RESEARCH
High-amplitude network co-fluctuations linked to
variation in hormone concentrations over
the menstrual cycle
Sarah Greenwell1, Joshua Faskowitz1,2, Laura Pritschet3, Tyler Santander3,
Emily G. Jacobs3,4, and Richard F. Betzel1,2,5,6
1Department of Psychological and Brain Sciences, Indiana University, Bloomington, IN, USA
2Program in Neurosciences, Indiana University, Bloomington, IN, USA
3Department of Psychological and Brain Sciences, University of California, Santa Barbara, Santa Barbara, CA, USA
4Neuroscience Research Institute, University of California, Santa Barbara, Santa Barbara, CA, USA
5Cognitive Science Program, Indiana University, Bloomington, IN, USA
6Network Science Institute, Indiana University, Bloomington, IN, USA
Keywords: Edge-centric, Functional connectivity, Time-varying networks
ABSTRACT
Many studies have shown that the human endocrine system modulates brain function,
reporting associations between fluctuations in hormone concentrations and brain connectivity.
Tuttavia, how hormonal fluctuations impact fast changes in brain network organization
over short timescales remains unknown. Here, we leverage a recently proposed framework
for modeling co-fluctuations between the activity of pairs of brain regions at a framewise
timescale. In previous studies we showed that time points corresponding to high-amplitude
co-fluctuations disproportionately contributed to the time-averaged functional connectivity
pattern and that these co-fluctuation patterns could be clustered into a low-dimensional set of
recurring “states.” Here, we assessed the relationship between these network states and
quotidian variation in hormone concentrations. Specifically, we were interested in whether the
frequency with which network states occurred was related to hormone concentration. Noi
addressed this question using a dense-sampling dataset (N = 1 brain). In this dataset, a single
individual was sampled over the course of two endocrine states: a natural menstrual cycle
and while the subject underwent selective progesterone suppression via oral hormonal
contraceptives. During each cycle, the subject underwent 30 daily resting-state fMRI scans
and blood draws. Our analysis of the imaging data revealed two repeating network states.
We found that the frequency with which state 1 occurred in scan sessions was significantly
correlated with follicle-stimulating and luteinizing hormone concentrations. We also
constructed representative networks for each scan session using only “event frames”—those
time points when an event was determined to have occurred. We found that the weights of
specific subsets of functional connections were robustly correlated with fluctuations in the
concentration of not only luteinizing and follicle-stimulating hormones, but also progesterone
and estradiol.
AUTHOR SUMMARY
We study a dense sampling dataset of one brain (N = 1) imaged across two complete menstrual
cycles (60 scan sessions). We identify network states—high-amplitude patterns of time-varying
connectivity that reoccur across scan sessions—and show that the frequency with which states
a n o p e n a c c e s s
j o u r n a l
Citation: Greenwell, S., Faskowitz, J.,
Pritschet, L., Santander, T., Jacobs,
E. G., & Betzel, R. F. (2023). High-
amplitude network co-fluctuations
linked to variation in hormone
concentrations over the menstrual
cycle. Network Neuroscience, 7(3),
1181–1205. https://doi.org/10.1162/netn
_a_00307
DOI:
https://doi.org/10.1162/netn_a_00307
Supporting Information:
https://doi.org/10.1162/netn_a_00307
Received: 8 settembre 2021
Accepted: 20 Dicembre 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Richard F. Betzel
richard.betzel@gmail.com
Handling Editor:
Michael Cole
Copyright: © 2023
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
T
/
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network dynamics are associated with reproductive hormones
occur is linked to endogenous fluctuations in follicle-stimulating and luteinizing hormones.
We further show that the weights of scan-specific and whole-brain co-fluctuation patterns are
broadly associated with hormone fluctuations.
INTRODUCTION
The human brain is a complex network composed of structurally connected neural elements
that help shape brain activity and give rise to widespread patterns of functional coupling
(Hermundstad et al., 2013; Sporns, Tononi, & Kötter, 2005). The organization and topology
of these structural and functional networks can be interrogated using tools from network sci-
ence (Bullmore & Sporns, 2009), revealing organizing principles including short processing
paths (Bassett & Bullmore, 2006), hubs and rich clubs (Hagmann et al., 2008; van den Heuvel
& Sporns, 2011), modular structure (Sporns & Betzel, 2016), and cost-efficient spatial embed-
ding (Bullmore & Sporns, 2012).
Recent work has shown that whole-brain patterns of functional coupling between brain
regions vary over short timescales (Allen et al., 2014; Barttfeld et al., 2015; R. F. Betzel,
Fukushima, Lui, Zuo, & Sporns, 2016). To reconstruct changes in network structure over time,
most studies use sliding-window analyses (Hindriks et al., 2016; Preti, Bolton, & Van De Ville,
2017). In this approach, a functional network is estimated using only those samples that fall
within a window of some fixed duration. The window is then advanced a certain number of
frames, resulting in a time series of functional networks. Although applied widely, Questo
approach has a number of drawbacks. Namely, it forces the user to specify parameters for
window length and amount of overlap between successive windows. The windowing proce-
dure, itself, also makes it impossible to precisely localize a network state to a specific moment
in time and resolve changes in network structure over short timescales (Hindriks et al., 2016;
Leonardi & Van De Ville, 2015; Liégeois, Laumann, Snyder, Zhou, & Sì, 2017; Lurie et al.,
2020).
Recentemente, we proposed edge time series (ETS) as a method for decomposing functional net-
works into time-varying components (Esfahlani et al., 2020; Faskowitz, Esfahlani, Jo, Sporns, &
Betzel, 2020; Sporns, Faskowitz, Teixeira, Cutts, & Betzel, 2021). This approach helps address
some of the limitations of sliding-window analyses, in that it is parameter-free and can resolve
changes in network structure at a framewise timescale. In previous studies, we used this
method to show that fast network dynamics are not smooth, but rather are bursty, identifying
long periods of quiescence punctuated by brief, network-wide, high-amplitude events (R. F.
Betzel, Cutts, Greenwell, Faskowitz, & Sporns, 2022; Esfahlani et al., 2020; Pope, Fukushima,
Betzel, & Sporns, 2021). These events are of particular interest, as time-averaged functional
networks can be accurately reconstructed from a very small number of events. We also found
that the patterns of co-fluctuation expressed during events contain disproportionate amounts of
information about an individual and that events can improve brain-behavior correlations.
Despite their apparent utility, the application of edge time series for linking brain dynamics
to cognitive, clinical, or physiological phenomena has been limited (Cooper, Kurkela, Davis,
& Ritchey, 2021; Esfahlani et al., 2022; Sorrentino et al., 2021; Tanner et al., 2022).
Edge time series are well situated for investigating relationships between brain connectivity
and physiological variables that also fluctuate over short timescales. A good example is the
human menstrual cycle, which is typified by variations in the sex steroid hormones
estradiol—the major form of estrogen in most mammals—and progesterone and
Edge or co-fluctuation time series:
The element-wise product of two
parcel time series; the average value
of an edge time series is exactly the
functional connectivity weight.
Event:
Frames when many edge time series
exhibit greater-than-expected
amplitudes; detected statistically by
comparing the global amplitude of
edge time series (root sum of squared
values) with a temporal null model.
Network Neuroscience
1182
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network dynamics are associated with reproductive hormones
Functional connectivity:
Operationalized as the correlation
coefficient between two parcel time
series.
gonadotropins follicle-stimulating hormone (FSH) and luteinizing hormone (LH). Briefly,
during the follicular phase of the menstrual cycle estradiol concentrations rise, prompting
the growth of the uterine lining. Immediately prior to ovulation, FSH encourages an immature
follicle to complete its development into an egg for release from the ovaries. LH promotes the
release of the egg into the fallopian tubes, followed by an increase in progesterone during
the luteal phase, during which the uterine lining thickens, creating a favorable environment
for the egg to be preserved. These hormones exhibit neuromodulatory effects and have been
associated with variation in brain connectivity, neural structure, and brain activity (Beltz &
Moser, 2020; Berman et al., 1997; Jacobs & D’Esposito, 2011; McEwen, Akama, Spencer-
Segal, Milner, & Waters, 2012; Taxier, Gross, & Frick, 2020). Tuttavia, most previous work
has involved sparse cross-sectional studies that capture one or more time points across the
menstrual cycle (Hidalgo-Lopez et al., 2020; Pletzer, Harris, Scheuringer, & Hidalgo-Lopez,
2019; Weis, Hodgetts, & Hausmann, 2019) or cross-sectional study designs with incomplete
sampling of participants’ cycles and comprised of heterogeneous cohorts of individuals
(Dubol et al., 2021).
Recentemente, a series of “dense sampling” studies have characterized quotidian variation in
these hormones and their relationships with functional connectivity and network community
structure (Fitzgerald, Pritschet, Santander, Grafton, & Jacobs, 2020; Mueller et al., 2021;
Pritschet et al., 2020; Taylor et al., 2020). The design of these studies parallels that of other
recent dense sampling studies (Braga & Buckner, 2017; Gordon et al., 2017; Laumann et al.,
2015), acquiring data from, in this case, a single individual over the course of two complete
menstrual cycles, one in which the participant was naturally cycling and another while the
same participant was placed on oral hormonal contraception that selectively suppressed
progesterone concentrations by ≈97% (referred to as Studies 1 E 2) (Pritschet, Taylor,
Santander, & Jacobs, 2021). Oral contraceptives are estimated to be used by over 100 million
women of reproductive age worldwide (Christin-Maitre, 2013). “The pill” generally contains
synthetic analogs of combined estrogen and progesterone or only progesterone. The pill
inhibits the spike in gonadotropins that induces ovulation, thereby preventing pregnancy. Questo
user followed the typical pattern of intake, taking the active hormonal pill for 3 weeks before
taking a nonhormonal ‘sugar pill’ during the fourth and final week. Previous studies using
these datasets have identified spatially diffuse increases in functional connectivity and modu-
lar flexibility coincident with peaks in estradiol; progesterone, by contrast, was predominantly
associated with reductions in connectivity (Mueller et al., 2021; Pritschet et al., 2020). How-
ever, the dynamic underpinnings of these associations have not been fully explored.
Here, we analyze these two fMRI datasets, one during the subject’s natural cycle and the
other while the subject was placed on an oral contraceptive regimen. We do not compare
between hormone states in this study. Invece, we pool data from both states in our analyses.
By pooling data from a synthetically altered hormonal state and a natural hormonal state in the
same individual, we can examine graded and continuous relationships between brain net-
works and the concentrations of individual hormones. In more detail, we follow the analysis
pipeline from R. F. Betzel, Cutts, et al. (2022), in which we use edge time series to detect
events from functional magnetic resonance imaging (fMRI) data and cluster whole-brain co-
fluctuation patterns into a low-dimensional set of states. Our primary aim is to assess whether
properties of these states can be statistically linked to hormone fluctuations and, if so, whether
those associations exhibit specificity such that they implicate specific subsets of hormones and
not others. In agreement with previous studies, we find that events can be subdivided into two
distinct clusters. In particular, we show that the frequency with which the first cluster appears
in a given scan is strongly correlated with quotidian variation in both FSH and LH.
Network Neuroscience
1183
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
/
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network dynamics are associated with reproductive hormones
Inoltre, we show that day-to-day variation in the edge- and system-level configuration of
the first cluster is broadly associated with both FSH and LH, as well as progesterone and estra-
diol. Collectively, our results establish a new link between high-amplitude, network-level
events and the human endocrine system, opening up new avenues for exploring the dynamic
interplay between brain-hormone relationships.
RESULTS
Clustering High-Amplitude Co-Fluctuations Reveal Distinct Patterns of Connectivity
Recent methodological advances have made it possible to track rapid fluctuations in func-
tional network architecture, revealing the presence of “events”—short-lived and high-
amplitude patterns of network-wide fluctuations (Esfahlani et al., 2020; Faskowitz et al.,
2020; Sporns et al., 2021). The results of previous studies suggested that events are low dimen-
sional, such that a small repertoire of patterns are reiterated (R. F. Betzel, Cutts, et al., 2022;
Pope et al., 2021). Here, we test whether this was also the case in an independently acquired
dataset of a single individual. Figura 1 depicts a schematic of our general analysis pipeline.
To detect and assess whether there were repeated patterns of high-amplitude events, we
used simple statistical procedure to identify sequences of temporally contiguous frames whose
root summed square (RSS) exceeded that of a null model in which regions’ activity time
courses were randomized (circular shifts). Note that this null model approximately preserves
local properties of regional activity, but destroys brain-wide correlation structure. Così, Questo
model is not, strictly speaking, a test of dynamics in co-fluctuations. We retained only those
sequences that were not obviously coincident with periods of excessive in-scanner movement,
questo è, did not include a high-motion frame and were at least two frames away from any high-
motion frame. For each such sequence, we extracted the co-fluctuations expressed during the
frame with the highest RSS value as a representative peak. In total, we detected 899 motion-
free events that occurred during low-motion intervals (14.98 ± 5.27 events per scan session).
Node time series:
The mean fMRI BOLD activity for a
region of interest, network node, O
atlas parcel.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
T
/
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Analysis pipeline. (UN) After preprocessing, we obtained parcellated regional time series from 60 scans (spanning two experiments).
(B) For a given scan, we transformed node time series into edge time series following Esfahlani et al. (2020). (C) Prossimo, we detected high-
amplitude events in each scan and for each event extracted its representative pattern (the frame with the greatest amplitude). Generalmente,
we obtained a different number of events per scan. (D) We aggregated event patterns from all scans and collectively clustered them using
modularity maximization. This procedure resulted in multiple community centroids (we analyze the two largest) and a count of how many
times a given community appeared on a given scan session. (E) In parallel, we analyzed hormone data that were collected concurrent with
each scan session. Our principal aim was to link features of communities (brain states) with hormone data. (F) Inoltre, we reconstructed
estimates of communities for each of the 60 scan sessions and, for each edge, computed the correlation of its co-fluctuation across sessions
with hormone concentrations.
Network Neuroscience
1184
Network dynamics are associated with reproductive hormones
Event community, cluster, or state:
After detecting events, we can group
them into clusters using data-driven
algorithms and based on their spatial
similarity to one another.
To detect putative states, clusters, or communities of events (note that here we use these
terms interchangeably), we calculated for each event its whole-brain co-fluctuation
pattern—the N × N matrix whose element {io, j} was equal to the product of z-scored activity
in regions i and j at the time of the event. We then vectorized the event co-fluctuation patterns
E, for all pairs of events, computed their spatial similarity (Pearson correlation), resulting in a
899 × 899 matrix. We then used a variant of modularity maximization to assign each co-
fluctuation pattern to a single cluster (see Materials and Methods for details). As in previous
studies, we found evidence for two large consensus communities that appeared in most scan
sessions and collectively accounted for ≈70.2% of all detected states (Figure 2A). These clus-
ters were highly reproducible across repeated runs of the modularity optimization algorithm
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
/
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. Modularity maximization and network states. We used an event detection algorithm to identify instances of “high-amplitude”
co-fluctuations, extracting 899 brain-wide patterns. Each putative event was represented as a region-by-region co-fluctuation matrix. Noi
vectorized each matrix by extracting its upper triangle elements and calculated the spatial similarity (Pearson correlation) between all pairs
of patterns. This resulted in a 899 × 899 similarity matrix. We used this matrix as input to a clustering algorithm (modularity maximization) A
detect groups or “communities” of mutually similar co-fluctuation patterns. Modularity maximization is nondeterministic, so we repeated the
algorithm 1,000 times with random restarts and assembled the partitions into a co-assignment matrix, whose elements counted the fraction of
IL 1,000 partitions in which any pair of co-fluctuation patterns were assigned to the same community. To resolve variability across runs, we
iteratively clustered the co-assignment matrix to obtain “consensus communities.” We refer to each consensus community as a “state.” (UN)
Similarity matrix, ordered by consensus communities. (B) Community co-assignment matrix ordered by consensus communities. (C) Vectorized
co-fluctuation patterns ordered by consensus communities. (D) Mean co-fluctuation matrix for cluster 1, ordered by canonical brain systems.
(E) First principal component of the co-fluctuation matrix. (F) Elements of first principal component grouped by brain system. Panels G–I are
analogous to D–F but for cluster 2.
Network Neuroscience
1185
Network dynamics are associated with reproductive hormones
(Figure 2B) and divided event patterns into cohesive clusters (Figure 2C). We note that these
clusters were also robust to variation in processing pipelines, after splitting the data by exper-
iment (see Supporting Information Figure S2), and after systematically excluding data from
individual scans in calculating the correlation (Supporting Information Figure S6). Finalmente,
we also assessed what effect using null distributions of RSS values from different scan sessions
had on the detected events. Generalmente, we found a near perfect correspondence, suggesting
that event structure is not strongly impacted by quotidian variation in the null distribution
(Supporting Information Figure S3).
È interessante notare, the spatial patterns of these clusters closely recapitulate those reported in pre-
vious studies (R. F. Betzel, Cutts, et al., 2022; Esfahlani et al., 2020). Comunità 1, for
instance, was characterized by opposed fluctuations between regions in the default mode
network with those in dorsal and salience/ventral attention networks (Figure 2D). To better
understand whether these co-fluctuations were underpinned by a specific mode or pattern
of activity, we performed a singular value decomposition of the mean co-fluctuation matrix,
revealing, as expected, a node-level pattern characterized by strong fluctuations in default
mode regions and opposed fluctuations in attentional and, to some extent, sensorimotor sys-
tems (Figure 2E and F). Allo stesso modo, the spatial pattern of community 2 was characterized by
opposed co-fluctuations of control and dorsal attention regions with the default mode network
(Figure 2G–I).
The remaining communities collectively accounted for <30% of all events, with the next
most frequent community accounting for 9.8% of all events (but appearing in ≈71.7% of scan
sessions). For this reason, we focus on the first two communities for all subsequent analyses.
In the Supporting Information we describe the remaining communities in greater detail (Sup-
porting Information Figure S1).
State Frequency Is Associated With LH and FSH Concentrations
Cluster analysis of co-fluctuation time series revealed the presence of repeating patterns or
states. However, the biological relevance of these states remains unclear. In this section, we
show that the frequency with which these states appear across scan sessions is robustly related
to endogenous variation in the gonadotropins, luteinizing, and follicle-stimulating hormone,
but not statistically associated with sex hormones estradiol and progesterone.
To link high-amplitude co-fluctuations with quotidian variation in hormone concentration
(see Figure 3C and D), we calculated for each scan session the number of times that each
community (1 and 2) appeared. We found that, on average, communities 1 and 2 appeared
6.15 ± 3.17 and 4.37 ± 2.57 times (≈41% and ≈29% of all events; maximum values of 18 and
11), respectively. Next, we computed the Spearman rank correlation of these frequencies with
hormone concentrations. Note that the rank correlation reduces statistical biases originating
from spikes in both luteinizing and follicle-stimulating hormone around ovulation. For all
reported correlations we corrected for multiple comparisons by fixing the false discovery rate
at q = 0.05, resulting in an adjusted critical value of padj = 0.0067. We found statistically sig-
nificant correlations between the frequency of community 1 with both gonadotropic hormones
(ρ1,FSH = 0.47 and ρ1,LH = 0.42; p = 0.0002 and p = 0.0007; effect sizes of d = 1.07 and d =
0.93, Cohen’s d ). Interestingly, the correlations between community 1 and progesterone
(ρ1,P = −0.33; p = 0.007) and community 2 with luteinizing hormone (ρ2,LH = 0.26; p =
0.02; effect sizes of d = 0.35 and d = 0.27, Cohen’s d ) were both statistically significant at
uncorrected levels, but failed to pass after multiple comparison corrections (see Supporting
Information Figure S8). The results described above were obtained after pooling data from
Network Neuroscience
1186
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
Figure 3. Correlations between state frequency and quotidian variation in gonadotropin concen-
tration. (A) Scatterplot showing concentration of follicle-stimulating hormone across scan sessions
versus the frequency with which cluster 1 appeared in a given scan. (B) Scatterplot showing con-
centration of luteinizing hormone across scan sessions versus the frequency with which cluster 1
appeared in a given scan. Panels C and D show variation of progesterone, estradiol, follicle-
stimulating hormone, and luteinizing hormone across the Study 1 and Study 2 datasets.
two separate experiments to obtain 60 observations in total (30 observations per experiment),
improving statistical power. However, the protocols differed between experiments: in one the
subject took an oral contraceptive and in the other the subject did not. To address the concern
that the results are driven by one of the experiments and not the other or from inappropriately
combining dissimilar protocol, we repeated our analyses separately for the two datasets (30
observations each). We find that those results are consistent with what was reported in the
main text with no discernible differences between the two datasets, suggesting that combining
the two is reasonable (Supporting Information Figure S4). We also compared hormone concen-
trations between experiments and found that, of the four hormones reported here, only pro-
gesterone exhibited significant differences (Supporting Information Figure S5). Note that we
also verify that results are not dependent on processing pipeline, replicating our main effects
using a pipeline in which global signal regression was not included (Supporting Information
Figure S9).
Notably, however, we found that both FSH and LH were correlated with total event count
(the number of events detected in a given scan session; ρFSH,total = 0.21 and ρLH,total = 0.33) and
that when we expressed the event count for state 1 as a proportion of the event count for each
session, the effect was in the same direction but attenuated (ρFSH,prop = 0.29 and ρLH,prop = 0.09).
These observations suggest that the relationship of hormone concentrations with event structure
is dually global—linked to the overall event count on any given scan session—but allows for
local associations with the frequency and proportion of specific co-fluctuation states.
Collectively, these results suggest that endogenous and exogenously induced changes in
gonadotropic hormone concentrations are related to the expression of distinct brain network
states. These findings address our stated aim of linking brain states defined based on edge time
Network Neuroscience
1187
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
series with hormonal fluctuations. Further, these findings suggest some specificity at both the
brain and endocrine levels, implicating only a single brain state and a pair of hormones (FSH
and LH, but not estradiol or progesterone). Finally, these findings also suggest that hormones
may have a role in modulating variation in high-amplitude network-level co-fluctuations.
Quotidian Variation in Edge-Level Co-Fluctuations and Hormone Concentrations Are Correlated
In the previous section, we demonstrated that day-to-day variation in gonadotropin concen-
tration was linked to the frequency with which particular high-amplitude “states” are
expressed. In that analysis, any co-fluctuation pattern assigned a given community label
was treated as a recurrence of the same state. In reality, however, the whole-brain patterns
classified by these state motifs exhibited variability between days and even within instances
during the same scan. Here, we demonstrate that within-state variation in edge co-fluctuation
magnitude is linked not only with gonadotropin concentration, but also with the concentra-
tions of sex hormones estradiol and progesterone.
To link edge-level co-fluctuations with hormones, we needed estimates of communities 1
and 2 for each scan session. To do this, we identified all co-fluctuation patterns assigned to a
given community on each scan session and averaged those. Note that in the case of commu-
nity 1, there was one scan in which it never appeared; in the case of community 2, there were
two. We omitted these scans from all analyses carried out in this section. For each of the
remaining scans, we created a representative version of community 1 and 2 centroids by aver-
aging all co-fluctuation assigned to that community. We also kept track of the number of sam-
ples used to compute the representative pattern (i.e., the number of times that a given state was
present in each scan) and after aggregating across all scans, regressed out this number from
each node pair, retaining the residuals and calculating their Spearman rank correlation with
the concentrations of progesterone, estradiol, luteinizing hormone, and follicle-stimulating
hormone.
Mass univariate edge-level analyses can lack statistical power to resolve certain effects.
Here, for instance, we performed 79,800 tests (the number of edges) with the aim of identifying
those that pass a criterion for statistical significance (for visualization only, we show the edges
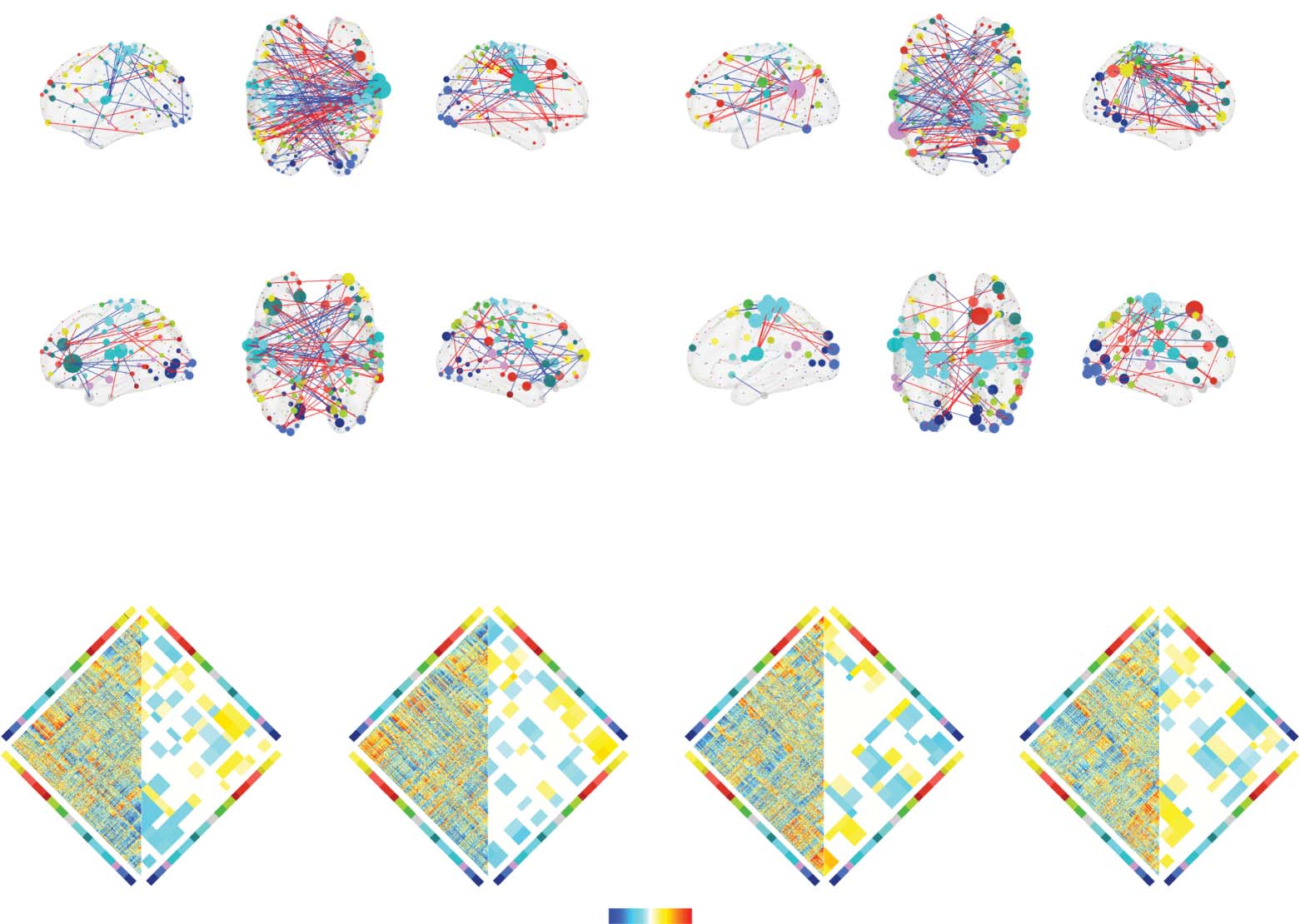
with the strongest positive and negative correlations embedded in anatomical space in
Figure 4A–D). An alternative strategy is to perform statistical testing at the level of brain sys-
tems or communities. Because nodes are aggregated by community, this approach necessarily
limits one’s ability to resolve focal, regional effects. However, because the number of compar-
isons is reduced by an order of magnitude or more, statistical power increases.
Here, we aggregated edge-level correlations by brain system (Schaefer et al., 2018), trans-
forming a correlation matrix of dimensions ρregion 2 [400 × 400] into a ρsystem 2 [16 × 16]
matrix. The elements of ρsystem represented the mean correlation coefficients between all pairs
of regions assigned to any two systems. We repeated this procedure for all four hormones
(Figure 4E–F), yielding four system-level correlation maps. To identify significant correlations,
we repeated the aggregation procedure after randomly permuting system labels using a “spin”
test to approximately preserve spatial dependencies between regions (1,000 permutations)
(Váša et al., 2018).
In general, we found that the regional correlation patterns for sex hormones were similar to
one another (rP,E = 0.27; p < 0.05). The same was true for gonadotropins (rFSH,LH = 0.38; p <
0.05) (Figure 5A). Due to their similarity and for ease of description, we combined system-level
correlation patterns for progesterone with estradiol and FSH with LH, focusing on shared
effects (these combined patterns are anticorrelated with one another; p < 0.05; Figure 5B).
Network Neuroscience
1188
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Edge- and system-level correlations with hormone concentration. We calculated the correlation of hormone concentrations with
Figure 4.
edge-level co-fluctuation magnitudes for cluster 1. This procedure resulted in a correlation coefficient at every edge (Nedges = 400 × 399/2 =
79,800) and separately for every hormone. Rather than perform statistical tests at the level of edges, we performed tests at the level of systems.
That is, for every pair of systems, we calculated the mean correlation of all edges that fall between those two systems and compare that value
against what would be expected by chance. This procedure results in Nsystempairs = 136 unique pairs of systems and offers greater statistical
power due to fewer independent tests. We corrected for multiple comparisons by fixing the accepted false discovery rate (the probability of
falsely detecting an effect when no such effect exists) to 5% and adjusted the critical p value to obtain this accepted rate. Thus, for every pair of
systems and for every hormone, we were able to assess whether the mean correlation of edges between those systems was statistically stronger
than expected. These results are shown in panels E–H. On the left-hand side of each diamond plot are the raw, uncorrected edge-level cor-
relations. On the right-hand side are system-level correlations that were greater than expected. System pairs that did not pass statistical testing
are shown in white. The matrices in panels E–H are not anatomically localized and may be difficult to interpret. To provide some intuition and
to anatomically ground these effects, in panels A–D, we show only the strongest edge-level correlations projected back into anatomical space.
In each panel, node size is proportional to the mean correlation magnitude of a node’s edges. Node color was determined by brain system.
Edge color denotes positive (red) and blue (negative) correlations. Note, however, that panels A–D are for provided for the sake of visualization
and intuition only. The statistical tests and their outcomes are shown in panels E–H.
To summarize shared effects, we classified every pair of systems based on the concordance of
correlation patterns. That is, whether both, one, or neither hormones exhibited effects in the
same direction (false discovery rate fixed at q = 0.05 resulting in adjusted critical value of
padj = 0.0036; Figure 5C and D).
In both cases, we found evidence of broad, brain-wide constellations formed by significant
system-level correlations. In the case of sex steroid hormones, correlations involving the con-
trol and default mode networks tended to be positive, while correlations involving attention,
somatomotor, and visual network tended to be negative (Figure 5C). On the other hand, in
the case of gonadotropins, positive and negative correlations were more uniformly distributed
across brain systems, with salience/ventral attention, somatomotor, and visual networks
ranking among the systems with the greatest overall number of significant correlations
(Figure 5D).
Network Neuroscience
1189
Network dynamics are associated with reproductive hormones
Figure 5. Similarity of brain-wide correlation patterns across hormones. In the main text we described a procedure in which we calculated
the correlation of hormone concentration with the co-fluctuation magnitude between every pair of nodes (edges). This procedure resulted in
four node × node matrices of correlation coefficients (one for each of the four hormones studied here). (A) Similarity of correlation patterns
between pairs of hormones. (B) Plotting hormones against each other yields dense scatterplots that are difficult to interpret. Here, we present
the same data as a two-dimensional histogram (by binning x and y axes and counting the number of points that fall within each bin). Here, the
two-dimensional histogram depicts the similarity of correlation patterns from combined sex hormones (progesterone + estradiol) and gonad-
otropins (FSH + LH). The color is linearly proportional to the number of points in each bin, with brighter colors corresponding to larger values.
(C) To further examine similarities and differences between the correlation patterns, we performed a concordance analysis. Specifically, we
identified pairs of brain systems in which the mean correlation was statistically greater/less than that of a permutation based null model. We
further examined similarities in sex and gonadotropic hormones separately. For every pair of brain systems, there were five possible outcomes:
both exhibited significant positive correlations, both exhibited significant negative correlations, one or the other exhibited a significant positive
correlation while the other did not, one or the other exhibited a significant negative correlation while the other did not, and neither exhibited a
significant correlation. In panel C, we plot the outcomes of this analysis for sex hormones (progesterone and estradiol). Large and small circles
indicate high and low levels of concordance. Red and blue colors indicate positive and negative correlations, respectively. The bar plot to the
right of the matrix is a count of the total number of high concordance interactions in which a given system interacts. Panel D depicts analogous
information for combined gonadotropins (FSH + LH).
We also repeated this analysis using daily estimates of edge-level co-fluctuations for com-
munity 2, resulting in four correlation maps (see Supporting Information Figure S7). Overall,
we find that LH and FSH exhibit similar whole-brain, edge-level correlation patterns (rFSH,LH =
0.28) but that progesterone and estradiol are largely dissimilar (rP,E = 0.06). Interestingly, LH
and estradiol also exhibit strong correlations (rLH,E = 0.49) while the relationship is reversed
with FSH and estradiol (rFSH,E = −0.35). As with community 1, the edge level correlations did
not clearly favor any particular set of systems; instead, they implicated a constellation of brain
areas and systems. In the case of progesterone, some of the strongest correlations involved
regions in the default mode and salience/ventral attention networks. Estradiol fluctuations,
on the other hand, exhibited strong correlations with edges incident upon visual and control
network nodes. In the case of gonadotropins, there was less consensus as to which specific
systems were implicated, as the correlation patterns were more diffuse and broadly distributed.
However, control, somatomotor, and visual networks appear particular prominent.
Here, we study brain-hormone correlations using cluster centroids from edge time series.
An important question is whether these correlational patterns are similar to those obtained in
previous studies using FC instead of centroids. Accordingly, we repeated our correlational
analysis, substituting static FC matrices for each scan session for the cluster centroids and
obtaining matrices of edge-level correlations. We then computed the similarity of these corre-
lation patterns with those obtained using cluster centroids (similarity is measured as the Spear-
man correlation of upper triangle elements). In general, we find a broad correspondence
between the two approaches (mean correlation of ρ = 0.30 ± 0.06). However, these results
also indicate that the majority of variance remains unexplained, suggesting that the cluster
centroids and FC analyses may reveal distinct brain-hormone relationships at the edge level.
Collectively, these findings suggest that the co-fluctuation between specific pairs of brain
regions are associated with concentrations of sex steroid hormones as well as gonadotropins.
Network Neuroscience
1190
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
As in the previous section, these results strengthen the link between features of edge time
series, which disclose fast fluctuations in coactivity, and hormonal fluctuations. These relation-
ships are expressed through distributed, brain-wide constellations of edges linking together
many brain systems.
DISCUSSION
Here, we aimed to link fast fluctuations in coactivity with hormonal fluctuations and to assess
the specificity of those effects. To this end, we built upon previous analyses of a dense-
sampling dataset in which a single participant underwent daily MRI scans and serological sam-
pling over the course of two full menstrual cycles (Fitzgerald et al., 2020; Mueller et al., 2021;
Pritschet et al., 2020; Taylor et al., 2020). Leveraging “edge time series” (Esfahlani et al., 2020;
Faskowitz et al., 2020), we detected high-amplitude events in each scan session, clustering
them into two large communities. The first community reflects opposed activation of default
mode and control regions with attentional and sensorimotor regions. We found that the fre-
quency with which it appears across scan sessions was strongly coupled to quotidian fluctu-
ations in the gonadotropic hormones, follicle-stimulating hormone and luteinizing hormone.
We find that variability in the scan-specific co-fluctuation pattern of community 1 was linked
to all four hormones at the level of connections. Our work sets the stage for future studies to
investigate relationships between fast fluctuations in network organization and hormones.
High-Amplitude Co-Fluctuations Are Linked to Endocrine System
Previous applications of edge time series analysis to functional imaging data have reported
brief, intermittent, and high-amplitude “events” (R. F. Betzel, Cutts, et al., 2022; Esfahlani
et al., 2020, 2022; Pope et al., 2021; Sporns et al., 2021). These studies have shown that
the co-fluctuation patterns expressed during events contribute disproportionately to the
time-averaged pattern of FC, are subject-specific, can be clustered into a small number of
putative “states,” and strengthen brain-behavior associations. For instance, in Esfahlani et al.
(2020), the authors show that the overall magnitude of brain-behavior correlations increases
when the brain measures are derived for high-amplitude co-fluctuations compared to low-
amplitude. However, the physiological underpinnings of high-amplitude co-fluctuations
remain unclear.
Here, we present evidence that both the frequency with which high-amplitude states occur
as well as their topology are strongly correlated with day-to-day fluctuations in hormone con-
centration. The findings suggest that hormones may have a role in modulating variation in
high-amplitude network-level co-fluctuations. While this observation is in line with previous
studies that have reported links between reproductive hormones and both brain activity
(Berman et al., 1997; Jacobs & D’Esposito, 2011) and functional connectivity (Arélin et al.,
2015; Dan et al., 2020; Fitzgerald et al., 2020; Hidalgo-Lopez, Zeidman, Harris, Razi, &
Pletzer, 2021; Liparoti et al., 2021; Lisofsky et al., 2015; Petersen, Kilpatrick, Goharzad, &
Cahill, 2014; Pritschet et al., 2020; Weis et al., 2019), our results extend this link to ultrafast
network dynamics. Changes in network structure at this timescale have been relatively unex-
plored in previous studies due to their cross-sectional study design and low temporal resolu-
tion of sliding-window methods for estimating time-varying functional connectivity (Dubol
et al., 2021) (although, see Mueller et al., 2021).
While the design of our study is not well-suited to support claims about causality, we spec-
ulate that by modulating receptor density during the menstrual cycle, hormones may shift the
sensitivity of neural circuits, yielding brain states that are increasingly excitable and more
Network Neuroscience
1191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
likely to produce high-amplitude events (Li et al., 2019; Liu & Herbison, 2013; Shine, Aburn,
Breakspear, & Poldrack, 2018). Indeed, recent studies have demonstrated that the sex hor-
mones estrogen and progesterone differentially influence the density of steroid hormone
receptors across the brain throughout the menstrual cycle (McEwen & Milner, 2017). More-
over, this view aligns with the nonlinear and nonstationary contribution of luteinizing
hormone to hormone dynamics across the menstrual cycle; large surges in LH near ovulation
disrupt the phasic coupling of sex steroid and gonadotropic hormones (Dubol et al., 2021),
suggesting that structural changes caused by acute changes in hormone concentration, such
as receptor density, influence network connectivity in addition to the direct effects of
hormones on cells. Sex hormones are intrinsically involved in micro-level structural changes
as well, given that estrogen- and progesterone-mediated changes in subcortical receptor
density induce the release of gonadotropin-releasing hormone to induce luteinizing hormone
spike (Sinchak, Mohr, & Micevych, 2020).
Another possible explanation for why fluctuations in reproductive hormones are associated
with brain network dynamics concerns their role as neurotransmitters. Previous studies have
established links between neurotransmitters and functional connectivity, demonstrating that
regional variation in the concentration of neurotransmitter-related neurons modulate large-
scale brain activity, shaping patterns of connectivity and the spontaneous emergence of
resting-state networks (Dang, O’Neil, & Jagust, 2012). For example, dopamine and serotonin
oppositionally influence anticorrelations between large-scale networks, such that dopamine is
associated with increases and decreases in functional connectivity of the somatomotor and
default mode, respectively, while serotonin is associated with the opposite (Conio et al.,
2020). Other studies have found that neurotransmitter effects on functional connectivity are
connected to regional differences in excitatory-inhibitory receptor and hormone ratios (van
den Heuvel et al., 2016). Gonadal hormones are increasingly recognized as critical neuro-
modulators of learning and memory (Taxier et al., 2020), and can pass through the blood-brain
barrier and are known to impact brain function, including memory and anxiety-level behavior
(Moraga-Amaro, van Waarde, Doorduin, & de Vries, 2018). These observations, combined
with the fact that edge time series are mathematically exact decompositions of functional con-
nectivity into time-varying contributions, opens the possibility for reproductive hormone to
impact patterns of connectivity across time.
Relationship With Results From Previous Analyses of Same Data
Notably, other studies have analyzed the 28andMe dataset but obtained dissimilar results. For
instance, Pritschet et al. (2020) identified a strong link between the default mode network and
estradiol. In contrast, we find evidence linking a specific network state to gonadotropins, FSH
and LH. An important question is why these results diverge. There are a number of possible
explanations, although the two most obvious deal with the timescales at which networks are
modeled and characterized and the measure used to assess connectivity. Here, we analyze
networks estimated using edge time series, which resolves cofluctuations at a timescale of
single frames (Esfahlani et al., 2020). Previous studies, on the other hand, have focused on
either static connectivity (Pritschet et al., 2020) or time-varying estimates of connectivity using
sliding windows (Mueller et al., 2021). Although sliding windows resolve time-varying
estimates of connectivity, the windowing procedure necessarily induces “blurring,” making
it impossible to precisely temporally localize networks (Jo et al., 2021). Additionally, edge time
series estimate connectivity using a correlation-based metric, which can result in opposed
co-fluctuations (negative weights in a connectivity matrix), whereas previous studies used
wavelet-based coherence to estimate interregional connectivity, which is bound to the interval
Network Neuroscience
1192
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
[0, 1]. These observations underscore a more general trend in the network neuroscience
community, wherein the widespread use of bespoke processing and analysis tools make it
challenging to directly compare or reproduce findings. Future work must focus on standard-
izing these practices.
Future Directions
The results of our study present several opportunities for future investigations. First, while we
identify a putative link between high-amplitude co-fluctuations and female reproductive
hormones, its implications for cognitive and clinical processes have not been fully explored.
For instance, the studies from which the data originated also collected behavioral data on the
participant’s level of stress, sleep, and affect across the menstrual cycle. Future studies should
investigate the role of brain-hormone coupling in mediating behavioral effects.
To our knowledge, this study is the first to our knowledge to identify a significant relation-
ship between luteinizing hormone and daily changes in high-amplitude co-fluctuations in
brain connectivity across the menstrual cycle. Many studies have demonstrated luteinizing
hormone’s negative association with cognition, and increased luteinizing hormone in females
after menopause is associated with poorer cognition and a higher risk of dementia (Bhatta,
Blair, & Casadesus, 2018; Blair et al., 2016). Future studies should examine how luteinizing
hormone’s modulatory effects on cognition relate to daily changes in brain network organiza-
tion and connectivity in both premenopausal and postmenopausal females.
While the dense-sampling framework allows for detailed analyses of single individuals or
small cohorts of individuals (Braga & Buckner, 2017; Gordon et al., 2017; Laumann et al.,
2015), its design makes generalizing to larger and more variable populations challenging.
Future studies should analyze relationships between network organization, hormone con-
centrations, and cognition within subjects across many days of their cycle and between
subjects in different endocrine states. Variability in sex hormone production occurs across
the female life-span, starting at the onset of puberty and continuing across the reproductive
cycle, pregnancy, and the menopausal transition. Targeting these major neuroendocrine tran-
sition states could yield greater insight into sex steroid hormones’ influence on the brain’s
network architecture, mood, and cognition. Throughout the life course, changes in women’s
reproductive status (e.g., puberty, use of hormonal contraceptives, pregnancy/postpartum,
and perimenopause) have been associated with increased risk for mood disturbance includ-
ing major depressive disorder. Yet, the neurobiological pathways by which endocrine
changes give rise to depressive symptoms in some women, but not others, is unclear. A
more detailed understanding of how sex hormone fluctuations produce rapid changes in
brain network structure could provide a framework for understanding risk and resilience
to mood disorders.
An interesting question that could be addressed in future work concerns what properties of
the fMRI BOLD signal itself contribute to events. One likely contributor is the correlation struc-
ture of BOLD data. Several studies have shown that strong modular correlation structure
supports high-amplitude events (Ladwig et al., 2022; Novelli & Razi, 2022; Pope et al.,
2021). Another possibility is signal to noise ratio. In R. F. Betzel, Chumin, Esfahlani, Tanner,
and Faskowitz (2022), the authors studied event structure in different brain systems, for exam-
ple, default mode, visual, somatomotor, and so forth, including subcortical areas that have
poor signal-to-noise ratio relative to neocortex. The authors found that events occurred less
frequently in subcortical systems than cortex, supporting the hypothesis that overall signal
quality plays a role in event generation.
Network Neuroscience
1193
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
An important question that arises from this paper is whether having more or fewer events on
any given day (or even in a different dataset) is necessarily a “good” or “bad” thing. In general,
this question should be addressed through subsequent studies that explicitly seek to relate
event frequencies with other phenotypes and behavioral data. This, however, is precisely
the advantage of edge time series. They provide a temporal dimension absent in the analysis
of static FC through which brain network data can be studied. There are many trajectories—
time-varying patterns of activity and connectivity—that lead to the same correlation structure,
but are lost when we analyze static FC. Edge time series offers a window into the statistics of
time-varying brain data, and possibly exposing new features.
Limitations
Here, we analyzed fMRI data from two studies of a single individual. Although these two stud-
ies provide ample information about daily changes in brain activity and hormone levels, the
single-subject nature of the study limits the generalizability of our results. It is possible that the
relationships between community organization and hormone concentrations observed in this
study are a result of individual processes in the subject that are not present in most of the
population. Further deep-phenotyping studies with larger cohorts should be conducted to
determine the generalizability of the results across the general population. This, however,
belies a broader limitation. With more studies adopting a dense-sampling design, it becomes
necessary to assess how much data is necessary to assess an effect of a given size. While the
expectation is that having lots of data from a single brain will improve statistical power, the
magnitude of this improvement remains untested or, at the very least, not well documented.
Even basic questions concerning the typical size of effects in dense-sampling studies have not
been fully reported—for example, the extent to which fluctuations in hormone levels are
coupled with brain-based variables. Thus, power estimates for these types of studies are
lacking and should be the focus of future research.
A second limitation concerns the detection and characterization of edge time series and
high-amplitude co-fluctuations. Recent studies have suggested that some of their properties
can be anticipated from the static, that is, time-invariant, functional connectivity matrix alone
(Novelli & Razi, 2022), downplaying their interpretation as dynamic events. Determining the
features of higher order network constructs like edge time series and edge connectivity remains
an active area of research (R. F. Betzel, Cutts, et al., 2022; Cooper et al., 2021; Esfahlani et al.,
2022). Additional studies are necessary to address this and related open questions.
We note, however, that while our findings suggest a hormonal contribution to high-
amplitude events, other factors likely also play important roles, including the underlying
anatomical connectivity, whose network organization shapes the spatial topography of event
co-fluctuations (Pope et al., 2021). Future work should be directed to tease apart the contri-
butions of structural connectivity (Seider et al., 2021) as well as other factors, including those
molecular, vascular (Bright, Whittaker, Driver, & Murphy, 2020), and hemodynamic (Chang &
Glover, 2009).
An additional limitation concerns the dynamic interpretation of edge time series. Here, we
use edge time series to track frame-by-frame changes in network structure, detect high-
amplitude frames, and partition those frames into repeating clusters. In this way, we use edge
time series, not a means of studying brain dynamics per se, but to decompose FC and filter
time points. Indeed, there remains ongoing debate, both in the edge time series literature (see
Novelli & Razi, 2022), but also in the time-varying FC literature (see Laumann et al., 2017;
Liégeois et al., 2017) concerning the extent to which brain activity and connectivity are truly
Network Neuroscience
1194
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
dynamic and marked by distinct changes in underlying correlation structure. While edge time
series do not require the parameterizations that sliding windows do (these may make detecting
“true” dynamics difficult), it remains unclear whether the fluctuations observed in edge time
series are, indeed, evidence of such dynamics. This remains an important area for future
research (Lurie et al., 2020).
We note, however, there are some important differences between our study and those pre-
vious studies. In particular, our study focuses on rapid, that is, framewise, changes in co-
fluctuation patterns. Even in sliding-window analyses, access to this timescale is limited and
the networks resulting from each technique exhibit dissimilar topological profiles (Esfahlani
et al., 2022). In addition, our study focused on gonadotropic and ovarian hormones, and
focused not on distinct phases of the menstrual cycle or on peaks in serum levels, but rather
on day-to-day fluctuations in hormone levels.
Here, we focus on endogenous hormones only. However, exogenous hormones—
introduced through the oral contraceptive—could, in principle, also be related to network
connectivity and brain states. However, we note that the serum concentrations of the exoge-
nous hormones ethinyl estradiol and levonorgestrel are very low and, consistent with previous
studies (Mueller et al., 2021; Pritschet et al., 2020), were not examined here. Future studies
should investigate the interplay of exogenous and endogenous hormones and their contribu-
tions to time-varying connectivity.
Another potential concern is related to pooling data from across two experiments. In prin-
ciple, this decision was made to enhance statistical power—it doubles the number of samples.
However, the data from each experiment was collected under different conditions (with versus
without oral contraceptive), raising concerns that the datasets are too dissimilar to warrant
aggregating. As a safeguard against the possibility that our results are biased by the pooling
procedure, we performed a number of supplementary analyses. Most importantly, we verified
that the main result—an association between events and FSH/LH concentrations—reproduces
when we analyzed either experimental dataset in isolation, that is, without pooling. A partial
explanation for this replication is that neither FSH nor LH exhibited significant differences in
mean concentration between experiments (see Supporting Information Figure S5). Another
explanation for the replication is that, rather than using product-moment correlations to link
events and hormones—a measure that is sensitive to amplitude changes and can be driven by
outliers or spikes in data—we used rank correlations which transforms data to an ordinal scale
before calculating correlations. Collectively, these and other efforts help reduce the likelihood
that the reported results are driven by differences in experimental conditions.
In general, we find evidence that hormones fluctuate from day to day and that the structure
of these fluctuations are preserved, irrespective of whether or not the subject took an oral con-
traceptive (see Supporting Information Figure S20). Though the causes of the fluctuations are
not always clear, they likely include lifestyle factors—for example, diet, sleep, and so
forth—and in the case of experiment 2, the effects of an oral contraceptive. Consider a “ground
truth” where the number of events observed on any scan session was proportional to hormone
concentration plus some random noise. Were this the case, then we would expect that any
fluctuation in hormones, irrespective of the cause, would yield a proportional change in the
number of events. Indeed, we find evidence that this is the case and that, at least for FSH and
LH, the relationship between events and hormone concentration is not interrupted by oral
contraceptives. However, they cannot conclusively rule out the possibility that there exist
experiment-dependent relationships between brain networks and hormone concentrations
that would appear with additional data from the same individual or by expanding the scope
Network Neuroscience
1195
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
of data collection to include more individuals. These two directions represent potentially fruit-
ful follow-ups to the present study.
A final limitation concerns our interpretation of endocrine effects on recorded brain activity
and connectivity. Here, we find that hormone concentrations are associated with high-
amplitude co-fluctuations that have been shown to shape whole-brain patterns of functional
connectivity (R. F. Betzel, Cutts, et al., 2022; Esfahlani et al., 2020, 2022; Pope et al., 2021;
Sporns et al., 2021). However, hormones also impact brain vasculature (Krause, Duckles, &
Pelligrino, 2006). Because the fMRI BOLD signal is an indirect measure of brain activity that
depends critically on neurovascular coupling, an alternative explanation is that true effect of
gonadotropins is on brain vasculature, which modulates the hemodynamic response. Future,
and more targeted, studies should aim to disentangle these effects.
Conclusions
In conclusion, our study posits a link between high-amplitude, network-level co-fluctuations
and the human endocrine system. Specifically, we report an association between the
frequency of dynamic network states and variation in luteinizing and follicle-stimulating
hormones. Our work addresses questions concerning the factors contributing to high-
amplitude co-fluctuations while opening up new opportunities for future studies.
MATERIALS AND METHODS
Datasets
Neuroimaging and endocrine data comes from a single subject (author L.P.) scanned over a
course of 30 days, on two separate occasions (Study 1 and Study 2). The subject had no history
of neuropsychiatric diagnosis, endocrine disorders, or prior head trauma and no history of
smoking. She had a history of regular menstrual cycles (no missed periods, cycle occurring
every 26–28 days). In the 12 months prior to the first 30-day data collection period, the subject
was free from hormone-based medication. In the second study, the participant was on a
hormone regimen (0.02 mg ethinyl-estradiol, 0.1 mg levonorgestrel, Aubra, Afaxys Pharma-
ceuticals), which she began 10 months prior to the start of data collection. The pharmacolog-
ical regimen used in Study 2 chronically and selectively suppressed progesterone while
leaving estradiol dynamics largely indistinguishable from Study 1. The participant gave written
informed consent and the study was approved by the University of California, Santa Barbara,
Human Subjects Committee.
Both studies consisted of similar design and included components that were analyzed here.
Test sessions began with a daily questionnaire (assessments of stress, sleep, anxiety, and
affective state), followed by a spatial navigation task (not reported here). Time-locked collec-
tion of serum and whole blood started each day at 10:00 am in Study 1 and 11:00 am in Study
2 (±30 min). Endocrine samples were collected, at minimum, after 2 hours of no food or drink
consumption (excluding water). Additionally, the participant abstained from caffeinated
beverages prior to test sessions. Following behavioral assessment, the participant underwent
a 1-hour MRI scan session that included the acquisition of structural and functional images.
Neuroimaging data was collected on a Siemens 3T Prisma scanner with a 64-channel
phased-array head coil. Scans were collected around the same time each day (11:00 am local
time). For each scanning session, a high-resolution T1-weighted anatomical sequence
(MPRAGE) was acquired (TR = 2,500 ms, TE = 2.31 ms, T1 = 934 ms, flip angle = 7°). Follow-
ing this, a 10-minute resting-state sequence was acquired, using a T2*-weighted multiband
echo-planar (EPI) sequence was acquired (72 oblique slices, TR = 720 ms, TE = 37 ms, voxel
Network Neuroscience
1196
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
size 2 mm3, flip angle = 52°, multiband factor = 8). To mitigate against motion, a custom
3D-printed foam head case was employed for days 8–30 of the first 30-day period, and for
days 1–30 of the second 30-day period.
Endocrine Procedures
A licensed phlebotomist inserted a saline-lock intravenous line into the dominant or nondom-
inant hand or forearm daily to evaluate hypothalamic-pituitary-gonadal axis hormones,
including serum levels of gonadal hormones (17β-estradiol, progesterone, and testosterone)
and the pituitary gonadotropins luteinizing hormone (LH) and follicle stimulating hormone
(FSH). One 10-cc-mL blood sample was collected in vacutainer SST (BD Diagnostic Systems)
each session. The sample clotted at room temperature for 45 minutes until centrifugation
(2,000 × g for 10 minutes) and were then aliquoted into three 1 mL microtubes. Serum samples
were stored at −20° until assayed. Serum concentrations were determined via liquid chroma-
tography mass spectrometry (for all steroid hormones) and immunoassay (for all gonadotro-
pins) at the Brigham and Women’s Hospital Research Assay Core. Assay sensitivities, dynamic
range, and intra-assay coefficients of variation (respectively) were as follows: estradiol 1 pg/mL,
1–500 pg/mL, <5% relative standard deviation (RSD), 0.05 ng/mL, 0.05–10 ng/mL, 9.33%
RSD; testosterone, 1.0 ng/dL 1–2,000 ng/dL, <4% RSD. FSH and LH levels were determined
via chemiluminescent assay (Beckman Coulter). The assay sensitivity, dynamic range, and
intra-assay coefficient of variation were as follows: FSH, 0.2 mlU/mL, 0.2–200 mIU/mL,
3.1–4.3%; LH, 0.2 mIU/mL, 0.2–250 mIU/mL, 4.3–6.4%. Importantly, we note that LC-MS
assessments of exogenous hormone concentrations (attributable to the hormone regime itself )
showed that serum concentrations of ethinyl estradiol were very low (M = 0.01 ng/mL; range
0.001–0.016 ng/mL) and below 1.5 ng/mL for levonorgestrel (M = 0.91 ng/mL; range = 0.03–
1.45 ng/mL). This ensures that the brain hormone associations reported in Study 2 are still due
to endogenous estradiol action.
Preprocessing was performed using fMRIPrep (20.2.0) (Esteban et al., 2018), which is based
on Nipype 1.5.1 (Gorgolewski et al., 2011). The following description of fMRIPrep’s prepro-
cessing is based on boilerplate distributed with the software covered by a ‘no rights reserved’
(CCO) license. Internal operations of fMRIPrep use Nilearn 0.6.2 (Abraham et al., 2014),
mostly within the functional processing workflow. The preprocessing pipelines employ func-
tions from ANTs (2.3.3), FreeSurfer (6.0.1), FSL (5.0.9), and AFNI (20160207).
T1-weighted (T1w) images were corrected for intensity nonuniformity (INU) with
N4BiasFieldCorrection (Tustison et al., 2010), distributed with ANTs (Avants, Epstein,
Grossman, & Gee, 2008), and used as T1w-reference throughout the workflow. The
T1w-reference was then skull-stripped with a Nipype implementation of the antsBrain-
Extraction.sh workflow, using NKI as target template. Brain tissue segmentation of
cerebrospinal fluid (CSF), white matter ( WM), and gray matter (GM) was performed on the
brain-extracted T1w using fast (Zhang, Brady, & Smith, 2001). Brain surfaces were recon-
structed using recon-all (Dale, Fischl, & Sereno, 1999), and the brain mask estimated pre-
viously was refined with a custom variation of the method to reconcile ANTs-derived and
FreeSurfer-derived segmentations of the cortical gray matter of Mindboggle (Klein et al.,
2017). Volume-based spatial normalization to one standard space (MNI152NLin2009cAsym)
was performed through nonlinear registration with antsRegistration, using brain-
extracted versions of both T1w reference and the T1w template. The following template
was selected for spatial normalization: ICBM 152 Nonlinear Asymmetrical template version
2009c (Fonov, Evans, McKinstry, Almli, & Collins, 2009). For each of the BOLD runs found
Network Neuroscience
1197
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
per subject (across all tasks and sessions), the following preprocessing was performed. First, a
reference volume and its skull-stripped version were generated using a custom methodology of
fMRIPrep. Susceptibility distortion correction (SDC) was omitted. The BOLD reference was
then co-registered to the T1w reference using bbregister (FreeSurfer), which implements
boundary-based registration (Greve & Fischl, 2009). Co-registration was configured with six
degrees of freedom. Head-motion parameters with respect to the BOLD reference (transforma-
tion matrices and six corresponding rotation and translation parameters) are estimated before
any spatiotemporal filtering using mcflirt (Jenkinson, Bannister, Brady, & Smith, 2002).
BOLD runs were slice-time corrected using 3dTshift from AFNI (Cox & Hyde, 1997).
The BOLD time series (including slice timing correction when applied) were resampled onto
their original, native space by applying the transforms to correct for head motion. These
resampled BOLD time series will be referred to as preprocessed BOLD in original space, or
just preprocessed BOLD. The BOLD time series were resampled into standard space, generat-
ing a preprocessed BOLD run in MNI152NLin2009cAsym space. First, a reference volume and
its skull-stripped version were generated using a custom methodology of fMRIPrep. Several
confounding time series were calculated based on the preprocessed BOLD: framewise dis-
placement (FD), DVARS and three region-wise global signals. FD was computed using two
formulations following Power (absolute sum of relative motions; Power et al., 2014) and
Jenkinson (relative root mean square displacement between affines; Jenkinson et al., 2002).
FD and DVARS are calculated for each functional run, both using their implementations in
Nipype following the definitions by Power et al. (2014). The three global signals are extracted
within the CSF, the WM, and the whole-brain masks. Additionally, a set of physiological
regressors were extracted to allow for component-based noise correction (Behzadi, Restom,
Liau, & Liu, 2007). Principal components are estimated after high-pass filtering the prepro-
cessed BOLD time series (using a discrete cosine filter with 128-s cutoff ) for the two CompCor
variants: temporal (tCompCor) and anatomical (aCompCor). tCompCor components are then
calculated from the top 2% variable voxels within the brain mask. For aCompCor, three prob-
abilistic masks (CSF, WM, and combined CSF + WM) are generated in anatomical space. The
implementation differs from that of Behzadi et al. in that instead of eroding the masks by two
pixels on BOLD space, the aCompCor masks are subtracted a mask of pixels that likely contain
a volume fraction of GM. This mask is obtained by dilating a GM mask extracted from the
FreeSurfer’s aseg segmentation, and it ensures components are not extracted from voxels con-
taining a minimal fraction of GM. Finally, these masks are resampled into BOLD space and
binarized by thresholding at 0.99 (as in the original implementation). Components are also
calculated separately within the WM and CSF masks. For each CompCor decomposition,
the k components with the largest singular values are retained, such that the retained compo-
nents’ time series are sufficient to explain 50 percent of variance across the nuisance mask
(CSF, WM, combined, or temporal). The remaining components are dropped from consider-
ation. The head-motion estimates calculated in the correction step were also placed within the
corresponding confounds file. The confound time series derived from head motion estimates
and global signals were expanded with the inclusion of temporal derivatives and quadratic
terms for each (Satterthwaite et al., 2013). Frames that exceeded a threshold of 0.5 mm FD
or 1.5 standardized DVARS were annotated as motion outliers. All resamplings can be per-
formed with a single interpolation step by composing all the pertinent transformations (i.e.,
head-motion transform matrices, susceptibility distortion correction when available, and
co-registrations to anatomical and output spaces). Gridded (volumetric) resamplings were
performed using antsApplyTransforms, configured with Lanczos interpolation to mini-
mize the smoothing effects of other kernels (Lanczos, 1964). Nongridded (surface)
resamplings were performed using mri_vol2surf (FreeSurfer).
Network Neuroscience
1198
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
The Schaefer parcellation (Schaefer et al., 2018) was used to delineate 400 regions on the
cortical surface. To transfer the parcellation from fsaverage to subject space, FreeSurfer’s
mris_ca_label function was used in conjunction with a pretrained Gaussian classifier sur-
face atlas (Fischl et al., 2004) to register cortical surfaces based on individual curvature and
sulcal patterns. The result is a volumetric parcellation rendered in subject anatomical space
that follows the cortical ribbon estimated via FreeSurfer’s recon-all process.
Each preprocessed BOLD image was linearly detrended, band-pass filtered (0.008–
0.08 Hz), confound regressed and standardized using Nilearn’s signal.clean function,
which removes confounds orthogonally to the temporal filters. The confound regression
strategy included six motion estimates, mean signal from a white matter, cerebrospinal fluid,
and whole brain mask, derivatives of these previous nine regressors, and squares of these
18 terms (Satterthwaite et al., 2013). Spike regressors were not applied. The 36 parameter
strategy (with and without spike regression) has been show to be a relatively effective option
to reduce motion-related artifacts (Parkes, Fulcher, Yücel, & Fornito, 2018). An alternative pre-
processesing strategy was also employed to evaluate the stability of the findings. This strategy
included six motion estimates, derivatives of these previous six regressors, and squares of
these 12 terms, in addition to five anatomical CompCor components (Behzadi et al., 2007).
Following these preprocessing operations, the mean signal was taken at each node in volu-
metric anatomical space.
Edge Time Series
Let zi = [zi (1), … zi (T )] be the z-scored time series for region i. Most network neuroscience
analyses define the functional connectivity between pairs of regions {i, j } as the Pearson
correlation of their activity, that is, rij = 1
T −1
t zi (t ) · zj (t ).
P
Recently, we proposed a “temporal unwrapping” of functional connection weights by
omitting the summation. That is, we calculate the instantaneous magnitude and sign of
co-fluctuation between pairs of brain regions as eij (t ) = zi (t ) · zj (t ) (R. F. Betzel, Cutts, et al.,
2022; Esfahlani et al., 2020; Faskowitz et al., 2020). This approach has the benefit of resolving
changes in pairwise interactions at a temporal resolution of single frames. It also is deeply
related to the Pearson correlation and functional connectivity—the temporal average of a
region pairs’ edge time series is exactly equal to its connection weight. In this way, edge time
series can be viewed as precise temporal decompositions of functional connectivity. In this
study, we calculated edge time series for every pair of cortical and subcortical regions for every
scan session.
Event Detection
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The brainwide level of network activity at any instant can be summarized as the root sum of
square over all edges co-fluctuations:
X
RSS tð Þ ¼
eij tð Þ:
(1)
i;j≠i
In previous studies we demonstrated that RSS exhibits “bursty” behavior, such that most time
points express low-amplitude co-fluctuations while a relatively small number exhibit large
RSS values (R. F. Betzel, Cutts, et al., 2022). These “events” are thought to reflect underlying
anatomical connectivity (Pope et al., 2021), are highly individualized (R. F. Betzel, Cutts, et al.,
2022), and contribute disproportionately to the time-averaged pattern of functional
connectivity (Esfahlani et al., 2020; Sporns et al., 2021).
Network Neuroscience
1199
Network dynamics are associated with reproductive hormones
Recently, we proposed a simple statistical test for detecting events. This test works by com-
paring observed RSS values with those generated under a temporal null model in which
regional time series are circularly shifted by a random offset in either direction. This procedure
generates a null distribution of RSS values against which an empirical value can be compared
statistically. For each frame we estimated a nonparametric p value by counting the fraction of
null RSS values that exceeded the observed value. We compared p values against an adjusted
critical value while fixing the false discovery rate at q = 0.05 (Benjamini & Hochberg, 1995).
Note that this null model necessarily destroys the correlation structure (functional connec-
tivity) and can be viewed as a principled method for selecting a threshold for events.
Once a statistical threshold for events is determined, we identify temporal contiguous
sequences of suprathreshold frames, which we refer to as event segments. We discard any such
segments that include a high-motion frame or are within two frames of a high-motion frame
and, from those segments, extract as a representative co-fluctuation pattern the frame corre-
sponding to the peak RSS. We repeat this procedure for all scans, retaining the peak co-
fluctuation patterns.
Cluster Definition
In previous studies, we demonstrated that high-amplitude co-fluctuation patterns can be clus-
tered into a series of states (R. F. Betzel, Cutts, et al., 2022). To do this, we compute the sim-
ilarity (correlation) between all pairs co-fluctuation patterns extracted during event segments.
This results in a pattern × pattern matrix, which we submitted to a generalized version of the
Louvain algorithm (Blondel, Guillaume, Lambiotte, & Lefebvre, 2008; Jutla, Jeub, & Mucha,
2011) for modularity maximization algorithm (Newman & Girvan, 2004). Modularity maximi-
zation is a computational heuristic for detecting community structure in networked data. It
defines communities (clusters) as groups elements whose internal density of connections
maximally exceed what would be expected. In this context, we defined the expected weight
of connections to be equal to the mean similarity between all pairs of patterns.
Modularity maximization with the Louvain algorithm is nondeterministic and, depending
upon initial conditions, can yield dissimilar results. Accordingly, we ran the algorithm 1,000
times with different random seeds. We resolved variability across these different seeds using a
consensus clustering algorithm in which we iteratively clustered the module co-assignment
matrix until convergence (see any of R. F. Betzel, Cutts, et al., 2022; R. F. Betzel, 2020;
R. F. Betzel et al., 2017; for details of this algorithm). The resulting consensus partition assigned
each co-fluctuation pattern to nonoverlapping clusters.
ACKNOWLEDGMENTS
We thank Haily Merritt and Jacob Tanner for providing feedback on an early version of this
manuscript.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00307.
AUTHOR CONTRIBUTIONS
Sarah Greenwell: Formal analysis; Investigation; Methodology; Software; Visualization;
Writing – original draft; Writing – review & editing. Joshua Faskowitz: Data curation;
Network Neuroscience
1200
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
Resources; Software; Writing – review & editing. Laura Pritschet: Conceptualization; Data
curation; Methodology; Resources; Writing – review & editing. Tyler Santander: Conceptual-
ization; Data curation; Methodology; Resources; Writing – review & editing. Emily Jacobs:
Conceptualization; Data curation; Methodology; Resources; Writing – review & editing.
Richard Betzel: Conceptualization; Data curation; Investigation; Methodology; Project
administration; Resources; Software; Supervision; Validation; Visualization; Writing – original
draft; Writing – review & editing.
FUNDING INFORMATION
Richard Betzel, National Science Foundation (https://dx.doi.org/10.13039/501100008982),
Award ID: 2023985.
REFERENCES
Abraham, A., Pedregosa, F., Eickenberg, M., Gervais, P., Mueller,
A., Kossaifi, J., … Varoquaux, G. (2014). Machine learning for
neuroimaging with scikit-learn. Frontiers in Neuroinformatics,
8, 14. https://doi.org/10.3389/fninf.2014.00014, PubMed:
24600388
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T., &
Calhoun, V. D. (2014). Tracking whole-brain connectivity
dynamics in the resting state. Cerebral Cortex, 24(3), 663–676.
https://doi.org/10.1093/cercor/bhs352, PubMed: 23146964
Arélin, K., Mueller, K., Barth, C., Rekkas, P. V., Kratzsch, J.,
Burmann, I., … Sacher, J. (2015). Progesterone mediates brain
functional connectivity changes during the menstrual cycle—A
pilot resting state MRI study. Frontiers in Neuroscience, 9, 44.
https://doi.org/10.3389/fnins.2015.00044, PubMed: 25755630
Avants, B., Epstein, C., Grossman, M., & Gee, J. (2008). Symmetric
diffeomorphic image registration with cross-correlation: Evaluat-
ing automated labeling of elderly and neurodegenerative brain.
Medical Image Analysis, 12(1), 26–41. https://doi.org/10.1016/j
.media.2007.06.004, PubMed: 17659998
Barttfeld, P., Uhrig, L., Sitt, J. D., Sigman, M., Jarraya, B., &
Dehaene, S. (2015). Signature of consciousness in the dynamics
of resting-state brain activity. Proceedings of the National Acad-
emy of Sciences, 112(3), 887–892. https://doi.org/10.1073/pnas
.1418031112, PubMed: 25561541
Bassett, D. S., & Bullmore, E. (2006). Small-world brain networks.
The Neuroscientist, 12(6), 512–523. https://doi.org/10.1177
/1073858406293182, PubMed: 17079517
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. NeuroImage, 37(1), 90–101. https://doi.org/10
.1016/j.neuroimage.2007.04.042, PubMed: 17560126
Beltz, A. M., & Moser, J. S. (2020). Ovarian hormones: A long
overlooked but critical contributor to cognitive brain structures
and function. Annals of the New York Academy of Sciences,
1464(1), 156–180. https://doi.org/10.1111/nyas.14255,
PubMed: 31639230
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society: Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Berman, K. F., Schmidt, P. J., Rubinow, D. R., Danaceau, M. A., Van
Horn, J. D., Esposito, G., … Weinberger, D. R. (1997). Modula-
tion of cognition-specific cortical activity by gonadal steroids: A
positron-emission tomography study in women. Proceedings of
the National Academy of Sciences, 94(16), 8836–8841. https://
doi.org/10.1073/pnas.94.16.8836, PubMed: 9238064
Betzel, R. F. (2020). Organizing principles of whole-brain functional
connectivity in zebrafish larvae. Network Neuroscience, 4(1),
234–256. https://doi.org/10.1162/netn_a_00121, PubMed: 32166210
Betzel, R. F., Chumin, E. J., Esfahlani, F. Z., Tanner, J., & Faskowitz,
J. (2022). System-level high-amplitude co-fluctuations. bioRxiv.
https://doi.org/10.1101/2022.07.26.501262
Betzel, R. F., Cutts, S. A., Greenwell, S., Faskowitz, J., & Sporns, O.
(2022). Individualized event structure drives individual differ-
ences in whole-brain functional connectivity. NeuroImage, 252,
118993. https://doi.org/10.1016/j.neuroimage.2022.118993,
PubMed: 35192942
Betzel, R. F., Fukushima, M., He, Y., Zuo, X.-N., & Sporns, O.
(2016). Dynamic fluctuations coincide with periods of high
and low modularity in resting-state functional brain networks.
NeuroImage, 127, 287–297. https://doi.org/10.1016/j
.neuroimage.2015.12.001, PubMed: 26687667
Betzel, R. F., Medaglia, J. D., Papadopoulos, L., Baum, G. L., Gur,
R., Gur, R., … Bassett, D. S. (2017). The modular organization of
human anatomical brain networks: Accounting for the cost of
wiring. Network Neuroscience, 1(1), 42–68. https://doi.org/10
.1162/NETN_a_00002, PubMed: 30793069
Bhatta, S., Blair, J. A., & Casadesus, G. (2018). Luteinizing hormone
involvement in aging female cognition: Not all is estrogen loss.
Frontiers in Endocrinology, 9, 544. https://doi.org/10.3389/fendo
.2018.00544, PubMed: 30319538
Blair, J. A., Palm, R., Chang, J., McGee, H., Zhu, X., Wang, X., &
Casadesus, G. (2016). Luteinizing hormone downregulation but
not estrogen replacement improves ovariectomy-associated cog-
nition and spine density loss independently of treatment onset
timing. Hormones and Behavior, 78, 60–66. https://doi.org/10
.1016/j.yhbeh.2015.10.013, PubMed: 26497249
Network Neuroscience
1201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
n
e
n
_
a
_
0
0
3
0
7
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network dynamics are associated with reproductive hormones
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks. Journal
of Statistical Mechanics: Theory and Experiment, 2008(10),
P10008. https://doi.org/10.1088/1742-5468/2008/10/P10008
Braga, R. M., & Buckner, R. L. (2017). Parallel interdigitated distrib-
uted networks within the individual estimated by intrinsic func-
tional connectivity. Neuron, 95(2), 457–471. https://doi.org/10
.1016/j.neuron.2017.06.038, PubMed: 28728026
Bright, M. G., Whittaker, J. R., Driver, I. D., & Murphy, K. (2020).
Vascular physiology drives functional brain networks. Neuro-
Image, 217, 116907. https://doi.org/10.1016/j.neuroimage.2020
.116907, PubMed: 32387624
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186–198. https://doi.org/10.1038
/nrn2575, PubMed: 19190637
Bullmore, E., & Sporns, O. (2012). The economy of brain network
organization. Nature Reviews Neuroscience, 13(5), 336–349.
https://doi.org/10.1038/nrn3214, PubMed: 22498897
Chang, C., & Glover, G. H. (2009). Relationship between respira-
tion, end-tidal CO2, and BOLD signals in resting-state fMRI.
NeuroImage, 47(4), 1381–1393. https://doi.org/10.1016/j
.neuroimage.2009.04.048, PubMed: 19393322
Christin-Maitre, S. (2013). History of oral contraceptive drugs and
their use worldwide. Best Practice & Research Clinical Endocri-
nology & Metabolism, 27(1), 3–12. https://doi.org/10.1016/j
.beem.2012.11.004, PubMed: 23384741
Conio, B., Martino, M., Magioncalda, P., Escelsior, A., Inglese, M.,
Amore, M., & Northoff, G. (2020). Opposite effects of dopamine
and serotonin on resting-state networks: Review and implications
for psychiatric disorders. Molecular Psychiatry, 25(1), 82–93.
https://doi.org/10.1038/s41380-019-0406-4 , PubMed:
30953003
Cooper, R. A., Kurkela, K. A., Davis, S. W., & Ritchey, M. (2021).
Mapping the organization and dynamics of the posterior medial
network during movie watching. NeuroImage, 236, 118075.
https://doi.org/10.1016/j.neuroimage.2021.118075, PubMed:
33910099
Cox, R. W., & Hyde, J. S. (1997). Software tools for analysis and
visualization of fMRI data. NMR in Biomedicine, 10(4–5),
171–178. https://doi.org/10.1002/(SICI )1099-1492(199706
/08)10:4/5<171::AID-NBM453>3.0.CO;2-l, PubMed: 9430344
Dale, UN. M., Fischl, B., & Sereno, M. IO. (1999). Cortical
surface-based analysis: IO. Segmentation and surface reconstruc-
zione. NeuroImage, 9(2), 179–194. https://doi.org/10.1006/nimg
.1998.0395, PubMed: 9931268
Dan, R., Reuveni, I., Canetti, L., Weinstock, M., Segman, R.,
Goelman, G., & Bonne, O. (2020). Trait-related changes in brain
network topology in premenstrual dysphoric disorder. Hormones
and Behavior, 124, 104782. https://doi.org/10.1016/j.yhbeh
.2020.104782, PubMed: 32470339
Dang, l. C., O’Neil, J. P., & Jagust, W. J. (2012). Dopamine supports
coupling of attention-related networks. Journal of Neuroscience,
32(28), 9582–9587. https://doi.org/10.1523/ JNEUROSCI.0909
-12.2012, PubMed: 22787044
Dubol, M., Epperson, C. N., Sacher, J., Pletzer, B., Derntl, B.,
Lanzenberger, R., … Comasco, E. (2021). Neuroimaging the
menstrual cycle: A multimodal systematic review. Frontiers in
Neuroendocrinology, 60, 100878. https://doi.org/10.1016/j
.yfrne.2020.100878, PubMed: 33098847
Esfahlani, F. Z., Byrge, L., Tanner, J., Sporns, O., Kennedy, D. P., &
Betzel, R. F. (2022). Edge-centric analysis of time-varying func-
tional brain networks with applications in autism spectrum disor-
der. NeuroImage, 263, 119591. https://doi.org/10.1016/j
.neuroimage.2022.119591, PubMed: 36031181
Esfahlani, F. Z., Jo, Y., Faskowitz, J., Byrge, L., Kennedy, D. P.,
(2020). High-amplitude cofluc-
Sporns, O., & Betzel, R. F.
tuations in cortical activity drive functional connectivity. Pro-
ceedings of the National Academy of Sciences, 117(45),
28393–28401. https://doi.org/10.1073/pnas.2005531117,
PubMed: 33093200
Esteban, O., Markiewicz, C. J., Blair, R. W., Moodie, C. A., Isik,
UN. I., Erramuzpe, A., … Gorgolewski, K. J. (2018). fMRIPrep: UN
robust preprocessing pipeline for functional MRI. Nature
Methods, 16(1), 111–116. https://doi.org/10.1038/s41592-018
-0235-4, PubMed: 30532080
Faskowitz, J., Esfahlani, F. Z., Jo, Y., Sporns, O., & Betzel, R. F.
(2020). Edge-centric functional network representations of
human cerebral cortex reveal overlapping system-level architec-
ture. Nature Neuroscience, 23(12), 1644–1654. https://doi.org/10
.1038/s41593-020-00719-y, PubMed: 33077948
Fischl, B., van der Kouwe, A., Destrieux, C., Halgren, E., Ségonne,
F., Salat, D. H., … Dale, UN. M. (2004). Automatically parcellating
the human cerebral cortex. Cerebral Cortex, 14(1), 11–22. https://
doi.org/10.1093/cercor/bhg087, PubMed: 14654453
Fitzgerald, M., Pritschet, L., Santander, T., Grafton, S. T., & Jacobs,
E. G. (2020). Cerebellar network organization across the human
menstrual cycle. Scientific Reports, 10(1), 20732. https://doi.org
/10.1038/s41598-020-77779-4, PubMed: 33244032
Fonov, V., Evans, A., McKinstry, R., Almli, C., & Collins, D. (2009).
Unbiased nonlinear average age-appropriate brain templates
from birth to adulthood. NeuroImage, 47(Suppl. 1), S102.
https://doi.org/10.1016/S1053-8119(09)70884-5
Gordon, E. M., Laumann, T. O., Gilmore, UN. W., Newbold, D. J.,
Greene, D. J., Berg, J. J., … Dosenbach, N. U. F. (2017). Precision
functional mapping of individual human brains. Neuron, 95(4),
791–807. https://doi.org/10.1016/j.neuron.2017.07.011,
PubMed: 28757305
Gorgolewski, K., Burns, C. D., Madison, C., Clark, D., Halchenko,
Y. O., Waskom, M. L., & Ghosh, S. S. (2011). Nipype: A flexible,
lightweight and extensible neuroimaging data processing frame-
work in Python. Frontiers in Neuroinformatics, 5, 13. https://doi
.org/10.3389/fninf.2011.00013, PubMed: 21897815
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain image
alignment using boundary-based registration. NeuroImage, 48(1),
63–72. https://doi.org/10.1016/j.neuroimage.2009.06.060,
PubMed: 19573611
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey,
C. J., Wedeen, V. J., & Sporns, O. (2008). Mapping the struc-
tural core of human cerebral cortex. PLoS Biology, 6(7), e159.
https://doi.org/10.1371/journal.pbio.0060159, PubMed:
18597554
Hermundstad, UN. M., Bassett, D. S., Brown, K. S., Aminoff, E. M.,
Clewett, D., Freeman, S., … Carlson, J. M. (2013). Structural
foundations of resting-state and task-based functional connec-
tivity in the human brain. Proceedings of the National Academy
Network Neuroscience
1202
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
.
/
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network dynamics are associated with reproductive hormones
of Sciences, 110(15), 6169–6174. https://doi.org/10.1073/pnas
.1219562110, PubMed: 23530246
Hidalgo-Lopez, E., Mueller, K., Harris, T., Aichhorn, M., Sacher, J.,
& Pletzer, B. (2020). Human menstrual cycle variation in subcor-
tical functional brain connectivity: A multimodal analysis
approach. Brain Structure and Function, 225(2), 591–605.
https://doi.org/10.1007/s00429-019-02019-z, PubMed:
31894405
Hidalgo-Lopez, E., Zeidman, P., Harris, T., Razi, A., & Pletzer, B.
(2021). Spectral dynamic causal modelling in healthy women
reveals brain connectivity changes along the menstrual cycle.
Communications Biology, 4(1), 954. https://doi.org/10.1038
/s42003-021-02447-w, PubMed: 34376799
Hindriks, R., Adhikari, M. H., Murayama, Y., Ganzetti, M., Mantini,
D., Logothetis, N. K., & Deco, G. (2016). Can sliding-window
correlations reveal dynamic functional connectivity in
resting-state fMRI? NeuroImage, 127, 242–256. https://doi.org
/10.1016/j.neuroimage.2015.11.055, PubMed: 26631813
Jacobs, E., & D’Esposito, M. (2011). Estrogen shapes dopamine-
dependent cognitive processes: Implications for women’s health.
Journal of Neuroscience, 31(14), 5286–5293. https://doi.org/10
.1523/JNEUROSCI.6394-10.2011, PubMed: 21471363
Jenkinson, M., Bannister, P., Brady, M., & Smith, S. (2002).
Improved optimization for the robust and accurate linear registra-
tion and motion correction of brain images. NeuroImage, 17(2),
825–841. https://doi.org/10.1006/nimg.2002.1132, PubMed:
12377157
Jo, Y., Esfahlani, F. Z., Faskowitz, J., Chumin, E. J., Sporns, O., &
Betzel, R. F. (2021). The diversity and multiplexity of edge com-
munities within and between brain systems. Cell Reports, 37(7),
110032. https://doi.org/10.1016/j.celrep.2021.110032, PubMed:
34788617
Jutla, IO. S., Jeub, l. G., & Mucha, P. J. (2011). A generalized Louvain
method for community detection implemented in MATLAB.
https://netwiki.amath.unc.edu/GenLouvain
Klein, A., Ghosh, S. S., Bao, F. S., Giard, J., Häme, Y., Stavsky, E., …
Keshavan, UN. (2017). Mindboggling morphometry of human
brains. PLoS Computational Biology, 13(2), e1005350. https://
doi.org/10.1371/journal.pcbi.1005350, PubMed: 28231282
Krause, D. N., Duckles, S. P., & Pelligrino, D. UN. (2006). Influence
of sex steroid hormones on cerebrovascular function. Journal of
Applied Physiology, 101(4), 1252–1261. https://doi.org/10.1152
/japplphysiol.01095.2005, PubMed: 16794020
Ladwig, Z., Seitzman, B. A., Dworetsky, A., Yu, Y., Adeyemo, B.,
Smith, D. M., … Gratton, C. (2022). BOLD cofluctuation ‘events’
are predicted from static functional connectivity. NeuroImage,
260, 119476. https://doi.org/10.1016/j.neuroimage.2022
.119476, PubMed: 35842100
Lanczos, C. (1964). Evaluation of noisy data. Journal of the Society
for Industrial and Applied Mathematics, Series B: Numerical
Analysis, 1(1), 76–85. https://doi.org/10.1137/0701007
Laumann, T. O., Gordon, E. M., Adeyemo, B., Snyder, UN. Z., Joo,
S. J., Chen, M.-Y., … Petersen, S. E. (2015). Functional system and
areal organization of a highly sampled individual human brain.
Neuron, 87(3), 657–670. https://doi.org/10.1016/j.neuron.2015
.06.037, PubMed: 26212711
Laumann, T. O., Snyder, UN. Z., Mitra, A., Gordon, E. M., Gratton,
C., Adeyemo, B., … Petersen, S. E. (2017). On the stability of
BOLD fMRI correlations. Cerebral Cortex, 27(10), 4719–4732.
https://doi.org/10.1093/cercor/bhw265, PubMed: 27591147
Leonardi, N., & Van De Ville, D. (2015). On spurious and real
fluctuations of dynamic functional connectivity during rest.
NeuroImage, 104, 430–436. https://doi.org/10.1016/j
.neuroimage.2014.09.007, PubMed: 25234118
Li, M., Han, Y., Aburn, M. J., Breakspear, M., Poldrack, R. A., Shine,
J. M., & Lizier, J. T. (2019). Transitions in information processing
dynamics at the whole-brain network level are driven by alter-
ations in neural gain. PLoS Computational Biology, 15(10),
e1006957. https://doi.org/10.1371/journal.pcbi.1006957,
PubMed: 31613882
Liégeois, R., Laumann, T. O., Snyder, UN. Z., Zhou, J., & Sì, B. T. T.
(2017). Interpreting temporal fluctuations in resting-state func-
tional connectivity MRI. NeuroImage, 163, 437–455. https://doi
.org/10.1016/j.neuroimage.2017.09.012, PubMed: 28916180
Liparoti, M., Troisi Lopez, E., Sarno, L., Rucco, R., Minino, R.,
Pesoli, M., … Sorrentino, P. (2021). Functional brain network
topology across the menstrual cycle is estradiol dependent and
correlates with individual well-being. Journal of Neuroscience
Research, 99(9), 2271–2286. https://doi.org/10.1002/jnr.24898,
PubMed: 34110041
Lisofsky, N., Mårtensson, J., Eckert, A., Lindenberger, U., Gallinat,
J., & Kühn, S. (2015). Hippocampal volume and functional con-
nectivity changes during the female menstrual cycle. Neuro-
Image, 118, 154–162. https://doi.org/10.1016/j.neuroimage
.2015.06.012, PubMed: 26057590
Liu, X., & Herbison, UN. E. (2013). Dopamine regulation of
gonadotropin-releasing hormone neuron excitability in male
and female mice. Endocrinology, 154(1), 340–350. https://doi
.org/10.1210/en.2012-1602, PubMed: 23239814
Lurie, D. J., Kessler, D., Bassett, D. S., Betzel, R. F., Breakspear, M.,
Kheilholz, S., … Calhoun, V. D. (2020). Questions and controver-
sies in the study of time-varying functional connectivity in resting
fMRI. Network Neuroscience, 4(1), 30–69. https://doi.org/10
.1162/netn_a_00116, PubMed: 32043043
McEwen, B. S., Akama, K. T., Spencer-Segal, J. L., Milner, T. A., &
Waters, E. M. (2012). Estrogen effects on the brain: Actions
beyond the hypothalamus via novel mechanisms. Behavioral
Neuroscience, 126(1), 4–16. https://doi.org/10.1037/a0026708,
PubMed: 22289042
McEwen, B. S., & Milner, T. UN. (2017). Understanding the broad
influence of sex hormones and sex differences in the brain. Jour-
nal of Neuroscience Research, 95(1–2), 24–39. https://doi.org/10
.1002/jnr.23809, PubMed: 27870427
Moraga-Amaro, R., van Waarde, A., Doorduin, J., & de Vries,
E. F. J. (2018). Sex steroid hormones and brain function: PET
imaging as a tool for research. Journal of Neuroendocrinology,
30(2), e12565. https://doi.org/10.1111/jne.12565, PubMed:
29237239
Mueller, J. M., Pritschet, L., Santander, T., Taylor, C. M., Grafton,
S. T., Jacobs, E. G., & Carlson, J. M. (2021). Dynamic community
detection reveals transient reorganization of functional brain net-
works across a female menstrual cycle. Network Neuroscience,
5(1), 125–144. https://doi.org/10.1162/netn_a_00169, PubMed:
33688609
Newman, M. E. J., & Girvan, M. (2004). Finding and evaluating
community structure in networks. Physical Review E, 69(2),
Network Neuroscience
1203
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
T
/
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network dynamics are associated with reproductive hormones
026113. https://doi.org/10.1103/PhysRevE.69.026113, PubMed:
14995526
Novelli, L., & Razi, UN. (2022). A mathematical perspective on
edge-centric brain functional connectivity. Nature Communica-
zioni, 13(1), 2693. https://doi.org/10.1038/s41467-022-29775
-7, PubMed: 35577769
Parkes, L., Fulcher, B., Yücel, M., & Fornito, UN. (2018). An evalua-
tion of the efficacy, reliability, and sensitivity of motion correc-
tion strategies for resting-state functional MRI. NeuroImage,
171, 415–436. https://doi.org/10.1016/j.neuroimage.2017.12
.073, PubMed: 29278773
Petersen, N., Kilpatrick, l. A., Goharzad, A., & Cahill, l. (2014).
Oral contraceptive pill use and menstrual cycle phase are asso-
ciated with altered resting state functional connectivity. Neuro-
Image, 90, 24–32. https://doi.org/10.1016/j.neuroimage.2013
.12.016, PubMed: 24365676
Pletzer, B., Harris, T.-A., Scheuringer, A., & Hidalgo-Lopez, E.
(2019). The cycling brain: Menstrual cycle related fluctuations
in hippocampal and fronto-striatal activation and connectivity
during cognitive tasks. Neuropsychopharmacology, 44(11),
1867–1875. https://doi.org/10.1038/s41386-019-0435-3,
PubMed: 31195407
Pope, M., Fukushima, M., Betzel, R. F., & Sporns, O. (2021).
Modular origins of high-amplitude cofluctuations in fine-scale
functional connectivity dynamics. Proceedings of the National
Academy of Sciences, 118(46), e2109380118. https://doi.org/10
.1073/pnas.2109380118, PubMed: 34750261
Energia, J. D., Mitra, A., Laumann, T. O., Snyder, UN. Z., Schlaggar,
B. L., & Petersen, S. E. (2014). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImage,
84, 320–341. https://doi.org/10.1016/j.neuroimage.2013.08
.048, PubMed: 23994314
Preti, M. G., Bolton, T. A., & Van De Ville, D. (2017). The dynamic
functional connectome: State-of-the-art and perspectives. Neuro-
Image, 160, 41–54. https://doi.org/10.1016/j.neuroimage.2016
.12.061, PubMed: 28034766
Pritschet, L., Santander, T., Taylor, C. M., Layher, E., Yu, S., Mugnaio,
M. B., … Jacobs, E. G. (2020). Functional reorganization of
brain networks across the human menstrual cycle. NeuroImage,
220, 117091. https://doi.org/10.1016/j.neuroimage.2020
.117091, PubMed: 32621974
Pritschet, L., Taylor, C. M., Santander, T., & Jacobs, E. G. (2021).
Applying dense-sampling methods to reveal dynamic endocrine
modulation of the nervous system. Current Opinion in Behavioral
Scienze, 40, 72–78. https://doi.org/10.1016/j.cobeha.2021.01
.012, PubMed: 35369044
Satterthwaite, T. D., Elliott, M. A., Gerraty, R. T., Ruparel, K.,
J., Calkins, M. E., … Wolf, D. H.
Loughead,
(2013). An
improved framework for confound regression and filtering for
control of motion artifact in the preprocessing of resting-state
functional connectivity data. NeuroImage, 64(1), 240–256.
https://doi.org/10.1016/j.neuroimage.2012.08.052, PubMed:
22926292
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, UN. J., … Yeo, B. T. T. (2018). Local-global parcellation of
the human cerebral cortex from intrinsic functional connectivity
MRI. Cerebral Cortex, 28(9), 3095–3114. https://doi.org/10.1093
/cercor/bhx179, PubMed: 28981612
Seider, N. A., Adeyemo, B., Mugnaio, R., Newbold, D. J., Hampton,
J. M., Scheidter, K. M., … Dosenbach, N. U. F. (2021). Precision
diffusion imaging. bioRxiv. https://doi.org/10.1101/2021.02.19
.432023
Shine, J. M., Aburn, M. J., Breakspear, M., & Poldrack, R. UN. (2018).
The modulation of neural gain facilitates a transition between
functional segregation and integration in the brain. eLife, 7,
e31130. https://doi.org/10.7554/eLife.31130, PubMed:
29376825
Sinchak, K., Mohr, M. A., & Micevych, P. E. (2020). Hypothalamic
astrocyte development and physiology for neuroprogesterone
induction of the luteinizing hormone surge. Frontiers in Endocri-
nology, 11, 420. https://doi.org/10.3389/fendo.2020.00420,
PubMed: 32670203
Sorrentino, P., Seguin, C., Rucco, R., Liparoti, M., Lopez, E. T.,
Bonavita, S., … Zalesky, UN. (2021). The structural connectome
constrains fast brain dynamics. eLife, 10, e67400. https://doi
.org/10.7554/eLife.67400, PubMed: 34240702
Sporns, O., & Betzel, R. F. (2016). Modular brain networks. Annual
Review of Psychology, 67, 613–640. https://doi.org/10.1146
/annurev-psych-122414-033634, PubMed: 26393868
Sporns, O., Faskowitz, J., Teixeira, UN. S., Cutts, S. A., & Betzel, R. F.
(2021). Dynamic expression of brain functional systems dis-
closed by fine-scale analysis of edge time series. Network Neu-
roscience, 5(2), 405–433. https://doi.org/10.1162/netn_a_00182,
PubMed: 34189371
Sporns, O., Tononi, G., & Kötter, R. (2005). The human connec-
tome: A structural description of the human brain. PLoS Compu-
tational Biology, 1(4), e42. https://doi.org/10.1371/journal.pcbi
.0010042, PubMed: 16201007
Tanner, J. C., Faskowitz, J., Byrge, L., Kennedy, D. P., Sporns, O., &
Betzel, R. F. (2022). Synchronous high-amplitude co-fluctuations
of functional brain networks during movie-watching. bioRxiv.
https://doi.org/10.1101/2022.06.30.497603
Taxier, l. R., Gross, K. S., & Frick, K. M. (2020). Oestradiol as a
neuromodulator of learning and memory. Nature Reviews Neu-
roscience, 21(10), 535–550. https://doi.org/10.1038/s41583-020
-0362-7, PubMed: 32879508
Taylor, C. M., Pritschet, L., Olsen, R. K., Layher, E., Santander, T.,
Grafton, S. T., & Jacobs, E. G. (2020). Progesterone shapes medial
temporal lobe volume across the human menstrual cycle. Neuro-
Image, 220, 117125. https://doi.org/10.1016/j.neuroimage.2020
.117125, PubMed: 32634592
Tustison, N. J., Avants, B. B., Cook, P. A., Zheng, Y., Egan, A.,
Yushkevich, P. A., & Gee, J. C. (2010). N4ITK: Improved N3
bias correction. IEEE Transactions on Medical Imaging, 29(6),
1310–1320. https://doi.org/10.1109/ TMI.2010.2046908,
PubMed: 20378467
van den Heuvel, M. P., Scholtens, l. H., Turk, E., Mantini, D.,
Vanduffel, W., & Feldman Barrett, l. (2016). Multimodal anal-
ysis of cortical chemoarchitecture and macroscale fMRI
resting-state functional connectivity. Human Brain Mapping,
37(9), 3103–3113. https://doi.org/10.1002/ hbm.23229,
PubMed: 27207489
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organization
of the human connectome. Journal of Neuroscience, 31(44),
15775–15786, https://doi.org/10.1523/ JNEUROSCI.3539-11
.2011, PubMed: 22049421
Network Neuroscience
1204
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network dynamics are associated with reproductive hormones
Váša, F., Seidlitz, J., Romero-Garcia, R., Whitaker, K. J., Rosenthal,
G., Vértes, P. E., … Bullmore, E. T. (2018). Adolescent tuning of
association cortex in human structural brain networks. Cerebral
Cortex, 28(1), 281–294. https://doi.org/10.1093/cercor/bhx249,
PubMed: 29088339
Weis, S., Hodgetts, S., & Hausmann, M. (2019). Sex differences and
menstrual cycle effects in cognitive and sensory resting state
networks. Brain and Cognition, 131, 66–73. https://doi.org/10
.1016/j.bandc.2017.09.003, PubMed: 29030069
Zhang, Y., Brady, M., & Smith, S. (2001). Segmentation of brain
MR images through a hidden Markov random field model
and the expectation-maximization algorithm. IEEE Transactions
on Medical Imaging, 20(1), 45–57. https://doi.org/10.1109/42
.906424, PubMed: 11293691
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
7
3
1
1
8
1
2
1
5
4
8
0
2
N
e
N
_
UN
_
0
0
3
0
7
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network Neuroscience
1205