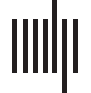RESEARCH
Structural covariance networks across the
life span, from 6 A 94 years of age
1
Elizabeth DuPre
1
and R. Nathan Spreng
1Laboratory of Brain and Cognition, Human Neuroscience Institute, Department of Human Development,
Cornell University, Ithaca, NY, USA
Keywords: Network, Structural covariance, Gray matter, Development, Aging, MRI
ABSTRACT
Structural covariance examines covariation of gray matter morphology between brain
regions and across individuals. Despite significant interest in the influence of age on
structural covariance patterns, no study to date has provided a complete life span
perspective—bridging childhood with early, middle, and late adulthood—on the
development of structural covariance networks. Here, we investigate the life span trajectories
of structural covariance in six canonical neurocognitive networks: default, dorsal attention,
frontoparietal control, somatomotor, ventral attention, and visual. By combining data from
five open-access data sources, we examine the structural covariance trajectories of these
networks from 6 A 94 years of age in a sample of 1,580 participants. Using partial least
piazze, we show that structural covariance patterns across the life span exhibit two
significant, age-dependent trends. The first trend is a stable pattern whose integrity declines
over the life span. The second trend is an inverted-U that differentiates young adulthood
from other age groups. Hub regions, including posterior cingulate cortex and anterior insula,
appear particularly influential in the expression of this second age-dependent trend. Overall,
our results suggest that structural covariance provides a reliable definition of neurocognitive
networks across the life span and reveal both shared and network-specific trajectories.
AUTHOR SUMMARY
The importance of life span perspectives is increasingly apparent in understanding normative
interactions of large-scale neurocognitive networks. Although recent work has made
significant strides in understanding the functional and structural connectivity of these
networks, there has been comparatively little attention to life span trajectories of structural
covariance networks. In this study we examine patterns of structural covariance across the
life span for six neurocognitive networks. Our results suggest that networks exhibit both
network-specific stable patterns of structural covariance as well as shared age-dependent
trends. Previously identified hub regions seem to show a strong influence on the expression
of these age-related trajectories. These results provide initial evidence for a multimodal
understanding of structural covariance in network structure-function interaction across the
life course.
INTRODUCTION
The human cerebral cortex is hierarchically organized into complex brain networks that can
be considered at multiple levels of analysis (Mesulam, 1998). One such level is structural
covariance, or how interindividual differences in regional brain structure covary with other
a n o p e n a c c e s s
j o u r n a l
Citation: DuPre, E., & Spreng, R. N.
(2017). Structural covariance networks
across the life span, from 6 A 94 years
of age. Network Neuroscience, 1(3),
302–323. https://doi.org/10.1162/
netn_a_00016
DOI:
https://doi.org/10.1162/netn_a_00016
Supporting Information:
https://doi.org/10.1162/netn_a_00016
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
R. Nathan Spreng
nathan.spreng@gmail.com
Handling Editor:
Olaf Sporns
Copyright: © 2017
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
N
e
N
_
UN
_
0
0
0
1
6
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Lifespan structural covariance networks
Structural covariance:
Interindividual differences in regional
brain structure covarying with other
brain structures across the
population.
Neurocognitive networks:
Large-scale, distributed neural
networks associated with cognition.
brain structures across the population (Alexander-Bloch, Giedd, & Bullmore, 2013; Mechelli,
Friston, Frackowiak, & Price, 2005). Structural covariance networks reflect shared variation in
gray matter morphology (Mechelli et al., 2005) and are assessed using measures such as corti-
cal thickness and regional volume. These networks exhibit reproducible organization at both a
population (Alexander-Bloch et al., 2013) and an individual (Tijms, Seris, Willshaw, & Lawrie,
2012) level and have been identified across species (Pagani, Bifone, & Gozzi, 2016), under-
scoring their role as an intrinsic feature of cortical organization. Despite this reliability, IL
source of gray matter shared covariance patterns is unclear and has been hypothesized to
reflect both genetic and plastic influences, including maturational timing (Alexander-Bloch,
Raznahan, Bullmore, & Geidd, 2013).
Age is a significant moderator of both anatomical
(Collin & van den Heuverl, 2013;
Hagmann et al., 2010) as well as functional
(Chan, Park, Savalia, Petersen, & Wig, 2014;
Dosenbach et al., 2010) connectivity. Some of the most extensive age effects occur in gray
matter (Giorgio et al., 2010). Gray matter organization undergoes significant structural change
with age, including synaptic proliferation, pruning, and eventual atrophy (Fjell et al., 2010;
Basso & Cheng, 2006). Normative gray matter changes do not occur simultaneously, Tuttavia,
and show variation across cortex (Krongold, Cooper, & Bray, 2017; Raz et al., 2005), yielding
significant differences in age-related trajectories across structural covariance networks. There
has therefore been substantial interest in the impacts of age on structural covariance networks,
and how these age-related trajectories may differ across neurocognitive networks.
Investigations of structural covariance trajectories have largely focused on specific devel-
opmental periods, including childhood and adolescence (Zielinski, Gennatas, Zhou, & Seeley,
2010) or aging (Montembeault et al., 2012). These studies have suggested the emergence of
increasing long-range structural covariance across early development (Zielinski et al., 2010)
and increased local covariance with advancing age (Montembeault et al., 2012). Importantly,
examining structural covariance networks in isolated developmental periods may limit our
understanding of the normative life cycle of each of these networks (Zuo et al., 2017). Initial
work examining trajectories over multiple developmental periods has found significant inter-
network variation (Hafkemeijer et al., 2014).
There has also been increasing interest in examining structural covariance networks from
a life span perspective. Tuttavia, to date existing life span structural covariance studies (cioè.,
those spanning a minimum of 35 years of development; Zuo et al., 2017) have only included
subjects with a minimum age of 18 years and considered differences between young-, middle-,
or older-adult groups (Li et al., 2013; Wu et al., 2012). Results from these studies have largely
been in agreement with those of individual developmental periods, with distributed structural
covariance shifting to more local topology in older adulthood, though the timing of this tran-
sition is unclear and has differed between middle- (Wu et al., 2012) and younger- (Li et al.,
2013) adulthood. Studies have also shown differences in structural covariance trajectories by
rete, with primary sensory and motor networks showing few to no age-related changes
across adulthood, while neurocognitive and semantic networks show a general shift from
distributed to local covariance (Li et al., 2013).
Despite this significant progress in understanding structural covariance during develop-
ment and aging, the authors are unaware of any studies that have examined the development
of large-scale structural covariance networks across the entire life span, including childhood
and adolescence to old age. The changes seen in the developmental trajectory of large-scale
functional networks (Zuo et al., 2017) suggest that a life span study of structural covariance
Network Neuroscience
303
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
N
e
N
_
UN
_
0
0
0
1
6
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Lifespan structural covariance networks
networks may provide an important complement, yielding insights into cortical organization
at the level of gray matter morphology.
Infatti, previous work by Zielinski and colleagues
(2010) supports the reflection of a network’s functional specialization in its age-specific struc-
tural covariance pattern. Based on previous findings, we hypothesized that the distributed
neurocognitive default, dorsal attention, frontoparietal control, and ventral attention networks
would show an inverted U-shaped trajectory of increasingly distributed structural covariance in
early development, before shifting to more local covariance in advanced aging. Following re-
sults reported by Li and colleagues (2013) of age-independent patterns of structural covariance
in somatomotor and visual networks across the adult life span, we predicted no age-dependent
patterns of structural covariance in these networks. To examine these hypothesized life span
trajectories, whole-brain structural covariance was assessed in a seed-based multivariate anal-
ysis (Persson et al., 2014; Spreng & Turner, 2013). This seed-based multivariate investigation
allowed for the data-driven identification of significant age-related trajectories, based on the
structural covariance of cortical gray matter with the chosen seed regions. We examine tra-
jectories of structural covariance networks across the life span to consider what these changes
might reveal about their developmental organization.
MATERIALS AND METHODS
In this study, our primary aim was to provide comprehensive mapping of the neurocognitive
large-scale structural covariance networks across the entire life span. We collapsed cross-
sectional data across five publicly available datasets to provide a normative sample ranging
from 6 A 94 years of age. This also afforded us sufficient power for reliable estimates of
structural covariance networks at six developmental epochs: Age Group 1 (6–15 years), Age
Group 2 (16–25 years), Age Group 3 (26–35 years), Age Group 4 (36–59 years), Age Group 5
(60–75 years), and Age Group 6 (76–94 years). We assessed the structural covariance of six
the default network
large-scale neurocognitive networks well represented in the literature:
(DN), dorsal attention network (DAN), frontoparietal control network (FPCN), somatomotor
rete (SM), ventral attention network (VAN), and visual systems.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
N
e
N
_
UN
_
0
0
0
1
6
P
D
.
T
Image Acquisition
Data were collated from five open-access data sources: National Institutes of Health Pedi-
atric MRI Data Repository (NIH-Peds; Brain Development Cooperative Group & Evans, 2007):
Release 5; Human Connectome Project (HCP): 500 subjects release; Nathan Kline Institute-
Rockland Sample (NKI-RS; Nooner et al., 2012): Release 5; Open Access Series of Imaging
Studi (OASIS); and Alzheimer’s Disease Neuroimaging Initiative (ADNI). A complete listing
of T1-weighted anatomical image acquisition procedures for each data source is provided in
Tavolo 1.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 1.
Image acquisition parameters by data source
Scanner
Strength (T)
1.5
3
3
1.5
1.5
NIH-Peds
HCP
NKI-RS
OASIS
ADNI
TR (ms)
22–25
2,400
1,900
9.7
2,400
TE (ms)
10–11
2.14
2.52
4.0
min. full
TI (ms)
TD (ms)
Flip
Angle (deg.)
1,000
900
20
1,000
0
0
200
0
8
9
10
8
FOV (mm)
256 × 160–180
224 × 224
250 × 250
256 × 256
240 × 240
Voxel Size (mm)
1.0 × 1.0 × 1.0–1.5
0.7 × 0.7 × 0.7
1.0 × 1.0 × 1.0
1.0 × 1.0 × 1.25
0.94 × 0.94 × 1.2
Network Neuroscience
304
Lifespan structural covariance networks
Participant Characteristics
From each sample, only healthy control participants older than six years of age with no di-
agnosed history of DSM (Diagnostic and Statistical Manual of Mental Disorders) Axis I or
II disorder were considered. Six years was chosen as the lowest estimate for life span char-
acterization, since previous work has indicated that normalization for children less than six
years old is likely to introduce significant artifacts (Muzik, Chugani, Juhász, Shen, & Chugani,
2000), as gray matter volume in younger children is less than 95% of that observed in adults
(Caviness, Kennedy, Richelme, Rademacher, & Filipek, 1996). For individuals meeting these
criteria, the T1-weighted anatomical image was selected.
In the case of longitudinal data,
only the first time point was selected for each participant.
All T1-weighted anatomical images ( n = 1, 667) were visually inspected for quality as-
surance: images that showed evidence of artifacts were excluded ( n = 87), yielding a final
sample size of n = 1, 580 (age M = 35 years, SD = 23 years, Range = 6–94 years; 659 males;
859 scanned at 1.5T and 721 at 3T). Participants were then sorted into the following age
groups: Age Group 1 (6–15 years), Age Group 2 (16–25 years), Age Group 3 (26–35 years),
Age Group 4 (36–59 years), Age Group 5 (60–75 years), and Age Group 6 (76–94 years). Vedere
Tavolo 2 for sample sizes and participant characteristics by age group.
Segmentation and Preprocessing
Each age group was separately submitted to voxel-based morphometry (Ashburner & Friston,
implemented in Matlab
2000) using the VBM8 toolbox (Kurth, Luders, & Gaser, 2010)
(MATLAB 8.0, MathWorks, Natick, MA, 2012).
Images were first segmented into gray mat-
ter, white matter, and cerebrospinal fluid using an extension of the New Segmentation algo-
rithm. Gray matter images for this age group were then affine registered to the MNI template
and carried to the Diffeomorphic Anatomical Registration through Exponentiated Lie Algebra
toolbox (DARTEL; Ashburner, 2007), where they were iteratively aligned to create an age-
group-specific template in MNI space. The six resulting age-group-specific templates were
themselves then iteratively aligned again using DARTEL to create a study-specific template in
MNI space. Importantly, this study-specific template equally weighted each of the age ranges
represented by the six age groups.
Finalmente, previously segmented images were aligned to the study-specific template of interest
using DARTEL high-dimensional normalization within VBM8. Nonlinear-only modulation was
applied to gray matter images to derive regional differences in gray matter volume, correcting
for total intracranial volume. Modulated gray matter images were iteratively smoothed to 8 mm
FWHM (full width at half maximum) using 3dBlurToFWHM in AFNI (Cox, 1996) and carried
forward for further analysis.
Age Group
6–15 years
16–25 years
26–35 years
36–59 years
60–75 years
76–94 years
Tavolo 2. Participant characteristics by age group
Sample Size
(Males)
330 (159)
302 (139)
472 (192)
139 (38)
203 (82)
134 (49)
Age in Years
M (SD)
10 (2.66)
21 (2.8)
30 (2.74)
49 (6.29)
70 (4.16)
81 (4.28)
Scanned at
1.5T/3T
306/24
176/126
31/441
68/71
157/46
121/13
Network Neuroscience
305
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
N
e
N
_
UN
_
0
0
0
1
6
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Lifespan structural covariance networks
Network Identification
In this study, we sought to examine the structural covariance of the large-scale neurocognitive
networks, including the DN, DAN, FPCN, SM, VAN, and visual networks. To examine each
of these six networks, gray matter volumes for selected high-confidence seeds reported in
Yeo et al. (2011) were extracted. Although Yeo and colleagues (2011) report high-confidence
seeds for seven networks, we chose to exclude the reported “limbic network” as recent work
has raised concerns regarding its test-retest reliability (Holmes et al., 2015).
For each of the six remaining networks we selected the top two high-confidence seeds
reported by Yeo et al. (2011) as well as the contralateral seed regions, where contralateral
seeds were chosen by changing the sign of the x-coordinate on each of the original high-
confidence seeds. An exception to this procedure was made for the DN, which is known to
separate into anterior and posterior components (Uddin, Kelly, Biswal, Castellanos, & Milham,
2009). Therefore, for the DN we selected the highest confidence seed and its contralateral
seed. We then selected the second-highest confidence seed in posterior cingulate cortex as
well as the fourth-highest confidence seed in medial prefrontal cortex, in order to ensure that
both the anterior and the posterior DN components were represented in structural covariance
estimates. A listing of all 24 seeds for the six networks examined is presented in Table 3 along
with their respective anatomical label, and a visual representation of their location on cortex
is presented in Figure 1.
Network Affiliation
Default
Dorsal attention
Frontoparietal control
Ventral attention
Somatomotor
Visual
Tavolo 3. Selected seeds for each network
X
−7
−7
−41
41
−22
22
−51
51
−40
40
−43
43
−5
5
−31
31
−41
41
−55
55
−3
3
−16
16
sì
49
−52
−60
−60
−8
−8
−64
−64
50
50
−50
−50
15
15
11
11
−20
−20
−4
−4
−74
−74
−74
−74
z
18
26
29
29
54
54
−2
−2
7
7
46
46
32
32
8
8
62
62
26
26
23
23
7
7
Laterality
l
l
l
R
l
R
l
R
l
R
l
R
l
R
l
R
l
R
l
R
l
R
l
R
Anatomical Label
Medial prefrontal cortex
Posterior cingulate cortex
Inferior parietal lobule
Inferior parietal lobule
Frontal eye fields
Frontal eye fields
Middle temporal motion complex
Middle temporal motion complex
Frontal pole
Frontal pole
Anterior inferior parietal lobule
Anterior inferior parietal lobule
Anterior cingulate cortex
Anterior cingulate cortex
Anterior insula
Anterior insula
Precentral gyrus (hand)
Precentral gyrus (hand)
Precentral gyrus (tongue)
Precentral gyrus (tongue)
Extrastriate visual cortex
Extrastriate visual cortex
Visual area 1
Visual area 1
Coordinates (X, sì, z) are in MNI stereotaxic space.
Network Neuroscience
306
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
N
e
N
_
UN
_
0
0
0
1
6
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Lifespan structural covariance networks
Figura 1. Selected seeded regions. The four selected seeded regions for each of the six neurocog-
nitive networks are depicted in colors corresponding to their Yeo et al. (2011) labeling.
For each seed, gray matter volumes were extracted from a 10.5 mm edge cubical region
of interest (ROI). Extracted gray matter volumes were then averaged across the four seeds for
each participant. We chose to average gray matter volumes from multiple seeds to provide
reliable, long-range estimates of network-specific structural covariance. This is in contrast to
the more local estimates of structural covariance provided by gray matter volume from a single
seed region. All six neurocognitive networks were examined by averaging the extracted gray
matter volumes for each of the network-specific seeds, resulting in a 1,580 × 1 vector for each
rete. For each of the analyses, this vector Y represented the average gray matter volume
for each participant of key nodes within the network. The resulting Y vectors were submitted
to partial least squares (PLS; McIntosh, Bookstein, Haxby, & Grady, 1996). Additionally sub-
mitted to PLS were matrices of participant structural images, X, where X is an N subjects × N
voxels matrix representing voxel-wise estimates of gray matter volume for each participant.
Partial least squares:
Multivariate decomposition
technique used to identify shared
patterns of covariance.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
N
e
N
_
UN
_
0
0
0
1
6
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Partial Least Squares Analyses
PLS is a data-driven multivariate statistical technique capable of identifying patterns of struc-
tural covariance (Persson et al., 2014; Spreng & Turner, 2013). We utilized seed PLS to identify
patterns of covariance between gray matter integrity in seed regions and whole-brain structural
MRI images (for a review, see Krishnan, Williams, McIntosh, & Abdi, 2011). Here, we adopt
the nomenclature used in Miši´c et al. (2016).
Derivation of covariance matrix. For experimental analyses, our seed value was the aver-
age gray matter volume of four selected high-confidence seeds reported in Yeo et al. (2011).
The vector Y representing this average gray matter volume was cross-correlated with a matrix
X of participant’s structural images. Importantly, this participant image matrix contained six
submatrices X1..6 corresponding to each age group. We retained this age group organization
in our PLS analyses in order to directly compare age groups in their structural covariance be-
tween average network and whole-brain gray matter volume. The vector Y can therefore be
considered as containing six subvectors, corresponding to the participant age groups. Both
the gray matter volume vector and the image matrix were centered and normalized within
age groups such that their cross-correlation resulted in a covariance vector Z according to the
following:
Y 1..6
T X 1..6 = Z1..6.
(1)
307
Network Neuroscience
Lifespan structural covariance networks
Latent variables:
Linear combinations of the input
variables that optimally explain
orthogonal patterns of covariance.
Brain scores:
In partial least squares nomenclature,
values measuring the extent to
which an individual aligns with
or contributes to the group spatial
pattern. The value is taken as the dot
product of the group result and the
individual subject gray matter
images.
Note that this covariance vector is equivalent to a correlation vector because of the de-
scribed within-group normalization. The resulting covariance vector Z measures the degree to
which the network average and whole-brain gray matter volumes covary at a voxel-wise level
across participants.
Singular value decomposition. Using singular value decomposition (SVD; Eckart & Young,
1936), the covariance vector Z from Equation 1 was then decomposed into the following:
Z1..6 = UΔV T,
(2)
where V is the orthonormal matrix of right singular vectors, U is the orthonormal matrix of left
singular vectors, and Δ is the diagonal matrix of singular values. The right and left singular
vectors represent the gray matter seed integrity profiles and spatial patterns that best charac-
terize the covariance vector Z. The triplet of the right and left singular vectors and the singular
values forms a set of mutually orthogonal latent variables (LVs), where the number of LVs
derived is equal to the rank of the covariance vector Z. In our analyses, this identified six LVs
for each network corresponding to the six submatrices of Z. Each LV was tested for statistical
significance with 5,000 permutations and cross-validated for reliability with 1,000 bootstraps.
Bootstrap ratios, derived from dividing the weight of the singular-vector by the bootstrapped
standard error, are equivalent to z scores and were used to threshold significant LV spatial
patterns at a 95% confidence interval for projection and interpretation.
Patterns were considered for further analysis based on two criteria. Primo, LVs must be statis-
tically significant by permutation testing at the level of p < 0.001. Second, LVs must account
for a minimum of 5% of the covariance in the data.
Derivation of subject scores. We also quantified individual contributions to each LV by de-
riving subject scores. Of particular interest in this work are the subject scores known in PLS
nomenclature as “brain scores,” which assess the contribution of each individual to the group
structural covariance pattern. Multiplying the original matrix X1..6 of participant structural
images by the matrix of right singular vectors V derives these brain scores as follows:
L = X 1..6V ,
(3)
where L is a matrix of brain scores. Recall from Equation 2 that the right singular vector V
represents the seed-integrity profiles that best characterize the covariance matrix Z, such that
multiplying this singular vector by the participant structural images derives the seed integrity
profiles for each participant that reflect their contribution to the group structural covariance
pattern. The matrix of brain scores L was extracted for each LV where, for each participant,
this brain score value represents a weighted value of gray matter integrity within the regions
identified in the group image.
By correlating brain score values for all subjects within each of the six age groups with
their input gray matter integrity values, we were able to assess gray matter integrity in these
regions for each age group separately. Computed confidence intervals on these correlations
provide a means to assess the reliability of the structural covariance patterns in each age group;
confidence intervals that cross zero are considered unreliable and are not interpreted in the
results. To account for potential confounds, we ran a multiple regression of these brain scores
controlling for scanner strength and gender. Although we present results corrected for age and
gender, controlling for these variables did not qualitatively affect the results (see Supplementary
Figure 1 for an exemplar network; DuPre & Spreng, 2017). Corrected brain scores were plotted
Network Neuroscience
308
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
against age to visualize the covariance of the associated spatial pattern across the population.
Because of the heterogeneity of resulting age-dependent trajectories, summary statistics for
models fit to these corrected brain scores are available in Supplementary Tables 1 and 2 (DuPre
& Spreng, 2017). For those models who show a “peak” in their age-dependent trajectory, the
age at which this functional maximum occurs is noted in Supplementary Table 2.
RESULTS
We investigated the structural covariance of previously identified large-scale neurocognitive
networks including the DN, DAN, FPCN, SM, VAN, and visual networks. Using PLS, we
identified patterns of structural covariance for each of the six networks examined.
Neurocognitive Network Structural Covariance Patterns
PLS analyses of each of the large-scale networks examined yielded multiple significant latent
variables (LVs), corresponding to reliable patterns of structural covariance within each network.
We review significant results for each of the networks in turn.
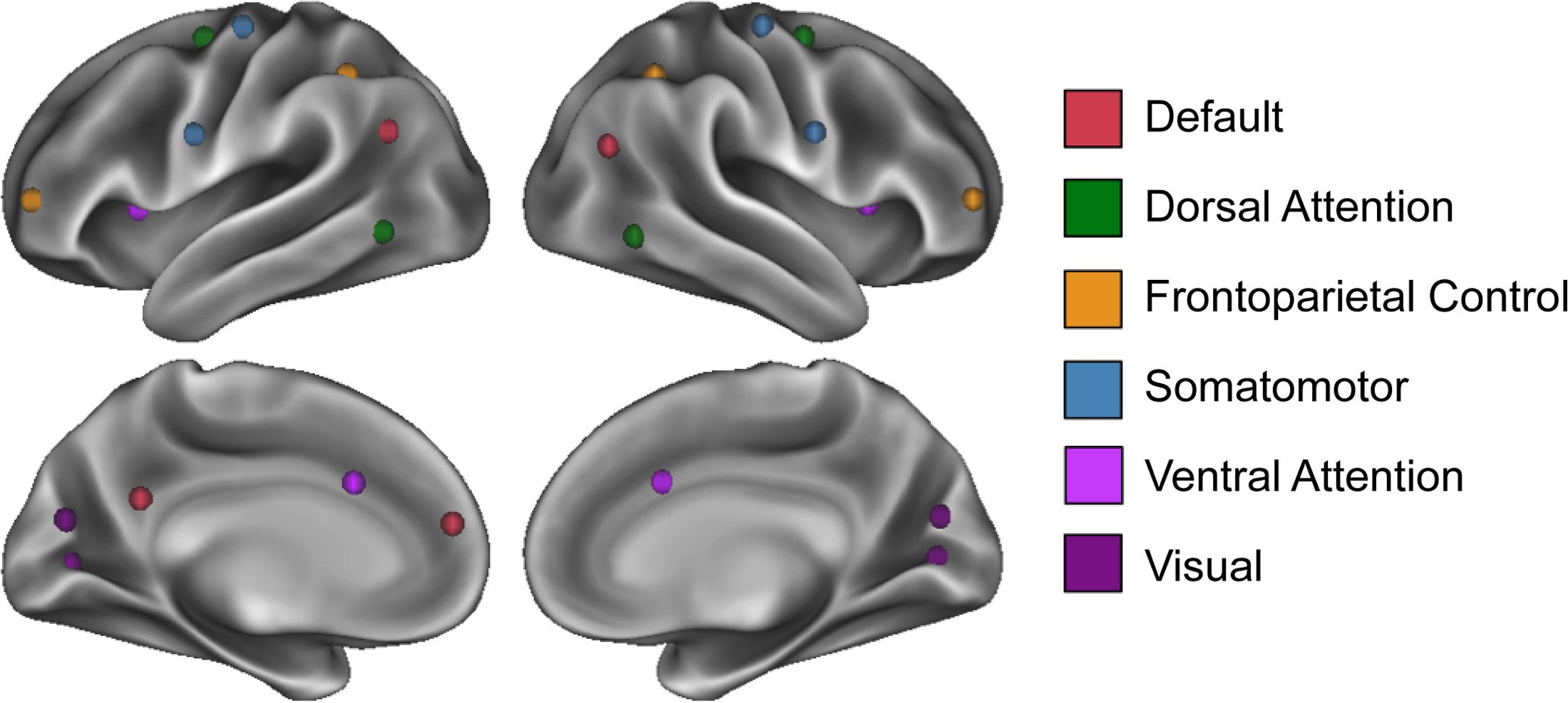
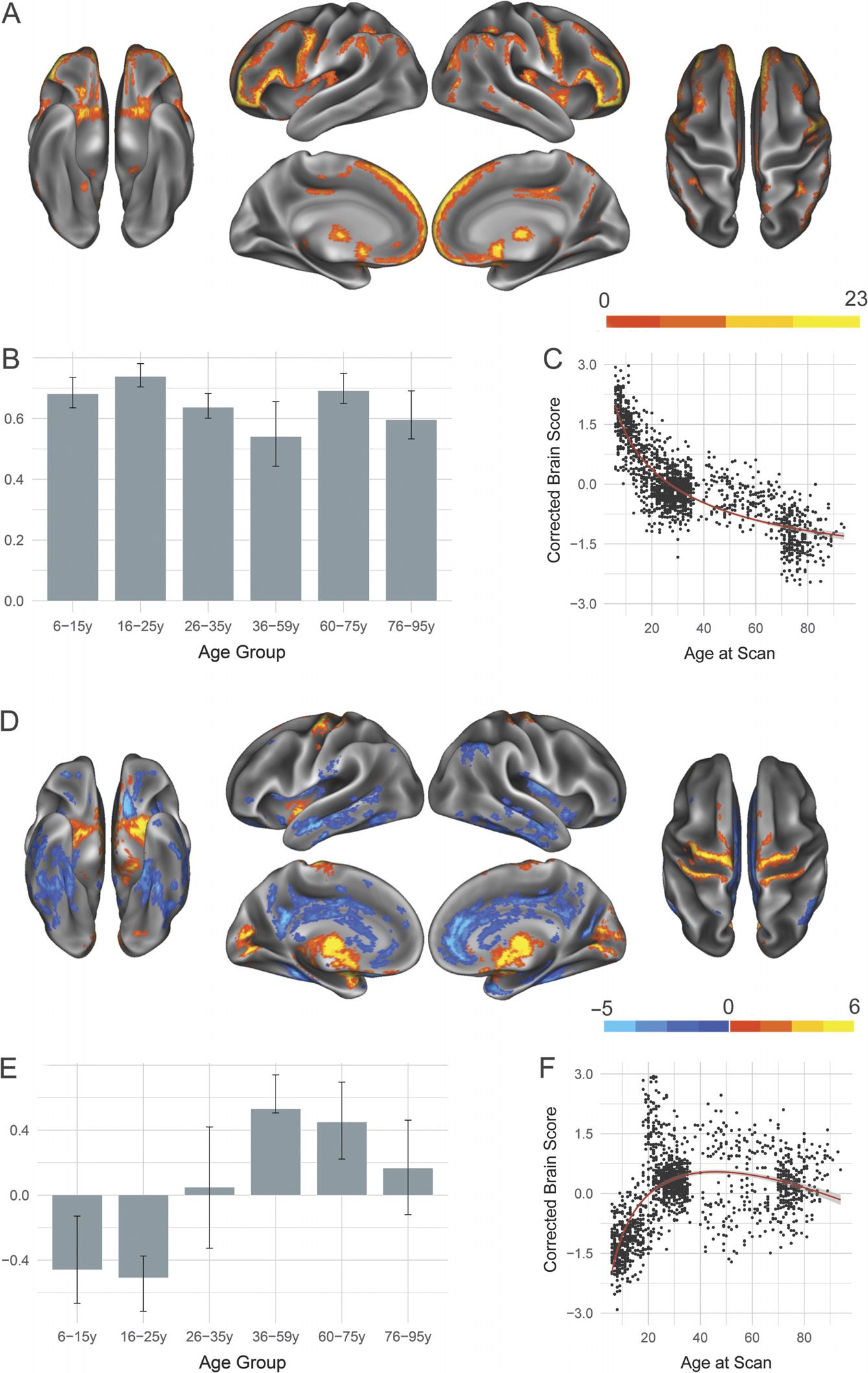
Two significant LVs were identified for the DN and are presented in Figure 2.
Default network.
In the first LV ( p < 0.0002; 61.57% covariance explained), seeded regions, along with homolo-
gous contralateral regions, covary together as well as with parahippocampal cortex and lateral
temporal cortex (Figure 2A). Covariance extended to noncanonical DN regions including pos-
terior insula. All age groups showed a robust positive association with this pattern (Figure 2B);
this suggests that this latent variable corresponds to the structural covariance of the DN as it
is preserved across the life span. Extracted brain scores (Figure 2C) revealed that the integrity
of this structural covariance pattern declines with advancing age rapidly before reaching a
plateau at approximately 70 years of age.
The second significant LV (p < 0.0002; 13.71% covariance explained) showed structural
covariance patterns of developmental change in the DN. Age Group 2 (16–25 years) showed
a unique pattern of increased structural covariance with medial prefrontal cortex and ante-
rior insula compared with all other age groups examined (Figure 2D, 2E). Age groups with
reliable correlations of brain scores and behavior—those for which the confidence interval
did not cross zero and were therefore considered interpretable—included the Age Group 1
(6–15 years), Age Group 3 (26–35 years), Age Group 5 (60–75 years), and Age Group 6 (76–
94 years) cohorts. Compared with Age Group 2 (16–25 years), each of these cohorts showed
relatively increased structural covariance between seeded DN regions and sensorimotor struc-
tures, including motor and visual cortices as well as thalamus. Across the life span, this pattern
shows a nearly linear decrease with advancing age (Figure 2F), suggesting that older adults are
less strongly aligning to the structural covariance pattern depicted in Figure 2D.
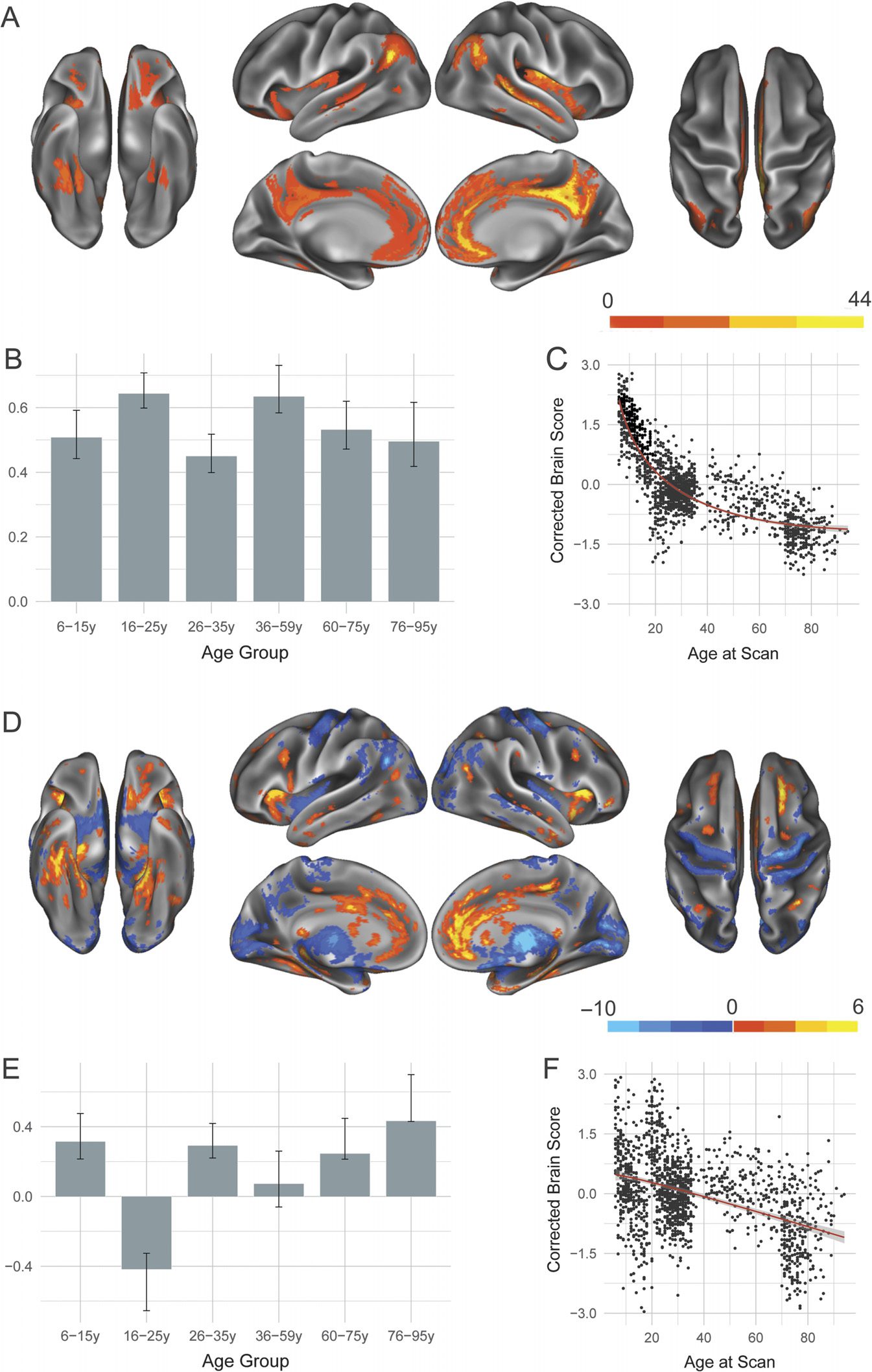
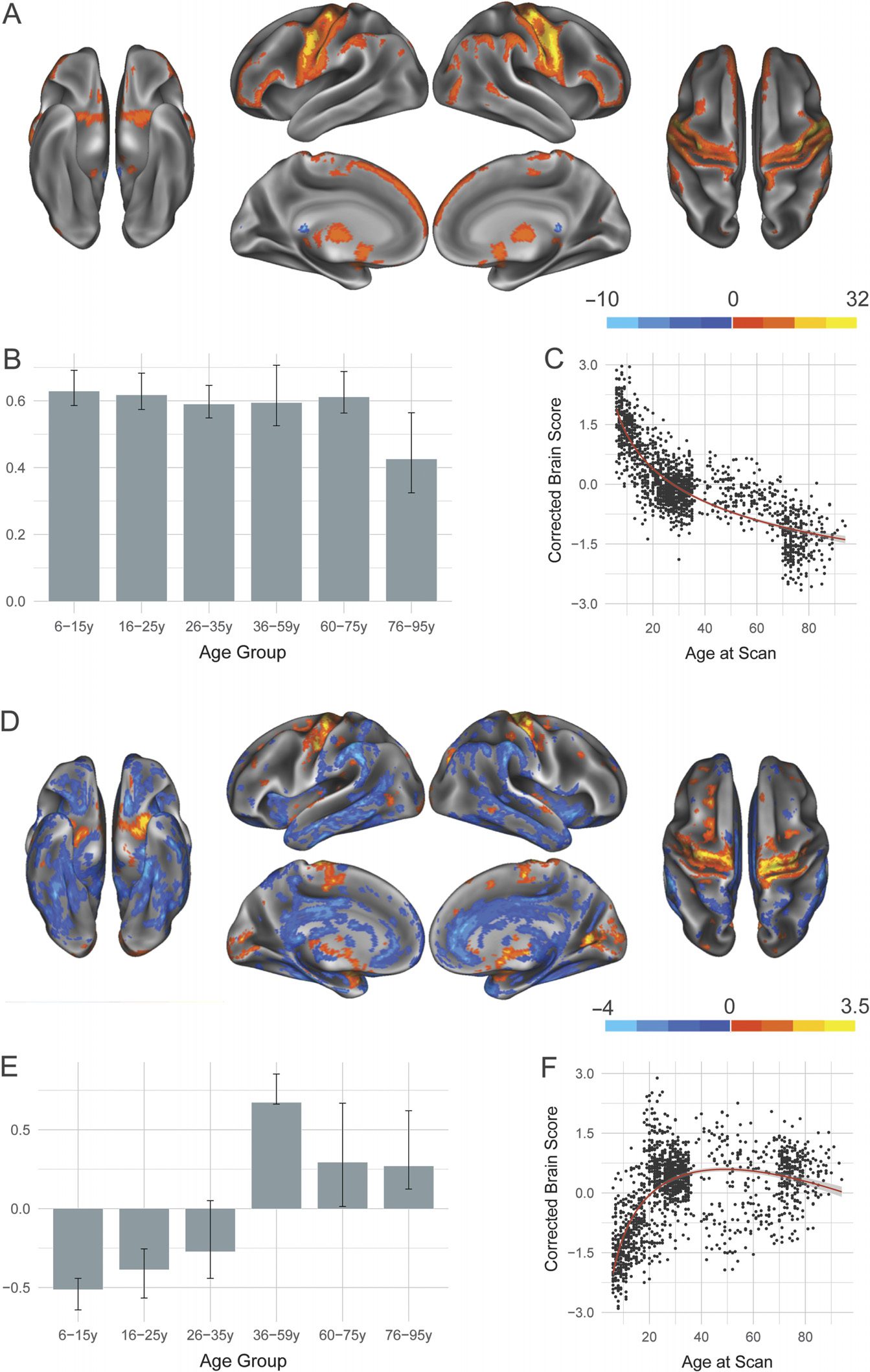
Two significant LVs were identified for the DAN and are presented
Dorsal attention network.
in Figure 3. In line with results presented for the DN, the first significant DAN LV ( p < 0.0002;
70.93% covariance explained) showed seeded regions positively covarying together as well as
with canonical DAN regions such as intraparietal sulcus (Figure 3A). Covariance also extended
to other, noncanonical DAN regions, including posterior insula and subgenual cingulate. All
age groups showed a robust association with this pattern (Figure 3B). Brain scores reveal that
the integrity of this pattern shows rapid decline with advancing age before plateauing at ap-
proximately 70 years (Figure 3C).
Network Neuroscience
309
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Structural covariance of the default network. (A) The spatial pattern for the first latent
variable, thresholded at 95% of the bootstrap ratio. (B) The bootstrapped correlation of brain scores
with the averaged gray matter volume estimates of default network seeds by age group for the first
latent variable. (C) The individual brain scores from the first latent variable corrected for scanner
strength and gender are plotted as a function of age. (D) Spatial pattern for the second latent vari-
able, thresholded at 95% of the bootstrap ratio. (E) The bootstrapped correlation of brain scores
with averaged gray matter volume estimates of default network seeds by age group for the second
latent variable. (F) The individual brain scores from the second latent variable corrected for scanner
strength and gender are plotted as a function of age.
Network Neuroscience
310
Lifespan structural covariance networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Structural covariance of the dorsal attention network. (A) The spatial pattern for the first
latent variable, thresholded at 95% of the bootstrap ratio. (B) The bootstrapped correlation of brain
scores with averaged gray matter volume estimates of dorsal attention network seeds by age group
for the first latent variable. (C) The individual brain scores from the first latent variable corrected
for scanner strength and gender are plotted as a function of age. (D) Spatial pattern for the second
latent variable, thresholded at 95% of the bootstrap ratio. (E) The bootstrapped correlation of brain
scores with averaged gray matter volume estimates of dorsal attention network seeds for the second
latent variable. (F) The individual brain scores from the second latent variable corrected for scanner
strength and gender are plotted as a function of age.
Network Neuroscience
311
Lifespan structural covariance networks
The second significant LV ( p < 0.0002; 9.52% covariance explained) revealed develop-
mental changes in the structural covariance pattern of the DAN. Age Group 2 (16–25 years)
showed uniquely increased structural covariance with medial prefrontal cortex and anterior
insula. Older age groups show relatively increased structural covariance between the seeded
DAN regions and areas including motor and visual cortices as well as subcortical structures.
Inspection of brain scores (Figure 3F) reveals an inverted U-shaped trajectory, with integrity of
the structural covariance pattern reaching its peak in middle adulthood, while very young and
very old individuals show significantly less integrity for the derived group structural covariance
patterns.
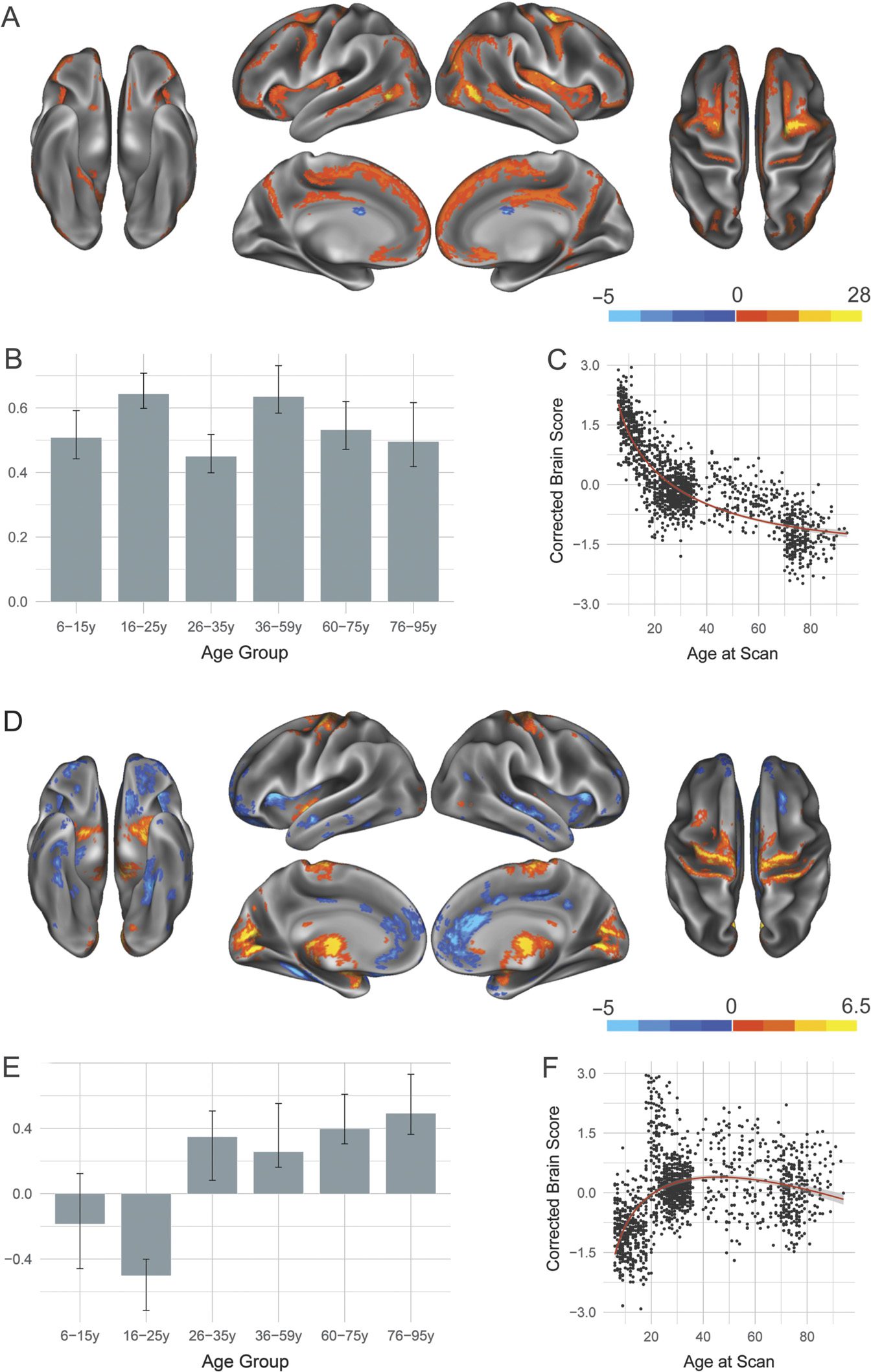
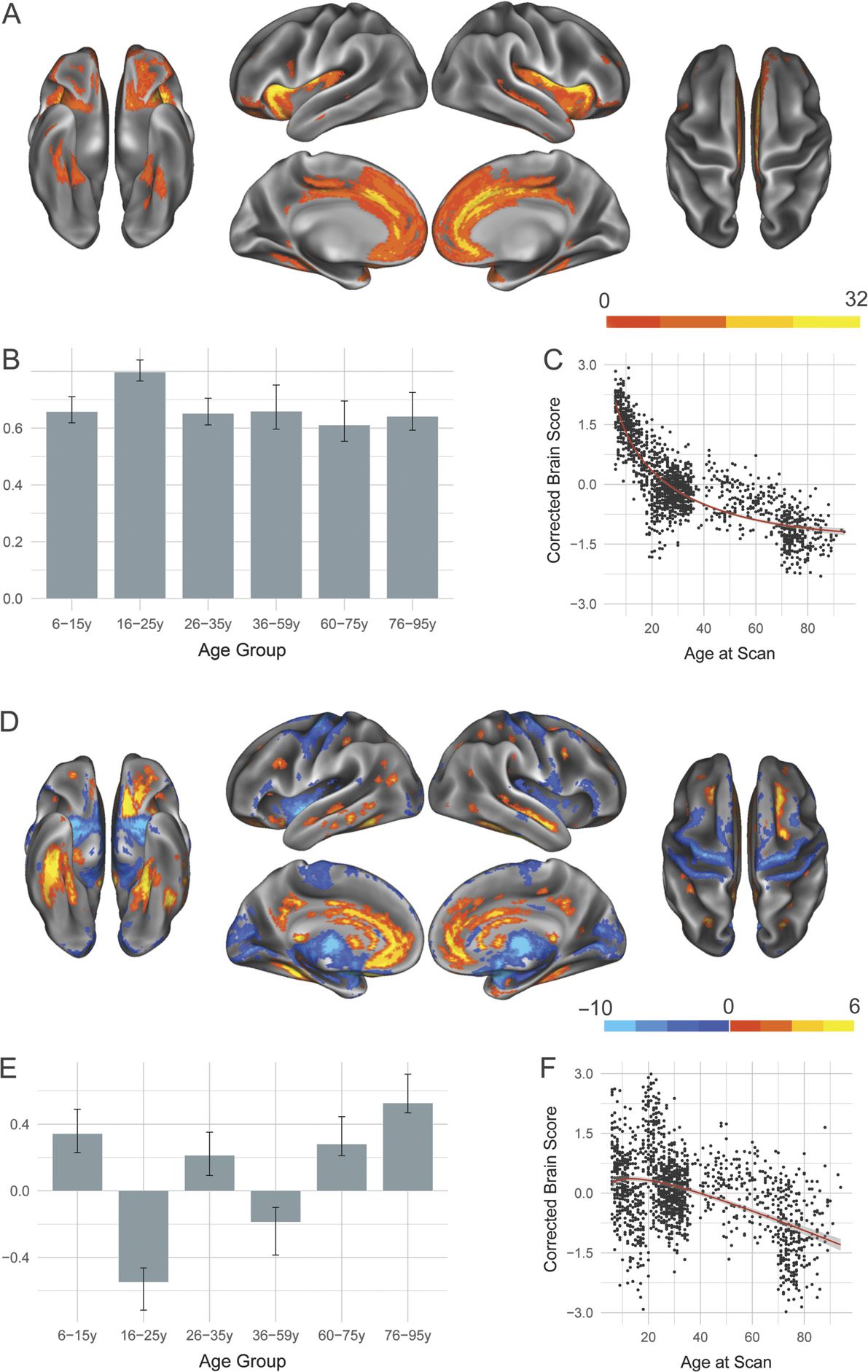
Two significant LVs were identified for the FPCN and are de-
Frontoparietal control network.
picted in Figure 4. Similar to results seen for the DN and DAN, the first significant LV (p <
0.0002; 78.05% covariance explained) showed a structural covariance pattern that was pos-
itively associated with all examined age groups, but showed a nonlinear decline in integrity
across the life span. Seeded FPCN regions positively covary together, as well as with structures
consistently associated with cognitive control, such as lateral prefrontal cortex, and noncanon-
ical FPCN regions, such as posterior insula.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The second LV ( p < 0.0002; 7.96% covariance explained) revealed developmental tra-
jectories of structural covariance patterns in the FPCN. There was a significant dissociation
between Age Groups 1 and 2 (6–25 years) as compared with middle and late Age Groups 4
and 5 (36–75 years). Younger age groups show increased structural covariance with structures
both within the canonical FPCN such as precuneus as well as with noncanonical regions such
as lateral temporal cortex. Older age groups, however, show relatively increased structural
covariance for sensorimotor structures such as motor cortex and thalamus. Brain scores sug-
gest an inverted U-shaped trajectory similar to that seen for the DAN, with the integrity of the
structural covariance pattern at its highest levels in middle adulthood.
Two significant LVs were identified for the SM and are depicted in
Somatomotor network.
In agreement with the previously reported networks, the first significant LV ( p <
Figure 5.
0.0002; 72.91% covariance explained) showed a structural covariance pattern that is positively
associated with all examined age groups and shows a nonlinear decline with advancing age.
Seeded regions covaried together as well as with the motor strip. Covariance extended to areas
outside the canonical motor network such as lateral prefrontal cortex and subcortical regions.
The second LV ( p < 0.0002; 9.17% covariance explained) showed a significant dissociation
between Age Groups 1 and 2 (6–25 years) as compared with Age Groups 4, 5, and 6 (36–
94 years). Younger age groups show increased structural covariance with structures outside
of the canonical motor network such as lateral temporal cortex and mid-insula, while older
age groups show relatively increased structural covariance local to the seed regions and to
thalamus. Similar to the individual subject score trajectories seen for the DAN and FPCN,
there is an inverted U-shaped trajectory in the integrity of this structural covariance pattern,
with integrity reaching a peak in middle adulthood before beginning to decline.
Two significant LVs were identified for the VAN and are presented
Ventral attention network.
in Figure 6. The first LV ( p < 0.0002; 70.93% covariance explained) again shows a structural
covariance pattern positively associated with all examined age groups. Seeded VAN regions
positively covary together and with the mid- and posterior insula as well as with the medial
Network Neuroscience
312
Lifespan structural covariance networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Structural covariance of the frontoparietal control network. (A) The spatial pattern for the
first latent variable, thresholded at 95% of the bootstrap ratio. (B) The bootstrapped correlation of
brain scores with averaged gray matter volume estimates of frontoparietal control network seeds. (C)
The individual brain scores from the first latent variable corrected for scanner strength and gender
are plotted as a function of age. (D) Spatial pattern for the second latent variable, thresholded at 95%
of the bootstrap ratio. (E) The bootstrapped correlation of brain scores with averaged gray matter
volume estimates of default network seeds by age group for the second latent variable.
(F) The
individual brain scores from the second latent variable corrected for scanner strength and gender
are plotted as a function of age.
Network Neuroscience
313
Lifespan structural covariance networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Structural covariance of the somatomotor network. (A) The spatial pattern for the first
latent variable, thresholded at 95% of the bootstrap ratio. (B) The bootstrapped correlation of brain
scores with bootstrapped averaged gray matter volume estimates of somatomotor network seeds by
age group for the first latent variable. (C) The individual brain scores from the first latent variable
corrected for scanner strength and gender are plotted as a function of age. (D) Spatial pattern for the
second latent variable, thresholded at 95% of the bootstrap ratio. (E) The bootstrapped correlation
of brain scores with averaged gray matter volume estimates of somatomotor network seeds by age
group for the second latent variable. (F) The individual brain scores from the second latent variable
corrected for scanner strength and gender are plotted as a function of age.
Network Neuroscience
314
Lifespan structural covariance networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Structural covariance of the ventral attention network. (A) The spatial pattern for the first
latent variable, thresholded at 95% of the bootstrap ratio. (B) The bootstrapped correlation of brain
scores with averaged gray matter volume estimates of ventral attention network seeds by age group
for the first latent variable. (C) The individual brain scores from the first latent variable corrected for
scanner strength and gender are plotted as a function of age. (D) Spatial pattern for the second latent
variable, thresholded at 95% of the bootstrap ratio. (E) The bootstrapped correlation of brain scores
with averaged gray matter volume estimates of ventral attention network seeds by age group for the
second latent variable. (F) The individual brain scores from the second latent variable corrected for
scanner strength and gender are plotted as a function of age.
prefrontal cortex. Extracted brain scores revealed a nonlinear decline in the integrity of this
structural covariance pattern across the life span.
The second significant LV ( p < 0.0002; 13.83% covariance explained) revealed a pat-
tern of developmental change similar to that seen in DN, with individuals in Age Group 2
Network Neuroscience
315
Lifespan structural covariance networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
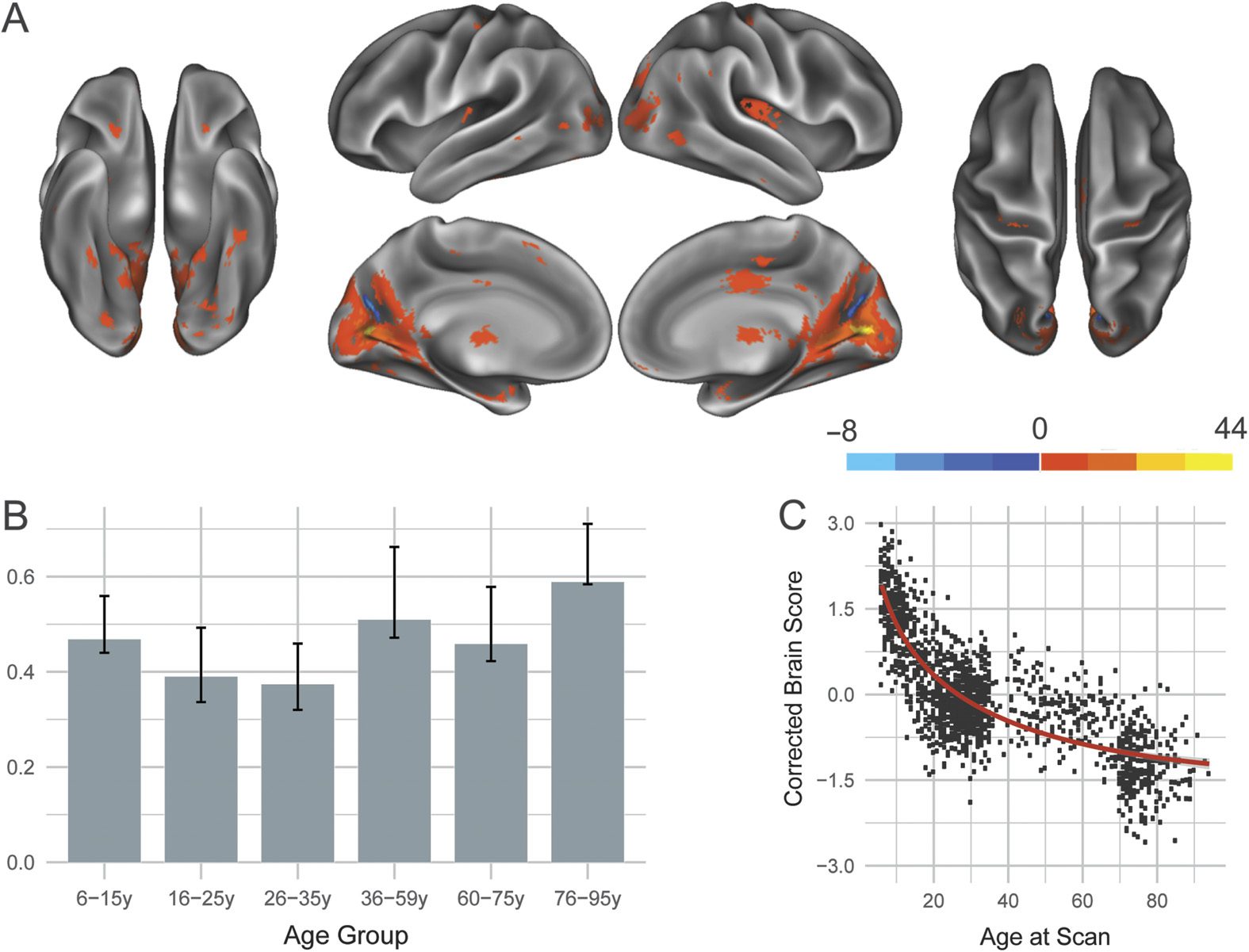
Figure 7. Structural covariance of the visual network. (A) The spatial pattern for the first latent
variable, thresholded at 95% of the bootstrap ratio. (B) The bootstrapped correlation of brain scores
with averaged gray matter volume estimates of visual network seeds by age group for the first latent
variable. (C) The individual brain scores from the first latent variable corrected for scanner strength
and gender are plotted as a function of age.
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
(16–25 years) and Age Group 4 (36–59 years) showing a unique structural covariance pattern
compared with all other age groups. Specifically, these two groups showed increased struc-
tural covariance with medial prefrontal as well as parahippocampal cortex. Other age groups
showed increased structural covariance with sensorimotor structures such as motor and visual
cortices. Similarly to the DN, there is a near linear decrease in structural integrity across the
life span, with older adults showing decreased structural covariance between seeded VAN
regions and sensorimotor structures.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Visual network. One significant LV ( p < 0.0002; 58.28% covariance explained) was identi-
fied for the visual network and is presented in Figure 7. As in previous networks, the significant
LV revealed a structural covariance pattern that was positively associated with all examined
age groups and nonlinearly declined with age. Seeded visual regions showed positive struc-
tural covariance with visual cortex as well as with noncanonical visual regions such as the
posterior insula and mid-cingulate.
DISCUSSION
In this study, we examined the life span trajectories of structural covariance with publicly
available cross-sectional data. For the six neurocognitive networks examined, our results
revealed two broad developmental patterns: a stable pattern of structural covariance that
Network Neuroscience
316
Lifespan structural covariance networks
reflects network-specific features and persists across the life span, and an age-dependent pat-
tern of structural covariance that reveals shared age-related trajectories of structural covariance
across networks.
Persistent Patterns of Structural Covariance
Across all networks, the first significant latent variable identified a structural covariance pattern
whose spatial extent was unique to the network of interest and persisted across age groups.
Despite the stability of these structural covariance patterns over the life span, inspection of
individual subject or “brain” scores (panel C, Figures 2– 7) revealed that integrity of these
patterns declines rapidly with advancing age before plateauing at approximately 70 years.
These findings extend on previous work showing a sharp decline in within-network struc-
tural covariance from young adulthood to middle age that persists into older adulthood (Li
et al., 2013). Although our results show that network-specific structural covariance patterns
were stable across the life span, we find that children and adolescents show even higher levels
of integrity to these structural covariance patterns compared with young adults. This decline in
integrity to structural covariance patterns over the life span may be related both to the increase
in myelination across early development and its effects on gray-white matter tissue contrast
(Lenroot & Giedd, 2006), as well as to the decline of cortical gray matter volume with age
(Allen, Bruss, Brown, & Damasio, 2005).
Age-Dependent Patterns of Structural Covariance
In addition to stable patterns of structural covariance, DN, DAN, FPCN, SM, and VAN
networks showed an additional, age-dependent pattern that differentiated young adulthood
from either end of the life span.
Examination of brain scores in the DAN, FPCN, and SM (panel F, Figures 3– 5) reveals
that these align with an inverted U-shaped trajectory. These latent variables also showed
overlapping features of structural covariance at a group level (panel D of Figures 3– 5).
In
young adulthood, seeded regions showed structural covariance with areas including medial
prefrontal cortex, posterior cingulate, insular cortex, and temporal cortex—association cor-
tices corresponding to functional hubs (van den Heuvel & Sporns, 2013). In both childhood
and older adulthood, however, seeded regions showed structural covariance with sensorimotor
structures including motor and visual cortices and thalamus. These findings support previous
work showing that structural covariance networks grow increasingly distributed over early de-
velopment before shifting to a more localized topology in advanced aging (Wu et al., 2012).
Our results in the DAN, FPCN, and SM suggest that distributed patterns of structural covari-
ance peak in middle adulthood before returning to a relatively localized topology in older
adulthood.
The second significant latent variables of the DN and VAN share spatial features of struc-
tural covariance with the second latent variables of the DAN, FPCN, and SM; however, their
trajectories (panel F, Figures 3and 6) do not show a reliable, inverted U-shape. One possible
explanation for this is that the selected seed regions for the DN and VAN included regions
such as the medial prefrontal cortex, posterior cingulate, and insular cortex. These regions are
known functional hubs (van den Heuvel & Sporns, 2013) and, in the second latent variable of
DAN, FPCN, and SM, their structural covariance reliably differentiates young adulthood from
other portions of the life span. In PLS, successive latent variables contribute unique, additional
portions of variance. Since these seed regions strongly contribute to the structural covariance
Network Neuroscience
317
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
of the DN and VAN first latent variables, it is possible that the appearance of a near linear
decline in the second latent variable—rather than an inverted U-shaped trajectory—is due to
the exclusion of medial prefrontal cortex, posterior cingulate, and insular cortex from the sec-
ond latent variables of the DN and VAN and their explained covariance. This would suggest
that these regions are particularly important in shaping age-dependent patterns of structural
covariance.
Previous investigations of structural covariance have found variation in the extent to which
networks show age-related changes. For example, relatively flat patterns of structural covari-
ance across adulthood have been seen in the visual network (Li et al., 2013) as well as in
temporal, auditory, and cerebellar networks (Hafkemeijer et al., 2014). Our finding that the
visual network did not have a significant second latent variable suggests that there is not a
significant age-dependent pattern of structural covariance for this network, in agreement with
this previous work.
Contrary to our initial hypotheses, we did find age-dependent structural covariance trajec-
tories for SM, where existing literature suggests that there are little to no age-related changes
(Li et al., 2013). In the present work, we find SM to exhibit the same age-dependent pattern of
structural covariance as DAN and FPCN. Future longitudinal studies of structural covariance
patterns will be important to address the impact of age on specific cortical networks.
Overall, our results therefore suggest that the structural covariance patterns of large-scale
neurocognitive networks each have a unique spatial topology; however, neurocognitive
networks also show overlapping patterns of age-dependent structural covariance.
Relationship of Structural Covariance to Function
Structural covariance networks have been extensively linked to neural function via their
marked disruptions in pathology and pathological aging (Bassett et al., 2008; Hafkemeijer
et al., 2016; Spreng & Turner, 2013; Valk, Martino, Milham, & Bernhardt, 2015). Alongside
functional connectivity, shared structural covariance has been suggested as a defining char-
acteristic of large-scale networks (Seeley, Crawford, Zhou, Miller, & Grecius, 2009; see also
Di et al., 2017). It is worth considering, therefore, these life span patterns of structural covari-
ance in light of the existing literature on the development of functional connectivity across the
life span.
In our work, the first significant latent variable seen in all examined networks showed a
stable pattern of structural covariance whose integrity declined across the life span. This is
similar to patterns of decreasing within-network functional connectivity with advancing age
(Betzel et al., 2014). The second latent variable seen in all networks—with the exception of the
visual network—showed an age-dependent pattern that distinguished young adulthood from
both childhood and advanced aging. These results mirror developmental trajectories com-
monly reported in functional connectivity studies with increased functional integration across
networks in childhood, peak functional segregation between networks in young adulthood,
and de-differentiation of network functional connectivity in older adulthood (Collin & van den
Heuvel, 2013).
The significant overlap of structural covariance trajectories found in the current investiga-
tion and those trajectories reported in the functional connectivity literature suggest that a life
span perspective may help illuminate the noted relationship between structural covariance
and neural function. Directly assessing the relationship between structural covariance and
Network Neuroscience
318
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
functional connectivity, however, is a topic for future research aided by the collection of
multimodal imaging data in life span samples (e.g., Glasser et al., 2016; Nooner et al., 2012).
Methodological Considerations
Although this study was able to leverage the increasing amount of anatomical data available in
open-access repositories, it included important methodological considerations related to age
group definition, scanner acquisition strength, and motion correction. Although we sought
to create cohorts representing neurobiologically meaningful age ranges, this resulted in un-
equal representation in both sample size and age range considered. Our smallest included
age group, Age Group 6 (76–94 years), included 134 participants, while our largest age group,
Age Group 3 (26–35 years), included 472 participants. Although differing sample sizes across
groups will invariably yield more variable estimates of group-wise covariance, we note that our
estimates have statistical power comparable to the smallest group size considered. At 134 sub-
jects, this is still significantly higher powered than current standards for MRI data collection,
particularly in life span samples. An additional consideration with our selection of age cohorts
is the age range considered in each age group. Age Group 4, defined here as ages 36–59,
spans a larger time period than any of the other cohorts considered. This was in large part
due to the paucity of openly available data for that cohort, particularly when compared with
other cohorts such as younger adulthood. The continuing collection of multimodal data for
life span initiatives such as NKI-RS (Nooner et al., 2012), the HCP Lifespan Project (Glasser
et al., 2016), and UK Biobank (Miller et al., 2016) will increase the availability of high-quality
data to investigate such questions.
Differences in scanner acquisition strength across data sources provide an additional im-
portant methodological consideration. Several data sources, including those representing the
youngest and oldest subjects, were acquired at 1.5T, while young adults were acquired at
3T. Although it is likely that subtle differences between groups may have been introduced
by MR field strength, inspection of individual subject scores from the two included life span
data sources (OASIS and NKI-RS) indicate that these subjects do not show divergent results
from those seen in the age-restricted datasets or from one another. This is suggestive of general
agreement in structural covariance trends across scanner field strength, as OASIS was collected
at 1.5T while NKI-RS was collected at 3T. Further, we controlled for MR field strength across
groups by adjusting individually derived subject scores and found similar results for both raw
and corrected subject scores. Future work assessing structural covariance across the life span
should nonetheless aim to examine scans acquired at the same MR field strength and ideally
on the same scanner.
A limitation of the current study is the inability to implement motion correction of structural
images. Recent work has shown that head motion may introduce artifacts into anatomical im-
ages, affecting automated estimates of structure (Alexander-Bloch et al., 2016; Savalia et al.,
2016). Although acquisition of a resting-state scan has been proposed to flag high-motion
subjects for exclusion from structural analyses (Alexander-Bloch et al., 2016; Savalia et al.,
2016), not all of the datasets utilized also provided at least one resting-state scan for each
subject. We therefore caution that estimates of age group differences may be inflated by un-
corrected motion. Each of these methodological considerations can be addressed in future
work, as comprehensive samples of participants across the life span with both structural and
functional imaging become increasingly available.
In this study we utilized open-access, cross-sectional data sources to examine structural co-
variance patterns of six neurocognitive networks across the life span. Using multivariate PLS
Network Neuroscience
319
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
analysis, we found that all networks exhibited stable patterns of network-specific structural
covariance, and with the exception of the visual network showed a second, age-dependent
pattern of structural covariance that mirrored developmental trends seen in the functional con-
nectivity literature. The present results confirm the utility of structural covariance in defining
neurocognitive networks and reveal both shared and network-specific trajectories of structural
covariance across the life span.
ACKNOWLEDGMENTS
NIH Peds data used in the preparation of this article were obtained from the NIH Pediatric MRI
Data Repository created by the NIH MRI Study of Normal Brain Development. This is a mul-
tisite, longitudinal study of typically developing children from ages newborn through young
adulthood conducted by the Brain Development Cooperative Group and supported by the
National Institute of Child Health and Human Development, the National Institute on Drug
Abuse, the National Institute of Mental Health, and the National Institute of Neurological
Disorders and Stroke (Contract numbers N01-HD02-3343, N01-MH9-0002, and N01-NS-
9-2314, -2315, -2316, -2317, -2319, and -2320). A listing of the participating sites and a
complete listing of
the study investigators can be found at http://pediatricmri.nih.gov/
nihpd/info/participating_centers.html. HCP data were provided by the Human Connectome
Project, WU-Minn Consortium (principal investigators: David Van Essen and Kamil Ugurbil;
1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint
for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at
Washington University. NKI data were obtained from the Nathan Kline Institute – Rockland
Sample, Release 5. Principal support for the enhanced NKI-RS project was provided by the
NIMH BRAINS R01MH094639-01 (principal investigator: Michael Milham). Funding for key
personnel was also provided in part by the New York State Office of Mental Health and Re-
search Foundation for Mental Hygiene. Funding for the decompression and augmentation of
administrative and phenotypic protocols was provided by a grant from the Child Mind Insti-
tute (1FDN2012-1). Additional personnel support was provided by the Center for the Devel-
oping Brain at the Child Mind Institute, as well as NIMH R01MH081218, R01MH083246,
and R21MH084126. Project support was also provided by the NKI Center for Advanced
Brain Imaging (CABI), the Brain Research Foundation, and the Stavros Niarchos Foundation.
OASIS data were supported by the following grants: P50 AG05681, P01 AG03991, R01
AG021910, P50 MH071616, U24 RR021382, R01 MH56584. ADNI data used in prepara-
tion of this article were obtained from the Alzheimer’s Disease Neuroimaging Initiative (ADNI)
database (http://adni.loni.usc.edu). As such, the investigators within the ADNI contributed
to the design and implementation of ADNI and/or provided data but did not participate in
analysis or writing of this report. A complete listing of ADNI investigators can be found
at http://adni.loni.usc.edu/wp-content/uploads/how_to_apply/ADNI_Acknowledgement_List.
pdf. Data collection and sharing for this project was funded by the Alzheimer’s Disease
Data collection and sharing for this project was funded by the Alzheimer’s Disease Neu-
roimaging Initiative (ADNI) (National Institutes of Health Grant U01 AG024904) and DOD
ADNI (Department of Defense award number W81XWH-12-2-0012). ADNI is funded by
the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengi-
neering, and through generous contributions from the following: AbbVie; Alzheimer’s Asso-
ciation; Alzheimer’s Drug Discovery Foundation; Araclon Biotech; BioClinica, Inc.; Biogen;
Bristol-Myers Squibb Company; CereSpir, Inc.; Eisai, Inc.; Elan Pharmaceuticals, Inc.; Eli Lilly
and Company; EuroImmun; F. Hoffmann-La Roche, Ltd., and its affiliated company, Genen-
tech, Inc.; Fujirebio; GE Healthcare; IXICO, Ltd.; Janssen Alzheimer Immunotherapy Research
Network Neuroscience
320
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
& Development, LLC.; Johnson & Johnson Pharmaceutical Research & Development, LLC.;
Lumosity; Lundbeck; Merck & Co., Inc.; Meso Scale Diagnostics, LLC.; NeuroRx Research;
Neurotrack Technologies; Novartis Pharmaceuticals Corporation; Pfizer, Inc.; Piramal Imag-
ing; Servier; Takeda Pharmaceutical Company; and Transition Therapeutics. The Canadian
Institutes of Health Research is providing funds to support ADNI clinical sites in Canada. Pri-
vate sector contributions are facilitated by the Foundation for the National Institutes of Health
(http://www.fnih.org). The grantee organization is the Northern California Institute for Research
and Education, and the study is coordinated by the Alzheimer’s Disease Cooperative Study at
the University of California, San Diego. ADNI data are disseminated by the Laboratory of
Neuro Imaging at the University of Southern California. This manuscript reflects the views of
the authors and may not reflect the opinions or views of the NIH.
AUTHOR CONTRIBUTIONS
Elizabeth DuPre: Conceptualization; Formal analysis; Visualization; Writing – original draft.
R. Nathan Spreng: Conceptualization; Formal analysis; Funding acquisition; Methodology;
Supervision; Writing – original draft.
FUNDING INFORMATION
This work was supported in part by an Alzheimer’s Association grant (NIRG-14-320049) to
R.N.S.
REFERENCES
Alexander-Bloch, A., Clasen, L., Stockman, M., Ronan, L., Lalonde,
F., Giedd, J., & Raznahan, A. (2016). Subtle in-scanner motion
biases automated measurement of brain anatomy from in vivo
MRI. Human Brain Mapping, 37, 2385–2397.
Alexander-Bloch, A., Giedd, J. N., & Bullmore, E.
Imag-
ing structural co-variance between human brain regions. Nature
Reviews: Neuroscience, 14, 322–336.
(2013).
Alexander-Bloch, A., Raznahan, A., Bullmore, E., & Giedd,
J.
(2013). The convergence of maturational change and structural
covariance in human cortical networks. Journal of Neuroscience,
33, 2889–2899.
Allen, J. S., Bruss, J., Brown, C. K., & Damasio, H.
(2005). Nor-
mal neuroanatomical variation due to age: The major lobes
and a parcellation of the temporal region. NeuroImage, 26,
1245–1260.
Ashburner, J.
(2007). A fast diffeomorphic registration algorithm.
NeuroImage, 38, 95–113.
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry—
The methods. NeuroImage, 11, 805–821.
Bassett, D. S., Bullmore, E., Verchinski, B. A., Mattay, V. S.,
Weinberger, D. R., & Meyer-Lindenberg, A. (2008). Hierarchical
organization of human cortical networks
in health and
schizophrenia. Journal of Neuroscience, 28, 9239–9248.
Betzel, R. F., Byrge, L., He, Y., Goñi, J., Zuo, X. N., & Sporns, O.
(2014). Changes in structural and functional connectivity among
resting-state networks across the human lifespan. NeuroImage,
102, 345–357.
Brain Development Cooperative Group & Evans, A. C. (2007). The
NIH MRI study of normal brain development. NeuroImage, 30,
184–202.
Caviness, J. V. S., Kennedy, D. N., Richelme, C., Rademacher, J.,
& Filipek, P. A. (1996). The human brain age 7–11 years: A vol-
umetric analysis based on magnetic resonance images. Cerebral
Cortex, 6, 726–736.
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig,
G. S. (2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences, 111, 4997–5006.
Collin, G., & van den Heuvel, M. P. (2013). The ontogeny of the hu-
man connectome: Development and dynamic changes of brain
connectivity across the life span. Neuroscientist, 19, 616–628.
Cox, R. W.
(1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29, 162–173.
Di, X., Gohel, S., Thielcke, A., Wehrl, H. F., Biswal, B. B., &
Alzheimer’s Disease Neuroimaging Initiative.
(2017). Do all
roads lead to Rome? A comparison of brain networks derived
from inter-subject volumetric and metabolic covariance and
moment-to-moment hemodynamic correlations in old individ-
uals. Brain Structure and Function. https://doi.org/10.1007/
s00429-017-1438-7
Dosenbach, N. U. F., Nardos, B., Cohen, A. L., Fair, D. A., Power,
J. D., Church, J. A., . . . Schlaggar, B. L.
(2010). Prediction of
individual brain maturity using fMRI. Science, 329, 1358–1361.
Network Neuroscience
321
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lifespan structural covariance networks
DuPre, E., & Spreng, R. N.
(2017). Supplemental material for
“Structural covariance networks across the life span from 6 to
94 years of age.” Network Neuroscience, 1(3), 302–323. https://
doi.org/10.1162/netn_a_00016
Eckart, C., & Young, G. (1936). The approximation of one matrix
by another of lower rank. Psychometrika, 1, 211–218.
Fjell, A. M., Westlye, L. T., Espeseth, T., Reinvang, I., Walhovd,
K. B., Dale, A. M., & Holland, D. (2010). Cortical gray matter at-
rophy in healthy aging cannot be explained by undetected incip-
ient cognitive disorders: A comment on Burgmans et al. (2009).
Neuropsychology, 24, 258–266.
Giorgio, A., Santelli, L., Tomassini, V., Bosnell, R., Smith, S.,
De Stefano, N., & Johansen-Berg, H. (2010). Age-related changes
in grey and white matter structure throughout adulthood. Neu-
roImage, 51, 943–951.
Glasser, M. F., Smith, S. M., Marcus, D. S., Andersson, J. L. R.,
Auerbach, E. J., Behrens, T. E. J., . . . Van Essen, D. C. (2016). The
Human Connectome Project’s neuroimaging approach. Nature
Neuroscience, 19, 1175–1187.
Hafkemeijer, A., Altmann-Schneider,
J. M.,
Slagboom, P. E., van der Grond, J., & Rombouts, S. A. R. B.
(2014). Associations between age and gray matter volume in
anatomical brain networks in middle-aged to older adults. Aging
Cell, 13, 1068–1074.
I., de Craen, A.
J. C.,
Hafkemeijer, A., Möller, C., Dopper, E. G. P., Jiskoot, L. C., van den
. . . Rombouts,
Berg-Huysmans, A. A., van Swieten,
S. A. R. B.
(2016). Differences in structural covariance net-
works between behavioral variant frontotemporal dementia and
Alzheimer’s disease. Human Brain Mapping, 37, 978–988.
Hagmann, P., Sporns, O., Madan, N., Cammoun, L., Pienarr, R.,
(2010). White matter matura-
Wedeen, V. J., . . . Grant, P. E.
tion reshapes structural connectivity in the late developing hu-
man brain. Proceedings of the National Academy of Sciences,
107, 19067–19072.
Holmes, A. J., Hollinshead, M. O., O’Keefe, T. M., Petrov, V. I.,
Fariello, G. R., Wald, L. L., . . . Buckner, R. L.
(2015). Brain
Genomics Superstruct Project initial release with structural, func-
tional, and behavioral measures. Scientific Data, 2. https://doi.
org/10.1038/sdata.2015.31
Krishnan, A., Williams, L. J., McIntosh, A. R., & Abdi, H.
(2011).
Partial Least Squares (PLS) methods for neuroimaging: A tutorial
and review. NeuroImage, 56, 455–475.
Krongold, M., Cooper, C., & Bray, S.
(2017). Modular develop-
ment of cortical gray matter across childhood and adolescence.
Cerebral Cortex, 27, 1125–1136.
Kurth, F., Luders, E., & Gaser, C.
(2010). VBM8 toolbox manual,
http://www.neuro.uni-jena.de/vbm/
Lenroot, R. K., & Giedd, J. N. (2006). Brain development in children
and adolescents: Insights from anatomical magnetic resonance
imaging. Neuroscience & Biobehavioral Reviews, 3(6), 718–729.
Li, X., Pu, F., Fan, Y., Niu, H., Li, S., & Li, D. (2013). Age-related
changes in brain structural covariance networks. Frontiers in
Human Neuroscience, 7. https://doi.org/10.3389/fnhum.2013.
00098
connections. Philosophical Transactions of the Royal Society B,
361, 1531–1544.
McIntosh, A., Bookstein, F., Haxby, J., & Grady, C. (1996). Spatial
pattern analysis of functional brain images using partial least
squares. NeuroImage, 3, 143–157.
Mechelli, A., Friston, K. J., Frackowiak, R. S., & Price, C. J. (2005).
Journal of Neuro-
Structural covariance in the human cortex.
science, 25, 8303–8310.
Mesulam, M. M. (1998). From sensation to cognition. Brain, 121,
1013–1052.
Miller, K. L., Alfaro-Almagro, F., Bangerter, N. K., Thomas, D. L.,
Yacoub, E., Xu, J., . . . Smith, S. M. (2016). Multimodal popula-
tion brain imaging in the UK Biobank prospective epidemiolog-
ical study. Nature Neuroscience, 19, 1523–1536.
Miši´c, B., Betzel, R. F., de Reus, M. A., van den Heuvel, M. P.,
Berman, M. G., McIntosh, A. R., & Sporns, O. (2016). Network-
structure-function relationships in human neocortex.
level
Cerebral Cortex, 26, 3285–3296.
Montembeault, M., Joubert, S., Doyon, J., Carrier, J., Gagnon, J. F.,
Monchi, O., . . . Brambati, S. M.
(2012). The impact of aging
on gray matter structural covariance networks. NeuroImage, 63,
754–759.
Muzik, O., Chugani, D. C., Juhász, C., Shen, C. G., & Chugani,
(2000). Statistical parametric mapping: Assessment of
H. T.
application in children. NeuroImage, 12, 538–549.
Nooner, K. B., Colcombe, S. J., Tobe, R. H., Mennes, M., Benedict,
M. M., Moreno, A. L., . . . Milham, M. P.
(2012). The NKI-
Rockland sample: A model for accelerating the pace of discovery
science in psychiatry. Frontiers in Neuroscience, 6. https://doi.
org/10.3389/fnins.2012.00152
Pagani, M., Bifone, A., & Gozzi, A. (2016). Structural covariance
networks in the mouse brain. NeuroImage, 129, 55–63.
Persson,
J., Spreng, R. N., Turner, G., Herlitz, A., Morell, A.,
Stening, E., . . . Söderlund, H.
Sex differences in
volume and structural covariance of the anterior and posterior
hippocampus. NeuroImage, 99, 215–225.
(2014).
Raz, N., Lindenberger, U., Rodrigue, K. M., Kennedy, K. M.,
Head, D., Williamson, A.,
(2005). Re-
gional brain changes in aging healthy adults: General trends,
Cerebral Cortex, 15,
individual differences and modifiers.
1676–1689.
. . . Acker,
J. D.
Savalia, N. K., Agres, P. F., Chan, M. Y., Feczko, E. J., Kennedy,
K. M., & Wig, G. S. (2016). Motion-related artifacts in structural
brain images revealed with independent estimates of in-scanner
head motion. Human Brain Mapping, 38, 472–492.
Seeley, W. W., Crawford, R. K., Zhou, J., Miller, B. L., & Grecius,
(2009). Neurodegenerative diseases target large-scale
M. D.
human brain networks. Neuron, 62, 42–52.
Spreng, R. N., & Turner, G. R.
(2013). Structural covariance of
the default network in healthy and pathological aging. Journal of
Neuroscience, 33, 15226–15234.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Tijms, B. M., Seris, P., Willshaw, D. J., & Lawrie, S. M.
(2012).
individual networks from gray
Similarity-based extraction of
matter MRI scans. Cerebral Cortex, 22, 1530–1541.
Low, L. K., & Cheng, H.-J.
(2006). Axon pruning: An essen-
tial step underlying the developmental plasticity of neuronal
Uddin, L. Q., Kelly, A. M. C., Biswal, B. B., Castellanos, F. X., &
Milham, M. P. (2009). Functional connectivity of default mode
Network Neuroscience
322
Lifespan structural covariance networks
network components: Correlation, anticorrelation, and causality.
Human Brain Mapping, 30, 625–637.
Valk, S. L., Martino, A. D., Milham, M. P., & Bernhardt, B. C. (2015).
Multicenter mapping of structural network alterations in autism.
Human Brain Mapping, 36, 2364–2373.
van den Heuvel, M. P., & Sporns, O. (2013). Network hubs in the
human brain. Trends in Cognitive Neurosciences, 17, 683–696.
Wu, K., Taki, Y., Sato, K., Kinomura, S., Goto, R., Okada, K., . . .
Fukuda, H. (2012). Age-related changes in topological organiza-
tion of structural brain networks in healthy individuals. Human
Brain Mapping, 33, 552–568.
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari,
D., Hollinshead, M., . . . Buckner, R. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
connectivity. Journal of Neurophysiology, 106, 1125–1165.
Zielinski, B. A., Gennatas, E. D., Zhou, J., & Seeley, W. W. (2010).
Network-level structural covariance in the developing brain.
Proceedings of the National Academy of Sciences, 107, 18191–
18196.
Zuo, X.-N., He, Y., Betzel, R. F., Colcombe, S., Sporns, O., &
Milham, M. P. (2017). Human connectomics across the life span.
Trends in Cognitive Sciences, 21, 32–45.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
3
3
0
2
1
0
9
2
0
6
2
n
e
n
_
a
_
0
0
0
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
323