RESEARCH
Cognition and connectomes in nondementia
idiopathic Parkinson’s disease
1
Luis M. Colon-Perez
2
, Jared J. Tanner
5
Thomas H. Mareci
, Michelle Couret
2
, and Catherine C. Price
3
4
, Shelby Goicochea
,
1
Department of Psychiatry, University of Florida, Gainesville, FL, USA
2Department of Clinical and Health Psychology, University of Florida, Gainesville, FL, USA
3Department of Medicine, Università della Columbia, New York, NY, USA
4Department of Medicine, University of Florida, Gainesville, FL, USA
5Department of Biochemistry and Molecular Biology, University of Florida, Gainesville, FL, USA
a n o p e n a c c e s s
j o u r n a l
Keywords: Connectome, Connectivity, Tractography, Structural networks, Parkinson’s disease,
Cognitive decline
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
ABSTRACT
In this study, we investigate the organization of the structural connectome in cognitively
well participants with Parkinson’s disease (PD-Well; n = 31) and a subgroup of
participants with Parkinson’s disease who have amnestic disturbances (PD-MI; n = 9).
We explore correlations between connectome topology and vulnerable cognitive
domains in Parkinson’s disease relative to non-Parkinson’s disease peers (controllo, n = 40).
Diffusion-weighted MRI data and deterministic tractography were used to generate
connectomes. Connectome topological indices under study included weighted indices of
node strength, path length, clustering coefficient, and small-worldness. Relative to controls,
node strength was reduced 4.99% for PD-Well (p = 0.041) E 13.2% for PD-MI (p = 0.004).
We found bilateral differences in the node strength between PD-MI and controls for inferior
parietal, caudal middle frontal, posterior cingulate, precentral, and rostral middle frontal.
Correlations between connectome and cognitive domains of interest showed that topological
indices of global connectivity negatively associated with working memory and displayed
more and larger negative correlations with neuropsychological indices of memory in PD-MI
than in PD-Well and controls. These findings suggest that indices of network connectivity are
reduced in PD-MI relative to PD-Well and control participants.
AUTHOR SUMMARY
Parkinson’s disease (PD) patients with amnestic mild cognitive impairment (per esempio., primary
processing-speed impairments or primary memory impairments) are at greater risk of
developing dementia. Recent evidence suggests that patients with PD and mild cognitive
impairment present an altered connectome connectivity. In this work, we further explore the
structural connectome of PD patients to provide clues to identify possible sensitive markers
of disease progression, and cognitive impairment, in susceptible PD patients. We employed a
weighted network framework that yields more stable topological results than the binary
network framework and is robust despite graph density differences, hence it does not require
thresholding to analyze the connectomes. As Supplementary Information (Colon-Perez et al.,
2017), we include databases sharing the results of the network data.
Citation: Colon-Perez, l. M., Tanner,
J. J., Couret, M., Goicochea, S., Mareci,
T. H., & Price, C. C. (2017). Cognition
and connectome in nondementia
idiopathic Parkinson’s disease.
Network Neuroscience. 2(1) 106–124.
https://doi.org/10.1162/netn_a_00027
DOI:
https://doi.org/10.1162/netn_a_00027
Supporting Information:
https://doi.org/10.1162/netn_a_00027
Received: 18 Febbraio 2017
Accepted: 18 settembre 2017
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Luis M. Colon-Perez
lcolon@ufl.edu
Copyright: © 2017
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
Connectomes in Parkinson’s disease
Structural connectome:
Brain connectivity graphs obtained
from diffusion MRI and tractography.
Tractography:
Three-dimensional representation of
the brain’s white matter tracts derived
from diffusion MRI data.
INTRODUCTION
Parkinson’s disease (PD) is a neurodegenerative disorder characterized by motor disruption
(cioè., tremors, unstable posture, bradykinesia) and cognitive symptoms (Chaudhuri & Schapira,
2009). Although James Parkinson initially proposed that PD did not progress to the cerebrum
(Parkinson, 2002), there is now substantial evidence that PD can compromise higher cortical
cognitive processes. The progression may not be the same for all individuals with PD, Tuttavia;
some individuals remain with primary processing-speed impairments while others may present
with primary memory impairments and are at greater risk for dementia (Henderson et al., 2016;
Janvin, Larsen, Aarsland, & Hugdahl, 2006). Neuroimaging investigations show regional frac-
tional anisotropy (FA) and volumetric gray and white matter (WM) differences in PD and
non-PD peers (Price et al., 2016; Tanner et al., 2015). PD may be a network-level disease
(Bellucci et al., 2016; Catani & Ffytche, 2005; Gratwicke, Jahanshahi, & Foltynie, 2015), E
the network heterogeneity possibly explains whether the subjects have memory impairment.
Novel applications with structural connectomes hold promise for improving our under-
standing of network heterogeneity within PD. Connectome studies represent the brain as a set
of nodes (brain areas) and edges (connecting white matter between brain areas) that quantify
the macroscopic topological organization of the brain network. The topological features of
the human connectome allow us to describe the complex interconnectedness of the human
brain in vivo. Connectome studies quantitatively describe the arrangement of connections in
the brain (Sporns, 2011B) and offer a novel approach to explore the brain in healthy and neu-
ropathological participants (Hagmann et al., 2010; Sporns, 2011UN). They have been used to
quantify the organization of connected white matter in neurological disorders such as Hunt-
ington’s disease (Odish et al., 2015), epilepsy (Taylor, Han, Schoene-Bake, Weber, & Kaiser,
2015), and Alzheimer’s disease (Daianu et al., 2015). Also, connectome topology has been
suggested as a sensitive biomarker for early stages of psychotic illness and the eventual de-
velopment of psychosis (Drakesmith et al., 2015). In individuals with PD who have a freezing
gait, maladaptive brain restructuring has been shown through the connectivity between lo-
comotor hubs, particularly in the supplementary motor area and mesencephalic locomotor
regions (Fling et al., 2014). A recent study showed that the connectome in patients with PD
with mild cognitive impairment (MCI) is altered (Galantucci et al., 2016). Hence, further struc-
tural connectome studies of PD may yield sensitive markers of disease progression, cognitive
impairment, and susceptibility to PD.
In this work, we performed global
(average values for the entire brain network for
each participant) and local
(average values for every individual node for each partici-
pant) connectome analyses. Given the elevated risk of developing dementia associated with
amnestic mild cognitive impairment in PD (Henderson et al., 2016), in this study we ex-
amined structural connectome differences in people with PD who are cognitively well
relative to individuals with PD who meet criteria for amnestic MCI. We also explored
the correlations between topological connectome indices and the most common cogni-
tive vulnerabilities of PD (cioè., processing speed, working memory, and episodic memory;
Zgaljardic, Borod, Foldi, & Mattis, 2003). We used diffusion MRI (Basser & Jones, 2002) E
tractography (Basser, Pajevic, Pierpaoli, Duda, & Aldroubi, 2000) to generate connectomes
comprising 82 brain regions (68 cortical and 14 subcortical). With this, we determined the
organization of structural connectivity of our three participant groups (PD, PD-MCI, non-PD
peers) and correlations between connectome indices and working memory, processing speed,
and verbal memory.
Network Neuroscience
107
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
Dementia Rating Scale-2:
A screening measure used to assess a
patient’s overall level of cognitive
functioning.
Unified Parkinson’s Disease Rating
Scale:
The most commonly used measure of
Parkinson’s disease clinical symptom
severity.
METHODS
Participants
This study was approved by the University of Florida Health Center Institutional Review Board
(Protocol #472-2007). Written consent was obtained from all participants, and all research
followed the Declaration of Helsinki.
to participate were included in the study.
Providers within the UF Center for Movement Disorders and Neurorestoration referred
nondemented individuals with idiopathic PD to the study. Structured telephone screening
was performed to verify inclusion/exclusion criteria. Potential participants were screened
in person with the Dementia Rating Scale-2 (DRS-2) (Matteau et al., 2011); a total DRS-2
score > 130 was required for participation. Only nondemented individuals who were able
to consent
right-handed
(Briggs & Nebes, 1975), DRS-2 raw score > 130, fluent English, diagnosis of PD by a move-
ment disorder neurologist, UK Parkinson’s Disease Society Brain Bank Clinical Diagnostic Cri-
teria (Hughes, Ben-Shlomo, Daniel, & Lees, 2001), and Hoehn and Yahr scale (Hoehn & Yahr,
1967) ranging from 1 A 3. Exclusion criteria: diseases likely to confound cognition (per esempio.,
cerebrovascular accident in the last six months), deep brain stimulation, secondary/atypical
Parkinsonism, and major psychiatric disorder. Depression and apathy were not exclusion cri-
teria because of their high prevalence in PD.
Inclusion criteria:
The final sample included 40 people with idiopathic PD and 40 non-PD peers. Diffusion
and gray matter structural data from some of these participants have also been seen in recent
publications (Crowley et al., 2017; Price et al., 2016; Schwab et al., 2015; Tanner, Levy, et al.,
2017; Tanner et al., 2015; Tanner, McFarland, et al., 2017).
Defining PD Subgroups
PD-Memory Impaired (PD-MI): From the measures described below, those with PD who had a
memory composite score ≤ −1.5 (relative to non-PD peers) were classified as PD-MI (n = 9).
All others with PD were classified as PD without memory impairment (PD-Well; n = 31).
Those with and without PD-MI are discussed elsewhere (Tanner et al., 2015).
Cognitive Measures
While on medication, participants completed cognitive testing, neuroimaging, and the Uni-
fied Parkinson’s Disease Rating Scale (UPDRS) to assess optimal performance and represent
normal functioning. All participants also completed tests on general cognition and mood,
PD symptoms and severity, comorbidity (Charlson, Pompei, Ales, & MacKenzie, 1987), E
a neuropsychological protocol. Medications were reverted to a common metric (levodopa
equivalency dose, LED; Tomlinson et al., 2010). Blinded raters scored the data twice. The UP-
DRS III is a measure of PD motor symptom severity and was used as a correlate with network
indices.
Primary cognitive domains of interest: PD is brain disorder of the frontal-subcortical areas
known to alter the frontal-striatal cognitive functions of processing speed and working memory.
These domains were assessed using composites of standardized neuropsychological measures:
Processing speed: based on standardized scores from the Trail Making Test, Part A (total
time; Heaton & Psychological Assessment Resources Inc., 2004), WAIS-III Digit Symbol
(total correct; Wechsler, 1991), and Stroop Color Word Test–Word Reading condition
(total correct; Golden & Freshwater, 2002).
Network Neuroscience
108
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
Prodromal dementia:
The earliest stage of
neurodegenerative disease when
there is a decline in memory or
cognition, but functional
independence remains intact.
Working memory: created from the Digit Span Backward (total span; Wechsler, 1991),
Spatial Span Backward (total score; Wechsler, 1991), and Letter Number Sequencing
(total correct; Wechsler, 1991).
In addition to frontal-striatal deficits, we examined connectome indices relative to declarative
memory abilities, as this is an essential domain of prodromal dementia.
Verbal memory: created from selected index scores from the 12-word version of the
Philadelphia (repeatable) Verbal Learning Test (P(R)VLT (Price et al., 2009) and Wechsler
Memory Scale 3rd Revision (WMS-III) Logical Memory (LM) (Wechsler, 1991).
MRI Acquisition and Processing
3
2
) E 64 directions (b = 1, 000 s/mm
Data were acquired with a Siemens 3 T Verio using an eight-channel head coil. Two T1-
weighted scans were used for node segmentation with scan parameters of 176 contiguous
slices, 1 mm
isotropic voxels, and TR/TE = 2,500/3.77 ms. Single-shot echo planar imag-
ing diffusion-weighted images were acquired for tractography with gradients applied along 6
2
(b = 100 s/mm
). Diffusion imaging parameters were set
3
at 73 contiguous axial slices with 8 mm
isotropic voxels and TR/TE = 17,300/81 ms. Node
segmentation was completed with FreeSurfer 5.3 and data were quality checked. The qual-
ity check for the diffusion data included visual inspection for artifacts (per esempio., signal loss in the
anterior and middle regions, Venetian blinding, checker boarding). No significant artifacts
were observed. The process was also repeated after eddy current correction (eddy_correct).
Participant head motion during diffusion sequences was quantified with four measures using
TRACULA (Yendiki, Koldewyn, Kakunoori, Kanwisher, & Fischl, 2013). Between-group regis-
tration and intensity-based metrics demonstrated no significant group differences in diffusion
sequence motion (Registration: average translation: t = 0.98, p = 0.33; average rotation:
χ2 = 1.25, p = 0.26; Intensity: Percentage bad slices χ2 = 0.26, p = 0.61; Average dropout
score χ2 = 0.26, p = 0.61), suggesting that data were appropriate for group comparisons.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
Diffusion data were preprocessed using in-house software written in IDL (Harris
Geospatial Solutions, Bloomfield, CO). Eddy current correction was performed using FSL
(Jenkinson, Beckmann, Behrens, Woolrich, & Smith, 2012). Diffusion tensor imaging met-
rics (fractional anisotropy, FA, and mean diffusivity, MD) were calculated using FSL.
For tractography, fiber orientation profiles were estimated based on the calculation of
diffusion displacement probability with a mixture of
the Wishart method outlined by
(2007). Diffusion images were interpolated
Jian, Vemuri, Ozarslan, Carney, and Mareci
3
(Meijering, Zuiderveld, & Viergever, 1999) A 1 mm
isotropic voxels using cubic convolution
and whole-brain deterministic fiber tracking initiated using 125 uniformly distributed stream-
line points per voxel.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge weight:
Graph theoretical representation of
the strength of the pairwise
connections between the nodes in a
graph.
Network Neuroscience
Network Preparation and Analysis
The network edges were weighted as described by Colon-Perez et al. (Colon-Perez, Spindler,
et al., 2015). The edge weight, w(e), defines connecting any two nodes is defined as
w(eij) =
(cid:3) (cid:4)
(cid:2)
Vvoxel
Pvoxel
2
Ai + Aj
(cid:5)
Pvoxel
∑
p=1
M
∑
m=1
χR( f p,M)
l( f p,M)
,
(1)
109
Connectomes in Parkinson’s disease
Characteristic function:
In probability and statistics, UN
function that precisely defines the
members of a set. It takes a value of
one for a member of the set and zero
for a nonmember.
Dove
χR( f p,M) =
(cid:6)
1, f p,m ∈ R
0, f p,m /∈ R
,
(2)
Vvoxel is the MR voxel volume, Pvoxel is the number of streamline seed points per voxel, UN
is the surface area of each node, M is the number of voxels making up the edge,
f p,m is
the streamline originating from seed point p in voxel m, l( f p,M) is the length of f p,M, R is
the set of streamlines that originate from the voxels making up the space occupied by the
WM path connecting nodes ni and nj, and χR( f p,M) is the characteristic function that en-
sures the streamlines connecting nodes ni and nj originate from the space (cioè., voxels) IL
streamlines traverse between nodes ni and nj. The edge weight (Equazione 1) eliminates the
bias effects of the length of the streamlines, the seeding paradigm (cioè., Pvoxel), image resolu-
zione (cioè., Vvoxel), and tractography-specific experimental factors from the calculation; for more
details we refer the reader to Colon-Perez, Spindler, et al. (2015). The characteristic function
(χR( f p,M), Equazione 2) eliminates those streamlines that originate within the nodes and voxels
that do not represent the WM path connecting the nodes. Also, this edge weight quantifies
the white matter strength between any two nodes in a dimensionless and scale-invariant man-
ner (Colon-Perez, Spindler, et al., 2015). The employed edge weight uses the strict criterion of
χR( f p,M) to determine the set streamlines used to quantify the strength of connectivity between
two nodes. Given the high level of false positives in tractography, this edge weight serves as a
layer of strict control to quantify the strength of connectivity between nodes.
The networks were analyzed in a weighted framework, as described in Colon-Perez, Couret,
Triplett, Price, and Mareci (2016). This approach is similar to the binary framework used by
Watts and Strogatz (1998), but with an additional degree of freedom from the edge weight-
ing (Equazione 1).
In our previous work, we showed that this weighted framework yields
topologically relevant features without the need for thresholds, therefore no threshold was
applied to generate the brain connectomes in this work (Colon-Perez et al., 2016). Weak
edges in a network are thought to provide a cohesive strength to networks (Granovetter, 1973).
The ability to obtain stable connectome results across thresholds, shown in Colon-Perez et al.
(2016), allows us to maintain these weak edges in our analysis. These networks were studied
with the following indices: (UN) graph density (Boccaletti, Latora, Moreno, Chavez, & Hwang,
2006), which is a binary metric that quantifies the fraction of edges in a graph (only non-
weighted index in this study); (B) node strength (Newman, 2001), which is a weighted topo-
logical index of the relative connectivity strength of the nodes with the rest of the network; (C)
clustering coefficient (Zhang & Horvath, 2005), which is a weighted metric that quantifies the
strength of connectivity between the neighbors of a node; (D) path length (Colon-Perez et al.,
2016), which is a weighted metric that quantifies the strength of the shortest path between two
nodes; E (D) small-worldness (Humphries & Gurney, 2008), which is a weighted metric that
estimates the likelihood that networks display similar path lengths and higher clustering to a
network connected by randomly assigned edges, as described by Erdös and Rényi (1959). For
a complete description of the weighted network analysis, refer to Colon-Perez et al. (2016).
The results of these network indices in this manuscript will be referred to as global for each
participant when the results are averaged into a single value for the entire brain network
(yields one value per participant). The local results for each participant refer to the average
values per node (yields 82 values per participant). Previous studies described global dif-
ferences in node strength and path length in patients with PD (Galantucci et al., 2016); In
this work, we use these indices to identify local changes in network connectivity in addition
to corroborate previous global changes described by Galantucci et al. (2016). The cluster-
ing coefficient is reduced globally in patients with PD (Luo et al., 2015); in this study, we
Network Neuroscience
110
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
Cognitive composites:
Averages of standardized scores from
multiple neurocognitive measures
theoretically measuring the same
cognitive domain.
identify the local changes in clustering coefficient and identify those nodes responsible for the
reduction. Several PD studies have confirmed the small-world topology in controls and pa-
tients with PD; in this study, we assess whether PD-Well or PD-MI displays a difference from
controls.
Statistics and Correlations
Cognitive composites and network variables were tested for statistical significance with a non-
parametric Mann-Whitney test using R (version 3.1.3) (Team, 2015). All node-specific network
results were controlled by the graph density to control the influence of global connectivity
features in local indices (cioè., node-specific; van Wijk, Stam, & Daffertshofer, 2010). To cor-
rect for multiple comparisons, we used the “fdrtool” package (Strimmer, 2008) and the false
nondiscovery rate, which estimates the proportion of nondiscovery rejections or type II errors
(Genovese & Wasserman, 2002). This tool works by first finding a suitable cutoff point using
an approximate null model, which is fitted; subsequently, a cutoff point is sought with a false
nondiscovery rate as small as possible. Scale parameters of the null model and proportion of
null values are then estimated from the data. The corresponding p values are computed, and a
modified Grenander algorithm (The Grenander, 1956, 1956, R implementation in fdrtool can
be found in https://cran.r-project.org/web/packages/fdrtool/fdrtool.pdf) is used to find the over-
all density and distribution function. Finalmente, adjusted p values are determined and reported.
Correlations were calculated between network indices (per esempio., node strength, path length, clus-
tering, small-worldness) and neuropsychological composites (per esempio., working memory, memory
speed, and verbal memory). Since our goal is to identify the network indices that correlate with
the different neurocognitive domains under study, no correlations were assessed between the
various network indices. The correlations were estimated using a Spearman’s partial correla-
tion method controlling for education level using the “ppcor” package in R. With significance
in our correlative analysis set at α = 0.05 and given our sample size of N = 80, we decided
to reduce potential false positives by considering only correlations larger than 0.50 (Cohen,
1992).
RESULTS
Demographics
Control and PD groups were not statistically different in their average age (Tavolo 1). The PD-
Well group did not show a significant difference from controls in education, working memory,
or memory composites (Tavolo 1).
The PD-Well participants’ working-memory scores did not show significant deficits, whereas
PD-MI was 0.88 standard deviations lower in working-memory score. The PD-MI group was
2.5 years less educated than controls (Tavolo 1) and PD-Well (p = 0.03, p value obtained be-
tween PD-MI and PW-Well). Relative to controls, PD-Well and PD-MI displayed a reduction
in their processing-speed scores (Tavolo 1).
The combined PD group (n = 40) had lower memory composite scores than controls
(P < 0.01). PD-Well had lower memory composite scores than controls, but this was not statis-
tically significant (p = 0.082; Table 1). PD-MI had lower memory composite scores than both
cohorts: controls (p < 0.001) and PD-Well (p < 0.001; Table 1). Group differences also
remained after controlling for processing speed and working memory (p < 0.001).
Network Neuroscience
111
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
n
e
n
_
a
_
0
0
0
2
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectomes in Parkinson’s disease
σσσ
Control (nnn = 40)
Mean
68.18
NA
16.75
2.75
1.15
0.16
0.00
Table 1. Demographics and cognitive indices
PD-Well (nnn = 31)
Mean
67.3
σσσ
ppp
PD-MI (nnn = 9)
Mean
69.4
σσσ
ppp
7.87
6.22
16.8
18.2
5.02
5.60
2.91
4.64
NA
2.35
3.36
0.55
0.47
1.00
Age (yrs)
Duration (yrs)
Education (yrs)
UPDRS III
Working mem
Proc speed
Mem comp
0.60
NA
8.23E-3*
4.52E-6*
6.70E-5*
3.90E-3*
3.54E-5*
Statistical significance was tested between PD subgroups (i.e., PD-Well and PD-MI) and controls.
Yrs = years, Duration = disease duration, UPDRS = Unified Parkinson’s Disease Rating Scale,
Working mem = working-memory score, Proc speed = processing-speed composite score, Mem
comp = memory composite score, and σ = standard deviation. Statistics performed with Mann-
Whitney statistics between PDs and controls. * correspond to statistically significant differences.
0.38
NA
0.86
1.34E-10*
0.148
3.23E-5*
0.082
6.77
3.07
2.74
7.25
0.57
0.76
0.20
0.89
−0.42
−0.32
0.27
−0.68
−1.76
0.77
0.56
0.70
14.3
15.7
11.6
Global Network Measures
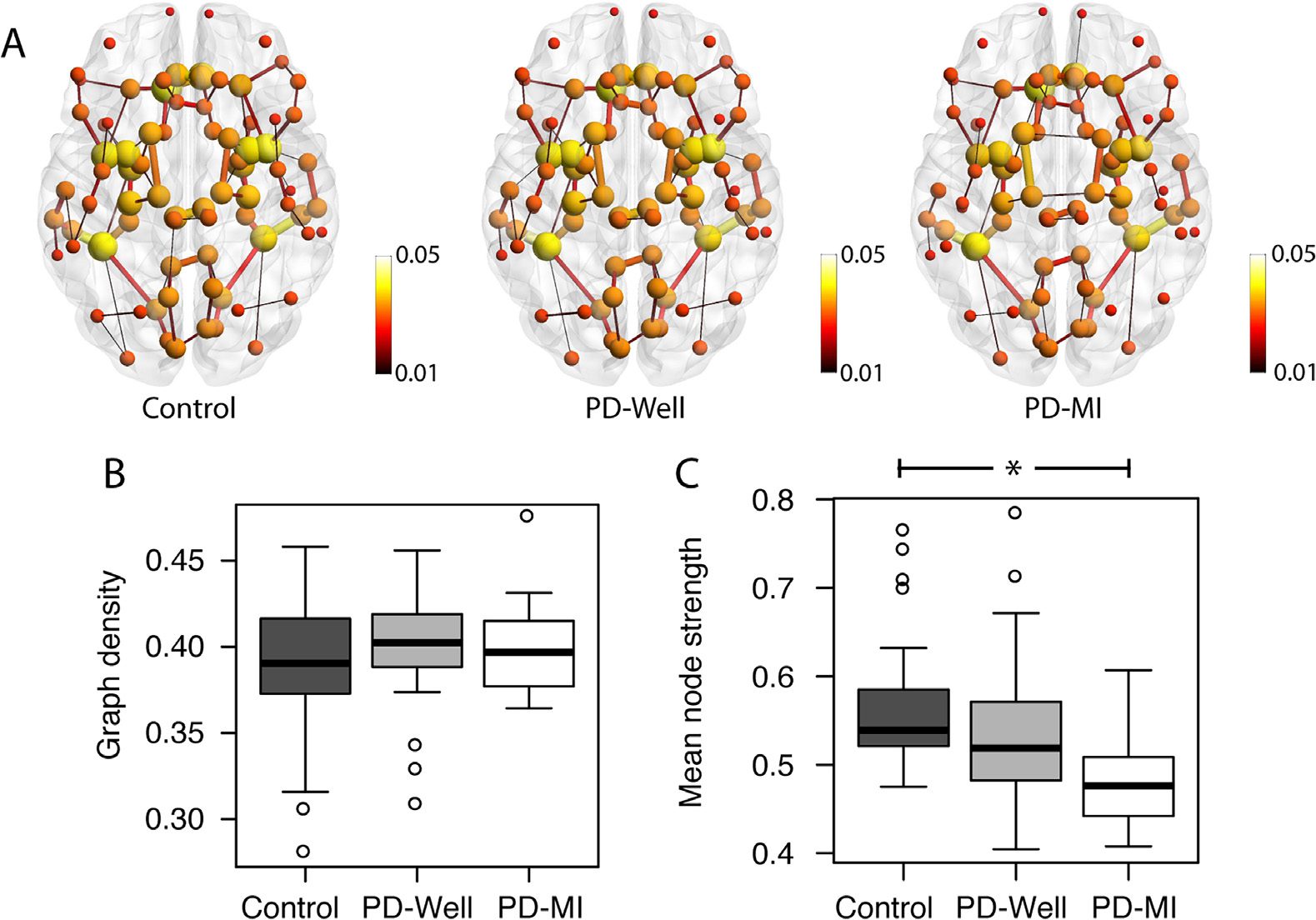
The connectomes in Figure 1A show the top 1% of the strongest con-
Results and differences
nections and the connectivity alterations in the PD-MI connectomes relative to controls. The
node size and color represent the node strength, and the edge thickness represents the edge
weight value. In particular, connections between nodes in the temporal lobe are smaller in
size (representing a reduction in edge weight). The graph density (Figure 1B) in all groups
was approximately 40% of all possible edges, with no significant differences between con-
trols and PD-Well (p = 0.249) or between controls and PD-MI (p = 0.624). To reduce the
bias of graph density on network indices, these will be controlled by the graph density for the
rest of the manuscript (van Wijk et al., 2010). A 4.99% reduction in mean node strength is
observed over the entire network between controls and PD-Well (p = 0.041), whereas mean
node strength in the PD-MI group was reduced relative to controls by 13.23% (p = 0.004;
Figure 1C). The mean path lengths for the entire network were reduced by 1.97% between
controls and PD-Well (p = 0.058), whereas the mean path lengths in the PD-MI group dis-
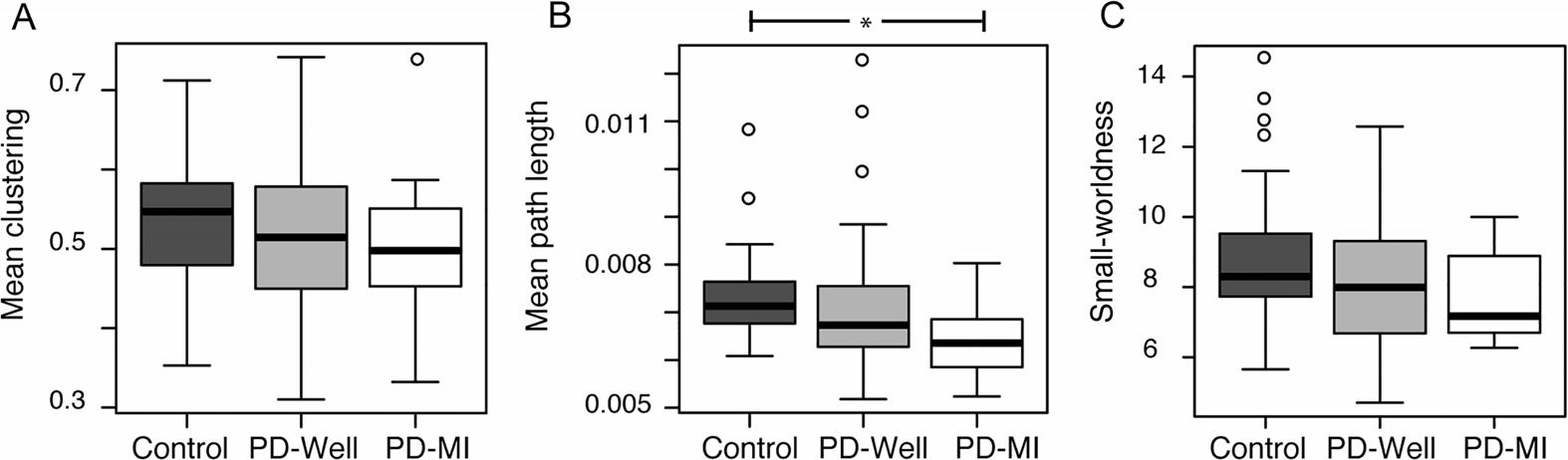
played a significant decrease of 11.7% (p = 0.014, Figure 2B). The mean clustering coefficient
for the entire network did not differ significantly between controls and PD-Well (p = 0.162) or
PD-MI (p = 0.320), with mean values approximately 0.55 (Figure 2A). The small-worldness in-
dex did not differ significantly between controls and PD-Well (p = 0.188) or PD-MI (p = 0.131),
with mean values approximately 8.00 (Figure 2C). Also, we determined node strength, clus-
tering coefficient, and path length with a more traditional weighting scheme of FA as the edge
weight. We did not find any differences between controls and PD-Well or controls and PD-MI
using FA as the edge weight (all p > 0.1; the database is available as an online supplement
and Supplementary Figure 3; see Colon-Perez et al., 2017).
For the controls, global network indices did not show any correlations with the
Correlations
primary neuropsychological composites of interest (Tavolo 2). The memory composite score did
not correlate with any network index for any of the groups. The working-memory composite
score showed negative correlations with node strength for PD-Well participants and control.
Path length also showed a negative correlation with working memory between PD-Well and
controls. The PD-MI group showed negative correlations between all network indices and the
processing-speed composite (except small-worldness); working memory negatively correlated
with path length. For PD-MI, there were positive correlations between the Unified Parkinson’s
Global network indices:
Average value of a network index
across all nodes for a single subject.
Network Neuroscience
112
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
Figura 1. The structural connectivity of controls, PD-Well, and PD-MI participants. (UN) Average
connectome per group; the node size represents the strength of the connections with the rest of
the network, and the edge width represents the relative strength of connections between pairs of
nodes. (B) Box-plot distributions of graph density values (number of connections in connectome).
No significant differences in the number of edges were observed between groups.
(C) Box-plot
distributions of average global node strength. The mean node strength group comparison between
Control and PD-MI was significantly reduced (p = 0.002). The connectome images in this figure
were prepared using BrainNet (Xia, Wang, & Lui, 2013).
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. Box plots of global network indices for all groups.
(UN) Mean clustering = mean
global clustering coefficient per brain averaged across subjects within each group. (B) Mean path
length = mean global path length per brain averaged across subjects within each group.
(C) Small-worldness = small-worldness index per brain averaged across subjects within each group.
◦
* = statistically significant difference (P < 0.05),
= outliers. Statistics performed with a nonpara-
metric Mann-Whitney test.
Network Neuroscience
113
Connectomes in Parkinson’s disease
Local network indices:
Value of a network index for every
node and a single subject.
Table 2. Correlations (Fisher’s z-score) between global network indices and neuropsychological
composites
Network index
Node strength
Clustering
Path length
Small-worldness
Network index
Node strength
Clustering
Path length
Small-worldness
Network index
Node strength
Clustering
Path length
Small-worldness
UPDRS
NA
NA
NA
NA
UPDRS
0.008
−0.287
0.070
−0.172
UPDRS
0.824#
−0.071
0.955#
−0.084
Controls (nnn = 40)
Working mem
Proc speed
Mem comp
0.225
0.079
0.100
−0.144
0.021
0.149
0.051
−0.076
PD-Well (nnn = 31)
Working mem
−0.574∗
0.018
−0.594∗
0.115
Proc speed
0.169
0.251
0.193
0.230
PD-MI (nnn = 9)
Working mem
−0.314
−0.328
−0.826∗
−0.109
Proc speed
−0.594#
−0.685∗#
−0.724∗#
−0.450
0.230
0.195
0.082
−0.148
Mem comp
−0.179
0.076
−0.229
−0.049
Mem comp
0.116
0.356
0.312
0.079
UPDRS = Unified Parkinson’s Disease Rating Scale, Working mem = working-memory score,
Proc speed = processing-speed composite score, Mem comp = memory composite score, and
clustering = clustering coefficient score. Partial correlation controlled for age using Spearman’s
method transformed to Fisher’s z-scores using the “psych” package in R. Black bins correspond
to |z| > 0.55 and correlated results. * corresponds to statistically significant difference to con-
trols; # corresponds to statistically significant difference to PD-Well, calculated from r values with
http://vassarstats.net/rdiff.html.
Disease Rating Scale (UPDRS) Part III (motor test) and node strength, as well as path length
(Tavolo 2; scatter plots are shown in Supplementary Figure 2; see Colon-Perez et al., 2017).
Local Network Measures
Group differences Node strength and path length box plots for each group are shown in
Figures 3 E 4. There were local network (cioè., node-specific) differences between controls
and PD-MI (no difference between controls and PD-Well; for specific node results, refer to ta-
bles in the Supplementary Information, Colon-Perez et al., 2017). After correcting for multiple
comparisons and controlling for graph density, statistically significant changes were observed
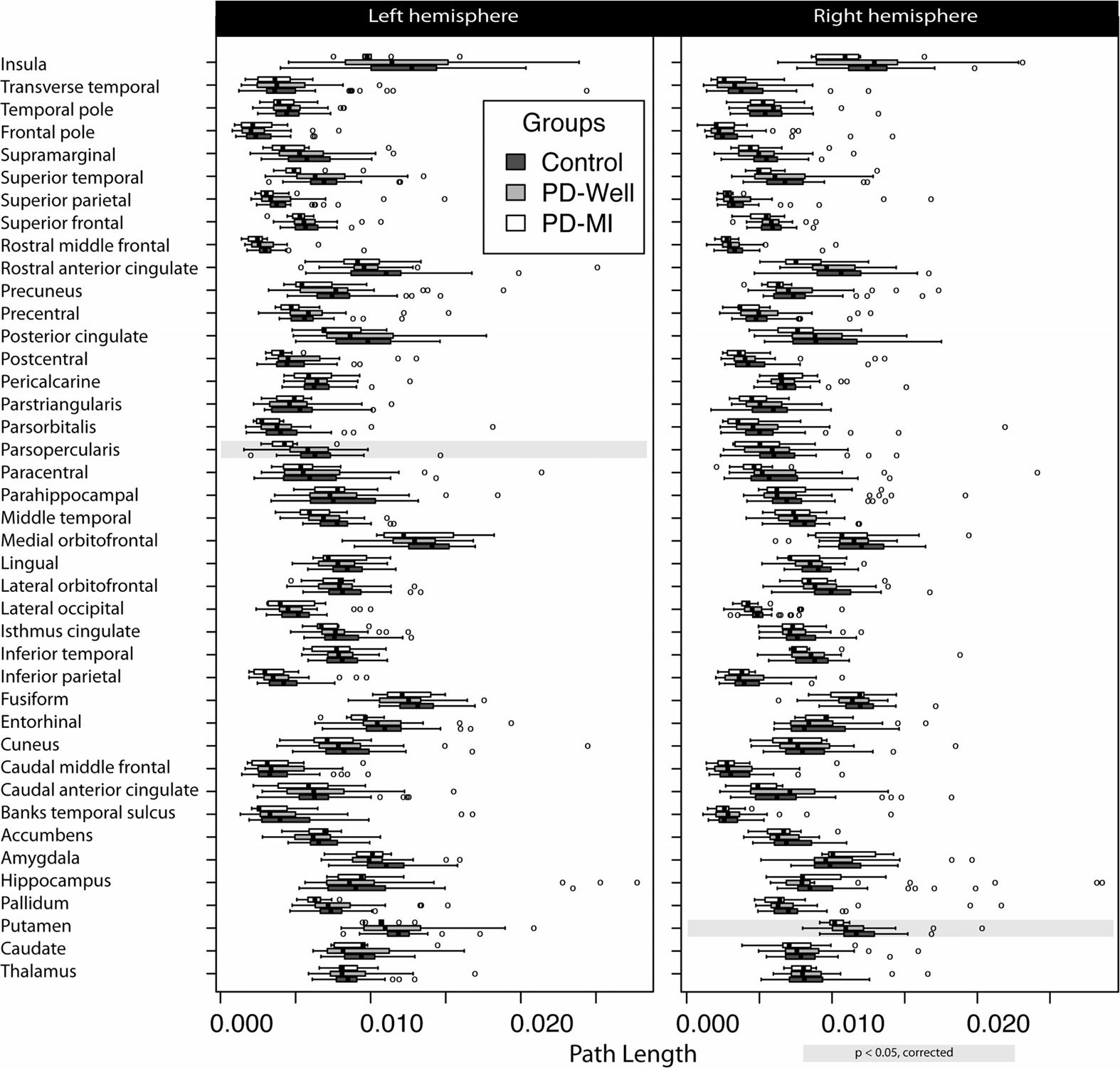
for 27 distinct nodes in node strength (Figura 3), and two nodes for path length (Figura 4). There
were no observed differences in clustering for any node. The left (Lf) pars opercularis and right
(Rt) putamen showed statistical differences in node strength and path length. The location of
the nodes with statistically significant differences can be seen in Supplementary Figure 1.
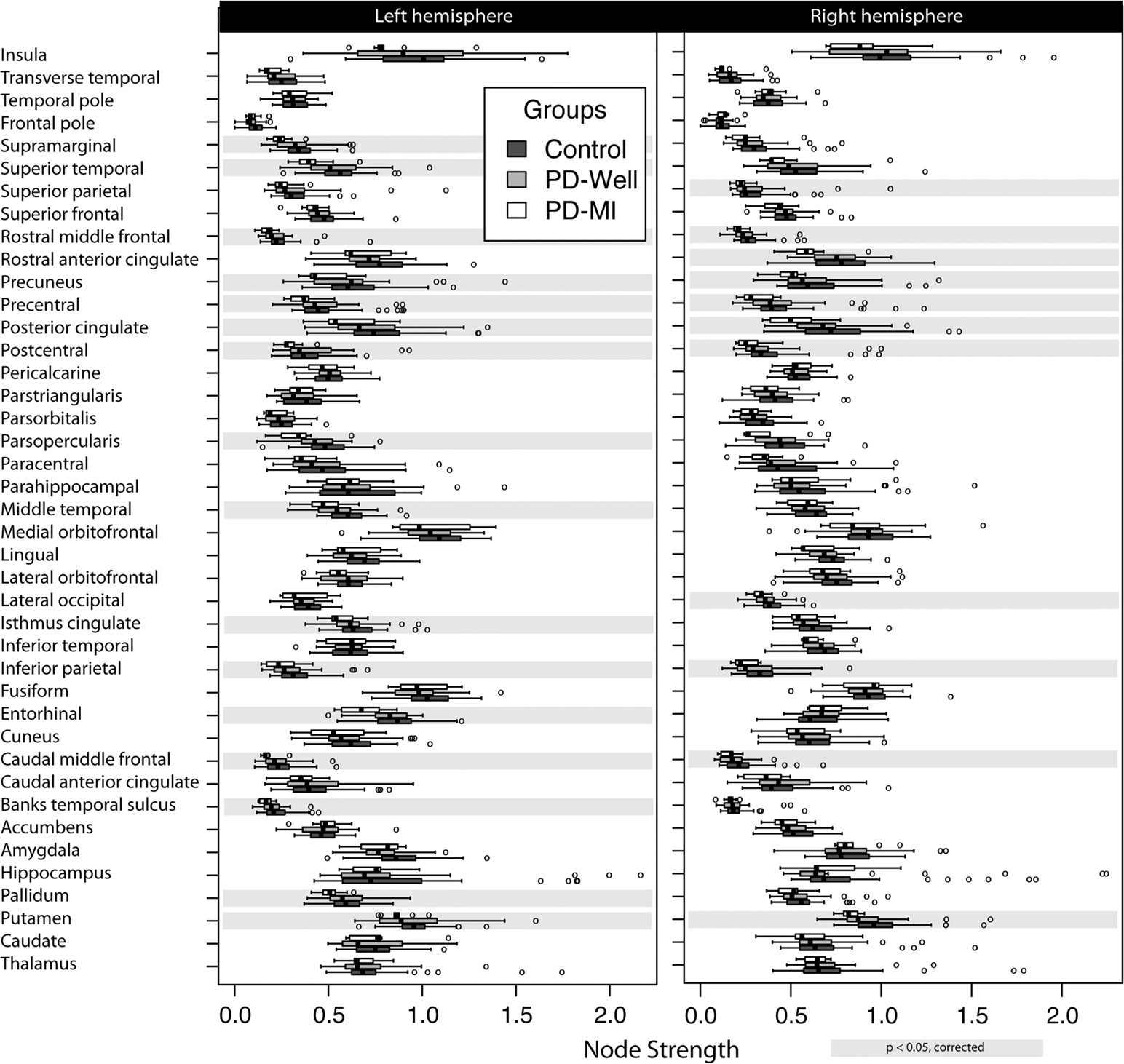
Bilateral differences in node strength between PD-MI and controls were found for the fol-
lowing: putamen (pLf = 0.043 and pRt = 0.002), caudal middle frontal (pLf = 0.018 E
pRt = 0.040), inferior parietal (pLf = 0.031 and pRt = 0.019), postcentral (pLf = 0.017 E
pRt = 0.045), posterior cingulate (pLf = 0.043 and pRt = 0.006), precentral (pLf = 0.017 E
pRt = 0.019), and precuneus (pLf = 0.019 and pRt = 0.043). Additional nodes with statisti-
cally significant differences were the following: Lf pallidum (p = 0.021), Lf entorhinal cortex
(p = 0.004), Lf isthmus cingulate (p = 0.027), Lf middle temporal (p = 0.016), Lf pars oper-
cularis (p = 0.003), Lf banks of the superior temporal sulcus (p = 0.019), Lf supramarginal
Network Neuroscience
114
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 3. Node strength results in a connectome of 82 cortical and subcortical nodes. Controls
(n = 40), Parkinson’s participants without memory impairment (n = 31), and Parkinson’s partic-
ipants with memory impairment (n = 9). The left column represents the left hemisphere nodes,
and the right column corresponds to the right hemisphere. Highlights in gray represent a significant
difference (corrected for multiple comparisons) between controls and PD-MI. No significant differ-
ences were observed between PD-Well and controls. Banks temporal sulcus is an abbreviation for
the banks of the superior temporal sulcus region, which is the name in the FreeSurfer nomenclature;
the full name is used in the main text.
(p = 0.002), Lf rostral middle frontal (p = 0.003), Lf superior temporal (p = 0.006), Rt lat-
eral occipital (p = 0.029), Rt rostral anterior cingulate (p = 0.019), Rt rostral middle frontal
(p = 0.007), and Rt superior parietal (p = 0.040). The path length was different for Lf pars
opercularis (p = 0.002) and Rt putamen (p = 0.001). Local indices of node strength, clustering
coefficient, and path length did not show any differences between controls and PD-Well or
controls and PD-MI using FA as the edge weight after correcting for multiple comparisons.
Correlations Given the large number of correlations (three network indices, three cognitive
indices, and UPDRS scores for PDs, 82 nodes, and three subject groups), the discussion is
restricted to the correlation analysis of those nodes that showed statistically significant differ-
ences between controls and PD-MI. Spearman correlations were performed for 27 nodes (cioè.,
those with statistically significant differences between PD-MI and controls; Figura 3), three
Network Neuroscience
115
Connectomes in Parkinson’s disease
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 4. Path length results per node in a connectome of 82 cortical and subcortical nodes.
Controls (n = 40), Parkinson’s participants without memory impairment (n = 31), and Parkinson’s
participants with memory impairment (n = 9). The left column represents the left hemisphere nodes,
and the right column corresponds to the right hemisphere. Highlights in gray represent a significant
difference (corrected for multiple comparisons) between controls and PD-MI. No significant differ-
ences were observed between PD-Well and controls. Banks temporal sulcus is an abbreviation for
the banks of the superior temporal sulcus region, which is the name in the FreeSurfer nomenclature;
the full name is used in the main text.
network indices, and three neuropsychological indices (PD participants also include UPDRS
scores). This analysis yields a maximum of 243 correlations in controls and 324 for individ-
uals with PD (the correlations for all nodes can be found in the online supplemental data;
Colon-Perez et al., 2017).
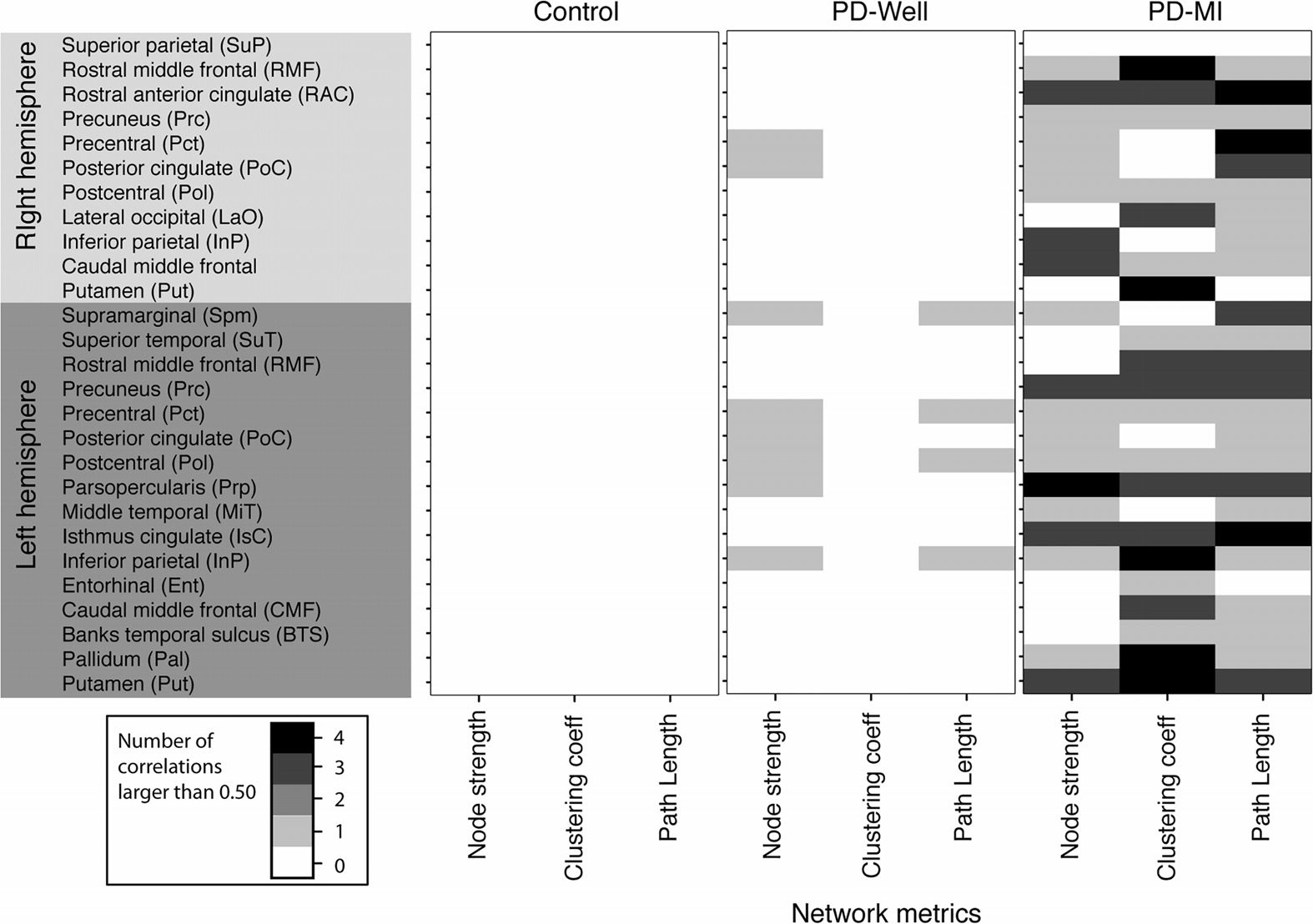
Controls: The control group did not exhibit significant correlations between any network
metric (cioè., node strength, clustering, or path length) and any neuropsychological composite
(working memory, memory composite, or processing-speed composite). See Figure 5, left
column.
PD-Well: There were 12 correlations with coefficient values larger than 0.5 between net-
work indices and all neuropsychological composites (composites described in cognitive mea-
sures). See Figure 5, middle column. For PD-Well, the node strength correlated with the
Network Neuroscience
116
Connectomes in Parkinson’s disease
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 5. The number of significant correlations (absolute value |R| > 0.5) between network
indices and each neuropsychological composite and UPDRS Part III scores (cioè., motor symptoms of
PD). The intensity of each pixel is the number of correlations larger than 0.5 for each network metric
and the specified node with the neuropsychological composites (cioè., working memory, memory
composite, speed composite, and UPDRS Part III). There is a maximum of four possible significant
correlations, computed between a single network metric and the four neurocognitive measures.
This is then reported for each node with significant differences in node strength between PD-MI
and controls (Guarda la figura 3). Banks temporal sulcus is an abbreviation for the banks of the superior
temporal sulcus region, which is the name in the FreeSurfer nomenclature; the full name is used in
the main text.
composites for the following nodes: Lf inferior parietal, Lf pars opercularis, Lf postcentral, Lf
posterior cingulate, Lf precentral, Lf supramarginal, Rt posterior cingulate, and Rt precentral.
The clustering coefficient did not correlate with any composite. The path length correlated for
the Lf inferior parietal, Lf precentral, Lf postcentral, and Lf supramarginal.
PD-MI: There were 63 correlations with values larger than 0.5 between network indices
and all neuropsychological composites. The majority of nodes correlated with at least one
neuropsychological composite for the PD-MI group (Figura 5); Perciò, in this section, we
will list the nodes that did not show correlations between network indices and composites.
The node strength of PD-MI did not correlate with Lf banks of the superior temporal sulcus, Lf
caudal middle frontal, Lf entorhinal, Lf rostral middle frontal, Lf superior temporal, Rt putamen,
Rt lateral occipital, and Rt superior parietal. The clustering coefficient did not correlate with
composites for Lf middle temporal, Lf posterior cingulate, Lf supramarginal, Rt inferior parietal,
Rt precentral, Rt superior parietal, and Rt posterior cingulate. The path length did not correlate
for the Lf entorhinal cortex, Rt putamen, and Rt superior parietal.
Network Neuroscience
117
Connectomes in Parkinson’s disease
DISCUSSION
In this study, we identified differences in the connectome organization for people with idio-
pathic PD with mild amnestic disturbance (PD-MI), and we explored correlations between
connectomes and the most vulnerable cognitive domains within PD relative to non-PD peers.
Global differences were identified in the PD-MI versus control groups, in mean node strength
and path length but not in clustering coefficients, small-worldness, or graph density. No dif-
ferences were observed for any of the indices (global or local) for the PD-Well group versus
controls. We found that 27 out of 82 nodes showed local differences of connectivity in node
strength in the PD-MI group relative to controls (Figura 3). Local node network differences in
node strength and path length were found in PD-MI brains and were specific to the Lf pars
opercularis, and Rt putamen (Supplementary Figure 1). Bilateral differences between controls
and PD-MI were found in node strength for the putamen, caudal middle frontal, inferior pari-
etal, postcentral, posterior cingulate, rostral middle frontal, precentral, and precuneus. IL
right putamen was the only subcortical region that displayed a significant connectome alter-
ation in the form of a reduction in node strength and path length. Node strength and cognitive
composites further showed a potential large-scale network connectivity reduction in PD-MI
relative to normal cognitive areas of vulnerability. This study shows key connectome indices
for consideration in PD-Well and PD-MI phenotypes.
Structural Differences
Robust connectome differences between PD-MI and controls were observed at the global level
with mean node strength and mean path length. In this study, groups’ structural connectomes
showed many similar levels of connectivity, as shown in the weighted top edges in Figure 1A
and their graph densities in Figure 1B. Relative to non-PD peers, mean node strength was al-
tered by 5% E 13% for PD-Well and PD-MI, rispettivamente, and mean path length was changed
2% E 11%, rispettivamente. These findings in node strength and path length suggest a reduction
in the integrity of white matter connectivity in PD. The path length alteration in PD is directly
related to the node strength reduction (cioè., r = 0.92). The edges (cioè., edge weights) connect-
ing nodes that in turn yield the shortest path between nodes possibly decreases because of
the neurodegeneration of white matter in PD (Tessitore, Giordano, Russo, & Tedeschi, 2016).
The absence of group differences in clustering coefficient and small-worldness, which is con-
firmed by a previous report (Galantucci et al., 2016), may suggest a method to compensate
for PD changes where the brain network adapts to preserve its small-worldness and clustering
caratteristiche. Therefore, the changes in node strength and path length could be markers of PD
progression and cognitive decline. Although cross-sectional and preliminary, these findings
suggest a local and global network reduction in connectivity with an amnestic disturbance
(cioè., PD-MI).
To further explore the relevance of particular node alterations, we performed local
topological analyses that revealed evidence of local network disruption in PD-MI. Node
strength and path length abnormalities were seen with the left pars opercularis and right
putamen. The frontal region is involved in networks of language, Attenzione, and work-
ing memory (Lezak, 2012; Petrides, 2000; Stuss et al., 2005; Zola-Morgan & Squire, 1986;
Zola-Morgan, Squire, & Amaral, 1986), while the subcortical nuclei of the putamen is in-
volved in PD motor symptomatology (Braak, Ghebremedhin, Rub, Bratzke, & Del Tredici,
2004; Lisanby et al., 1993; Nemmi, Sabatini, Rascol, & Peran, 2015; Price et al., 2016). Al-
though preliminary, these connectivity reductions primarily in left cortical areas validate the
profile of our PD-MI, particularly given that the PD-MI group was classified with verbal memory
Network Neuroscience
118
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
measures, which involve left hemisphere regions more than the right (Golby et al., 2001). Questo
profile is also consistent with other reports showing anterior and lateral temporal thinning and
volume reduction in PD-MI (Crowley et al., 2017; Pagonabarraga et al., 2013; Tanner et al.,
2015).
this point, we would like to illustrate a larger issue in the connectomics litera-
A
ture:
In this work, we found a discrepancy be-
the lack of a ground truth edge weight.
tween our weighted framework (Colon-Perez et al., 2016) and an FA weighting scheme.
Several works have been published that bring attention to the inadequacies of tradition-
ally used edge weighting schemes in network neuroscience. Cheng et al. (2012) first de-
scribed how increasing seed density improves the stability of network metrics. Tuttavia,
there is a caveat that higher seed densities lead to a larger number of spurious streamlines
and thus affect the connectome.
In our edge weight scheme, we employ a large num-
ber of seeds per voxel to increase its stability and use the characteristic function (Equa-
zione 2) to mitigate the effects of spurious streamlines (Colon-Perez, Spindler, et al., 2015).
Buchanan, Pernet, Gorgolewski, Storkey, and Bastin (2014) suggest
that some measure of
streamline density (as our edge weight) is superior to FA since it produces better test-retest
performance. These biases and others are attempted to be controlled or reduced by our
weighting scheme (Colon-Perez, Spindler, et al., 2015). Our work and that of many others
are under way to develop new and better ways to weigh networks in connectome studies that
improve the stability of network metrics derived from tractography (Colon-Perez et al., 2016;
Colon-Perez, Spindler, et al., 2015; Girard, Whittingstall, Deriche, & Descoteaux, 2014). Noi
are not claiming that our weighting scheme is a more accurate representation of the underlying
anatomical connectivity than others; but with our previous articles and this one, we hope to
continue the discussion to find new and novel ways to weigh connectomes.
Functional MRI studies have shown that disruptions in the motor networks of patients with
PD correlate with disease severity (Wu et al., 2009). These functional changes are likely ac-
companied by structural disruption, potentially like the ones described here. Microstructural
changes have been reported in PD using diffusion tensor imaging. Lower FA has been observed
in the substantia nigra and the striatum in patients with PD (Tessitore et al., 2016). In contrasto,
FA increases have been noted in the corpus callosum and the superior longitudinal fascicu-
lus (Gattellaro et al., 2009). This white matter change leads us to hypothesize that interhemi-
spheric connectivity alterations are mediated via the corpus callosum and within-hemisphere
alterations through the superior longitudinal fasciculus (Northam et al., 2012). Reductions in
FA have also been reported in participants with PD in the putamen, substantia nigra, striatum,
frontal lobes, and motor areas (Zhan et al., 2012). These brain connectivity changes are not
only restricted to MRI but also have been observed with SPECT (Booij et al., 1997) and PET
(Brooks, 1995). Altogether, the research findings, including our study, suggest many changes
in PD may coalesce into a reorganization of the structural brain connectome. We recognize
that these connectome differences may not be unique to memory-impaired PD because simi-
lar differences have been reported in patients with epilepsy after anterior temporal lobectomy
(Ji et al., 2015), and in individuals with PD after deep brain stimulation (van Hartevelt et al.,
2014).
Cognitive-Network Correlations
Cognitive-network analyses help to validate group-level structural differences. We examined
key cognitive domains compromised in PD: processing speed, working memory, and episodic
Network Neuroscience
119
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
memory. Our observations suggest that PD-MI had more correlations between neuropsycho-
logical composites and connectivity indices than PD-Well and non-PD peers. For PD-Well,
correlations were found between working memory and node strength, as well as for working
memory and path length. The PD-Well group’s working memory showed a negative corre-
lation with node strength and path length (cioè., higher node strength and path lengths were
associated with lower working-memory scores). The topological changes suggest a possible
maladaptation in the brain networks of those with PD, as suggested in other neurological dis-
orders (Doucet et al., 2015; Drakesmith et al., 2015; Wu et al., 2009). Così, these topological
indices might be used as markers for cognitive performance in PD.
There were no significant local network correlations between the network indices and neu-
ropsychology composites in the control group (Figura 5). This negative finding is expected
given this group’s relative cohesiveness, intact performance, and restricted range of scores. In
contrasto, the PD-Well group showed an increased number of strong correlations, while the
PD-MI showed an even higher number of strong correlations. The increase in the number of
correlations might reflect a progressive alteration of brain connectivity mediating the cognitive
decline. Although we observed increasing numbers of correlations between network indices
and neuropsychological composites, we did not find verbal memory correlations to any of
the network indices. The lack of correlations between verbal memory and network indices in
PD-MI was possibly due to the restricted range of their scores (all impaired).
Study Strengths and Limitations
Overall, the study strengths include the presentation of a comprehensive correlational analysis
between network topology and cognition using various neuropsychological indices, prospec-
tive PD and control matching on demographic variables (cioè., age and education), and robust
identification of memory impairment in PD. The PD subgroup cognitive profiles showed ex-
pected reductions in processing speed for PD relative to non-PD controls, with PD-MI show-
ing reductions in memory and processing speed. In this work, we used a weighted network
method that yields more stable topological metric results than binary network methods and is
robust despite graph density differences; hence, it does not require thresholding to generate
the connectomes (Colon-Perez et al., 2016). Topological features of binary network connec-
tomes may be affected by their graph density and ultimately may hinder comparisons between
groups (Langer, Pedroni, & Jancke, 2013; Sporns, 2011B). We could circumvent the problem
of thresholding by employing a weighted framework that reduces the effects of thresholding in
network indices results (Colon-Perez et al., 2016).
The authors recognize some of the study limitations. MR tractography is susceptible to
false positives and false negatives; hence, care is required when analyzing and interpreting
tractography-derived results (Alhourani & Richardson, 2015). In this study, we used a different
edge weight to calculate the strength of connectivity between nodes. This approach reduces
the tractography bias effects of seed density and length (Colon-Perez, Spindler, et al., 2015).
Also, the edge weight is defined only along the path of the streamlines, which reduces the
possibility of obtaining streamlines from extraneous areas. Also, the use of a weighted net-
work defines connectomes as a set of weak and strong connections, which has been shown
to reduce the effects of false-positive connections in the quantification of connectome indices
using binary networks (Colon-Perez et al., 2016). Inoltre, false-positive connections yield
connectome edges with a small number of streamlines, which in turn corresponds to a low
edge weight value; these spurious links will only produce changes in node strength of less than
1% (Colon-Perez et al., 2016). Another limitation is the relatively low spatial resolution (2 mm
Network Neuroscience
120
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
isotropic for diffusion images) in tractography, which is a limiting factor when estimating small
tracts and may lead to false negatives (Colon-Perez, King, et al., 2015; Ford et al., 2013). In
the current work, we used high angular-resolution diffusion imaging to increase the angular
resolution and enable better estimation of the fiber orientation on each voxel (Jian et al., 2007;
Tuch et al., 2002). It also has been reported that the connectome changes with age (Wu et al.,
2007). Tuttavia, the mean and range of age in each group is similar to ensure there were no
age differences between groups. Così, the current results may only apply to similarly aged
populations. We also did not consider genetics, which has been shown to modulate connec-
tome topology in patients with PD through the rs405509 risk allele (TT) (Shu et al., 2015). An
additional limitation is the relatively small sample size of this study, particularly within the
PD-MI group (n = 9). Future studies are encouraged using larger sample sizes and additional
explorations of network differences in memory versus other cognitive (per esempio., executive working,
Attenzione) deficits.
CONCLUDING REMARKS
Our connectome analyses suggest a loss and reorganization of brain white matter structure
in PD, particularly PD with memory impairment. We identified a reduction of connectome
topological and neuropsychological indices. Our results show a relationship between cog-
nitive deficits and connectome structure alteration in PD with mild cognitive impairments.
It remains to be determined whether the observed network changes are causal of the neu-
rocognitive deficits or vice versa. Also, further studies are needed to assess the mechanisms
relating to the observed topological changes in brain structure to the neurocognitive deficits.
The data suggest a broader change, at the level of the connectome, associated with the clinical
manifestations of cognitive phenotypes, particularly the memory phenotype of PD.
ACKNOWLEDGMENTS
We are grateful to the participants involved in the current investigation. We are also grateful to
Tony Mancuso, MD, for his help in securing the MR Siemens Verio, and the UF Radiology team
for their guidance. We would like to acknowledge Irene Malaty, MD, Ramon Rodriguez, MD,
Janet Romrell, ARNP, Pam Zeilman, ARNP, and all the faculty in the UF Center for Movement
Disorders and Neurorestoration, Gainesville, Florida, for referring individuals to the investiga-
zione. We also thank Jessica Williams, Cassie Catania, Jade Ward, Katie Rodriguez, and Breana
Wallace, for their assistance with participant recruitment.
AUTHOR CONTRIBUTIONS
Luis M. Colon-Perez: Conceptualization; Formal analysis; Investigation; Methodology; Project
administration; Software; Writing – original draft; Writing – review & editing. Jared J. Tanner:
Conceptualization; Data curation; Formal analysis; Methodology; Writing – original draft;
Writing – review & editing. Michelle Couret: Formal analysis; Investigation; Writing – review
& editing. Shelby Goicochea: Formal analysis; Investigation. Thomas H. Mareci: Project
administration; Supervision; Conceptualization; Methodology; Writing – review & editing.
Catherine C. Price: Conceptualization; Formal analysis; Funding acquisition; Investigation;
Methodology; Project administration; Supervision; Writing – original draft; Writing – review
& editing.
Network Neuroscience
121
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
FUNDING INFORMATION
Funding for this research was supported by the National Institutes of Health (grant nos. K23
NS060660, RO1 NR014181, and R01 NS082386), by the National Institutes of Health Clini-
cal and Translational Science Award program (grant nos. UL1TR001427, KL2TR001429, E
TL1TR001428), and by the UF Center for Movement Disorders and Neurorestoration and the
Brain and Spinal Cord Injury Research Trust Fund of the State of Florida. The content is solely
the responsibility of the authors and does not necessarily represent the official views of the
National Institutes of Health.
REFERENCES
Alhourani, A., & Richardson, R. M. (2015). Inherent limitations of
tractography for accurate connectivity maps. Neurosurgery, 76,
N11–N12.
Basser, P. J., & Jones, D. K. (2002). Diffusion-tensor MRI: Theory,
experimental design and data analysis – a technical review. NMR
in Biomedicine, 15, 456–467.
Basser, P. J., Pajevic, S., Pierpaoli, C., Duda, J., & Aldroubi, UN.
(2000). In vivo fiber tractography using DT-MRI data. Magnetic
Resonance in Medicine, 44, 625–632.
Bellucci, A., Mercuri, N. B., Venneri, A., Faustini, G., Longhena,
F., Pizzi, M., . . . Spano, P. (2016). Review: Parkinson’s disease:
From synaptic loss to connectome dysfunction. Neuropathology
and Applied Neurobiology, 42, 77–94.
Boccaletti, S., Latora, V., Moreno, Y., Chavez, M., & Hwang, D. U.
(2006). Complex networks: Structure and dynamics. Physics
Reports, 424, 175–308.
Booij, J., Tissingh, G., Boer, G. J., Speelman, J. D., Stoof, J. C.,
Janssen, UN. G., . . . van Royen, E. UN. (1997). [123IO]FP-CIT SPECT
shows a pronounced decline of striatal dopamine transporter
Journal of
labelling in early and advanced Parkinson’s disease.
Neurology, Neurosurgery, and Psychiatry, 62, 133–140.
Braak, H., Ghebremedhin, E., Rub, U., Bratzke, H., & Del Tredici,
(2004). Stages in the development of Parkinson’s disease-
K.
related pathology. Cell and Tissue Research, 318, 121–134.
Briggs, G. G., & Nebes, R. D. (1975). Patterns of hand preference
in a student population. Cortex, 11, 230–238.
Brooks, D. J. (1995). The role of the basal ganglia in motor control:
Journal of the Neurological Sciences,
Contributions from PET.
128, 1–13.
Buchanan, C. R., Pernet, C. R., Gorgolewski, K. J., Storkey, UN. J.,
(2014). Test-retest reliability of structural brain
& Bastin, M. E.
networks from diffusion MRI. NeuroImage, 86, 231–243.
Catani, M., & Ffytche, D. H. (2005). The rises and falls of discon-
nection syndromes. Brain, 128, 2224–2239.
Charlson, M. E., Pompei, P., Ales, K. L., & MacKenzie, C. R. (1987).
A new method of classifying prognostic comorbidity in longitu-
Journal of Chronic
dinal studies: Development and validation.
Diseases, 40, 373–383.
Chaudhuri, K. R., & Schapira, UN. H. (2009). Non-motor symptoms
of Parkinson’s disease: Dopaminergic pathophysiology and treat-
ment. The Lancet Neurology, 8, 464–474.
Cheng, H., Wang, Y., Sheng, J., Sporns, O., Kronenberger, W. G.,
Mathews, V. P., . . . Saykin, UN. J. (2012). Optimization of seed
density in DTI tractography for structural networks.
Neuroscience Methods, 203, 264–272.
Journal of
Cohen, J.
(1992). A power primer. Psychological Bulletin, 112,
155–159.
Colon-Perez, l. M., Couret, M., Triplett, W., Price, C., & Mareci,
T. H. (2016). Small worldness in dense and weighted connec-
tomes. Frontiers in Physics, 4.
Colon-Perez, l. M., King, M., Parekh, M., Boutzoukas, A.,
(2015). High-
Carmona, E., Couret, M., . . . Carney, P. R.
field magnetic resonance imaging of the human temporal lobe.
NeuroImage: Clinical, 9, 58–68.
Colon-Perez, l. M., Spindler, C., Goicochea, S., Triplett, W., Parekh,
M., Montie, E., . . . Mareci, T. H. (2015). Dimensionless, scala
invariant, edge weight metric for the study of complex structural
networks. PLoS ONE, 10, e0131493.
Colon-Perez, l. M., Tanner, J. J., Couret, M., Goicochea, S., Mareci,
T. H., & Price, C. C.
for
(2017).
“Cognition and connectome in nondementia idiopathic Parkin-
son’s disease.” Network Neuroscience, 2(1), 106–124. https://
doi.org/10.1162/netn_a_00027
Supplemental material
Crowley, S. J., Huang, H., Tanner, J. J., Zho, Q., Schwab, N. A.,
(2017). Considering total in-
Hizel, l. P., . . . Price, C. C.
tracranial volume and other nuisance variables in brain voxel
based morphometry in idiopathic PD. Brain Imaging and
Behavior, 1–12.
Daianu, M., Jahanshad, N., Nir, T. M., Jack, C. R., Jr., Weiner, M. W.,
(2015). Rich club anal-
Bernstein, M. A., & Thompson, P. M.
ysis in the Alzheimer’s disease connectome reveals a relatively
undisturbed structural core network. Human Brain Mapping, 36,
3087–3103.
Doucet, G. E., Rider, R., Taylor, N., Skidmore, C., Sharan, A.,
Sperling, M., & Tracy, J. IO.
(2015). Presurgery resting-state lo-
cal graph-theory measures predict neurocognitive outcomes after
brain surgery in temporal lobe epilepsy. Epilepsia, 56, 517–526.
Drakesmith, M., Caeyenberghs, K., Dutt, A., Zammit, S., Evans,
C. J., Reichenberg, A., . . . Jones, D. K. (2015). Schizophrenia-
like topological changes in the structural connectome of
individuals with subclinical psychotic experiences. Human Brain
Mapping, 36, 2629–2643.
Erdös, P., & Rényi, UN. (1959). On random graphs, IO. Publicationes
Mathematicae (Debrecen), 6, 290–297.
Fling, B. W., Cohen, R. G., Mancini, M., Carpenter, S. D., Fair, D. A.,
Nutt, J. G., & Horak, F. B. (2014). Functional reorganization of
Network Neuroscience
122
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
the locomotor network in Parkinson patients with freezing of gait.
PLoS ONE, 9, e100291.
Ford, UN. A., Colon-Perez, L., Triplett, W. T., Gullett, J. M., Mareci,
T. H., & Fitzgerald, D. B. (2013). Imaging white matter in human
brainstem. Frontiers in Human Neuroscience, 7, 400.
Galantucci, S., Agosta, F., Stefanova, E., Basaia, S., van den Heuvel,
M. P., Stojkovic, T., . . . Filippi, M. (2016). Structural brain con-
nectome and cognitive impairment in Parkinson disease. Radiol-
ogy, 160274.
Gattellaro, G., Minati, L., Grisoli, M., Mariani, C., Carella, F., Osio,
M., . . . Bruzzone, M. G. (2009). White matter involvement in
idiopathic Parkinson disease: A diffusion tensor imaging study.
American Journal of Neuroradiology, 30, 1222–1226.
Genovese, C., & Wasserman, l. (2002). Operating characteristics
and extensions of the false discovery rate procedure. Journal of
the Royal Statistical Society: Series B (Statistical Methodology),
64, 499–517.
Girard, G., Whittingstall, K., Deriche, R., & Descoteaux, M. (2014).
Towards quantitative connectivity analysis: Reducing tracto-
graphy biases. NeuroImage, 98, 266–278.
Golby, UN. J., Poldrack, R. A., Brewer, J. B., Spencer, D., Desmond,
J. E., Aron, UN. P., & Gabrieli, J. D. (2001). Material-specific lateral-
ization in the medial temporal lobe and prefrontal cortex during
memory encoding. Brain, 124, 1841–1854.
Golden, C. J., & Freshwater, S. M.
(2002). The Stroop Color and
Word Test: A manual for clinical and experimental uses. Chicago,
IL: Stoelting.
Granovetter, M. S.
(1973). The strength of weak ties. American
Journal of Sociology, 78, 1360–1380.
Gratwicke, J., Jahanshahi, M., & Foltynie, T.
(2015). Parkinson’s
disease dementia: A neural networks perspective. Brain, 138,
1454–1476.
Grenander, U.
(1956). On the theory of mortality measurement.
Scandinavian Actuarial Journal, 1956, 125–153.
Hagmann, P., Cammoun, L., Gigandet, X., Gerhard, S., Ellen Grant,
P., Wedeen, V., . . . Sporns, O. (2010). MR connectomics: Prin-
Journal of Neuroscience Methods, 194,
ciples and challenges.
34–45.
Heaton, R. K., & Psychological Assessment Resources Inc. (2004).
Revised comprehensive norms for an expanded Halstead-Reitan
battery: Demographically adjusted neuropsychological norms
for African American and Caucasian adults, professional manual
[Updated ed.]. Lutz, FL: Psychological Assessment Resources.
Henderson, E. J., Chu, H., Gaunt, D. M., Whone, UN. L., Ben-
Shlomo, Y., & Lyell, V. (2016). Comparison of Test Your Memory
and Montreal Cognitive Assessment measures in Parkinson’s dis-
ease. Parkinson’s Disease, 2016, 1012847.
Hoehn, M. M., & Yahr, M. D.
(1967). Parkinsonism: Onset, pro-
gression and mortality. Neurology, 17, 427–442.
Hughes, UN. J., Ben-Shlomo, Y., Daniel, S. E., & Lees, UN. J. (2001).
What features improve the accuracy of clinical diagnosis in
Parkinson’s disease: A clinicopathologic study. 1992. Neurology,
57, S34–38.
Humphries, M. D., & Gurney, K.
(2008). Network “small-world-
ness”: A quantitative method for determining canonical network
equivalence. PLoS ONE, 3, e0002051.
Janvin, C. C., Larsen, J. P., Aarsland, D., & Hugdahl, K. (2006). Sub-
types of mild cognitive impairment in Parkinson’s disease: Pro-
gression to dementia. Movement Disorders, 21, 1343–1349.
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W., &
Smith, S. M. (2012). Fsl. NeuroImage, 62, 782–790.
Ji, G. J., Zhang, Z., Xu, Q., Wei, W., Wang, J., Wang, Z., . . . Lu,
G. (2015). Connectome reorganization associated with surgical
outcome in temporal lobe epilepsy. Medicine (Baltimore), 94,
e1737.
Jian, B., Vemuri, B. C., Ozarslan, E., Carney, P. R., & Mareci,
T. H. (2007). A novel tensor distribution model for the diffusion-
weighted MR signal. NeuroImage, 37, 164–176.
Langer, N., Pedroni, A., & Jancke, l. (2013). The problem of thresh-
olding in small-world network analysis. PLoS ONE, 8, e53199.
(2012). Neuropsychological assessment (5th ed.).
Lezak, M. D.
New York, NY: Oxford University Press.
Lisanby, S. H., McDonald, W. M., Massey, E. W., Doraiswamy,
P. M., Rozear, M., Boyko, O. B., . . . Nemeroff, C. (1993). Di-
minished subcortical nuclei volumes in Parkinson’s disease by
MR imaging. Journal of Neural Transmission, 40(Suppl.), 13–21.
Luo, C. Y., Guo, X. Y., Song, W., Chen, Q., Cao, B., Yang, J., . . .
(2015). Functional connectome assessed using
Shang, H. F.
graph theory in drug-naive Parkinson’s disease. Journal of Neu-
rology, 262, 1557–1567.
Matteau, E., Dupre, N., Langlois, M.,
Jean, L., Thivierge, S.,
(2011). Mattis Dementia Rating
Provencher, P., & Simard, M.
Scale 2: Screening for MCI and dementia. American Journal of
Alzheimer’s Disease and Other Dementias, 26, 389–398.
Meijering, E. H. W., Zuiderveld, K. J., & Viergever, M. UN. (1999). Im-
age reconstruction by convolution with symmetrical piecewise
IEEE Transactions on Image Pro-
nth-order polynomial kernels.
cessazione, 8, 192–201.
Nemmi, F., Sabatini, U., Rascol, O., & Peran, P.
(2015). Parkin-
son’s disease and local atrophy in subcortical nuclei: Insight from
shape analysis. Neurobiology of Aging, 36, 424–433.
Newman, M. E. J.
(2001). Scientific collaboration networks. II.
Shortest paths, weighted networks, and centrality. Physical Re-
view E, 64.
Northam, G. B., Liegeois, F., Tournier, J. D., Croft, l. J., Johns,
P. N., Chong, W. K., . . . Baldeweg, T. (2012). Interhemispheric
temporal lobe connectivity predicts language impairment in ado-
lescents born preterm. Brain, 135, 3781–3798.
Odish, O. F., Caeyenberghs, K., Hosseini, H., van den Bogaard,
S. J., Roos, R. A., & Leemans, UN. (2015). Dynamics of the con-
nectome in Huntington’s disease: A longitudinal diffusion MRI
study. NeuroImage: Clinical, 9, 32–43.
Pagonabarraga, J., Corcuera-Solano, I., Vives-Gilabert, Y., Llebaria,
G., Garcia-Sanchez, C., Pascual-Sedano, B., . . . Gomez-Anson,
(2013). Pattern of regional cortical thinning associated with
B.
cognitive deterioration in Parkinson’s disease. PLoS ONE, 8,
e54980.
Parkinson, J. (2002). An essay on the shaking palsy. 1817. Journal of
Neuropsychiatry and Clinical Neurosciences, 14, 223–236; dis-
cussion 222.
Petrides, M. (2000). Dissociable roles of mid-dorsolateral prefrontal
and anterior inferotemporal cortex in visual working memory.
Journal of Neuroscience, 20, 7496–7503.
Network Neuroscience
123
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Connectomes in Parkinson’s disease
Price, C. C., Garrett, K. D., Jefferson, UN. L., Cosentino, S., Tanner,
J. J., Penney, D. L., . . . Libon, D. J. (2009). Leukoaraiosis sever-
ity and list-learning in dementia. Clinical Neuropsychologist, 23,
944–961.
Price, C. C., Tanner, J., Nguyen, P. T., Schwab, N. A., Mitchell,
S., Slonena, E., . . . Bowers, D.
(2016). Gray and white matter
contributions to cognitive frontostriatal deficits in non-demented
Parkinson’s disease. PLoS ONE, 11, e0147332.
Schwab, N. A., Tanner, J. J., Nguyen, P. T., Schmalfuss, IO. M.,
Bowers, D., Okun, M., & Price, C. C.
(2015). Proof of princi-
ple: Transformation approach alters caudate nucleus volume and
structure-function associations. Brain Imaging and Behavior, 9,
744–753.
Shu, N., Li, X., Mamma, C., Zhang, J., Chen, K., Liang, Y., . . . Zhang,
Z. (2015). Effects of APOE promoter polymorphism on the topo-
logical organization of brain structural connectome in nonde-
mented elderly. Human Brain Mapping, 36, 4847–4858.
Sporns, O. (2011UN). The human connectome: A complex network.
Annals of the New York Academy of Scienes, 1224, 109–125.
Sporns, O. (2011B). Networks of the brain. Cambridge, MA: MIT
Press.
Strimmer, K. (2008). Fdrtool: A versatile R package for estimating
local and tail area-based false discovery rates. Bioinformatics,
24, 1461–1462.
Stuss, D. T., Alexander, M. P., Shallice, T., Picton, T. W.,
Binns, M. A., Macdonald, R., . . . Katz, D. IO. (2005). Multiple
frontal systems controlling response speed. Neuropsychologia,
43, 396–417.
Tanner, J. J., Levy, S. A., Schwab, N. A., Hizel, l. P., Nguyen, P. T.,
Okun, M. S., & Price, C. C. (2017). Marked brain asymmetry with
intact cognitive functioning in idiopathic Parkinson’s disease: UN
longitudinal analysis. Clinical Neuropsychologist, 31, 654–675.
Tanner, J. J., Mareci, T. H., Okun, M. S., Bowers, D., Libon, D. J.,
& Price, C. C.
(2015). Temporal lobe and frontal-subcortical
dissociations in non-demented Parkinson’s disease with verbal
memory impairment. PLoS ONE, 10, e0133792.
Tanner, J. J., McFarland, N. R., & Price, C. C. (2017). Striatal and
hippocampal atrophy in idiopathic Parkinson’s disease patients
without dementia: A morphometric analysis. Frontiers in Neu-
rology, 8, 139.
Taylor, P. N., Han, C. E., Schoene-Bake, J. C., Weber, B., & Kaiser,
M.
(2015). Structural connectivity changes in temporal lobe
epilepsy: Spatial features contribute more than topological mea-
sures. NeuroImage: Clinical, 8, 322–328.
Team, R. C.
(2015). R: A language and environment for statisti-
cal computing. Vienna, Austria: R Foundation for Statistical
Computing.
Tessitore, A., Giordano, A., Russo, A., & Tedeschi, G. (2016). Struc-
tural connectivity in Parkinson’s disease. Parkinsonism and Re-
lated Disorders, 22(Suppl. 1), S56–S59.
Tomlinson, C. L., Stowe, R., Patel, S., Rick, C., Gray, R., &
Clarke, C. E. (2010). Systematic review of levodopa dose equiva-
lency reporting in Parkinson’s disease. Movement Disorders, 25,
2649–2653.
Tuch, D. S., Reese, T. G., Wiegell, M. R., Makris, N., Belliveau,
(2002). High angular resolution diffu-
J. W., & Wedeen, V. J.
sion imaging reveals intravoxel white matter fiber heterogeneity.
Magnetic Resonance in Medicine, 48, 577–582.
van Hartevelt, T. J., Cabral, J., Deco, G., Moller, A., Verde, UN. L.,
Aziz, T. Z., & Kringelbach, M. l. (2014). Neural plasticity in hu-
man brain connectivity: The effects of long term deep brain stim-
ulation of the subthalamic nucleus in Parkinson’s disease. PLoS
ONE, 9, e86496.
van Wijk, B. C., Stam, C. J., & Daffertshofer, UN. (2010). Comparing
brain networks of different size and connectivity density using
graph theory. PLoS ONE, 5, e13701.
Watts, D. J., & Strogatz, S. H.
(1998). Collective dynamics of
“small-world” networks. Nature, 393, 440–442.
Wechsler, D.
(1991). Wechsler Memory Scale (3rd ed.). San
Antonio, TX: Psychological Corporation.
Wu, T., Wang, L., Chen, Y., Zhao, C., Li, K., & Chan, P.
(2009).
Changes of functional connectivity of the motor network in the
resting state in Parkinson’s disease. Neuroscience Letters, 460,
6–10.
Wu, T., Zang, Y., Wang, L., Lungo, X., Hallett, M., Chen, Y., . . .
Chan, P. (2007). Aging influence on functional connectivity of
the motor network in the resting state. Neuroscience Letters, 422,
164–168.
Xia, M., Wang, J., & Lui, Y.
(2013). BrainNet Viewer: A network
visualization tool for human brain connectomics. PLoS ONE, 8,
e68910.
Yendiki, A., Koldewyn, K., Kakunoori, S., Kanwisher, N., & Fischl,
B. (2013). Spurious group differences due to head motion in a
diffusion MRI study. NeuroImage, 88C, 79–90.
Zgaljardic, D. J., Borod, J. C., Foldi, N. S., & Mattis, P.
(2003). UN
review of the cognitive and behavioral sequelae of Parkinson’s
disease: Relationship to frontostriatal circuitry. Cognitive and
Behavioral Neurology, 16, 193–210.
Zhan, W., Kang, G. A., Glass, G. A., Zhang, Y., Shirley, C., Millin,
R., . . . Schuff, N.
(2012). Regional alterations of brain micro-
structure in Parkinson’s disease using diffusion tensor imaging.
Movement Disorders, 27, 90–97.
Zhang, B., & Horvath, S. (2005). A general framework for weighted
gene co-expression network analysis. Statistical Applications in
Genetics and Molecular Biology, 4, Article 17.
Zola-Morgan, S., & Squire, l. R.
(1986). Memory impairment in
monkeys following lesions limited to the hippocampus. Behav-
ioral Neuroscience, 100, 155–160.
Zola-Morgan, S., Squire, l. R., & Amaral, D. G.
(1986). Umano
amnesia and the medial temporal region: Enduring memory
impairment following a bilateral lesion limited to field CA1 of
the hippocampus. Journal of Neuroscience, 6, 2950–2967.
Network Neuroscience
124
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
2
1
1
0
6
1
9
5
9
4
4
3
N
e
N
_
UN
_
0
0
0
2
7
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3