij
(cid:5)
δMi Mj
(2)
Equation 2 gives the Louvain modularity algorithm, where v is the total weight of the network
(sum of all negative and positive connections), wij is the weighted and signed connection
Network Neuroscience
390
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
between regions i and j, eij is the strength of a connection divided by the total weight of the
network, and δMiMj is set to 1 when regions are in the same community and 0 otherwise.
Superscripts
denote all positive and negative connections, respectively.
and
+
−
For each temporal window, the community assignment for each region was assessed 500 times,
and a consensus partition was identified using a fine-tuning algorithm from the Brain Connec-
tivity Toolbox (http://www.brain-connectivity-toolbox.net/ ). This then afforded an estimate of
both the time-resolved modularity (QT) and cluster assignment (CiT) within each temporal
window for each participant in the study. To define an appropriate value for the γ parameter,
we iterated the Louvain algorithm across a range of values (0.5–2.5 in steps of 0.1) for 100
iterations of a single subjects’ time-averaged connectivity matrix and then estimated the sim-
ilarity of the resultant partitions by using mutual information. Across the cohort there were
on average 3.9 ± 1.2 communities found in each window. A γ parameter of 1.1 provided the
most robust estimates of topology across these iterations, both at the group and individual sub-
ject level. Alternatively, a multilayer implementation of the Louvain algorithm could be used,
although this would require the tuning of a separate parameter, ω, that defines the strength of
connection between layers (Mucha, Richardson, Macon, Porter, & Onnela, 2010).
Based on time-resolved community assignments, we estimated within-
Cartographic profiling.
module connectivity by calculating the time-resolved module degree Z-score (WT; within-
module strength) for each region in our analysis (Equation 3) (Guimerà & Nunes Amaral, 2005).
WiT =
κiT − ´κsiT
sκsiT
(3)
Equation 3 gives the module degree Z-score, WiT, where κiT is the strength of the connections
of region i to other regions in its module si at time T, ´κsiT is the average of κ over all the regions
in si at time T, and σκsiT is the standard deviation of κ in si at time T.
To estimate between-module connectivity (BT), we used the participation coefficient, BT,
which quantifies the extent to which a region connects across all modules (i.e., between-
module strength):
BiT = 1 − ∑nM
s=1
(cid:6)
(cid:7)2
κisT
κiT
(4)
Equation 4 gives the participation coefficient BiT, where κ
isT is the strength of the positive
connections of region i to regions in module s at time T, and κ
iT is the sum of strengths of all
positive connections of region i at time T. The participation coefficient of a region is therefore
close to 1 if its connections are uniformly distributed among all the modules and 0 if all of its
links are within its own module.
To track fluctuations in cartography over time, for each temporal window, we computed a joint
histogram of within- and between-module connectivity measures, which we refer to here as
a “cartographic profile” (Figure 1A). Code for this analysis is freely available at https://github.
com/macshine/integration/.
Pupil size was measured from the right eye at 500 Hz with an MRI-compatible
Pupilometry.
Eyelink 1000 eye tracker. Blinks and other artifacts were interpolated offline by using shape-
preserving piecewise cubic interpolation. Pupil data were low-pass filtered at 5 Hz to re-
move high-frequency noise and Z-scored across conditions. Five participants were excluded
from pupil-related analyses because of poor signal quality ( ≥50% of continuous time series
Network Neuroscience
391
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
interpolated) or missing data. Of the remaining participants, on average 20% ± 9% of the data
were interpolated. We replicated our previous observation (Shine et al., 2016) of a positive
relationship between pupil diameter and integrated network topology (Supporting Information
Figure S2a, Shine et al., 2018).
To determine whether the integrative signature of the brain was more
Null model creation.
dynamic than predicted by a stationary null model (Supporting Information Figure S2b, Shine
et al., 2018; Laumann, Snyder, Mitra, & Gordon, 2016); surrogate data was created using a
stationary Vector Auto Regressive model (order was set at 6 to match the expected temporal
signature of the BOLD response in 2.2 s TR data). The mean covariance matrix across the
entire experiment was used to generate 2,500 independent null datasets, which allows for the
appropriate estimation of the tails of nonparametric distributions (Nichols & Holmes, 2002).
These time series were then preprocessed using the same approach outlined for the BOLD data.
For each analysis, we estimated the kurtosis of the mean BT time series for each of the 2,500
simulations. We then calculated the 95th percentile of this distribution and used this value
to determine whether the resting-state data fluctuated more frequently than the null model. We
found that the dynamic network structure within the fMRI data had a higher kurtosis than the
95th percentile of the stationary null model (Supporting Information Figure S2b, Shine et al.,
2018), suggesting the presence of time-varying patterns in the brain. However, this inter-
pretation requires some caution, given that stationary null models are often unable to de-
termine whether interesting time-varying changes are indeed occurring in time series data
(Liégeois et al., 2017; Miller et al., 2017).
Statistical analyses.
Framework (https://osf.io/utqq2/ ):
The following hypotheses were preregistered with the Open Science
Hypothesis 1: To explicitly test whether the resting brain fluctuates more frequently than a
stationary null model, we calculated the kurtosis of the window-to-window difference in the
mean BT score for each iteration of a vector autoregression (VAR) null model (model order = 6).
The mean covariance matrix across all 24 subjects from the preplacebo session was be used
to generate 2,500 independent null datasets, which allows for the appropriate estimation of
the tails of nonparametric distributions (Nichols & Holmes, 2002). These time series was then
be filtered in a similar fashion to the BOLD data. For each analysis, we created a statistic for
each independent simulation that summarized the extent of fluctuations in the null dataset.
We then calculated the 95th percentile of this distribution and used this value to determine
whether the resting-state data fluctuated more frequently than the null model (i.e., whether
there deviations as extreme as the 95th percentile of the null dataset occur more than 5% of
the time).
Hypothesis 2: We estimated the Spearman’s rho correlation between the convolved pupil
diameter and the time series of each bin of the cartographic profile. We then fitted a linear
mixed-effects model with random intercept to determine whether the correlation between each
bin of the cartographic profile was more extreme than chance levels (FDR q ≤ 0.05).
Hypothesis 3: Group level differences were investigated by comparing each bin of the carto-
graphic profile for all subjects prior to and postatomoxetine administration, as well as prior
to and postplacebo using a 2 × 2 ANOVA design. Specifically, the percentage of time that
each bin of the cartographic profile was occupied during the resting state for each of the
four sessions was be entered into a 2 × 2 ANOVA. Our hypothesis predicted a significant
Network Neuroscience
392
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
interaction effect between pre- and postplacebo and pre- and postatomoxetine administration.
We corrected for multiple comparisons by using a false discovery rate of q ≤ 0.05. Similar
interaction effects were assessed at the regional level (i.e., regional topological measures, such
as participation coefficient) by using a series of 2 × 2 ANOVA designs.
Task-Based Study
Based on our interim results, we hypothesized that if phasic and tonic noradrenaline release
differentially alter the balance between integration and segregation, then integration should
be stronger following ATX during cognitive task performance. This hypothesis was not identi-
fied as part of our preregistration, but arose as a post hoc interrogation of the data. To test this
hypothesis, we analyzed data from a different dataset, in which 19 participants (age 18–30,
all right-handed males) underwent a cognitively challenging N-back task following either ATX
(60 mg) or placebo in a double-blind, randomized placebo-controlled crossover design
(PLC-ATX n = 8; ATX-PLC n = 11; Hernaus et al., 2017). The study was carried out in
accordance with the Declaration of Helsinki and was approved by the local medical ethics
committee of Maastricht University Medical Centre (NL53913.068.15). All participants gave
written, informed consent prior to each session and were reimbursed for participation.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Participants performed a parametrically modulated N-back task, in which they
Cognitive task.
were required to identify target letters that were presented for 1,000 ms on an LCD screen
during MRI scanning. Targets consisted of letters that were the same as the letter presented one,
two, or three trials previously (i.e., 1-back, 2-back, or 3-back, respectively). A further control
condition was also involved, in which participants were asked to detect the letter ‘X’ (i.e.,
0-back). Every task condition was presented three times in pseudorandom order (3–4 targets
per block). Participants responded to targets and distractors with right index and middle finger
button presses, respectively. Task effects were modeled for fMRI analysis by using a block
design.
All MRI data were collected with a Siemens 3T MRI scanner.
MRI data and preprocessing.
In each of the scanning sessions, we collected T2*-weighted EPI images (echo time 30 ms,
repetition time 2.0 s, flip angle 77◦, and 286 volumes). To allow magnetic equilibrium to
be reached, the first five volumes were automatically discarded.
In addition, each time the
participant entered the scanner, we collected a b0 field inhomogeneity scan (echo time 3.2 ms,
, and FOV 256 × 256 × 80 voxels with a reconstructed
repetition time 200 ms, flip angle 30
size of 0.86 × 0.86 mm with 3-mm-thick slices). Finally, at the start of the first session, we
collected a high-resolution anatomical T1 image (echo time 4.6 ms, repetition time 9.77 ms,
flip angle 8◦
, and FOV 256 × 256 × 140 voxels with size 0.88 × 0.88 mm with 1.2-mm-thick
slices) for image normalization and registration. Data were preprocessed in a similar fashion to
the resting-state analysis, albeit without correction for physiological parameters, which were
not collected in the study.
◦
Preprocessed BOLD data were subjected to the same time-resolved network
fMRI analysis.
analysis pipeline as to the one utilized for the resting-state analysis. Following this step, both
regional (WT and BT) and global (cartographic profile) time series were modeled against the
blocks of the 0-, 1-, 2-, and 3-back conditions in both the post-ATX and postplacebo ses-
sions. Instruction screens, rest blocks, and head motion parameters (6 linear parameters and
their temporal derivatives) were also modeled. We then statistically compared the resultant
β weights for each of the blocks separately using a series of F tests (FDR q ≤ 0.05) with the
Network Neuroscience
393
Catecholaminergic manipulation and cognitive states
following two contrasts: i) main effects, which were modeled as the mean activity in the 1-,
2-, and 3-back blocks versus the 0-back block; and ii) load effects, which were modeled as a
parametric increase in activity as a function of cognitive load across the four blocks. None of
the effects were significantly correlated with head motion, either within- or between-subjects
(p > 0.5 ).
Finalmente, we correlated the β weights for the main effect of ATX > PLC during the task for
each parcel with the interaction effect of ATX[Post>Pre] > PLC[Post>Pre] on resting-state topology
by using a Pearson’s correlation (the task data did not contain a “predrug” condition). IL
significance of this correlation was determined by randomly permuting the task-based effects
5,000 times and then reestimating the Pearson’s correlation between the shuffled effects and
the original regional effects in the resting state. The inverse correlation between the two parcel
values was more extreme than the 0.2nd percentile of the null distribution (cioè., P < 0.002). A
nonparametric bootstrapping approach was also used to estimate a 95% confidence interval
for the correlation (1,000 iterations).
Regional flexibility. We estimated the flexibility of each brain parcel by calculating the per-
centage of temporal windows in which an individual region “switched” between modules,
normalized to the total number of modules in the data (as estimated in the previous step).
Code was obtained directly from the original author (http://www.danisbassett.com/resources/).
As the modular assignment was essentially arbitrary within each unique temporal window, we
used a version of the Hungarian algorithm (Kuhn, 1955), which is a combinatorial optimization
algorithm that calculates the most efficient path between different layers of a multislice net-
work, to assign regions to modules with consistent values over time. Regional flexibility values
were then compared across atomoxetine and placebo trials. Pearson’s correlations were later
used to compare the regional signature of flexibility with regional BT values, and the Dunn
and Clark statistic was used to compare the correlations between rest and task in both datasets
(Bayer, 2016).
AUTHOR CONTRIBUTIONS
James Shine: Conceptualization; Data curation; Formal analysis; Investigation; Methodol-
ogy; Visualization. Rudy van den Brink: Data curation; Investigation; Methodology. Sander
Niewenhuis: Data curation; Investigation; Methodology. Russell Poldrack: Conceptualization;
Methodology; Visualization.
FUNDING INFORMATION
James Shine was supported by an NHMRC CJ Martin Fellowship (GNT1072403).
REFERENCES
Aston-Jones, G., & Cohen, J. D. (2005). An integrative theory of lo-
cus coeruleus-norepinephrine function: Adaptive gain and opti-
mal performance. Annual Review of Neuroscience, 28(1), 403–450.
Bari, A., & Aston-Jones, G. (2013). Atomoxetine modulates spon-
taneous and sensory-evoked discharge of locus coeruleus nor-
adrenergic neurons. Neuropharmacology, 64, 53–64.
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T. (2015).
Learning-induced autonomy of sensorimotor systems. Nature
Neuroscience, 18(5), 744–751. https://doi.org/10.1038/nn.3993
Bayer, A. (2016). Applied statistics analysis of variance and regres-
sion. Applied Statistics.
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. NeuroImage, 37(1), 90–101.
Berridge, C. W., & Waterhouse, B. D. (2003). The locus coeruleus–
noradrenergic system: Modulation of behavioral state and state-
dependent cognitive processes. Brain Research Reviews, 42(1),
33–84.
Network Neuroscience
394
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
Bullmore, E., & Sporns, O. (2012). The economy of brain network
organization. Nature Reviews Neuroscience, 13(5), 336–349.
Carter, M. E., Yizhar, O., Chikahisa, S., Nguyen, H., Adamantidis,
(2010). Tuning arousal with optogenetic
A., Nishino, S., et al.
modulation of locus coeruleus neurons. Nature Neuroscience,
13(12), 1526–1533.
Chen, N., & Reith, M. (2000). Structure and function of the dopamine
transporter. European Journal of Pharmacology, 405, 329–339.
Cole, M. W., Reynolds, J. R., Power, J. D., Repovs, G., Anticevic,
A., & Braver, T. S. (2013). Multi-task connectivity reveals flexi-
ble hubs for adaptive task control. Nature Neuroscience, 16(9),
1348–1355.
Cools, R., & D’Esposito, M. (2011). Inverted-U-shaped dopamine
actions on human working memory and cognitive control.
Biological Psychiatry, 69(12), e113–e125.
de Pasquale, F., Penna, Della S., Sporns, O., Romani, G. L., &
Corbetta, M. (2016). A dynamic core network and global efficiency
in the resting human brain. Cerebral Cortex, 26(10), 4015–4033.
Devilbiss, D. M., & Waterhouse, B. D. (2011). Phasic and tonic pat-
terns of locus coeruleus output differentially modulate sensory
network function in the awake rat. Journal of Neurophysiology,
105(1), 69–87.
Devoto, P., Flore, G., Pira, L., & Longu, G.
(2004). Alpha2-
adrenoceptor mediated co-release of dopamine and noradrena-
line from noradrenergic neurons in the cerebral cortex. Journal
of Neuroscience, 88, 1003–1009.
Diedrichsen, J., Balsters, J. H., Flavell, J., Cussans, E., & Ramnani,
(2009). A probabilistic MR atlas of the human cerebellum.
N.
NeuroImage, 46(1), 39–46.
Ding, Y. S., Naganawa, M., Gallezot, J.-D., Nabulsi, N., Lin, S.-F.,
Ropchan, J., et al. (2014). Clinical doses of atomoxetine signifi-
cantly occupy both norepinephrine and serotonin transports: Im-
plications on treatment of depression and ADHD. NeuroImage,
86, 164–171.
Eldar, E., Cohen, J. D., & Niv, Y. (2013). The effects of neural gain on
attention and learning. Nature Neuroscience, 16(8), 1146–1153.
Gordon, E. M., Laumann, T. O., Adeyemo, B., Huckins, J. F., Kelley,
W. M., & Petersen, S. E. (2014). Generation and evaluation of a
cortical area parcellation from resting-state correlations. Cerebral
Cortex, bhu239.
Guimerà, R., & Nunes Amaral, L. A. (2005). Functional cartography
of complex metabolic networks. Nature, 433(7028), 895–900.
Hermans, E. J., van Marle, H., & Ossewaarde, L. (2011). Stress-
related noradrenergic activity prompts large-scale neural net-
work reconfiguration. Science, 334, 1151–1153.
Hernaus, D., Casales Santa, M. M., Offermann,
J. S., &
Van Amelsvoort, T. (2017). Noradrenaline transporter block-
ade increases fronto-parietal functional connectivity relevant for
working memory. European Neuropsychopharmacology, 27(4),
399–410.
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M., et al. (2013). Dynamic functional
connectivity: Promise, issues, and interpretations. NeuroImage,
80, 360–378.
Invernizzi, R. W., & Garattini, S. (2004). Role of presynaptic α2-
recent findings from
adrenoceptors in antidepressant action:
microdialysis studies. Progress in Neuro-Psychopharmacology
and Biological Psychiatry, 28(5), 819–827.
Joshi, S., Li, Y., Kalwani, R. M., & Gold, J. I. (2016). Relationships
between pupil diameter and neuronal activity in the locus coeru-
leus, colliculi, and cingulate cortex. Neuron, 89(1), 221–234.
Kitzbichler, M. G., Henson, R. N. A., Smith, M. L., Nathan, P. J.,
& Bullmore, E. T. (2011). Cognitive effort drives workspace con-
figuration of human brain functional networks. The Journal of
Neuroscience, 31(22), 8259–8270.
Kuhn, H. W.
(1955). The Hungarian method for the assignment
problem. Naval Research Logistics, 2(1), 83–97.
Laumann, T. O., Snyder, A. Z., Mitra, A., & Gordon, E. M. (2016).
On the stability of bold fmri correlations. Cerebral Cortex, 27(10),
4719–4732.
Liégeois, R., Laumann, T. O., Snyder, A. Z., Zhou, J., & Yeo, B. T. T.
(2017). Interpreting temporal fluctuations in resting-state func-
tional connectivity MRI. NeuroImage, 163, 437–455.
Liu, L.-L., Yang, J., Lei, G.-F., Wang, G.-J., Wang, Y.-W., & Sun,
R.-P. (2008). Atomoxetine increases histamine release and im-
proves learning deficits in an animal model of attention-deficit
hyperactivity disorder: the spontaneously hypertensive rat. Basic
& Clinical Pharmacology & Toxicology, 102(6), 527–532.
Mattar, M. G., Cole, M. W., Thompson-Schill, S. L., & Bassett,
D. S. (2015). A functional cartography of cognitive systems. PLoS
Computational Biology, 11(12), e1004533.
McGinley, M. J., David, S. V., & McCormick, D. A. (2015a). Cortical
membrane potential signature of optimal states for sensory signal
detection. Neuron, 87(1), 179–192.
McGinley, M. J., Vinck, M., Reimer, J., Batista-Brito, R., Zagha,
E., Cadwell, C. R., et al. (2015b). Waking state: Rapid varia-
tions modulate neural and behavioral responses. Neuron, 87(6),
1143–1161.
Medaglia, J. D., Lynall, M. E., & Bassett, D. S. (2015). Cognitive
network neuroscience. Journal of Cognitive Neuroscience, 27(8),
1471–1491.
Miller, R. L., Adali, J., Levin-Schwarz, Y., & Calhoun, V. D. (2017).
Resting-state fMRI dynamics and null models: Perspectives,
sampling variability, and simulations. biorxiv.
Morón, J. A., Brockington, A., & Wise, R. A. (2002). Dopamine up-
take through the norepinephrine transporter in brain regions with
low levels of the dopamine transporter: Evidence from knock-out
mouse lines. Journal of Neuroscience, 22, 389–395.
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J.-P. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Science, 328(5980), 876–878.
Nichols, T. E., & Holmes, A. P. (2002). Nonparametric permuta-
tion tests for functional neuroimaging: a primer with examples.
Human Brain Mapping, 15(1), 1–25.
Nieuwenhuis, S., De Geus, E. J., & Aston-Jones, G. (2011). The
anatomical and functional relationship between the P3 and auto-
nomic components of the orienting response. Psychophysiology,
48(2), 162–175.
Power, J. D., Mitra, A., Laumann, T. O., Snyder, A. Z., Schlaggar,
B. L., & Petersen, S. E. (2014). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImage,
84, 320–341.
Network Neuroscience
395
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Catecholaminergic manipulation and cognitive states
Reimer, J., Froudarakis, E., Cadwell, C. R., Yatsenko, D., Denfield,
G. H., & Tolias, A. S. (2014). Pupil fluctuations track fast switch-
ing of cortical states during quiet wakefulness. Neuron, 84(2),
355–362.
Reimer, J., McGinley, M. J., Liu, Y., Rodenkirch, C., Wang, Q.,
McCormick, D. A., & Tolias, A. S. (2016). Pupil fluctuations track
rapid changes in adrenergic and cholinergic activity in cortex.
Nature Communications, 7, 13289.
Robbins, T. W., & Arnsten, A. F. T. (2009). The neuropsychophar-
macology of fronto-executive function: Monoaminergic modu-
lation. Annual Review of Neuroscience, 32(1), 267–287.
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069.
Safaai, H., Neves, R., Eschenko, O., Logothetis, N. K., & Panzeri, S.
(2015). Modeling the effect of locus coeruleus firing on corti-
cal state dynamics and single-trial sensory processing. Proceed-
ings of the National Academy of Sciences of the United States of
America, 112(41), 12834–12839.
Sako˘glu, U., Pearlson, G. D., Kiehl, K. A., Wang, Y. M., Michael,
A. M., & Calhoun, V. D.
(2010). A method for evaluating
dynamic functional network connectivity and task-modulation:
application to schizophrenia. Magma (New York, N.Y.), 23(5-6),
351–366.
Salimi-Khorshidi, G., Douaud, G., & Beckmann, C. F. (2014). Auto-
matic denoising of functional MRI data: Combining indepen-
dent component analysis and hierarchical fusion of classifiers.
NeuroImage, 90, 449–468.
Samuels, E. R., & Szabadi, E. (2008). Functional neuroanatomy of
the noradrenergic locus coeruleus: Its roles in the regulation of
arousal and autonomic function Part II: Physiological and phar-
macological manipulations and pathological alterations of locus
coeruleus activity in humans. Current Neuropharmacology, 6(3),
254–285.
Sara, S. J. (2009). The locus coeruleus and noradrenergic modula-
tion of cognition. Nature Reviews Neuroscience, 10(3), 211–223.
Sara, S. J., & Bouret, S. (2012). Orienting and reorienting: The locus
coeruleus mediates cognition through arousal. Neuron, 76(1),
130–141.
Servan-Schreiber, D., Printz, H., & Cohen, J. (1990). A network
model of catecholamine effects- gain, signal-to-noise ratio, and
behavior. Science, 249, 892–895.
Shine, J. M., & Poldrack, R. A. (in press). Principles of dynamic net-
work reconfiguration across diverse brain states. NeuroImage.
Retrieved from https://www.sciencedirect.com/science/article/
pii/s1053811917306572?via%3dihub
Shine,
J. M., Aburn, M. J., Breakspear, M., & Poldrack, R. A.
(in press). The modulation of neural gain facilitates a transi-
tion between functional segregation and integration in the brain.
eLife. Retrieved from https://elifesciences.org/articles/31130
Shine, J. M., Bissett, P. G., Bell, P. T., Koyejo, O., Balsters, J. H.,
Gorgolewski, K. J., et al. (2016). The dynamics of functional brain
networks: Integrated network states during cognitive task perfor-
mance. Neuron, 92(2), 544–554.
Shine, J. M., Koyejo, O., Bell, P. T., Gorgolewski, K. J., Gilat, M., &
Poldrack, R. A. (2015). Estimation of dynamic functional connec-
tivity using Multiplication of Temporal Derivatives. NeuroImage,
122, 399–407.
Shine, J. M., van den Brink, R. L., Hernaus, D., Nieuwenhuis,
S., & Poldrack, R. A. (2018). Supporting Information for “Cat-
echolaminergic manipulation alters dynamic network topology
across cognitive states.” Network Neuroscience, 2(3), 381–396.
https://doi.org/10.1162/netn_a_00042
Toussay, X., Basu, K., Lacoste, B., & Hamel, E. (2013). Locus coe-
ruleus stimulation recruits a broad cortical neuronal network and
increases cortical perfusion. The Journal of Neuroscience, 33(8),
3390–3401.
van den Brink, R. L., Pfeffer, T., Warren, C. M., Murphy, P. R.,
Tona, K.-D., van der Wee, N. J. A., et al. (2016). Catecholamin-
ergic neuromodulation shapes intrinsic MRI functional connec-
tivity in the human brain. The Journal of Neuroscience, 36(30),
7865–7876.
J., & Stamatakis, E. A.
Vatansever, D., Menon, D. K., Manktelow, A. E., Sahakian,
B.
(2015). Default mode dynamics
for global functional integration. The Journal of Neuroscience,
35(46), 15254–15262.
Warren, C. M., van den Brink, R. L., Nieuwenhuis, S., & Bosch,
J. A. (2017). Norepinephrine transporter blocker atomoxetine
increases salivary alpha amylase. Psychoneuroendocrinology,
78, 233–236.
Waterhouse, B. D., Moises, H. C., & Woodward, D. J. (1980). No-
radrenergic modulation of somatosensory cortical neuronal re-
sponses to lontophoretically applied putative neurotransmitters.
Experimental Neurology, 69(1), 30–49.
Network Neuroscience
396
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
3
3
8
1
1
0
9
2
0
8
2
n
e
n
_
a
_
0
0
0
4
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3