RESEARCH
A mixed-modeling framework for whole-brain
dynamic network analysis
Mohsen Bahrami1,2 , Paul J. Laurienti1,2, Heather M. Shappell1,3,
Dale Dagenbach1,4, and Sean L. Simpson1,3
1Laboratory for Complex Brain Networks, Wake Forest School of Medicine, Winston-Salem, NC, USA
2Department of Radiology, Wake Forest School of Medicine, Winston-Salem, NC, USA
3Department of Biostatistics and Data Science, Wake Forest School of Medicine, Winston-Salem, NC, USA
4Department of Psychology, Wake Forest University, Winston-Salem, NC, USA
Keywords: Mixed models, fMRI, Dynamic brain networks, Multivariate, Simulation
a n o p e n a c c e s s
j o u r n a l
ABSTRACT
The emerging area of dynamic brain network analysis has gained considerable attention in
recent years. Tuttavia, development of multivariate statistical frameworks that allow for
examining the associations between phenotypic traits and dynamic patterns of system-level
properties of the brain, and drawing statistical inference about such associations, has largely
lagged behind. To address this need we developed a mixed-modeling framework that
allows for assessing the relationship between any desired phenotype and dynamic patterns
of whole-brain connectivity and topology. This novel framework also allows for simulating
dynamic brain networks with respect to desired covariates. Unlike current tools, Quale
largely use data-driven methods, our model-based method enables aligning neuroscientific
hypotheses with the analytic approach. We demonstrate the utility of this model in identifying
the relationship between fluid intelligence and dynamic brain networks by using resting-state
fMRI (rfMRI) data from 200 participants in the Human Connectome Project (HCP) study. Noi
also demonstrate the utility of this model to simulate dynamic brain networks at both group
and individual levels. To our knowledge, this approach provides the first model-based
statistical method for examining dynamic patterns of system-level properties of the brain and
their relationships to phenotypic traits as well as simulating dynamic brain networks.
AUTHOR SUMMARY
In recent years, a growing body of studies have aimed at analyzing the brain as a complex
dynamic system by using various neuroimaging data. This has opened new avenues to answer
compelling questions about the brain function in health and disease. Tuttavia, methods that
allow for providing statistical inference about how the complex interactions of the brain are
associated with desired phenotypes are to be developed for a more profound insight. Questo
study introduces a promising regression-based model to relate dynamic brain networks to
desired phenotypes and provide statistical inference. Inoltre, it can be used for simulating
dynamic brain networks with respect to desired phenotypes at the group and individual levels.
Citation: Bahrami, M., Laurienti, P. J.,
Shappell, H. M., Dagenbach, D., &
Simpson, S. l. (2022). A mixed-
modeling framework for whole-brain
dynamic network analysis. Network
Neuroscience, 6(2), 591–613. https://doi
.org/10.1162/netn_a_00238
DOI:
https://doi.org/10.1162/netn_a_00238
Supporting Information:
https://doi.org/10.1162/netn_a_00238
Received: 22 settembre 2021
Accepted: 9 Febbraio 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Mohsen Bahrami
mbahrami@wakehealth.edu
Handling Editor:
Vince Calhoun
Copyright: © 2022
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
Dynamic brain networks:
Large-scale brain networks that vary
over short periods of time on the
order of seconds.
INTRODUCTION
The past two decades have witnessed an explosion of studies aimed at examining the brain as
a complex system through analysis of neuroimaging data, particularly data from functional
MRI (fMRI). Complex functional systems of the brain are often analyzed through graph
theoretical measures of the brain’s functional networks (Bullmore & Sporns, 2009). Nodes
in brain networks often represent brain regions, and edges represent functional connections
(statistical associations) between the blood oxygen level–dependent (BOLD) signals in differ-
ent brain regions. Until recent years, most network studies of the brain focused on static func-
tional networks, in which the functional connections between brain regions were defined
over the entire scanning period or condition. Although such studies have provided promising
insights into functional organization and abnormalities of the brain (Bassett & Bullmore,
2009; Park & Friston, 2013), recent studies indicate that functional connectivity patterns
are not stationary and fluctuate over very short periods of time on the order of seconds
(Chang & Glover, 2010; Handwerker, Roopchansingh, Gonzalez-Castillo, & Bandettini,
2012; Hutchison et al., 2013; Parr, Rees, & Friston, 2018). This has resulted in a new and
rapidly evolving line of studies examining dynamic networks or time-varying functional con-
nectivity patterns of the brain.
Studies of brain dynamics are critical for establishing a profound understanding of the brain
given that the brain is a complex multiscale dynamic system rather than a stationary one (Lurie
et al., 2020). As noted in Breakspear (2017), analytical frameworks that allow for examining
the dynamical systems of the brain, from differential equations and state space analyses of
populations of neurons to larger scale network science methods, are essential for understanding
how behavioral and cognitive processes emerge from neural activities. Dynamic brain networks
have been associated with a wide range of cognitive and behavioral responses (Cole, Bassett,
Energia, Braver, & Petersen, 2014; Shine et al., 2016; Vidaurre, Abeysuriya, et al., 2018). More
specifically, they have been used to determine the engagement of a participant in a specific cog-
nitive task (Gonzalez-Castillo et al., 2015; Shirer, Ryali, Rykhlevskaia, Menon, & Greicius,
2012), and have been associated with consciousness (Barttfeld et al., 2015; Godwin, Barry, &
Marois, 2015), apprendimento (Bassett et al., 2011), and various neuropsychiatric and neurological
disorders, such as schizophrenia (Rashid et al., 2016; Sakoglu et al., 2010), depression (Lungo
et al., 2020; Martinez, Deco, Ter Horst, & Cabral, 2020), Alzheimer’s disease (Gu et al.,
2020; Jones et al., 2012), and Parkinson’s disease (Diez-Cirarda et al., 2018; Zhu et al.,
2019). New studies indicate that dynamic brain networks may provide more sensitive biomark-
ers for detecting differences between study populations or individuals than static networks
(Rashid et al., 2016). Per esempio, dynamic patterns of brain connectivity at rest have been
shown to better characterize people with post-traumatic stress disorder (Jin et al., 2017) or pre-
dict weight loss among older adults (Mokhtari, Laurienti, Rejeski, & Ballard, 2019).
Despite such insights, substantial challenges remain to enable more accurate analysis of
dynamic brain networks and accurate interpretation of results. Dynamic changes in the sys-
temic organization of brain networks confers much of our brains’ functional abilities (Bressler
& Menon, 2010; Buzsaki & Draguhn, 2004). If functional connections are lost or rendered
dynamically rigid due to an adverse health condition, compensatory connections may develop
to maintain organizational consistency and functional abilities. Consequently, brain network
analysis necessitates a suite of tools including a multivariate modeling framework for dynamic
brain network data to assess effects of multiple variables of interest and topological network
features on the overall network structure. Tuttavia, most current methods rudimentarily com-
pare the dynamic patterns of connection strength or networks measures across study popula-
zioni (Elton & Gao, 2015; Fukushima et al., 2018; Sizemore & Bassett, 2018), failing to fully
Network Neuroscience
592
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
System-level properties:
Properties of the brain as a system
comprising multiple interacting
regions.
harness the wealth of information about dynamics of system-level properties of the brain,
which can be obtained via a validated multivariate statistical method.
To date, despite having a broad range, most analytical approaches have aimed at identify-
ing similar FC patterns (per esempio., through clustering methods; Allen et al., 2014) or hidden brain
stati (often through hidden Markov models; Shappell, Caffo, Pekar, & Lindquist, 2019;
Vidaurre, Hunt, et al., 2018) with respect to desired outcomes. Although such methods have
provided useful insights, there remain many gaps in the suite of analytic methods available,
including the following. (1) One gap is the lack of a well-defined statistical framework to align
the neuroscientific hypothesis with the analytical approach and to assess the effects of multiple
phenotypes of interest simultaneously. As pointed out earlier, most current methods use
clustering-based approaches on FC patterns or hidden Markov models on time series to iden-
tify brain states and their transitions with respect to behavioral and cognitive outcomes. Such
data-driven methods have allowed for identifying complex patterns of dynamic changes in the
brain, providing profound insight into dynamic brain networks. Così, they may still be the
most appropriate methods for studies aimed at examining state-based changes in dynamic
brain networks or for comparing study populations in the absence of a well-defined hypoth-
esis. Tuttavia, to conduct statistical inference (cioè., hypothesis testing) on a well-posited neu-
roscientific hypothesis about dynamic brain networks, a model-based method that allows
selectively incorporating the desired topological properties of the brain or FC patterns as well
as combinations of desired variables, will provide a more fruitful framework. Other gaps in the
suite of analytic methods include (2) difficulty in controlling for confounding effects, ad esempio
age and gender (this often requires matching the study populations, which is a daunting task
for most neuroimaging studies); (3) the limitation of examining fluctuations of single network
measures instead of capturing the complex dynamics of brain networks as a system; E (4)
inability to simulate dynamic brain networks with respect to changes in system-level properties
of the brain and desired covariates. Such critical methodological gaps may be addressed
through parsimonious multivariate statistical frameworks.
As noted by Shine et al. (2019), the neurobiological mechanisms underlying brain network
dynamics (dynamic changes in functional architecture) remain poorly understood; and as
pointed out in Liu (2017), “novel methods are urgently needed for a better quantification of
temporal dynamics in resting-state fMRI.” The development of rigorous statistical methods
within a multivariate framework are among such urgent needs. Developing and disseminating
explainable, validated multivariate statistical methods are paramount for relating phenotypic
traits to dynamic changes in network properties of the brain, which will greatly aid in provid-
ing profound insights into normal and abnormal brain function.
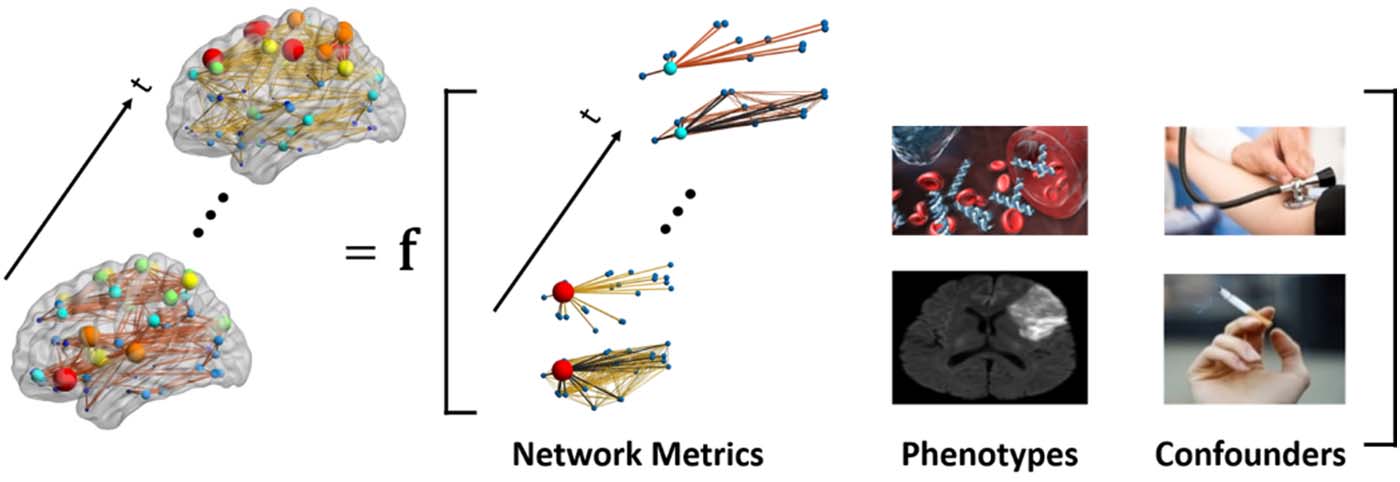
For the modeling framework, if we have
(cid:1)
Data
Y it :network of participant i at time point t
X it :covariate information
we wish to accurately estimate the probability density function of the networks given the
covariates P(Yit|Xit, βit), where βit are the parameters that relate the covariates to the network
structure as shown in Figure 1. Tuttavia, the development of such methods has vastly lagged
behind, mainly due to the same challenges which exist in developing multivariate statistical
tools for static networks (Bahrami, Laurienti, & Simpson, 2019UN; Shehzad et al., 2014; Simpson
& Laurienti, 2016). Here we introduce a novel multivariate statistical framework for assessing
phenotype-dynamic brain network pattern relationships and drawing inference from such
relationships.
Network Neuroscience
593
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
Figura 1. Dynamic brain networks as a function of endogenous and exogenous variables of interest.
Dynamic patterns of brain connectivity (presence/absence and strength) is modeled as a function of
(dynamic) nodal and global network metrics (per esempio., clustering coefficient, global efficiency, eccetera.) E
exogenous covariates, including phenotypes (per esempio., blood measurements and brain damage) E
possible confounding effects (per esempio., hypertension and smoking).
This model-based framework is a fundamentally different approach toward analyzing
dynamic brain networks when compared to current approaches, which often use data-driven
methodologies to identify “brain states” and their transitions with respect to health and behav-
ioral outcomes (Allen et al., 2014; Shappell et al., 2019; Shine & Poldrack, 2018; Vidaurre,
Smith, & Woolrich, 2017). We develop this method by advancing a promising statistical
mixed-modeling framework for static networks (Simpson & Laurienti, 2015). Several exten-
sions of the original framework (Bahrami et al., 2019UN; Simpson, Bahrami, & Laurienti,
2019), as well as a MATLAB toolbox (Bahrami, Laurienti, & Simpson, 2019B) have recently
been introduced. Tuttavia, it has yet to be extended to the dynamic network context. A
our knowledge, this proposed extension will be the first to allow relating group- E
individual-level characteristics to time-varying changes in spatial and topological brain
network properties while also maintaining the capabilities of the original model, ad esempio
accounting for variance associated with confounders.
We will demonstrate the utility of this framework by identifying the relationship between
fluid intelligence and dynamic brain network patterns by using data from 200 participants from
the Human Connectome Project (HCP) (Van Essen et al., 2013) study. Fluid intelligence (gF)
refers to reasoning ability and the capacity of an individual to discern patterns or solve prob-
lems when that individual doesn’t have or has minimal resources or acquired knowledge to act
upon (Cattell, 1987). Understanding the neurobiological underpinnings of gF is of great inter-
est, as it has been associated with a variety of cognitive abilities (Colom & Flores-Mendoza,
2007; Unsworth, Fukuda, Awh, & Vogel, 2014; Ye et al., 2019). We also demonstrate the utility
of this new framework for simulating dynamic brain networks, which can be critical for a
better understanding of topological variability in time and across individuals with respect to
desired covariates. To our knowledge, this simulation capability is also the first to enable
simulating representative group-level dynamic networks from an array of desired global/local
network measures and phenotypic characteristics.
MATERIALS AND METHODS
Motivating Data
We used the rich dataset provided by the HCP study (Van Essen et al., 2013) to be able to
explore dynamic functional brain network differences in cognitively variable populations as
a function of phenotype, while maintaining continuity with previous analyses to contrast and
clearly distinguish the novel utilities of our proposed method. We specifically focused on
Network Neuroscience
594
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
demonstrating the utility of our framework in assessing the relationship between dynamic
functional networks and intelligence due to the great interest in identifying such relationship.
The HCP data released to date include 1,200 individuals. Of those, 1,113 (606 female; 283
minority) have complete MRI images, cognitive testing, and detailed demographic informa-
zione. Participants in the HCP were screened to rule out neurological and psychiatric disorders.
All data were collected on 3T Siemens MRI scanners located at Washington University or the
University of Minnesota, using identical scanning parameters. The HCP performed extensive
testing and development to ensure comparable imaging at the two sites. The BOLD-weighted
images were collected using the following parameters: TR = 720 ms, TE = 33.1 ms, voxel size
2 mm3, 72 slices, E 1,200 volumes. In this study, we selected a subsample comprising 389
individuals with unique family identification numbers that also passed our image processing
quality control assessments. For multiple individuals with the same family identification num-
ber, one individual was selected randomly. We initially used the entire 389 individuals, but we
further reduced this to 200 individuals (randomly chosen from our subsample) after we faced
convergence issues in modeling one of the two-part mixed-effect models. We have provided
the HCP identification numbers for the 200 individuals used in this study in Supporting Infor-
mation Appendix S1 (Table S1). The HCP analyses are an exemplar; importantly, our methods
can be applied to any network-based neuroimaging study.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Dynamic Networks Generation
We used minimally preprocessed resting fMRI (rfMRI) data from HCP (Glasser et al., 2013). Noi
used two scans for each individual, the left-to-right (LR) and right-to-left (RL). For each scan,
we used ICA-AROMA (Pruim et al., 2015) to correct for any motion artifact in the rfMRI data. UN
band-pass filter (0.009–0.08 Hz) was applied to each scan. The LR and RL scans for each indi-
vidual were then concatenated temporally, and transient artifacts were removed using a win-
dowed wavelet transformation. Then a regression was performed with the mean tissue signals
(GM, WM, and CSF), the six movement parameters and derivatives, as well as a binary regres-
sor to account for any mean signal differences between the two scan conditions (LR and RL
scans). Our quality control process removed 116 individuals from the analysis. QC included
manually checking the rfMRI for warping irregularities as well as remaining motion artifact
after the above processing steps. For the remaining individuals, among those with the same
family identification number, one individual was selected randomly. This provided a final
dataset comprising 389 individuals with unique family identification numbers. For all 389
individuals, we divided the brain into 268 regions based on a functional atlas (Shen, Tokoglu,
Papademetris, & Constable, 2013) and averaged all time series within each region to create a
single time series for that region. We used a continuous wavelet transform to filter artifact
resulting from the LR and RL concatenation with a window size of 30 S (covering 15 s from
the ending and starting points of LR and RL time series, rispettivamente). We then prewhitened the
time series to avoid undesired autocorrelation effects for our regression analyses and as rec-
ommended by (Honari, Choe, Pekar, & Lindquist, 2019) for dynamic network analyses by
using a sliding window correlation approach.
Dynamic brain networks for each participant were constructed through a sliding window
correlation approach. We used a modulated rectangular window (Mokhtari, Akhlaghi, Simpson,
Wu, & Laurienti, 2019) with a length of 120 volumes and the same shift size (cioè., 120 volumes)
to generate windows with no overlap between consecutive networks. We understand that
this is not a commonly used shift size as most studies use overlapping windows with 1 TR
shift size; Tuttavia, unlike other methods, we subsequently use the dynamic networks in a
regression framework, thus we used windows with no overlap to further reduce temporal
Network Neuroscience
595
A mixed model for dynamic brain networks
autocorrelation. Tuttavia, in an additional analysis provided in Supporting Information
Appendix S2, we used windows with 50% overlap between consecutive networks, questo è,
using a shift size of 60 volumes, which yielded similar results. Since at least one other study
that uses dynamic networks in a regression framework has used windows with 50% sovrapposizione
between consecutive networks (Chang, Liu, Chen, Liu, & Duyn, 2013), we conducted this
additional analysis to see how our reported results would be affected when using this
approach. Despite similar results, we present results for windows with no overlap between
consecutive networks to ensure that temporal autocorrelation effects are minimized. The rea-
sons and implications of our choices for window type, window length, and shift size will be
further explained in the Discussion. The dynamic networks for each participant were gener-
ated by moving the window across the time series and computing the Pearson’s correlation
between time series of all pairs of 268 regions at each shift. This yielded 19 dynamic net-
works for each participant. We then thresholded all dynamic networks to remove negative
correlations as multiple network measures, particularly clustering, remain poorly understood
in networks with negative correlations (Friedman, Landsberg, Owen, Li, & Mukherjee, 2014;
Telesford, Simpson, Burdette, Hayasaka, & Laurienti, 2011). Inoltre, distributions of net-
work variables (such as degree) and the neurobiological interpretation of positive and nega-
tive edges are different (Parente et al., 2018; UN. J. Schwarz & McGonigle, 2011). When
including positive and negative edges in an analysis, networks should be generated and
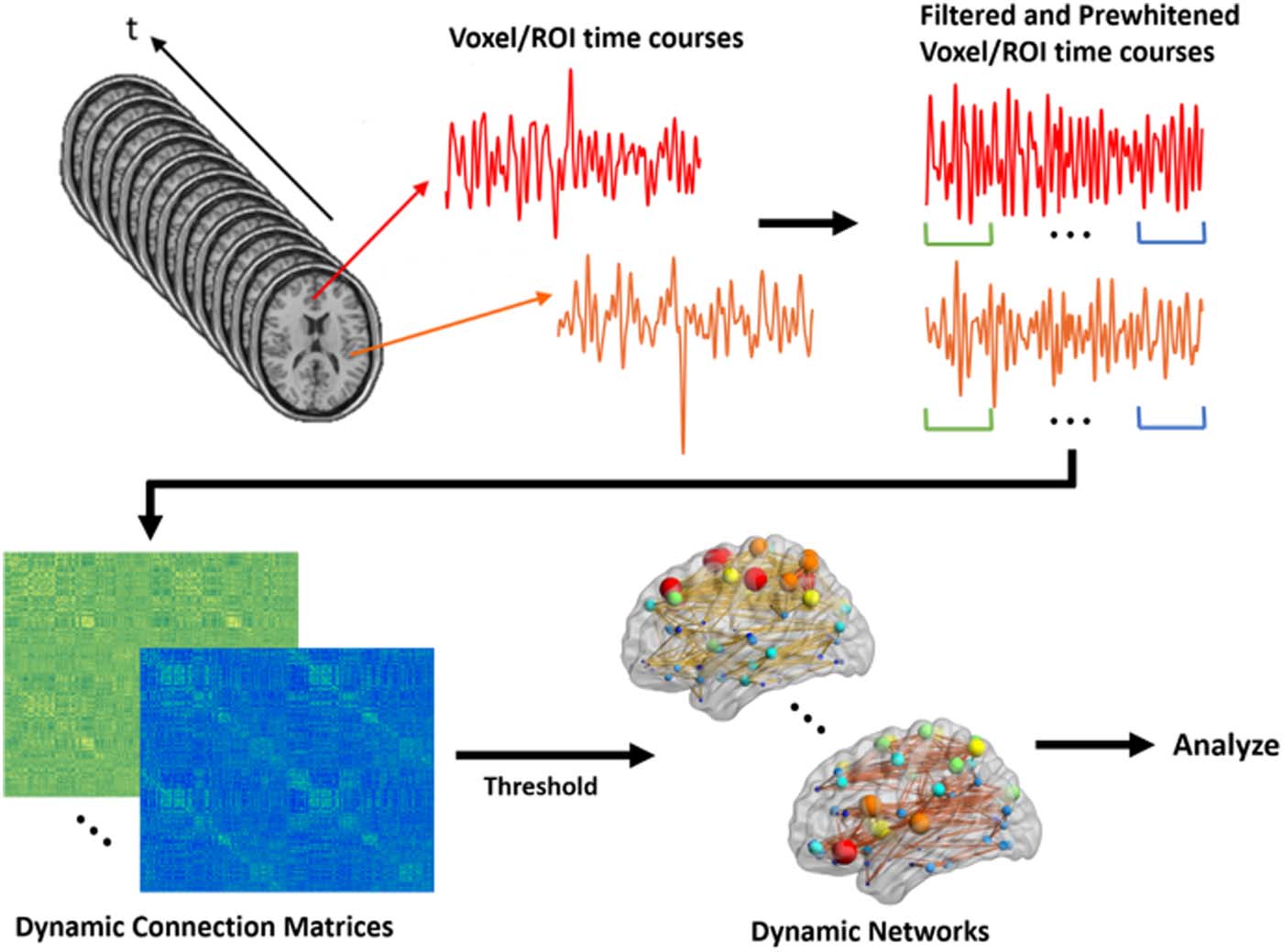
assessed separately (UN. J. Schwarz & McGonigle, 2011). Figura 2 shows a schematic exhibit-
ing this dynamic network generation process.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. Schematic for generating dynamic brain networks from fMRI time series. Time series are
first filtered and prewhitened to remove the undesired undershoots/overshoots in the middle as well
as undesired effects of autocorrelation. Then, using a sliding window correlation approach, func-
tional connectivity between brain areas is estimated between all time series pairs at each shift to
produce a connection matrix at that shift. By moving the window across the entire length of time
series, a series of dynamic connection matrices will be produced for each participant. A threshold is
applied to the matrix to remove negative connections. These networks are subsequently used for
analyses.
Network Neuroscience
596
A mixed model for dynamic brain networks
Mixed-effects (or mixed) models:
Statistical regression models that
include both fixed (cioè., population-
level) and random (cioè., individual-
level) parameters.
We also used the original models (Simpson & Laurienti, 2015) to conduct the same analyses
but with static networks to further examine whether/how dynamic and static networks provide
different insight into fluid intelligence–brain network associations (see Supporting Information
Appendix S3).
Mixed-Effects Modeling Framework for Weighted Dynamic Brain Networks
Given that we have sparse weighted networks, a two-part mixed-effects model will be
employed to model both the probability of a connection (presence/absence) and the strength
of a connection, if it exists (Simpson & Laurienti, 2015). The model includes the entire brain
connectivity matrix of each participant, endogenous covariates, and exogenous covariates (Vedere
Figura 1). The endogenous covariates are summary variables extracted from the network to
summarize global topology. The exogenous covariates are the biologically relevant pheno-
typic variables (per esempio., for our data, fluid intelligence, sex, race, and education among others).
This statistical framework allows for the evaluation of group and individual effects. Another
key feature of the model is the multivariate nature of the statistics. Inclusion of the actual con-
nectivity matrices allows the statistics to be performed on the entire network simultaneously,
rather than performing edge-by-edge analyses in a massively univariate fashion.
More specifically, let Yijkt represent the strength of the connection (quantified as the corre-
lation in our case) and Rijkt indicate whether a connection is present (presence variable)
between node j and node k for the ith participant at time t. Così, Rijkt = 0 if Yijkt = 0, and Rijkt =
1 if Yijkt > 0 with conditional probabilities:
(cid:4)
(cid:5)
(cid:6)
(cid:3)
P Rijkt ¼ rijktjβ
; bri; γ
R
; d ri
R
¼
ð
1 − pijkt β
R
ð
; bri; γ
pijkt β
R
; bri; γ
R
; d ri
R
; d ri
Þ if
rijkt ¼ 0
Þ if
rijkt ¼ 1
;
(1)
where pijkt( βr; bri; γr; dri) is the probability of a connection between nodes j and k for partic-
ipant i at time t. We then have the following logistic mixed model (part I model) for the prob-
ability of this connection:
(cid:3)
logit pijkt β
ð
R
; bri ; γ
R
; d ri
(cid:4)
Þ
¼ X
0
ijkt
β
R
þ
Xn
o¼1
!
!
γ
ros oð Þ Xtð
Þ
þ Z
0
ijktbri þ
drios oð Þ Ztð
Þ
;
(2)
Xn
o¼1
Fixed effects:
Variables in the mixed models whose
effects are constant across all
individuals.
Random effects:
Variables in the mixed models whose
effects change across individuals.
Orthonormal polynomials:
A family of orthogonal polynomials
where the inner product of the same
polynomials is one.
N
P
where βr (note that r and s subscripts simply denote the probability and strength models in
Equations 2 E 3, rispettivamente, and have no relationship with Rijkt or Sijkt in the next equation)
is a vector of population parameters (fixed effects) that relate the probability of a connection to
a set of covariates (Xijkt) for each participant and nodal pair (dyad), bri is a vector of participant-
and node-specific parameters (random effects) that capture how this relationship varies about
the population average (βr) by participant and node (Zijkt), Zijkt is the design matrix for the ran-
o¼1 γros(o)(Xt) corresponds to a population-level nth-order orthonormal polyno-
dom effects,
P
N
mial model capturing the dynamic trend in the presence of connections across time, E
o¼1
drios(o)(Zt) corresponds to an individual-level nth-order orthonormal polynomial model capturing
how much the participant-specific trends deviate from the population trend. S(o) is the value of
the oth degree polynomial generated from a set of orthonormal polynomials with maximum degree
poynomial of n, Xt is the design matrix for the population-level orthonormal polynomials at time t,
and Zt is the deisgn matrix for the individual-level orthonormal polynomials at time t (see Supporting
Information Appendix S4 for a simple example of defining the design matrix and orthonormal poly-
nomials). Employing an orthonormal polynomial model in this manner has been shown to accu-
rately represent the trend in time series data while avoiding the computational issues resulting from
the use of natural polynomials (Edwards & Simpson, 2014; Simpson & Edwards, 2013). Note that
our βr and βs are population estimates showing if/how the desired covariates are associated with
Network Neuroscience
597
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
dynamic networks, questo è, after accounting for the dynamic trends through incorporating ortho-
normal polynomials, βr and βs, show if/how the relationships between desired covariates and
dynamic networks, on average, are significant.
For the part II model, which aims to model the strength of a connection given that there is
one, we let Sijkt = [Yijkt|Rijkt = 1]. In our case, the Sijkt will be the values of the correlation
coefficients between nodes j and k for participant i at time t. We can then use Fisher’s
Z-transform, denoted as FZT, to induce normality for the following mixed model (part II model)
(cid:3)
FZT Sijkt β
ð
S
; bsi; γ
; d ri
S
(cid:4)
Þ
¼ X
0
ijkt
β
S
þ
Xn
o¼1
!
γ
sos oð Þ Xtð
Þ
þ Z
0
ijktbsi þ
!
Xn
o¼1
dsios oð Þ Ztð
Þ
þ eijkt;
(3)
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
where βs is a vector of population parameters that relate the strength of a connection to the
same set of covariates (Xijkt) for each participant and nodal pair (dyad), bsi is a vector of
participant- and node-specific parameters that capture how this relationship varies about
the population average (βs) by participant and node (Zijkt),
o¼1 γsos(o)(Xt) corresponds to a
population-level nth-order orthonormal polynomial model capturing the dynamic trend in
N
o¼1 dsios(o)(Zt) corresponds to an individual-level
the strength of connections across time,
nth-order orthonormal polynomial model capturing how much the participant-specific trends
deviate from the population trend, and eijkt accounts for the random noise in the connection
strength of nodes j and k for participant i at time t.
P
P
N
In this study, the covariates (Xijkt) used to explain and predict both the presence and strength
of connection include the following. (1) Net: the average of the following network variables
(categorized and further detailed in Table 1 and in Rubinov and Sporns [2010] and Simpson,
Bowman, and Laurienti [2013]) in each dyad: Clustering Coeficient (C ), Global Efficiency
(Eglob), Degree (k) (difference between connected nodes instead of average to capture “assor-
tativity”), Modularity (Q), and Leverage Centrality (Joyce, Laurienti, Burdette, & Hayasaka, 2010)
(l ). Clustering coefficient and global efficiency have been widely used as hallmark measures
of segregation (presence of highly interconnected regions supporting regional specialization)
and integration (widespread connectivity interconnecting specialized regions) in the brain,
rispettivamente (Rubinov & Sporns, 2010).
We have used degree difference to capture assortativity, which provides a profound character-
ization of network resilience (Rubinov & Sporns, 2010). Leverage centrality measures the extent of
connectivity of a node relative to its neighbors’ connectivity (Joyce, Laurienti, Burdette, &
Hayasaka, 2010). This characterizes the importance of each node for information flow in the
brain. Modularity is a hallmark measure of community structure in the brain which has been
associated with cognitive performance and intelligence as will be discussed in more detail in
the Discussion. (2) COI: Covariates of Interest (fluid intelligence (gF) in our study – we modeled
Tavolo 1. Network measures by category
Category
Functional segregation
Measure(S)
Clustering coefficient
Type
Local measure
Functional integration
Global efficiency
Globale(/Local) measure
Resilience
Degree difference
Local measure
Centrality and information flow
Leverage centrality
Local measure
Community structure
Modularity
Global measure
Network Neuroscience
598
A mixed model for dynamic brain networks
gF as a continuous covariate to maximize power, Tuttavia, in separate analysis, we used gF as a
median split binary variable (low/high) which yielded similar results. The binary results can be
found in the Supporting Information Appendix S5). gF in the HCP protocol has been assessed
using the Raven’s progressive matrices with 24 items with scores being integers representing num-
ber of correct items (Bilker et al., 2012); (3) Int: Interactions of the Covariate of Interest with the
variables in 1; E (4) Contro: Confounders (for our data: Sex (binary), Age (continuous), years of
Education (categorical with three levels − level 1 (≤11), level 2 (12–16), and level 3 (≥17)),
BMI (continuous), Race (categorical with six categories − cat 1 (Am. Indian/Alaskan Nat.), cat 2
(Asian/Nat. Hawaiian/Other Pacific Is.), cat 3 (Black or African Am.), cat 4 (White), cat 5 (More
than one), cat 6 (Unknown or Not Reported)), Ethnicity (categorical with three categories − cat 1
(Hispanic/Latino), cat 2 (Not Hispanic/Latino), cat 3 (Unknown or Not Reported)), Handedness
(continuous − ranging from −100 to +100, with negative and positive numbers indicating whether
participants were more left- or right-handed, rispettivamente, assessed using the Edinburgh
Handedness Inventory (Oldfield, 1971), Income (Continuous − Total household income),
Alcohol abuse (Binary − Indicating whether participant met DSM4 criteria for alcohol abuse),
Alcohol dependence (Binary − Indicating whether participant met DSM4 criteria for alcohol
dependence), Smoking status (Binary − Indicating whether participant smoked or not), Spatial
distance between nodes (importance of spatial distance as potential geometric confounders has
been discussed in Friedman et al. (2014), and square of spatial distance between nodes). We used
these confounding variables given their widely studied effects on intelligence. Così, we can
decompose βr and βs into βr = [βr,0 βr,net βr,coi βr,int βr,con] and βs = [βs,0 βs,net βs,coi βs,int βs,con] A
correspond with the population intercepts and these covariates. For the random-effects vectors we
have that bri = [bri,0 bri,net bri,dist δri,jδri,k] and bsi = [bsi,0 bsi,net bsi,dist δsi,jδsi,k], where bri,0 and bsi,0
quantify the deviation of participant-specific intercepts from the population intercepts (βr,0 E
βs,0), bri,net, and bsi,net contain the participant-specific parameters that capture how much the
relationships between the network variables in (1) and the presence and strength of a
connection vary about the population relationships (βr,net and βs,net), rispettivamente, bri,dist and bsi,dist
contain the participant-specific parameters that capture how much the relationship between
spatial distance (and square of spatial distance) and the presence and strength of a connection vary
about the population relationships, rispettivamente, δri,j and δsi,j contain nodal-specific
parameters that represent the propensity for node j (of the given dyad) to be connected and the
magnitude of its connections, rispettivamente, and δri,k and δsi,k contain nodal-specific parameters
that represent the propensity for node k (of the given dyad) to be connected and the magnitude
of its connections, rispettivamente. Parameters for all 19 time points (number of networks per
individual) (t = 1, 2, …, 19) are estimated or predicted simultaneously from the model. In
general, additional covariates can also be incorporated as guided by the biological context.
Specifying a reasonable covariance model (balancing appropriate complexity with parsi-
mony and computational feasibility) is paramount for a unified dynamic model such as the
one developed here. Toward this end, we assume that bri, dri, bsi, dsi, and ei are normally
distributed and mutually independent, with variance component covariance structures for
bri, dri, bsi, and dsi, and the standard conditional independence structure for ei. Questo è, bri ∼
(cid:3)
br;node2 , …,
N(0, Σ b r i(τ b r) = diag (τ b r)), where τbr ¼
dr;N)0, bsi ∼ N(0,
σ2
, E
, dri ∼ N(0, Σdri(τdr) = diag (τdr)), where τdr = (σ2
dr;1, …, σ2
(cid:3)
bs;net , σ2
bs;0, σ2
σ2
bs;node2, …, σ2
Σbsi(τbs) = diag (τbs)), where τbs ¼
(cid:3)
(cid:4)0—yielding (276 + (N + 1))
ds;0, σ2
σ2
dsi ∼ N(0, Σdsi(τds) = diag (τds)) where τds ¼
random effects variance parameters for both the presence and strength models—and ei ∼
N(0, Σei = σ 2I). These variance and covariance parameters will provide insight into whether
individual and group differences in variability in dynamics relate to health and behavioral
dr;0, σ2
bs;node1, σ2
br;node1 , σ2
ds;1, …, σ2
br;dist , σ2
br ;0 , σ2
σ2
br ;net , σ2
bs;dist, σ2
br;node268
bs;node268
ds;N
(cid:4)0
(cid:4)0
Network Neuroscience
599
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
outcomes. Parameter estimation is conducted via restricted pseudo-likelihood (Wolfinger &
Oconnell, 1993) with the residual approximation of the F test for a Wald statistic employed
for inference.
We implemented the models (Equations 2 E 3) above to describe and compare brain net-
work dynamics as a function of fluid intelligence. For both models, we started model fitting
with the entire set of random effects, questo è, random effects for intercept, nodal network mea-
sures (clustering, global efficiency, degree, and leverage centrality), distance, and nodal pro-
pensities. Tuttavia, after facing convergence issues, we dropped nodal propensities from our
random effects. We assessed model goodness-of-fit (GOF) and consistency of estimates (A
further avoid overfitting) to determine the orthonormal polynomial degree yielding the best
model fits. We fit the two-part model defined above with the mentioned fixed- and random-
effect parameters by using orthonormal polynomial models of degrees ranging from 3–18
(giving 16 model fits), and determined the “best” model based on a composite approach
employing the Akaike information criterion (AIC) (Akaike, 1981), Bayesian information crite-
rion (Schwarz, 1978), modified AIC (AICc) (Hurvich & Tsai, 1989), Hannan–Quinn informa-
tion criterion (Hannan & Quinn, 1979), and consistent AIC (CAIC) (Bozdogan, 1987) GOF
measures as well as the consistency of the obtained parameter estimates and p values
to further avoid overfitting. We used MATLAB to generate the appropriate data frame for
our framework and used SAS v9.4 on a Linux operating system with 330 GB of RAM and
2.60 GH processor to perform the model fitting. We have provided the essential SAS macro
employed in fitting the statistical mixed models for both Equations 2 E 3 in Supporting
Information Appendix S6.
Simulations
Bernoulli distribution:
Discrete probability distribution for a
random variable with binary (cioè., 0
O 1) outcome.
We used the fitted models from Equations 2 E 3 to simulate dynamic brain networks. Noi
used covariates from 50 participants (selected randomly from 200) E 10 dynamic networks
(selected randomly from 19), and simulated 10 realizations for each dynamic network of each
participant. This yielded a total of 5,000 simulated dynamic networks. To simulate each
dynamic network, we first simulated the existence of edges (presence/absence) for all
35,778 node pairs (vectorized symmetric network with 268 nodes) from a Bernoulli distribution
with the probability from the fitted model (pijkt(βr; bri; γr; dri) from Equation 2 and the covariates
used for each participant’s dynamic network. We simulated the random-effect coefficients
(γr, dri) for each participant from a normal distribution with mean zero and the covariance matrix
obtained from the estimated parameters for random effects in Equation 2. To simulate the strength
values, we first simulated continuous values from a normal distribution with the mean and covari-
ance obtained from the fitted model in Equation 3 and the covariates for each dynamic network
(cid:7)
(cid:7)
N; μ
. Noi
then used the inverse Fisher’s Z-transform to get the untransformed values, and finally multiplied
the resulting vector by the simulated binary vector to get the simulated strength values for the
weighted network. We then calculated several (weighted) descriptive measures often used in
analyzing brain networks to compare the simulated and observed networks with respect to such
topological measures as this provides the most appropriate way to assess the GOF of statistical
models in the network context (Hunter, Goodreau, & Handcock, 2008). All simulations were
done in MATLAB. The MATLAB script is provided in the Supporting Information Appendix S7.
We have also provided the HCP identification numbers for the 50 randomly chosen participants
used in our simulation along with the numbers of their 10 randomly chosen dynamic networks
in Supporting Information Appendix S8.
¼ Z ijktΣbsi τbs
þ Z t Σdsi τds
sos oð Þ Xtð
þ σ2I
o¼1 γ
¼X 0
ÞZ0
T
ÞZ0
; σ2
(cid:4)
Þ
β
S
P
þ
sim
sim
ijkt
ijkt
(cid:8)
(cid:8)
(cid:3)
ð
ð
N
Network Neuroscience
600
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
RESULTS
Here, we show our framework’s ability in identifying the relationship between fluid intelligence
and dynamic brain networks and its utility for simulating dynamic brain networks. For orthonormal
polynomial models of degrees ranging from 3 A 18, all GOF measures for the strength model
(Equazione 3) slightly improved with increasing degree, providing good fits for almost all degrees.
Tuttavia, the models with polynomial degrees ranging from 9 A 16 provided the most consistent
Tavolo 2.
Fixed-effect estimates, SEs, and p values for 12th degree orthonormal polynomial fit
Parameters Ortho Poly Degree
Modelli
Probability Model
Strength Model
γr,0
γr,1
γr,2
γr,3
γr,4
γr,5
γr,6
γr,7
γr,8
γr,9
γr,10
γr,11
γr,12
γs,0
γs,1
γs,2
γs,3
γs,4
γs,5
γs,6
γs,7
γs,8
γs,9
γs,10
γs,11
γs,12
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Intercept
1
2
3
4
5
6
7
8
9
10
11
12
Estimate
−0.13770
SE
0.02022
*P Value
<.0001
0.00121
0.004361
0.7817
−0.01694
0.004914
0.0021
0.02394
0.004304
<.0001
−0.00593
−0.00508
−0.01185
0.004641
0.3805
0.004429
0.4498
0.004382
0.0232
0.00794
0.004019
0.1259
0.00392
0.004106
0.4716
−0.00433
−0.01063
0.004443
0.4716
0.004236
0.0374
0.00774
0.004203
0.1583
−0.00481
0.004897
0.4716
Intercept
0.31190
0.00786
<.0001
1
2
3
4
5
6
7
8
9
10
11
12
0.00148
0.00214
0.4906
−0.01415
0.00221
<.0001
0.01782
0.00203
<.0001
−0.00110
−0.00057
−0.01012
0.00197
0.5785
0.00214
0.7917
0.00211
<.0001
0.01077
0.00226
<.0001
0.00388
0.00228
0.1106
−0.00440
−0.00726
0.00204
0.0426
0.00200
0.0005
0.00134
0.00199
0.5007
−0.00907
0.00257
0.0007
* Adjusted using the adaptive false discovery rate procedure described in Benjamini and Hochberg (2000).
Network Neuroscience
601
A mixed model for dynamic brain networks
estimates and p values. Thus, to avoid overfitting while still using a model with a relatively good fit
as indicated by the GOF measures, we used the model with polynomial degree of 12 as a middle
ground between 9 and 16. For the probability model, although all GOF measures slightly
improved with increasing degree too, the differences were negligible. Thus, we used the same
polynomial degree (12) for consistency. The estimates, standard errors, and p values for the poly-
nomial parameters are presented in Table 2 ( p values presented in this table and other tables are
corrected for multiple comparison using an adaptive false discovery rate procedure detailed in
Benjamini & Hochberg, 2000). The parameter estimates, standard errors, and p values (based
on the residual approximation of the F test for a Wald statistic) associated with the important
fixed-effect covariates are presented in Table 3. The estimates for other parameters (e.g., gender,
age, etc.) are fully explained in the Supporting Information (Table S6). The estimates quantify the
relationship between dynamic patterns of probability (presence/absence) and strength of (present)
connections between nodes (brain regions), as dependent variables, and the previously men-
tioned sets of covariates, including (dynamic patterns of ) endogenous network measures, fluid
intelligence as our covariate of interest, and confounders. The estimates for confounding covari-
ates, including sex, age, education, BMI, race, ethnicity, handedness, income, DSM4 alcohol
abuse, DSM4 alcohol dependence, smoking status, spatial distance, and square of spatial distance
between nodes are fully explained in the Supporting Information as pointed out above.
The estimates for interaction covariates shows if (and how) the relationship between dynamic
patterns of probability/strength of connections and dynamic patterns of endogenous network
measures are affected by fluid intelligence. Notable results are detailed in the following sections.
Dynamic Network Analysis
As Table 3 presents, dynamic changes of clustering (functional segregation), global efficiency
(functional integration), degree difference (functional resilience), and leverage centrality
Table 3.
Parameter estimates, standard errors, and p values for dynamic networks
Parameter
βr,0
βr,COI
βr,C
βr,Eglob
βr,D
βr,L
βr,Q
βr,COI×C
βr,COI×Eglob
βr,COI×D
βr,COI×L
βr,COI×Q
Probability Model Outputs
Estimate
−0.1377
0.00319
−7.22530
12.5799
−0.07686
0.04332
2.10910
0.10290
0.24720
−0.00140
−0.01690
−0.02684
SE
0.02022
0.00295
0.11490
0.43460
0.00389
0.02006
0.01471
0.11500
0.43470
0.00389
0.02006
0.01514
*P value
<.0001
Parameter
βs,0
Strength Model Outputs
SE
Estimate
0.00786
0.31190
0.4716
<.0001
<.0001
<.0001
0.0872
<.0001
0.4836
0.5696
0.7177
0.4836
0.1619
βs,COI
βs,C
βs,Eglob
βs,D
βs,L
βs,Q
βs,COI×C
βs,COI×Eglob
βs,COI×D
βs,COI×L
βs,COI×Q
−0.00086
3.07330
3.86090
−0.07111
−0.21300
−1.35550
−0.02418
−0.00301
−0.00002
0.00127
0.03078
0.00115
0.02008
0.04105
0.00672
0.00273
0.00241
0.02009
0.04106
0.00672
0.00273
0.00248
*P value
<.0001
0.4540
<.0001
<.0001
<.0001
<.0001
<.0001
0.2478
0.9416
0.9978
0.6423
<.0001
* Adjusted using the adaptive false discovery rate procedure described in Benjamini and Hochberg (2000). Bold values show fluid intelligence–related
inferential results discussed here.
Network Neuroscience
602
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
n
e
n
_
a
_
0
0
2
3
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
A mixed model for dynamic brain networks
(information flow) all play important roles in explaining dynamic patterns of both connection
probability and strength, but with leverage centrality having a marginal effect on dynamic pat-
terns of connection probability.
Fluid intelligence, our covariate of interest (COI), is neither directly related to dynamic pat-
terns of connection probability (presence/absence) nor connection strength as indicated by the
p values associated with βr,COI (p value = 0.4716) and βs,COI (p value = 0.4540), respectively.
However, it has a significant effect on the relationship between dynamic changes of connec-
tion strength and dynamic changes of whole-brain modularity as indicated by the p value asso-
ciated with βs,COI ×Q (p value < 0.0001), while having no effect on other relationships (using the
gF as a median split binary [low/high] variable rather than a continuous one, provided the same
results as shown in Supporting Information Appendix S5). Dynamic changes of whole-brain
modularity and connection strength are negatively associated with each other (βs,Q), which
implies the dominance of between-community (rather than within-community) connections
in driving the dynamic changes of whole-brain modularity. Fluid intelligence interacts with this
relationship—as intelligence increases, dynamic changes of modularity are less driven by
dynamic changes in between-community (and more by within-community) connectivity as
indicated by the positive and significant estimate for βs,COI ×Q. (However, the dynamics of
between-community connections are still the dominant factor in driving the dynamic changes
of whole-brain modularity.)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
n
e
n
_
a
_
0
0
2
3
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
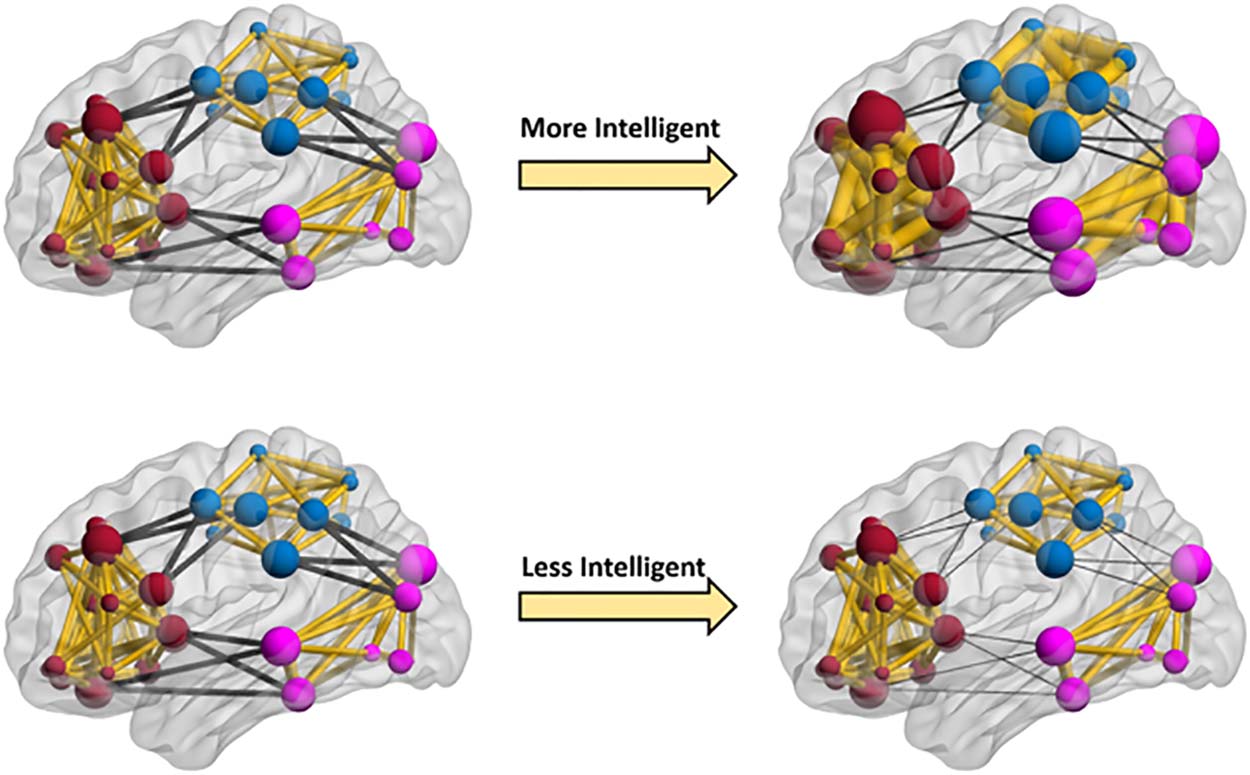
Figure 3. Cartoon depiction of fluid intelligence effects on dynamic brain networks. The nodes
represent brain regions, and edges represent dynamic functional connections. To illustrate the
effects of fluid intelligence on dynamic changes of modularity as interpreted from Table 3, three
artificial communities marked with separate colors (dark red, light blue, and purple) are shown
in this figure. The within- and between-community connections are shown with the yellow and
black colors, respectively. As this figure illustrates, dynamic changes of modularity are predomi-
nantly determined by between-community connections for any level of intelligence (here two levels
are shown: low and high). However, when comparing the more and less intelligent participants
(networks on the right), in more intelligent participants, dynamic changes of modularity are less
determined by between-community connections (thicker dark edges in top right), and dynamic
changes of within-community connections also play more important roles in changing the modu-
larity (thicker yellow edges in top right network). We should note that fluid intelligence was mod-
eled as a continuous variable, but for illustrative purposes we show higher and lower intelligence
levels. We have made a movie that better illustrates how dynamic patterns of whole-brain modu-
larity are affected across a range of fluid intelligence values (Supporting Information Movie S1).
Network Neuroscience
603
A mixed model for dynamic brain networks
Table 4. Weighted network measures of observed and simulated dynamic networks
Observed (N = 50 × 10)
Simulated (N = 50 × 10 × 10)
Metric
Clustering coefficient (C)
Mean
0.1778
Global efficiency (Eglob)
0.2948
Degree (K)
39.229
SD
0.0217
0.0096
2.8391
Mean
0.1463
0.3263
40.847
SD
0.0296
0.0517
7.3629
Our results might imply that brain networks in people with higher fluid intelligence are
more flexible with respect to changes in their modularity at rest. These changes are associated
with both stronger within-community connections (more specialized neural communities) and
weaker between-community connections (more segregated neural communities). This is illus-
trated in Figure 3 and Supporting Information Movie S1. We provide more detail on possible
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
n
e
n
_
a
_
0
0
2
3
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
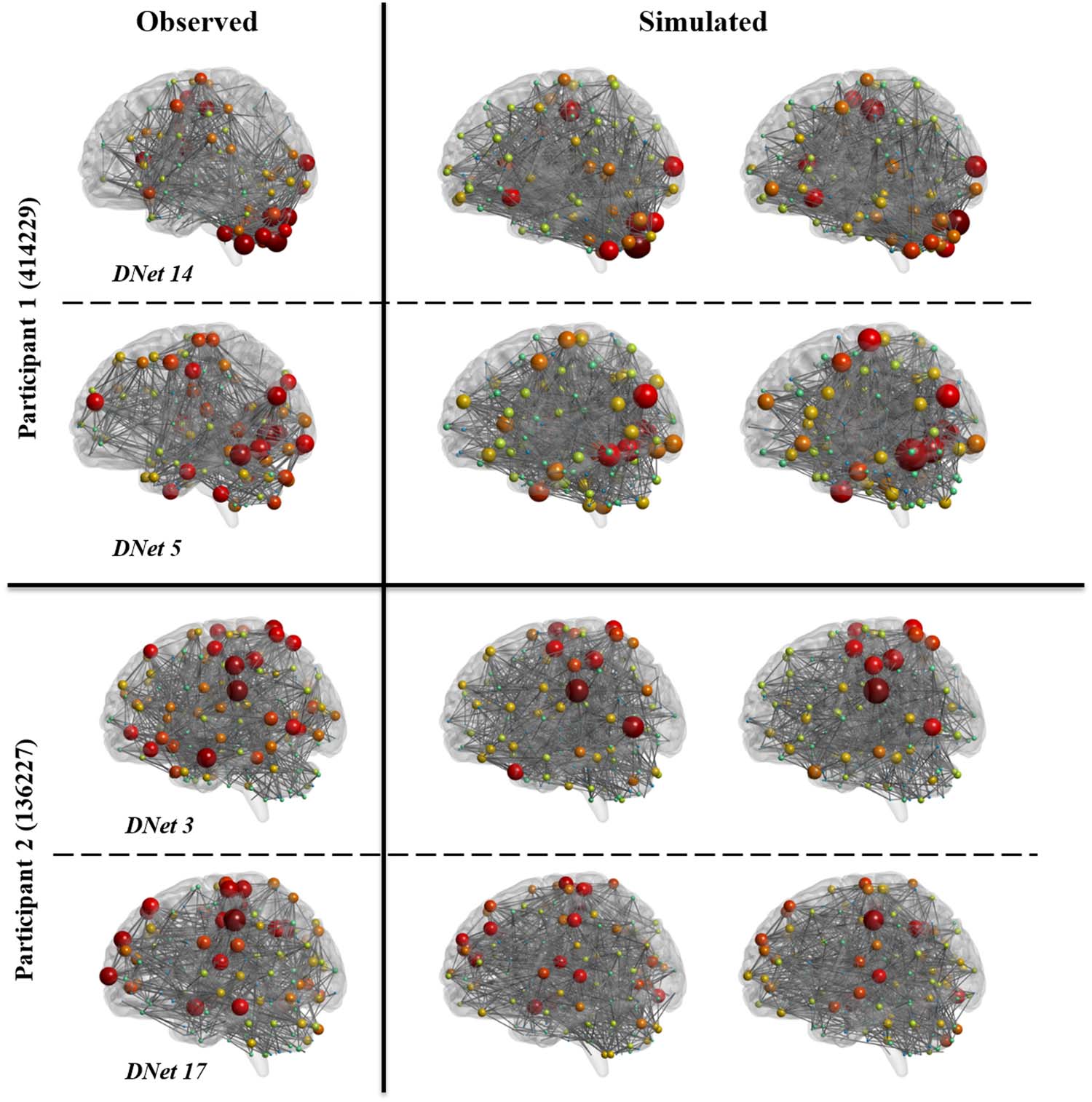
Figure 4. Observed and simulated dynamic networks for two randomly selected participants. For
each participant, two randomly selected dynamic networks (DNet #) are shown on the left, and for
each dynamic network, two randomly selected simulated networks (from the 10 simulation reali-
zations) are shown on the right. We have shown the HCP individual IDs on the left. All networks are
thresholded to maintain the top 5% of strongest connections. The size and color of each node rep-
resent the degree of that node.
Network Neuroscience
604
A mixed model for dynamic brain networks
interpretations and implications of this result in the Discussion. The results for using windows
with 50% overlap between consecutive networks were similar, but with gF modifying the rela-
tionship between dynamic patterns of whole-brain modularity and connection probability as
well. For more detail and a brief interpretation see Supporting Information Appendix S2.
Dynamic Network Simulation
Using the estimated parameters from fitted models in Equations 2 and 3, we simulated 5,000
dynamic networks: 10 realizations for each one of 10 (randomly selected) dynamic networks
from 50 (randomly selected) individuals. We then calculated descriptive graph measures
including: C, Eglob, and k. Table 4 presents the average values across all dynamic networks
and all regions for both observed and simulated networks.
As evidenced by Table 4, average network measures are very close between simulated and
observed networks, indicating the ability of this model to simulate representative group-level
dynamic networks. However, to further illustrate that simulated networks represent observed
dynamic networks at multiple resolutions beyond average values, that is, at the individual and
nodal level, in Figure 4, we have shown two realizations of two dynamic networks for two
participants (all chosen randomly from the 5,000 simulated networks). Networks in this figure
represent the top 5% of strongest connections. As this figure shows, even after thresholding the
networks, the simulated networks well represent the observed networks.
DISCUSSION
As the interest in dynamic brain networks continues to grow, new methods are needed to
enable gleaning neurobiological insight into this complex and big data. Development of mul-
tivariate statistical methods, particularly model-based ones, which allow quantifying relation-
ships between phenotypic traits and dynamic patterns of brain connectivity and topology, as
well as drawing inference from such relationships, is among the urgent needs. Development of
such methods even for static networks has remained a challenge given the size, complexity,
and multiscale dependence inherent in brain network data. However several model-based
methods (Shehzad et al., 2014; Simpson, Hayasaka, & Laurienti, 2011; Simpson & Laurienti,
2015) and various data-driven multivariate methods (Allen et al., 2011; Beckmann & Smith,
2004; Calhoun, Adali, Pearlson, & Pekar, 2001; Smith, Hyvarinen, Varoquaux, Miller, &
Beckmann, 2014) have been introduced and extensively used for static networks. Dynamic
changes in the systematic organization of our brain networks confer much of our brains’
functions abilities due to the fact that our brain is a complex multiscale dynamic system with
known and unknown compensatory mechanisms at multiple scales. Thus, methods that allow
analyzing the brain within a multivariate framework can provide much deeper insights into
dynamic patterns of brain networks in health and disease. In addition, multivariate model-
based tools enable aligning neuroscientific hypotheses with the analytic approach, which is
ideal for dynamic brain network analysis (Preti, Bolton, & Van De Ville, 2017). Nevertheless,
no model-based multivariate method has been introduced for dynamic network analyses to
our knowledge.
Here we provided a model-based multivariate method to relate phenotypic traits to
dynamic patterns of brain connectivity and topology. We developed this model by advancing
a two-part mixed-effects regression framework for static brain networks (Simpson & Laurienti,
2015). Our proposed model allows accounting for the connectivity/network dynamics when
assessing group differences and phenotype-health outcome relationships, to avoid confound-
ing and drawing erroneous conclusions. The incorporation of endogenous network measures
Network Neuroscience
605
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
n
e
n
_
a
_
0
0
2
3
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
A mixed model for dynamic brain networks
such as clustering coefficient and global efficiency, as independent variables, allows simulta-
neous analyses of connectivity and topology dynamics. There is a long history of modeling a
network as a function of endogenous network metrics to identify how nodal properties are
related to the probability (and strength) of connections (O’Malley, 2013; Robins, Pattison, Kalish,
& Lusher, 2007; Simpson et al., 2011, 2012). Part of the motivation for our modeling framework
was the desire to port this approach into the time-varying multiple-network context and blend
it with the more standard exogenous covariate approach to create a hybrid method that allows
examining and accounting for both an individual’s endogenous network structure and exog-
enous phenotypic characteristics in a manner suitable for dynamic brain network analyses.
The topological network covariates allow examining how nodal properties influence the con-
nection between two brain areas. Having both the endogenous and exogenous covariates in
the model allows us to assess how these topology-connection relationships vary by individual
and group characteristics. Additionally, it allows simulating more realistic dynamic networks
by incorporating the dynamics of an array of explanatory network measures and their interplay
with desired covariates.
Most current methods used to assess dynamic brain networks reduce this data into dynamic
patterns of individual brain connections (Schmlazle et al., 2017; Simony et al., 2016; Tewarie
et al., 2019) or commonly used topological summary variables, such as node degree or mod-
ularity (Jones et al., 2012; Kabbara et al., 2019), rather than analyzing the systemic dynamics
of the brain networks. Such methods not only fail to model the brain as a multiscale dynamic
system (Lurie et al., 2020), but often entail matching study populations to perform group com-
parisons, which is a daunting task for most neuroimaging studies. Our model provides a frame-
work to assess the systemic dynamics of brain networks and thus to account for complex
dynamics of the brain via the simultaneous modeling of brain connectivity and topological
network variables. The multivariate nature of this framework reduces demands for matching
study populations as any number of confounding effects can be incorporated as covariates,
and the effects of multiple covariates of interest can be studied in a single model. Another
important utility of this model is its ability to simulate dynamic brain networks, which is critical
for a better understanding of brain function in health and disease (Tikidji-Hamburyan,
Narayana, Bozkus, & El-Ghazawi, 2017). To our knowledge, our framework is the first model
that allows simulating dynamic brain networks from system-level properties of the brain, and
with respect to desired covariates. An important utility of the simulation capability of this
model is to generate representative group-level dynamic networks. The need for reliably
generating representative group-level networks has been well documented (Jirsa, Sporns,
Breakspear, Deco, & McIntosh, 2010; Meunier, Achard, Morcom, & Bullmore, 2009; Valencia
et al., 2009; Zuo et al., 2012). Additionally, the simulation capability also provides a scientif-
ically appropriate way to assess GOF (as shown in the Dynamic Network Simulation section)
and simulate individual-level networks.
It is important to note that our framework itself, which aims to account for continuous time-
varying network changes when relating covariates to topology, is not designed to identify
latent states like hidden Markov models are, for example. But adding a latent state analysis
to the preprocessing steps prior to implementing our model would provide a complementary
and insightful extension to our overall approach. Our method will work for many approaches
used to generate the networks, allowing for great flexibility in the network generation method
that one chooses to use. A standard approach for assessing the performance of regression-
based methods is through examining the quality of model fit. Since our method is also a
regression-based framework, we used different GOF measures to examine the performance
of our method. More importantly, our simulation analyses allowed for a more profound
Network Neuroscience
606
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
n
e
n
_
a
_
0
0
2
3
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
A mixed model for dynamic brain networks
assessment of the performance of our method as the most appropriate method to assess the
GOF of statistical methods in the network context is through simulation analyses (Hunter
et al., 2008). In addition, with respect to identifying the association between phenotypic traits
and dynamic brain networks, our method uses a fundamentally different approach, and pro-
vides different (complementary) insight, than current data-driven methods, and thus a compar-
ison between our model and current data-driven models would not be appropriate.
We demonstrated the utility of our model in identifying the relationship between fluid intel-
ligence and dynamic patterns of brain connectivity and topological network variables by using
the rich data set provided by the HCP study (Van Essen et al., 2013). Our model allowed
accounting for various sources of potential confounding effects, such as sex, education,
age, and alcohol abuse, among others. Our results indicated that dynamic patterns of
whole-brain modularity and connection strength are significantly affected by fluid intelligence.
More specifically, our results showed that for any level of fluid intelligence, dynamic patterns
of modularity are predominantly associated with between-community, rather than within-
community, connections. However, fluid intelligence modulates this trend such that, across
an entire spectrum of fluid intelligence, dynamics of whole-brain modularity play a less impor-
tant role in driving changes in between-community connections for higher fluid intelligence
values (with dynamics of within-community connections probably being affected more).
While the ultimate neurobiological interpretations of such effects is speculative at this point,
our results may suggest that at lower levels of intelligence, distinct network modules necessary
for cognition (such as the module comprised of areas forming the central executive attention
network, or CEN) are formed primarily by segregating information. Thus, distinct subnetworks
are formed by decreasing connectivity to other subnetworks, which could result in relatively
poor distribution of information between subnetworks. As intelligence increases, the formation
of distinct modules is driven more by strengthening connections within the module and less by
segregating modules. Increased intramodular connectivity could enhance processing within
subnetworks like the CEN while maintaining communication between subnetworks for opti-
mal information distribution. Other studies have reported associations between brain modu-
larity and intelligence (Chaddock-Heyman et al., 2020), as well as significant correlations
between creativity and learning and dynamic patterns of brain modularity (Bassett et al.,
2011; Kenett, Betzel, & Beaty, 2020).
In the absence of a “gold standard” in sliding window approach, the optimal choice for
window type, window length, and step size is challenging. We used a modulated rectangular
window due to its superior performance in examining dynamic brain networks when com-
pared to other conventional window types (Mokhtari, Akhlaghi et al., 2019). We used 120
volumes for our window length for multiple reasons, including (1) to provide more stabilized
correlation values while not losing the variability of the brain dynamics, (2) due to its wider use
which makes comparing and contrasting our method with currently used methods easier, and
(3) model fit and convergence considerations of our proposed method. It is also important to
note that no commonly used window length can accurately identify different states of corre-
lation (Shakil, Lee, & Keilholz, 2016), and that the sliding window correlation approach is only
used to demonstrate the utility of our method rather than to provide comprehensive analyses of
fluid intelligence-dynamic brain network associations. Also, typical shift sizes used in the lit-
erature range from 1 TR to 50% of the window length (Chang et al., 2013; Kucyi & Davis,
2014; Shakil, Magnuson, Keilholz, & Lee, 2014), with the 1 TR being the most commonly used
shift size (Shakil et al., 2016). However, as our proposed method subsequently uses the
dynamic networks in a regression framework, we used a shift size of 120 volumes, equal to
the window length, to create nonoverlapping windows and thus further reduce
Network Neuroscience
607
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
n
e
n
_
a
_
0
0
2
3
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
A mixed model for dynamic brain networks
autocorrelation. Additionally, using nonoverlapping windows allowed using a smaller, but suf-
ficient number of dynamic networks for each participant and thus helped avoiding possible
convergence issues. Nevertheless, for windows with 50% overlap between consecutive net-
works, our modeling framework yielded similar outcomes.
Incorporating all brain connections and modeling the dependence among multiple vari-
ables across time is computationally time intensive and can lead to convergence issues for
datasets with large numbers of participants and regions of interest. This prevented us from
using more complex variance-covariance structures for the random effects that could, in turn,
yield even more accurate estimates and better simulations of dynamic brain networks. We
plan to develop complementary data reduction methods to address this in future work. More-
over, there is no agreement on a well-accepted atlas for functional connectivity studies. While
many studies have used clustering-based methods or canonical correlation analysis to define
nodes for their own data (Allen et al., 2014), other studies have used publicly available par-
cellation schemes generated from rich datasets with higher reproducibility across individuals
(Glasser et al., 2016). Even newer studies indicate that using an individual atlas rather than a
group-level parcellation may provide more reliable results (Salehi et al., 2020). However, as
with the parameters used for generating the dynamic networks (sliding window type, length,
etc.), our method is independent of the parcellation scheme used for defining the brain
regions. The sensitivity to the method used to generate dynamic networks as well as the par-
cellation scheme (Bahramf & Hossein-Zadeh, 2014; Glasser et al., 2016; Power et al., 2011)
will be assessed in future studies as for each parameter, multiple models should be run to find
the best orthonormal polynomial degree for that particular parameter, and thus, a composite
grid search over all combinations of such parameters and polynomial degrees will be required,
which is beyond the scope of this paper. The proposed mixed-effects framework can be used
for predicting dynamic networks based on participant characteristics as well. However, we
didn’t demonstrate this capability here. These capabilities will be demonstrated in future work,
given that they require extensive analytical assessment and exposition, which lie beyond the
scope of this paper. We have also made our proposed framework accessible to neuroimaging
researchers by incorporating new graphical user interfaces into WFU_MMNET (https://www
.nitrc.org/projects/wfu_mmnet), the software developed for the application of the original static
model (Bahrami et al., 2019b).
This study is not without limitations. Incorporating all brain connections as well modeling
the dependence structure between multiple variables across individuals’ brain connections
and network metrics (e.g., clustering coefficient, global efficiency) across time is computation-
ally intensive and can lead to convergence issues (particularly for the probability model; Equa-
tion 2) for datasets with large numbers of participants and network nodes. This precludes using
our method for studies analyzing voxel level dynamic networks and limits its utility for studies
with very large numbers of subjects or very spatially resolved (>1,000 nodes) brain networks.
We have developed a clustering-based data reduction method to resolve this issue for the orig-
inal model for static networks and are currently working on extending it to be applicable for
dynamic brain networks as well. This will greatly aid in resolving the convergence issue. Also,
parcellation schemes generated from a clustering or canonical correlation analysis approach
with a fewer number of regions of interest are also appealing alternatives to be used within our
framework. Interpreting the results produced by our method may not always be straightfor-
ward, as both connectivity and network topology dynamics should be considered simulta-
neously. This can complicate interpretation of the results for continuous variables and when
particular network metrics with both local and global implications are examined, such as clus-
tering coefficient. Another limitation of our method is that its only applicable for whole-brain
Network Neuroscience
608
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
networks at present, and not for analyzing dynamics of local brain networks or subnetworks
(per esempio., the default mode network) unless they are extracted from the whole-brain network. How-
ever, we are working on devising a procedure to allow for analyzing dynamics of local brain
networks within the context of their whole networks as was done for the original framework for
static networks in Bahrami et al. (2019UN). Finalmente, future studies should conduct a thorough
sensitivity analysis of our framework with respect to the parameters used in generating the
dynamics networks and the parcellation scheme used.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00238.
AUTHOR CONTRIBUTIONS
Mohsen Bahrami: Data curation; Formal analysis; Methodology; Validation; Visualization;
Writing – original draft; Writing – review & editing. Paul J. Laurienti: Conceptualization; Fund-
ing acquisition; Validation; Writing – review & editing. Heather M. Shappell: Conceptualiza-
zione; Writing – review & editing. Dale Dagenbach: Conceptualization; Writing – review &
editing. Sean L. Simpson: Conceptualization; Funding acquisition; Methodology; Supervision;
Validation; Writing – original draft; Writing – review & editing.
FUNDING INFORMATION
Sean L. Simpson, National Institute of Biomedical Imaging and Bioengineering (https://
grantome.com/grant/NIH/R01-EB024559-04), Award ID: R01EB024559.
REFERENCES
Akaike, H. (1981). A new look at the statistical-model identification.
Current Contents/Engineering Technology & Applied Sciences,
12(51), 22.
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T., &
Calhoun, V. D. (2014). Tracking whole-brain connectivity
dynamics in the resting state. Cerebral Cortex, 24(3), 663–676.
https://doi.org/10.1093/cercor/bhs352, PubMed: 23146964
Allen, E. A., Erhardt, E. B., Damaraju, E., Gruner, W., Segall, J. M.,
Silva, R. F., … Calhoun, V. D. (2011). A baseline for the multivar-
iate comparison of resting-state networks. Frontiers in Systems
Neuroscience, 5, 2. https://doi.org/10.3389/fnsys.2011.00002,
PubMed: 21442040
Bahramf, M., & Hossein-Zadeh, G.-A. (2014). Functional parcellations
affect the network measures in graph analysis of resting-state fMRI.
Paper presented at the 2014 21st Iranian Conference on Biomed-
ical Engineering (ICBME). https://doi.org/10.1109/ICBME.2014
.7043933
Bahrami, M., Laurienti, P. J., & Simpson, S. l. (2019UN). Analysis of
brain subnetworks within the context of their whole-brain net-
works. Human Brain Mapping, 40(17), 5123–5141. https://doi
.org/10.1002/hbm.24762, PubMed: 31441167
Bahrami, M., Laurienti, P. J., & Simpson, S. l. (2019B). A MATLAB
toolbox for multivariate analysis of brain networks. Human Brain
Mapping, 40(1), 175–186. https://doi.org/10.1002/hbm.24363,
PubMed: 30256496
Barttfeld, P., Uhrig, L., Sitt, J. D., Sigman, M., Jarraya, B., &
Dehaene, S. (2015). Signature of consciousness in the dynamics
of resting-state brain activity. Proceedings of the National Academy
of Sciences of the United States of America, 112(3), 887–892.
https://doi.org/10.1073/pnas.1418031112, PubMed: 25561541
Bassett, D. S., & Bullmore, E. T. (2009). Human brain networks in
health and disease. Current Opinion in Neurology, 22(4), 340–347.
https://doi.org/10.1097/wco.0b013e32832d93dd, PubMed:
19494774
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of human
brain networks during learning. Proceedings of the National
Academy of Sciences of the United States of America, 108(18),
7641–7646. https://doi.org/10.1073/pnas.1018985108,
PubMed: 21502525
Beckmann, C. F., & Smith, S. UN. (2004). Probabilistic independent
component analysis for functional magnetic resonance imaging.
IEEE Transactions on Medical Imaging, 23(2), 137–152. https://
doi.org/10.1109/TMI.2003.822821, PubMed: 14964560
Benjamini, Y., & Hochberg, Y. (2000). On the adaptive control of
the false discovery rate in multiple testing with independent
statistics. Journal of Educational and Behavioral Statistics, 25(1),
60–83. https://doi.org/10.3102/10769986025001060
Bilker, W. B., Hansen, J. A., Brensinger, C. M., Richard, J., Gur,
R. E., & Gur, R. C. (2012). Development of abbreviated
Network Neuroscience
609
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
nine-item forms of the Ravenʼs Standard Progressive Matrices
test. Assessment, 19(3), 354–369. https://doi.org/10.1177
/1073191112446655, PubMed: 22605785
Bozdogan, H. J. P. (1987). Model selection and Akaikeʼs information
criterion (AIC): The general theory and its analytical extensions. Psy-
chometrika, 52(3), 345–370. https://doi.org/10.1007/BF02294361
Breakspear, M. (2017). Dynamic models of large-scale brain activ-
ità. Nature Neuroscience, 20(3), 340–352. https://doi.org/10
.1038/nn.4497, PubMed: 28230845
Bressler, S. L., & Menon, V. (2010). Large-scale brain networks in
cognition: Emerging methods and principles. Trends in Cognitive
Scienze, 14(6), 277–290. https://doi.org/10.1016/j.tics.2010.04
.004, PubMed: 20493761
Bullmore, E. T., & Sporns, O. (2009). Complex brain networks:
Graph theoretical analysis of structural and functional systems.
Nature Reviews Neuroscience, 10(3), 186–198. https://doi.org
/10.1038/nrn2575, PubMed: 19190637
Buzsaki, G., & Draguhn, UN. (2004). Neuronal oscillations in cortical
networks. Scienza, 304(5679), 1926–1929. https://doi.org/10
.1126/science.1099745, PubMed: 15218136
Calhoun, V. D., Adali, T., Pearlson, G. D., & Pekar, J. J. (2001). UN
method for making group inferences from functional MRI data
using independent component analysis. Human Brain Mapping,
14(3), 140–151. https://doi.org/10.1002/ hbm.1048, PubMed:
11559959
Cattell, R. B. (1987). Intelligenza: Its structure, growth and action.
Amsterdam, the Netherlands: Elsevier.
Chaddock-Heyman, L., Weng, T. M. B., Kienzler, C., Weisshappel,
R., Drollette, E. S., Raine, l. B., … Kramer, UN. F. (2020). Brain net-
work modularity predicts improvements in cognitive and scho-
lastic performance in children involved in a physical activity
intervention. Frontiers in Human Neuroscience, 14. https://doi
.org/10.3389/fnhum.2020.00346, PubMed: 33100988
Chang, C., & Glover, G. H. (2010). Time-frequency dynamics of
resting-state brain connectivity measured with fMRI. Neuro-
Image, 50(1), 81–98. https://doi.org/10.1016/j.neuroimage.2009
.12.011, PubMed: 20006716
Chang, C., Liu, Z. M., Chen, M. C., Liu, X., & Duyn, J. H. (2013).
EEG correlates of time-varying BOLD functional connectivity.
NeuroImage, 72, 227–236. https://doi.org/10.1016/j.neuroimage
.2013.01.049, PubMed: 23376790
Cole, M. W., Bassett, D. S., Energia, J. D., Braver, T. S., & Petersen,
S. E. (2014). Intrinsic and task-evoked network architectures of
the human brain. Neuron, 83(1), 238–251. https://doi.org/10
.1016/j.neuron.2014.05.014, PubMed: 24991964
Colom, R., & Flores-Mendoza, C. E. (2007). Intelligence predicts
scholastic achievement irrespective of SES factors: Evidence from
Brasile. Intelligenza, 35(3), 243–251. https://doi.org/10.1016/j
.intell.2006.07.008
Diez-Cirarda, M., Strafella, UN. P., Kim, J., Pena,
J., Ojeda, N.,
Cabrera-Zubizarreta, A., & Ibarretxe-Bilbao, N. (2018). Dynamic
functional connectivity in Parkinsonʼs disease patients with mild
cognitive impairment and normal cognition. NeuroImage –
Clinical, 17, 847–855. https://doi.org/10.1016/j.nicl.2017.12
.013, PubMed: 29527489
Edwards, l. J., & Simpson, S. l. (2014). An analysis of 24-h ambulatory
blood pressure monitoring data using orthonormal polynomials
in the linear mixed model. Blood Pressure Monitoring, 19(3),
153–163. https://doi.org/10.1097/ MBP.0000000000000039,
PubMed: 24667908
Elton, A., & Gao, W. (2015). Task-related modulation of functional
connectivity variability and its behavioral correlations. Umano
Brain Mapping, 36(8), 3260–3272. https://doi.org/10.1002/hbm
.22847, PubMed: 26015070
Friedman, E. J., Landsberg, UN. S., Owen, J. P., Li, Y.-O., & Mukherjee,
P. (2014). Stochastic geometric network models for groups of
functional and structural connectomes. NeuroImage, 101,
473–484. https://doi.org/10.1016/j.neuroimage.2014.07.039,
PubMed: 25067815
Fukushima, M., Betzel, R. F., Lui, Y., de Reus, M. A., van den
Heuvel, M. P., Zuo, X. N., & Sporns, O. (2018). Fluctuations
between high- and low-modularity topology in time-resolved
functional connectivity. NeuroImage, 180, 406–416. https://doi
.org/10.1016/j.neuroimage.2017.08.044, PubMed: 28823827
Glasser, M. F., Coalson, T. S., Robinson, E. C., Hacker, C. D., Harwell,
J., Yacoub, E., … Van Essen, D. C. (2016). A multi-modal parcellation
of human cerebral cortex. Nature, 536(7615), 171–178. https://doi
.org/10.1038/nature18933, PubMed: 27437579
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S.,
Fischl, B., Andersson, J. L., … for the WU-Minn HCP Consortium.
(2013). The minimal preprocessing pipelines for the Human
Connectome Project. NeuroImage, 80, 105–124. https://doi.org
/10.1016/j.neuroimage.2013.04.127, PubMed: 23668970
Godwin, D., Barry, R. L., & Marois, R. (2015). Breakdown of the
brainʼs functional network modularity with awareness. Proceedings
of the National Academy of Sciences of the United States of
America, 112(12), 3799–3804. https://doi.org/10.1073/pnas
.1414466112, PubMed: 25759440
Gonzalez-Castillo, J., Hoy, C. W., Handwerker, D. A., Robinson,
M. E., Buchanan, l. C., Saad, Z. S., & Bandettini, P. UN. (2015).
Tracking ongoing cognition in individuals using brief, whole-brain
functional connectivity patterns. Proceedings of the National
Academy of Sciences of the United States of America, 112(28),
8762–8767. https://doi.org/10.1073/pnas.1501242112,
PubMed: 26124112
Gu, Y., Lin, Y., Huang, l. L., Mamma, J. J., Zhang, J. B., Xiao, Y., …
Alzheimer’s Disease Neuroimaging Initiative. (2020). Abnormal
dynamic functional connectivity in Alzheimerʼs disease. CNS
Neuroscience & Therapeutics, 26(9), 962–971. https://doi.org
/10.1111/cns.13387, PubMed: 32378335
Handwerker, D. A., Roopchansingh, V., Gonzalez-Castillo, J., &
Bandettini, P. UN. (2012). Periodic changes in fMRI connectivity.
NeuroImage, 63(3), 1712–1719. https://doi.org/10.1016/j
.neuroimage.2012.06.078, PubMed: 22796990
Hannan, E. J., & Quinn, B. G. (1979). Determination of the order of
an autoregression. Journal of the Royal Statistical Society Series
B-Methodological, 41(2), 190–195. https://doi.org/10.1111/j
.2517-6161.1979.tb01072.x
Honari, H., Choe, UN. S., Pekar, J. J., & Lindquist, M. UN. (2019).
Investigating the impact of autocorrelation on time-varying con-
nectivity. NeuroImage, 197, 37–48. https://doi.org/10.1016/j
.neuroimage.2019.04.042, PubMed: 31022568
Hunter, D. R., Goodreau, S. M., & Handcock, M. S. (2008). Good-
ness of fit of social network models. Journal of the American
Statistical Association, 103(481), 248–258. https://doi.org/10
.1198/016214507000000446
Network Neuroscience
610
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
Hurvich, C. M., & Tsai, C. l. (1989). Regression and time-series
model selection in small samples. Biometrika, 76(2), 297–307.
https://doi.org/10.1093/biomet/76.2.297
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M., … Chang, C. (2013). Dynamic
functional connectivity: Promise, issues, and interpretations.
N e u ro I m a g e , 8 0 , 3 6 0 – 3 7 8 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neuroimage.2013.05.079, PubMed: 23707587
Jin, C. F., Jia, H., Lanka, P., Rangaprakash, D., Li, l. J., Liu, T. M., …
Deshpande, G. (2017). Dynamic brain connectivity is a better
predictor of PTSD than static connectivity. Human Brain
Mapping, 38(9), 4479–4496. https://doi.org/10.1002/ hbm
.23676, PubMed: 28603919
Jirsa, V. K., Sporns, O., Breakspear, M., Deco, G., & McIntosh, UN. R.
(2010). Towards the virtual brain: Network modeling of the intact
and the damaged brain. Archives Italiennes de Biologie, 148(3),
189–205. https://doi.org/10.4449/aib.v148i3.1223, PubMed:
21175008
Jones, D. T., Vemuri, P., Murphy, M. C., Gunter, J. L., Senjem, M. L.,
Machulda, M. M., … Jack, C. R. (2012). Non-stationarity in the
“resting brainʼs” modular architecture. PLoS One, 7(6). https://
doi.org/10.1371/journal.pone.0039731, PubMed: 22761880
Joyce, K. E., Laurienti, P. J., Burdette, J. H., & Hayasaka, S. (2010). UN
new measure of centrality for brain networks. PLoS One, 5(8).
https://doi.org/10.1371/journal.pone.0012200, PubMed:
20808943
Kabbara, A., Khalil, M., OʼNeill, G., Dujardin, K., El Traboulsi, Y.,
Wendling, F., & Hassan, M. (2019). Detecting modular brain
states in rest and task. Network Neuroscience, 3(3), 878–901.
https://doi.org/10.1162/netn_a_00090, PubMed: 31410384
Kenett, Y. N., Betzel, R. F., & Beaty, R. E. (2020). Community struc-
ture of the creative brain at rest. NeuroImage, 210. https://doi.org
/10.1016/j.neuroimage.2020.116578, PubMed: 31982579
Kucyi, A., & Davis, K. D. (2014). Dynamic functional connectivity
of the default mode network tracks daydreaming. NeuroImage,
100, 471–480. https://doi.org/10.1016/j.neuroimage.2014.06
.044, PubMed: 24973603
Liu, X. (2017). Dynamic fcMRI: Approaches. Sunrise Educational
Session: Dynamic Functional Connectivity MRI: Approaches &
Mechanisms. Proceedings of the International Society for
Magnetic Resonance in Medicine.
Lungo, Y. C., Cao, H. Y., Yan, C. G., Chen, X., Li, L., Castellanos,
F. X., … Liu, Z. N. (2020). Altered resting-state dynamic func-
tional brain networks in major depressive disorder: Findings from
the REST-meta-MDD consortium. NeuroImage: Clinical, 26.
https://doi.org/10.1016/j.nicl.2020.102163, PubMed: 31953148
Lurie, D. J., Kessler, D., Bassett, D. S., Betzel, R. F., Breakspear, M.,
Kheilholz, S., … Calhoun, V. D. (2020). Questions and controver-
sies in the study of time-varying functional connectivity in resting
fMRI. Network Neuroscience, 4(1), 30–69. https://doi.org/10
.1162/netn_a_00116, PubMed: 32043043
Martinez, S. A., Deco, G., Ter Horst, G. J., & Cabral, J. (2020).
The dynamics of functional brain networks associated with
depressive symptoms in a nonclinical sample. Frontiers in Neural
Circuits, 14. https://doi.org/10.3389/fncir.2020.570583, PubMed:
33071760
Meunier, D., Achard, S., Morcom, A., & Bullmore, E. T. (2009).
Age-related changes in modular organization of human brain
functional networks. NeuroImage, 44(3), 715–723. https://doi
.org/10.1016/j.neuroimage.2008.09.062, PubMed: 19027073
Mokhtari, F., Akhlaghi, M. I., Simpson, S. L., Wu, G. R., & Laurienti,
P. J. (2019). Sliding window correlation analysis: Modulating
window shape for dynamic brain connectivity in resting state.
NeuroImage, 189, 655–666. https://doi.org/10.1016/j
.neuroimage.2019.02.001, PubMed: 30721750
Mokhtari, F., Laurienti, P. J., Rejeski, W. J., & Ballard, G. (2019).
Dynamic functional magnetic resonance imaging connectivity
tensor decomposition: A new approach to analyze and interpret
dynamic brain connectivity. Brain Connectivity, 9(1), 95–112.
https://doi.org/10.1089/brain.2018.0605, PubMed: 30318906
Oldfield, R. C. (1971). The assessment and analysis of handedness:
The Edinburgh Inventory. Neuropsychologia, 9(1), 97–113. https://
doi.org/10.1016/0028-3932(71)90067-4, PubMed: 5146491
O’Malley, UN. J. (2013). The analysis of social network data: An
exciting frontier for statisticians. Statistics In Medicine, 32(4),
539–555. https://doi.org/10.1002/sim.5630, PubMed: 23023735
Parente, F., Frascarelli, M., Mirigliani, A., Di Fabio, F., Biondi, M., &
Colosimo, UN. (2018). Negative functional brain networks. Brain
Imaging and Behavior, 12(2), 467–476. https://doi.org/10.1007
/s11682-017-9715-x, PubMed: 28353136
Park, H.-J., & Friston, K. J. (2013). Structural and functional brain
networks: From connections to cognition. Scienza, 342(6158).
https://doi.org/10.1126/science.1238411, PubMed: 24179229
Parr, T., Rees, G., & Friston, K. J. (2018). Computational neuropsy-
chology and Bayesian inference. Frontiers in Human Neurosci-
ence, 12. https://doi.org/10.3389/fnhum.2018.00061, PubMed:
29527157
Energia, J. D., Cohen, UN. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., … Petersen, S. E. (2011). Functional network orga-
nization of the human brain. Neuron, 72(4), 665–678. https://doi
.org/10.1016/j.neuron.2011.09.006, PubMed: 22099467
Preti, M. G., Bolton, T. UN. W., & Van De Ville, D. (2017). IL
dynamic functional connectome: State-of-the-art and perspec-
tives. NeuroImage, 160, 41–54. https://doi.org/10.1016/j
.neuroimage.2016.12.061, PubMed: 28034766
Pruim, R. H. R., Mennes, M., van Rooij, D., Llera, A., Buitelaar, J. K.,
& Beckmann, C. F. (2015). ICA-AROMA: A robust ICA-based
strategy for removing motion artifacts from fMRI data. Neuro-
Image, 112, 267–277. https://doi.org/10.1016/j.neuroimage
.2015.02.064, PubMed: 25770991
Rashid, B., Arbabshirani, M. R., Damaraju, E., Cetin, M. S., Mugnaio,
R., Pearlson, G. D., & Calhoun, V. D. (2016). Classification of
schizophrenia and bipolar patients using static and dynamic
resting-state fMRI brain connectivity. NeuroImage, 134, 645–657.
https://doi.org/10.1016/j.neuroimage.2016.04.051, PubMed:
27118088
Robins, G. L., Pattison, P. E., Kalish, Y., & Lusher, D. (2007). An
introduction to exponential random graph (p*) models for social
networks. Social Networks, 29(2), 173–191. https://doi.org/10
.1016/j.socnet.2006.08.002
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Sakoglu, U., Pearlson, G. D., Kiehl, K. A., Wang, Y. M., Michael,
UN. M., & Calhoun, V. D. (2010). A method for evaluating
Network Neuroscience
611
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
dynamic functional network connectivity and task-modulation:
application to schizophrenia. Magnetic Resonance Materials in
Physics Biology and Medicine, 23(5–6), 351–366. https://doi
.org/10.1007/s10334-010-0197-8, PubMed: 20162320
Salehi, M., Greene, UN. S., Karbasi, A., Shen, X. L., Scheinost, D., &
Constable, R. T. (2020). There is no single functional atlas even
for a single individual: Functional parcel definitions change with
task. NeuroImage, 208. https://doi.org/10.1016/j.neuroimage
.2019.116366, PubMed: 31740342
Schmlazle, R., OʼDonnell, M. B., Garcia, J. O., Cascio, C. N.,
Bayer, J., Bassett, D. S., … Falk, E. B. (2017). Brain connectivity
dynamics during social interaction reflect social network struc-
ture. Proceedings of the National Academy of Sciences of the
United States of America, 114(20), 5153–5158. https://doi.org
/10.1073/pnas.1616130114, PubMed: 28465434
Schwarz, UN. J., & McGonigle, J. (2011). Negative edges and soft
thresholding in complex network analysis of resting state func-
tional connectivity data. NeuroImage, 55(3), 1132–1146.
https://doi.org/10.1016/j.neuroimage.2010.12.047, PubMed:
21194570
Schwarz, G. (1978). Estimating the dimension of a model. Annals of
Statistics, 6(2), 461–464. https://doi.org/10.1214/aos/1176344136
Shakil, S., Lee, C. H., & Keilholz, S. D. (2016). Evaluation of sliding
window correlation performance for characterizing dynamic
functional connectivity and brain states. NeuroImage, 133,
111–128. https://doi.org/10.1016/j.neuroimage.2016.02.074,
PubMed: 26952197
Shakil, S., Magnuson, M. E., Keilholz, S. D., & Lee, C. H. (2014).
Cluster-based analysis for characterizing dynamic functional
connectivity. Annual International Conference of the IEEE Engi-
neering in Medicine and Biology Society, 2014, 982–985.
https://doi.org/10.1109/ EMBC.2014.6943757, PubMed:
25570125
Shappell, H., Caffo, B. S., Pekar, J. J., & Lindquist, M. UN. (2019).
Improved state change estimation in dynamic functional connec-
tivity using hidden semi-Markov models. NeuroImage, 191,
243–257. https://doi.org/10.1016/j.neuroimage.2019.02.013,
PubMed: 30753927
Shehzad, Z., Kelly, C., Reiss, P. T., Craddock, R. C., Emerson, J. W.,
McMahon, K., … Milham, M. P. (2014). A multivariate
distance-based analytic framework for connectome-wide associ-
ation studies. NeuroImage, 93, 74–94. https://doi.org/10.1016/j
.neuroimage.2014.02.024, PubMed: 24583255
Shen, X., Tokoglu, F., Papademetris, X., & Constable, R. T. (2013).
Groupwise whole-brain parcellation from resting-state fMRI data
for network node identification. NeuroImage, 82, 403–415.
https://doi.org/10.1016/j.neuroimage.2013.05.081, PubMed:
23747961
Shine, J. M., Bissett, P. G., Campana, P. T., Koyejo, O., Balsters, J. H.,
Gorgolewski, K. J., … Poldrack, R. UN. (2016). The dynamics of
functional brain networks: Integrated network states during cog-
nitive task performance. Neuron, 92(2), 544–554. https://doi.org
/10.1016/j.neuron.2016.09.018, PubMed: 27693256
Shine, J. M., Breakspear, M., Campana, P. T., Martens, K. UN. E., Shine, R.,
Koyejo, O., … Poldrack, R. UN. (2019). Human cognition involves
the dynamic integration of neural activity and neuromodulatory
systems. Nature Neuroscience, 22(6), 1036–1036. https://doi.org
/10.1038/s41593-019-0347-x, PubMed: 30792538
Shine, J. M., & Poldrack, R. UN. (2018). Principles of dynamic net-
work reconfiguration across diverse brain states. NeuroImage,
180, 396–405. https://doi.org/10.1016/j.neuroimage.2017.08
.010, PubMed: 28782684
Shirer, W. R., Ryali, S., Rykhlevskaia, E., Menon, V., & Greicius, M. D.
(2012). Decoding subject-driven cognitive states with whole-brain
connectivity patterns. Cerebral Cortex, 22(1), 158–165. https://doi
.org/10.1093/cercor/bhr099, PubMed: 21616982
Simony, E., Honey, C. J., Chen, J., Lositsky, O., Yeshurun, Y., Wiesel, A.,
& Hasson, U. (2016). Dynamic reconfiguration of the default mode
network during narrative comprehension. Nature Communications,
7. https://doi.org/10.1038/ncomms12141, PubMed: 27424918
Simpson, S. L., Bahrami, M., & Laurienti, P. J. (2019). UN
mixed-modeling framework for analyzing multitask whole-brain
network data. Network Neuroscience, 3(2), 307–324. https://doi
.org/10.1162/netn_a_00065, PubMed: 30793084
Simpson, S. L., Bowman, F. D., & Laurienti, P. J. (2013). Analyzing
complex functional brain networks: Fusing statistics and network
science to understand the brain. Statistics Surveys, 7, 1–36.
https://doi.org/10.1214/13-SS103, PubMed: 25309643
Simpson, S. L., & Edwards, l. J. (2013). A circular LEAR correlation
structure for cyclical longitudinal data. Statistical Methods in
Medical Research, 22(3), 296–306. https://doi.org/10.1177
/0962280210395741, PubMed: 21216801
Simpson, S. L., Hayasaka, S., & Laurienti, P. J. (2011). Exponential
random graph modeling for complex brain networks. PLoS One,
6(5). https://doi.org/10.1371/journal.pone.0020039, PubMed:
21647450
Simpson, S. L., & Laurienti, P. J. (2015). A two-part mixed-effects
modeling framework for analyzing whole-brain network data.
NeuroImage, 113, 310–319. https://doi.org/10.1016/j
.neuroimage.2015.03.021, PubMed: 25796135
Simpson, S. L., & Laurienti, P. J. (2016). Disentangling brain graphs:
A note on the conflation of network and connectivity analyses.
Brain Connectivity, 6(2), 95–98. https://doi.org/10.1089/ brain
.2015.0361, PubMed: 26414952
Simpson, S. L., Moussa, M. N., & Laurienti, P. J. (2012). An exponen-
tial random graph modeling approach to creating group-based
representative whole-brain connectivity networks. NeuroImage,
60(2), 1117–1126. https://doi.org/10.1016/j.neuroimage.2012
.01.071, PubMed: 22281670
Sizemore, UN. E., & Bassett, D. S. (2018). Dynamic graph metrics:
Tutorial, toolbox, and tale. NeuroImage, 180, 417–427. https://
doi.org/10.1016/j.neuroimage.2017.06.081, PubMed: 28698107
Smith, S. M., Hyvarinen, A., Varoquaux, G., Mugnaio, K. L., & Beckmann,
C. F. (2014). Group-PCA for very large fMRI datasets. NeuroImage,
101, 738–749. https://doi.org/10.1016/j.neuroimage.2014.07.051,
PubMed: 25094018
Telesford, Q. K., Simpson, S. L., Burdette, J. H., Hayasaka, S., &
Laurienti, P. J. (2011). The brain as a complex system: Using
network science as a tool for understanding the brain. Brain
Connectivity, 1(4), 295–308. https://doi.org/10.1089/brain.2011
.0055, PubMed: 22432419
Tewarie, P., Liuzzi, L., OʼNeill, G. C., Quinn, UN. J., Griffa, A., Woolrich,
M. W., … Brookes, M. J. (2019). Tracking dynamic brain networks
using high temporal resolution MEG measures of functional con-
nectivity. NeuroImage, 200, 38–50. https://doi.org/10.1016/j
.neuroimage.2019.06.006, PubMed: 31207339
Network Neuroscience
612
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
A mixed model for dynamic brain networks
Tikidji-Hamburyan, R. A., Narayana, V., Bozkus, Z., & El-Ghazawi,
T. UN. (2017). Software for brain network simulations: A compar-
ative study. Frontiers in Neuroinformatics, 11. https://doi.org/10
.3389/fninf.2017.00046, PubMed: 28775687
Unsworth, N., Fukuda, K., Awh, E., & Vogel, E. K. (2014). Working
memory and fluid intelligence: Capacity, attention control, and sec-
ondary memory retrieval. Cognitive Psychology, 71, 1–26. https://
doi.org/10.1016/j.cogpsych.2014.01.003, PubMed: 24531497
Valencia, M., Pastor, M. A., Fernandez-Seara, M. A., Artieda, J.,
Martinerie, J., & Chavez, M. (2009). Complex modular structure
of large-scale brain networks. Chaos, 19(2). https://doi.org/10
.1063/1.3129783, PubMed: 19566254
Van Essen, D. C., Smith, S. M., Barch, D. M., Behrens, T. E. J.,
Yacoub, E., Ugurbil, K., & WU-Minn HCP Consortium. (2013).
The WU-Minn Human Connectome Project: An overview. Neu-
roImage, 80, 62–79. https://doi.org/10.1016/j.neuroimage.2013
.05.041, PubMed: 23684880
Vidaurre, D., Abeysuriya, R., Becker, R., Quinn, UN. J., Alfaro-Almagro,
F., Smith, S. M., & Woolrich, M. W. (2018). Discovering dynamic
brain networks from big data in rest and task. NeuroImage,
180(Pt B), 646–656. https://doi.org/10.1016/j.neuroimage.2017
.06.077, PubMed: 28669905
Vidaurre, D., Hunt, l. T., Quinn, UN. J., Hunt, B. A., Brookes, M. J.,
Nobre, UN. C., & Woolrich, M. W. (2018). Spontaneous cortical
activity transiently organises into frequency specific phase-
coupling networks. Nature Communications, 9(1), 1–13. https://
doi.org/10.1038/s41467-018-05316-z, PubMed: 30061566
Vidaurre, D., Smith, S. M., & Woolrich, M. W. (2017). Brain net-
work dynamics are hierarchically organized in time. Proceedings
of the National Academy of Sciences of the United States of
America, 114(48), 12827–12832. https://doi.org/10.1073/pnas
.1705120114, PubMed: 29087305
Wolfinger, R., & Oconnell, M. (1993). Generalized linear mixed
models a pseudo-likelihood approach. Journal of Statistical Com-
putation and Simulation, 48(3–4), 233–243. https://doi.org/10
.1080/00949659308811554
Ye, T., Li, P., Zhang, Q., Gu, Q., Lu, X. Q., Gao, Z. F., & Shen,
M. W. (2019). Relation between working memory capacity of
biological movements and fluid intelligence. Frontiers in Psy-
chology, 10. https://doi.org/10.3389/fpsyg.2019.02313,
PubMed: 31749726
Zhu, H., Huang, J., Deng, l. F., Lui, N. Y., Cheng, L., Shu, P., … Ling,
H. W. (2019). Abnormal dynamic functional connectivity associ-
ated with subcortical networks in Parkinsonʼs disease: A tempo-
ral variability perspective. Frontiers in Neuroscience, 13. https://
doi.org/10.3389/fnins.2019.00080, PubMed: 30837825
Zuo, X. N., Ehmke, R., Mennes, M., Imperati, D., Castellanos, F. X.,
Sporns, O., & Milham, M. P. (2012). Network centrality in the human
functional connectome. Cerebral Cortex, 22(8), 1862–1875.
https://doi.org/10.1093/cercor/bhr269, PubMed: 21968567
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
9
1
2
0
2
8
1
1
3
N
e
N
_
UN
_
0
0
2
3
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Network Neuroscience
613