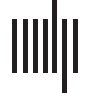RESEARCH
Pattern of frustration formation in the
functional brain network
Majid Saberi1, Reza Khosrowabadi1, Ali Khatibi2, Bratislav Misic3, and Gholamreza Jafari1,4,5
1Institute for Cognitive and Brain Sciences, Shahid Beheshti University, G.C. Tehran, Iran
2Centre of Precision Rehabilitation for Spinal Pain (CPR Spine), School of Sport, Exercise and Rehabilitation Sciences,
University of Birmingham, Birmingham, UK
3McConnell Brain Imaging Centre, Montréal Neurological Institute, McGill University, Montréal, QC, Canada
4Physics Department, Shahid Beheshti University, Tehran, Iran
5Institute of Information Technology and Data Science, Irkutsk National Research Technical University, Irkutsk, Russia
a n o p e n a c c e s s
j o u r n a l
Keywords: Brain network, Functional brain network, Functional connectivity, Negative link,
Frustration, Frustrated system, Signed network, Life-span, Subcortical regions
ABSTRACT
The brain is a frustrated system that contains conflictual link arrangements named frustration.
The frustration as a source of disorder prevents the system from settling into low-energy states
and provides flexibility for brain network organization. In this research, we tried to identify the
pattern of frustration formation in the brain at the levels of region, connection, canonical
rete, and hemisphere. We found that frustration formation has no uniform pattern. Some
subcortical elements have an active role in frustration formation, despite low contributions
from many cortical elements. Frustrating connections are mostly between-network
connections, and triadic frustrations are mainly formed between three regions from three
distinct canonical networks. We did not find any significant differences between brain
hemispheres or any robust differences between the frustration formation patterns of various
life-span stages. Our results may be interesting for those who study the organization of brain
links and promising for those who want to manipulate brain networks.
AUTHOR SUMMARY
Brain network analysis approaches commonly ignore the signs of links. Frustration is a
fascinating phenomenon referring to the conflictual arrangements of signed links. As a source
of instability, it can give valuable information on altering components of a network. It can
specify which brain network elements intend to drive brain network alterations. Accordingly,
we tried to identify the pattern of frustration formation in the brain network, which brain
network elements are more frustrated and which ones are less frustrated. We provided some
maps for frustration formation in the levels of region, connection, canonical network, E
hemisphere. The introduced concept of frustration and our results may be interesting for brain
network scientists.
INTRODUCTION
The brain is an integrative system in that its components cooperate to execute advanced func-
zioni. So considering components of the brain independently risks oversimplification and may
Citation: Saberi, M., Khosrowabadi, R.,
Khatibi, A., Misic, B., & Jafari, G. (2022).
Pattern of frustration formation in the
functional brain network. Network
Neuroscience, 6(4), 1334–1356.
https://doi.org/10.1162/netn_a_00268
DOI:
https://doi.org/10.1162/netn_a_00268
Supporting Information:
https://doi.org/10.1162/netn_a_00268;
https://github.com/majidsaberi
/ BrainNetFrustration
Received: 1 Febbraio 2022
Accepted: 5 Luglio 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Majid Saberi
ma_saberi@sbu.ac.ir
Handling Editor:
Alex Fornito
Copyright: © 2022
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
result in misinterpreting their role during such functions. In recent years, network modeling has
been facilitating the study of the collective behavior of brain elements. Several classes of net-
works including simple connected-disconnected, weighted, and directed networks are being
applied to clear brain mechanisms such as functional segregation and neural integration and
help to discriminate brain disorders (Bassett & Bullmore, 2009; Bassett & Sporns, 2017; Liu
et al., 2017UN; Mišić & Sporns, 2016; Rubinov & Sporns, 2010). Most of these works ignore
the sign of links, either by taking the absolute value of connections or by positive thresholding
on connection values (Garrison et al., 2015; Theis et al., 2021; Wang et al., 2021). They min-
imize the impact of negative connections and disregard interactions between positive and neg-
ative connections.
Considering the brain as a signed network is also a promising approach to investigating
the collective behavior of brain regions, where synchronous and antisynchronous coactiva-
tions of brain regions determine positive and negative links, rispettivamente (Saberi et al., 2021UN)
(Figura 1). Signed networks are commonly used for social system modeling in the context of
friendship and hostility between entities (Facchetti et al., 2011; Tang et al., 2016; Tomasso
et al., 2022; Yang et al., 2007, 2012). Approaches such as the structural balance theory (con-
sisted of social balance and social imbalance) are utilized to investigate signed networks for the
optimized arrangement of relationships (Alabandi et al., 2021; Anchuri & Magdon-Ismail,
2012; Antal et al., 2005; Bagherikalhor et al., 2021; Derr et al., 2018; Facchetti et al., 2011).
Signed network:
A network where links have positive
and negative attributes, per esempio., a social
network with friendship and hostility
links.
Structural balance theory:
A mathematical framework for
studying the balance of relationships
in a system of entities with positive
and negative relations.
Social balance:
When a friend’s friend or an enemy’s
enemy is an enemy and everything
is well.
Social imbalance:
When a friend’s friend or an enemy’s
enemy is an enemy. It is nontrivial;
entities endure a tension and tend to
change their relationships.
Formation of frustrated (imbalanced) and satisfied (balanced) triadic relations in the
Figura 1.
brain. Every region has a specific time course. Nodes of triads correspond to the regions by color.
Correlation coefficients between every pair of activity patterns are also denoted on links. Blue and
red links denote synchrony and antisynchrony between time courses.
Network Neuroscience
1335
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
/
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
Signed links:
Network links with positive and
negative qualities.
Frustration:
A topological constraint that prevents
system energy minimization, it brings
flexibility to the system.
Balance theory:
An attitude change theory developed
by Fritz Heider that explains how the
quality of relationships affects
people’s mind-sets.
Brain signed network:
A brain network made based on
functional neuroimage data where
synchronous and antisynchronous
brain coactivations determine the
positive and negative links between
brain nodes.
Network frustration:
A closed cycle of signed links having
an odd number of negative links and
frustrating properties.
In this context, the optimized state happens when signed links have fewer higher order con-
flictual relations and the network has less tendency to alteration and lower energy. In this
modo, we recently employed the theory to study the structural balance of the resting-state net-
lavoro (Saberi et al., 2021UN). We found that minor negative brain connections get together in a
way to make negative hubs to reduce the number of conflictual signed links arrangements,
which decreases brain energy and brings more stability to the brain network. Our results high-
light the role of negative connections in brain network organization.
Frustration is another interesting phenomenon that can be studied in signed networks. IL
concept of frustration originates from the study of order-disorder systems in many-particle
physics (Toulouse, 1987; Vannimenus & Toulouse, 1977; Villain et al., 1980) where it helps
to understand the mechanism behind system ordering and phase transition (Goremychkin
et al., 2008; Zhao et al., 2019). In spin systems, frustration is defined as topological constraints
between spin neighbors that prevent minimizing system energy (Vannimenus & Toulouse,
1977). Generally, frustration as a source of disorder matters for systemic organization, alter-
ation, and optimization.
In signed networks, frustration refers to nontrivial cycles of signed links, unstable assemblies
that are seldomly found in real networks (Antal et al., 2005; Kirkley et al., 2019; Saberi et al.,
2021UN). Triadic relation is the smallest cycle where other elements influence the quality of a
link, which makes a sense of the system (Winkler & Reichardt, 2013). It is analogous to the
imbalanced triads of Heider’s balance theory (Heider, 1946; Rapoport, 1963): “the friend of a
friend is an enemy” and “the enemy of an enemy is an enemy.” The theory states that entities
of nontrivial relationships are frustrated about their conditions and endure pressure to change
the type of relationships to become balanced: “the friend of a friend is a friend” and “the
enemy of an enemy is a friend.” These conflictual arrangements have been extended to any
cycles with odd numbers of negative links (Aref & Wilson, 2019; Cartwright & Harary, 1956;
Estrada, 2019).
Previous works showed that the resting-state brain signed network locates in a glassy state
containing triadic frustrations (Saberi et al., 2021UN, 2021B). These frustrations prevent the brain
network from reaching minimum energy (absolute stable state). In other words, the resting-
state network is a state that can potentially transition to other brain network states. This result
highlights the role of brain network frustrations in the systemic reorganization of brain links.
Identifying brain network frustrations provides us with the opportunity to control the organi-
zation of brain links that affect the functionality and efficiency of the brain system. It is helpful
for those who are interested in inducing functional changes by the use of neurostimulation and
may promote understanding of the mechanism of brain system functions and dysfunctions. So
we decided to identify the pattern of frustration formation in the brain network system in the
current study. In this regard, we investigated the contribution level of brain elements including
regions, functional connections, canonical networks, and hemispheres in frustration forma-
zione. We explored them for each life-span stage separately and then compared the patterns
of stages.
RESULTS
We designed this study to investigate the pattern of frustration formation in the brain. So we
explored how brain components contribute to brain network frustrations. We preprocessed
anatomical and resting-state fMRI images of healthy subjects from two online repositories,
ABIDE (Di Martino et al., 2014) and Southwest University (Wei et al., 2018), which can ensure
the reliability of the results. Then we extracted regional activations of each functional image
Network Neuroscience
1336
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
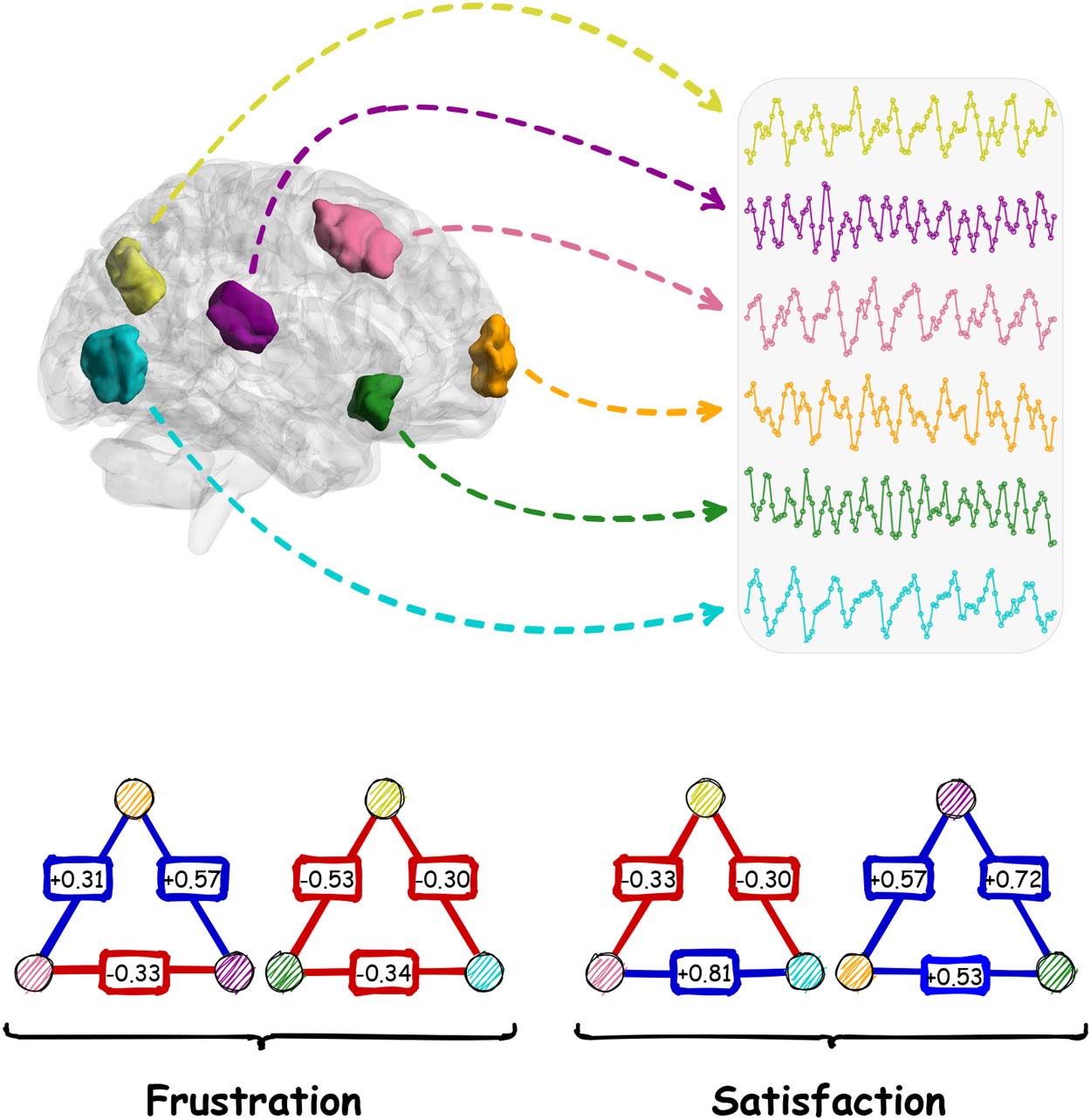
Figura 2. Categorizing Shen’s regions of interest into canonical networks.
based on the parcels of Shen’s atlas (Shen et al., 2013) where the regions were projected to
canonical brain networks (Yeo et al., 2011) (Figura 2). After that, we calculated functional con-
nectivities and formed a signed network for each subject based on the signs of connections.
Then we identified the triadic frustrations of each network and measured the contribution level
of brain regions, functional connections, canonical networks, and hemispheres in their forma-
zioni. On another side, we estimated the null contribution values of mentioned elements
according to the number of appeared frustrations for each subject’s signed network. Finalmente,
we performed a group-level paired comparison for the contribution of every mentioned ele-
ment between actual and null values to find out which regions, connections, canonical net-
works, and hemispheres were significantly involved in the frustration formation for each stage
separately (Figure 3–6). Finalmente, we provided a multimodal map for frustration formation in the
brain stagewise and without considering stages and investigated any significant differences
between contribution patterns of life-span stages.
Contribution of Brain Regions
For each subject, we identified presented frustrations and determined the number of frustra-
tions that each region is involved in. We also calculated the expected values of regional
Network Neuroscience
1337
Pattern of frustration formation in the functional brain network
involvements in the case of a uniform engagement of regions in frustration formation. IL
expected contribution value of a region of interest (ROI) for a subject’s whole-brain network
with NROI regions and NFrust frustrations is derived as follows:
3
NROI
(cid:1) NFrus
Numero 3 is multiplied since a frustration occurs between three regions. In this way, we
provided actual and null values of contributions in frustration formation for 268 regions of
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
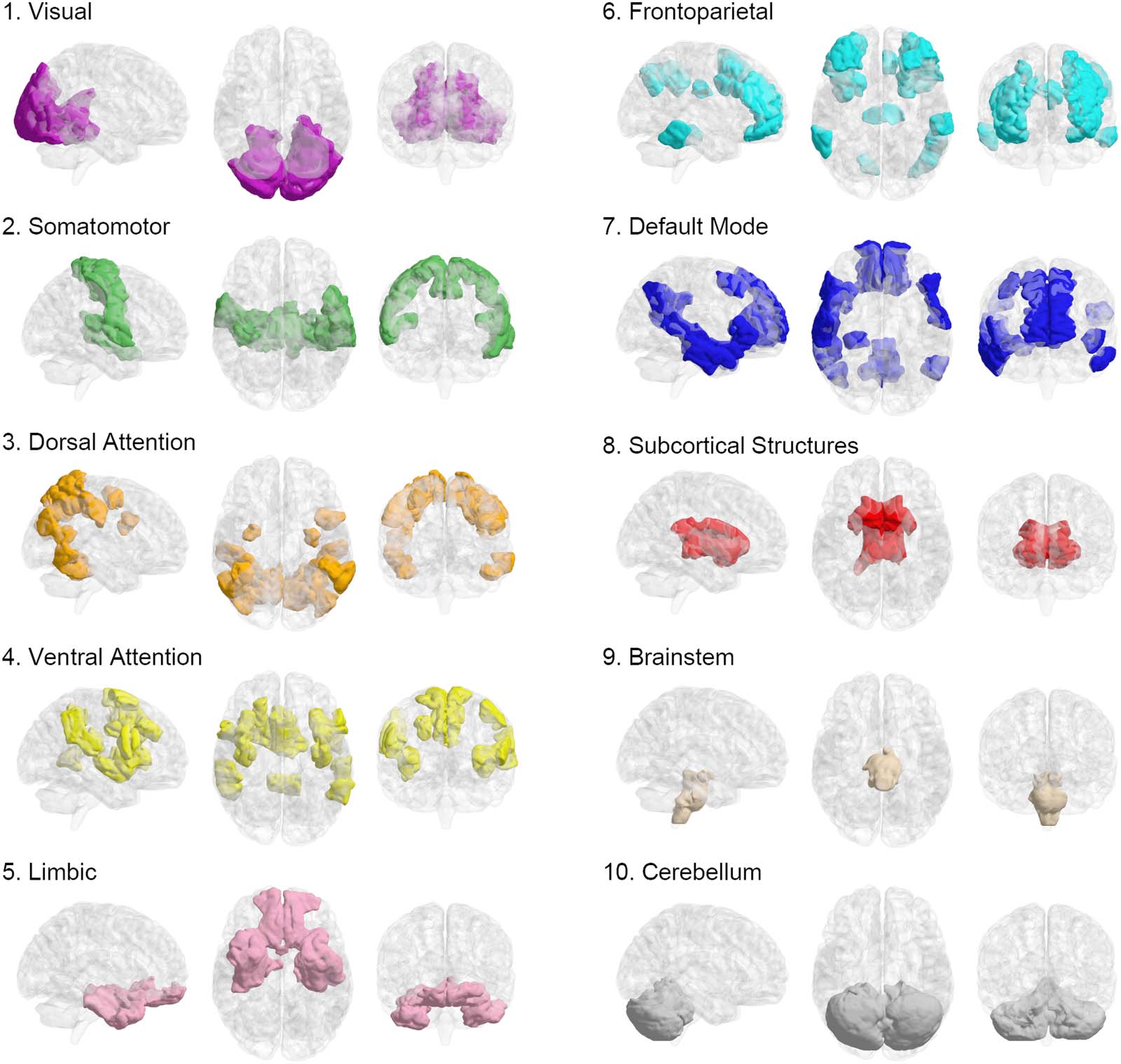
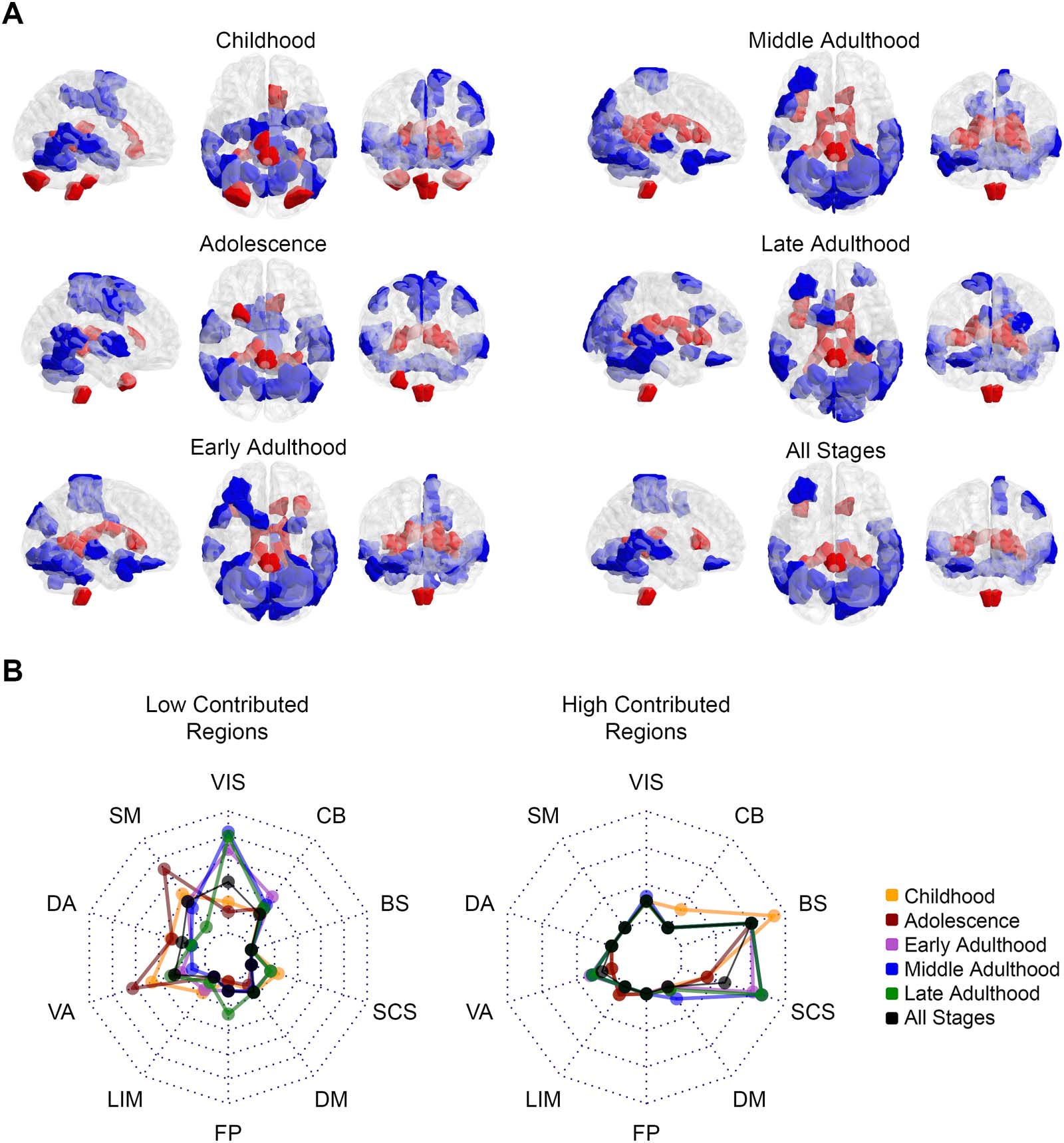
Figura 3. Pattern of regional contribution in frustration formation. (UN) Red-colored and blue-colored areas of the brain maps indicate Shen’s
regions with significantly greater and significantly lower contributions in frustration formation. Brain maps demonstrate the patterns for various
life-span stages and all stages in three representational planes of sagittal, axial, and coronal. (B) Radar charts show what percentage of canon-
ical brain networks are involved in significant areas of panel A. The left radar plot relates to blue areas and the right one corresponds to red
areas. Each dashed polygon indicates 10%. Various line colors are related to life-span stages. VIS = visual; SM = somatomotor; DA = dorsal
Attenzione; VA = ventral attention; LIM = limbic; FP = fronto-parietal; DM = default mode; SCS = subcortical structures; BS = brainstem; CB =
cerebellum.
Network Neuroscience
1338
Pattern of frustration formation in the functional brain network
each subject. Then we performed a paired group-level comparison between actual and null
values of each region to find which regions have a significantly different contribution. Noi
analyzed subjects of each stage separately in addition to disregarding the life-span stages
(Figura 3). Since contribution values were distributed nonnormally, we used Wilcoxon
matched-pairs signed-rank test for this goal. Figure 3A indicates regions with significantly
inferiore (blue-colored) and significantly greater (red-colored) involvements in frustration forma-
tion compared to expected values after multiple comparison corrections on p values using
false discovery rate (FDR) and with large effect sizes (greater than 0.6). We reported the
corrected p values and effect sizes in the Supporting Information. Since the frustration
expresses disorganization in the network, the blue and red areas of the figure represent
regions that have well-organized and disorganized relations with other regions, rispettivamente.
To better interpret the results, we mapped significant brain regions to canonical networks of
Figura 2 to show how red and blue areas belong to the canonical networks. The Methods
section “Parcellation Atlas Projection” describes the projection process that we used to
obtain Figure 2 in detail. Radar plots of Figure 3B represent what percentages of canonical
networks have significant contributions where dotted polygons divide the whole network
volume by 10%. The left and right radar charts correspond to blue-colored and red-colored
regions of Figure 3A. The left radar chart shows low contributed blue regions mostly belong
to the visual network in adulthood and they involve somatomotor and ventral attention net-
works in the early stages. More than 40% of the visual network in middle adulthood and
Di 35% of the somatomotor network and ventral attention network in adolescence have
regions with well-organized relations to other brain regions. The right radar chart also shows
that most of the high contributed red regions of Figure 3A belong to subcortical structures
and the brain stem. Despite early stages, more than 20% of subcortical structures in adult-
hood have areas with disorganized relations to other regions. Also, more than 20% del
brain stem volume is significantly involved in frustration formations, an inferior portion of
the brain stem according to Figure 3A. For all stages on the brain map, radar charts indicate
moderate values, which we desired in case of disregarding stages. We also used the non-
parametric Kruskal–Wallis test to investigate between-stage differences of regional contribu-
zioni. We performed the test between contribution values of various stages for each region
separately and then corrected p values from false positives that occurred by multiple com-
parisons using FDR besides effect size measurement. There is no region with a significant
corrected p value and large effect size (greater than 0.14) that indicates no robust contribution
differences between stages. Although, Supporting Information Figure S1 shows regions with
corrected p value lower than 0.05 and medium effect size (between 0.06 E 0.14). To test
whether significant regions of Figure 3A appears randomly or not, we compared the entropy of
the all stage pattern with the entropy of its shuffled pattern for greater and lower contributions
separately. We explained the way we calculated entropies in the Methods section “Entropy
Calculation.” Supporting Information Figure S2 shows the histogram of entropies related to
shuffled patterns compared to the entropy of actual patterns. It indicates that both lower con-
tributed regions (blue-colored) and greater contributed regions (red-colored) have signifi-
cantly lower entropies that validate the nonrandomness of the regional significant patterns
of Figure 3.
Contribution of Functional Connections
After regional investigations, we explored the contribution of functional connections in frus-
tration formation. So for each subject, we counted the number of triadic frustrations that
Network Neuroscience
1339
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
each connection is involved in. Also, we estimated the expected contribution values of each
connection in case of uniform involvement of connections as follows:
(cid:3) (cid:1) NFrus
3
(cid:1)
2
NROI
where NFrust and NROI are the number of presented frustration and the number of nodes in
the subject’s network.
also denotes the 2-combinations of NROI that is equals to the
(cid:1)
(cid:3)
2
NROI
number of links in a fully connected network with NROI regions. The factor of 3 also appears
because frustration engages three connections. In this way, we calculated actual and null
contribution values for each connection of each subject. After that, we performed a paired
group-level analysis between actual and null values for each functional connection using
Wilcoxon matched-pairs signed-rank test. We chose this nonparametric test since the values
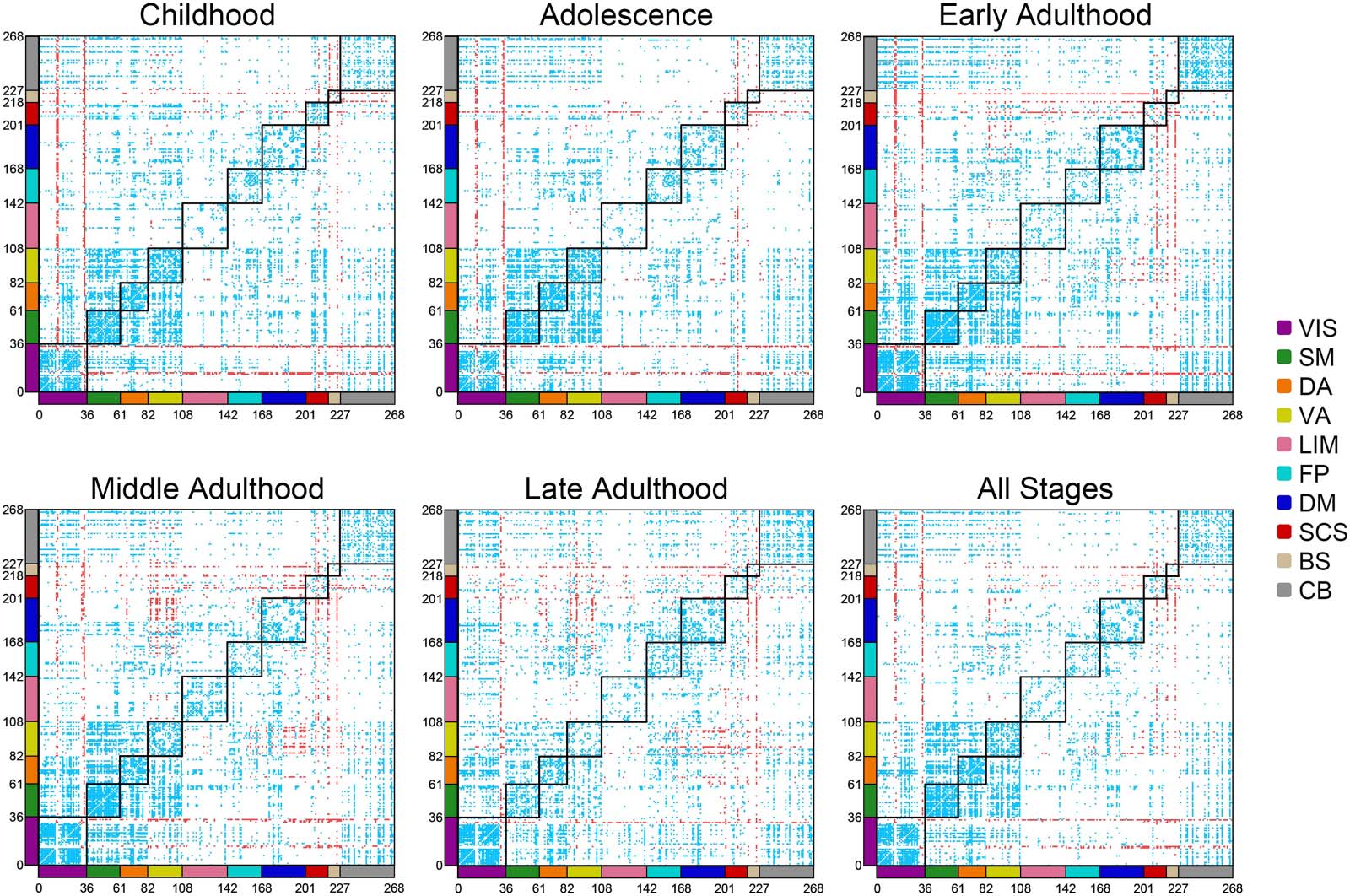
were not distributed normally. Figura 4 displays contribution maps of functional connec-
tions in frustration formation for each stage separately and for all stages. Each cell corre-
sponds to a functional connection between two regions of interest, so heat maps represent
mutual connections between 268 of Shen’s regions. The regions are categorized based on
networks of Figure 2 by segmenting the axes of heat maps. Also, black squares discriminate
between-network connections. Blue and red cells indicate connections with significantly
lower and significantly greater contributions in frustration formation that have FDR cor-
rected p values smaller than 0.05 and large effect sizes (greater than 0.6). The figure does
not exhibit a distributed pattern of high contributed connections (red-colored cells). It seems
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
/
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 4. Contribution maps of functional connections in frustration formation. The first five maps demonstrate the maps related to life-span
stages and the last one corresponds without considering the stage. Shen’s 268 ROIs are categorized into the 10 networks of Figure 2 by axes
coloring. Cells also represent functional connections between every two ROI. Red cells display those connections with a significantly greater
contribution to frustration formation, and blue cells indicate those connections with significantly lower involvement. Black squares discrim-
inate within-network connections. VIS = visual; SM = somatomotor; DA = dorsal attention; VA = ventral attention; LIM = limbic; FP = fronto-
parietal; DM = default mode; SCS = subcortical structures; BS = brainstem; CB = cerebellum.
Network Neuroscience
1340
Pattern of frustration formation in the functional brain network
that connections of several visual and subcortical regions with other regions have signifi-
cantly great contributions to frustration formation. Many default mode to ventral attention
connections of adulthood are also high contributed. On the contrary, lower contributed
connections are more distributed in the brain network. Most of these connections are
within-network type, especially those located in visual, somatomotor, and attentional net-
works. There are also lots of between-network connections that have lower contributions in
frustration formation, especially connections between visual, somatomotor, cerebellar, E
attentional regions. We reported corrected p values and effect sizes of the comparisons in
the Supporting Information.
As well as the last section, we also wanted to explore the randomness of the pattern of the
significantly contributed connections. Based on what we described in the Methods section,
we estimated the entropy of the all stage subfigure of Figure 4 and its shuffled patterns for
significantly lower connections (blue-colored) and significantly greater connections (red-
colored) separately. Supporting Information Figure S3 shows that the pattern of blue-colored
connections and the pattern of red-colored connections have lower entropies compared to
shuffled patterns that states significant connections are not random.
Inoltre, a question has arisen whether high contributed and low contributed connec-
tions that are denoted in Figure 4 are within-network type or between-network type. To answer
this question and better interpret the heat maps of Figure 4, we created a metric and named it
WBR. WBR measures the ratio of within-network connectivity to between-network connectiv-
ità. As such, we first consider our interesting connections, Per esempio, higher contributed
connections, then count the number of appearing between-network connections and
within-network connections. On another side, we calculate the number of possible
between-network connections and within-network connections based on the number of
nodes. Finalmente, we divide the calculated values as below:
WBR ¼
(cid:4)
WC
WC0 =
BC
BC0
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
/
T
where WC and BC stand for the number of within-network connections and the number of
between-network connections, and WC0 and BC0 are also their maximum possible values.
WBR would be more than one in the case of intense within-network connections, lower than
one in the case of extreme between-network connections, and equal to one in the balance
between them. Tavolo 1 demonstrates WBR of high contributed connections and low contrib-
uted connections in frustration formation calculated based on heat maps of Figure 4. IL
results show that the low contributed connections tend to be within-network type and high
contributed connections are mostly between-network type. We also investigated the differ-
ence in the contribution of connections in frustration formation between life-span stages. So
we performed a multigroup Kruskal–Wallis test for each connection between observed con-
tribution values of different stages subjects. Then we corrected multiple comparison effects
on p values using the FDR method. We did not find any significant corrected p values with
large effect sizes (greater than 0.14) for various functional connections. Supporting
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 1. WBRs calculated based on low contributed and high contributed connections of Figure 4 heat maps
Functional connections
Low contributed (blue cells)
Childhood
3.962
Adolescence
4.239
High contributed (red cells)
0.507
0.718
Early adulthood Middle adulthood
4.163
0.615
4.579
0.273
Late adulthood
3.903
All stages
4.966
0.253
0.269
Network Neuroscience
1341
Pattern of frustration formation in the functional brain network
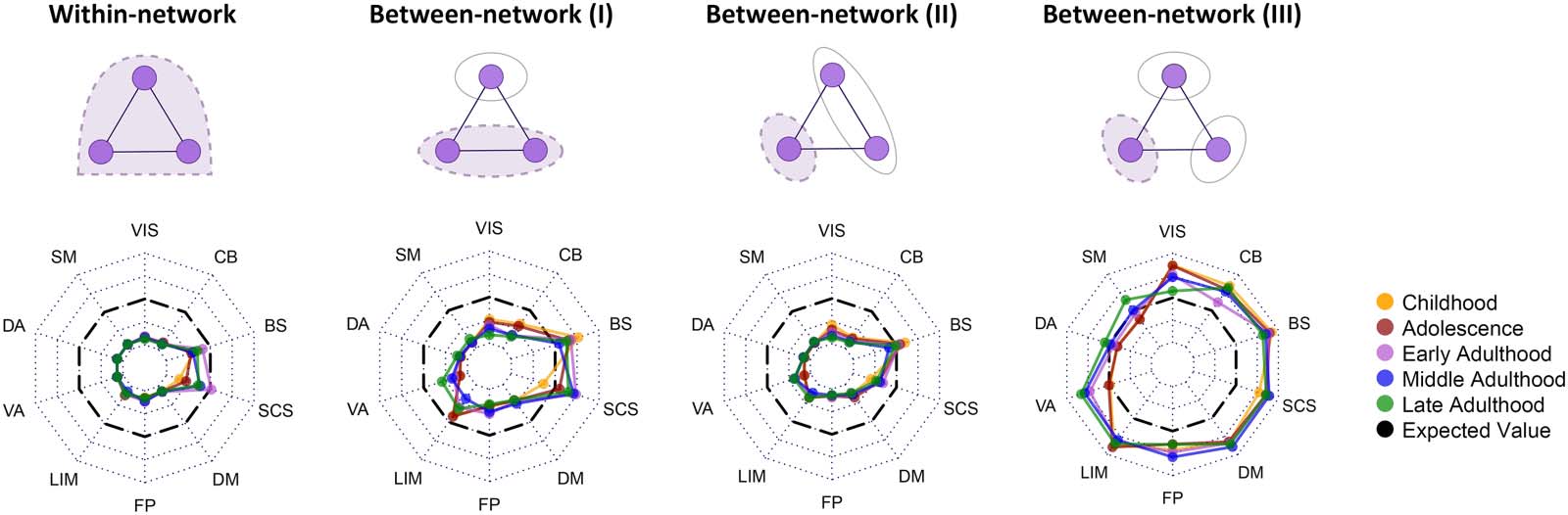
Figura 5. Contribution of canonical networks in frustration formation. The above row displays four possible frustration formation types
between regions of canonical networks, from left to right: all regions located in the network, two regions in the network and another outside,
one region in the network and two other regions in another network, one region in the network and other regions in two distinct networks.
Dashed violet binds denote the elements of interest in analysis. Radar charts correspond to every frustration formation type displayed above
them and show the difference between actual and expected contribution values. Dotted lines of radar charts segment r effect size from −1 to
1, where black dashed lines indicate zero r effect size. Line colors demonstrate different life-span stages. Effect size—small: |R | < 0.4;
medium: 0.4 < |r | < 0.6; large: |r | > 0.6. VIS = visual; SM = somatomotor; DA = dorsal attention; VA = ventral attention; LIM = limbic;
FP = fronto-parietal; DM = default mode; SCS = subcortical structures; BS = brain stem; CB = cerebellum.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
Information Figure S4 only shows connections with corrected p values lower than 0.05 con
medium effect sizes (between 0.06 E 0.14).
Contribution of Canonical Networks
In the last sections, we explored the involvement of brain regions and their functional connec-
tions in frustration formation. In this section, we wanted to study the contribution of canonical
networks. So we used the projection of Shen’s ROIs into Yeo’s networks (Figura 2 and Methods
section “Parcellation Atlas Projection”). There are four types of frustration formations that
regions of a canonical network can be involved in (Figura 5), (1) within-network: all three
regions belong to the target network; (2) between-network (IO): two regions located in the target
network and one another outside; (3) between-network (II): one region located in the target
network and two other regions in another network; E (4) between-network (III): one region
located in the target network and two other regions in two other distinct networks. For every
combination of formation type, stage, and canonical network, we performed a pairwise group
comparison using the Wilcoxon matched-pairs signed-rank test between actual and expected
contribution values of subjects and then integrated all results into Figure 5 and Supporting
Information Tables S1–S4. Formulas to calculate null contribution values of the target network
of T in mentioned frustration formation types are described as follows:
(cid:1) (cid:3)
3
NT
(cid:5) (cid:6) (cid:1) NFrus ðwithin(cid:3)networkÞ
3
N
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
T
/
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
where N and NT denote total ROI number and those ROI numbers located in canonical net-
work of T;
E
equal to the possible triangle numbers in whole-brain network and the
(cid:5) (cid:6)
3
N
(cid:1) (cid:3)
3
NT
possible triangle numbers in the target network. NFrus is also the number of appeared frustra-
tions in corresponding actual whole-brain network and we wanted to calculate its null values.
(cid:1) (cid:3)
2
NT
Þ
(cid:1) N − NT
ð
(cid:5) (cid:6)
3
N
(cid:1) NFrus ðbetween(cid:3)networkðIÞÞ
Network Neuroscience
1342
Pattern of frustration formation in the functional brain network
The numerator of the fraction is equaled to the number of triangles that their two ROIs located
in the target network of T and one another outside.
P
(cid:1) (cid:3)
i≠T NT (cid:1) 2
Ni
(cid:5) (cid:6)
3
N
(cid:1) NFrus ðbetween(cid:3)network ðIIÞÞ
The summation performs over all the networks except network of T, and the numerator of the
fraction is equaled to the number of triangles that their one ROI located in the target network
and two ROIs in another networks.
P
i≠j≠T NT (cid:1) Ni (cid:1) Nj
(cid:5) (cid:6)
3
N
(cid:1) NFrus ðbetween(cid:3)network ðIIIÞÞ
The summation performs over every distinct pair of networks except network of T. IL
numerator of fraction denotes possible triangles that one of their ROIs is located in the target
network and two other ROIs belong to two other distinct networks. After counting contribu-
tion values for formation types and calculating null values, we performed paired group anal-
ysis. All the effect sizes and corrected p values are reported in the Supporting Information.
Radar charts of Figure 5 demonstrate the r effect size of Wilcoxon matched-pairs signed-rank
tests between actual and null contribution values. The results demonstrate that all networks
in all of the stages except all stages’ brain stem and adults’ subcortical structures have sig-
nificantly lower within-network types of frustration formation with large effect sizes (greater
di 0.6). Visual, somatomotor, dorsal, and ventral attention, fronto-parietal, and default
mode networks have significantly lower contributions in between-network frustration forma-
tion type I, although their effect sizes are medium (between 0.4 E 0.6) in some stages.
Also, early adults’ limbic networks and adults’ cerebellums have significantly lower contri-
butions in between-network type I and large effect sizes. Subcortical structures in early and
middle adulthood and brain stem in childhood have significantly higher contributions in the
formation of between-network type I with medium effect sizes. Investigation of between-
network type II also shows that most of the networks in all stages have significantly lower
contributions in frustration formation with large effect sizes except brain stem and subcorti-
cal structures, where subcortical structures just in childhood, adolescence, and late adult-
hood have medium effect sizes. The last radar chart (between-network III) shows that most of
the higher contributions occur between three distinct networks. Although, it indicates that
somatomotor and dorsal attention are not contributed to this type of frustration formation.
Most other networks of all stages have significantly higher contributions in this type of frus-
tration formation with large or medium effect sizes except visual network in late adulthood,
ventral attention in childhood and adolescence, and fronto-parietal in late adulthood that
have no significant p values, and fronto-parietal in adolescence and cerebellum in early
adulthood that have small effect sizes. We also compared contribution values of different
stages for every pair of formation types and canonical networks using the Kruskal–Wallis test.
The test obtained no significant corrected p values with large effect sizes. P values and effect
sizes of these analyses are also presented in the Supporting Information.
Contribution of Hemispheres
In the end, we investigated the contribution of the brain hemispheres in frustration formation.
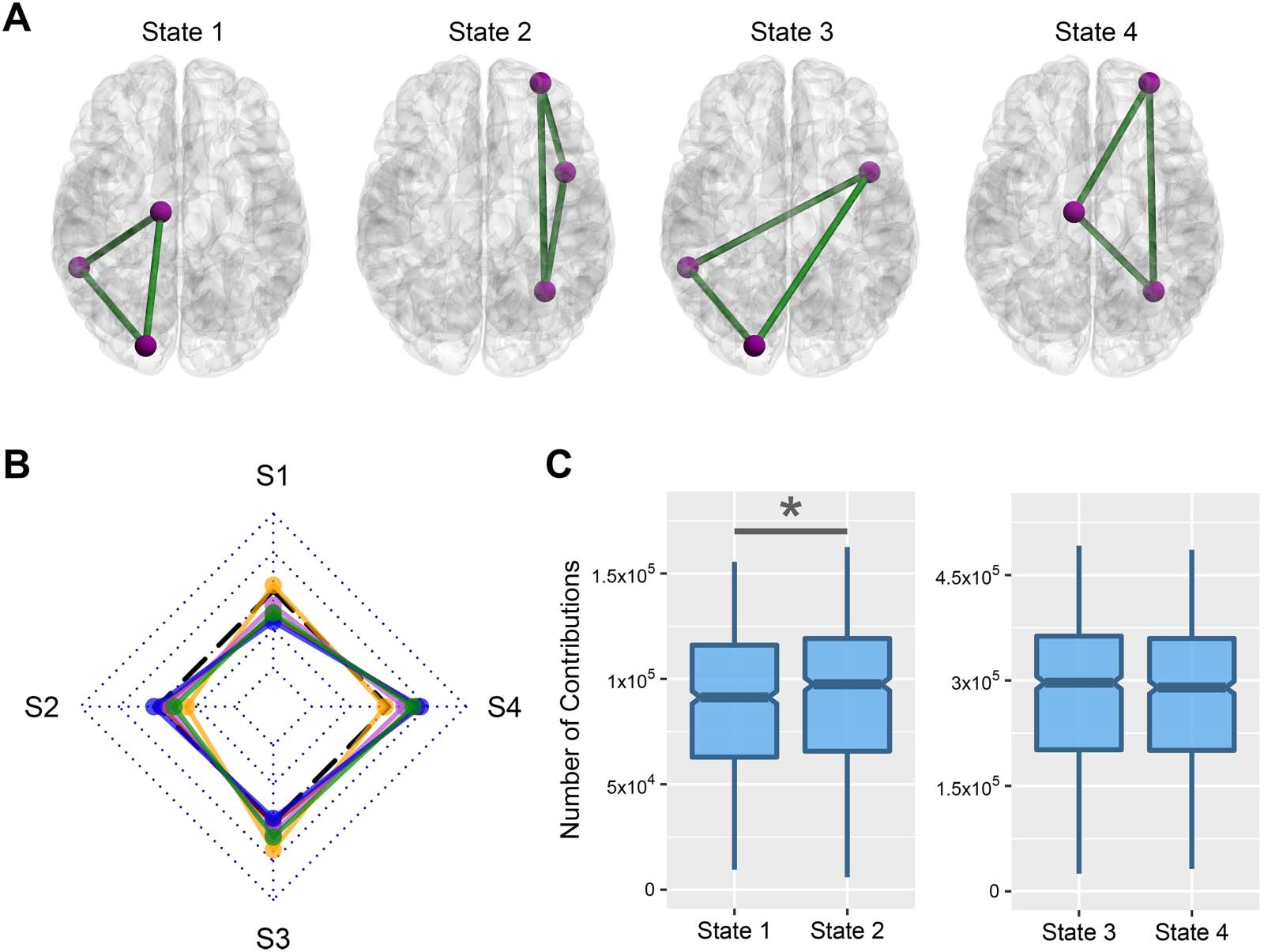
There are four possible states for the contribution (Figure 6A): (1) all three regions of frustration
belong to the right hemisphere, (2) all three regions belong to the left hemisphere, (3) only two
regions located in the right hemisphere, E (4) and only two regions located in the left hemi-
sphere. We compared actual and null contribution values for every combination of state and
Network Neuroscience
1343
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
/
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
/
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 6. Contribution of brain hemispheres in frustration formation. (UN) Four possible frustration formation types between brain hemispheres.
(B) The difference between actual and null contribution values of every stage and state. Dotted rhombuses segment r effect sizes from −1 to 1,
where black dashed lines denote zero value. S1–S4 corresponds to State 1–State 4. Lines colors—orange: childhood; red: adolescence; violet:
early adulthood; blue: middle adulthood; green: late adulthood. Effect size—small: |R | < 0.4; medium: 0.4 < |r | < 0.6; large: |r | > 0.6. (C)
Comparison between the contribution of hemispheres without considering the stage. Horizontal lines of boxes indicate medians, and notches
determine the 95% confidence interval for the medians. Asterisk (*) denotes the significant corrected p value with a small effect size.
stage using the Wilcoxon matched-pairs signed-rank test. We estimated the expected contri-
bution values of four states for a network as follows:
(cid:1)
(cid:3)
3
NLH
(cid:5) (cid:6) (cid:1) NFrus State 1
3
N
ð
Þ
(cid:1)
(cid:3)
3
NRH
(cid:5) (cid:6) (cid:1) NFrus State 2
Þ
3
N
ð
(cid:1)
(cid:3)
2
NLH
(cid:1)
(cid:3)
2
NRH
Þ
(cid:1) N − NLH
ð
(cid:5) (cid:6)
3
N
Þ
(cid:1) N − NRH
ð
(cid:5) (cid:6)
3
N
(cid:1) NFrus State 3
ð
Þ
(cid:1) NFrus State 4
ð
Þ
where NRH and NLH are the number of regions in right and left hemispheres that are equaled to
133 E 135 according to Shen’s atlas. Figure 6B shows the result of comparisons. Noi
reported their statistics in Supporting Information Table S5. The results show no significant
Network Neuroscience
1344
Pattern of frustration formation in the functional brain network
corrected p values with large effect size (greater than 0.6) for the comparisons. All effect sizes
were small except State 4 in middle adulthood that has medium effect sizes (r effect size =
0.408). We also compared contribution values of different stages in every state and did not
find any nonsmall effect sizes for between-stage differences. To investigate the main effect
of the hemisphere on frustration formation disregarding life-span stage, we also performed
two Wilcoxon matched-pairs signed-rank tests, one between State 1 and State 2 contribu-
tion values and the other between State 3 and State 4 contribution values (Figure 6C). Only
the comparison between State 1 and State 2 had a significant p value, although its effect
size was small (r effect size = −0.38). Generally, we did not find any robust hemispherical
effect on frustration formation.
DISCUSSION
We wanted to find out the role of different brain elements in frustration formation. Noi veniamo-
pared the number of frustrations that each element contributes with the null contribution value
of that element. The null values are estimated based on the hypothesis of the uniform contri-
bution of the elements in frustration formation. We performed the comparison in four levels of
the brain region, functional connection, canonical network, and hemisphere. We did it for
each life-span stage separately as well as without considering stages. We also compared the
involvement of different stages. Regional level comparisons show that some brain regions have
greater contributions, and some regions have lower contributions in frustration formation;
although, we did not find any robust differences between the regional contributions of different
stages (Figura 3). Investigation of functional connections also indicated that many functional
connections have significantly lower or greater contributions to frustration formation (Figura 4).
Low contributed connections are mostly within-network types and high contributed connec-
tions are usually the between-network type (Tavolo 1). We also did not find any robust variation
in the contribution of connections between stages. Inoltre, the results show that the most
significant regions and connections are lower contributed. Entropy analysis also indicated that
regional and connectional significant patterns are not random. In the following, we studied the
contribution of canonical networks in frustration formations. We found that most frustrations
appear between three regions of three distinct networks (Figura 5), and there are no powerful
differences between-network-based contributions of life-span stages. We also did not find any
strong hemisphere-related effects on frustration formation and no difference between the role
of the right and left hemispheres (Figura 6).
Role of Subcortical Structures
Our results suggest that subcortical regions have a prominent role in frustration formation in the
brain network at both nodal and connectional levels. So they can bring instability and altering
properties that facilitate systemic level neural changes and provide adaptive characteristics to
the brain. Certamente, we cannot understand brain reorganization without taking into account the
white matter tract located in subcortical areas. Older theories considered subcortical alterations
as a passive consequence of cortical functional reshaping; Tuttavia, new mechanisms under-
line the active reorganization of subcircuits within the large network of the brain (Duffau,
2009). Animal studies posited brain stem–related and subcortical plasticity under exposure
to visual and auditory stimuli (Chandrasekaran et al., 2014; Duménieu et al., 2021; Miranda
et al., 2014). Human studies reported such subcortical rewiring mechanisms for skill learning
such as second language and motor training (Liu et al., 2020; Sampaio-Baptista et al., 2013;
Scholz et al., 2009). A recent study also indicates subcortical short-term plasticity yielded by
deep brain stimulation pulse (Awad et al., 2021). All these observations are agreeing with
Network Neuroscience
1345
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
the frustrating essence of the brain subcortex. Inoltre, we know that many studies
reveal malfunction and abnormal structural changes in the subcortex of neural disorders
(Arnold Anteraper et al., 2014; Cerliani et al., 2015; Hoogman et al., 2017; Lee et al., 2018;
Rosenberg-Katz et al., 2016) while subcortical small changes may have extensive effects at the
cortical level (Jones, 2000). Therefore, controlling the activation of frustrating subcortical
regions can provide a favorable systemic reorganization of the brain network. It may be appli-
able by focusing on our detected frustrating regions and connections using sophisticated sub-
cortical stimulation methods (Awad et al., 2021; Colle et al., 2021; Folloni et al., 2019; Shi
et al., 2021). Inoltre, the prominent role of the subcortex in frustration formation may be
related to its unidirectional projections (Shi et al., 2014) or indirect pathways (Lanciego et al.,
2012), which need more exploration.
Between-Network Frustrations
The brain is a self-organized system (Dresp-Langley, 2020) such that functional signed links
adopt a topology to reduce network frustration (Saberi et al., 2021UN). We found that this
property is consistent within canonical networks as well, where canonical networks have
lower numbers of frustrations compared to null (Figura 5). Most of the frustrating connec-
tions are formed between networks (Tavolo 1), and frustrated triangles mostly engage three
distinct networks (Figura 5). This is what we expect to happen since large-scale brain net-
works consist of coactivating local brain seeds (Yeo et al., 2011). They decrease the chance
of negative functional connection appearance and subsequently frustration formation. Conse-
quently, if someone is interested in handling brain network frustration he/she should pursue a
multinetwork approach.
Visual Network Contribution
Our results indicate that most visual regions of adults have a low contribution to frustration
formation (Figura 3), which may be a consequence of visual maturation (Siu & Murphy,
2018). Although, several ROIs belonging to the hippocampus show frustrating connections
to most brain areas (Figura 4). It is an interesting result since the prominent role of the hippo-
campus in learning and memory and its association with broad areas of the brain to play this
role (Anand & Dhikav, 2012) are consistent with the frustrating essence of its connections. Noi
should mention that frustration facilitates adaptive processes due to their unstable nature.
Somatomotor Network Contribution
As Figure 3 shows, nearly 40% of somatomotor and ventral attention regions of adolescents
have a lower contribution to frustration formation than adults. We wonder why this happens
while neural flexibility is an essential developmental feature of adolescence. We could not
find a proper answer or any biological relevance for this effect.
Ventral Attention to Default Mode Connections
We also saw some ventral attention to default mode connections are frustrating (Figura 4). Questo
effect is enhanced in the adulthood range. We could not find a good psych-neural interpreta-
tion for this observation. Although, a recent paper highlighted connectivity between ventral
attention and default mode areas in Bulimia Nervosa (Domakonda et al., 2019). Because of
the high comorbidity of Bulimia Nervosa with other mental disorders such as depression and
bipolar (McElroy et al., 2005; Walsh et al., 1985) and the frequency of mood disorders in
Network Neuroscience
1346
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
adulthood range, we think the frustrating properties of these connections may be related to
these types of disorders.
Life-Span Comparisons
In one of our previous works, when we compared the number of frustrations between life-span
stages to track the requirement to change of brain network, we reported a significant difference
between stages (Saberi et al., 2021B). Because of the large sample size of stages, we decided to
consider effect size besides corrected p value in current work comparisons to specify more
robust differences. We did not find any strong differences between frustration formation pat-
terns of various stages; in other words, all significant comparisons with corrected p values
lower than 0.05 had corresponding small and medium effect sizes.
Shen’s Atlas to Yeo’s Atlas Projection
As we wanted to investigate the quality of canonical brain networks’ contribution to frustra-
tion formation, we needed projection of Shen’s 268 ROIs into them. Finn et al. (2015) utilized
Shen’s clustering algorithm (Shen et al., 2013) to group the ROIs into eight networks of the
medial frontal network, fronto-parietal network, default mode network, subcortical and cer-
ebellar regions, motor network, visual I network, visual II network, and the visual association
rete. The labeling is presented at https://bioimagesuiteweb.github.io/webapp/connviewer
.html and has been used for the network-based analysis of studies (Chen et al., 2020;
Rosenberg et al., 2016). When we explore the output of the categorization, we can observe
many implausible labeling, for instance: some superior regions labeled as subcortical and
cerebellar networks, some temporal regions categorized into medial frontal, and some cere-
bellar regions classified as fronto-parietal, visual, and default mode networks. Also, the limbic
rete, which is one of the most important functional brain subsystems, is not regarded.
Since three networks are nominated based on the sense of vision, a question has arisen: if
the sensory system has been considered a major element of the labeling, why is there no sign
of other sensations such as auditory in the ROIs categorization. As there seem to be some
shortages in the categorization process, we decided to use a new standard procedure (Lawrence
et al., 2021) to project Shen’s ROIs into Yeo’s seven large-scale brain networks (Yeo et al., 2011)
based on the coincidence index of the two atlases (Supporting Information Figure S5). Since Yeo
et al. (2011) excluded subcortical areas, we specified them into three distinct networks accord-
ing to their anatomical characteristics. We described the procedure and the intermediate results
in the Methods section “Parcellation Atlas Projection.” So our optimized model classified Shen’s
regions into 10 subnetworks of visual, somatomotor, dorsal attention, ventral attention, limbic,
fronto-parietal, default mode, subcortical structures, brain stem, and cerebellum (Figura 2).
Global Signal Effects
There is much conflictual evidence on the quality of removing global brain signals; some stud-
ies suggest this step in preprocessing and some others decline it (Aquino et al., 2020; Fox et al.,
2009; Liu et al., 2017B; Murphy et al., 2009; Schölvinck et al., 2010). It is clear that global
signal regression produces such antisynchronous connections with negative correlations (Fox
et al., 2009) (Supporting Information Figure S6) that matter for signed networks and can affect
the structural balance. It is also a fact that increasing negative links grows the number of frus-
trations since they have more negative links as compared to satisfaction (Figura 1). In the anal-
ysis of the main manuscript as well as our previous research (Saberi et al., 2021UN, 2021B), we
ignored global signal regression and explored the formation of frustration in the presence of
minor negative links. Besides, we performed the same analysis on the global signal regressed
Network Neuroscience
1347
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
functional images to check the difference. Supporting Information Figures S7 and S8 show the
result of the analysis for the contribution of regions and contribution of connections in frustra-
tion formation. Figure S7 shows a lower number of significant regions compared to Figure 3
and indicates some other regions. Low contributed regions belong to somatomotor and atten-
tional networks in adulthood and default mode in all stages, and high contributed regions are
mostly located in the subcortex in early adulthood and outspread in other stages. Investigation
of connection contribution on the regressed images (Supporting Information Figure S8) also
shows fewer numbers of significant connections compared to Figure 4, in which most of them
are lower contributed and within-network type. In summary, it seems that global signal regres-
sion gives little but different information about frustration formation. Inoltre, we did not
find any significant differences between frustration formation of life-span stages as well as
without the global signal regression approach.
Parcellation-Based Reliability
Many parcellation atlases were developed based on the anatomical and functional attributes
of the brain and using different algorithms. In connectivity studies where brain parcellations
are used, a question always arises about whether the results are reliable under other atlases
(Domhof et al., 2021; Popovych et al., 2021). Actually, we utilized Shen’s parcellation atlas
(Shen et al., 2013) and categorized its 268 regions into canonical networks (Yeo et al., 2011)
in our analysis. To check the mentioned reliability, we chose Desikan–Killiany–Tourville
(DKT) atlas (Klein & Tourville, 2012) as one of the most similar ones to Shen’s parcellation
con 101 cortical and subcortical ROIs, although it has different ROI boundaries and lower
resolution and was developed based on another algorithm. We also categorized DKT ROIs
into canonical networks based on the projection manner that we explained in the Methods
section and Supporting Information Figure S9. Since our analysis was focused on brain ele-
ments and considering elemental features of brain atlases are not the same in various atlases,
obtaining similar global features and moderate similar local features may be satisfying. Supporto-
ing Information Figure S10 shows regional contribution analysis for DKT ROIs and Figure S11
demonstrates that for corresponding functional connections. Both of them do not indicate any
significant differences between the contribution map of life-span stages, as the same results were
obtained based on Shen’s atlas. Also, subcortical regions and their connections with other brain
regions show high contributions to frustration formation that emphasizes the effective role of the
subcortex. Most low contributed connections are also within-network types and high contrib-
uted connections are almost between subcortical structures and other brain regions. Somatomo-
tor and visual areas also show a low contribution to frustration formation. Tuttavia, we can see
contradictory observations compared to Shen’s related results. Per esempio, a medial large
region of the default mode network shows a high contribution opposite to Shen’s results, E
the limbic network has some low contributed regions. The differences may be routed in the dis-
similarity of the atlases.
Neural Essence of Frustration
Frustration is formed by a combination of positive and negative links (Figura 1). Positive and
negative links refer to regional synchronous and regional antisynchronous coactivations,
rispettivamente. We do not know which mechanism is behind appearing negative links; maybe
it is related to time delay due to axonal propagation where influential subcortical regions that
are highly contributed to frustration formation have large axonal wiring (Petkoski & Jirsa, 2019;
Petkoski et al., 2018). It is an open question that needs more investigation, and computational
modeling can be helpful to solve it. Positive and negative links conceptually look the same as
Network Neuroscience
1348
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
/
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
inphase and antiphase synchronizes (Petkoski & Jirsa, 2019, 2022; Petkoski et al., 2016),
whereas Petkoski and his colleagues showed that inphase synchrony (or perfectly aligned
in/antiphase clustering) makes the lowest energy that is similar to a brain signed network that
has no negative links and frustrations located in the lowest balance energy. They also found
that when giving the distribution of the time delays in the brain, it is more probable that the
brain minimizes the disorders that are consistent with our previous results (Saberi et al., 2021UN)
and the self-organizing essence of the brain (Dresp-Langley, 2020). So we propose their com-
putational approach as a high-potential way to investigate the mechanism behind negative
link and frustration appearances.
Frustration as a Brain Network Measure
Some graph measures are defined based on the arrangement of graph links; Per esempio, we
can calculate the clustering of a network by counting triangles, and motifs as subgraphs are
considered the building blocks of the networks. They are commonly investigated in brain net-
works (Liu et al., 2017UN; Rubinov & Sporns, 2010) and extracted from connected-disconnected
graphs where either we have a link between two nodes or we do not have any. Although frus-
tration is defined in a signed network where presented links have two states of positive and
negative and gives information on the system disordering to us.
Disregarding Threshold on Connections
Thresholding is a common way of making a brain network (Garrison et al., 2015; Theis et al.,
2021; Wang et al., 2021). To provide a brain signed network, we claim that although a func-
tional brain network has a low number of antisynchronous coactivations (Supporting Informa-
tion Figure S6), the impact of positive and negative connections are not the same, so we
should not consider the same threshold for both of them, so disregarding the thresholding
process is a good way. Consequently, a question emerges that maybe most negative links
randomly appear. To answer it we checked the randomness of the contribution pattern
(Figures S2 and S3). Since we found that they are not random we can conclude that disregarding
the threshold is not vulnerable and negative connections’ appearance is meaningful.
Conclusione
In summary, many brain elements play an active role in the frustration formation of the brain
rete; Tuttavia, many other elements are less contributed to frustration formation. We iden-
tified both of them at the level of the node, connection, and network. The subcortical areas
and hippocampus are the most influential region for frustration formation. Matured visual
regions have less propensity to get involved in frustrations. Ventral attention to default mode
connections are frustrating, especially in adulthood. Generally, regional and connectional
contribution patterns are not random, and the frustrations are mainly formed between three
distinct networks. Also, there is no robust difference between the contribution pattern of brain
elements in frustration formation between life-span stages. The study of network frustration can
reveal the mechanisms behind neural alteration and brain disfunction. Localization of the frus-
trations also provides the possibility of brain network reorganization.
METHODS
Neuroimage Data
We collected functional and anatomical T1 images from two public repositories of ABIDE (Di
Martino et al., 2014) and Southwest (Wei et al., 2018). The Southwest database contains early
Network Neuroscience
1349
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
adulthood to late adulthood subjects, and ABIDE contains childhood to early adulthood sub-
jects. We selected all healthy subjects whose functional repetition times were equal to 2 sec-
onds (most frequent repetition times). Selected subjects aged from 6 A 80 (mean: 31.31; SD:
19.78) E 44% were female. We classified subjects into five life-span stages of childhood
(age: 6–12), adolescence (age: 12–18), early adulthood (age: 18–40), middle adulthood
(age: 40–65), and late adulthood (age: greater than 65) according to Erikson’s stages (Sharleen,
2013). After preprocessing, we also excluded subjects whose images could not pass the quality
check. Supporting Information Table S6 represents the demography of 793 finalized subjects
based on the stage and neuroimaging site. Table S7 also describes site-specific imaging pro-
tocols. We should mention that the neuroimaging procedures were carried out in compliance
with the Declaration of Helsinki. All adult subjects and parents (legal guardians) of subjects
under the age of 18 provided informed consent before starting the procedure. Neuroimages
are collected in several sites, and the acquisition protocols were approved by their licensing
committees including the Research Ethics Committee of the Brain Imaging Center of Southwest
Università, Institutional Review Boards of the New York University School of Medicine, IL
Institutional Review Board of San Diego State University, the Institutional Review Board of
University of Michigan, Yale University Institutional Review Board, Ethics Commission of
ETH Zurich, Georgetown University Institutional Review Board, Hospital of Trinity College,
and The Institutional Review Board of University of Utah School of Medicine.
Preprocessing Functional Images
We employed FSL (Jenkinson et al., 2012) and AFNI (Cox, 1996) to preprocess images. At first,
we extracted the brain tissue from the T1 image, then segmented it into gray matter (GM),
white matter ( WM), and cerebral spinal fluid (CSF). Then we removed the first five volumes
of the functional image to assure magnetic stability and then performed slice timing correction.
After that, we registered volumes of the functional image to the extracted brain of the T1 image
using the least square optimization with three translational and three rotational variables. Then
we conducted spatial smoothing on registered volumes using a Gaussian kernel (FWHM =
5 mm). In the following, we interpolated spiking outliers of every voxel’s time series and
applied band-pass filtering (0.01–0.09 Hz) to them to exclude nonrelevant information. We also
regressed out three translational and three rotational confounds of motions as well as WM and
CSF signals from the time series of every voxel. Finalmente, we normalized volumes of functional
images to MNI152 standard space (2 × 2 × 2 mm3) by optimization of 12 variables including 3
translational, 3 rotational, 3 scaling, E 3 shearing variables. We did not regress out the global
signal from functional image for analysis of main manuscripts, although we brought a version of
the results that regressed out global signal from the image in the Supporting Information. In
the end, we inspected the quality of preprocessing. So we excluded subjects whose images
had low extraction and registration quality and those with movement parameters greater
than one voxel size. It is beneficial to mention that we had used the procedure in other
studies (Saberi et al., 2021UN, 2021B; Sadeghi et al., 2017).
Regional Brain Activations
We used MATLAB software to extract 268 regional activity patterns from every preprocessed
functional image based on Shen’s atlas (Shen et al., 2013). Although all functional images
were acquired with the same repetition time, imaging sites had different acquisition times, vol-
ume numbers, and regional time points. The shortest acquisition time belongs to the George-
town University site of ABIDE with 147 volumes. Accordingly, we chose 147 first time points
of all regional time series. So we obtained 268 time series with 147 time points as activity
Network Neuroscience
1350
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
/
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
patterns for each subject. Since all repetition times were equal, adjusted activity patterns cor-
responded to an equal acquisition time. The equality of the number of time points and equality
of acquisition times matter in connectivity matrix formation and help to improve the validity of
the comparisons.
Frustration Formation
We constructed a connectivity matrix for each subject according to Pearson’s correlation of
pairs of regional time series. We only considered signs of correlation coefficients to obtain the
adjacency matrix of the subject’s signed networks denoting positive and negative links. Then
we identified the triadic frustrations of the subject’s networks. After identifying frustrations, we
counted the number of triadic frustrations that every element contributed to its formation. Noi
measured the contribution value for every nodal, connectional, network-based, and hemi-
sphere element. We explained the way to estimate the expected contribution values of the
elements in the Results section.
Structural Balance Theory
Attitude change is a social psychology topic that studies how people change their beliefs about
concepts and objects. Fritz Heider’s attitude change theory is known as the balance theory
(Heider, 1946), a triangle between two individuals and one object. It describes how the rela-
tionship between individuals affects the quality of individual attitude toward the object that we
consider trivial nowadays, Per esempio, business owners introduce their products by using
social influencers to better change the attitude of customers. Heider’s theory was extended
to interpersonal relations: when a friend’s friend or an enemy’s enemy is a friend, the triadic
relation is balanced; and when a friend’s friend or an enemy’s enemy is an enemy the triadic
relation is imbalanced (Rapoport, 1963). The imbalanced triads are nontrivial and unstable, so
their entities are frustrated about their condition and try to alter their relationships to become
balanced (Heider, 1946). Generally, components of a balanced triad are satisfied with their
situation, and components of an imbalanced triad are frustrated about their situation (Aref &
Wilson, 2019; Estrada, 2019). Also, Cartwright and Harary (1956) generalized Heider’s theory
to a wider range of interactions. They developed structural balance theory using mathematical
graphs. In this context, we can explore the balance of a system containing lots of positive and
negative interactions. In recent years, network scientists used the structural balance theory to
analyze various signed networks with a large set of entities and signed links (Anchuri &
Magdon-Ismail, 2012; Kirkley et al., 2019; Saberi et al., 2021UN; Zahedian et al., 2022). In this
context, we can define energy and phase states for the signed network, discriminate between
different state types, and investigate criticality and transitions between states (Antal et al., 2005;
Bagherikalhor et al., 2021; Derr et al., 2018; Marvel et al., 2009).
Parcellation Atlas Projection
As we wanted to study the contribution of the canonical networks in frustration formation, we
needed to know which Shen’s ROIs belong to which canonical networks. So we calculated the
Dice coefficient between any pair of Yeo’s networks and Shen’s ROIs as follows (Dice, 1945;
Lawrence et al., 2021):
Cij ¼ 2hij
ai þ bj
where ai and bj correspond to the total voxels in region i of Shen’s atlas and the total voxels in
network j of Yeo’s atlas, rispettivamente; hij also denotes the total number of overlapped voxels
Network Neuroscience
1351
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
N
e
N
_
UN
_
0
0
2
6
8
P
D
.
/
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Pattern of frustration formation in the functional brain network
between them. So we obtained a matrix where each cell indicates a Dice coefficient for a pair
of ROI and canonical network (Supporting Information Figure S5). According to the values, we
estimated that every ROI belongs to what network based on the largest coefficient. Since Yeo
only parcellated the cerebral cortex, we grouped subcortical ROIs into three networks of sub-
cortical structures, brain stem, and cerebellum based on their anatomical information. Figura 2
shows the result of ROI categorization. We attached labeled ROIs in the Supporting Informa-
zione. In the same way, we also categorized regions of DKT atlases (Figure S9) and provided
them to check reliability of results under changing parcellation.
Entropy Calculation
Entropy is a measure of system randomness. When a system has some possible microstates
with identical probabilities, Shannon’s entropy determines randomness of variable X as follows:
H Xð Þ ¼ −
Xn
i¼1
P xið
Þ logP xið
Þ
where summation applies on all states and P(xi) denotes the probability of the state xi. IL
entropy is maximum in the case of equality of states probabilities. We wanted to know whether
the appearance of significant nodes and connections in Figure 3 and Figure 4 are random or not.
So we calculated Shannon’s entropy for actual patterns and compared them with the entropy of
1,000 shuffled patterns. In this regard, we measured the entropy of regional patterns as follows.
Firstly, we considered canonical networks as states, counted the number of significant regions
located in each network, normalized counted numbers to bin size (total number of ROIs located
in networks), then divided the outputs by the total number of significant ROIs. In this way, we
provided probabilities of states and calculated Shannon’s entropy for the apparent pattern of
significant brain regions (Figure S2). Allo stesso modo, we measured Shannon’s entropy for the pattern
of significant brain connections, but we regarded connections between networks as states
(Figure S3).
Statistical Analysis
In this study, we had two types of comparison for each element: the two-group paired com-
parison between actual and null contribution values, and the multiple-group comparison
between contribution values of life-span stages. We used Wilcoxon matched-pairs signed-rank
test for the first one and the Kruskal–Wallis test for the last one. We utilized nonparametric
statistical analysis because contribution values were not distributed normally. We performed
multiple comparison corrections on p values using FDR and considered effect sizes to improve
the validity of the statistical analysis. We also used two different algorithms for calculating
effect sizes of between-group and multiple-group analyses with different thresholds (Tavolo 2).
Also, we calculated the p values of entropy analysis based on the null hypothesis of “entropy
of actual pattern is larger than entropies of shuffled patterns.” We should mention that we car-
ried out all statistical analyses in R software (Kassambara, 2020; Mangiafico & Mangiafico,
2017; RC Team, 2013; Whitcher et al., 2013). We provided Figure 1 using “draw.io” and
Tavolo 2.
Effect size thresholds and corresponding algorithms
Nonparametric test
Paired two-group
Effect size algorithm
Wilcoxonpairedr
R package
R companian
Small
0.1 – <0.4
Medium
0.4 – <0.6
Multiple-group
Kruskal_effsize
Rstatix
0.01 – <0.06
0.06 – <0.14
Network Neuroscience
Large
≥0.6
≥0.14
1352
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
n
e
n
_
a
_
0
0
2
6
8
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pattern of frustration formation in the functional brain network
statistical figures and brain maps by the advance of “BrainNet Viewer” (Xia et al., 2013) and
some other R packages (Nakazawa, 2019; Wickham et al., 2016). In addition, we shared all of
the information and codes at https://github.com/majidsaberi/ BrainNetFrustration (Saberi,
2022) so everyone can publicly access, replicate, and develop our research.
ACKNOWLEDGMENTS
We would like to thank ABIDE and Southwest University for generously sharing the data and
David Matthews for reading the manuscript and language checking.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00268 and
https://github.com/majidsaberi/BrainNetFrustration.
AUTHOR CONTRIBUTIONS
Majid Saberi: Conceptualization; Formal analysis; Methodology; Project administration;
Writing – original draft. Reza Khosrowabadi: Validation; Visualization; Writing – review &
editing. Ali Khatibi: Validation; Visualization; Writing – review & editing. Bratislav Mišić:
Validation; Visualization; Writing – review & editing. Gholamreza Jafari: Validation; Visualiza-
tion; Writing – review & editing.
REFERENCES
Alabandi, G., Tešić, J., Rusnak, L., & Burtscher, M. (2021). Discov-
ering and balancing fundamental cycles in large signed graphs.
In Proceedings of the International Conference for High Perfor-
mance Computing, Networking, Storage and Analysis (pp. 1–17).
https://doi.org/10.1145/3458817.3476153
Anand, K. S., & Dhikav, V. (2012). Hippocampus in health and disease:
An overview. Annals of Indian Academy of Neurology, 15(4), 239–246.
https://doi.org/10.4103/0972-2327.104323, PubMed: 23349586
Anchuri, P., & Magdon-Ismail, M. (2012). Communities and bal-
ance in signed networks: A spectral approach. In 2012
IEEE/ACM International Conference on Advances in Social Net-
works Analysis and Mining (pp. 235–242). IEEE. https://doi.org
/10.1109/ASONAM.2012.48
Antal, T., Krapivsky, P. L., & Redner, S. (2005). Dynamics of social
balance on networks. Physical Review E, 72(3), 036121. https://
doi.org/10.1103/PhysRevE.72.036121, PubMed: 16241529
Aquino, K. M., Fulcher, B. D., Parkes, L., Sabaroedin, K., & Fornito,
A. (2020). Identifying and removing widespread signal deflec-
tions from fMRI data: Rethinking the global signal regression
problem. NeuroImage, 212, 116614. https://doi.org/10.1016/j
.neuroimage.2020.116614, PubMed: 32084564
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aref, S., & Wilson, M. C. (2019). Balance and frustration in signed
networks. Journal of Complex Networks, 7(2), 163–189. https://
doi.org/10.1093/comnet/cny015
Arnold Anteraper, S., Triantafyllou, C., Sawyer, A. T., Hofmann,
S. G., Gabrieli, J. D., & Whitfield-Gabrieli, S. (2014). Hyper-con-
nectivity of subcortical resting-state networks in social anxiety
disorder. Brain Connectivity, 4(2), 81–90. https://doi.org/10
.1089/brain.2013.0180, PubMed: 24279709
Awad, M. Z., Vaden, R. J., Irwin, Z. T., Gonzalez, C. L., Black, S.,
Nakhmani, A., … Walker, H. C. (2021). Subcortical short-term
plasticity elicited by deep brain stimulation. Annals of Clinical
and Translational Neurology, 8(5), 1010–1023. https://doi.org
/10.1002/acn3.51275, PubMed: 33826240
Bagherikalhor, M., Kargaran, A., Shirazi, A. H., & Jafari, G. R.
(2021). Heider balance under disordered triadic interactions.
Physical Review E, 103(3), 032305. https://doi.org/10.1103
/PhysRevE.103.032305, PubMed: 33862685
Bassett, D. S., & Bullmore, E. T. (2009). Human brain networks
in health and disease. Current Opinion in Neurology, 22(4),
340–347. https://doi.org/10.1097/ WCO.0b013e32832d93dd,
PubMed: 19494774
Network Neuroscience
1353
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
n
e
n
_
a
_
0
0
2
6
8
p
d
t
/
.
Pattern of frustration formation in the functional brain network
Bassett, D. S., & Sporns, O. (2017). Network neuroscience. Nature
Neuroscience, 20(3), 353–364. https://doi.org/10.1038/nn.4502,
PubMed: 28230844
Cartwright, D., & Harary, F. (1956). Structural balance: A generali-
zation of Heider’s theory. Psychological Review, 63(5), 277–293.
https://doi.org/10.1037/h0046049, PubMed: 13359597
Cerliani, L., Mennes, M., Thomas, R. M., Di Martino, A., Thioux, M.,
& Keysers, C. (2015). Increased functional connectivity between
subcortical and cortical resting-state networks in autism spec-
trum disorder. JAMA Psychiatry, 72(8), 767–777. https://doi
.org/10.1001/jamapsychiatry.2015.0101, PubMed: 26061743
Chandrasekaran, B., Skoe, E., & Kraus, N. (2014). An integrative model
of subcortical auditory plasticity. Brain Topography, 27(4), 539–552.
https://doi.org/10.1007/s10548-013-0323-9, PubMed: 24150692
Chen, Y.-L., Tu, P.-C., Huang, T.-H., Bai, Y.-M., Su, T.-P., Chen, M.-H.,
& Wu , Y. - T.
( 2 0 2 0 ) . U s i n g m i n i m a l - r e d u n d a n t a n d
maximal-relevant whole-brain functional connectivity to classify
bipolar disorder. Frontiers in Neuroscience, 14, 563368. https://
doi.org/10.3389/fnins.2020.563368, PubMed: 33192250
Colle, H., Colle, D., Noens, B., Dhaen, B., Alessi, G., Muller, P., …
van der Linden, C. (2021). Subcortical stimulation with tip of
ultrasound aspirator. Journal of Neurological Surgery Part A: Cen-
tral European Neurosurgery, 82(6), 581–584. https://doi.org/10
.1055/s-0039-1691824, PubMed: 34433223
Cox, R. W. (1996). AFNI: Software for analysis and visualization of
functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29(3), 162–173. https://doi.org/10.1006
/cbmr.1996.0014, PubMed: 8812068
Derr, T., Aggarwal, C., & Tang, J. (2018). Signed network modeling
based on structural balance theory. In Proceedings of the 27th
ACM International Conference on Information and Knowledge Man-
agement (pp. 557–566). https://doi.org/10.1145/3269206.3271746
Di Martino, A., Yan, C. G., Li, Q., Denio, E., Castellanos, F. X.,
Alaerts, K., … Milham, M. P. (2014). The autism brain imaging data
exchange: Towards a large-scale evaluation of the intrinsic brain
architecture in autism. Molecular Psychiatry, 19(6), 659–667.
https://doi.org/10.1038/mp.2013.78, PubMed: 23774715
Dice, L. R. (1945). Measures of the amount of ecologic association
between species. Ecology, 26(3), 297–302. https://doi.org/10
.2307/1932409
Domakonda, M. J., He, X., Lee, S., Cyr, M., & Marsh, R. (2019).
Increased functional connectivity between ventral attention and
default mode networks in adolescents with bulimia nervosa.
Journal of the American Academy of Child & Adolescent Psychi-
atry, 58(2), 232–241. https://doi.org/10.1016/j.jaac.2018.09.433,
PubMed: 30738550
Domhof, J. W., Jung, K., Eickhoff, S. B., & Popovych, O. V. (2021).
Parcellation-induced variation of empirical and simulated brain
connectomes at group and subject levels. Network Neurosci-
ence, 5(3), 798–830. https://doi.org/10.1162/netn_a_00202,
PubMed: 34746628
Dresp-Langley, B. (2020). Seven properties of self-organization in
the human brain. Big Data and Cognitive Computing, 4(2), 10.
https://doi.org/10.3390/bdcc4020010
Duffau, H. (2009). Does post-lesional subcortical plasticity exist in
the human brain? Neuroscience Research, 65(2), 131–135.
https://doi.org/10.1016/j.neures.2009.07.002, PubMed: 19616045
Duménieu, M., Marquèze-Pouey, B., Russier, M., & Debanne, D.
(2021). Mechanisms of plasticity in subcortical visual areas.
Cells, 10(11), 3162. https://doi.org/10.3390/cells10113162,
PubMed: 34831385
Estrada, E. (2019). Rethinking structural balance in signed social
networks. Discrete Applied Mathematics, 268, 70–90. https://
doi.org/10.1016/j.dam.2019.04.019
Facchetti, G., Iacono, G., & Altafini, C. (2011). Computing global
structural balance in large-scale signed social networks. Pro-
ceedings of the National Academy of Sciences, 108(52),
20953–20958. https://doi.org/10.1073/pnas.1109521108,
PubMed: 22167802
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., … Constable, R. T. (2015). Functional connectome
fingerprinting: Identifying individuals using patterns of brain con-
nectivity. Nature Neuroscience, 18(11), 1664–1671. https://doi
.org/10.1038/nn.4135, PubMed: 26457551
Folloni, D., Verhagen, L., Mars, R. B., Fouragnan, E., Constans, C.,
Aubry, J. F., … Sallet, J. (2019). Manipulation of subcortical and
deep cortical activity in the primate brain using transcranial
focused ultrasound stimulation. Neuron, 101(6), 1109–1116.
https://doi.org/10.1016/j.neuron.2019.01.019, PubMed:
30765166
Fox, M. D., Zhang, D., Snyder, A. Z., & Raichle, M. E. (2009). The
global signal and observed anticorrelated resting state brain net-
works. Journal of Neurophysiology, 101(6), 3270–3283. https://
doi.org/10.1152/jn.90777.2008, PubMed: 19339462
Garrison, K. A., Scheinost, D., Finn, E. S., Shen, X., & Constable,
R. T. (2015). The (in)stability of functional brain network mea-
sures across thresholds. NeuroImage, 118, 651–661. https://
doi.org/10.1016/j.neuroimage.2015.05.046, PubMed:
26021218
Goremychkin, E. A., Osborn, R., Rainford, B. D., Macaluso, R. T.,
Adroja, D. T., & Koza, M. (2008). Spin-glass order induced by
dynamic frustration. Nature Physics, 4(10), 766–770. https://doi
.org/10.1038/nphys1028
Heider, F. (1946). Attitudes and cognitive organization. The Journal
of Psychology, 21(1), 107–112. https://doi.org/10.1080
/00223980.1946.9917275, PubMed: 21010780
Hoogman, M., Bralten, J., Hibar, D. P., Mennes, M., Zwiers, M. P.,
Schweren, L. S., … Franke, B. (2017). Subcortical brain volume
differences in participants with attention deficit hyperactivity dis-
order in children and adults: A cross-sectional mega-analysis.
The Lancet Psychiatry, 4(4), 310–319. https://doi.org/10.1016
/S2215-0366(17)30049-4, PubMed: 28219628
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W., &
Smith, S. M. (2012). FSL. NeuroImage, 62(2), 782–790. https://
doi.org/10.1016/j.neuroimage.2011.09.015, PubMed: 21979382
Jones, E. G. (2000). Cortical and subcortical contributions to
activity-dependent plasticity in primate somatosensory cortex.
Annual Review of Neuroscience, 23(1), 1–37. https://doi.org/10
.1146/annurev.neuro.23.1.1, PubMed: 10845057
Kassambara, A. (2020). rstatix: Pipe-friendly framework for basic
statistical tests. R package version 0.6.0.
Kirkley, A., Cantwell, G. T., & Newman, M. E. (2019). Balance in
signed networks. Physical Review E, 99(1), 012320. https://doi
.org/10.1103/PhysRevE.99.012320, PubMed: 30780212
Network Neuroscience
1354
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
n
e
n
_
a
_
0
0
2
6
8
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pattern of frustration formation in the functional brain network
Klein, A., & Tourville, J. (2012). 101 labeled brain images and a
consistent human cortical labeling protocol. Frontiers in Neuro-
science, 6, 171. https://doi.org/10.3389/fnins.2012.00171,
PubMed: 23227001
Lanciego, J. L., Luquin, N., & Obeso, J. A. (2012). Functional neu-
roanatomy of the basal ganglia. Cold Spring Harbor Perspectives
in Medicine, 2(12), a009621. https://doi.org/10.1101
/cshperspect.a009621, PubMed: 23071379
Lawrence, R. M., Bridgeford, E. W., Myers, P. E., Arvapalli, G. C.,
Ramachandran, S. C., Pisner, D. A., … Vogelstein, J. T. (2021).
Standardizing human brain parcellations. Scientific Data, 8(1),
1–9. https://doi.org/10.1038/s41597-021-00849-3, PubMed:
33686079
Lee, Y. J. G., Kim, S., Kim, N., Choi, J. W., Park, J., Kim, S. J., …
Lee, Y. J. (2018). Changes in subcortical resting-state functional
connectivity in patients with psychophysiological insomnia
after cognitive–behavioral therapy. NeuroImage: Clinical, 17,
115–1 23. https://doi.org/10.1016/j.nicl.2017.1 0.013,
PubMed: 29062688
Liu, D., Wang, S., Gao, Q., Dong, R., Fu, X., Pugh, E., & Hu, J.
(2020). Learning a second language in adulthood changes sub-
cortical neural encoding. Neural Plasticity, 2020, 8836161.
https://doi.org/10.1155/2020/8836161, PubMed: 33144853
Liu, J., Li, M., Pan, Y., Lan, W., Zheng, R., Wu, F. X., & Wang, J.
(2017a). Complex brain network analysis and its applications
to brain disorders: A survey. Complexity, 2017, 8362741.
https://doi.org/10.1155/2017/8362741
Liu, T. T., Nalci, A., & Falahpour, M. (2017b). The global signal in
fMRI: Nuisance or information? NeuroImage, 150, 213–229.
https://doi.org/10.1016/j.neuroimage.2017.02.036, PubMed:
28213118
Mangiafico, S., & Mangiafico, M. S. (2017). Package ‘rcompanion’.
CRAN Repos, 20, 1–71.
McElroy, S. L., Kotwal, R., Keck, Jr., P. E., & Akiskal, H. S. (2005).
Comorbidity of bipolar and eating disorders: Distinct or related
disorders with shared dysregulations? Journal of Affective Disor-
ders, 86(2–3), 107–127. https://doi.org/10.1016/j.jad.2004.11
.008, PubMed: 15935230
Marvel, S. A., Strogatz, S. H., & Kleinberg, J. M. (2009). Energy
landscape of social balance. Physical Review Letters, 103(19),
198701. https://doi.org/10.1103/ PhysRevLett.103.198701,
PubMed: 20365960
Miranda, J. A., Shepard, K. N., McClintock, S. K., & Liu, R. C.
(2014). Adult plasticity in the subcortical auditory pathway of
the maternal mouse. PLoS One, 9(7), e101630. https://doi.org
/10.1371/journal.pone.0101630, PubMed: 24992362
Mišić, B., & Sporns, O. (2016). From regions to connections and
networks: New bridges between brain and behavior. Current
Opinion in Neurobiology, 40, 1–7. https://doi.org/10.1016/j
.conb.2016.05.003, PubMed: 27209150
Murphy, K., Birn, R. M., Handwerker, D. A., Jones, T. B., & Bandettini,
P. A. (2009). The impact of global signal regression on resting state
correlations: Are anti-correlated networks introduced? Neuro-
Image, 44(3), 893–905. https://doi.org/10.1016/j.neuroimage
.2008.09.036, PubMed: 18976716
Nakazawa, M. (2019). Package ‘fmsb’. https://cran.r-project.org
/web/packages/fmsb/fmsb.pdf.
Petkoski, S., & Jirsa, V. K. (2019). Transmission time delays organize
the brain network synchronization. Philosophical Transactions of
the Royal Society A, 377(2153), 20180132. https://doi.org/10
.1098/rsta.2018.0132, PubMed: 31329065
Petkoski, S., & Jirsa, V. K. (2022). Normalizing the brain connec-
tome for communication through synchronization. Network
Neuroscience, 6(3), 722–744. https://doi.org/10.1162/netn_a
_00231
Petkoski, S., Palva, J. M., & Jirsa, V. K. (2018). Phase-lags in large
scale brain synchronization: Methodological considerations and
in-silico analysis. PLoS Computational Biology, 14(7), e1006160.
https://doi.org/10.1371/journal.pcbi.1006160, PubMed:
29990339
Petkoski, S., Spiegler, A., Proix, T., Aram, P., Temprado, J. J., & Jirsa,
V. K. (2016). Heterogeneity of time delays determines synchroni-
zation of coupled oscillators. Physical Review E, 94(1), 012209.
https://doi.org/10.1103/ PhysRevE.94.012209, PubMed:
27575125
Popovych, O. V., Jung, K., Manos, T., Diaz-Pier, S., Hoffstaedter, F.,
Schreiber, J., … Eickhoff, S. B. (2021). Inter-subject and
inter-parcellation variability of resting-state whole-brain dynam-
ical modeling. NeuroImage, 236, 118201. https://doi.org/10
.1016/j.neuroimage.2021.118201, PubMed: 34033913
Rapoport, A. (1963). Mathematical models of social interac-
tion. In D. Luce (Ed.), Handbook of mathematical psychology
(pp. 492—493). New York, NY: John Wiley & Sons.
RC Team. (2013). R: A language and environment for statistical
computing. Vienna, Austria: R Foundation for Statistical
Computing.
Rosenberg, M. D., Finn, E. S., Scheinost, D., Papademetris, X.,
Shen, X., Constable, R. T., & Chun, M. M. (2016). A neuromarker
of sustained attention from whole-brain functional connectivity.
Nature Neuroscience, 19(1), 165–171. https://doi.org/10.1038/nn
.4179, PubMed: 26595653
Rosenberg-Katz, K., Herman, T., Jacob, Y., Kliper, E., Giladi, N., &
Hausdorff, J. M. (2016). Subcortical volumes differ in Parkinson’s
disease motor subtypes: New insights into the pathophysiol-
ogy of disparate symptoms. Frontiers in Human Neuroscience,
10, 356. https://doi.org/10.3389/fnhum.2016.00356, PubMed:
27462214
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Saberi, M. (2022). BrainNetFrustration, GitHub, https://github.com
/majidsaberi/BrainNetFrustration.
Saberi, M., Khosrowabadi, R., Khatibi, A., Misic, B., & Jafari, G.
(2021a). Topological impact of negative links on the stability of
resting-state brain network. Scientific Reports, 11(1), 1–14.
https://doi.org/10.1038/s41598-021-81767-7, PubMed: 33500525
Saberi, M., Khosrowabadi, R., Khatibi, A., Misic, B., & Jafari, G.
(2021b). Requirement to change of functional brain network
across the lifespan. PLoS One, 16(11), e0260091. https://doi
.org/10.1371/journal.pone.0260091, PubMed: 34793536
Sadeghi, M., Khosrowabadi, R., Bakouie, F., Mahdavi, H., Eslahchi,
C., & Pouretemad, H. (2017). Screening of autism based on
task-free fMRI using graph theoretical approach. Psychiatry
Network Neuroscience
1355
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
n
e
n
_
a
_
0
0
2
6
8
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pattern of frustration formation in the functional brain network
Research: Neuroimaging, 263, 48–56. https://doi.org/10.1016/j
.pscychresns.2017.02.004, PubMed: 28324694
Sampaio-Baptista, C., Khrapitchev, A. A., Foxley, S., Schlagheck, T.,
Scholz, J., Jbabdi, S., … Johansen-Berg, H. (2013). Motor skill
learning induces changes in white matter microstructure and
myelination. Journal of Neuroscience, 33(50), 19499–19503.
https://doi.org/10.1523/ JNEUROSCI.3048-13.2013, PubMed:
24336716
Schölvinck, M. L., Maier, A., Frank, Q. Y., Duyn, J. H., & Leopold,
D. A. (2010). Neural basis of global resting-state fMRI activity.
Proceedings of the National Academy of Sciences, 107(22),
10238–10243. https://doi.org/10.1073/pnas.0913110107,
PubMed: 20439733
Scholz, J., Klein, M. C., Behrens, T. E., & Johansen-Berg, H. (2009).
Training induces changes in white-matter architecture. Nature
Neuroscience, 12(11), 1370–1371. https://doi.org/10.1038/nn
.2412, PubMed: 19820707
Sharleen, L. K. (2013). Lifespan development. Tinley Park, IL:
Goodheart-Wilcox.
Shen, X., Tokoglu, F., Papademetris, X., & Constable, R. T. (2013).
Groupwise whole-brain parcellation from resting-state fMRI data
for network node identification. NeuroImage, 82, 403–415.
https://doi.org/10.1016/j.neuroimage.2013.05.081, PubMed:
23747961
Shi, S., Xu, A. G., Rui, Y. Y., Zhang, X., Romanski, L. M., Gothard,
K. M., & Roe, A. W. (2021). Infrared neural stimulation with 7T
fMRI: A rapid in vivo method for mapping cortical connections of
primate amygdala. NeuroImage, 231, 117818. https://doi.org/10
.1016/j.neuroimage.2021.117818, PubMed: 33548458
Shi, Y., Ikrar, T., Olivas, N. D., & Xu, X. (2014). Bidirectional global
spontaneous network activity precedes the canonical unidirec-
tional circuit organization in the developing hippocampus. Jour-
nal of Comparative Neurology, 522(9), 2191–2208. https://doi
.org/10.1002/cne.23528, PubMed: 24357090
Siu, C. R., & Murphy, K. M. (2018). The development of human visual
cortex and clinical implications. Eye and Brain, 10, 25–36. https://
doi.org/10.2147/EB.S130893, PubMed: 29760575
Tang, J., Chang, Y., Aggarwal, C., & Liu, H. (2016). A survey of
signed network mining in social media. ACM Computing Sur-
veys, 49(3), 1–37. https://doi.org/10.1145/2956185
Theis, N., Rubin, J., Cape, J., Iyengar, S., Gur, R. E., Gur, R. C., …
Prasad, K. M. (2021). Evaluating network threshold selection for
structural and functional brain connectomes. bioRxiv. https://doi
.org/10.1101/2021.10.09.463759
Tomasso, M., Rusnak, L. J., & Tešić, J. (2022). Advances in scaling
community discovery methods for signed graph networks. Jour-
nal of Complex Networks, 10(3), cnac013. https://doi.org/10
.1093/comnet/cnac013
Toulouse, G. (1987). Theory of the frustration effect in spin glasses:
I. In M. Mézard, G. Parisi, & M. Á. Virasoro (Eds.), Spin glass the-
ory and beyond: An introduction to the replica method and its
applications. Teaneck, NJ: World Scientific. https://doi.org/10
.1142/9789812799371_0009
Vannimenus, J., & Toulouse, G. (1977). Theory of the frustration
effect. II. Ising spins on a square lattice. Journal of Physics C:
Solid State Physics, 10(18), L537. https://doi.org/10.1088/0022
-3719/10/18/008
Villain, J., Bidaux, R., Carton, J. P., & Conte, R. (1980). Order as an
effect of disorder. Journal de Physique, 41(11), 1263–1272.
https://doi.org/10.1051/jphys:0198000410110126300
Walsh, B. T., Roose, S. P., Glassman, A. H., Gladis, M., & Sadik, C.
(1985). Bulimia and depression. Psychosomatic Medicine, 47,
123–131. https://doi.org/10.1097/00006842-198503000-00003,
PubMed: 3863157
Wang, Z., Jie, B., Feng, C., Wang, T., Bian, W., Ding, T. X., … Liu, M.
(2021). Distribution-guided network thresholding for functional con-
nectivity analysis in fMRI-based brain disorder identification. IEEE
Journal of Biomedical and Health Informatics, 26(4), 1602–1613.
https://doi.org/10.1109/JBHI.2021.3107305, PubMed: 34428167
Wei, D., Zhuang, K., Ai, L., Chen, Q., Yang, W., Liu, W., … Qiu, J.
(2018). Structural and functional brain scans from the
cross-sectional Southwest University adult lifespan dataset. Sci-
entific Data, 5(1), 1–10. https://doi.org/10.1038/sdata.2018.134,
PubMed: 30015807
Whitcher, B., Schmid, V., Thornton, A., Tabelow, K., Clayden, J., &
Muschelli, J. (2013). Package ‘oro. nifti’. https://CRAN.R-project
.org/package=oro.nifti
Wickham, H., Chang, W., Henry, L., Pedersen, T. L., Takahashi, K.,
Wilke, C., … Dunnington, D. (2016). Package ‘ggplot2’. Create
elegant data visualisations using the grammar of graphics.
https://CRAN.R-project.org/package=ggplot2
Winkler, M., & Reichardt, J. (2013). Motifs in triadic random graphs
based on Steiner triple systems. Physical Review E, 88(2),
022805. https://doi.org/10.1103/PhysRevE.88.022805, PubMed:
24032881
Xia, M., Wang, J., & He, Y. (2013). BrainNet Viewer: A network
visualization tool for human brain connectomics. PLoS One,
8(7), e68910. https://doi.org/10.1371/journal.pone.0068910,
PubMed: 23861951
Yang, B., Cheung, W., & Liu, J. (2007). Community mining from
signed social networks. IEEE Transactions on Knowledge and
Data Engineering, 19(10), 1333–1348. https://doi.org/10.1109
/TKDE.2007.1061
Yang, S.-H., Smola, A. J., Long, B., Zha, H., & Chang, Y. (2012).
Friend or frenemy? Predicting signed ties in social networks.
In Proceedings of the 35th international ACM SIGIR confer-
ence on research and development in information retrieval
(pp. 555–564). https://doi.org/10.1145/2348283.2348359
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., … Buckner, R. L. (2011). The organization of the
human cerebral cortex estimated by intrinsic functional connec-
tivity. Journal of Neurophysiology, 106(3), 1125–1165. https://doi
.org/10.1152/jn.00338.2011, PubMed: 21653723
Zahedian, M., Bagherikalhor, M., Trufanov, A., & Jafari, G. R. (2022).
Financial crisis in the framework of non-zero temperature balance
theory. arXiv:2202.03198. https://doi.org/10.48550/arXiv.2202.03198
Zhao, H., Zhang, J., Lyu, M., Bachus, S., Tokiwa, Y., Gegenwart, P.,
… Sun, P. (2019). Quantum-critical phase from frustrated mag-
netism in a strongly correlated metal. Nature Physics, 15(12),
1261–1266. https://doi.org/10.1038/s41567-019-0666-6
Network Neuroscience
1356
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
3
4
2
0
5
9
8
1
7
n
e
n
_
a
_
0
0
2
6
8
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3