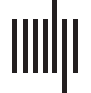RESEARCH
Normalization effect of levodopa on hierarchical
brain function in Parkinson’s disease
Tao Guo1#, Min Xuan1#, Cheng Zhou1, Jingjing Wu1, Ting Gao2, Xueqin Bai1, Xiaocao Liu1,
Luyan Gu2, Ruiqi Liu3, Zhe Song2, Quanquan Gu1, Peiyu Huang1, Jiali Pu2,
Baorong Zhang2, Xiaojun Xu1, Xiaojun Guan1, and Minming Zhang1
1Department of Radiology, Second Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, China
2Department of Neurology, Second Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, China
3School of Information Science and Technology, ShanghaiTech University, Shanghai, China
# Tao Guo and Min Xuan should be considered joint first author.
a n o p e n a c c e s s
j o u r n a l
Keywords: Parkinson’s disease, Graph theory, Diverse club, Rich club, Levodopa
ABSTRACT
Hierarchical brain organization, in which the rich club and diverse club situate in core
position, is critical for global information integration in the human brain network. Parkinson’s
disease (PD), a common movement disorder, has been conceptualized as a network disorder.
Levodopa is an effective treatment for PD. Whether there is a functional divergence in the
hierarchical brain system under PD pathology, and how this divergence is regulated by
immediate levodopa therapy, remains unknown. We constructed a functional network in
61 PD patients and 89 normal controls and applied graph theoretical analyses to examine the
neural mechanism of levodopa short response from the perspective of brain hierarchical
configuration. The results revealed the following: (UN) PD patients exhibited disrupted function
within rich-club organization, while the diverse club preserved function, indicating a
differentiated brain topological organization in PD. (B) Along the rich-club derivate
hierarchical system, PD patients showed impaired network properties within rich-club and
feeder subnetworks, and decreased nodal degree centrality in rich-club and feeder nodes,
along with increased nodal degree in peripheral nodes, suggesting distinct functional patterns
in different types of nodes. E (C) levodopa could normalize the abnormal network
architecture of the rich-club system. This study provides evidence for levodopa effects on the
hierarchical brain system with divergent functions.
AUTHOR SUMMARY
Many studies of brain networks have revealed densely connected regions forming the rich
club and diverse club, which occupy the central position of the hierarchical brain system.
Here, we explore the hierarchical topology in Parkinson’s disease (PD) and investigate the
neural effect of levodopa on it. We show that within the core position of the hierarchical
system, the function of the diverse club is preserved while the function of the rich club is
impaired. Along the rich-club hierarchical system, the function of biologically costly rich-club
and feeder subnetworks is disrupted, together with an increased function of peripheral nodes,
which could be normalized by levodopa. Our study provides evidence of a disparity
pattern between different levels of brain hierarchical systems under PD pathology.
Citation: Guo, T., Xuan, M., Zhou, C.,
Wu, J., Gao, T., Bai, X., … Zhang, M.
(2022). Normalization effect of levodopa
on hierarchical brain function in
Parkinson’s disease. Network
Neuroscience, 6(2), 552–569. https://doi
.org/10.1162/netn_a_00232
DOI:
https://doi.org/10.1162/netn_a_00232
Supporting Information:
https://doi.org/10.1162/netn_a_00232
Received: 5 May 2021
Accepted: 10 Gennaio 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Minming Zhang
zhangminming@zju.edu.cn
Xiaojun Guan
xiaojunguan1102@zju.edu.cn
Handling Editor:
Mikail Rubinov
Copyright: © 2022
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
N
e
N
_
UN
_
0
0
2
3
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Levodopa-induced brain normalization of PD
INTRODUCTION
Parkinson’s disease (PD) is a common neurodegenerative disorder characterized by hypodo-
paminergic neurotransmission within the nigrostriatal dopamine pathway (Braak et al., 2003;
Lees, Hardy, & Revesz, 2009), leading to classic motor deficits (Kalia & Lang, 2015). Clinically,
levodopa has become the most effective and widespread treatment for controlling PD symp-
toms (Cotzias, Van Woert, & Schiffer, 1967; Hauser, 2009). In recent years, network neurosci-
ence approaches pointed to PD being a network-disconnection syndrome (Cronin-Golomb,
2010), and the network dysfunction could be represented by the functional abnormalities cou-
pling various brain regions (Luo et al., 2015; Suo et al., 2017). Inoltre, some preliminary
exploration had demonstrated the normalization effect of dopaminergic drugs on PD func-
tional brain architecture (Ballarini et al., 2018; Berman et al., 2016). Tuttavia, brain function
is not solely attributable to the properties of individual regions but rather emerges from the
network organization of the brain as a whole (Sporns, 2011). Specifically, the human brain
is a hierarchical system in which different levels of the brain region jointly preserve overall
brain function, but without much knowledge currently available. Therefore, although the dys-
function in PD connectome and the normalization effect of dopaminergic therapy on PD over-
all network measures have been indicated, whether there is a functional divergence between
different levels of brain connectome under PD pathology, and how this divergence is regulated
by immediate levodopa therapy, remains unknown.
To elucidate these questions, the first step was to characterize the brain hierarchical system.
Network neuroscience revealed that there are a number of highly connected regions situated
in the core position of the brain network (Hagmann et al., 2008). As indexed by a high degree
centrality or high participation, these brain regions play a central role in overall network orga-
nization and have been identified as “brain hubs” (Sporns, Honey, & Kotter, 2007). Previous
studies have demonstrated that some of these brain hub regions could generate a rich-club
organization, in which these regions tend to be more densely connected among themselves
than regions with a lower degree (van den Heuvel & Sporns, 2011). The rich club is function-
ally valuable for global neural signaling and interregional brain communication, and for pro-
viding information about the network’s hierarchical ordering (van den Heuvel, Kahn, Goni, &
Sporns, 2012). Nel frattempo, a group of brain regions with a high participation coefficient show-
ing diverse connectivity are also strongly interconnected, forming the diverse club, Quale
allows for integrating information and coordinating connectivity between communities,
enabling local modular processing (Bertolero, Sì, & D’Esposito, 2017). Based on these highly
functionally connected organizations, the hierarchical brain system is depicted, which may
contribute to the investigation of functional divergence of different brain levels in PD.
Next is how to transform these theories into practice. Neuroimaging analyses provide a pow-
erful approach to map the brain network in vivo (Rubinov & Sporns, 2010). By employing resting-
state functional magnetic resonance imaging (rs-fMRI), we could construct a large-scale functional
rete, wherein brain regions serve as nodes and the interregional functional connectivity rep-
resents edges. Combining the graph theory approaches, researchers have successfully addressed
the hierarchical brain system in several disease statuses (R. Li et al., 2016; Stellmann et al., 2017;
Verhelst, Vander Linden, De Pauw, Vingerhoets, & Caeyenberghs, 2018). This hierarchical brain
model provides an avenue to investigate the functional divergence of different levels in the PD
brain connectome, and by integrating with the immediate dopaminergic therapy, we could
explore the neural effect of acute dopaminergic administration on various brain subsystems.
This study aimed to illustrate the neural mechanism of levodopa short response from the
perspective of the brain hierarchical configuration. We hypothesized that PD patients would
Brain hubs:
Some brain regions play a central role
in the overall network organization,
as indexed by a high degree or high
participation in multiple communities
across the network.
Rich club:
When the hubs of a network tend to
be more densely connected among
themselves than nodes of a lower
degree, the network present a
rich-club organization.
Diverse club:
Nodes with a high participation
coefficient are connected densely,
forming the diverse club.
Network Neuroscience
553
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
N
e
N
_
UN
_
0
0
2
3
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Levodopa-induced brain normalization of PD
exhibit a disparity pattern between different levels of the brain hierarchical system and that
immediate levodopa supplementation would exert a flexible modulation effect on them.
MATERIALS AND METHODS
Participants
All PD patients and normal controls signed informed consent forms in accordance with the
approval of the Medical Ethics Committee of the Second Affiliated Hospital of Zhejiang Uni-
versity School of Medicine.
A total of 61 PD patients and 89 normal controls were included in this study. The diagnosis
of PD was made by an experienced neurologist (B. Z.) according to the UK Parkinson’s Dis-
ease Society Brain Bank criteria (Hughes, Daniel, Kilford, & Lees, 1992). Normal controls and
PD patients with a history of other neurologic or psychiatric disorders, brain trauma, or general
exclusion criteria for MRI scanning were excluded from this study. Demographic information,
including age, sex, and education, was obtained from each participant. The neurologic assess-
menti, including disease duration, Unified Parkinson’s Disease Rating Scale (UPDRS), E
Hoehn-Yahr stage, were recorded from all PD patients in practically defined OFF-medication
condition (>12 hr after last dopaminergic medication). Additionally, motor symptoms were
reevaluated in an ON-medication condition, defined as 1 hr following antiparkinsonian treat-
ment (one tablet of immediate-release benserazide/levodopa 50/200 mg) immediately after
initial clinical assessment and MRI scanning.
MRI Data Acquisition and Processing
All participants were scanned on a 3.0 T MRI scanner (GE Health, Discovery 750) equipped
with an eight-channel head coil. During MRI scanning, the head was stabilized using resting
foam pads, and earplugs were provided to reduce the noise. Structural T1-weighted images
were acquired using a fast spoiled gradient recalled sequence: repetition time (TR) = 7.336 ms;
echo time (TE) = 3.036 ms; inversion time = 450 ms; flip angle (FA) = 11°; field of view (FOV) =
260 × 260 mm2; matrix = 256 × 256; slice thickness = 1.2 mm; 196 continuous sagittal slices.
Rs-fMRI images were acquired using a gradient recalled echo–echo planar imaging sequence:
TR= 2,000 ms; TE = 30 ms; FA = 77°; FOV = 240 × 240 mm2; matrix = 64 × 64; slice thickness =
4 mm; slice gap = 0 mm; 38 interleaved axial slices. After completing an initial rs-fMRI scan-
ning session in the OFF-medication condition, PD patients were advised to take one tablet of
benserazide/levodopa and were re-scanned one hour afterward.
The rs-fMRI data processing was performed using fMRIPrep v1.5.9 (https://fmriprep.org/en/1
.5.9/; Esteban et al., 2019) with the default processing steps. To summarize: Each T1-weighted
image was corrected for intensity nonuniformity and skull-stripped. Brain surfaces were recon-
structed using recon-all from FreeSurfer software. Spatial normalization to the ICBM 152 Non-
linear Asymmetrical template version 2009c was performed through nonlinear registration,
using brain-extracted versions of both the T1-weighted images and template. Brain tissue seg-
mentation of cerebrospinal fluid, white matter, and gray matter was performed on the brain-
extracted T1-weighted images. Functional data were corrected for slice-timing, motion, E
field distortion. This was followed by coregistration to the corresponding T1-weighted images
using boundary-based registration with 9 df. All processed rs-fMRI data were denoised by
fMRIDenoise (https://github.com/compneuro-ncu/fmridenoise) with the 24HMP8PhysSpikeReg
pipeline, including temporal band-pass filtering (0.008–0.08 Hz), detrending, and confound
regression. The confound regression employed 24 head motion parameters (three translations,
three rotations, their temporal derivatives, and their quadratic term), eight physiological noise
Network Neuroscience
554
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
N
e
N
_
UN
_
0
0
2
3
2
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Levodopa-induced brain normalization of PD
parameters (mean signals from white matter signal and cerebrospinal fluid, their temporal
derivatives, and quadratic terms), and spike regressors based on framewise displacement
(FD) and DVARS thresholds. After this, all functional data were resampled to 3-mm isotropic
and smoothed with a 5-mm full width at half maximum (FWHM) Gaussian kernel, masked by
gray matter.
Network Construction
The functional network was constructed as in a previous study (Guan et al., 2019), Dove
nodes represented brain regions and edges represented interregional functional connectivity
between every pair of nodes. The anatomical automatic labeling (AAL) atlas with 90 regions of
interest was used to generate network nodes. The mean time course of each node was
extracted, and interregional resting-state functional connectivity was calculated based on
the Pearson correlation between the time courses of each pair of nodes. To implement graph
analyses relevant to the functional network, negative correlations were omitted, and networks
were thresholded at a connection sparsity that showed the best discrimination ability of PD
patients and normal controls (see the next section).
Defining the Connection Sparsity for Network Analyses
Since the human brain network shows the characteristic of sparsity, each functional network
was thresholded by applying a set of sparsity from 0.1 A 0.5 with an interval of 0.02. We used
network density to refer sparsity, which is defined as the ratio of the number of connections
existing in the network to the maximum possible number of connections (Liao, Vasilakos, &
Lui, 2017). To implement the graph analyses in a specific sparse network, we employed
support vector machine (SVM) to identify the specific sparsity for functional networks that
was mostly discriminative and favorable for PD classification. We used the functional connec-
tivity features of normal controls and PD patients in OFF-medication status to minimize the
potential drug effects on the PD classification. To remove the redundant features, the func-
tional connectivity features selected by a two-sample t test with p < 0.05 were applied in
the SVM model using LIBSVM (https://www.csie.ntu.edu.tw/~cjlin/libsvm/) with default set-
ting. Specifically, the hyperparameter C of SVM was set to 1, and radial basis function
(RBF) kernel was used. The raw functional connectivity features were scaled individually to
range [−1, +1]. A leave-one-out cross-validation (LOOCV) scheme was used to evaluate the
performance of the SVM classifier. For each sparsity across the range of 0.1 to 0.5 with an
interval of 0.02, the same SVM procedure was performed. The sparsity with the highest
accuracy for PD classification was used to perform the network analyses. Statistical signifi-
cance of the highest classification accuracy was determined by a permutation test, which
involved repeating the classification procedure 1,000 times with a different random permuta-
tion of the training group labels (F. Li et al., 2014). Moreover, we also validated the results
within the connection sparsity range of 0.1–0.5 with an interval of 0.1. The results are shown
in the Supporting Information (Table S9 and Figures S1–S5).
Network Analyses
The final functional network was thresholded by applying the sparsity with the best discrim-
inative ability for PD classification. All network analyses were performed using Brain Connec-
tivity Toolbox (BCT, https://sites.google.com/site/bctnet/).
Rich-club organization. A brain network is thought to have a rich-club organization if nodes
with a high degree are more densely and strongly interconnected than what would be
Network Neuroscience
555
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
Rich-club nodes:
Here, defined as the top 13 brain
regions with the highest degree.
Feeder nodes:
Showing the connections with
rich-club nodes.
Peripheral nodes:
The remaining nodes except
rich-club nodes and feeder nodes
in the whole brain.
expected by chance (van den Heuvel & Sporns, 2011). The presence of a rich-club organiza-
tion was examined by calculating the weighted rich-club coefficient using BCT (Rubinov &
Sporns, 2010). The weighted rich-club coefficient φw(k) across a range of degree k of the
individual brain network was computed; a detailed description is given in the Supporting Infor-
mation. After the calculation of the weighted rich-club coefficient, the φw(k) was normalized
by comparing it with the mean weighted rich-club coefficient of 1,000 random networks.
By definition, φw
norm(k) > 1 for a range of k was indicative of a rich-club organization within
a network.
Rich-club nodes and subnetwork analyses. The rich-club node definition was based on normal
controls. To define the rich-club nodes, a group-averaged network within the normal control
group was computed as follows: Primo, from the set of individual group matrices, only connec-
tions that were present in at least 60% of the population of the group were selected for
medie, while all other connections were set to 0. Then, the group-averaged matrix was
computed by averaging only across the nonzero values of the individual subject matrices
(R. Li et al., 2016). Based on the functional group-averaged network of normal controls, IL
rich-club regions were defined as the top 15% (n = 13) of brain regions with the highest
degree (R. Li et al., 2016; Yan et al., 2018).
Identification of the rich-club regions allowed for the categorization of the whole-brain
nodes of the connectome into three types: (UN) rich-club nodes, the top 13 brain regions with
the highest degree; (B) feeder nodes, showing the connections with rich-club nodes; E (C)
peripheral nodes, the remaining nodes except rich-club nodes and feeder nodes. Based on
these three types of nodes, we categorized three types of subnetworks: (UN) rich-club subnet-
lavoro, a subgraph with rich-club nodes and the edges linking members of the rich-club nodes;
(B) feeder subnetwork, a subgraph with feeder nodes and the edges linking members of the
feeder nodes; E (C) peripheral subnetwork, a subgraph with peripheral nodes and the edges
linking members of the peripheral nodes. These three types of nodes and the derived subnet-
works based on normal controls were applied into the PD group in both OFF- and ON-
medication status.
The further subnetwork analyses focused on these three types of nodes and subnetworks.
Primo, we evaluated the nodal properties for three types of nodes, which were defined by the
sum of degree centrality of all nodes belonging to a specific node category. Then, we assessed
the intra-subnetwork properties for each type of subnetwork, including (UN) network-based sta-
tistic (NBS, https://www.nitrc.org/projects/nbs/) analysis to identify the subnetwork difference
between groups; (B) computation of the subnetwork connection strength, which was defined
by the sum of all the weights of the connections within each type of subnetwork, rispettivamente;
E (C) calculation of the global efficiency for each type of subnetwork. Finalmente, we further
analyzed the inter-subnetwork interactions, including (UN) interactions between the rich-club
subnetwork and feeder subnetwork, which were defined by the sum of all the weights of
the connections linking the rich-club nodes and feeder nodes; E (B) interactions between
the feeder subnetwork and peripheral subnetwork, which were defined by the sum of all the
weights of the connections linking the feeder nodes and peripheral nodes.
In addition to the rich-club organization, there is
Diverse-club nodes and subnetwork analyses.
another set of nodes that have edges diversely distributed across the network communities,
forming a diverse club in human brain networks (Bertolero et al., 2017). The participation
coefficient is an indicator of the diversity of each node’s connections across the network’s
communities, where nodes with a high participation coefficient exhibit diverse connectivity
Network Neuroscience
556
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
N
e
N
_
UN
_
0
0
2
3
2
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Levodopa-induced brain normalization of PD
and form the diverse club. Similar to the rich-club node definition, the diverse-club regions
were generated based on normal control groups. We first detected the community structure
based on the group-averaged network. Then, based on the community structure, we calcu-
lated the participation coefficient of each node. Allo stesso modo, we considered nodes with a high
participation coefficient (top 15%) to be the diverse-club regions. The diverse-club nodes and
the interconnections linking the members of diverse-club nodes comprised the diverse-club
subnetwork, which was applied to PD groups. We calculated the connection strength and
global efficiency of this diverse-club subnetwork.
Statistical Analyses
Statistical analyses of demographic and clinical data were performed using SPSS 19.0 statisti-
cal software. The one-sample Kolmogorov-Smirnov test was used to check the data normality.
Differences in the age, sex distribution, and education between groups were compared with
the unpaired t tests, the Mann-Whitney U tests, and Pearson chi-squared test as appropriate.
Wilcoxon signed-rank test was used to compare the difference of UPDRS motor scores
between the OFF-medication condition and ON-medication condition. Statistical significance
was set at p < 0.05.
To check whether a rich-club organization was present in the groups, a one-sample t test
was performed at each level of k to examine whether the normalized rich-club coefficient
φw
norm(k) statistically exceeded 1 in each group separately. False discovery rate (FDR) correc-
tion was applied to correct for multiple comparison across all examined levels of k. To
determine the significance levels of altered connectivity networks in NBS analysis, a general
linear model controlling for age and sex as covariates at each edge independently was
employed to test for group differences in subnetwork connectivity. A threshold (p = 0.05)
was used to form a set of suprathreshold edges (connections) among which any connected
components and their size (number of edges) could be determined. The statistical signifi-
cance of the size of each observed component was assessed with respect to an empirical
null distribution of maximal component sizes obtained under the null hypothesis of random
group membership (5,000 permutations). Significant components in each subnetwork were
determined at p < 0.05.
Group differences either in rich-club coefficient or other network properties between PD
patients and normal controls were assessed using the permutation test (with 10,000 permuta-
tions) with age and sex as covariates. A paired t test or Wilcoxon signed-rank test was used
appropriately to compare the network differences between patients in OFF-medication and
ON-medication. Tests were two-tailed with a significance level of p < 0.05, and FDR correc-
tion (with q < 0.05) was applied to correct for multiple comparisons. Finally, the relationships
between subnetwork properties or the clinical scores were examined via partial correlation
analyses, taking age and sex into account. Specifically, the relationships between subnetwork
properties and motor symptoms in OFF- and ON-medication status, as well as the relationships
between subnetwork changes and motor symptom improvement, were examined. Statistical
significance was set at p < 0.00185 (Bonferroni corrected, 3 subnetworks × 3 properties × 3
kinds of relationships, 27 correlations in total, p < 0.05/27 = 0.00185). Regarding the relation-
ships between network properties, statistical significance was set at p < 0.0125 (Bonferroni
corrected, 4 correlations in total: correlations between peripheral node degree and rich-
club/feeder node degree in OFF-medication status, and correlations between peripheral node
degree change rate and rich-club/feeder node degree improvement rate after levodopa
administration).
Network Neuroscience
557
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
In order to assess whether the results could be explained by motion, we also conducted
permutation tests with age, sex, and mean FD as covariates. Further, correlation analyses
between motion parameters and network metrics in both the normal control group and
patients in either OFF or ON status were performed. Results are shown in the Supporting Infor-
mation (Tables S4 and S5). The comparison results with mean FD as covariates between
groups were similar to the main results; in addition, there were no correlations between
motion parameters and network metrics in both the control group and PD patients whether
in OFF or ON status (FDR corrected), indicating that the network alterations were not
explained by motion.
Atlas-Based Validation on Hierarchical Topology
A functional brain network is constructed by defining the synchronization of rs-fMRI signals
between predefined brain regions, which could be potentially affected by the different brain
parcellations (atlas) employed in the computations. To minimize the potential influence of
brain atlas selection, we recruited a newly constructed brain atlas (200 parcellations) that
was derived from rs-fMRI data and that was in agreement with certain architectonic and visuo-
topic boundaries (https://github.com/ThomasYeoLab/CBIG/tree/master/stable_projects/brain
_parcellation/Schaefer2018_LocalGlobal; Schaefer et al., 2018) to validate our results. Based
on this atlas, we replicated the same procedures to define different levels of the brain hierar-
chical system and explored the neural substrate of immediate levodopa effect from the view of
brain hierarchical configuration.
RESULTS
Demographic and Clinical Characteristics
Demographic and clinical features of PD patients and normal controls are shown in Table 1.
Age, sex distribution, or education was not significantly different between PD patients and
normal controls. After levodopa administration, motor symptoms were significantly relieved
in PD patients ( p < 0.001).
Connection Sparsity for Network Analyses
The final functional networks were thresholded at a sparsity of 0.2, in which the discriminative
ability for PD classification reached the highest accuracy of 88.7% (Table 2) and was highly
Table 1. Demographic and clinical information
Normal controls
60.6 (7.0)
42/47
8.5 (3.3)
–
–
–
Parkinson’s disease patients
60.9 (8.8)
35/26
8.1 (4.6)
4.7 (3.6)
23.6 (15.0)/15.4 (12.5)
2.5 (1–5)
p value
0.805a
0.220b
0.504c
–
<0.001d
–
558
Age, mean (SD)
Sex (M/F)
Education, mean (SD)
Disease duration, mean (SD)
UPDRS-III (OFF/ON), mean (SD)
Hoehn-Yahr stage, median (range)
a Unpaired t tests.
b Pearson chi-squared test.
c Mann-Whitney U tests.
d Wilcoxon signed-rank test.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
Table 2.
Accuracy for PD classification across different sparsities
Sparsity
0.10
Accuracy (%)
88
Sparsity
0.24
Accuracy (%)
86
Sparsity
0.38
Accuracy (%)
80
0.12
0.14
0.16
0.18
0.20
0.22
88
86.7
86
85.3
88.7
86.7
0.26
0.28
0.30
0.32
0.34
0.36
85.3
85.3
85.3
84.7
83.3
82
0.40
0.42
0.44
0.46
0.48
0.50
79.3
78
78.7
76
78
78
significant at p = 0.001. Thus, all of the following network analyses were performed based on
the network sparsity of 0.2.
Rich-Club Organization
Figure 1 depicts the averaged rich-club coefficients and normalized rich-club coefficients for
both groups. In the whole-brain network, the rich-club coefficient was significantly lower in
PD patients than in normal controls in the range k = 12–15 (FDR corrected); after the levodopa
administration, the rich-club coefficient showed no difference between PD patients and nor-
mal controls. Compared with the OFF-medication condition, patients in the ON-medication
condition showed an increased rich-club coefficient in the range k = 13–15 (p < 0.05, uncor-
rected; Figure 1A). The normalized rich-club coefficient increases as a function of degree (k)
higher than 1 in both normal controls and PD patients in the OFF- and ON-medication con-
dition (Figure 1B), indicating a rich-club organization of the functional network in both groups.
Comparisons of the normalized rich-club coefficient showed that PD patients in either OFF-
medication condition or ON-medication condition exhibited a higher normalized rich-club
coefficient than normal controls in the range k = 13–25 for OFF-medication condition and
k = 10–23 for ON-medication condition, FDR corrected. No difference was observed for
the normalized rich-club coefficient in PD patients between OFF-medication and ON-
medication conditions.
Rich-club nodes. The rich-club nodes, selected on the basis of the group-averaged network,
ranking top 15% highest degree, included the following regions: left precentral gyrus, left
orbital part of inferior frontal gyrus, right Rolandic operculum, right supplementary motor area,
bilateral lingual gyrus, bilateral superior occipital gyrus, left middle occipital gyrus, right fusi-
form, bilateral superior parietal gyrus, and left supramarginal gyrus (Figure 1C, red nodes).
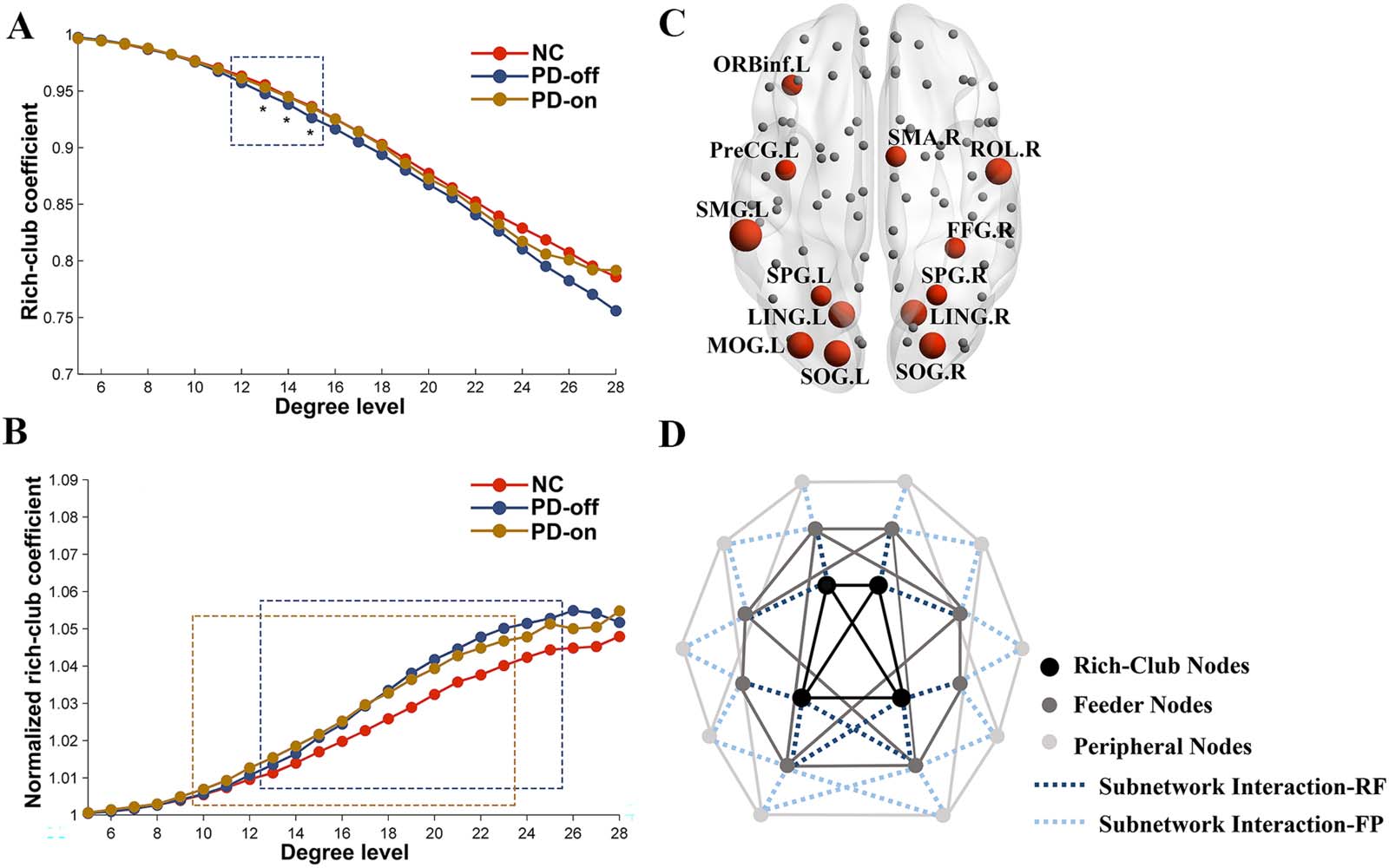
Intra-subnetwork analyses. Results of the intra-subnetwork analyses are shown in Figure 2 and
Supporting Information Table S1. First, in the comparison between normal controls and PD
patients in OFF-medication condition, NBS analysis revealed a component showing signifi-
cantly lower functional connectivity in PD patients in the rich-club subnetwork (p = 0.008)
and feeder subnetwork (p = 0.0002), respectively (Figure 2, column 1). Analyses of the
intra-subnetwork connection strength showed that in the OFF-medication condition, PD
patients exhibited decreased functional connectivity strength in the rich-club subnetwork
(p = 0.0019) and feeder subnetwork (p < 0.001) compared with normal controls. There was
Network Neuroscience
559
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Rich-club organization of functional connectome. Group-averaged rich-club curve of weighted rich-club coefficient (A) and nor-
malized weighted rich-club coefficient (B) for NC (red), PD-off (blue), and PD-on (yellow). The dashed box indicates the differences between
NC and PD patients (blue for PD-off, yellow for PD-on) after the FDR correction. * indicates the difference between PD-off and PD-on with p <
0.05. (C) Red nodes represent the functional rich-club regions. This figure is based on the functional group-averaged network in controls. The
size of the red nodes indicates the degree centrality. (D) A simplified example of the three types of nodes: rich-club nodes (black nodes), feeder
nodes (gray nodes), and peripheral nodes (light gray nodes). Three types of nodes could form three classes of subnetworks. Dashed lines
indicate the subnetwork interaction (dark blue represents the interactions between rich-club subnetwork and feeder subnetwork; light blue
shows the interactions between feeder subnetwork and peripheral subnetwork). Abbreviations: NC, normal controls; PD-off, PD patients in
OFF-medication condition; PD-on, PD patients in ON-medication condition; ORBinf.L, left orbital part of inferior frontal gyrus; PreCG.L,
left precentral gyrus; SMG.L, left supramarginal gyrus; SPG.L, left superior parietal gyrus; LING.L, left lingual gyrus; MOG.L, left middle
occipital gyrus; SOG.L, left superior occipital gyrus; SMA.R, right supplementary motor area; ROL.R, right Rolandic operculum; FFG.R, right
fusiform; SPG.R, right superior parietal gyrus; LING.R, right lingual gyrus; SOG.R, right superior occipital gyrus; subnetwork interaction-RF,
interactions between rich-club subnetwork and feeder subnetwork; subnetwork interaction-FP, interactions between feeder subnetwork and
peripheral subnetwork.
no difference in connection strength in the peripheral subnetwork between PD patients in the
OFF-medication condition and normal controls (p = 0.1205). After levodopa administration,
there was no difference in connection strength between PD patients and normal controls in
three types of subnetworks (p = 0.1326, 0.0526, and 0.1390 for rich-club subnetwork, feeder
subnetwork, and peripheral subnetwork, respectively). Compared with the OFF-medication
condition, PD patients in the ON-medication condition showed increased connection strength
in the feeder subnetwork (p = 0.009) and decreased connection strength in the peripheral sub-
network (p = 0.0383) (Figure 2, column 2, and Supporting Information Table S1). These results
indicated that levodopa administration could improve the disrupted functional connection
strength in the rich-club subnetwork and feeder subnetwork.
Regarding the global efficiency of the subnetwork, compared with normal controls, PD
patients in the OFF-medication condition showed decreased global efficiency in the rich-club
subnetwork and feeder subnetwork (both p < 0.001); PD patients in the ON-medication con-
dition showed decreased global efficiency in the feeder subnetwork (p = 0.0061). Compared
with the OFF-medication condition, PD patients in the ON-medication condition exhibited
increased global efficiency in the feeder subnetwork (p = 0.0203). No difference for global
Network Neuroscience
560
Levodopa-induced brain normalization of PD
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Comparisons of network properties for the rich-club subnetwork (A), feeder subnetwork (B), and peripheral subnetwork (C). The
blue nodes represent three types of nodes identified in this study. The edges concatenating nodes in (A) and (B) indicate a significant com-
ponent detected by network-based statistic (NBS) analysis in the rich-club subnetwork and feeder subnetwork, respectively. ** indicates the
differences corrected by FDR correction; * indicates the uncorrected differences with p < 0.05.
efficiency in the peripheral subnetwork was observed between normal controls and PD
patients in either the OFF-medication condition or the ON-medication condition (p = 0.48
and 0.0531, respectively; Figure 2, column 3, and Supporting Information Table S1). Similarly,
levodopa administration relieved the damaged efficiency in the rich-club subnetwork and
feeder subnetwork.
Nodal property analyses. For the analyses of nodal property, compared with normal controls,
we found that PD patients in the OFF-medication condition showed decreased nodal degree
centrality in rich-club nodes and feeder nodes (p = 0.0075 and 0.0029, respectively), and
increased degree centrality in peripheral nodes (p < 0.001); no difference was observed
between patients in the ON-medication condition and normal controls in three types of nodes
(p = 0.4188, 0.3745, and 0.3840 for rich-club nodes, feeder nodes, and peripheral nodes,
respectively). Compared with the OFF-medication condition, patients in the ON-medication
condition showed increased nodal degree centrality in rich-club nodes and feeder nodes (p =
0.0209 and 0.0257, respectively), which suggested a corrected effect of levodopa on rich-club
nodes and feeder nodes; intriguingly, the degree centrality in peripheral nodes was decreased
(p = 0.0130), which suggested a potentially compensatory effect of peripheral nodes on PD
pathology (Figure 2, column 4, and Supporting Information Table S1).
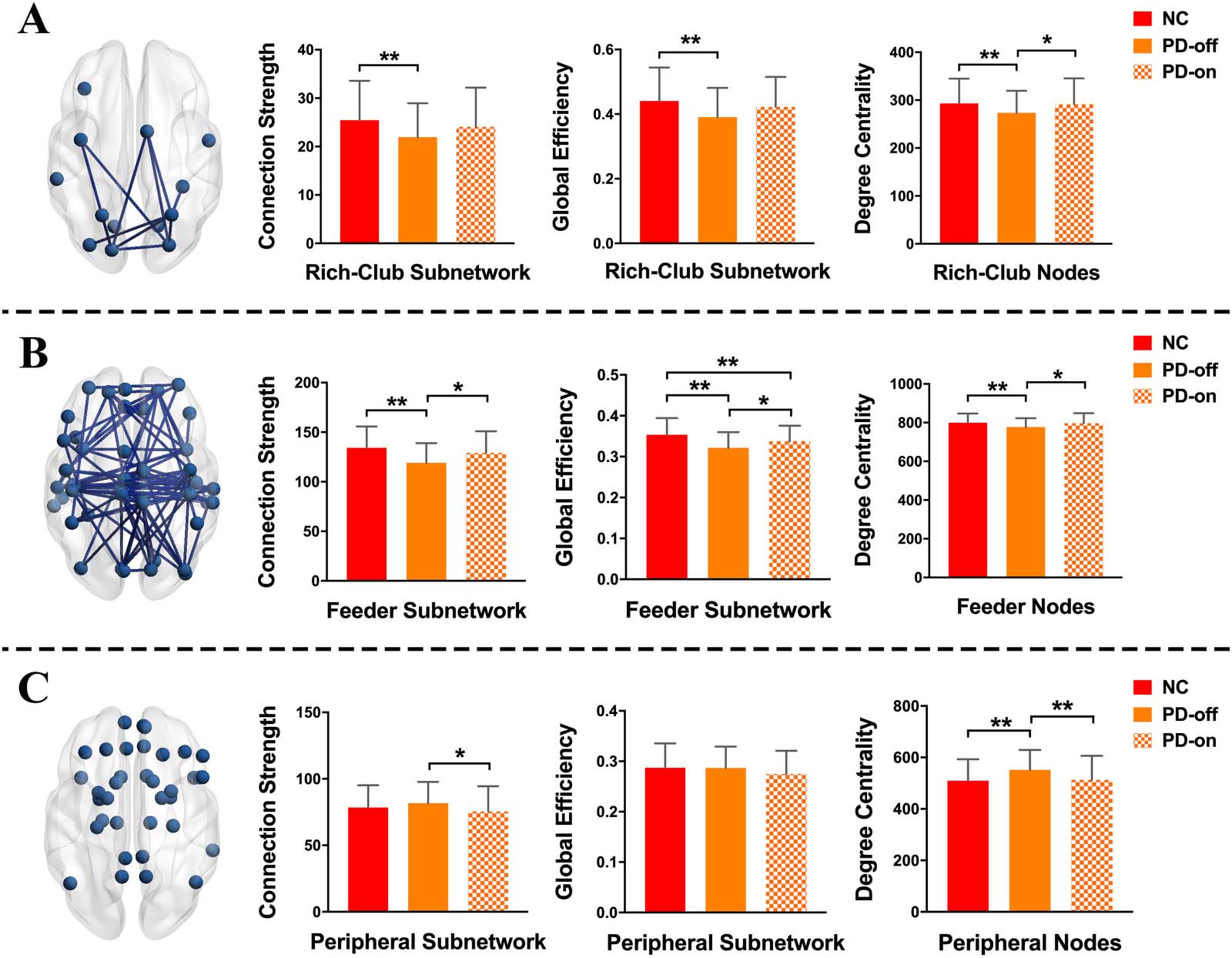
Inter-subnetwork interaction. We analyzed the interactions between subnetworks. We found
that compared with normal controls, the interactions between the rich-club subnetwork and
Network Neuroscience
561
Levodopa-induced brain normalization of PD
Figure 3. Comparisons of subnetwork interactions between the rich-club subnetwork and feeder
subnetwork (A), and between the feeder subnetwork and peripheral subnetwork (B). ** indicates the
differences corrected by FDR correction.
feeder subnetwork was decreased in PD patients in the OFF-medication condition (p < 0.001),
while PD patients in the ON-medication condition and normal controls showed no difference
in these interactions (p = 0.1687); directly compared with PD patients in the OFF-medication
condition, patients in the ON-medication condition exhibited increased interactions between
the rich-club and feeder subnetwork (p = 0.0202; Figure 3A and Supporting Information Table
S2). For the interactions between the feeder subnetwork and peripheral subnetwork, there was
no difference between normal controls and patients in either the OFF-medication condition or
ON-medication condition (p = 0.3018 and 0.0893, respectively; Figure 3B and Supporting
Information Table S2).
Relationships among network properties and clinical scores. We did not find any relationships
between subnetwork properties and motor symptom scores in PD patients in either the
OFF-medication condition or the ON-medication condition, and there was no correlation
between network changes and motor symptom improvements (Supporting Information Table
S8). Interestingly, we found that in the OFF-medication condition, the degree centrality of
peripheral nodes was negatively correlated with degree centrality of both rich-club nodes
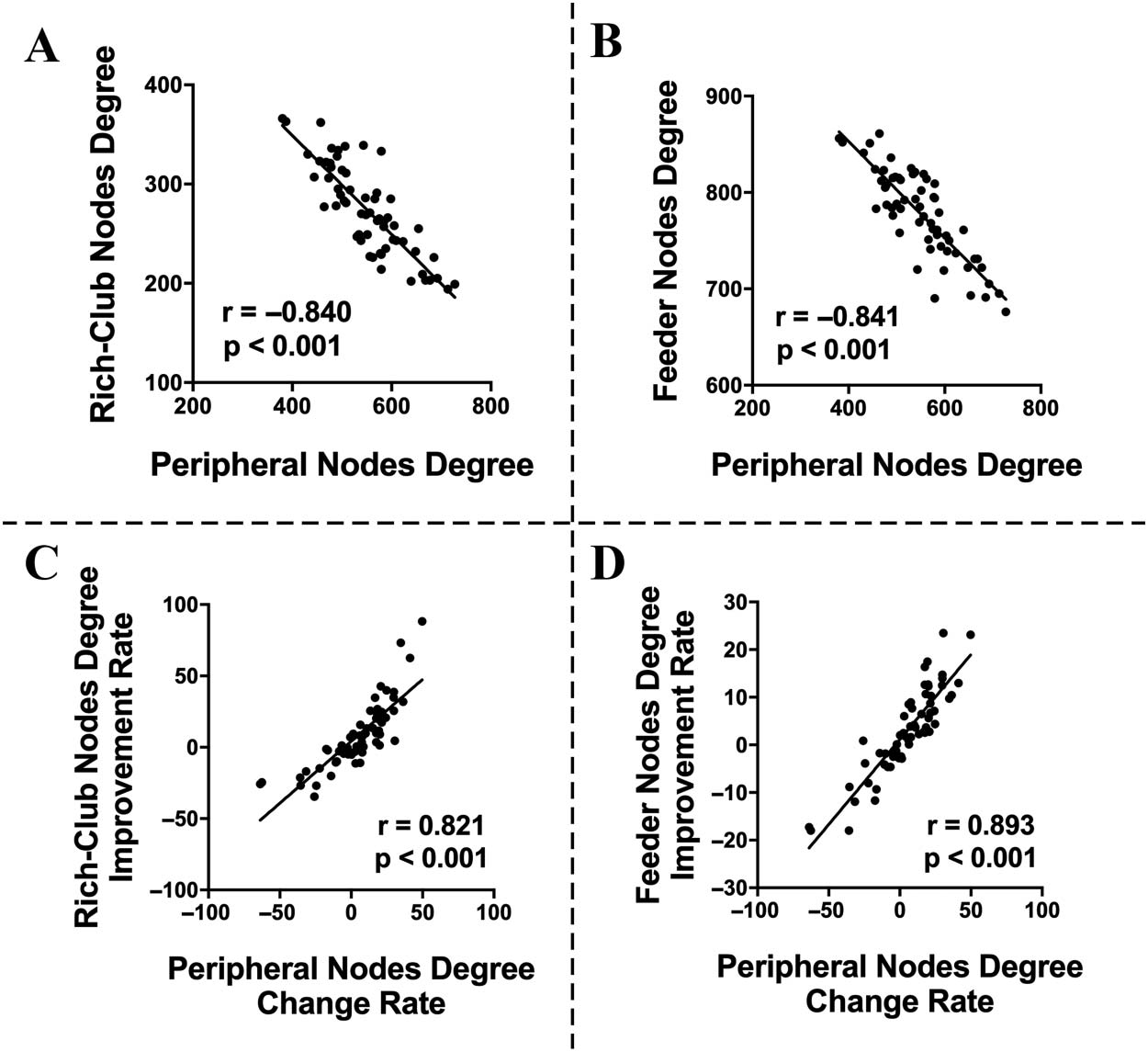
and feeder nodes (r = −0.840, p < 0.001, and r = −0.841, p < 0.001, respectively;
Figures 4A and 4B). After levodopa administration, the change rate of degree in peripheral
nodes, computed as (degreeOFF − degreeON)/degreeOFF, was positively correlated with the
improvement rate of the degree in both rich-club nodes and feeder nodes, computed as
(degreeON − degreeOFF)/degreeOFF (r = 0.821, p < 0.001, and r = 0.893, p < 0.001, respec-
tively; Figures 4C and 4D). These relationships between nodal degree centrality suggested that
the peripheral nodes may serve as a positive role to compensate the disrupted function of core
nodes (including rich-club nodes and feeder nodes).
Diverse-Club Analyses
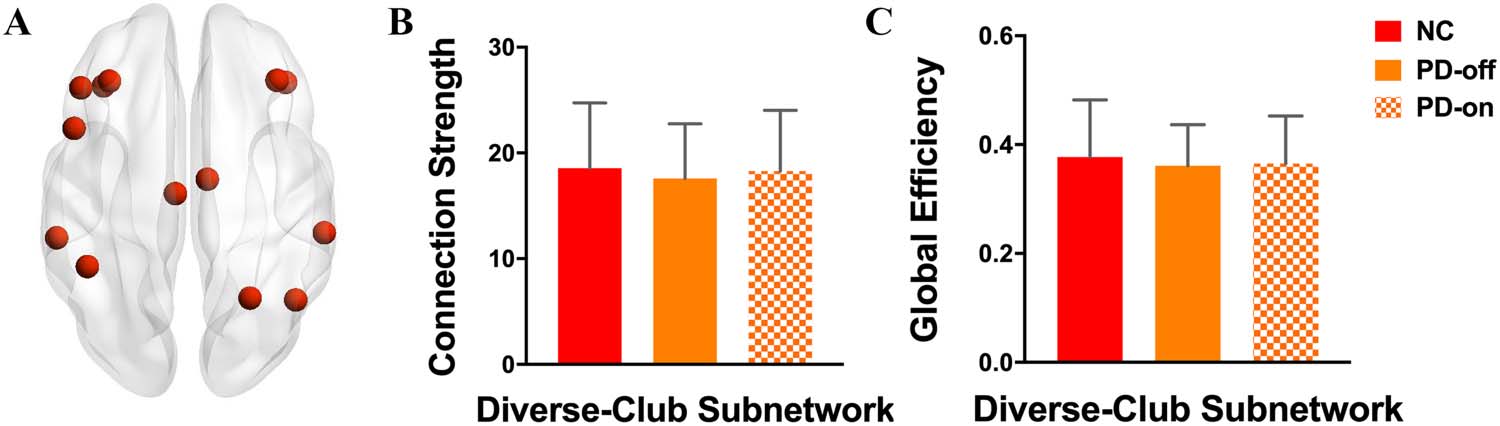
The diverse-club nodes, selected by the top 15% highest participation coefficients based on
the group-averaged network, included the following regions: bilateral middle frontal gyrus, left
opercular part of inferior frontal gyrus, left triangular part of inferior frontal gyrus, bilateral
orbital part of inferior frontal gyrus, bilateral middle cingulum, right superior parietal gyrus,
left inferior gyrus, bilateral supramarginal gyrus, and right angular (Figure 5A, red nodes).
Analyzing the connection strength and the global efficiency within the diverse-club subnet-
work, we found that there was no difference between normal controls and PD patients in
either the OFF-medication condition or the ON-medication condition (Figures 5B and 5C,
Network Neuroscience
562
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 4. Correlations between network properties. Negative correlations between peripheral
node degree and rich-club node degree (A) or feeder nodes degree (B) in OFF-medication condi-
tion. Positive correlations between the change rate of the peripheral node degree and the improve-
ment rate of rich-club node degree (C) and feeder node degree (D).
and Supporting Information Table S3), indicating a relatively reserved function of the diverse
club in PD patients.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
Robustness of Normalization Effect of Levodopa on Hierarchical Brain Organization in PD
We used the same network analysis procedures by employing another atlas to validate the
main findings. First, both normal controls and PD patients in the OFF- and ON- medication
status showed a rich-club organization in their functional network, as the normalized rich-club
coefficient higher than 1 across a range of degree (Supporting Information Figure S6). Second,
we analyzed hierarchical brain organization along the rich-club core structure as in previous
procedures. We observed impaired function within the rich-club and feeder subnetwork in PD
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Comparisons of diverse-club properties among groups. (A) Red nodes represent diverse-
club regions in the brain. (B) Connection strength and (C) global efficiency difference within the
diverse-club subnetwork were compared among groups.
Network Neuroscience
563
Levodopa-induced brain normalization of PD
patients with OFF-medication status, represented by decreased connection strength and global
efficiency, as well as decreased nodal degree within rich-club and feeder subnetworks. For the
peripheral subnetwork, PD patients in OFF-medication status showed increased connection
strength and nodal degree. After levodopa administration, these abnormal network properties
tend to reach a relatively normal state (Supporting Information Table S11). In summary, by
recruiting a new brain atlas, we found similar hierarchical brain topography and levodopa
normalizing effects as the previous findings, verifying that these results were independent of
the specific brain parcellations.
DISCUSSION
In this study, we applied a graph theory–based approach to analyze the topological organiza-
tion of the functional connectome in PD patients and the impact of dopaminergic therapy on
its functional reorganization. The main findings were as follows: (a) PD patients in the OFF-
medication condition showed impaired global network property for rich-club organization,
while the diverse club preserved function, indicating a differentiated brain topological organi-
zation in PD patients. (b) Decreased nodal degree centrality in rich-club nodes and feeder
nodes were observed in PD patients in the OFF-medication condition, while the peripheral
nodal degree showed an increase, suggesting the distinct functional patterns in different types
of nodes. And (c) levodopa could exert a normalizing effect on abnormal network architecture
of the rich-club system.
Differentiated Brain Topological Organization in PD Patients
Rich-club organization is a property common to complex networks and is considered to be a
basis for efficient global information transfer and complex neurological function in the brain
(van den Heuvel et al., 2012; van den Heuvel & Sporns, 2011). In the present study, the func-
tional brain networks of both normal controls and PD patients showed a rich-club organiza-
tion, which meant that normal controls as well as PD patients both had a subset of highly
connected nodes that were more interconnected than what would be expected by chance
(van den Heuvel & Sporns, 2011). This finding was consistent with previous research reporting
the existence of rich-club organization in PD patients (C. Li et al., 2017). Notwithstanding the
presence of a rich-club organization, PD patients showed a significant reduction in rich-club
interconnectivity compared with normal controls, which suggested that it was difficult for
patients to maintain or repair this core subnetwork composed of rich-club nodes. Previous
studies had showed that the high-degree nodes had higher blood flow, higher glucose meta-
bolic rate, and longer connection distance than other nodes (Collin, Sporns, Mandl, & van den
Heuvel, 2014; Tomasi, Wang, & Volkow, 2013). Their high topological centrality and high
biological cost could make these nodes particularly vulnerable to pathogenic factors (Crossley
et al., 2014). This phenomenon was strengthened by studies in different clinical populations,
for example, in multiple sclerosis (Stellmann et al., 2017), schizophrenia (van den Heuvel
et al., 2013), and epilepsy (R. Li et al., 2016). Our findings of disrupted interconnectivity
among rich-club regions were in line with the previous evidence and supported the proposal
that the high-degree regions are generally more susceptible to the pathology of PD. Addition-
ally, the existence of a rich-club organization has been proposed to underlie important
network properties, such as global efficiency (Xu, Zhang, & Small, 2010). The topologically
central role of the rich club may indicate that pathological attack on pivotal regions will have
an impact on the network’s global efficiency of information processing (Albert, Jeong, &
Barabasi, 2000). Therefore, the reduced global efficiency of the rich-club subnetwork suggests
that the detected abnormalities may be partly due to impaired rich-club organization.
Network Neuroscience
564
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
As opposed to the high connectivity that high-degree nodes exhibited, nodes with a high
participation coefficient exhibited diverse connectivity. These nodes are also highly intercon-
nected and form a diverse club (Bertolero et al., 2017). In the present study, in contrast to the
rich club, the connection strength and global efficiency of the diverse club showed no disrup-
tion in PD, indicating a preserved function of the diverse club. The diverse club is another
topological configuration of the human brain network showing distinct roles in network
communication that differ from the rich club. Specifically, the function of the rich club may
predominately be to maintain stability in the entire network via slow processing, potentially
using its high degree to integrate information at slower timescales, while the diverse club may
act on shorter timescales (Bertolero et al., 2017; Gollo, Zalesky, Hutchison, van den Heuvel, &
Breakspear, 2015). These different functions of the rich club and the diverse club may underlie
the observed phenomenon—impaired rich club and preserved diverse club—indicating that
the topological dysfunction in PD has an intrinsic, distinct pattern.
Distinct Functional Pattern Attributes of Different Levels of Nodes
Identification of rich-club nodes allows us to class the whole-brain nodes into different levels,
including rich-club nodes, feeder nodes, and peripheral nodes. Subsequently, the whole-brain
network could be subdivided into three subnetworks based on node classification. In the pres-
ent study, we observed reduced connection strength and global efficiency within the rich-club
and feeder subnetwork, whereas the properties within the peripheral subnetwork were
unaffected, which suggested that PD is characterized by selective disruption in central
node-related brain configuration. As mentioned before, the rich-club subnetwork and to a
lesser extent the feeder network are biologically costly, making it harder to maintain or further
develop, and therefore they are more likely to be affected in the pathological condition (Collin
et al., 2014; Crossley et al., 2014). On the other hand, the peripheral network might have a
lower biological cost and therefore is less vulnerable and less affected in the same pathological
burden (Verhelst et al., 2018). Therefore, our results underline the importance of subdividing
the brain into subnetworks and uncovering differential effects of PD pathology on the hierar-
chical brain subnetworks’ properties.
Further, analyses of the nodal property reinforced the proposal of distinct functional pat-
terns for different levels of nodes. A differential pattern of nodal degree centrality was observed
in three levels of nodes. In particular, we found that PD patients in OFF-medication status
showed a decrease of nodal degree in rich-club nodes and feeder nodes together with an
increase of nodal degree in peripheral nodes. These results suggested a likely divergence of
nodal function in hierarchical network architecture. Since the rich-club nodes and to a lesser
extent the feeder nodes are more likely to exhibit pathology (Crossley et al., 2014), the reduc-
tion of nodal degree in rich-club and feeder nodes may represent the direct pathologic influ-
ence. The increased degree centrality in peripheral nodes might reflect an attempt to restore or
compensate reduced rich-club and feeder node degree. The human brain is an integrative net-
work; brain nodes are working together to maintain the overall function of whole brain. Given
the vulnerability of topology of central nodes under the pathologic condition, other nodes with
a lower biological cost (e.g., peripheral nodes) may up-regulate their function to balance the
overall communication within the whole brain (Crossley et al., 2014; Verhelst et al., 2018). A
similar mechanism was reported in patients with traumatic brain injury (Verhelst et al., 2018);
the authors found that the increased strength was confined to the local subnetwork and may
compensate the reduced rich-club connectivity. Interestingly, the inverse associations between
reduced degree centrality in rich-club nodes as well as feeder nodes and increased degree
centrality in peripheral nodes were observed in the present study. This relationship further
Network Neuroscience
565
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
theoretically supported the notion of peripheral nodes’ compensatory role. Taken together,
analyses of nodal properties as well as subnetwork properties suggested a distinct functional
pattern for different levels of nodes: disrupted function in topological central-related nodes
accompanied by a compensatory effect in topological peripheral nodes.
Since different network sparsities may influence the network analyses, we repeated the
same network analyses over a range of network sparsities (0.1:0.1:0.5). As a result, over the
range of sparsity thresholds, normalized rich-club coefficients of normal controls and PD
patients in either OFF- or ON-medication status were more than 1, indicating the existence
of the rich-club organization in both groups. Moreover, the findings of impaired function of
rich-club and feeder subnetworks in PD OFF-medication status consistently existed over the
range of sparsity thresholds. The changes in peripheral subnetworks in PD patients with
OFF-medication status were increased overall (e.g., increased peripheral node degree) along
the range of sparsity thresholds, indicating a compensatory role of the peripheral subnetwork,
while the specific metric (e.g., global efficiency) of the peripheral subnetwork showed a
decrement when the network was denser (corresponding to a higher sparsity, e.g., sparsity =
0.4/0.5). The human brain network shows an ability to balance cost and efficiency; a denser
network often indicates a higher wiring cost, which could result in a decrement of network
efficiency (Liao et al., 2017). In our study, we showed that the core hierarchical structures
of the network were consistently impaired along a range of sparsity in PD patients, which
was consistent with our main findings and may be a result of the PD pathology regardless
of the network density. Similarly, the peripheral subnetwork with lower biological cost
exhibited an overall compensatory effect under the PD pathology, while the slight variation
of efficiency at a higher density reflected the flexibility to dynamically balance the network
cost (density) and the network efficiency, which further indicated a lower vulnerability of
the peripheral subnetwork under the same pathological burden.
Levodopa Modulates Abnormal Network Architecture
To further investigate the effect of dopaminergic medication on the abnormal brain network,
we compared the normal controls and PD patients in ON-medication status. The improvement
of decreased network properties in the rich-club subnetwork and feeder subnetwork were
observed after levodopa administration. Furthermore, the direct comparisons of network
properties between OFF- and ON-medication conditions yielded a significant increment of
network properties mainly in the feeder subnetwork. These results implied that levodopa
administration could improve the disrupted brain topology.
Previous studies investigated the effect of dopaminergic therapy on brain function (Esposito
et al., 2013; Gao, Zhang, Chan, & Wu, 2017; Zhong et al., 2019), and the results showed that
levodopa had a significant impact on restoring impaired functional connectivity of the senso-
rimotor network, default mode network, and basal ganglia motor circuit. The findings in this
study were consistent with the previous results and indicated the restorative effects of levo-
dopa on brain function. In addition, we found that levodopa could reduce the increased
degree centrality of peripheral nodes in the OFF-medication condition, making the brain’s
organization reach a relatively normal state. Consistent with our study, Berman et al. (2016)
found that levodopa can reduce the local efficiency of specific subnetworks that show signif-
icant increments in the PD OFF-medication state, suggesting a normalizing effect of levodopa
on brain topology. Thus, these findings indicated that levodopa could modulate abnormal
brain architecture; it could not only improve impaired brain function but also normalize the
abnormally increased brain topological properties. Furthermore, a positive correlation
Network Neuroscience
566
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
between the change rate of peripheral node degree and the improvement rate of rich-club (and
feeder) node degree was observed in this study; this finding reinforced the speculation of the
compensatory role of peripheral nodes as we discussed above, and also suggested that
levodopa could impart a flexible modulation effect on different parts of the brain’s organiza-
tion, making the whole brain reach a normal state.
Limitations
Several limitations of this study should be acknowledged. One major limitation was that PD
patients only took levodopa without any placebo, and normal controls were scanned only one
time. Although we ensured that all participants were in a relatively stable state, the time
effects may potentially influence the results. Both drug, placebo, and time effects should be
considered in future studies to optimize the experimental design. Second, there may be a long
duration response for dopaminergic drugs since we only withdraw antiparkinsonian drugs for
at least 12 hr; even though this withdrawal time has been widely used to reflect PD OFF-
medication status (Albanese et al., 2001; Zach et al., 2020). Third, this study was a cross-
sectional study with a moderate sample size; further prospective and longitudinal studies with
a larger sample size are warranted to validate these findings and, importantly, to explore the
longitudinal alterations of the different types of nodes along the disease progression, which is
expected to give an in-depth understanding of the topological organization in PD patients.
Finally, PD patients in this study were under multiple antiparkinsonian treatment, which
may have a potential influence on the investigation of levodopa effect; future studies including
drug-naïve PD patients could purify the effect of levodopa and contribute to exploring the
levodopa-induced brain alteration.
Conclusion
This study revealed differentiated brain organization in PD patients: The function of the
rich-club organization was disrupted, while the function of the diverse club was preserved.
Furthermore, a functional divergence existed in the PD hierarchical brain system, character-
ized by disrupted function in topological central nodes along with the compensatory effect in
topological peripheral nodes. Finally, dopaminergic therapy could modulate the brain archi-
tecture to make it reach a normal state.
ACKNOWLEDGMENTS
The authors thank all the normal volunteers and Parkinson’s disease patients recruited in this
project. The authors appreciate clinical assistance from other neurologists in the Department
of Neurology at the Second Affiliated Hospital of Zhejiang University School of Medicine.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00232.
AUTHOR CONTRIBUTIONS
Tao Guo: Conceptualization; Data curation; Formal analysis; Writing – original draft; Writing –
review & editing. Min Xuan: Conceptualization; Data curation; Formal analysis; Writing –
original draft; Writing – review & editing. Cheng Zhou: Data curation. Jingjing Wu: Data
curation. Ting Gao: Data curation. Xueqin Bai: Data curation. Xiaocao Liu: Data curation.
Luyan Gu: Data curation. Ruiqi Liu: Software. Zhe Song: Writing – review & editing.
Network Neuroscience
567
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Levodopa-induced brain normalization of PD
Quanquan Gu: Funding acquisition; Writing – review & editing. Peiyu Huang: Funding acqui-
sition; Writing – review & editing. Jiali Pu: Investigation. Baorong Zhang: Investigation. Xiaojun
Xu: Writing – review & editing. Xiaojun Guan: Data curation; Funding acquisition. Minming
Zhang: Conceptualization; Funding acquisition; Supervision; Writing – review & editing.
FUNDING INFORMATION
Minming Zhang, 13th Five-Year Plan for National Key Research and Development Program of
China, Award ID: 2016YFC1306600. Minming Zhang, National Natural Science Foundation
of China (https://dx.doi.org/10.13039/501100001809), Award ID: 81971577. Xiaojun Guan,
National Natural Science Foundation of China (https://dx.doi.org/10.13039/501100001809),
Award ID: 82001767. Xiaojun Guan, China Postdoctoral Science Foundation (https://dx.doi
.org/10.13039/501100002858), Award ID: 2021T140599 and 2019M662082. Xiaojun Xu,
National Natural Science Foundation of China (https://dx.doi.org/10.13039/501100001809),
Award ID: 82171888. Xiaojun Guan, Natural Science Foundation of Zhejiang Province
(https://dx.doi.org/10.13039/501100004731), Award ID: LQ21H180008. Min Xuan, Natural
Science Foundation of Zhejiang Province (https://dx.doi.org/10.13039/501100004731), Award
ID: LQ20H180012.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Albanese, A., Bonuccelli, U., Brefel, C., Chaudhuri, K. R., Colosimo,
C., Eichhorn, T., … Zappia, M. (2001). Consensus statement on
the role of acute dopaminergic challenge in Parkinson’s disease.
Movement Disorders, 16(2), 197–201. https://doi.org/10.1002
/mds.1069, PubMed: 11295770
Albert, R., Jeong, H., & Barabasi, A. L. (2000). Error and attack
tolerance of complex networks. Nature, 406(6794), 378–382.
https://doi.org/10.1038/35019019, PubMed: 10935628
Ballarini, T., Ruzicka, F., Bezdicek, O., Ruzicka, E., Roth, J., Villringer,
A., … Jech, R. (2018). Unraveling connectivity changes due to
dopaminergic therapy in chronically treated Parkinson’s disease
patients. Scientific Reports, 8(1), 14328. https://doi.org/10.1038
/s41598-018-31988-0, PubMed: 30254336
Berman, B. D., Smucny, J., Wylie, K. P., Shelton, E., Kronberg, E.,
Leehey, M., & Tregellas, J. R. (2016). Levodopa modulates
small-world architecture of functional brain networks in
Parkinson’s disease. Movement Disorders, 31(11), 1676–1684.
https://doi.org/10.1002/mds.26713, PubMed: 27461405
Bertolero, M. A., Yeo, B. T. T., & D’Esposito, M. (2017). The diverse
club. Nature Communications, 8(1), 1277. https://doi.org/10
.1038/s41467-017-01189-w, PubMed: 29097714
Braak, H., Del Tredici, K., Rub, U., de Vos, R. A., Jansen Steur,
E. N., & Braak, E. (2003). Staging of brain pathology related to
sporadic Parkinson’s disease. Neurobiology of Aging, 24(2),
197–211. https://doi.org/10.1016/s0197-4580(02)00065-9,
PubMed: 12498954
Collin, G., Sporns, O., Mandl, R. C., & van den Heuvel, M. P.
(2014). Structural and functional aspects relating to cost and
benefit of rich club organization in the human cerebral cortex.
Cerebral Cortex, 24(9), 2258–2267. https://doi.org/10.1093
/cercor/bht064, PubMed: 23551922
Cotzias, G. C., Van Woert, M. H., & Schiffer, L. M. (1967). Aromatic
amino acids and modification of parkinsonism. New England
Journal of Medicine, 276(7), 374–379. https://doi.org/10.1056
/NEJM196702162760703, PubMed: 5334614
Cronin-Golomb, A. (2010). Parkinson’s disease as a disconnection
syndrome. Neuropsychology Review, 20(2), 191–208. https://doi
.org/10.1007/s11065-010-9128-8, PubMed: 20383586
Crossley, N. A., Mechelli, A., Scott, J., Carletti, F., Fox, P. T.,
McGuire, P., & Bullmore, E. T. (2014). The hubs of the human
connectome are generally implicated in the anatomy of brain
disorders. Brain, 137(Pt. 8), 2382–2395. https://doi.org/10.1093
/brain/awu132, PubMed: 25057133
Esposito, F., Tessitore, A., Giordano, A., De Micco, R., Paccone, A.,
Conforti, R., … Tedeschi, G. (2013). Rhythm-specific modulation
of the sensorimotor network in drug-naive patients with
Parkinson’s disease by levodopa. Brain, 136(Pt. 3), 710–725.
https://doi.org/10.1093/brain/awt007, PubMed: 23423673
Esteban, O., Markiewicz, C. J., Blair, R. W., Moodie, C. A., Isik,
A. I., Erramuzpe, A., … Gorgolewski, K. J. (2019). fMRIPrep: A
robust preprocessing pipeline for functional MRI. Nature
Methods, 16(1), 111–116. https://doi.org/10.1038/s41592-018
-0235-4, PubMed: 30532080
Gao, L. L., Zhang, J. R., Chan, P., & Wu, T. (2017). Levodopa effect
on basal ganglia motor circuit in Parkinson’s disease. CNS
Neuroscience and Therapeutics, 23(1), 76–86. https://doi.org/10
.1111/cns.12634, PubMed: 27663605
Gollo, L. L., Zalesky, A., Hutchison, R. M., van den Heuvel, M., &
Breakspear, M. (2015). Dwelling quietly in the rich club: Brain
network determinants of slow cortical fluctuations. Philosophical
Transactions of the Royal Society B: Biological Sciences,
370(1668). https://doi.org/10.1098/rstb.2014.0165, PubMed:
25823864
Guan, X., Zhang, Y., Wei, H., Guo, T., Zeng, Q., Zhou, C., …
Zhang, M. (2019). Iron-related nigral degeneration influences
functional topology mediated by striatal dysfunction in
Network Neuroscience
568
Levodopa-induced brain normalization of PD
Parkinson’s disease. Neurobiology of Aging, 75, 83–97. https://
doi.org/10.1016/j.neurobiolaging.2018.11.013, PubMed:
30554085
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Wedeen, V. J., & Sporns, O. (2008). Mapping the structural core
of human cerebral cortex. PLoS Biology, 6(7), e159. https://doi
.org/10.1371/journal.pbio.0060159, PubMed: 18597554
Hauser, R. A. (2009). Levodopa: Past, present, and future. European
Neurology, 62(1), 1–8. https://doi.org/10.1159/000215875,
PubMed: 19407449
Hughes, A. J., Daniel, S. E., Kilford, L., & Lees, A. J. (1992). Accu-
racy of clinical diagnosis of idiopathic Parkinson’s disease: A
clinico-pathological study of 100 cases. Journal of Neurology,
Neurosurgery, and Psychiatry, 55(3), 181–184. https://doi.org/10
.1136/jnnp.55.3.181, PubMed: 1564476
Kalia, L. V., & Lang, A. E. (2015). Parkinson’s disease. Lancet,
386(9996), 896–912. https://doi.org/10.1016/S0140-6736(14)
61393-3
Lees, A. J., Hardy, J., & Revesz, T. (2009). Parkinson’s disease.
Lancet, 373(9680), 2055–2066. https://doi.org/10.1016/S0140
-6736(09)60492-X
Li, C., Huang, B., Zhang, R., Ma, Q., Yang, W., Wang, L., … Huang,
R. (2017). Impaired topological architecture of brain structural
networks in idiopathic Parkinson’s disease: A DTI study. Brain
Imaging and Behavior, 11(1), 113–128. https://doi.org/10.1007
/s11682-015-9501-6, PubMed: 26815739
Li, F., Huang, X., Tang, W., Yang, Y., Li, B., Kemp, G. J., … Gong, Q.
(2014). Multivariate pattern analysis of DTI reveals differential
white matter in individuals with obsessive-compulsive disorder.
Human Brain Mapping, 35(6), 2643–2651. https://doi.org/10
.1002/hbm.22357, PubMed: 24048702
Li, R., Liao, W., Li, Y., Yu, Y., Zhang, Z., Lu, G., & Chen, H. (2016).
Disrupted structural and functional rich club organization of the
brain connectome in patients with generalized tonic-clonic
seizure. Human Brain Mapping, 37(12), 4487–4499. https://doi
.org/10.1002/hbm.23323, PubMed: 27466063
Liao, X., Vasilakos, A. V., & He, Y. (2017). Small-world human brain
networks: Perspectives and challenges. Neuroscience and Biobe-
havioral Reviews, 77, 286–300. https://doi.org/10.1016/j
.neubiorev.2017.03.018, PubMed: 28389343
Luo, C. Y., Guo, X. Y., Song, W., Chen, Q., Cao, B., Yang, J., …
Shang, H. F. (2015). Functional connectome assessed using
graph theory in drug-naive Parkinson’s disease. Journal of
Neurology, 262(6), 1557–1567. https://doi.org/10.1007/s00415
-015-7750-3, PubMed: 25929663
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X. N.,
Holmes, A. J., … Yeo, B. T. T. (2018). Local-global parcellation of
the human cerebral cortex from intrinsic functional connectivity
MRI. Cerebral Cortex, 28(9), 3095–3114. https://doi.org/10.1093
/cercor/bhx179, PubMed: 28981612
Sporns, O. (2011). The human connectome: A complex network.
Annals of the New York Academy of Sciences, 1224, 109–125.
https://doi.org/10.1111/j.1749-6632.2010.05888.x, PubMed:
21251014
Sporns, O., Honey, C. J., & Kotter, R. (2007). Identification and classi-
fication of hubs in brain networks. PLoS ONE, 2(10), e1049. https://
doi.org/10.1371/journal.pone.0001049, PubMed: 17940613
Stellmann, J. P., Hodecker, S., Cheng, B., Wanke, N., Young, K. L.,
Hilgetag, C., … Siemonsen, S. (2017). Reduced rich-club connec-
tivity is related to disability in primary progressive MS. Neurology:
Neuroimmunology and Neuroinflammation, 4(5), e375. https://
doi.org/10.1212/NXI.0000000000000375, PubMed: 28804744
Suo, X., Lei, D., Li, N., Cheng, L., Chen, F., Wang, M., … Gong, Q.
(2017). Functional brain connectome and its relation to Hoehn and
Yahr stage in Parkinson disease. Radiology, 285(3), 904–913.
https://doi.org/10.1148/radiol.2017162929, PubMed: 28873046
Tomasi, D., Wang, G. J., & Volkow, N. D. (2013). Energetic cost of
brain functional connectivity. Proceedings of the National Acad-
emy of Sciences, 110(33), 13642–13647. https://doi.org/10.1073
/pnas.1303346110, PubMed: 23898179
van den Heuvel, M. P., Kahn, R. S., Goni, J., & Sporns, O. (2012).
High-cost, high-capacity backbone for global brain communica-
tion. Proceedings of the National Academy of Sciences, 109(28),
11372–11377. https://doi.org/10.1073/pnas.1203593109,
PubMed: 22711833
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organization
of the human connectome. Journal of Neuroscience, 31(44),
15775–15786. https://doi.org/10.1523/ JNEUROSCI.3539-11
.2011, PubMed: 22049421
van den Heuvel, M. P., Sporns, O., Collin, G., Scheewe, T., Mandl,
R. C., Cahn, W., … Kahn, R. S. (2013). Abnormal rich club orga-
nization and functional brain dynamics in schizophrenia. JAMA
P s y c h i a t r y, 7 0 ( 8 ) , 7 8 3 – 7 9 2 . h t t p s : / / d o i . o r g / 1 0 . 1 0 0 1
/jamapsychiatry.2013.1328, PubMed: 23739835
Verhelst, H., Vander Linden, C., De Pauw, T., Vingerhoets, G., &
Caeyenberghs, K. (2018). Impaired rich club and increased local
connectivity in children with traumatic brain injury: Local sup-
port for the rich? Human Brain Mapping, 39(7), 2800–2811.
https://doi.org/10.1002/hbm.24041, PubMed: 29528158
Xu, X. K., Zhang, J., & Small, M. (2010). Rich-club connectivity
dominates assortativity and transitivity of complex networks.
Physical Review E, 82(4 Pt. 2), 046117. https://doi.org/10.1103
/PhysRevE.82.046117, PubMed: 21230355
Yan, T., Wang, W., Yang, L., Chen, K., Chen, R., & Han, Y. (2018).
Rich club disturbances of the human connectome from subjective
cognitive decline to Alzheimer’s disease. Theranostics, 8(12),
3237–3255. https://doi.org/10.7150/thno.23772, PubMed:
29930726
Zach, H., Dirkx, M. F., Roth, D., Pasman, J. W., Bloem, B. R., &
Helmich, R. C. (2020). Dopamine-responsive and dopamine-
resistant resting tremor in Parkinson disease. Neurology, 95(11),
e1461–e1470. https://doi.org/10.1212/ WNL.0000000000010316,
PubMed: 32651292
Zhong, J., Guan, X., Zhong, X., Cao, F., Gu, Q., Guo, T., … Zhang,
M.
(2019). Levodopa imparts a normalizing effect on
default-mode network connectivity in non-demented Parkinson’s
disease. Neuroscience Letters, 705, 159–166. https://doi.org/10
.1016/j.neulet.2019.04.042, PubMed: 31026534
Network Neuroscience
569
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
5
5
2
2
0
2
8
1
3
9
n
e
n
_
a
_
0
0
2
3
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3