RESEARCH
Reconfigurations in brain networks upon
awakening from slow wave sleep: Interventions
and implications in neural communication
Cassie J. Hilditch1*
, Kanika Bansal2,3*, Ravi Chachad1, Lily R. Wong1,
Nicholas G. Bathurst4, Nathan H. Feick1, Amanda Santamaria5,
Nita L. Shattuck6, Javier O. Garcia3**, and Erin E. Flynn-Evans4**
1Fatigue Countermeasures Laboratory, Department of Psychology, San José State University, San José, CA, USA
2Department of Biomedical Engineering, Università della Columbia, New York, NY, USA
3US DEVCOM Army Research Laboratory, Humans in Complex Systems Division, Aberdeen Proving Ground, MD, USA
4Fatigue Countermeasures Laboratory, Human Systems Integration Division, NASA Ames Research Center,
Moffett Field, CA, USA
5Cognitive and Systems Neuroscience Research Hub, University of South Australia, Adelaide, SA, Australia
6Operations Research Department, Naval Postgraduate School, Monterey, CA, USA
*Co-first authors.
**Co-senior authors.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
N
e
N
_
UN
_
0
0
2
7
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Keywords: Sleep inertia, Short-wavelength-enriched light, Network communication, Graph
theoretical framework
ABSTRACT
Sleep inertia is the brief period of impaired alertness and performance experienced
immediately after waking. Little is known about the neural mechanisms underlying this
phenomenon. A better understanding of the neural processes during sleep inertia may offer
insight into the awakening process. We observed brain activity every 15 min for 1 hr following
abrupt awakening from slow wave sleep during the biological night. Using 32-channel
electroencephalography, a network science approach, and a within-subject design, we
evaluated power, clustering coefficient, and path length across frequency bands under both a
control and a polychromatic short-wavelength-enriched light intervention condition. Noi
found that under control conditions, the awakening brain is typified by an immediate
reduction in global theta, alpha, and beta power. Simultaneously, we observed a decrease in
the clustering coefficient and an increase in path length within the delta band. Exposure to
light immediately after awakening ameliorated changes in clustering. Our results suggest that
long-range network communication within the brain is crucial to the awakening process and
that the brain may prioritize these long-range connections during this transitional state. Nostro
study highlights a novel neurophysiological signature of the awakening brain and provides a
potential mechanism by which light improves performance after waking.
AUTHOR SUMMARY
Using a graphical framework approach, our findings suggest that following awakening from
slow wave sleep: (UN) a prioritization scheme may underlie recovery rates for different
behaviors; (B) long-range neural connections orchestrating local-global operations are
uniquely disrupted; E (C) light is able to minimize disruption to long-range connections,
revealing the potential mechanism through which light acts as a countermeasure. Questo
research (UN) advances the knowledge of neural processes during the transition from sleep to
wakefulness; (B) demonstrates a mechanism underlying a brain-behavior relationship that may
a n o p e n a c c e s s
j o u r n a l
Citation: Hilditch, C. J., Bansal, K.,
Chachad, R., Wong, l. R., Bathurst,
N. G., Feick, N. H., Santamaria, A.,
Shattuck, N. L., Garcia, J. O., &
Flynn-Evans, E. E. (2023).
Reconfigurations in brain networks
upon awakening from slow wave
sleep: Interventions and implications
in neural communication. Network
Neuroscience, 7(1), 102–121. https://
doi.org/10.1162/netn_a_00272
DOI:
https://doi.org//10.1162/netn_a_00272
Supporting Information:
https://doi.org/10.1162/netn_a_00272
Received: 11 April 2022
Accepted: 5 agosto 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Cassie J. Hilditch
cassie.j.hilditch@nasa.gov
Handling Editor:
Bratislav Misic
Copyright: © 2022
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
Reconfigurations in brain networks upon awakening from slow wave sleep
serve as a target for future countermeasure research; E (C) applies a novel methodological
approach to sleep-wake brain states. Further research is needed to apply this analytical
method to alternative interventions and sleep-wake transition scenarios.
Sleep inertia:
The brief period of impaired alertness
and cognitive performance
experience immediately after
waking.
Homeostatic sleep pressure:
The increase in sleep propensity with
increasing sleep loss or extended
wakefulness.
Circadian rhythm:
The change in sleep propensity
across a near-24-hr cycle as
determined by a circadian
pacemaker (suprachiasmatic
nucleus).
Graph theoretical framework:
A mathematical framework used to
describe the relationships between
connected objects.
Clustering coefficient:
Estimates the tendency of a node’s
neighbors within a network to also be
linked.
Path length:
Estimates the number of edges, SU
average, that must be traversed to
connect any two nodes within a
rete.
Small-world network:
A network lying in between a
random network and a regular
lattice, reflecting intermediate values
of both path length and clustering.
Network Neuroscience
INTRODUCTION
Immediately after waking from sleep there is a temporary period of reduced alertness and per-
formance. The impact of this so-called sleep inertia on behavioral performance measurements
has been well described, including impaired reaction times (Hilditch et al., 2016; Van Dongen
et al., 2001), memory (Achermann et al., 1995; Santhi et al., 2013), decision-making (Bruck &
Pisani, 1999; Horne & Moseley, 2011), and a variety of other cognitive functions (Burke et al.,
2015). These behaviors are also associated with a perceived state of sleepiness (Burke et al.,
2015; Santhi et al., 2013), disorientation (Dinges, 1990), poor mood (Hilditch et al., 2022),
and misperceptions of performance (Hilditch et al., 2016).
Existing sleep inertia research has associated the waking process with several neural
the brain (Ferrara
changes that include increased delta power over posterior regions of
et al., 2006; Marzano et al., 2011), reduced beta power across all brain regions (Marzano
et al., 2011), and increased functional connectivity of the default mode network (Vallat et al.,
2019). È interessante notare, the links between observed impaired performance and the neural
behavior during sleep inertia are also commonly associated with sleep-related neural pro-
cesses and states of sleepiness due to homeostatic and circadian pressures (Aeschbach et al.,
1997). These observations suggest a complex orchestration of neural elements supporting the
transition from sleep to wakefulness spanning these oppositional constructs. Preliminary
research investigating this complexity in neural network changes has suggested broad func-
tional connectivity changes post-sleep, with the default mode network and the delta and
beta bands playing a critical role in the network changes transitioning from sleep to wake-
fulness (Chen et al., 2020; Vallat et al., 2019). These connectivity changes, Tuttavia, Avere
yet to be characterized and it is unknown whether an intervention may moderate these brain
i cambiamenti.
Understanding how heterogeneous neural elements of the brain coalesce to produce
behavior and subjective experience may be understood via a graph theoretical framework.
Using this framework, the brain is visualized as a graph or network made up of a collection
of nodes (specified brain regions) and edges (connections) that represent brain elements and
the corresponding statistical relationship between them. Topological description of brain net-
works within this framework can provide quantitative insights into the underlying mechanisms
that give rise to emerging neural properties such as specialization and efficiency of information
processing (Bassett & Sporns, 2017), a variety of cognitive phenomena (Bassett & Sporns,
2017; Garcia et al., 2018; Medaglia et al., 2015), transitioning brain states (Miraglia et al.,
2021), and abnormalities in neurological disorders (Y. Liu et al., 2008). Two common metrics,
clustering coefficient and path length, have been used to describe properties of many complex
systems, from biological phenomena (Watts & Strogatz, 1998) to higher level systems (Telesford
et al., 2011). Clustering coefficient estimates the tendency of a node’s neighbors within a net-
work to also be linked. Path length, on the other hand, estimates the number of edges, SU
average, that must be traversed to connect any two nodes within a network. When these met-
rics are at intermediate levels, associated with neither random nor regular networks, Essi
describe the properties of a small-world network, which has been established as a popular
103
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
N
e
N
_
UN
_
0
0
2
7
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
model to describe functional brain networks by facilitating both localized and distributed pro-
cessing of information.
In the current study, we describe the neurophysiological profile of the awakening brain
under ecologically relevant sleep and circadian pressures using graph theoretical analysis of
functional connectivity with 32-channel electroencephalography (EEG). Using clustering and
path length, we see that sleep inertia is characterized by a global shift in these metrics imme-
diately after awakening. Inoltre, we confirm, using a within-subject, randomized, crossover
intervention design, that polychromatic short-wavelength-enriched light exposure alleviates
this sleep inertia effect. We interpret our findings within the context of discontinuity of neural
elements and efficiency of brain processes while the brain transitions from sleep to
wakefulness.
RESULTS
EEG was analyzed from 11 participants (6 female; 23.1 ± 4.4 years, range 19–35 years). Data
were recorded while participants performed a reaction time task (psychomotor vigilance task,
PVT) following nocturnal awakenings from slow wave sleep (SWS). Table S1 in the Supporting
Information contains sleep history information; see Hilditch et al. (2022) for details of sleep
macrostructure prior to awakenings. Briefly, because of the functional associations with
defined categories of oscillations within the brain (Nunez & Srinivasan, 2006) and previous
associations of band-specific spectral power and sleep inertia (Ferrara et al., 2006; Marzano
et al., 2011; Vallat et al., 2019), we first assessed the evolution of global spectral power (cioè.,
average across all channels) of the EEG recorded during the four separate test bouts and com-
pared them with the pre-sleep baseline assessment. Then, via pairwise connectivity (wPLI) A
estimate the phase-based relationship between channels, we estimated graph metrics that pur-
posefully targeted the segregating and integrating aspects of the network. The temporal evo-
lution of these estimates were then assessed to evaluate the duration of sleep inertia. Previous
studies assessing the impact of sleep inertia typically report severe behavioral impairments that
resolve within 15–30 min of waking (Hilditch & McHill, 2019). Remaining mild impairment
may take up to an hour or more to dissipate, depending on conditions such as sleep pressure,
sleep stage at awakening, and time of day (Hilditch & McHill, 2019; Jewett et al., 1999). Given
these previous findings and our behavioral results under similar conditions (Hilditch et al.,
2016; Hilditch et al., 2022), we consider a neural change to be due to sleep inertia when,
compared with baseline, there is a significant change in the assessed metric at the first test
bout (cioè., 2 min after awakening; Figura 1, BL vs. T1C).
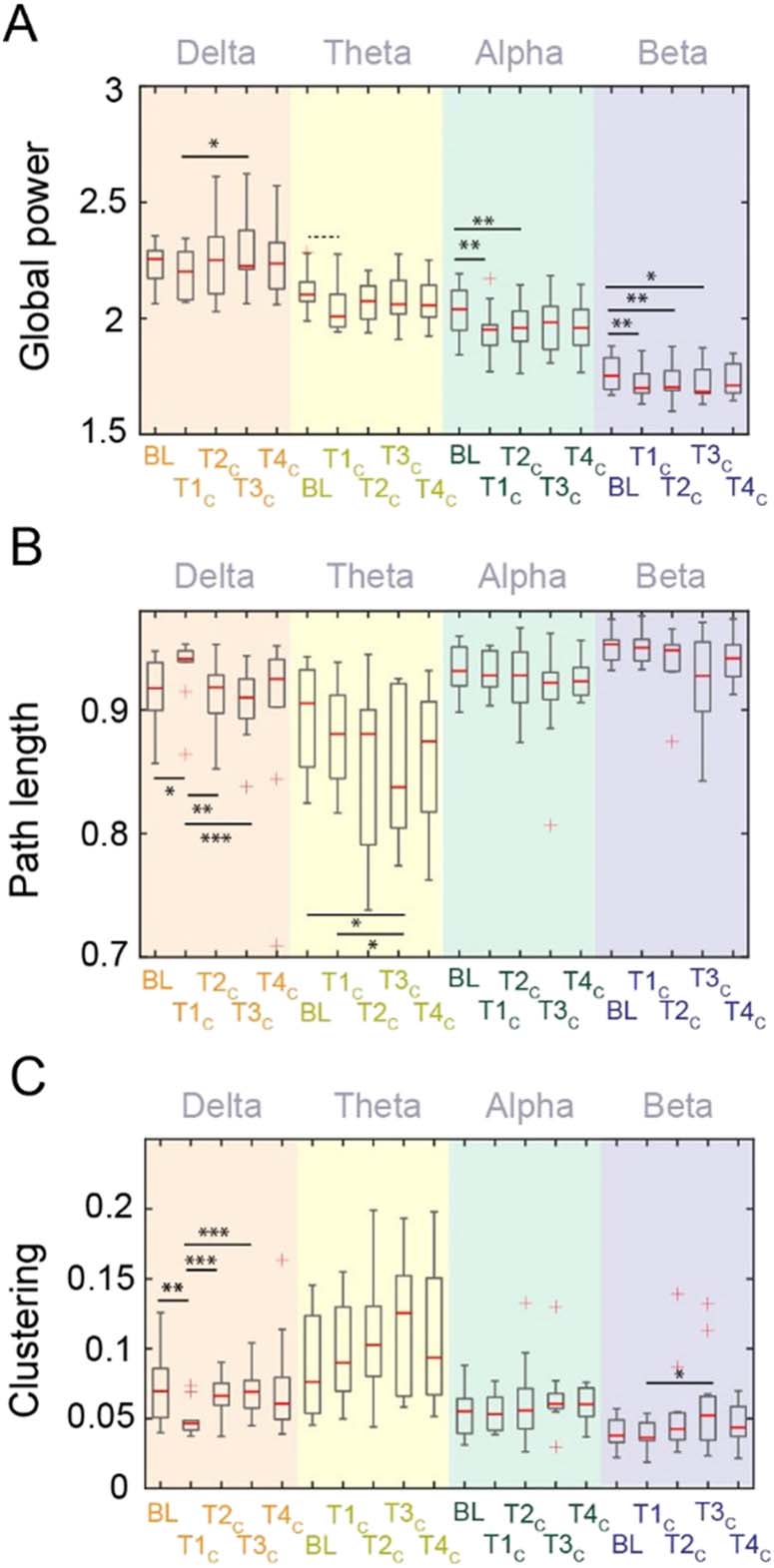
Global Power of Lower Frequencies Recovers Faster Than Higher Frequencies During Sleep Inertia
Figure 1A displays the average spectral power across all channels as a function of test bout.
Statistical comparisons between these test bouts within each frequency band describe a com-
plex coordination of neural firing that may be initially disturbed upon awakening but then
gradually recovers. Compared with participants’ global beta power prior to sleep (following
moderate sleep restriction), beta global power was significantly reduced at T1C (T (10) =
3.47, p = 0.006, effect size Hedge’s g = 0.6), T2C (T (10) = 3.54, p = .005, g = 0.56), E
T3C (T (10) = 2.98, p = 0.014, g = 0.47). Allo stesso modo, compared with baseline, global alpha power
was significantly reduced in the first two test bouts in the control (dim red light) condition (T1C:
T (10) = 3.84, p = 0.003, g = 0.76; T2C: T (10) = 3.56, p = 0.005, g = 0.66). In the theta band,
compared with baseline, we observed that the global power was marginally lower only at T1C
(T (10) = 2.2, p = 0.053, g = 0.71). There were no significant differences between baseline and
Network Neuroscience
104
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
N
e
N
_
UN
_
0
0
2
7
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
N
e
N
_
UN
_
0
0
2
7
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Comparison for (UN) power and (B), (C) brain network properties across test bouts for each
frequency band in the control condition (dim red light). BL = baseline, T#C = Test bout # during the
control condition. Asterisks represent significant difference on a paired t test without any further
correction applied such that *p < 0.05, **p < 0.01, ***p < 0.001. Dashed line denotes marginally
significant difference (p = 0.053).
post-awakening test bouts within the delta band (all p > 0.05). Similar patterns of global power
were observed under equivalent testing conditions performed one week apart (see Figure S1A
in the Supporting Information). To better understand the specificity of these findings, we also
explored aperiodic components effects (Donoghue et al., 2020) with the preprocessing steps
used here (Figure S2A in the Supporting Information) and those specific to the aperiodicity
analysis (Figure S2C in the Supporting Information). We find that the aperiodic components
using the preprocessing steps here displayed sustained differences between the pre-sleep base-
line and subsequent temporal intervals after awakening, most similar to the findings within the
higher frequencies of global power (see Figure S2 in the Supporting Information). Overall,
Anche se, our results presented here are not driven by aperiodic signals in the EEG; Tuttavia,
using another preprocessing pipeline did show promise for this analysis. For an extended
discussion on the effects of preprocessing on these aperiodic results, see the Supporting
Information.
Network Neuroscience
105
Reconfigurations in brain networks upon awakening from slow wave sleep
Network Properties of Delta Band Connectivity Display Unique Characteristics During Sleep Inertia
Prossimo, we considered the global graph metrics of average shortest path length as a measure of
integration and communication efficiency, in addition to average clustering coefficient as a
metric of segregation. Under control conditions, path length was significantly increased in
the delta band immediately after awakening compared with the pre-sleep baseline (T (10) =
−2.52, p = 0.03, g = 0.73). Path length reached baseline levels at T2C (Figure 1B). Allo stesso modo,
we observed a significant reduction of clustering coefficient immediately after awakening from
SWS at night (T1C) compared with pre-sleep baseline (T (10) = 4.0, p = 0.002, g = 1.17;
Figure 1C). The clustering coefficient also returned to baseline levels at T2C and persisted at
that level. Except for a single effect of path length in the theta band significantly decreasing
more than 30 min after awakening (T (10) = 2.65, p = 0.02, g = 0.76), there were no significant
differences in these metrics compared with baseline in other frequency bands. Così, sentiero
length and clustering coefficient within the delta band reflect a robust sleep inertia signal, con
an initial significant change immediately after awakening, followed by recovery towards base-
line at later time points. Similar patterns in delta band connectivity were observed under
equivalent testing conditions performed one week apart (see Figure S1B–C in the Supporting
Information).
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
N
e
N
_
UN
_
0
0
2
7
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Polychromatic Short-Wavelength-Enriched Light Exposure at Awakening Attenuates Neural Network
Changes Associated With Sleep Inertia in the Delta Band
We also assessed the effect of polychromatic short-wavelength-enriched light as a way to
potentially alleviate the neural changes associated with sleep inertia. Infatti, previous studies
have shown that short-wavelength-enriched light has acute alerting properties, especially at
night (Lok et al., 2018; Souman et al., 2018), and this effect has been demonstrated during
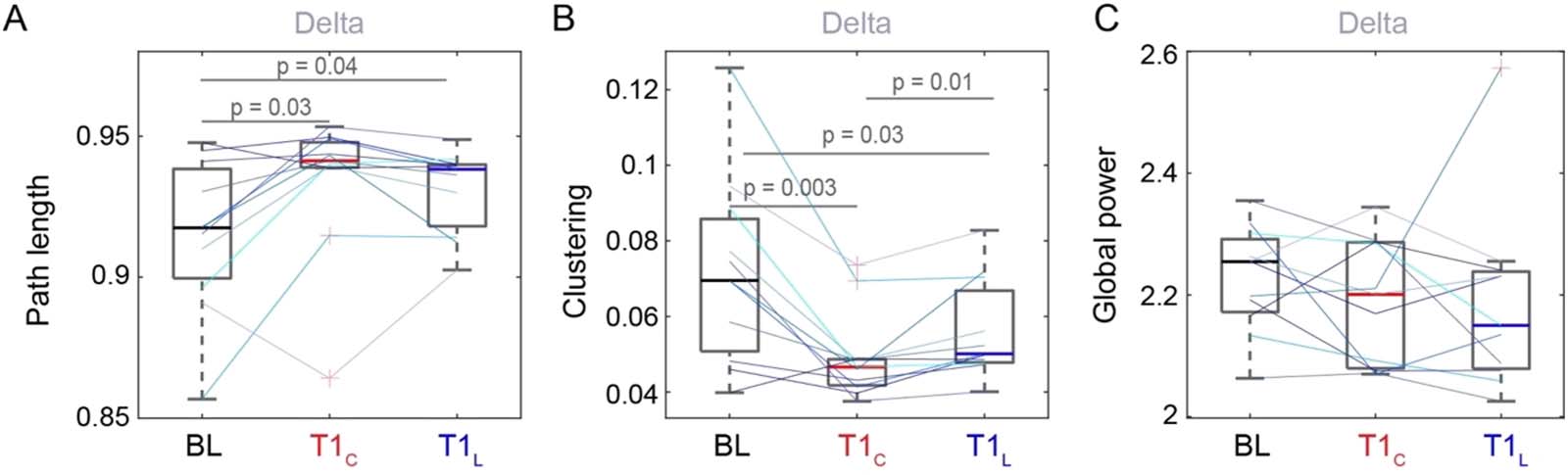
the sleep inertia period (Hilditch et al., 2022). Figura 2 displays the effect of polychromatic
short-wavelength-enriched light on sleep inertia for each of the estimated global metrics across
the frequency bands of interest for the baseline condition, the control condition at T1 (T1C),
and the light intervention condition also at T1 (T1L). Importantly, we were interested in the
effects that showed a difference between the control condition and the light intervention con-
dition at this first time point at which the sleep inertia process is maximally influencing neural
networks and behavior. See Figure S3 in the Supporting Information for a visualization of all
time points under the light condition.
Under the polychromatic short-wavelength-enriched light intervention condition, sentiero
length at T1 in the delta band was not significantly different from control (p > 0.05;
Figure 2A). The clustering coefficient at T1 in the delta band, on the other hand, significantly
increased in the light condition compared with control (T (10) = −3.04, p = 0.01, g = 0.54);
Tuttavia, the clustering coefficient was still significantly lower than baseline at T1 in the light
condition (T (10) = 2.56, p = 0.03, g = 0.8; Figure 2B). Individual lines show a similar pattern for
most participants. There were no significant changes in power between the control and the
polychromatic short-wavelength-enriched light conditions for any frequency band, suggesting
that polychromatic short-wavelength-enriched light has little impact on power immediately
after awakening from SWS (Figure 2C; Figures S2B and S4 in the Supporting Information). Fur-
ther, Figure S4 shows that power at T1L remains significantly lower than baseline in the alpha
and beta bands. Figure S5 in the Supporting Information shows that the clustering coefficient
recovered to baseline levels at T2 in both conditions. Just as delta was the only frequency band
reflecting sleep inertia under control conditions, the changes observed for delta path length
and clustering under the light condition were not observed in other frequency bands,
Network Neuroscience
106
Reconfigurations in brain networks upon awakening from slow wave sleep
Figura 2. Brain network properties comparing pre-sleep baseline (BL), control at T1 (T1C), and light at T1 (T1L) for (UN), (B) the delta frequency
band and (C) delta power. Colored lines represent individual participants. Here, p represents the p value on a paired t test without any further
correction applied.
indicating a frequency-specific role of delta following awakening from SWS and in response to
polychromatic short-wavelength-enriched light.
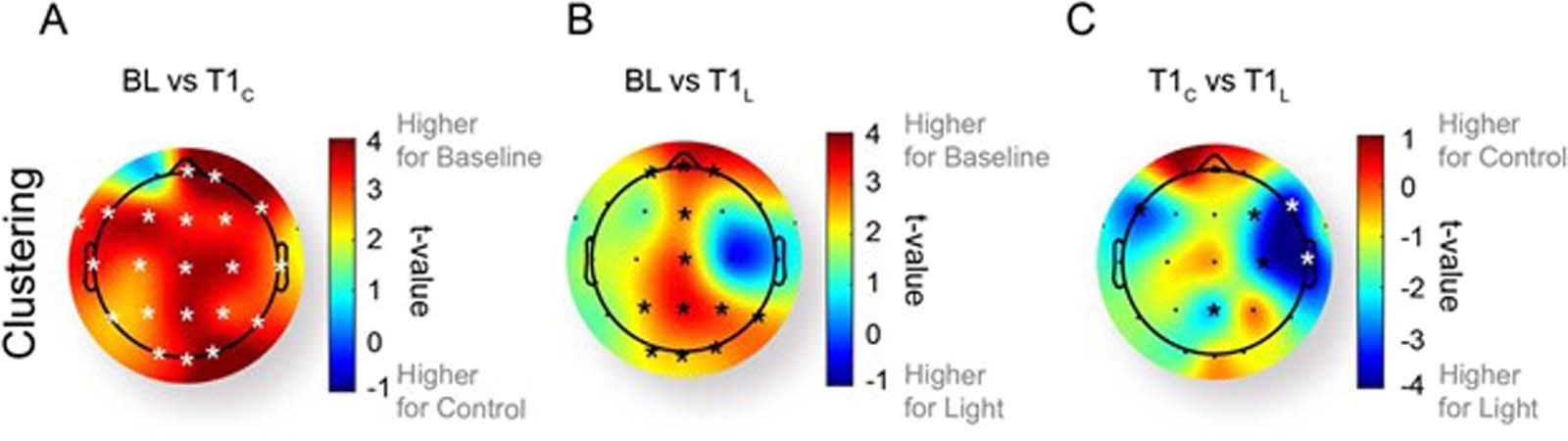
Sleep Inertia Is Characterized by a Global Reduction in Clustering and Region-Specific Rescue
With Light
To understand regional contributions to the changes in the clustering coefficient, we next com-
pared each electrode’s clustering coefficient between the baseline, controllo, and light condi-
tions in the delta band. Compared with the pre-sleep baseline, in the control condition all but
one electrode showed a significant reduction in the clustering coefficient immediately after
awakening (q < 0.05 after correction for multiple comparisons; Figure 3A). When exposed
to light immediately after awakening, there was a trend for reduced clustering across the mid-
line regions of the scalp (Figure 3B), but this did not survive correction for multiple comparisons
(q > 0.05). When comparing the control and light conditions immediately after awakening (at
T1), significant differences were observed in the right hemisphere, with higher clustering in the
light intervention condition (Electrode F8: T (10) = −3.91; Electrode T8: T (10) = −3.95). We also
inspected the regional differences in power and degree at each electrode (see Figures S6–S8 in
the Supporting Information). È interessante notare, in inspecting all of the different effects that we
observe at the global average level, we similarly see a variety of electrodes (scattered across
the scalp) that may be contributing to this effect.
Subjective Sleepiness and Behavioral Performance Are Associated With Changes in Small-Worldness
To further attempt to understand these results, we performed correlational analysis at the crit-
ical time period for the sleep inertia effects following awakening (T1) and some behavioral and
subjective measures collected within the experiment. We concentrate on the control period
(T1C) and specifically compare global power, average clustering, average path length with the
average PVT speed and number of lapses and one subjective measurement of sleepiness
(Karolinska Sleepiness Scale; KSS). È interessante notare, for this T1C period, global power had no sig-
nificant associations with either behavioral or subjective data estimated or collected from par-
ticipants ( p > 0.05); Tuttavia, clustering and path length showed several significant effects for
both behavioral and subjective ratings. For the subjective data (KSS rating), during the T1C
period, we observed a significant positive relationship with path length (R = 0.67, p =
0.034) within the alpha frequency range, suggesting that higher path length is associated with
perceived sleepiness. Also, within the alpha range, clustering coefficient was associated with
the KSS rating as well (R = −0.64, p = 0.048), suggesting that subjects perceived more sleep-
iness when their brains were displaying lower average clustering coefficient behavior. These
Network Neuroscience
107
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
N
e
N
_
UN
_
0
0
2
7
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
Figura 3. Change in clustering between (UN) baseline (BL) and control at T1 (T1C), (B) baseline and light at T1 (T1L), E (C) control and light at
T1 across scalp regions in the delta band. Asterisks represent electrodes with significant difference on a paired t test (P < 0.05). Electrodes that
survived an additional correction for multiple comparisons are highlighted in white (q < 0.05). (See Figures S6–S8 in the Supporting Information
for an additional analysis across the metrics and frequency bands shown within the main text.)
results suggest that a link between small-worldness in the brain within the alpha range may be
linked to perceived sleepiness.
Behavioral responses, such as PVT speed and lapses, also displayed interesting associa-
tions, but within the theta range for clustering coefficient and path length, where higher clus-
tering and lower path length are associated with a faster speed (R = 0.7, p = 0.024; R = −0.7,
p = 0.022). Lapses show a similar trend where lower clustering and higher path length are
associated with more lapses (R = −0.67, p = 0.034; R = 0.67, p = 0.032). These results again
suggest that metrics associated with small-worldness not only are playing a role in perceived
sleepiness, but also are associated with performance fluctuations on a PVT within the theta
range.
DISCUSSION
This is the first study to describe the changes to small-world network dynamics in the waking
brain and the impact of polychromatic short-wavelength-enriched light on this profile. Our
analyses revealed significant reductions in clustering and increases in path length in the
delta band immediately after awakening from SWS at night. Exposure to polychromatic
short-wavelength-enriched light attenuated the changes observed in clustering, suggesting
a specific regime of segregation and integration driving the waking process. Network analyses,
together with our investigation of an intervention and associations with both subjective sleepi-
ness and objective behavioral performance, provide unique insight into the neurophysiological
profile of sleep inertia beyond previous approaches.
Small-Worldness Is Altered During Sleep Inertia
Studies of characteristically small-world activity have shown that small-world properties are
higher during sleep, especially in the slower oscillatory schemes, compared with wakefulness
(Ferri et al., 2007, 2008). Given this finding, we might expect higher small-worldness features
immediately after waking from sleep if there is a slow transition from sleep to wakefulness. We
observed, however, a significant reduction in clustering coefficient and increased path length
immediately following sleep compared with pre-sleep levels. This change suggests a distinc-
tive scheme towards a more random network with both an observed lower clustering of
channels and simultaneously more segregation across them, indicating a relative decrease
in small-worldness. Moreover, with fMRI-estimated connectivity, Liu et al. (2014) observed
that under conditions of sleep deprivation, the estimated small-worldness increased relative
to rested wakefulness, suggesting that the brain may compensate for sleep deprivation via this
Network Neuroscience
108
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
mechanism. These findings may appear contradictory to what we observed in this study, but
important experimental manipulations may provide a key understanding to how small-
worldness shapes neural behavior during the critically important state of sleep, including
how engaged participants are during a task, and the targeted network, as defined by nuances
of the methodology or analysis (e.g., fMRI BOLD vs. EEG power or EEG oscillatory scheme).
Interestingly, with EEG, Koenis et al. (2013) observed a decrease in small-worldness within
the alpha band while awake but sleep-deprived, but reported no changes in the delta band.
Importantly, these findings were reported while participants were engaged in a reaction time
task. Given the similarities with our study design, the differences in our outcomes may reflect a
unique profile of the awakening brain that is distinct from the state of sleep deprivation itself. It
is notable that immediately after awakening, the primary features of small-worldness describe
a unique aspect of brain segregation and integration, independent of both sleep-like features
and the influence of sleep deprivation. This finding may indicate that, while participants are
engaged in a task, sleep inertia is characterized by neural network reconfigurations that arise
due to the disruption of SWS into an awake state. These reconfigurations place the brain net-
work into a state of high segregation.
Inspecting the relationship of behavioral measurements to the graph metrics, PVT perfor-
mance, as estimated with clustering coefficient and path length, did not show a relationship
with global power; however, these metrics showed a relationship with clustering coefficient
and path length within the theta band, suggesting that changes in small-worldness are related
to behavioral performance fluctuations across subjects. This adds to previous research that has
found a significant relationship between performance measurements of this task with fluctua-
tions in power within the theta band within subjects (Gorgoni et al., 2014; Jung et al., 1997)
and expands our understanding of network changes associated with this task (Hoedlmoser
et al., 2011).
Complementary to the aforementioned study (Koenis et al., 2013), our subjective measure-
ment of sleepiness (KSS) was associated with our metrics of small-worldness within the alpha
band, but the alpha band, taking all observed effects into account, did not display a sleep
inertia effect. There is a unique relationship between the alpha oscillation and awareness, gen-
erally, as it has been shown, in certain cases, to halt the processing of visual information
(Mathewson et al., 2009) acting as a gatekeeper of information and even plays a critical role
in temporal expectation (Rohenkohl & Nobre, 2011). While these effects represent rapid infor-
mational processing, these findings have been generalized to suggest that the alpha band may
be required to access knowledge of previous events and grasp contextual (“semantic”) infor-
mation. Our results seem to provide some general support for this theory, but future research is
needed to explore these findings, disentangling these effects on perceived sleepiness, sleep
deprivation, and fluctuations within the alpha band.
Global Power Suggests a Prioritization Scheme May Underlie Sleep Inertia
Previous research seeking to describe the waking brain has focused on EEG power. Oscilla-
tions within the brain characterize rhythmic activity of subpopulations of neurons. It has been
observed that specific cognitive functions are associated with different oscillatory schemes
(Buzsáki, 2006; Neuper & Klimesch, 2006; Nunez & Srinivasan, 2006), where specific fre-
quencies have been associated with top-down processes like executive actions and attention
(Engel et al., 2001; Varela et al., 2001), others with motor control or maintenance of sensori-
motor behaviors (Engel & Fries, 2010), and others with localized and rapid computations (Fries
et al., 2007). Studies of sleep inertia have observed the waking brain exhibiting “sleep-like”
Network Neuroscience
109
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
attributes like high delta power and low beta power, compared with rested wakefulness
(Ferrara et al., 2006; Gorgoni et al., 2015; Marzano et al., 2011). Notably, our baseline testing
was conducted following mild sleep restriction, which may have elevated delta power in our
baseline measures relative to rested wakefulness (Åkerstedt & Gillberg, 1986), and thus
dampened our ability to detect a difference between pre-sleep and post-sleep delta power.
This suggests, however, that delta power itself is not a unique signature of the awakening
brain. Further, we extended observations of EEG power to show the relative rate of recovery
of these frequencies beyond time points previously tested (i.e., beyond 10–25 min post-sleep)
and with greater temporal resolution (i.e., more frequent testing points). We observed a mar-
ginally significant difference in the global power between baseline and the control condition
in the theta band approximately 2 min after awakening and significant differences in global
power between baseline and the control condition up to approximately 17 min in the alpha
band, and approximately 32 min in the beta band after awakening. While none of these obser-
vations were different from each other across post-awakening test bouts, this finding suggests
that there is a measurable change in global power in these bands that is sustained for different
lengths of time. For example, the recovery to baseline was faster for theta frequencies and
longest for beta power, taking at least 30 min to return to pre-sleep levels. These observations
support and extend findings from others who also reported reductions in alpha and beta activ-
ity immediately following awakening (Ferrara et al., 2006; Gorgoni et al., 2015; Marzano
et al., 2011), but did not report the subsequent time course of recovery.
Our observations, overall, suggest that the broader organization of the brain may underlie
the slower dissipation rate of impairment typically observed after approximately 15 min that
continues across the next hour (Jewett et al., 1999; Wertz et al., 2006). These findings suggest
that cognitive functions associated with slower oscillations (i.e., theta band, 3–7 Hz) have a
more rapid recovery than those associated with faster oscillations (i.e., beta band 13–25 Hz).
In addition to these findings, correlational analyses with PVT behavioral performance suggest a
critical role of the theta band small-worldness and its associations with behavioral performance.
Taken together with the findings that specific cognitive functions are associated with
different oscillatory schemes (Buzsáki, 2006; Neuper & Klimesch, 2006; Nunez & Srinivasan,
2006)—slower frequencies generally with top-down processes like executive actions and
attention (Engel et al., 2001; Varela et al., 2001); higher frequencies with motor control or
maintenance of sensorimotor behaviors (Engel & Fries, 2010); and even higher frequencies
with localized, specific, and rapid computations (Fries et al., 2007)—it follows that sleep
inertia is also characterized by different time courses in cognitive subsystem recovery (e.g.,
working memory task vs. simple math task; Achermann et al., 1995; Jewett et al., 1999). It
may also signal a prioritization scheme of the waking brain, from high-level executive func-
tions to motor coordination; however, further research is necessary to understand the potential
differences in cognitive system recovery.
Long-Range Connections Orchestrating Local-Global Operations Are Uniquely Disrupted Within the
Brain Shortly After Awakening
The suggested prioritization scheme in power, in addition to the delta band specificity in net-
work changes, could suggest how this prioritization scheme is implemented in the brain.
Oscillations emanating from the brain, as measured via EEG, are a consequence of short-
and long-range connections within the brain that coalesce to support cognition (Buzsáki
et al., 2013). Slower oscillations often represent the coordination of distal regions of the cortex
that modulate higher frequency oscillations within the brain (Arnulfo et al., 2015; Bragin et al.,
Network Neuroscience
110
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
1995; Buzsáki & Schomburg, 2015; Buzsáki & Wang, 2012; Canolty et al., 2006; Chrobak &
Buzsáki, 1998). In other words, oscillatory activity and the associated cognitive functions
may be understood as a consequence of the ever-present need and importance of global
coordination of local processes (Bressler & Kelso, 2001). This local-global coordination of
neural activity is critical to a variety of cognitive processes (Sauseng & Klimesch, 2008), is
the hallmark of several diseases (Schnitzler & Gross, 2005), and is an organizing principle
of brain activity that has been suggested to be foundational even across multiple species
(Buzsáki et al., 2013). It has even been suggested that the oscillatory synchronization across
brain sites is related to brain size (i.e., distance between coordinated regions; Nunez et al.,
1978; Valdés-Hernández et al., 2010), suggesting that this coordination across distal and
proximal brain regions may not only underlie cognitive processing but also may be associated
with short- and long-range connections within the brain. Within this framework, our global
power results might suggest that long-range connections, as indicated by slower oscillations
(delta and theta) and large-scale cortical integration (Bruns & Eckhorn, 2004), recover more
rapidly than the higher frequency bands (e.g., more localized computation; Buzsáki & Wang,
2012). While global power in lower frequency bands is influenced by ensemble synaptic
action across long ranges and averaged across all channels, our graph theoretic analysis is
derived from the statistical dependencies between nodes, specifically estimating phase-based
relationships between different channels. In other words, while global power in slower
oscillations is sensitive to both amplitude and phase-based relationships aggregated across
the long-range connections within the brain, our graph theoretic results are narrowly sensitive
to phase-based coordination within longer connections, or the communication structure
rather than fluctuations in synchronous neural activity. Taken together, our results display a
prioritization of longer range—perhaps higher cognitive—coordinated activity after awakening,
while simultaneously increasing communication efficiency across them. Future research
may investigate the role of higher segregation and long-range integration of networks during
this process.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Polychromatic Short-Wavelength-Enriched Light Serves as an Intervention to Mitigate Neural Effects of
Sleep Inertia
We also studied an intervention condition in which participants were exposed to a polychro-
matic short-wavelength-enriched light immediately after being awakened from SWS and
throughout the 1-hr testing period. Light, particularly bright, short-wavelength-enriched light,
is known to have acute alerting properties when administered under conditions of sleep dep-
rivation and is particularly effective when used during the biological night (Lok et al., 2018;
Souman et al., 2018). EEG studies have shown that acute exposure to short-wavelength-
enriched light at night during continuous wakefulness reduces delta-theta power (0.5–
5.5 Hz, a biomarker of sleepiness; Lockley et al., 2006; Phipps-Nelson et al., 2009; Rahman
et al., 2014) and increases alpha and high-alpha power (9.5–10.5 Hz, a biomarker of alertness;
Cajochen et al., 1998; Lavoie et al., 2003; Lockley et al., 2006; Rahman et al., 2014).
Recently, we reported the effects of light during the sleep inertia period following nocturnal
awakenings. Our study showed that light modestly improved performance on a psychomotor
vigilance task as well as subjective outcomes such as alertness and mood (Hilditch et al.,
2022). In the current paper, we report the potential mechanisms underlying these neurobe-
havioral effects of light during the sleep inertia period. In contrast to previous findings during
continuous wakefulness at night, we observed only modest, regional changes in power with
light exposure during the sleep inertia period (see the Supporting Information). This
Network Neuroscience
111
Reconfigurations in brain networks upon awakening from slow wave sleep
contradiction in findings may suggest that light acts through a different mechanism during the
sleep inertia period.
We observed that the significant decreases in clustering in the delta band following awaken-
ing in the control condition were attenuated with exposure to polychromatic short-wavelength-
enriched light. Thus, our observed effects of light on the awakening brain appear to counteract
the unique hallmark of the sleep inertia period illustrated by our novel network analysis. Taken
together with our lack of changes in global power with polychromatic short-wavelength-
enriched light exposure, this finding strengthens the specificity of the long-range communication
aspect of our results. During the sleep inertia period, polychromatic short-wavelength-
enriched light may help to restore or protect against the dis-coordination of long-range com-
munication within the brain. This manipulation of delta networks may represent a mechanism
through which polychromatic short-wavelength-enriched light exposure influences neuro-
physiological properties to improve alertness, mood, and performance immediately after
awakening (Hilditch et al., 2022). To our knowledge, no previous studies have evaluated
the effect of a light stimulus on these network dynamics in different brain states; our study
is the first to describe and tentatively interpret these effects.
Methodological Considerations and Limitations
Although our study involved a randomized, within-subject, crossover design with frequent
testing points and 32-channel EEG, it is not without limitations. First, our study did not include
a measure of melatonin to explore whether manipulation of its secretion was a potential con-
tributing factor to the differences observed in the polychromatic short-wavelength-enriched
light condition. We assume based on prior literature, however, that melatonin was suppressed
for the duration of the light exposure (Lewy et al., 1980; Lockley et al., 2006; Zeitzer et al.,
2000) and, therefore, may act as a mechanism for the acute effects of light. Second, while a
strength of our study is that we controlled the sleep stage at awakening, this was traded for
potential changes in circadian sleep pressure between the two awakenings. The order of con-
dition was randomized to limit any differences, and the mean time between awakenings was
90 min (Hilditch et al., 2022), so we do not expect large differences from this design. Further,
the likelihood of a circadian effect in the current study is low as we only observed the reduc-
tion in clustering and path length immediately after awakening, with a rapid return to baseline
levels, which is not characteristic of a circadian effect. We acknowledge that we are unable to
directly disentangle the relative contributions from, or effect of light on, the three sleep pro-
cesses (homeostatic, circadian, inertia) in the current study. However, our first testing point
(T1 at 2 min post-awakening) is a robust proxy for sleep inertia. Importantly, we have, for
the first time, demonstrated the neurophysiological profile of the awakening brain and the
effect of polychromatic short-wavelength-enriched light exposure following awakening from
SWS during a nocturnal sleep episode, which is a common scenario for on-call and emer-
gency service workers.
Conclusions
The current study extends prior research investigating the waking brain by building a more
comprehensive description of the neurophysiological profile of the brain following awakening
from SWS at night. Our results suggest that long-range network communication within
the brain is crucial to the waking process and, further, that the brain may prioritize these
long-range connections, adding to the evolutionary importance of the coordination of local
and global activity within the brain. Moreover, the addition of a within-subject assessment
of the effects of polychromatic short-wavelength-enriched light exposure provides more
Network Neuroscience
112
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
insight into our understanding of sleep inertia, adds a causal aspect to our findings, and
suggests how we might mitigate its effects to improve alertness and performance in safety-
critical scenarios.
MATERIALS AND METHODS
Participants
Twelve healthy young adults participated in the study, having met the following inclusion cri-
teria: healthy (General Health Screening Questionnaire, personal physician’s permission to
participate, approval from onsite physician upon review of urinalysis and blood work screen-
ing); normal sleepers (Pittsburgh Sleep Quality Index ≤ 5; no self-reported sleep problems;
habitual sleep of 7–9 hours); no shiftwork or travel > 3 time zones in the past 3 months
(self-report); free of illicit substances and nicotine (urine toxicology screen); and free of alcohol
during the study period (breathalyzer). All participants provided written informed consent. IL
protocol was approved by the NASA Ames Research Center Institutional Review Board (HRII-
371 and HRII-20-04). One participant’s dataset was incomplete; Perciò, results presented
here reflect a sample population of n = 11.
A priori power calculations were based on anticipated changes in our primary outcome
measure, PVT performance, the results of which are presented elsewhere (Hilditch et al.,
2022). The PVT used in this study is a 5-min reaction time task used to measure vigilant atten-
zione (Loh et al., 2004; Roach et al., 2006). Participants are required to monitor a screen and
respond to stimuli presented at random intervals as soon as possible by pressing a button with
their dominant thumb. Based on previous studies in similar populations, we expected that the
effect size for the change in PVT during sleep inertia would be approximately 0.75. Using
these assumptions, we estimated that we would need 10 participants to detect a change in
performance with 80% power at an alpha level of 0.05. Prior studies investigating sleep inter-
ventions including similar sample sizes have observed significant and equivalent changes in
both reaction time test and EEG connectivity outcome measures (Koenis et al., 2013).
Procedure
The results presented here come from a 2-week study. Here we report a 1-week within-subject,
crossover intervention study with the presentation order of intervention randomized by sex. UN
summary of relevant results from the alternate week are presented in the Supporting
Information.
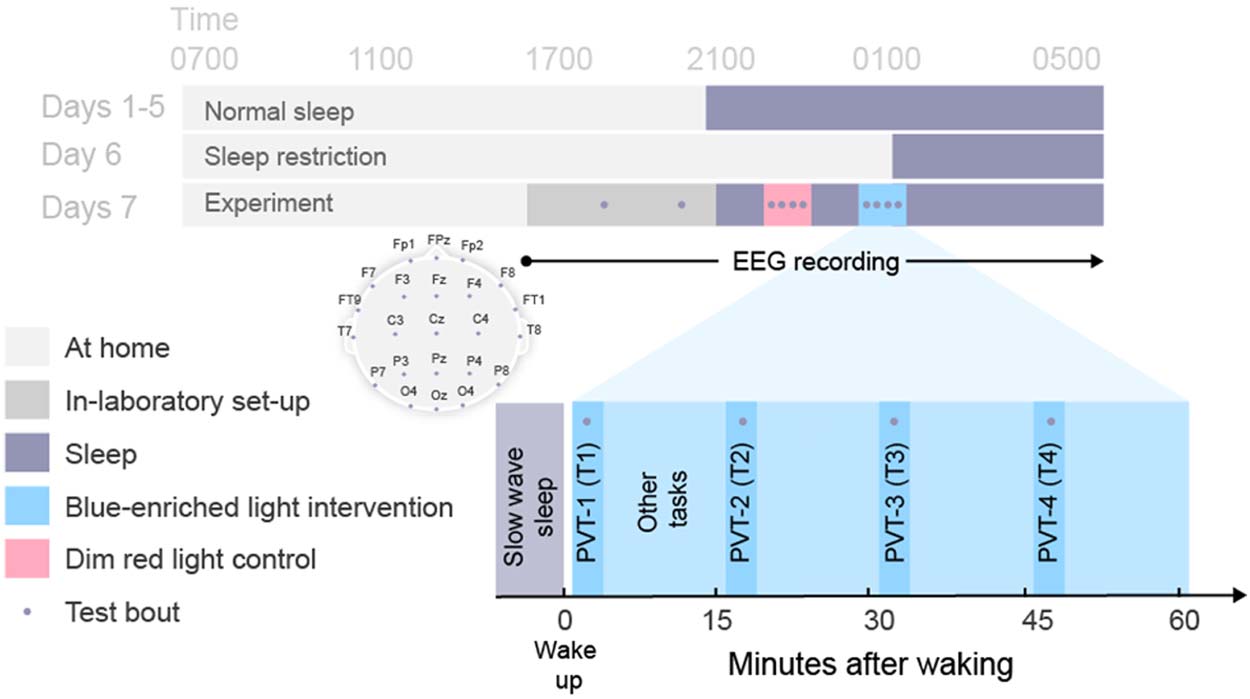
Participants were required to follow a fixed sleep-wake schedule based on habitual sleep
timing for the 6 nights leading up to the in-laboratory visit (Guarda la figura 4). Following a night of
at-home sleep restriction (5 hr), participants were brought into the sound attenuated, leggero- E
temperature-controlled laboratory for pre-sleep procedures that included task familiarization,
electrode setup, and baseline tests prior to overnight observation and testing. Baseline EEG
measures were taken during a PVT performed 2.5 hr before habitual bedtime.
At the participant’s habitual bedtime, all lights were turned off (<0.3 lux) and the participant
was instructed to sleep. EEG was monitored during the sleep period to identify slow wave
sleep (SWS) stages (Stage 3 and 4; Rechtschaffen & Kales, 1968). Participants were awakened
after a minimum of 5 consecutive min of SWS. Immediately upon awakening, a dim red ambi-
ent light was illuminated in the room. In the intervention condition only, at 1 minute post-
awakening, a polychromatic short-wavelength-enriched light was illuminated and remained
on for the hour of testing. The dim red ambient light remained on during the testing period
Network Neuroscience
113
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
Figure 4. Protocol schematic. Light gray shading indicates wakefulness during the at-home portion
of study. Dark gray shading indicates in-laboratory pre-sleep activities including baseline testing (•).
Black shading indicates sleep opportunities (<0.3 lux). Blue and red shading indicate intervention
and control sleep inertia testing periods, respectively. Inset shows electrode montage and post-
awakening test bouts. Clock times shown are approximate and varied depending on habitual
sleep-wake times and appearance of slow wave sleep periods.
in both conditions. A 5-min PVT was performed four times (T1, T2, T3, and T4 at +2, +17, +32,
and +47 min after the awakening, respectively), during which EEG was recorded. At the end of
the testing period, all lights were turned off and the participant was instructed to return to
sleep. EEG was monitored again to identify the next period of 5 consecutive min of SWS, at
which time the participant was awoken again and exposed to the opposite condition (poly-
chromatic short-wavelength-enriched light or control). Following the second testing period, all
lights were turned off and the participant was instructed to sleep until they were awakened at
their habitual wake time.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
Intervention
A 12″ × 24″ canvas of light-emitting diodes (Circadian Positioning Systems, Inc., Newport, RI)
was positioned at 15 degrees to the horizontal angle of gaze and approximately 56 cm away
from the participant. Light levels during the intervention and control conditions were
confirmed via Spectroradiometer ILT950 (International Lighting Technologies, Peabody,
MA). Illuminance, irradiance, equivalent daylight (D65) illuminance (EDI), and peak spectra
during the intervention were 242.77 lux, 0.95 W/m2, 338.03 melanopic lux, and 456 nm,
respectively, measured at the angle of gaze. An ambient dim red light served as the control
(0.26 lux, 0.00 W/m2, 0.10 melanopic lux, 714 nm).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG Analysis
Preprocessing. EEG was recorded during the baseline and post-awakening testing periods
using BrainVision 32-channel caps with sintered Ag/AgCl electrodes (Brain Products GmbH,
Munich, Germany) and BrainVision Recorder software (Brain Products GmbH, Munich,
Germany) recording at a sampling rate of 500 Hz. Additional electrodes included bipolar
horizontal electrooculogram (EOG: left/right placed on the outer canthus of the eye 1 cm
above and below the horizon, respectively), and submental electromyogram (EMG), for stan-
dard monitoring with sleep periods. For visualization while the experimenter monitored the
Network Neuroscience
114
Reconfigurations in brain networks upon awakening from slow wave sleep
EEG, a 70 Hz high-pass filter in conjunction with a notch filter at 60 Hz was used online so
that the experimenter could easily determine whether the participant was in SWS and primed
for an awakening. After the recording sessions, the raw, unfiltered EEG recordings were then
subjected to a thorough artifact editing scheme offline. After a preliminary filtering of the raw
EEG data, using a third-order zero-phase bandpass Butterworth filter (0.5–50 Hz) in EEGLAB
(Delorme & Makeig, 2004), the EEG data were subjected to artifact subspace reconstruction
(ASR; Kothe & Makeig, 2013; Kothe & Jung, 2016; Mullen et al., 2015). This method removes
extremes in data using a time-evolving blind source separation method; importantly, this
method has been shown to be particularly resilient to artifact encountered in real-world sce-
narios (Mullen et al., 2013). To deploy ASR on the dataset, we first created a “clean” reference
signal from each participant’s EEG data by concatenating EEG segments that were at least
1,000 ms long with amplitude below 100 μV, most likely not contaminated by artifacts due
to muscle activity following the awakening. Following the creation of the reference signal, ASR
was then used to clean the EEG that contained large fluctuations greater than 5 standard
deviations beyond the reference signal (in 500-ms chunks). Finally, EEG data from the begin-
ning to the end of each PVT (approximately 5 min each) were identified and then filtered via a
third-order zero-phase bandpass Butterworth filter within the frequency bands of interest
(delta: 1–3 Hz, theta: 4–7 Hz, alpha: 8–12 Hz, beta: 15–25 Hz). This produced four sets of
continuous time courses for each of the time segments following the awakening (T1, T2, T3,
T4) in addition to the baseline pre-sleep time period (BL) for each participant.
Global power spectral density. Power spectral density (PSD) was estimated using a standard
approach of Welch’s average modified periodogram method of spectral estimation (Welch,
1967) in MATLAB (MathWorks, Inc.). The log-transformed PSD values were then standardized
for each electrode before analysis by mean-centering each channel and dividing by standard
deviation across the entire frequency range (0 to 250 Hz). To represent a given frequency band
of interest, standardized PSD values were averaged over the frequency range of that band. For
the sake of simplicity, we use the term power to represent the standardized PSD values and the
term global power to represent the average standardized PSD across the scalp for a given
frequency band of interest.
Network connectivity. To estimate the functional network connectivity between EEG sensors,
we computed the pairwise weighted phase lag index (wPLI), which is known to be highly sen-
sitive to linear and nonlinear interactions (Imperatori et al., 2019); it is well established that
phase-based measurements of connectivity are less susceptible to nuisance artifacts (Lau et al.,
2012). wPLI belongs to a suite of phase-based measurements often deployed with EEG signals
to mitigate the effects of volume conduction (Vinck et al., 2011); however, amplitude-based
connectivity measurements have been shown to display robust effects in particular systems
(e.g., motor; Wei et al., 2021). Importantly, wPLI, when used as the backbone for graphical
analyses, has been shown to produce highly reproducible graph metrics within subjects while
simultaneously capturing individual differences between subjects (Hardmeier et al., 2014).
Thus, within a participant, we calculated the wPLI matrix for all the time points and frequency
bands by using the band-wise filtered EEG activity for the entire 5-min trial epoch. This pro-
duced one weighted and undirected connectivity matrix for T1, T2, T3, T4, and the baseline
pre-sleep time period.
Network analysis. Three network measurements were estimated on the wPLI matrices, includ-
ing clustering coefficient, path length, and (in the Supporting Information) degree. These
common metrics have been used to describe properties of many complex systems including
a variety of biological, social, and other phenomena (Bassett & Sporns, 2017; Collins & Chow,
Network Neuroscience
115
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
1998; Seaton & Hackett, 2004) and are often evoked when describing small-world phenom-
ena (Collins & Chow, 1998). Here, we use clustering coefficient and path length to describe
network changes related to small-worldness/randomness of the network and degree as a visu-
alization of changes in network connectivity.
Specifically, clustering coefficient (C ) estimates the tendency of a node’s neighbors within a
network to also be linked, and may be mathematically described from a connectivity matrix,
W (here estimated via wPLI; Onnela et al., 2005; Rubinov & Sporns, 2010).
Ci ¼
2
Þ
Ki Ki − 1
ð
X
j;k
(cid:2)
wi;jwj;k wk;i
(cid:3)1=3;
(1)
where wi,j represents an element of the connectivity matrix W implying the strength of con-
nectivity between nodes i and j. Ci and Ki represent the clustering coefficient and degree for
node i. Degree of a node is defined as the sum of all the edge weights connected to it and is a
general representation of a node’s connectivity across the network:
Ki ¼
X
j
wi;j:
(2)
Path length, on the other hand, estimates the number of edges, on average, that must be tra-
versed to connect any two nodes within a network. If d(i, j ) represents the shortest path
through edges between nodes i and j, path length λ is given by (Rubinov & Sporns, 2010)
λ ¼
1
ð
n n − 1
Þ
X
i≠j
ð
d i; j
Þ;
(3)
where n is the total number of nodes. In general, the distance between two nodes with strong
connectivity is lower than the distance between nodes with relatively weaker connectivity.
Therefore, to estimate d(i, j ), we used (1 − wi,j), and then the path length was calculated
using Brain Connectivity Toolbox function charpath.m.
Neurobehavioral analysis. As a final analysis, we explored the associations between three
behavioral metrics and the global neural metrics found to be susceptible to the state of sleep
inertia. The three behavioral metrics were subjective sleepiness (KSS), and objective cognitive
performance as assessed by mean PVT speed (1/reaction time, RT) and number of PVT lapses
(RT > 500 ms). Pearson’s correlation coefficient was obtained in MATLAB (MathWorks, Inc.)
via the corr.m function (see the Results section and Supporting Information Figure S9).
Analysis and statistics. Our primary analysis explored the network configuration of the awak-
ening brain under control conditions as compared with the pre-sleep baseline period across
four frequency bands (described above). Our secondary analysis explored the impact of poly-
chromatic short-wavelength-enriched light on this network profile by comparing conditions
(control and light). Specifically, to assess any changes in the metrics of power and network
connectivity we used paired t tests (in MATLAB, MathWorks, Inc.) with a significance criterion
of p < 0.05. Therefore, we compared pre-sleep baseline period and each of the post-sleep test
bouts for both control and light using paired t tests for all the frequencies of interest. Addition-
ally, we compared the respective test bouts between conditions (control and light) to explore
the impact of polychromatic short-wavelength-enriched light across frequencies. Significance
criterion was applied on uncorrected p values. Where appropriate for multiple comparisons,
false discovery rate was used with a q threshold set to 0.05 within a given frequency band. To
estimate the effect size of significant findings, we computed the Hedges’ g, defined as the dif-
ference between the sample means normalized by the pooled standard deviation. Hedges’ g
Network Neuroscience
116
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
was used to express the effect size here, as it better characterizes the effect size with samples
lower than 20 (Lakens, 2013).
Visualization. All figures, boxplots, and topographic plots were created in MATLAB (Math-
Works, Inc.) with common core functionality from MATLAB and some additional functions
with EEGLAB (Delorme & Makeig, 2004). Then, they were imported into Adobe Illustrator
(version 25.3) and combined into panels for visualization.
ACKNOWLEDGMENTS
This work was supported in part by the Naval Medical Research Center’s Naval Advanced
Medical Development Program (MIPR N3239820WXHN007) and the NASA Airspace Oper-
ations and Safety Program, System-Wide Safety. This work was also supported in part through
mission funding from the US DEVCOM Army Research Laboratory (ARL). K. B. also acknowl-
edges support from ARL through Cooperative Agreement Number W911NF-16-2-0158. The
views and conclusions contained in this document are those of the authors and should not
be interpreted as representing the official policies, either expressed or implied, of the ARL
or the US government. The authors would like to thank Aditi Periyannan for her help with
figure design and layout. The authors would also like to thank all the participants who volun-
teered their time for this study.
DATA AVAILABILITY
Data are accessible upon reasonable request as far as allowed by the data-sharing policy and
guidelines established by NASA Ames Research Center.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00272.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
t
.
AUTHOR CONTRIBUTIONS
Cassie J. Hilditch: Conceptualization; Data curation; Funding acquisition; Investigation; Meth-
odology; Writing – original draft; Writing – review & editing. Kanika Bansal: Data curation;
Formal analysis; Methodology; Visualization; Writing – original draft; Writing – review & edit-
ing. Ravi G. Chachad: Data curation; Investigation. Lily R. Wong: Investigation; Writing –
review & editing. Nicholas G. Bathurst: Project administration; Writing – review & editing.
Nathan H. Feick: Investigation; Methodology. Amanda Santamaria: Methodology; Writing –
review & editing. Nita L. Shattuck: Funding acquisition; Writing – review & editing. Javier
O. Garcia: Data curation; Formal analysis; Methodology; Writing – original draft; Writing –
review & editing. Erin E. Flynn-Evans: Conceptualization; Funding acquisition; Investigation;
Methodology; Supervision; Writing – review & editing.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
FUNDING INFORMATION
Cassie Hilditch and Erin Flynn-Evans, NASA Airspace Operations and Safety Program, System-
Wide Safety. Kanika Bansal and Javier Garcia, US DEVCOM Army Research Laboratory.
Kanika Bansal, US DEVCOM Army Research Laboratory, Award ID (Cooperative Agreement
Number): W911NF-16-2-0158. Nita Shattuck, Naval Medical Research Center’s Naval
Advanced Medical Development Program, Award ID: MIPR N3239820WXHN007.
Network Neuroscience
117
Reconfigurations in brain networks upon awakening from slow wave sleep
REFERENCES
Achermann, P., Werth, E., Dijk, D.-J., & Borbely, A. A. (1995). Time
course of sleep inertia after nighttime and daytime sleep epi-
sodes. Archives Italiennes de Biologie, 134(1), 109–119. https://
doi.org/10.4449/aib.v134i1.650, PubMed: 8919196
Aeschbach, D., Dijk, D.-J., & Borbély, A. A. (1997). Dynamics of
EEG spindle frequency activity during extended sleep in humans:
Relationship to slow-wave activity and time of day. Brain
Research, 748(1–2), 131–136. https://doi.org/10.1016/S0006
-8993(96)01275-9, PubMed: 9067453
Åkerstedt, T., & Gillberg, M. (1986). A dose-response study of sleep
loss and spontaneous sleep termination. Psychophysiology,
23(3), 293–297. https://doi.org/10.1111/j.1469-8986.1986
.tb00635.x, PubMed: 3749409
Arnulfo, G., Hirvonen, J., Nobili, L., Palva, S., & Palva, J. M. (2015).
Phase and amplitude correlations in resting-state activity in human
stereotactical EEG recordings. NeuroImage, 112, 114–127. https://
doi.org/10.1016/j.neuroimage.2015.02.031, PubMed: 25721426
Bassett, D. S., & Sporns, O. (2017). Network neuroscience. Nature
Neuroscience, 20(3), 353–364. https://doi.org/10.1038/nn.4502,
PubMed: 28230844
Bragin, A., Jandó, G., Nádasdy, Z., Hetke, J., Wise, K., & Buzsáki, G.
(1995). Gamma (40–100 Hz) oscillation in the hippocampus of
the behaving rat. Journal of Neuroscience, 15(1), 47–60. https://
doi.org/10.1523/ JNEUROSCI.15-01-00047.1995, PubMed:
7823151
Bressler, S. L., & Kelso, J. A. S. (2001). Cortical coordination
dynamics and cognition. Trends in Cognitive Sciences, 5(1),
26–36, https://doi.org/10.1016/S1364-6613(00)01564-3,
PubMed: 11164733
Bruck, D., & Pisani, D. L. (1999). The effects of sleep inertia on
decision-making performance. Journal of Sleep Research, 8(2),
95–103. https://doi.org/10.1046/j.1365-2869.1999.00150.x,
PubMed: 10389091
Bruns, A., & Eckhorn, R. (2004). Task-related coupling from high- to
low-frequency signals among visual cortical areas in human
subdural recordings. International Journal of Psychophysiology,
51(2), 97–116. https://doi.org/10.1016/j.ijpsycho.2003.07.001,
PubMed: 14693360
Burke, T. M., Scheer, F. A., Ronda, J. M., Czeisler, C. A., & Wright
Jr., K. P. (2015). Sleep inertia, sleep homeostatic and circadian
influences on higher-order cognitive functions. Journal of Sleep
Research, 24(4), 364–371. https://doi.org/10.1111/jsr.12291,
PubMed: 25773686
Buzsáki, G. (2006). Rhythms of the brain. Oxford University Press.
https://doi.org/10.1093/acprof:oso/9780195301069.001.0001
Buzsáki, G., Logothetis, N., & Singer, W. (2013). Scaling brain size,
keeping timing: Evolutionary preservation of brain rhythms. Neu-
ron, 80(3), 751–764. https://doi.org/10.1016/j.neuron.2013.10
.002, PubMed: 24183025
Buzsáki, G., & Schomburg, E. W. (2015). What does gamma coher-
ence tell us about inter-regional neural communication? Nature
Neuroscience, 18(4), 484–489. https://doi.org/10.1038/nn.3952,
PubMed: 25706474
Buzsáki, G., & Wang, X.-J. (2012). Mechanisms of gamma oscilla-
tions. Annual Review of Neuroscience, 35, 203–225. https://doi
.org/10.1146/annurev-neuro-062111-150444, PubMed:
22443509
Cajochen, C., Kräuchi, K., Danilenko, K. V., & Wirz-Justice, A.
(1998). Evening administration of melatonin and bright light:
Interactions on the EEG during sleep and wakefulness. Journal
of Sleep Research, 7(3), 145–157. https://doi.org/10.1046/j.1365
-2869.1998.00106.x, PubMed: 9785269
Canolty, R. T., Edwards, E., Dalal, S. S., Soltani, M., Nagarajan,
S. S., Kirsch, H. E., Berger, M. S., Barbaro, N. M., & Knight,
R. T. (2006). High gamma power is phase-locked to theta oscil-
lations in human neocortex. Science, 313(5793), 1626–1628.
https://doi.org/10.1126/science.1128115, PubMed: 16973878
Chen, X., Hsu, C. F., Xu, D., Yu, J., & Lei, X. (2020). Loss of frontal
regulator of vigilance during sleep inertia: A simultaneous
EEG-fMRI study. Human Brain Mapping, 41(15), 4288–4298.
https://doi.org/10.1002/hbm.25125, PubMed: 32652818
Chrobak, J., & Buzsáki, G. (1998). Gamma oscillations in the ento-
rhinal cortex of the freely behaving rat. Journal of Neuroscience,
18(1), 388–398. https://doi.org/10.1523/ JNEUROSCI.18-01
-00388.1998, PubMed: 9412515
Collins, J. J., & Chow, C. C. (1998). It’s a small world. Nature,
393(6684), 409–410. https://doi.org/10.1038/30835, PubMed:
9623993
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source tool-
box for analysis of single-trial EEG dynamics including indepen-
dent component analysis. Journal of Neuroscience Methods,
134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009,
PubMed: 15102499
Dinges, D. F. (1990). Are you awake? Cognitive performance and
reverie during the hypnopompic state. In R. R. Bootzin, J. F.
Kihlstrom, & D. L. Schacter (Eds.), Sleep and cognition
(pp. 159–175). American Psychological Association. https://doi
.org/10.1037/10499-012
Donoghue, T., Haller, M., Peterson, E. J., Varma, P., Sebastian, P.,
Gao, R., Noto, T., Lara, A. H., Wallis, J. D., Knight, R. T.,
Shestyuk, A., & Voytek, B. (2020). Parameterizing neural power
spectra into periodic and aperiodic components. Nature Neuro-
science, 23, 1655–1665. https://doi.org/10.1038/s41593-020
-00744-x, PubMed: 33230329
Engel, A. K., & Fries, P. (2010). Beta-band oscillations—Signalling
the status quo? Current Opinion in Neurobiology, 20(2),
156–165. https://doi.org/10.1016/j.conb.2010.02.015, PubMed:
20359884
Engel, A. K., Fries, P., & Singer, W. (2001). Dynamic predictions:
Oscillations and synchrony in top-down processing. Nature
Reviews Neuroscience, 2(10), 704–716. https://doi.org/10.1038
/35094565, PubMed: 11584308
Ferrara, M., Curcio, G., Fratello, F., Moroni, F., Marzano, C.,
Pellicciari, M. C., & De Gennaro, L. (2006). The electroenceph-
alographic substratum of the awakening. Behavioural Brain
Research, 167(2), 237–244. https://doi.org/10.1016/j.bbr.2005
.09.012, PubMed: 16242789
Ferri, R., Rundo, F., Bruni, O., Terzano, M. G., & Stam, C. J. (2007).
Small-world network organization of functional connectivity of
EEG slow-wave activity during sleep. Clinical Neurophysiology,
Network Neuroscience
118
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
118(2), 449–456. https://doi.org/10.1016/j.clinph.2006.10.021,
PubMed: 17174148
Ferri, R., Rundo, F., Bruni, O., Terzano, M. G., & Stam, C. J. (2008).
The functional connectivity of different EEG bands moves
towards small-world network organization during sleep. Clinical
Neurophysiology, 119(9), 2026–2036. https://doi.org/10.1016/j
.clinph.2008.04.294, PubMed: 18571469
Fries, P., Nikolić, D., & Singer, W. (2007). The gamma cycle. Trends
in Neurosciences, 30(7), 309–316. https://doi.org/10.1016/j.tins
.2007.05.005, PubMed: 17555828
Garcia, J. O., Ashourvan, A., Muldoon, S., Vettel, J. M., & Bassett,
D. S. (2018). Applications of community detection techniques to
brain graphs: Algorithmic considerations and implications for
neural function. Proceedings of the IEEE, 106(5), 846–867.
https://doi.org/10.1109/ JPROC.2017.2786710, PubMed:
30559531
Gorgoni, M., Ferlazzo, F., Ferrara, M., Moroni, F., D’Atri, A.,
Fanelli, S., Torriglia, I. G., Lauri, G., Marzano, C., Rossini,
P. M., & Rossini, P. M. (2014). Topographic electroencephalo-
gram changes associated with psychomotor vigilance task
performance after sleep deprivation. Sleep Medicine, 15(9),
1132–1139. https://doi.org/10.1016/j.sleep.2014.04.022,
PubMed: 25087194
Gorgoni, M., Ferrara, M., D’Atri, A., Lauri, G., Scarpelli, S., Truglia,
I., & De Gennaro, L. (2015). EEG topography during sleep inertia
upon awakening after a period of increased homeostatic sleep
pressure. Sleep Medicine, 16(7), 883–890. https://doi.org/10
.1016/j.sleep.2015.03.009, PubMed: 26004680
Hardmeier, M., Hatz, F., Bousleiman, H., Schindler, C., Stam, C. J.,
& Fuhr, P. (2014). Reproducibility of functional connectivity and
graph measures based on the phase lag index (PLI) and weighted
phase lag index (wPLI) derived from high resolution EEG. PLoS
ONE, 9(10), e108648. https://doi.org/10.1371/journal.pone
.0108648, PubMed: 25286380
Hilditch, C. J., Centofanti, S. A., Dorrian, J., & Banks, S. (2016). A
30-minute, but not a 10-minute, nighttime nap is associated with
sleep inertia. Sleep, 39(3), 675–685. https://doi.org/10.5665
/sleep.5550, PubMed: 26715234
Hilditch, C. J., & McHill, A. W. (2019). Sleep inertia: Current
insights. Nature and Science of Sleep, 11, 155–165. https://doi
.org/10.2147/NSS.S188911, PubMed: 31692489
Hilditch, C. J., Wong, L. R., Bathurst, N. G., Feick, N. H., Pradhan,
S., Santamaria, A., Shattuck, N. L., & Flynn-Evans, E. E. (2022).
Rise and shine: The use of polychromatic short-wavelength-
enriched light to mitigate sleep inertia at night following awak-
ening from slow-wave sleep. Journal of Sleep Research, 31(5),
e13558. https://doi.org/10.1111/jsr.13558, PubMed: 35102669
Hoedlmoser, K., Griessenberger, H., Fellinger, R., Freunberger, R.,
Klimesch, W., Gruber, W., & Schabus, M. (2011). Event-related
activity and phase locking during a psychomotor vigilance task
over the course of sleep deprivation. Journal of Sleep Research,
20(3), 377–385. https://doi.org/10.1111/j.1365-2869.2010
.00892.x, PubMed: 20977513
Horne, J., & Moseley, R. (2011). Sudden early-morning awakening
impairs immediate tactical planning in a changing “emergency”
scenario. Journal of Sleep Research, 20(2), 275–278. https://doi
.org/10.1111/j.1365-2869.2010.00904.x, PubMed: 21518064
Imperatori, L. S., Betta, M., Cecchetti, L., Canales-Johnson, A.,
Ricciardi, E., Siclari, F., Pietrini, P., Chennu, S., & Bernardi, G.
(2019). EEG functional connectivity metrics wPLI and wSMI
account for distinct types of brain functional interactions. Scien-
tific Reports, 9(1), 1–15. https://doi.org/10.1038/s41598-019
-45289-7, PubMed: 31222021
Jewett, M. E., Wyatt, J. K., Ritz-De Cecco, A., Khalsa, S. B., Dijk,
D. J., & Czeisler, C. A. (1999). Time course of sleep inertia dissi-
pation in human performance and alertness. Journal of Sleep
Research, 8(1), 1–8. https://doi.org/10.1111/j.1365-2869.1999
.00128.x, PubMed: 10188130
Jung, T.-P., Makeig, S., Stensmo, M., & Sejnowski, T. J. (1997). Esti-
mating alertness from the EEG power spectrum. IEEE Transactions
on Biomedical Engineering, 44(1), 60–69. https://doi.org/10.1109
/10.553713, PubMed: 9214784
Koenis, M. M. G., Romeijn, N., Piantoni, G., Verweij, I., Van der
Werf, Y. D., Van Someren, E. J. W., & Stam, C. J. (2013). Does
sleep restore the topology of functional brain networks? Human
Brain Mapping, 34(2), 487–500. https://doi.org/10.1002/ hbm
.21455, PubMed: 22076871
Kothe, C. A., & Makeig, S. (2013). BCILAB: A platform for brain–
computer interface development. Journal of Neural Engineering,
10(5), 056014. https://doi.org/10.1088/1741-2560/10/5/056014,
PubMed: 23985960
Kothe, C. A. E., & Jung, T.-P. (2016). Artifact removal techniques with
signal reconstruction. US Patent 2016/0113587-A1, April 28, 2016.
Lakens, D. (2013). Calculating and reporting effect sizes to facilitate
cumulative science: A practical primer for t-tests and ANOVAs.
Frontiers in Psychology, 4, 863. https://doi.org/10.3389/fpsyg
.2013.00863, PubMed: 24324449
Lau, T. M., Gwin, J. T., McDowell, K. G., & Ferris, D. P. (2012).
Weighted phase lag index stability as an artifact resistant measure
to detect cognitive EEG activity during locomotion. Journal of
Neuroengineering and Rehabilitation, 9(1), 1–9. https://doi.org
/10.1186/1743-0003-9-47, PubMed: 22828128
Lavoie, S., Paquet, J., Selmaoui, B., Rufiange, M., & Dumont, M.
(2003). Vigilance levels during and after bright light exposure
in the first half of the night. Chronobiology International, 20(6),
1019–1038. https://doi.org/10.1081/CBI-120025534, PubMed:
14680141
Lewy, A. J., Wehr, T. A., Goodwin, F. K., Newsome, D. A., &
Markey, S. P. (1980). Light suppresses melatonin secretion in
humans. Science, 210(4475), 1267–1269. https://doi.org/10
.1126/science.7434030, PubMed: 7434030
Liu, H., Li, H., Wang, Y., & Lei, X. (2014). Enhanced brain
small-worldness after sleep deprivation: A compensatory effect.
Journal of Sleep Research, 23(5), 554–563. https://doi.org/10
.1111/jsr.12147, PubMed: 24673840
Liu, Y., Liang, M., Zhou, Y., He, Y., Hao, Y., Song, M., Yu, C., Liu,
H., Liu, Z., & Jiang, T. (2008). Disrupted small-world networks in
schizophrenia. Brain, 131(4), 945–961. https://doi.org/10.1093
/brain/awn018, PubMed: 18299296
Lockley, S. W., Evans, E. E., Scheer, F. A., Brainard, G. C., Czeisler,
C. A., & Aeschbach, D. (2006). Short-wavelength sensitivity for
the direct effects of light on alertness, vigilance, and the waking
electroencephalogram in humans. Sleep, 29(2), 161–168. https://
doi.org/10.1093/sleep/29.2.161, PubMed: 16494083
Network Neuroscience
119
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
Loh, S., Lamond, N., Dorrian, J., Roach, G., & Dawson, D. (2004).
The validity of psychomotor vigilance tasks of less than
10-minute duration. Behavior Research Methods, Instruments, and
Computers, 36(2), 339–346. https://doi.org/10.3758/BF03195580,
PubMed: 15354700
Lok, R., Smolders, K. C., Beersma, D. G., & de Kort, Y. A. (2018).
Light, alertness, and alerting effects of white light: A literature over-
view. Journal of Biological Rhythms, 33(6), 589–601. https://doi
.org/10.1177/0748730418796443, PubMed: 30191746
Marzano, C., Ferrara, M., Moroni, F., & De Gennaro, L. (2011).
Electroencephalographic sleep inertia of the awakening brain.
Neuroscience, 176, 308–317. https://doi.org/10.1016/j
.neuroscience.2010.12.014, PubMed: 21167917
Mathewson, K. E., Gratton, G., Fabiani, M., Beck, D. M., & Ro, T.
(2009). To see or not to see: Prestimulus α phase predicts visual
awareness. Journal of Neuroscience, 29(9), 2725–2732. https://
doi.org/10.1523/ JNEUROSCI.3963-08.2009, PubMed:
19261866
Medaglia, J. D., Lynall, M.-E., & Bassett, D. S. (2015). Cognitive net-
work neuroscience. Journal of Cognitive Neuroscience, 27(8),
1471–1491. https://doi.org/10.1162/jocn_a_00810, PubMed:
25803596
Miraglia, F., Tomino, C., Vecchio, F., Gorgoni, M., De Gennaro, L.,
& Rossini, P. M. (2021). The brain network organization during
sleep onset after deprivation. Clinical Neurophysiology, 132(1),
36–44. https://doi.org/10.1016/j.clinph.2020.10.016, PubMed:
33254098
Mullen, T., Kothe, C. A., Chi, Y. M., Ojeda, A., Kerth, T., Makeig, S.,
Cauwenberghs, G., & Jung, T.-P. (2013). Real-time modeling and
3D visualization of source dynamics and connectivity using
wearable EEG. In 35th Annual International Conference of the
IEEE Engineering in Medicine and Biology Society (EMBC).
Osaka, Japan. https://doi.org/10.1109/ EMBC.2013.6609968,
PubMed: 24110155
Mullen, T. R., Kothe, C. A., Chi, Y. M., Ojeda, A., Kerth, T., Makeig,
S., Jung, T.-P., & Cauwenberghs, G. (2015). Real-time neuroim-
aging and cognitive monitoring using wearable dry EEG. IEEE
Transactions on Biomedical Engineering, 62(11), 2553–2567.
https://doi.org/10.1109/ TBME.2015.2481482, PubMed:
26415149
Neuper, C., & Klimesch, W. (Eds.). (2006). Event-related dynamics
of brain oscillations. Elsevier.
Nunez, P. L., Reid, L., & Bickford, R. G. (1978). The relationship of
head size to alpha frequency with implications to a brain wave
model. Electroencephalography and Clinical Neurophysiology,
44(3), 344–352. https://doi.org/10.1016/0013-4694(78)90309-7,
PubMed: 76540
Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain: The
neurophysics of EEG (2nd ed.). Oxford University Press. https://
doi.org/10.1093/acprof:oso/9780195050387.001.0001
Onnela, J.-P., Saramäki, J., Kertész, J., & Kaski, K. (2005). Intensity
and coherence of motifs in weighted complex networks. Physical
Review E, 71(6), 065103. https://doi.org/10.1103/PhysRevE.71
.065103, PubMed: 16089800
Phipps-Nelson, J., Redman, J. R., Schlangen, L. J., & Rajaratnam,
S. M. (2009). Blue light exposure reduces objective measures
of sleepiness during prolonged nighttime performance testing.
Chronobiology International, 26(5), 891–912. https://doi.org/10
.1080/07420520903044364, PubMed: 19637049
Rahman, S. A., Flynn-Evans, E. E., Aeschbach, D., Brainard, G. C.,
Czeisler, C. A., & Lockley, S. W. (2014). Diurnal spectral sensi-
tivity of the acute alerting effects of light. Sleep, 37(2), 271–281.
https://doi.org/10.5665/sleep.3396, PubMed: 24501435
Rechtschaffen, A., & Kales, A. (1968). A manual of standardized ter-
minology, techniques and scoring system for sleep stages of
human subjects. Brain Information Service/ Brain Research
Institute.
Roach, G. D., Dawson, D., & Lamond, N. (2006). Can a shorter
psychomotor vigilance task be used as a reasonable substitute
for the ten-minute psychomotor vigilance task? Chronobiology
International, 23(6), 1379–1387. https://doi.org/10.1080
/07420520601067931, PubMed: 17190720
Rohenkohl, G., & Nobre, A. C. (2011). Alpha oscillations related to
anticipatory attention follow temporal expectations. Journal of
Neuroscience, 31(40), 14076–14084. https://doi.org/10.1523
/JNEUROSCI.3387-11.2011, PubMed: 21976492
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Santhi, N., Groeger, J. A., Archer, S. N., Gimenez, M., Schlangen,
L. J., & Dijk, D.-J. (2013). Morning sleep inertia in alertness and
performance: Effect of cognitive domain and white light condi-
tions. PLoS ONE, 8(11), e79688. https://doi.org/10.1371/journal
.pone.0079688, PubMed: 24260280
Sauseng, P., & Klimesch, W. (2008). What does phase information
of oscillatory brain activity tell us about cognitive processes?
Neuroscience and Biobehavioral Reviews, 32(5), 1001–1013.
https://doi.org/10.1016/j.neubiorev.2008.03.014, PubMed:
18499256
Schnitzler, A., & Gross, J. (2005). Normal and pathological oscillatory
communication in the brain. Nature Reviews Neuroscience, 6(4),
285–296. https://doi.org/10.1038/nrn1650, PubMed: 15803160
Seaton, K. A., & Hackett, L. M. (2004). Stations, trains and
small-world networks. Physica A: Statistical Mechanics and Its
Applications, 339(3–4), 635–644. https://doi.org/10.1016/j
.physa.2004.03.019.
Souman, J. L., Tinga, A. M., Te Pas, S. F., van Ee, R., & Vlaskamp,
B. N. S. (2018). Acute alerting effects of light: A systematic liter-
ature review. Behavioural Brain Research, 337, 228–239. https://
doi.org/10.1016/j.bbr.2017.09.016, PubMed: 28912014
Telesford, Q. K., Joyce, K. E., Hayasaka, S., Burdette, J. H., &
Laurienti, P. J. (2011). The ubiquity of small-world networks.
Brain Connectivity, 1(5), 367–375. https://doi.org/10.1089/brain
.2011.0038, PubMed: 22432451
Valdés-Hernández, P. A., Ojeda-González, A., Martínez-Montes, E.,
Lage-Castellanos, A., Virués-Alba, T., Valdés-Urrutia, L., & Valdes-
Sosa, P. A. (2010). White matter architecture rather than cortical
surface area correlates with the EEG alpha rhythm. NeuroImage,
49(3), 2328–2339. https://doi.org/10.1016/j.neuroimage.2009
.10.030, PubMed: 19850139
Vallat, R., Meunier, D., Nicolas, A., & Ruby, P. (2019). Hard to
wake up? The cerebral correlates of sleep inertia assessed using
combined behavioral, EEG and fMRI measures. NeuroImage,
Network Neuroscience
120
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Reconfigurations in brain networks upon awakening from slow wave sleep
184, 266–278. https://doi.org/10.1016/j.neuroimage.2018.09
.033, PubMed: 30223060
Van Dongen, H. P., Price, N. J., Mullington, J. M., Szuba, M. P.,
Kapoor, S. C., & Dinges, D. F. (2001). Caffeine eliminates psycho-
motor vigilance deficits from sleep inertia. Sleep, 24(7), 813–819.
https://doi.org/10.1093/sleep/24.7.813, PubMed: 11683484
Varela, F., Lachaux, J.-P., Rodriguez, E., & Martinerie, J. (2001). The
brainweb: Phase synchronization and large-scale integration.
Nature Reviews Neuroscience, 2(4), 229–239. https://doi.org/10
.1038/35067550, PubMed: 11283746
Vinck, M., Oostenveld, R., van Wingerden, M., Battaglia, F., &
Pennartz, C. M. A. (2011). An improved index of phase-
synchronization for electrophysiological data in the presence of
volume-conduction, noise and sample-size bias. NeuroImage,
55(4), 1548–1565. https://doi.org/10.1016/j.neuroimage.2011
.01.055, PubMed: 21276857
Watts, D. J., & Strogatz, S. H. (1998). Collective dynamics of
“small-world” networks. Nature, 393(6684), 440–442. https://
doi.org/10.1038/30918, PubMed: 9623998
Wei, H. T., Francois-Nienaber, A., Deschamps, T., Bellana, B.,
Hebscher, M., Sivaratnam, G., Zadeh, M., & Meltzer, J. A.
(2021). Sensitivity of amplitude and phase based MEG measures
of interhemispheric connectivity during unilateral finger move-
ments. NeuroImage, 242, 118457. https://doi.org/10.1016/j
.neuroimage.2021.118457, PubMed: 34363959
Welch, P. (1967). The use of fast Fourier transform for the estimation
of power spectra: A method based on time averaging over short,
modified periodograms. IEEE Transactions on Audio and Electro-
acoustics, 15(2), 70–73. https://doi.org/10.1109/ TAU.1967
.1161901
Wertz, A. T., Ronda, J. M., Czeisler, C. A., & Wright Jr., K. P. (2006).
Effects of sleep inertia on cognition. JAMA, 295(2), 159–164.
https://doi.org/10.1001/jama.295.2.163, PubMed: 16403927
Zeitzer, J. M., Dijk, D. J., Kronauer, R. E., Brown, E. N., & Czeisler,
C. A. (2000). Sensitivity of the human circadian pacemaker to
nocturnal light: Melatonin phase resetting and suppression. Jour-
nal of Physiology, 526(3), 695–702. https://doi.org/10.1111/j
.1469-7793.2000.00695.x, PubMed: 10922269
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
0
2
2
0
7
2
0
0
4
n
e
n
_
a
_
0
0
2
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
121