Randomized Manipulation of Early Cognitive
Experience Impacts Adult Brain Structure
Martha J. Farah1, Saul Sternberg1, Thomas A. Nichols1, Jeffrey T. Duda1,
Terry Lohrenz2, Yi Luo2, Libbie Sonnier2, Sharon L. Ramey2,
Read Montague2, and Craig T. Ramey2
Astratto
■ Does early exposure to cognitive and linguistic stimulation im-
pact brain structure? Or do genetic predispositions account for
the co-occurrence of certain neuroanatomical phenotypes and
a tendency to engage children in cognitively stimulating activities?
Low socioeconomic status infants were randomized to either
5 years of cognitively and linguistically stimulating center-based
care or a comparison condition. The intervention resulted in large
and statistically significant changes in brain structure measured in
midlife, particularly for male individuals. These findings are the
first to extend the large literature on cognitive enrichment effects
on animal brains to humans, and to demonstrate the effects of
uniquely human features such as linguistic stimulation. ■
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
C
N
_
UN
_
0
1
7
0
9
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
How does early life experience shape the human brain? IL
question is surprisingly difficult to answer, as it concerns
the causes, rather than merely the correlates, of individual
differences in human development. Studies of such differ-
ences are normally observational and thus silent on the
subject of causality. Animal studies, in contrast, have dem-
onstrated causal influence of environmental stimulation on
brain structure using random assignment to physical envi-
ronments with low or high complexity. Tuttavia, Essi
cannot tell us about the features of the environment that
matter most for human development: linguistic and cogni-
tive stimulation.
The role of the environment in shaping brain develop-
ment is a central issue for neuroscience, and a significant
open question concerns the impact of uniquely human
features of the environment, namely, linguistic and cogni-
tive stimulation (Lenroot & Giedd, 2011). Whereas a large
animal literature shows that more complex cage environ-
ments lead to microscopic and macroscopic brain changes,
including larger cortex (Diamond, 2001), such manipula-
tions provide an incomplete model for the environmental
differences that may matter most in human development.
These include differences in complex forms of cognitive
and linguistic experience.
Understanding how experience shapes human develop-
ment is also a central issue for social science and policy.
Does early experience drive socioeconomic stratification
across generations? Can environmental interventions
1University of Pennsylvania, 2Virginia Polytechnic Institute and
State University
enhance the development in individuals of lower socio-
economic status (SES) and disrupt intergenerational cy-
cles of disadvantage (Peterson, Loeb, & Chamberlain,
2018; Duncan, Magnuson, & Votruba-Drzal, 2014)? In an
era of growing inequality and persistent child achievement
gaps, the response of the human brain to early childhood
cognitive and linguistic experiences has societal, as well as
scientific, importance (Farah, 2018).
To address these questions an experiment is needed,
with human infants randomly assigned to environments
of high versus low cognitive and linguistic stimulation,
ideally starting early in life and comprising a substantial
portion of their early childhood years. Although it would
be unethical and unfeasible to experimentally assign a
group of children to low cognitive and linguistic stimula-
zione, below what it would otherwise have been, there is
an alternative way to achieve the equivalent contrast. It
has long been reported that children growing up in lower
SES families, on average, receive less cognitive and linguis-
tic stimulation compared with their higher SES peers
(Hoff, 2013; Bradley & Corwyn, 2002). By randomizing
such infants into one group that continues to receive the
expected low stimulation and one that receives higher
linguistic and cognitive stimulation, the effect of randomly
assigned high versus low stimulation can be observed.
This was the intervention design of the Abecedarian
Project (Ramey et al., 2000). Starting between 3 E 21 weeks
of age, and continuing through age 5 years, participants in
the intervention group engaged in a program designed to
promote linguistic interactions and age-appropriate learning
opportunities. Randomization was constrained to equate
the two groups for multiple poverty-associated risk factors,
and the two groups eventually scanned four decades later
© 2021 Istituto di Tecnologia del Massachussetts. Published under a
Creative Commons Attribution 4.0 Internazionale (CC BY 4.0) licenza.
Journal of Cognitive Neuroscience 33:6, pag. 1197–1209
https://doi.org/10.1162/jocn_a_01709
remained well-matched on these factors as described in
Methods section. Both the early intervention group and
the comparison group received enhanced medical care
and social services.
Participants were evaluated throughout the period of the
intervention and over the subsequent decades. Cognitive
benefits of the early intervention, assessed with IQ and
academic achievement tests, were significant through the
latest evaluation at age 21 years, although smaller than
when measured in childhood (Campbell, Pungello, Mugnaio-
Johnson, Burchinal, & Ramey, 2001). Larger and enduring
effects occurred in real-world behavioral achievements in-
cluding additional years of education completed, greater
likelihood of a 4-year college graduation, lower reliance on
public assistance, older age at first child, and greater rates of
full-time employment (Campbell, Pan, & Burchinal, 2019).
Sex shapes life trajectories and health in myriad ways,
biological and social, in many cases with boys more affected
by adverse environments than girls, even within the same
family (Golding & Fitzgerald, 2017; Bale et al., 2010). Within
the Abecedarian project, sex differences emerged over time
in some but not all analyses. Different outcomes showed fe-
male advantage, male advantage, or no sex difference
(Campbell et al., 2019), a mix of outcomes that has also been
observed in other early childhood education programs
(Magnuson et al., 2016). These differences in intervention ef-
fects have yet to be satisfactorily explained, but their existence
motivates the inclusion of sex as a moderator in this study.
For the present research, structural MRI scans were
obtained from 47 of the Abecedarian sample, 29 from the
early intervention group and 18 from the comparison group.
As shown in Table 1, the groups were closely matched on a
number of characteristics that would be expected to cor-
relate with brain structure, including mothers’ IQ, educa-
tional attainment and age at birth, infant gestational age
and head circumference at birth, composite risk index
(see Methods section), sex (15/29 O 52% E 9/18 O
50% male in the early intervention and comparison
groups, rispettivamente), and race (all African American).
The size of the sample deserves comment. Primo, as de-
tailed in Methods section, power analysis indicates that the
sample is adequate under the assumption that the effect to
be detected is large (by Cohen’s classification of effect sizes;
Cohen, 1992). Previous research, summarized in Methods
section, suggests that the effects of sustained environmental
stimulation will indeed be large. Secondo, even if sample size
were a concern, the sustained randomized manipulation
of cognitive and linguistic stimulation followed by brain
imaging is unprecedented. The unique research opportu-
nity presented by a full-time, 5 days/week intervention
lasting the first 5 years of life warrants examination.
The brain measures of primary interest were the vol-
umes of five specific ROIs, summed to create a primary
summary measure, as well as the volume of cortex more
generally. Four of the specific ROIs were selected for their
a priori relevance to the intervention, which emphasized
language for communication and as scaffolding for cogni-
tive control (see Methods): left inferior frontal gyrus (LIFG)
and left superior temporal gyrus (LSTG) relevant to lan-
guage (Friederici, 2011), right inferior frontal gyrus and
bilateral ACC, relevant to cognitive control (Aron, Robbins,
& Poldrack, 2004). The fifth, bilateral hippocampus, era
Tavolo 1. Characteristics of the Participants in the Successfully Scanned Comparison and Early Intervention Groups
Variable
Comparison Group
Early Intervention Group
Maternal Characteristics at Enrollment
Maternal educational attainment
Maternal IQ
Maternal age at birth
Participant Characteristics at Enrollment
Sex
Race/ethnicity
High-risk index (see Methods)
Gestational age at birth
Head circumference at birth
Participant Characteristics at Time of Scan
Age at scan
SAI status
AA = African American; SAI = School Age Intervention.
10.50 (2.04) sì
85.5 (9.64)
21.28 (6.91) sì
15/29, 52% male
18/18, 100% AA
19.83 (5.42)
39.44 (3.52) wks
34.06 (2.21) cm
41.22 (1.67) sì
9/18, 50% SAI
10.46 (1.53) sì
84.62 (9.02)
18.72 (2.42) sì
9/18, 50% male
29/29, 100% AA
19.93 (5.91)
39.32 (2.50) wks
34.33 (1.52) cm
41.38 (1.57) sì
15/29, 52% SAI
1198
Journal of Cognitive Neuroscience
Volume 33, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
C
N
_
UN
_
0
1
7
0
9
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 2. Factors Included in the Computation of Each
Participant’s High Risk Index, Measuring Poverty and
Poverty-associated Risk Factors at Time of Enrollment,
with Contributions of Each
Tavolo 2. (continued )
Factor and Level
Weighted
Contribution
Factor and Level
Low Maternal Educational Attainment
(Highest Grade Completed)
Weighted
Contribution
6
7
8
9
10
11
12
Low Paternal Educational Attainment
(Highest Grade Completed)
6
7
8
9
10
11
12
Family Income (Per Year), dollars
>1000
1,001–2,000
2,001–3,000
3,001–4,000
4,001–5,000
5,001–6,000
Father absent for reasons other than health
or death
Absence of maternal relatives in local area
(cioè., parents, grandparents, or brothers or
sisters of majority age)
Siblings of school age who are one or more
grades behind age-appropriate grade or
who score equivalently low on school-
administered achievement test
Payments received from welfare agencies
within past 3 years
8
7
6
3
2
1
0
8
7
6
3
2
1
0
8
7
6
5
4
0
3
3
3
3
Record of father’s work indicated unstable
and unskilled or semi-skilled labor
Records of mother’s or father’s IQ indicate
score of 90 or below
Records of sibling’s IQ indicate score of
90 or below
Relevant social agencies in the community
indicate that the family is in need of
assistance
One or more members of the family has
sought counseling or professional help
in the past 3 years
Special circumstances not included in any
of the above that are likely contributors
to cultural or social disadvantage
3
3
3
3
1
1
added because its volume is frequently associated with
early life adversity including poverty (Hanson et al., 2015).
Unless otherwise specified, volumes were expressed as
percentages of the mean comparison group volume for
the same sex, allowing us to report intervention effects
in percentage differences, a more meaningful measure
than cubic centimeters.
The effects of the early intervention on brain structure
were assessed using standard and permutation-based
tests. Initially, we assessed the effects of the early interven-
tion and possible moderation by sex, using standard and
robust ANOVA on the two relatively global measures: IL
summed volumes of ROIs and total cortical volume. A later
School-Age Intervention (SAI), not found to affect behav-
ioral measures in the long term (Campbell et al., 2019) E
balanced between the two early intervention groups, era
also included as a covariate. The five a priori ROIs were
then tested individually. Additional exploratory analyses
included early intervention effects on the surface areas
and thicknesses of the cortical ROIs and the relations of
brain measures to selected psychological measures. Finalmente,
the volumes, surface areas, and thicknesses of all brain
areas from the Desikan–Killiany atlas (Desikan et al., 2006)
were also assessed.
METHODS
Participants
The Abecedarian Project (Ramey et al., 2000) was estab-
lished in North Carolina in the early 1970s and enrolled
112 prevalentemente (98%) African American infants from
homes of very low SES (low income and maternal educa-
zione) with multiple associated risk factors such as paternal
absence, welfare receipt, and low parental IQ (Tavolo 2),
but free of neurodevelopmental disorder. In order to
Farah et al.
1199
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
C
N
_
UN
_
0
1
7
0
9
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
equate the early intervention and comparison groups on
various demographic and risk factors, pairs of children
with equivalent baseline measures were randomly allocated
to each condition. One of the 112 infants initially ran-
domized to early intervention later received a diagnosis of
a congenital condition that was disqualifying based on the
exclusionary criteria, resulting in 111 infants participating
in the study. The intervention was a comprehensive pro-
gram of developmentally appropriate cognitive and lin-
guistic enrichment embedded within a positive and
responsive, university-based childcare setting for five full
days (6–8 hr) per week, 50 weeks per year.
The Abecedarian Early Intervention was designed to
provide consistently high levels of individually paced cog-
nitive and language experiences, of the kind more com-
mon in higher SES families. The program utilized the
Learning Games curriculum (Sparling & Lewis, 1984),
which is based on the Vygotskian view of the centrality
of language in cognitive development—that children learn
self-regulation by internalizing speech. Infant activities in-
cluded talking to the child, playing with cause-and-effect
toys or picture books, and offering infants an opportunity
to react to sights and sounds in the environment. As chil-
dren grew, the curriculum shifted toward more concep-
tual and skill-based learning games and interactions,
always using language, even in motor skill activities,
and eliciting language from the child.
Both the early intervention group and the comparison
group were provided with free iron-fortified baby formula
(because none were breastfed), and with social workers to
facilitate access to free or low-cost healthcare for the first
5 years of life, as well as family social services. Di conseguenza,
outcome differences would not be attributable to these
factors and both groups correctly viewed themselves as
part of a treatment group.
Over the ensuing decades, participants were evaluated via
blinded assessment on their functioning in various important
spheres of life including cognitive, educational, social–
emotional, occupational, economic, and health outcomes.
Cognition was assessed with IQ and academic achievement
tests of reading and mathematical skill. Tests of more specific
neurocognitive abilities such as executive function were not
administered. Although the IQ and academic skills advantage
faded with time, more enduring benefits were observed in
other important behavioral outcomes including years of ed-
ucation completed, likelihood of college graduation, reliance
on public assistance, age at first child, and continuity of
employment (Campbell et al., 2019; Ramey et al., 2000).
Seventy-eight study participants (42 intervention and
36 comparison) traveled to Roanoke for follow-up testing
between the ages of 38 E 44 years. Eighteen were not
scanned because of anxiety or claustrophobia, in some
cases related to their girth relative to the scanner bore
(8 interventions and 10 comparisons); eight were not
scanned because of metal in the body (three interventions
and five comparisons); one was not scanned because of
weight alone (comparison group); and one was not scanned
because of recent neurological symptoms (intervention
group). One intervention participant declined with no rea-
son offered, and one comparison participant’s scan failed
because of hardware error. Finalmente, del 48 completed
scans, one was of poor quality (comparison), leaving a total
Di 47 images. Twenty-nine of these (15 male, 14 female)
came from the intervention group, E 18 (9 male, 9 female)
came from the comparison group. Mean age at time of scan
era 41.4 E 41.2 years (SD = 1.6 E 1.7) for the interven-
tion and comparison groups, rispettivamente.
Imaging
Imaging was conducted on a 3.0-T Siemens Trio scanner. High-
resolution T1-weighted scans (voxel size: 1.0 × 1.0 × 1.0 mm)
were acquired using an MPRAGE sequence (Siemens). Each
participant’s T1 data were processed using Advanced
Normalization Tools (ANTs; Avants, Epstein, Grossman, &
Gee, 2008). The antsMultivariateTemplateConstruction.sh
script was used to build a template using all participants’
T1 images. The population-specific template was pro-
cessed using the antssCorticalThickness.sh tool in order
to obtain a set of tissue segmentation priors. Each scan
was then processed using the population-specific template
along with antsCorticalThickness.sh (Tustison et al., 2014).
This pipeline produces a brain extraction mask and a
six-tissue segmentation. Jacobian images of volume were
calculated from the nonlinear warp fields that align each
participant to the template. To obtain cortical labels for
each participant, the antsJointLabelFusion.sh script was
used along with an existing population of labeled images
to perform multi-atlas label fusion, which provides both
cortical labels as well as deep gray labels (Wang et al., 2013).
Behavioral Measures
Although not the primary focus of this research, behavioral
data were also analyzed. These analyses were aimed at as-
sessing the relation of the brain measures used in this study
to individual psychological outcomes. Two behavioral mea-
sures were selected for these analyses. One, contempora-
neous with the early intervention, was the Stanford–Binet
intelligence test (Form L-M), administered at age 4 years
by staff blind to group assignment. All but two of the
scanned participants had taken this test and therefore had
scores available. The other behavioral measure, obtained
the day of scanning, was a midlife strengths and risk index.
This was extracted from structured interviews conducted by
research staff blind to participants’ group assignments. IL
index was computed by adding together two checklists,
each out of 10, of strengths (such as high school graduate
and current full-time employment) and reverse-coded risk
(such as unfavorable self-rated health and first child before
age 20 years; Sonnier-Netto, 2018). We note that four other
assessments were administered on this occasion but were
not analyzed in relation to brain structure because they did
not measure an ability or quality of performance along a
quantitative dimension of better or worse. These were an
1200
Journal of Cognitive Neuroscience
Volume 33, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
C
N
_
UN
_
0
1
7
0
9
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
open-ended interview by a staff member not blind to group
assignment, an Ultimatum Game, a Multi-round Trust
Game, and a Locus of Control questionnaire (see Luo
et al., 2018, for a report on the economic games and
Sonnier-Netto, 2018, for a report on locus of control).
Analyses
All statistical analyses were carried out in R (2019), con
additional permutation analyses assisted by R package
lmPerm ( Wheeler & Torchiano, 2016), and bias-corrected
and accelerated confidence intervals calculated by boot-
strapping (Canty & Ripley, 2019; Peng, 2019; Weiss, 2016).
For analyses reported here, unless otherwise noted,
brain measures were normalized as percentages of the
mean control participant of the corresponding sex as
follows: Relative volume = 100(v − (cid:1)vs)/ (cid:1)vs, Dove (cid:1)vs is
the corresponding same-sex mean volume.
The resulting proportions provide a more intuitive mea-
sure of the intervention effect than absolute volumes mea-
sured in cc. Inoltre, by using the comparison mean
from the same-sex participants, we eliminate the size dif-
ference between male and female brains from these per-
centage increase measures.
To determine whether or not these measures would
need to be corrected for other participant characteristics
that could affect brain outcomes, we examined the rich
array of baseline measures available on the study partici-
pants and their families that are shown in Table 1. These
included age at scan, gestational age at birth, head circum-
ference at birth, maternal IQ, maternal educational attain-
ment, and overall “high risk” score. The groups were
highly similar on all measures, as shown in Table 1, so that
any difference in brain outcomes cannot be attributed to
differences in these baseline measures.
The hypothesis testing sequence progressed from ana-
tomically general to specific, questo è, normalized cortex and
summed normalized ROIs and then normalized individual
ROIs. The analyses of cortex and summed ROIs consisted
of standard and permutation-based ANOVA, including the
following variables: Early Intervention, SAI, Sex, and all
interactions among these variables. The effect of the early
intervention on the five ROIs was then assessed separately
for male and female individuals, with false discovery rate
(FDR) correction for the 10 multiple comparisons.
The sample size, although modest, provided adequate
power for detecting a large effect by Cohen’s classification.
Specifically, power analysis for the present sample, con
groups of 28 E 19 participants, indicates adequate power
by the conventional criteria of 80% power and p < .05
for an effect size of d = 0.84 (G*Power 3; Faul, Erdfelder,
Lang, & Buchner, 2007). The most similar research with
humans, comparing the effects of Romanian versus UK
orphanage environments, shows an effect of size d =
1.13 on gray matter volume (Mackes et al., 2020; Appendix).
Research that varies sustained environmental stimulation
experimentally has been carried out only with animals.
Although contemporary animal research in this area focuses
on molecular and cellular effects, some early work reported
macroscopic differences roughly analogous to those stud-
ied here, focused on cortical weight, length, width, and
thickness, largely in male rats (Diamond, 2001). Based on
an early publication that included a table with data required
for calculating a standardized effect size (Rosenzweig, 1966;
Table 1), the increase in cortex weight for rats given envi-
ronmental enrichment was d = 0.87.
Additional exploratory analyses were undertaken to
learn as much as possible from this unique data set.
First, effects of the early intervention with 95% confidence
intervals were assessed with normalized measures of total
surface area and mean cortical thickness of cortex as a
whole and the four a priori cortical ROIs for male and
female participants. Second, the normalized volumes of
all 134 regions of the Desikan–Killany atlas identified by
joint label fusion were then analyzed in the same manner
to gauge the effects of early intervention on them. Third,
to assess the relation of brain measures to psychological
outcomes, Pearson correlations were computed between
normalized brain measures on the one hand, and IQ and
midlife strengths and risk index on the other. Brain mea-
sures selected for testing were confined to the two rela-
tively global measures, namely, the sum of ROIs and
total cortex volume, to be augmented by any regional mea-
sure that showed significant effects of the intervention for
participants of both sexes. The results of these correlations
were FDR corrected. The goal of these analyses was to
determine whether the brain measures studied here
are related to psychological outcomes of interest.
RESULTS
Descriptive Overview of Anatomical Sequelae
Table 3 presents basic descriptive data concerning partic-
ipants’ brain volumes, separated by the manipulation of
interest, the early intervention, as well as by sex, given
other findings of sex differences in outcomes from early
intervention programs. In addition to regions selected
for a priori testing, included are also whole-brain volume
and the remainders of cortical and brain volumes when a
priori ROIs have been subtracted.
Table 4 presents the same results expressed as normal-
ized volumes relative to mean of same-sex comparison
participants.
Observe that 18 of the 20 entries in the Intervention
columns of Table 4 are positive, indicating that the early
intervention is associated with increased size of the whole
brain, the cortex, and most of the ROIs. Observe also
that, except for one region (the left inferior frontal gyrus),
the group treatment effects for males were substantially
greater than for females.
In order to visualize the distributions of these volume
measurements over participants, we plotted raw (as op-
posed to normalized) volumes separated by sex and
Farah et al.
1201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 3. Means and Standard Errors of Brain Region Volumes for Early Intervention and Comparison Participants
Female Mean (SE)
Male Mean (SE)
Percent Volume Increase
Comparison
Intervention
Comparison
Intervention
Areas of a priori Interest
Cortex
Sum of ROIs
ACC (bilat)
HC (bilat)
IFG (L)
IFG (R)
STG (L)
Remaining Compartments
400.77 (12.57)
402.09 (6.56)
25.59 (0.84)
26.49 (0.63)
5.4 (.11)
6.63 (0.48)
4.47 (0.17)
4.52 (0.16)
4.57 (0.30)
5.47 (0.28)
6.59 (0.11)
5.11 (0.17)
4.91 (0.19)
4.41 (0.13)
433.78 (7.8)
25.72 (0.6)
4.79 (0.27)
7.16 (0.28)
4.72 (0.13)
4.32 (0.15)
4.71 (0.16)
480.33 (9.29)
29.7 (.52)
6.24 (0.19)
7.61 (0.23)
5.30 (0.16)
5.36 (0.18)
5.11 (0.15)
Total brain
868.53 (27.91)
870 (14.66)
938.11 (14.84)
1030.91 (20.45)
Cortex net of ROIs
381.81 (12.21)
382.19 (6.18)
Brain net of cortex and HC
461.13 (16.77)
461.32 (9.99)
415.21 (7.68)
497.17 (8.81)
458.24 (9.13)
542.97 (12.69)
Abbreviations: ACC = anterior cingulate gyrus; HC = hippocampus; IFG = inferior frontal gyrus; STG = superior temporal gyrus.
intervention group for the two relatively global measures
of a priori interest: summed ROIs and total cortex. Figure 1
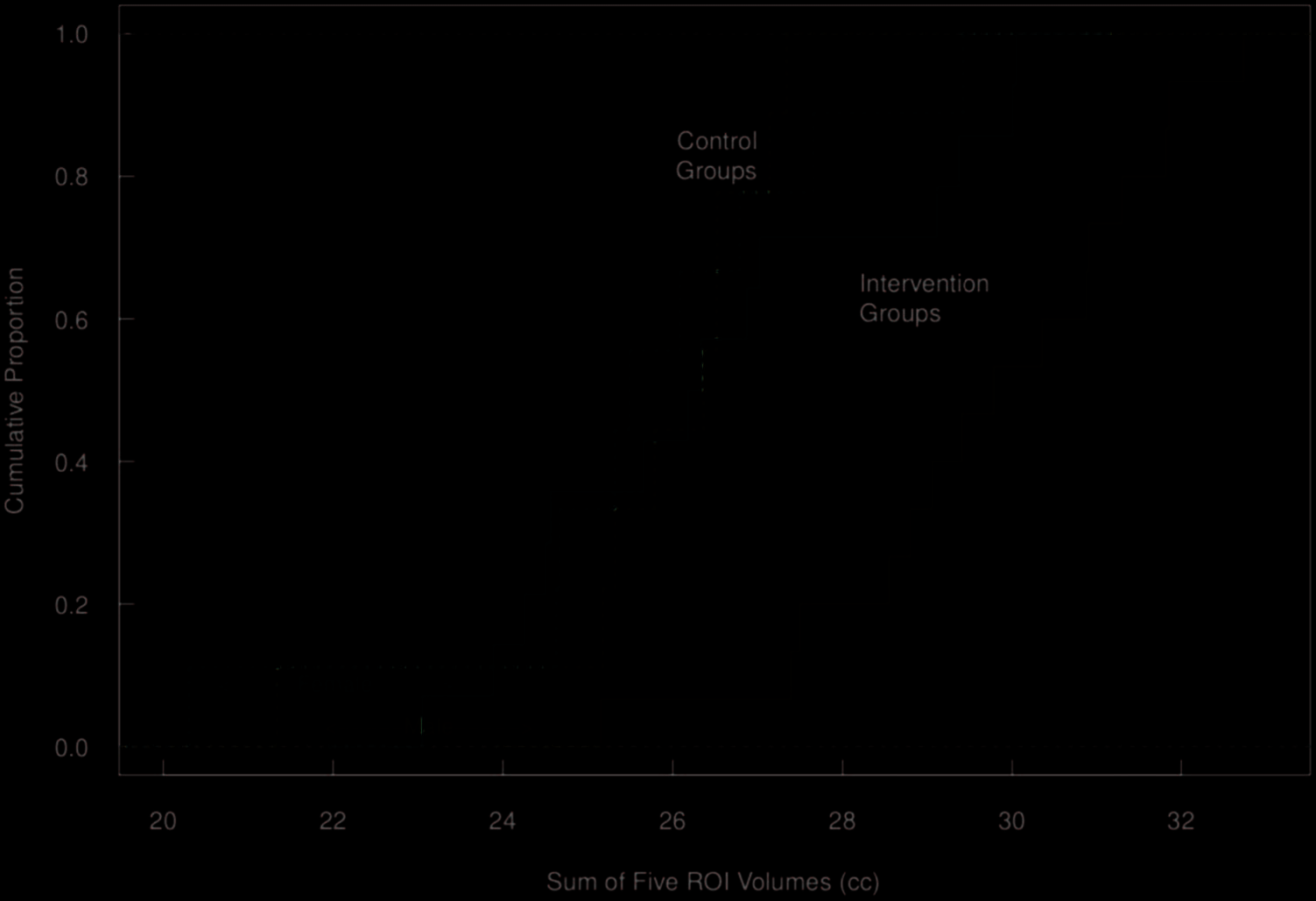
shows the effect of the intervention on the sum of ROIs,
substantially more pronounced in males, as well as the ex-
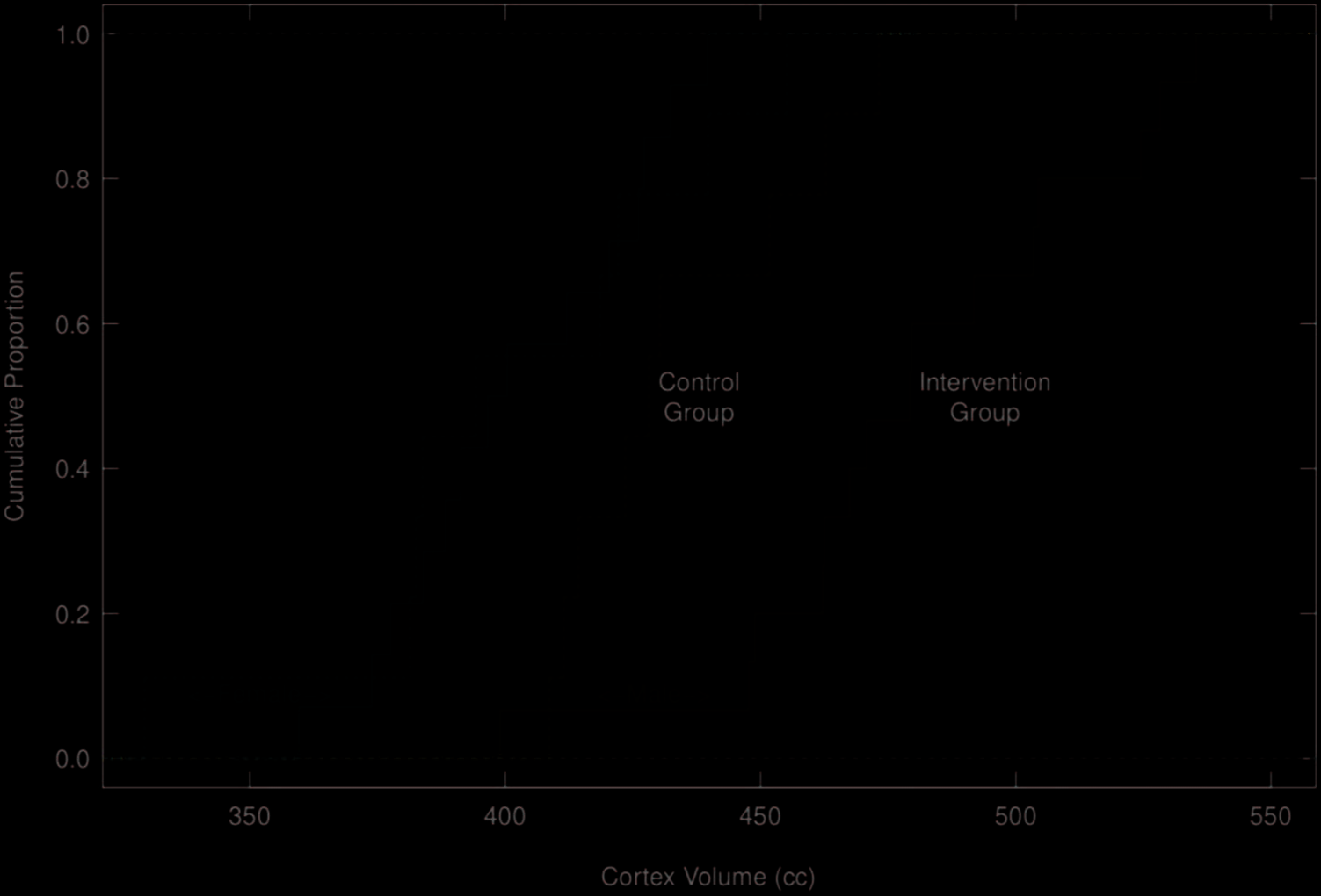
pected sex differences in volume. Figure 2 displays the
same relations for cortex volume, again showing sex differ-
ences in both volume and effect of intervention.
Size and Reliability of Intervention Effects
To assess the early intervention effects on the two most
global of the a priori measures, namely, the sum of the pre-
dicted ROIs and total cortical volume, we conducted anal-
yses of variance. Tables 5 and 6 show the results of these
analyses for summed ROI volumes and cortex volume,
Table 4. Means and Standard Errors of Brain Region Volume Percentage Increase, Relative to Same Sex Comparison Mean, for Early
Intervention and Comparison Participants
Percent Volume Increase
Comparison
Intervention
Comparison
Intervention
Female Mean (SE)
Male Mean (SE)
Areas of a priori Interest
Cortex
Sum of ROIs
ACC (bilat)
HC (bilat)
IFG (L)
IFG (R)
STG (L)
Remaining Compartments
Total brain
Cortex net of ROIs
Brain net of cortex and HC
Abbreviations as in Table 3.
0 (3.14)
0 (3.27)
0 (2.08)
0 (7.22)
0 (3.80)
0 (3.58)
0 (6.47)
0 (3.21)
0 (3.20)
0 (3.64)
+0.33 (1.64)
+3.50 (2.47)
+1.29 (5.12)
−0.67 (1.61)
+14.24 (3.88)
+8.61 (4.22)
−3.40 (2.84)
+0.17 (1.69)
+0.10 (1.62)
+0.04 (2.17)
0 (1.80)
0 (2.34)
0 (5.67)
0 (3.96)
0 (2.74)
0 (3.51)
0 (3.45)
0 (1.58)
0 (1.85)
0 (1.77)
+10.73 (2.14)
+15.46 (2.04)
+30.37 (4.04)
+6.27 (3.22)
+14.19 (3.31)
+23.88 (4.29)
+7.84 (3.22)
+9.89 (2.18)
+10.36 (2.20)
+9.21 (2.55)
1202
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Cumulative plot of
individual participants’ summed
ROI volume in each group,
separated by sex.
respectively, with factors Sex, SAI, and all interactions.
In these tables, PConv values are based on conventional
analyses of variance, using the F-distribution and the ob-
tained values of the F-statistic; PPerm values are based on
permutation-based ANOVA.
Standardized effect sizes for the early intervention effect,
expressed as Cohen’s d based on F values (Thalheimer &
Cook, 2002), are substantial: 1.61 for summed ROIs and
0.80 for cortex. These large effects sizes are for all partici-
pants combined, male and female.
As can be seen, for both summed ROI volumes and cor-
tex volume, analyzed with standard and permutation-
based ANOVA, the analyses agree on which differences
are important and which are negligible. Specifically, Early
Intervention, Sex, and their interaction are all significant,
consistent with the means shown in Table 4. The later, SAI
and all of its interactions are nonsignificant.
The size and reliability of the intervention effects in spe-
cific ROIs were then assessed for male and female participants.
Correcting for multiple comparisons across the 10 tests, male
Figure 2. Cumulative plot of
individual subjects’ cortex
volume in each group,
separated by sex.
Farah et al.
1203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 5. Analysis of Variance of Normalized Sum of ROI Volumes
Source
Early Intervention
Sex
School-Age Intervention (SAI)
Early Int. × Sex
Early Int. × SAI.
Sex × SAI
Early Int. × Sex × SAI
Residuals
Df
Mean Sq.
1
1
1
1
1
1
1
39
1042
638
127
431
4
0
100
74
F Value
14.12
8.65
1.72
5.84
0.06
0
1.36
PConv
0.001
0.006
0.200
0.020
0.807
0.986
0.251
PPerm
0.000
0.023
0.190
0.025
0.902
0.583
0.251
participants showed significant increases in three of the five
areas and female participants showed an increase in one.
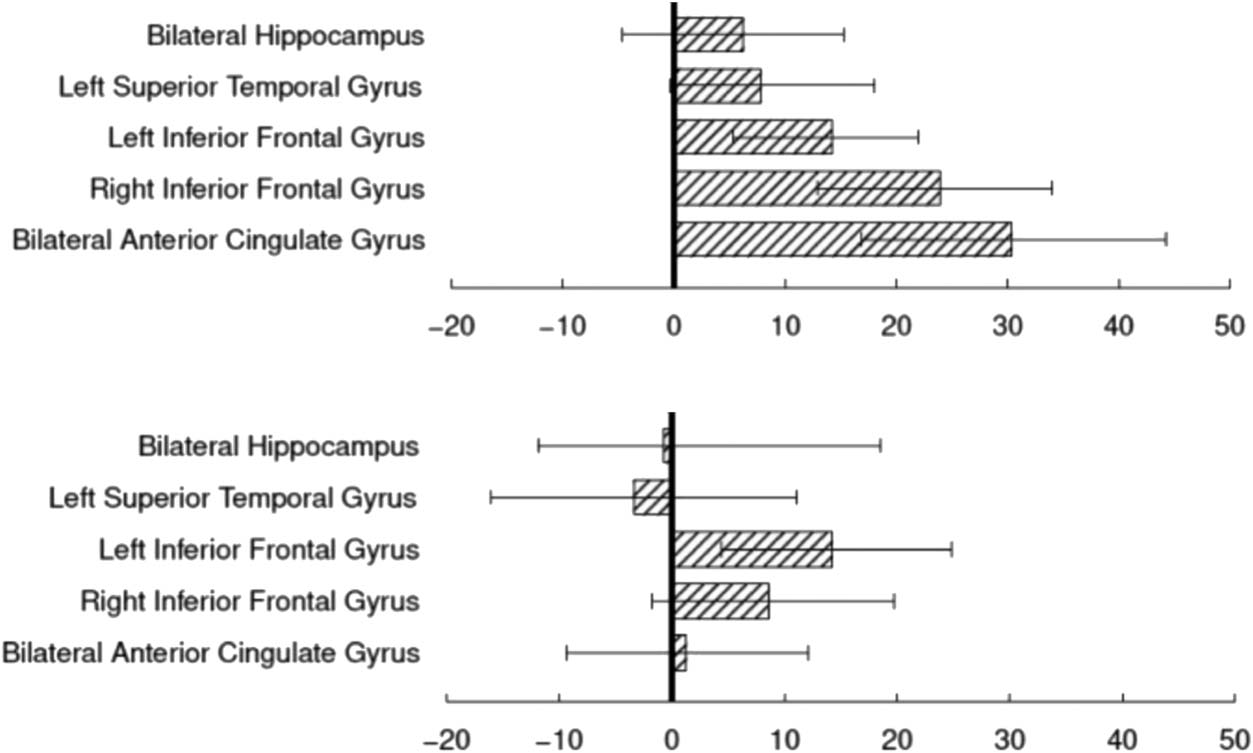
Figure 3 depicts the relationship of the early interven-
tion on percentage volume increase in the sample, sepa-
rated by sex, for volumes of the five individual a priori
ROIs. The 95% bootstrap confidence intervals show that,
for male individuals, the intervention had a positive effect
on bilateral ACC, LIFG, and RIFG, with smaller positive nu-
merical differences observed on LSTG and bilateral hippo-
campus. For female individuals, only the LIFG shows a
relationship that is comparable to that of the male partic-
ipants. Applying FDR correction to the 10 tests together,
the areas just noted were significant at q = 0.0025, 0.0025,
0.0233, and 0.0488, respectively.
Exploratory Analyses: Beyond the Volumes of
Selected ROIs and Relations to Behavior
Cortical surface area and thickness index different devel-
opmental processes, with surface area assumed to reflect
the development of cortical columns and cortical thick-
ness reflecting the development of cells within a column
as well as synapse formation, pruning, and myelination
( Johnson & de Haan, 2015). As with SES effects (Noble
& Giebler, 2020), the intervention effects were more pro-
nounced for cortical surface area than thickness.
Table 7A provides the numerical values corresponding
to the volume effects shown in Figure 3, for comparison
with the cortical surface area and cortical thickness results
reported next. Table 7B shows that males had significantly
expanded surface areas for cortex, bilateral ACC, and RIFG,
similar to the volume findings, as indicated by confidence
intervals that did not cross zero, along with LSTG; for LIFG
surface area, the confidence interval just crossed zero.
Female participants showed surface area effects only for
LIFG, similar to the findings for volume. In contrast, as
shown in Table 7C, the intervention had little effect on cor-
tical thickness, with the only one confidence interval fail-
ing to cross zero, indicating thinning of LSTG for males.
An exploratory analysis sought to assess the relations of
brain to behavioral measures of psychology in this sample.
Brain anatomy has a priori relevance to psychological func-
tion, which is one reason to study it in animals and humans.
Although further testing of this relation was not a goal of this
study, we attempted a brief confirmation that the relation
was present for the participants studied here. As detailed
in the Methods section, two psychological outcomes were
selected and examined in relation to the two relatively
global brain volumes of interest, as well as the most reliably
affected ROI, which was LIFG. Pearson correlations (and
bootstrapped p values) of the six brain–behavior relation-
ships are shown in Table 8, demonstrating each brain
Table 6. ANOVA of Cortex Volume
Source
Early Intervention
Sex
School-Age Intervention (SAI)
Early Int. × Sex
Early Int. × SAI
Sex × SAI
Early Int. × Sex × SAI
Residuals
Df
Mean Sq.
F Value
1
1
1
1
1
1
1
39
362
483
52
320
19
119
71
54
6.69
8.93
0.96
5.91
0.35
2.21
1.32
PConv
0.014
0.005
0.334
0.020
0.560
0.146
0.258
PPerm
0.019
0.037
0.121
0.017
0.902
0.065
0.295
1204
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Percent differences in
volume of individual ROIs
resulting from treatment in
male (top) and female (bottom)
participants, with 95%
confidence intervals.
Table 7. Cortex and Regional Effects of Early Intervention Relative to Comparison Groups for (A) Volume, (B) Cortical Surface Area,
and (C) Cortical Thickness
Female Participants
Male Participants
Mean %
Difference
95% CI
Lower Limit
95% CI
Upper Limit
Mean %
Difference
95% CI
Lower Limit
95% CI
Upper Limit
(A) Regional Volume
Cortex
Left Superior Temporal Gyrus
Left Inferior Frontal Gyrus
Right Inferior Frontal Gyrus
Bilateral Anterior Cingulate Gyrus
Bilateral Hippocampus
(B) Regional Surface Area
Cortex
Left Superior Temporal Gyrus
Left Inferior Frontal Gyrus
Right Inferior Frontal Gyrus
Bilateral Anterior Cingulate Gyrus
(C) Regional Mean Thickness
Cortex
Left Superior Temporal Gyrus
Left Inferior Frontal Gyrus
Right Inferior Frontal Gyrus
Bilateral Anterior Cingulate Gyrus
0.33
−3.40
14.24*
8.61
1.29
−0.67
0.87
−0.09
20.04*
1.13
−5.33
−0.38
−1.4
−4.97
1.13
6.08
−6.03
−16.05
4.33
−1.77
−9.35
−4.62
−6.45
−15.04
4.17
−7.50
−18.66
−5.72
−12.33
−15.67
−7.50
−3.03
* Indicate differences whose 95% confidence intervals do not cross zero.
7.36
11.12
24.88
19.74
12.16
15.32
9.57
17.51
38.52
8.41
7.75
5.48
8.79
5.55
8.41
14.36
10.73*
7.84
14.19*
23.88*
30.37*
6.27
13.13*
17.34*
12.63
21.99*
30.41*
−1.94
−7.38*
1.04
2.83
0.39
5.02
−0.34
5.28
12.88
16.90
−4.62
7.39
8.03
−0.39
8.56
18.62
−4.99
−12.82
−5.93
−3.50
−5.81
15.73
18.02
21.96
33.98
44.22
15.32
18.43
26.96
24.85
37.47
42.59
0.83
−1.79
8.61
9.12
6.93
Farah et al.
1205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 8. Pearson Correlations between Brain Volumes and
Psychological Measures (Bootstrapped, FDR-Corrected q
Values) of Six Brain–Behavior Relationships
Brain Region
Stanford–Binet
Midlife Strengths
and Risks Index
Summed ROIs
+0.29 (0.045)
+0.21 (0.191)
Cortex
LIFG
+0.42 (0.015)
+0.23 (0.191)
+0.36 (0.015)
+0.35 (0.016)
measure was significantly associated in the expected direc-
tion with one or both of two psychological outcomes.
Finally, the relative volumes of all 134 regions delineated
by the Desikan–Killiany atlas, as well as the surface areas and
thicknesses of cortical regions, were compared for early
intervention and comparison participants, separated by
sex. The three large tables containing these results and their
95% confidence intervals are included with raw data at the
Figshare link listed at the end of the paper. We offer these
data for descriptive purposes to readers seeking additional
information. Given the many regions tested, caution re-
garding potential false positives is warranted.
Of the regions subjected to exploratory analysis, a num-
ber showed substantial volume increases. More regions
overall showed volume increases from the intervention
in male than female participants, 42 (all positive in sign)
versus six (of which five were positive). Similarly, surface
area showed numerous positively signed differences for
male individuals and many fewer such differences observed
for female individuals. Intervention effects on cortical thick-
ness were overall fewer in number for both sexes and in-
cluded both positive and negative differences.
One question of interest addressed by the exploratory
analyses is whether the sex difference observed with the
a priori brain measures is specific to those measures.
Before examining the different anatomical dimensions of
the rest of the cortex and brain in all brain regions, one
might have thought that anatomy is affected for both sexes
equally, but with different regional distributions or differ-
ent manifestations in volume, surface area, or thickness.
The findings here indicate that this is not the case.
Rather, the results obtained throughout the brain suggest
that macroscopic brain structure is more affected by early
life cognitive and linguistic stimulation in male than in fe-
male individuals.
DISCUSSION
Here we report the first evidence that normal variation in
early life experience impacts human brain structure.
Specifically, we show that the cognitive and linguistic envi-
ronment of young humans affects macroscopic brain struc-
ture. Unlike previous observational research, which cannot
address causality, the present data show that early life expe-
rience shapes brain structure, through its immediate causal
effects and continuing chains of causal consequences.
Only one other randomized experimental study of early
life experience in humans has reported brain measures,
the Bucharest Early Intervention Project. It differs
from this study in two ways. First, it could not shed light
on the earliest years of human development, because its
randomized component started at 2 years, as opposed to
early infancy. Second, the manipulation involved a general
and severe perturbation of childhood experience, includ-
ing limited social, emotional, motoric experience in addi-
tion to limited cognitive and linguistic experience in
Romanian orphanages. At 2 years of age, children were
randomly assigned to foster care or continued institutional
care. The impact of home rearing improved, but did not
fully restore, later cognitive abilities, psychological adjust-
ment, or brain structure (Mackes et al., 2020; Sheridan, Fox,
Zeanah, McLaughlin, & Nelson, 2012). The inability of the
fostering experience to “rescue” the brain from pathological
treatment in the first 2 years does not address the question
of interest here: whether experience changes brain struc-
ture in the context of normal human development, such
that higher versus lower levels of cognitive and linguistic
stimulation in the earliest years of life make a difference
in brain structure. The present study therefore provides
unique information about the causal relationship between
early life experience and human brain structure, and the
specific effect of cognitive and linguistic stimulation.
The present findings are also relevant to understanding
the recently observed relation between brain structure and
socioeconomic status (Noble & Giebler, 2020). Two general
types of explanation have been put forward for this relation.
On the one hand, it may be that environmental causes, such
as the well-documented disparities in opportunities for
cognitive stimulation and child-directed speech, are
responsible, which is called a “social causation” account
because the social environment causes the observed differ-
ences (Dunham, 1961). On the other hand, genetic inheri-
tance of neural and cognitive differences may operate, and
insofar as these differences influence SES, they could
accountwhich is called a for the relations between brain,
cognition, and SES (Murray, 2020; Wax, 2017), called a
“social selection” account because different levels of SES
select individuals based on their innate capabilities
(Dunham, 1961). In order for brain disparities to be
accounted for by the first type of account, it must be the
case that cognitive and linguistic experience impacts brain
structure. The present results provide the first evidence
that this is true.
The results showed a pronounced sex difference in the
effect of the early intervention, with larger effects on
males. The only a priori ROI for which the intervention
benefitted females to the same degree as males was the
left inferior frontal gyrus; female participants showed non-
significant trends in some but not all other a priori areas.
Of note, animal studies measuring gross anatomical effects
of environmental stimulation frequently include only
males, and a variety of differences have been reported
when both sexes are included (Diamond, 2001).
1206
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
For humans, it is not uncommon for childhood interven-
tion studies to find differences in efficacy for the behavior of
male and female individuals (García, Heckman, & Ziff, 2018;
Chetty, Hendren, & Katz, 2016). A recent meta-analysis of
sex differences in response to early childhood education
across studies found small sex differences favoring female in-
dividuals for most outcomes, with a more pronounced effect
of reduced grade retention and special education referral fa-
voring male individuals (Magnuson et al., 2016). For the
Abecedarian Early Intervention, many of the young adult be-
havioral measures showed more lasting effects for female
than male participants (Campbell et al., 2019), with the ex-
ception that later cardiovascular and metabolic health
indicators showed more benefits to male participants
(Campbell et al., 2014).
The reasons for sex differences in the Abecedarian and
other program outcomes are poorly understood. They
could involve biological differences between the sexes or
social differences in their lives or both. Furthermore, these
differences could moderate the intervention effect
through either their effects on sensitivity to environmental
enrichment in the intervention group or to the conditions
of poverty in the comparison group or both (García et al.,
2018; Golding & Fitzgerald, 2017).
Limitations of this study include those intrinsic to the
sample and those intrinsic to MRI. Regarding the sample,
it is small compared to most current studies in cognitive
neuroscience. The trend toward larger samples has been
motivated in part by the realization that they reduce the
risk of false positives, in addition to the more obvious
reduction in risk of false negatives (Button et al., 2013).
Sample size impacts false positives by its relation to statis-
tical power, and power in turn depends on expected effect
size. Crucially, power and replicability are not determined
simply by sample size per se, but rather by sample size in
relation to the size of the effect being tested. As discussed
in the Methods section, our sample is adequately powered
to detect a large effect, and a large effect is plausible given
effect sizes from comparable studies in humans and ani-
mals. On this basis, it was appropriate to proceed with
analyses of the sample. The effects we found were also
large, and the possibility that they were false positives,
by chance yielding p values below 0.05, is unlikely. As
shown in Table 5, the main effect of the intervention on
the a priori summary measure was highly significant by
conventional and permutation testing; in the latter case,
the precise value was 0.000000004, truncated to 0.000 in
the table. In summary, although recent concerns about
sample size in neuroscience are well-justified in general,
they do not call into question this study and its findings.
Regarding MRI, it does not reveal changes in the brain
at the cellular level. On the basis of our data, we cannot
know whether the intervention affected size or number
of neuronal or glial cell bodies, of dendrites, synapses, or
other experience-dependent features of brain tissue doc-
umented by animal research. Furthermore, the study is
limited by having images from just one stage of life, long
after the conclusion of the intervention. While the endur-
ing nature of the effect adds to its potential practical im-
portance, it would have been ideal to scan participants
longitudinally, starting in infancy, in order to further con-
strain the ways in which early childhood experience and
the causal chain of its later life effects impact the brain.
In the absence of such data, we can nevertheless con-
clude that early life cognitive and linguistic stimulation im-
pacts brain structure, in the form of larger volumes of brain
regions associated with cognition. At present, only a small
number of human beings in the world have ever under-
gone early, intensive, and sustained cognitive and linguis-
tic intervention with random assignment, namely, the
participants of the Abecedarian project. Their brain struc-
ture findings extend, in a qualitative way, our knowledge of
experiential effects on the brain. They also argue for in-
vestment in future randomized intervention studies, with
longitudinal, multimodal imaging and behavioral mea-
sures starting in infancy.
Acknowledgments
The authors thank Carrie Bynum and Laura Bateman for their
assistance in data collection and Vincent Hurtubise for computer
systems support.
Reprint requests should be sent to Martha J. Farah, Center for
Neuroscience & Society, University of Pennsylvania, 3710
Hamilton Walk, Goddard Labs 506, Philadelphia, PA 19104, or
via e-mail: mfarah@psych.upenn.edu.
Author Contributions
Martha J. Farah: Conceptualization; Formal analysis;
Investigation; Methodology; Writing—Original draft. Saul
Sternberg: Conceptualization; Formal analysis; Software;
Visualization; Writing—Review & editing. Thomas A.
Nichols: Data curation; Formal analysis; Writing—Original
draft. Jeffrey T. Duda: Data curation; Writing—Original draft.
Terry Lohrenz: Data curation; Investigation; Methodology;
Writing—Review & editing. Yi Luo: Investigation;
Methodology; Writing—Review & editing. Libbie Sonnier:
Data curation; Writing—Review & editing. Sharon L.
Ramey: Conceptualization; Resources; Writing—Review &
editing. Read Montague: Conceptualization; Investigation;
Methodology; Resources; Writing—Review & editing. Craig
T. Ramey: Conceptualization; Resources; Writing—Review
& editing.
Funding Information
This work was supported by a Principal Research Fellowship
from the Wellcome Trust (R. M.), Virginia Tech (R. M.) and
the School of Arts and Sciences Research Fund, University
of Pennsylvania (M. J. F.).
Data and Materials Availability
Anonymized brain measures analyzed here with group
membership, age, and sex, as well as analyzed regional
Farah et al.
1207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
differences in volume, surface area, and thickness, are
a v a i l a b l e a t h t t p s : / / f i g s h a r e . c o m / a r t i c l e s / E a r l y
_Experience_Volume_Cortical_Thickness_Surface_Area
_data/9161894.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publish-
ing in the Journal of Cognitive Neuroscience ( JoCN )
during this period were M(an)/M = .408, W(oman)/M =
.335, M/W = .108, and W/W = .149, the comparable pro-
portions for the articles that these authorship teams cited
were M/M = .579, W/M = .243, M/W = .102, and W/W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender
citation balance. The authors of this article report its pro-
portions of citations by gender category to be as follows:
M/M = .542, W/M = .125, M/W = .167, and W/W = .167.
REFERENCES
Aron, A. R., Robbins, T. W., & Poldrack, R. A. (2004). Inhibition
and the right inferior frontal cortex. Trends in Cognitive
Sciences, 8, 170–177. DOI: https://doi.org/10.1016/j.tics.2004
.02.010, PMID: 15050513
Avants, B. B., Epstein, C. L., Grossman, M., & Gee, J. C. (2008).
Symmetric diffeomorphic image registration with cross-
correlation: Evaluating automated labeling of elderly and
neurodegenerative brain. Medical Image Analysis, 12, 26–41.
DOI: https://doi.org/10.1016/j.media.2007.06.004, PMID:
17659998, PMCID: PMC2276735
Bale, T. L., Baram, T. Z., Brown, A. S., Goldstein, J. M., Insel, T. R.,
McCarthy, M. M., et al. (2010). Early life programming and
neurodevelopmental disorders. Biological Psychiatry, 68,
314–319. DOI: https://doi.org/10.1016/j.biopsych.2010.05.028,
PMID: 20674602, PMCID: PMC3168778
Bradley, R. H., & Corwyn, R. F. (2002). Socioeconomic status
and child development. Annual Review of Psychology, 53,
371–399. DOI: https://doi.org/10.1146/annurev.psych.53
.100901.135233, PMID: 11752490
Button, K. S., Ioannidis, J. P. A., Mokrysz, C., Nosek, B. A., Flint,
J., Robinson, E. S. J., et al. (2013). Power failure: Why small
sample size undermines the reliability of neuroscience.
Nature Reviews Neuroscience, 14, 365–376. DOI: https://
doi.org/10.1038/nrn3475, PMID: 23571845
Campbell, F. A., Conti, G., Heckman, J. J., Moon, S. H., Pinto, R.,
Pungello, E., et al. (2014). Early childhood investments
substantially boost adult health. Science, 343, 1478–1485.
DOI: https://doi.org/10.1126/science.1248429, PMID:
24675955, PMCID: PMC4028126
Campbell, F. A., Pan, Y., & Burchinal, M. (2019). Sustaining
gains from early childhood intervention: The Abecedarian
program. In A. J. Reynolds & J. Temple (Eds.), Sustaining
early childhood learning gains: Program, school, and family
influences (pp. 268–286). New York: Cambridge University
Press. DOI: https://doi.org/10.1017/9781108349352.013
Campbell, F. A., Pungello, E. P., Miller-Johnson, S., Burchinal,
M., & Ramey, C. T. (2001). The development of cognitive and
academic abilities: Growth curves from an early childhood
educational experiment. Developmental Psychology, 37,
231–242. DOI: https://doi.org/10.1037/0012-1649.37.2.231,
PMID: 11269391
Canty, A., & Ripley, B. D. (2019). boot: Bootstrap R (S-Plus)
functions (R package version 1.3-23). Retreived from https://
cran.r-project.org/web/packages/boot/index.html.
Chetty, R., Hendren, N., & Katz, L. F. (2016). The effects
of exposure to better neighborhoods on children: New
evidence from the Moving to Opportunity experiment.
American Economic Review, 106, 855–902. DOI: https://
doi.org/10.1257/aer.20150572, PMID: 29546974
Cohen, J. (1992). A power primer. Psychological Bulletin, 112,
155–159. DOI: https://doi.org/10.1037/0033-2909.112.1.155,
PMID: 19565683
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., et al. (2006). An automated labeling system
for subdividing the human cerebral cortex on MRI scans into
gyral based regions of interest. Neuroimage, 31, 968–980.
DOI: https://doi.org/10.1016/j.neuroimage.2006.01.021,
PMID: 16530430
Diamond, M. C. (2001). Response of the brain to enrichment.
Anais da Academia Brasileira de Ciências, 73, 211–220.
DOI: https://doi.org/10.1590/S0001-37652001000200006,
PMID: 11404783
Duncan, G. J., Magnuson, K. A., & Votruba-Drzal, E. (2014).
Boosting family income to promote child development.
Future of Children, 24, 99–120. DOI: https://doi.org/10.1353
/foc.2014.0008, PMID: 25518705
Dunham, H. W. (1961). Social structures and mental disorders:
Competing hypotheses of explanation. Milbank Memorial
Fund Quarterly, 39, 259–311. DOI: https://doi.org/10.2307
/3348602, PMID: 13888471
Farah, M. J. (2018). Socioeconomic status and the brain:
Prospects for neuroscience-informed policy. Nature Reviews
Neuroscience, 19, 428–438. DOI: https://doi.org/10.1038
/s41583-018-0023-2
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007).
G*Power 3: A flexible statistical power analysis program for
the social, behavioral, and biomedical sciences. Behavior
Research Methods, 39, 175–191. DOI: https://doi.org/10
.3758/BF03193146, PMID: 17695343
Friederici, A. D. (2011). The brain basis of language processing:
From structure to function. Physiological Reviews, 91,
1357–1392. DOI: https://doi.org/10.1152/physrev.00006.2011,
PMID: 22013214
García, J. L., Heckman, J. J., & Ziff, A. L. (2018). Gender
differences in the benefits of an influential early childhood
program. European Economic Review, 109, 9–22. DOI:
https://doi.org/10.1016/j.euroecorev.2018.06.009, PMID:
30410186, PMCID: PMC6217989
Golding, P., & Fitzgerald, H. E. (2017). Psychology of boys at
risk: Indicators from 0–5. Infant Mental Health Journal,
38, 5–14. DOI: https://doi.org/10.1002/imhj.21621, PMID:
27959473
Hanson, J. L., Nacewicz, B. M., Sutterer, M. J., Cayo, A. A.,
Schaefer, S. M., Rudolph, K. D., et al. (2015). Behavioral
problems after early life stress: Contributions of the
hippocampus and amygdala. Biological Psychiatry, 77,
314–323. DOI: https://doi.org/10.1016/j.biopsych.2014
.04.020, PMID: 24993057, PMCID: PMC4241384
Hoff, E. (2013). Interpreting the early language trajectories
of children from low-SES and language minority homes:
Implications for closing achievement gaps. Developmental
Psychology, 49, 4–14. DOI: https://doi.org/10.1037/a0027238,
PMID: 22329382, PMCID: PMC4061698
1208
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Johnson, M. H., & de Haan, M. (2015). Developmental
cognitive neuroscience (4th ed.). Malden, MA: Wiley.
Lenroot, R. K., & Giedd, J. N. (2011). Annual research review:
Developmental considerations of gene by environment
interactions. Journal of Child Psychology and Psychiatry,
52, 429–441. DOI: https://doi.org/10.1111/j.1469-7610.2011
.02381.x, PMID: 21391998, PMCID: PMC3268527
Luo, Y., Hétu, S., Lohrenz, T., Hula, A., Dayan, P., Ramey, S. L.,
et al. (2018). Early childhood investment impacts social
decision-making four decades later. Nature Communications,
9, 4705. DOI: https://doi.org/10.1038/s41467-018-07138-5,
PMID: 30459305, PMCID: PMC6246600
Mackes, N. K., Golm, D., Sarkar, S., Kumsta, R., Rutter, R.,
Fairchild, G., et al. (2020). Early childhood deprivation is
associated with alterations in adult brain structure despite
subsequent environmental enrichment. Proceedings of the
National Academy of Sciences, U.S.A., 117, 641–649. DOI:
https://doi.org/10.1073/pnas.1911264116, PMID: 31907309,
PMCID: PMC6955353
Magnuson, K. A., Kelchen, R., Duncan, G. J., Schindler, H. S.,
Shager, H., & Yoshikawa, H. (2016). Do the effects of early
childhood education programs differ by gender? A meta-
analysis. Early Childhood Research Quarterly, 36, 521–536.
DOI: https://doi.org/10.1016/j.ecresq.2015.12.021, PMID:
31576062, PMCID: PMC6771425
Murray, C. A. (2020). Human diversity: The biology of gender,
race, and class. New York: Hachette Books.
Noble, K. G., & Giebler, M. A. (2020). The neuroscience of
socioeconomic inequality. Current Opinion in Behavioral
Sciences, 36, 23–28. DOI: https://doi.org/10.1016/j.cobeha
.2020.05.007, PMID: 32719820, PMCID: PMC7384696
Peng, R. D. (2019). simpleboot: Simple bootstrap routines
(R package version 1.1-7). Retrieved from https://CRAN.R
-project.org/package=simpleboot.
Peterson, J. W., Loeb, S., & Chamberlain, L. J. (2018). The
intersection of health and education to address school
readiness of all children. Pediatrics 142, e20181126. DOI:
https://doi.org/10.1542/peds.2018-1126, PMID: 30366953
Ramey, C. T., Campbell, F. A., Burchinal, M., Skinner, M. L.,
mothers. Applied Developmental Science, 4, 2–14. DOI:
https://doi.org/10.1207/S1532480XADS0401_1
Rosenzweig, M. R. (1966). Environmental complexity, cerebral
change, and behavior. American Psychologist, 21, 321–332.
DOI: https://doi.org/10.1037/h0023555, PMID: 5910063
Sheridan, M. A., Fox, N. A., Zeanah, C. H., McLaughlin, K. A., &
Nelson, C. A. (2012). Variation in neural development as a
result of exposure to institutionalization early in childhood.
Proceedings of the National Academy of Sciences, U.S.A.,
109, 12927–12932. DOI: https://doi.org/10.1073/pnas
.1200041109, PMID: 22826224, PMCID: PMC3420193
Sonnier-Netto, M. E. (2018). The association between early care
and education and midlife outcomes: The Abecedarian 5th
Decade Follow-up (Doctoral dissertation. Blacksburg, Virginia:
Virginia Polytechnic Institute and State University.
Sparling, J., & Lewis, I. (1984). LearningGames for threes and
fours: A guide to adult/child play. New York: Berkley Books.
DOI: https://doi.org/10.1007/BF01617063
Thalheimer, W., & Cook, S. (2002). How to calculate effect sizes
from published research articles: A simplified methodology.
Retrieved from http://work-learning.com/effect_sizes.htm.
Tustison, N. J., Cook, P. A., Klein, A., Song, G., Das, S. R., Duda,
J. T., et al. (2014). Large-scale evaluation of ANTs and
FreeSurfer cortical thickness measurements. Neuroimage,
99, 166–179. DOI: https://doi.org/10.1016/j.neuroimage
.2014.05.044, PMID: 24879923
Wang, H., Suh, J. W., Das, S. R., Pluta, J. B., Craige, C., &
Yushkevich, P. A. (2013). Multi-atlas segmentation with
joint label fusion. IEEE Transactions on Pattern Analysis
and Machine Intelligence, 35, 611–623. DOI: https://doi
.org/10.1109/TPAMI.2012.143, PMID: 22732662, PMCID:
PMC3864549
Wax, A. L. (2017). The poverty of the neuroscience of poverty:
Policy payoff or false promise? (SSRN Scholarly Paper ID
2888600). Rochester, NY: Social Science Research Network.
Weiss, N. A. (2016). wBoot: Bootstrap methods (R package
version 1.0.3). Retrieved from https://CRAN.Rproject.org
/package=wBoot.
Wheeler, B., & Torchiano, M. (2016). lmPerm: Permutation
Gardner, D. M., & Ramey, S. L. (2000). Persistent effects of
early childhood education on high-risk children and their
tests for linear models (R package version 2.1.0). Retrieved
from https://CRAN.R-project.org/package=lmPerm.
Farah et al.
1209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
1
9
7
1
9
5
8
7
6
8
/
/
j
o
c
n
_
a
_
0
1
7
0
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3